retro-Tango enables versatile retrograde circuit tracing in Drosophila
- Department of Neuroscience, Brown University, United States
- Carney Institute for Brain Science, Brown University, United States
Figures
Figure 1 with 2 supplements
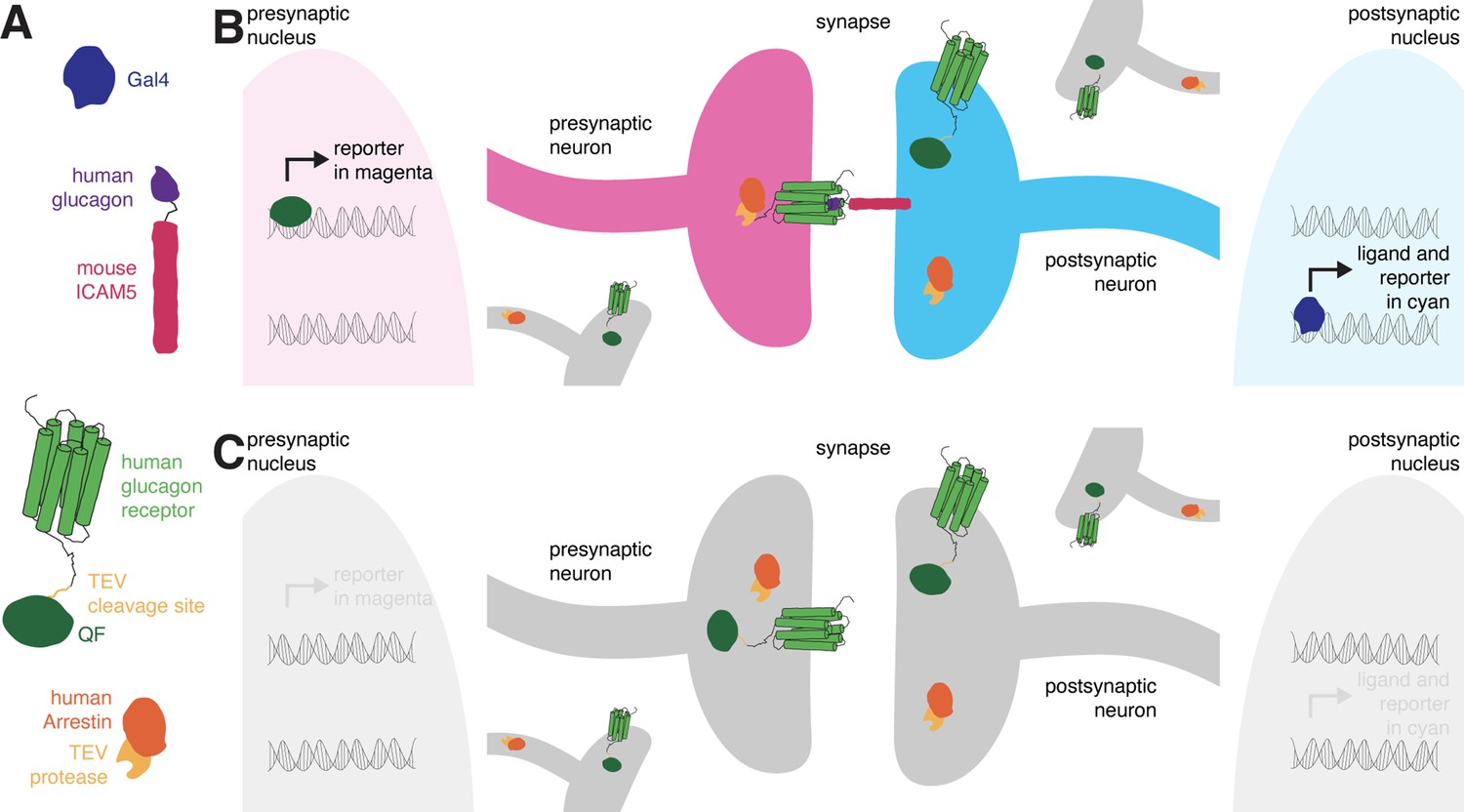
The design of retro-Tango.
(A) The components of retro-Tango. (B) In retro-Tango, all neurons express two of the components of the signaling cascade: human glucagon receptor::TEV cleavage site::QF and human β-arrestin2::TEV protease. They also carry the gene encoding the presynaptic mtdTomato reporter (magenta) under the control of QF. Therefore, all neurons are capable of expressing the reporter. In starter neurons expressing Gal4, the ligand (human glucagon::mouse ICAM5) is expressed along with the GFP reporter (cyan) marking the postsynaptic starter neurons. The mICAM5 fusion localizes the ligand to the postsynaptic sites such that the ligand activates its receptor only across the synapse. Upon activation of the receptor in the presynaptic neuron, the Arrestin-TEV fusion is recruited. TEV-mediated proteolytic cleavage then releases the transcription factor QF from the receptor. QF in turn translocates to the nucleus and initiates transcription of the presynaptic magenta reporter. In neurons that are not presynaptic to the starter neurons, the reporter is not expressed. (C) In the absence of a Gal4 driver, the ligand is not expressed, and the signaling cascade is not triggered, resulting in no expression of the reporters.
Figure 1—figure supplement 1
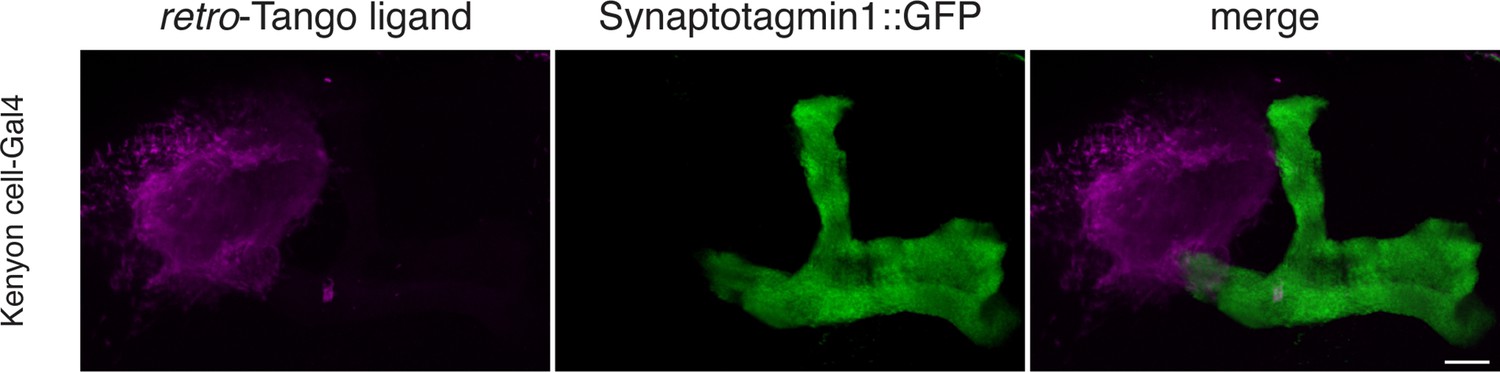
The retro-Tango ligand localizes to dendrites and somata.
The retro-Tango ligand and GFP-tagged Synaptotagmin1 was expressed in Kenyon cells of the mushroom body. The retro-Tango ligand localizes to the cell bodies and the mushroom body calyx where the dendrites of Kenyon cells reside. It is however absent in axons as it does not colocalize with the GFP-tagged Synaptotagmin1. Subset of the z-stack is shown for clarity. Syt::GFP (green), myc (magenta). Scale bar, 10 μm.
Figure 1—figure supplement 2

The genetic components of retro-Tango.
Details of the genetic components used in retro-Tango are shown. Schematics are not drawn to scale. Elav: Drosophila melanogaster panneuronal promoter; polyA: polyadenylation signal; nSyb: Drosophila melanogaster panneuronal promoter; DSCP: Drosophila Synthetic Core Promoter; hGCGR: human Glucagon Receptor; TEVcs: cleavage site for N1a protease from the Tobacco Etch Virus; UAS: Upstream Activating Sequence for Gal4; hGCG: human Glucagon analogue with enhanced receptor binding; mICAM5: mouse intercellular adhesion molecule 5; P2A: 2A peptide from porcine teschovirus-1; GFPfar: farnesylated Green Fluorescent Protein; QUAS: Upstream Activating Sequence for QF.
Figure 2 with 4 supplements
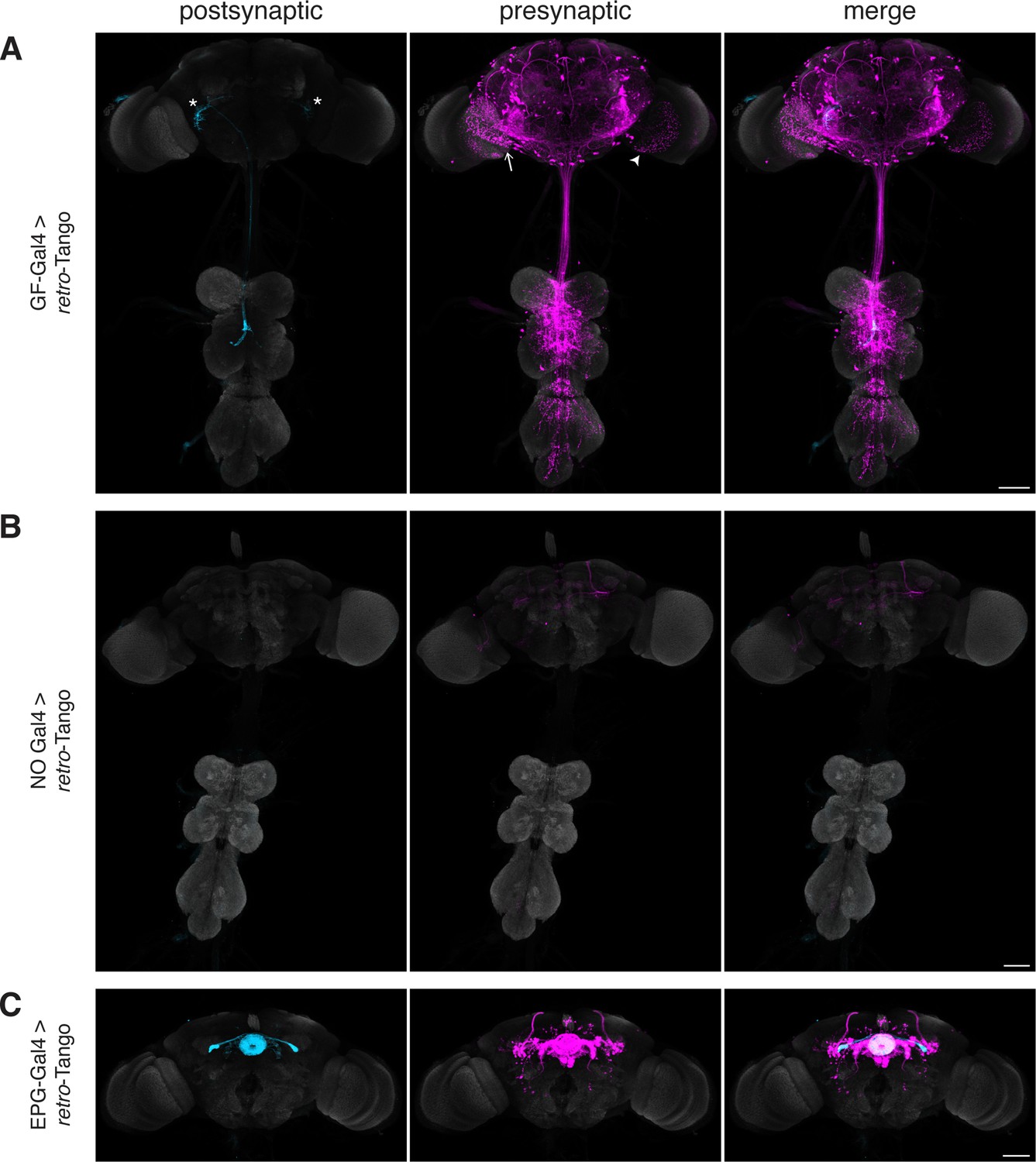
Implementation of retro-Tango in the giant fiber and central complex circuits.
(A) Initiating retro-Tango from the GFs (asterisks mark the cell bodies) results in presynaptic signal in the brain and VNC (223±59 neurons in 5 brains, 1±3 neurons in 5 VNCs). Both LC4 (arrow) and LPLC2 (arrowhead) neurons, known presynaptic partners of GFs, are identified by retro-Tango. Note the asymmetry between hemispheres in the signal in the postsynaptic starter neurons and their corresponding presynaptic partners. (B) retro-Tango exhibits little background noise in the absence of a Gal4 driver. Background is observed in the mushroom bodies, in the central complex, and in a few neurons in the VNC (68±10 neurons in 4 brains, 1±1 neurons in 4 VNCs). (C) Ligand expression in EPG neurons of the central complex leads to retro-Tango signal in their known presynaptic partners: PEN, PFR and Δ7 neurons (170±24 neurons in 5 brains). The signal in these neurons can be easily discerned from the background noise. 15do males were analyzed for all panels. Postsynaptic GFP (cyan), presynaptic mtdTomato (magenta) and neuropil (grey). Scale bars, 50 μm.
Figure 2—figure supplement 1
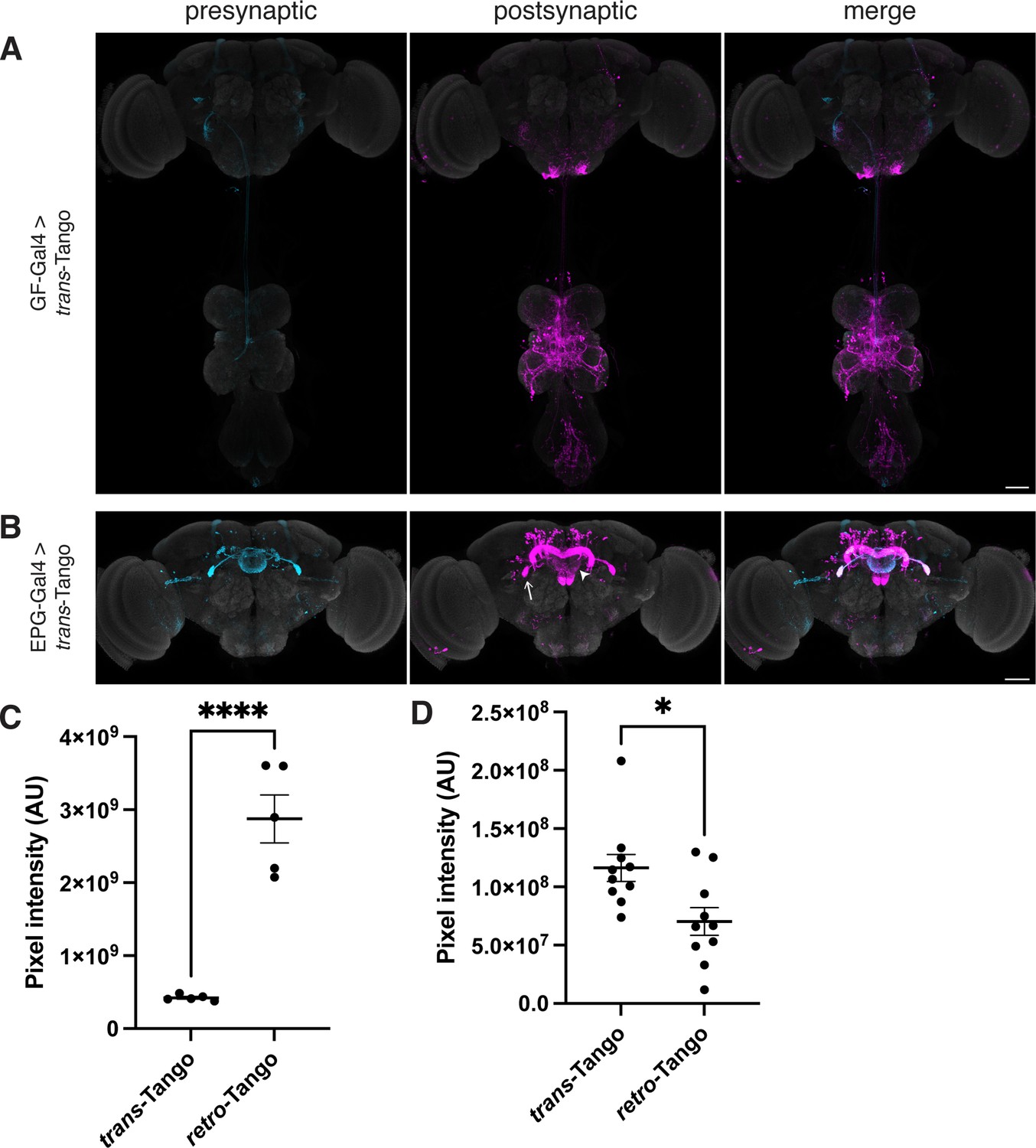
trans-Tango in the giant fiber and central complex circuits.
(A) Initiating trans-Tango from the GFs results in strong postsynaptic signal in the VNC and little to no signal in the brain (4±2 neurons in 4 brains, 48±16 neurons in 4 VNCs). (B) Expression of the trans-Tango ligand in the EPG neurons of the central complex reveals their postsynaptic partners (255±22 neurons in 5 brains). Note the stronger signal in the LAL (arrow) and the weaker signal in the EB (arrowhead) compared to retro-Tango results (Figure 2C). 20do males were analyzed for both panels. Presynaptic GFP (cyan), postsynaptic mtdTomato (magenta) and neuropil (grey). Scale bars, 50 μm. (C) Comparison of the pixel intensities for the signals of retro-Tango and trans-Tango for the EPG circuit in the ellipsoid body (n=5 brains each). (D) Comparison of the pixel intensities for the signals of retro-Tango and trans-Tango for the EPG circuit in the lateral accessory lobes (n=10 hemibrains each). Dots represent data points, the horizonal lines represent the mean and the error bars represent the standard error of the mean. Student’s t-test, *: p<0.05, ****: p<0.0001.
Figure 2—figure supplement 2
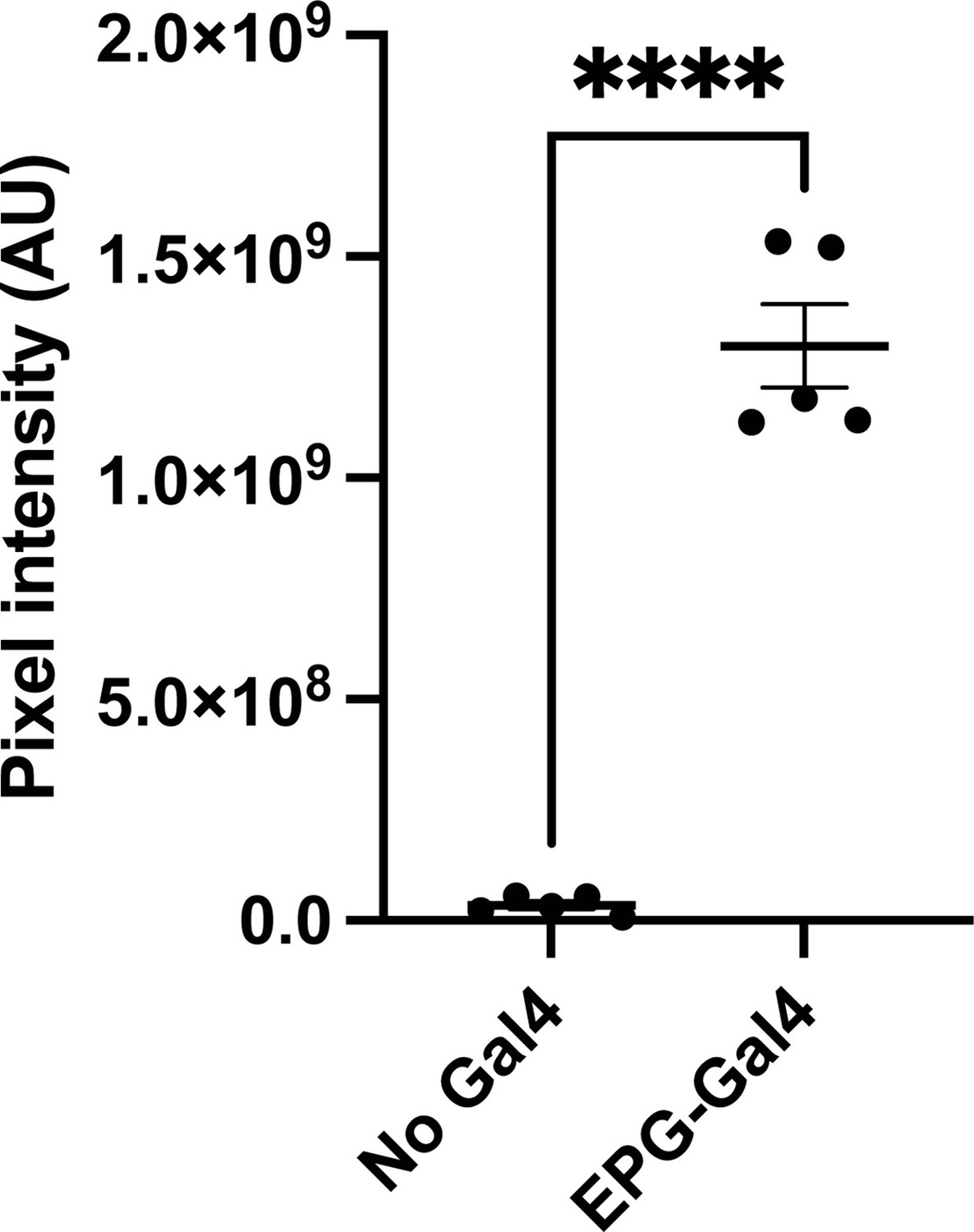
The retro-Tango signal in the EPG circuit is far stronger than the background noise.
Comparison of the pixel intensities in the central complex for the background noise signal of retro-Tango and retro-Tango signal when initiated from EPG neurons (n=5 brains each). Dots represent data points, the horizonal lines represent the mean and the error bars represent the standard error of the mean. Student’s t-test, ****: p<0.0001.
Figure 2—figure supplement 3
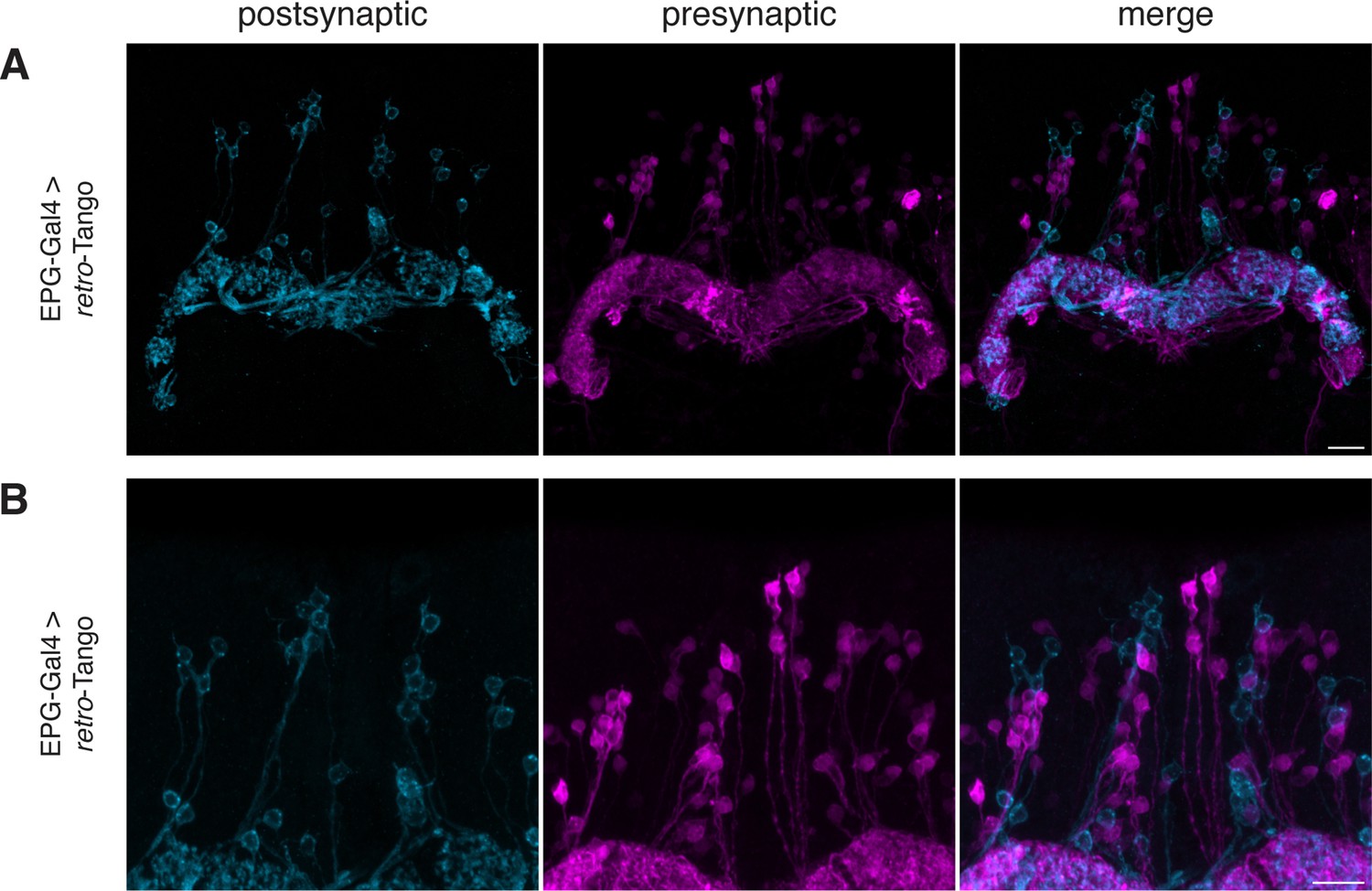
retro-Tango does not yield false positive signal in neighboring neurons in the EPG circuit.
(A–B) When retro-Tango is initiated from EPG neurons, the ligand present in the cell bodies does not lead to false positive presynaptic signal in neighboring neurons. For clarity, only a subset of the z-stack projection is shown. 15do males were analyzed. Postsynaptic GFP (cyan), presynaptic mtdTomato (magenta). Scale bars, 10 μm.
Figure 2—video 1
retro-Tango does not yield false positive signal in neighboring neurons in the EPG circuit.
Video through the z-stack sections of the image in Figure 2—figure supplement 3B. Postsynaptic GFP (cyan), presynaptic mtdTomato (magenta).
Figure 3 with 3 supplements

Age dependence of retro-Tango.
The retro-Tango signal is observed in 5 day intervals upon ligand expression in the EPGs. The signal accumulates with time and saturates around day 10 post-eclosion. Males were analyzed for all panels. Presynaptic mtdTomato (magenta) and neuropil (grey). Scale bars, 50 μm.
Figure 3—figure supplement 1

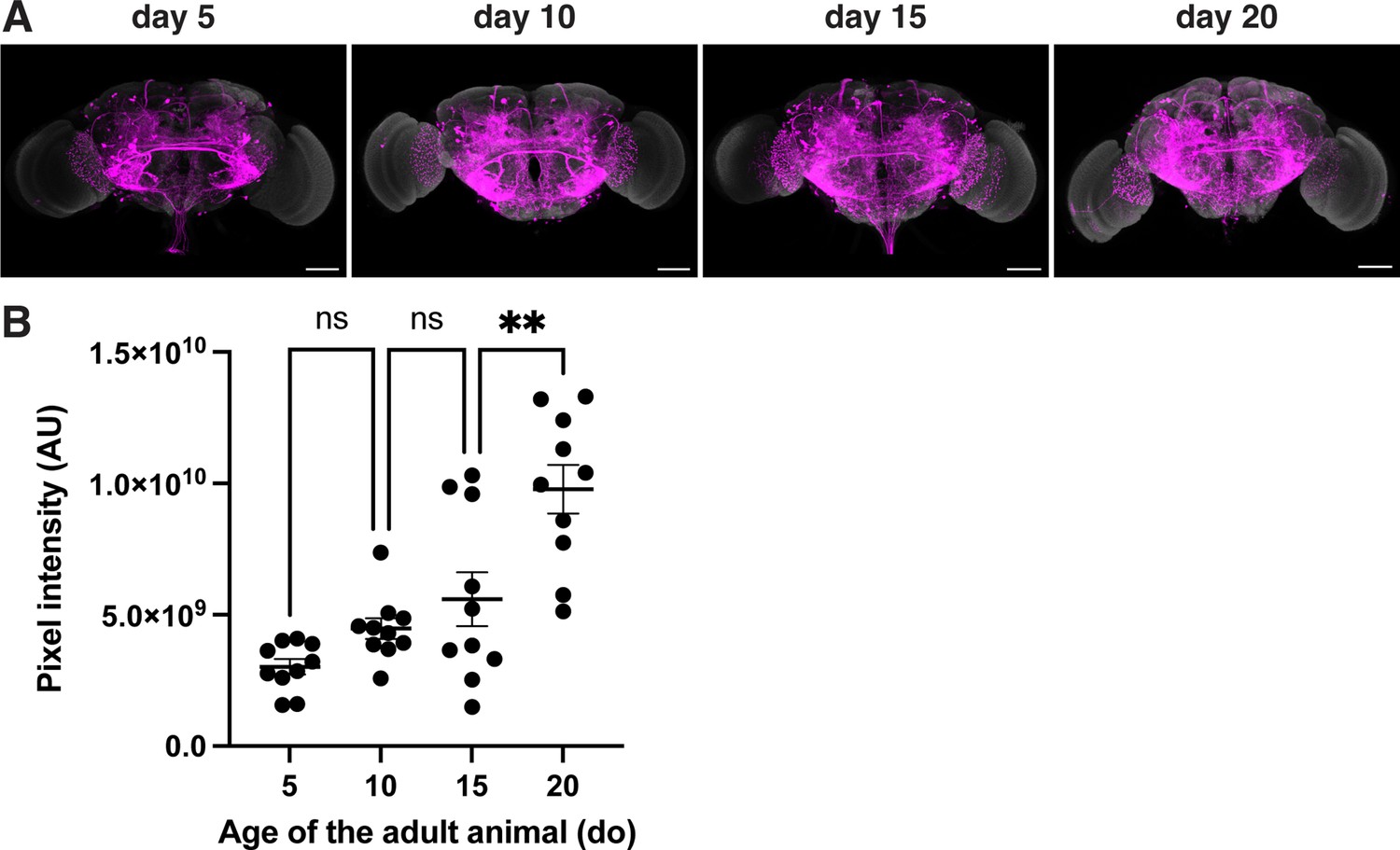
Quantification of the pixel intensity for the signal of retro-Tango when initiated from the EPG neurons.
Comparison of the pixel intensities in the central complex for the presynaptic signal in males of different ages where the retro-Tango was initiated from the EPG neurons (n=5 brains each). Dots represent data points, the horizonal lines represent the mean and the error bars represent the standard error of the mean. One-way ANOVA, *: p<0.05, ns: not significant.
Figure 3—figure supplement 2
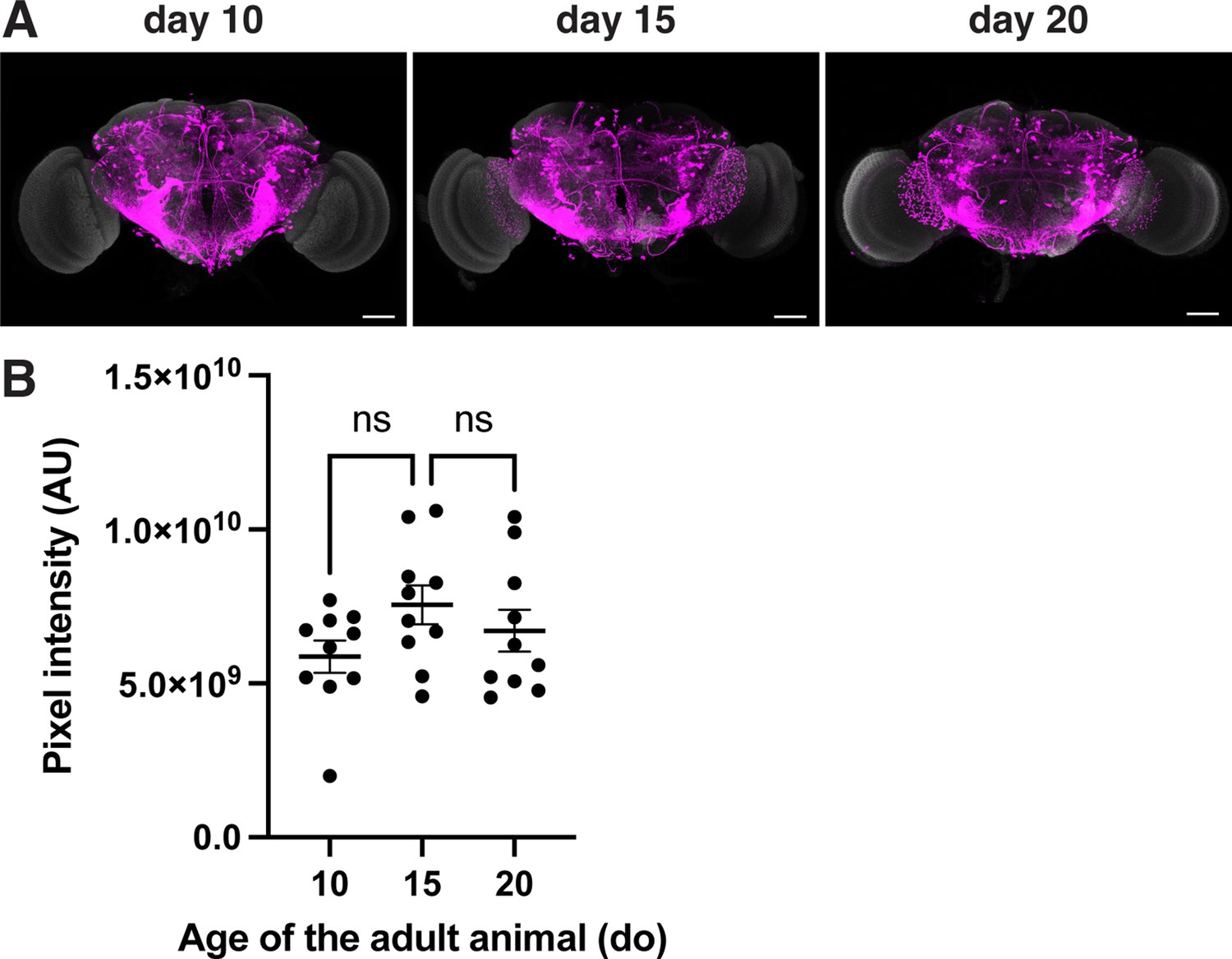
Age dependence of the retro-Tango signal in the presynaptic partners of the GFs in males.
(A) retro-Tango signal is observed in 5-day intervals upon ligand expression in the GFs. The signal accumulates over time. Males were analyzed for all panels. Presynaptic mtdTomato (magenta) and neuropil (grey). Scale bars, 50 μm. (B) Comparison of the pixel intensities in the whole brain for the presynaptic signal in males of different ages where the retro-Tango was initiated from the GF neurons (n=10 hemibrains each). Dots represent data points, the horizonal lines represent the mean and the error bars represent the standard error of the mean. One-way ANOVA, **: p<0.01, ns: not significant.
Figure 3—figure supplement 3
Age dependence of the retro-Tango signal in the presynaptic partners of the GFs in females heterozygous for the reporter.
(A) retro-Tango signal is observed in 5-day intervals upon ligand expression in the GFs. The signal does not seem to change significantly over time. Females heterozygous for the reporter were analyzed for all panels. Presynaptic mtdTomato (magenta) and neuropil (grey). Scale bars, 50 μm. (B) Comparison of the pixel intensities in the whole brain for the presynaptic signal in females of different ages where the retro-Tango was initiated from the GF neurons (n=10 hemibrains each). Dots represent data points, the horizonal lines represent the mean and the error bars represent the standard error of the mean. One-way ANOVA, ns: not significant.
Figure 4 with 2 supplements
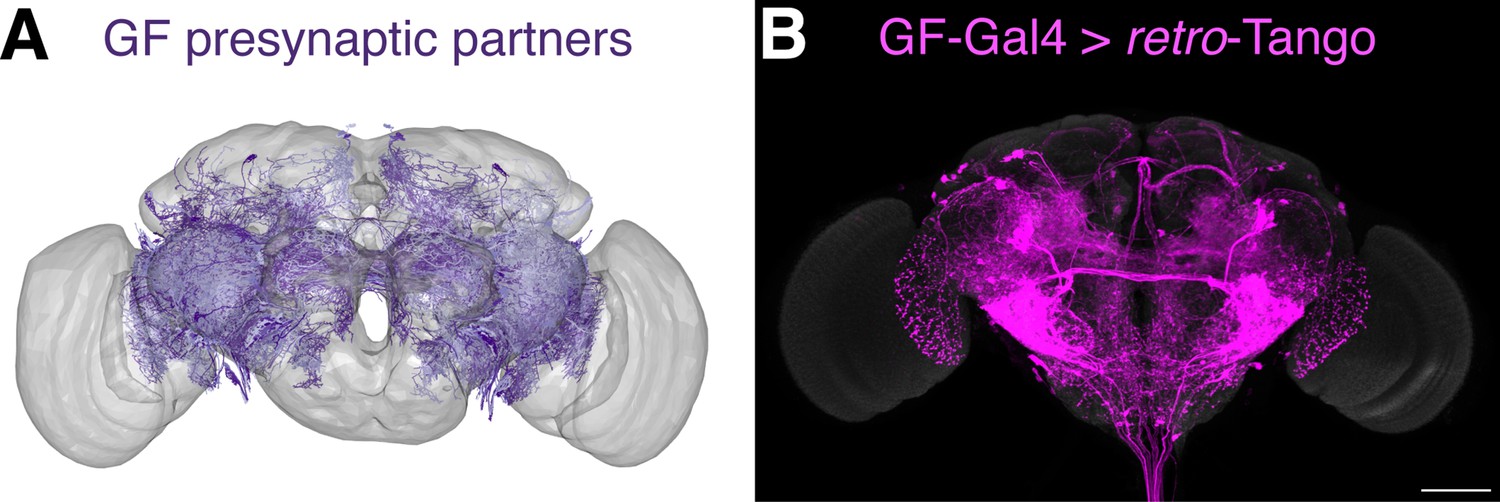
Comparison of the retro-Tango signal with the EM reconstruction of the female hemibrain.
(A) Plotting of the skeletonizations of the EM segmentations of presynaptic partners that connect with the GF via 17 synapses or more. (B) Presynaptic partners of the GFs in a female fly as revealed by retro-Tango. 15do females heterozygous for the tdTomato reporter were analyzed for panel (B). Presynaptic mtdTomato (magenta) and neuropil (grey). Scale bar, 50 μm. Note the high similarity between the patterns in both panels.
Figure 4—figure supplement 1
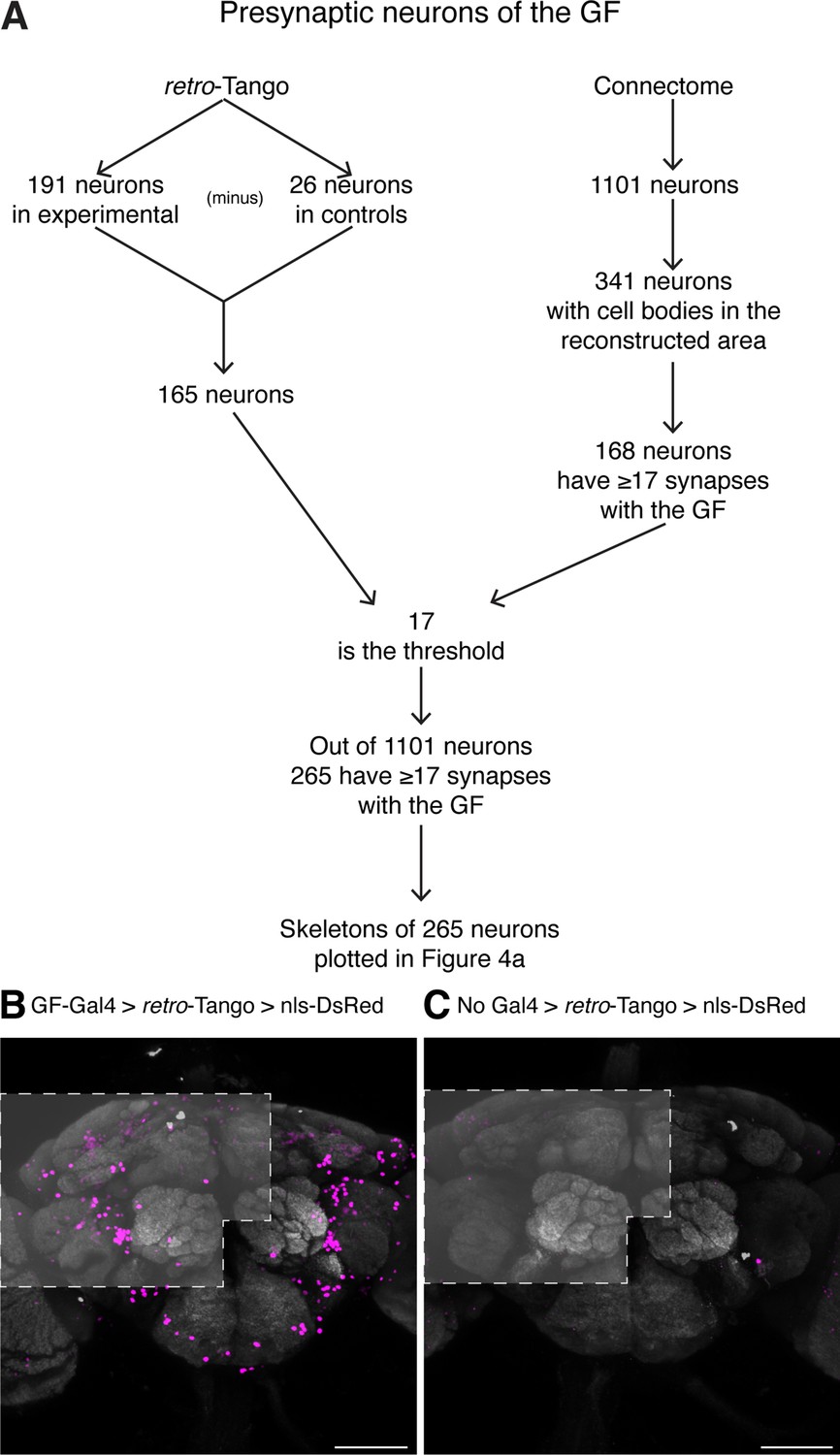
Methodology for the comparison of retro-Tango results with the hemibrain connectome.
(A) Flowchart explaining the steps in the comparison. (B) Driving retro-Tango from the GFs results in nuclear staining in an average of 191 neurons in ten hemibrains. (C) In the absence of a Gal4 driver, retro-Tango has background nuclear staining in 26 neurons. The areas analyzed are marked in light grey based on the approximate regions covered by the published hemibrain connectome. 15do females heterozygous for the nls-DsRed reporter were analyzed for panels (B) and (C). Presynaptic DsRed (magenta) and neuropil (grey). Scale bars, 50 μm.
Figure 4—figure supplement 2
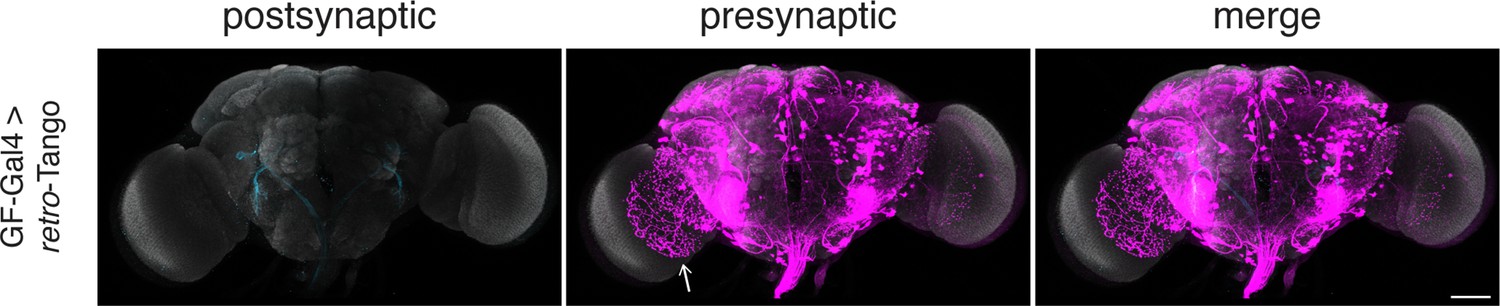
retro-Tango reveals LPLC2s as presynaptic partners of the GF in females when the reporter is homozygous.
Initiating retro-Tango from the GFs in females homozygous for the reporter results in presynaptic signal in LPLC2 (arrow) neurons (157±20 neurons in 5 brains). 15do females homozygous for the tdTomato reporter were analyzed. Presynaptic mtdTomato (magenta) and neuropil (grey). Scale bar, 50 μm.
Figure 5
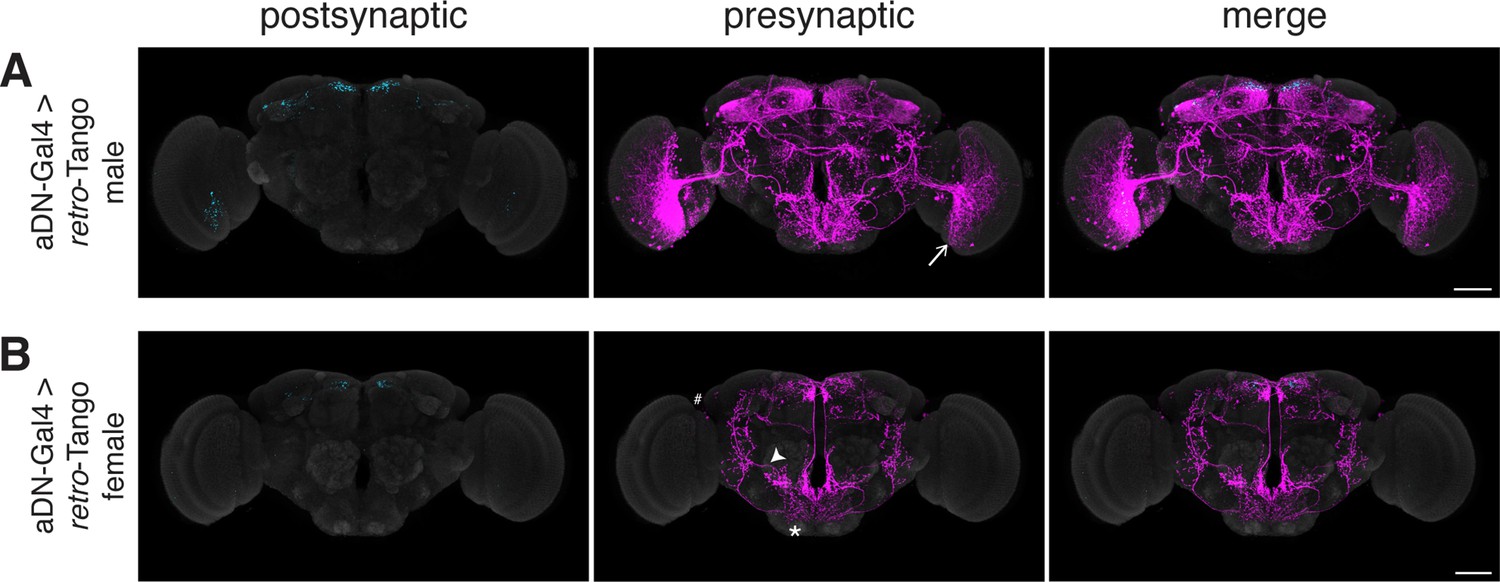
Assessing the specificity of retro-Tango in a sexually dimorphic circuit.
(A) Initiating retro-Tango in aDNs in male flies reveals visual projection neurons (arrow) as presynaptic partners (223±59 neurons in 5 brains). (B) Initiating retro-Tango in aDNs in females results in presynaptic reporter expression in the lateral antennal lobe tract (arrowhead), the SEZ (asterisk), and the LH (hash) (24±11 neurons in 5 brains). 15do males hemizygous for the tdTomato reporter (A) and females heterozygous for the reporter (B) were analyzed. Postsynaptic GFP (cyan), presynaptic mtdTomato (magenta) and neuropil (grey). Scale bars, 50 μm.
Figure 6 with 1 supplement
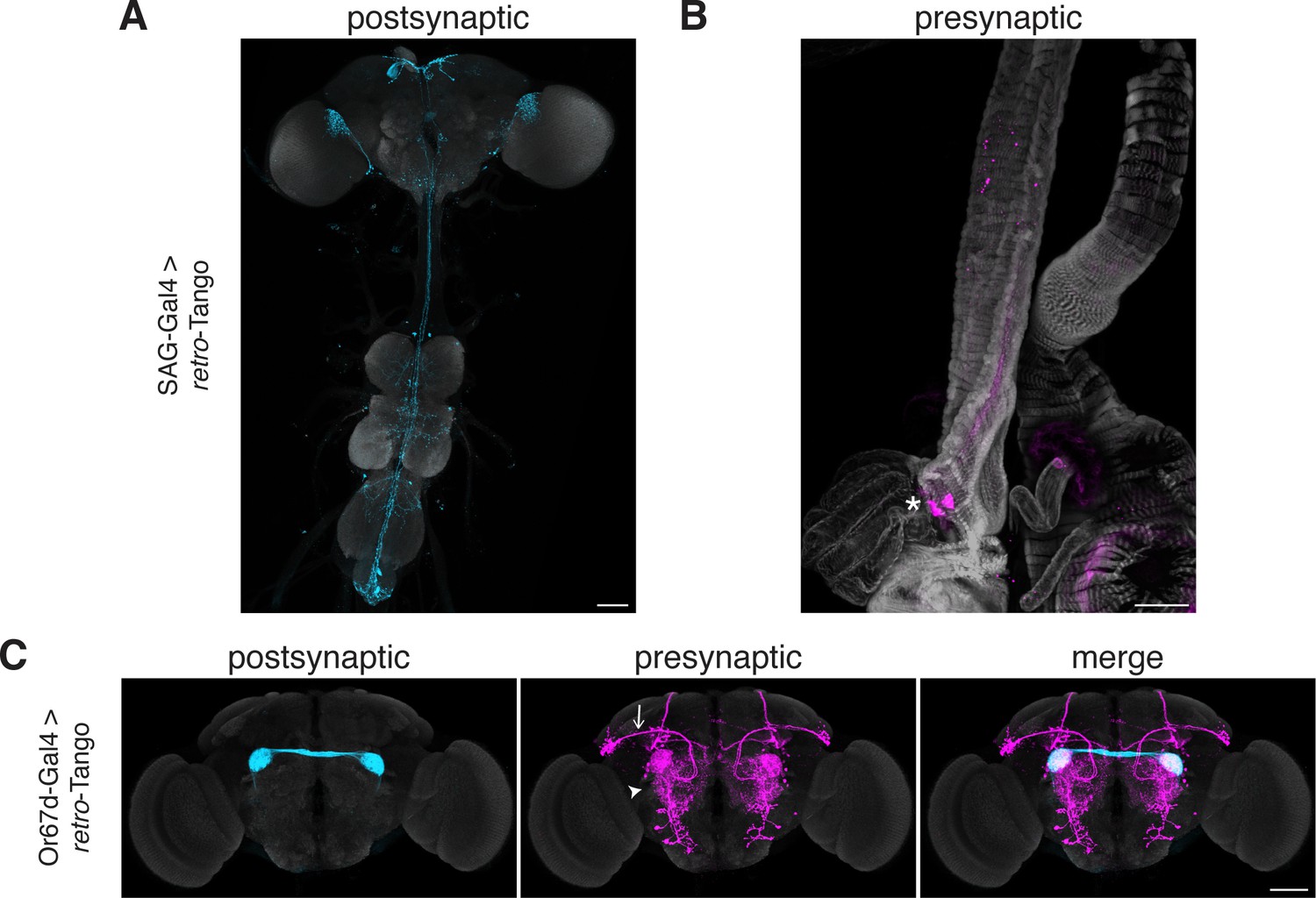
Tracing connections between the periphery and the CNS with retro-Tango.
(A) Expression of the retro-Tango ligand in SAG neurons reveals (B) SPSNs (asterisk) as presynaptic partners. (C) When retro-Tango is initiated from Or67d-expressing ORNs, OPNs (arrow) and LNs (arrowhead) are revealed as their presynaptic partners(134±17 neurons in 5 brains). 15do females heterozygous for the tdTomato reporter (A) and males (B) were analyzed. Postsynaptic GFP (cyan), presynaptic mtdTomato (magenta) and neuropil (A, C), or phalloidin (B) (grey). Scale bars, 50 μm.
Figure 6—figure supplement 1
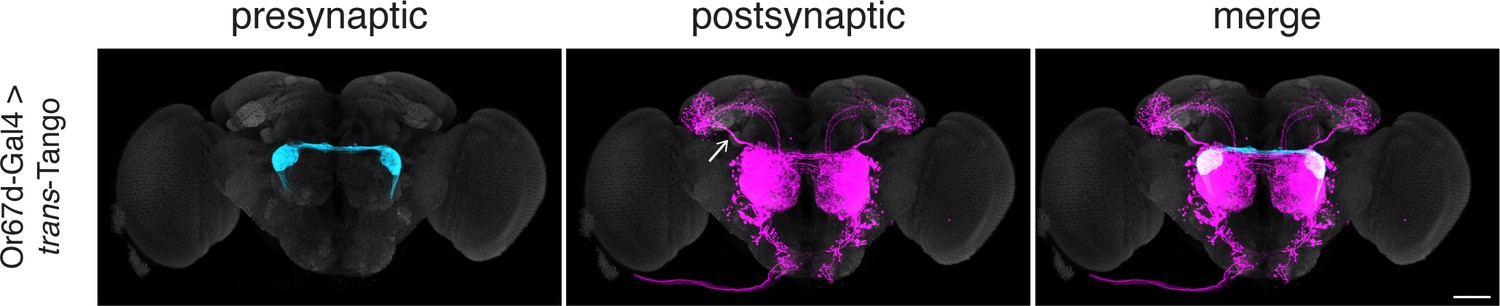
Initiating trans-Tango from the Or67d-expressing ORNs.
Initiating trans-Tango from the Or67d-expressing ORNs results in strong postsynaptic signal in OPNs and LNs (102±17 neurons in 5 brains). Note the labeling in the mediolateral antennal lobe tract (arrow). 20do males were analyzed for both panels. Presynaptic GFP (cyan), postsynaptic mtdTomato (magenta) and neuropil (grey). Scale bars, 50 μm.
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Genetic Reagent (D. melanogaster) | GF-split-Gal4 | von Reyn et al., 2014 | RRID: BDSC#79602 | Flybase symbols: P{R17A04-p65.AD} P{R68A06-GAL4.DBD} |
| Genetic Reagent (D. melanogaster) | Or67dGal4 | Kurtovic et al., 2007 | FlyBase: FBti0168583 | Flybase symbol: TI{GAL4}Or67dGAL4-1 |
| Genetic Reagent (D. melanogaster) | ss00090-Gal4 | Wolff and Rubin, 2018 | RRID: BDSC#75849 | Flybase symbols: P{R15C03-GAL4.DBD} P{R19G02-p65.AD} |
| Genetic Reagent (D. melanogaster) | SAG-split-Gal4 | Feng et al., 2014 | RRID: BDSC#66875 | Flybase symbols: P{VT007068-GAL4.DBD} P{VT050405-p65.AD} |
| Genetic Reagent (D. melanogaster) | aDN-split-Gal4 | Nojima et al., 2021 | FlyBase:FBal0243326 FlyBase: FBal0325783 | Flybase symbols: P{dVP16AD}VGlutOK371-dVP16AD TI{GAL4(DBD)::Zip-}dsxGAL4-DBD |
| Genetic Reagent (D. melanogaster) | QUAS-nls-DsRed | Snell et al., 2022 | RRID: BDSC#95315 | Isolated from BDSC#95315 Flybase symbol: P{5xQUAS-nlsDsRedT4}su(Hw)attP8 |
| Genetic Reagent (D. melanogaster) | QUAS-mtdTomato(3xHA) | This study | Will be deposited to Bloomington Drosophila Stock Center | |
| Genetic Reagent (D. melanogaster) | retro-Tango(panneuronal) | This study | Will be deposited to Bloomington Drosophila Stock Center | |
| Genetic Reagent (D. melanogaster) | retro-Tango(ligand) | This study | Will be deposited to Bloomington Drosophila Stock Center | |
| Genetic Reagent (D. melanogaster) | MB247-Gal4 | Aso et al., 2009 | RRID: BDSC#50742 | Flybase symbol: P{Mef2-GAL4.247} |
| Genetic Reagent (D. melanogaster) | UAS-syt::GFP | Zhang et al., 2002 | RRID: BDSC#6924 | Flybase symbol: P{UAS-syt.eGFP} |
| Genetic Reagent (D. melanogaster) | Reporters +trans-Tango | Talay et al., 2017 | RRID: BDSC#77124 | Flybase symbols: P{trans-Tango} P{UAS-myrGFP.QUAS-mtdTomato-3xHA} |
| Antibody | α-GFP (chicken polyclonal) | Gift from Susan Brenner-Morton (Columbia University) | IHC (1:10000) | |
| Antibody | α-RFP (guinea pig polyclonal) | Gift from Susan Brenner-Morton (Columbia University) | IHC (1:10000) | |
| Antibody | α-Brp (mouse monoclonal) | DSHB | RRID: AB_2314866 | IHC (1:20) |
| Antibody | α-chicken 488 (donkey polyclonal) | Jackson ImmunoResearch # 703-546-155 | RRID: AB_2340376 | IHC (1:1000) |
| Antibody | α-guinea pig 555 (donkey polyclonal) | Jackson ImmunoResearch # 706-165-148 | RRID: AB_2340460 | IHC (1:1000) |
| Antibody | α-mouse 647 (donkey polyclonal) | Thermo Fisher Scientific #A-31571 | RRID: AB_162542 | IHC (1:1000) |
| Chemical compound, drug | Phalloidin 647 | Thermo Fisher Scientific | Catalog number: A22287 | (1:500) |
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
retro-Tango enables versatile retrograde circuit tracing in Drosophila
eLife 12:e85041.
https://doi.org/10.7554/eLife.85041




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}