Lack of CCDC146, a ubiquitous centriole and microtubule-associated protein, leads to non-syndromic male infertility in human and mouse
- Institute for Advanced Biosciences (IAB), INSERM 1209, France
- Institute for Advanced Biosciences (IAB), CNRS UMR 5309, France
- Institute for Advanced Biosciences (IAB), Université Grenoble Alpes, France
- UM GI-DPI, CHU Grenoble Alpes, France
- Cell Biology/ Electron Microscopy, University of Bayreuth, Germany
- Department of Genetics, Reproductive Biomedicine Research Center, Royan Institute for Reproductive Biomedicine, ACECR, Islamic Republic of Iran
- UM de Génétique Chromosomique, Hôpital Couple-Enfant, CHU Grenoble Alpes, France
- Department of Genetic Medicine and Development, University of Geneva Medical School, Switzerland
- University of Geneva, Department of Molecular and Cellular Biology, Sciences III, Switzerland
- Polyclinique les Jasmins, Centre d'Aide Médicale à la Procréation, Centre Urbain Nord, Tunisia
- Laboratoire TIMC/MAGe, CNRS UMR 5525, Pavillon Taillefer, Faculté de Medecine, France
Figures
Figure 1 with 1 supplement
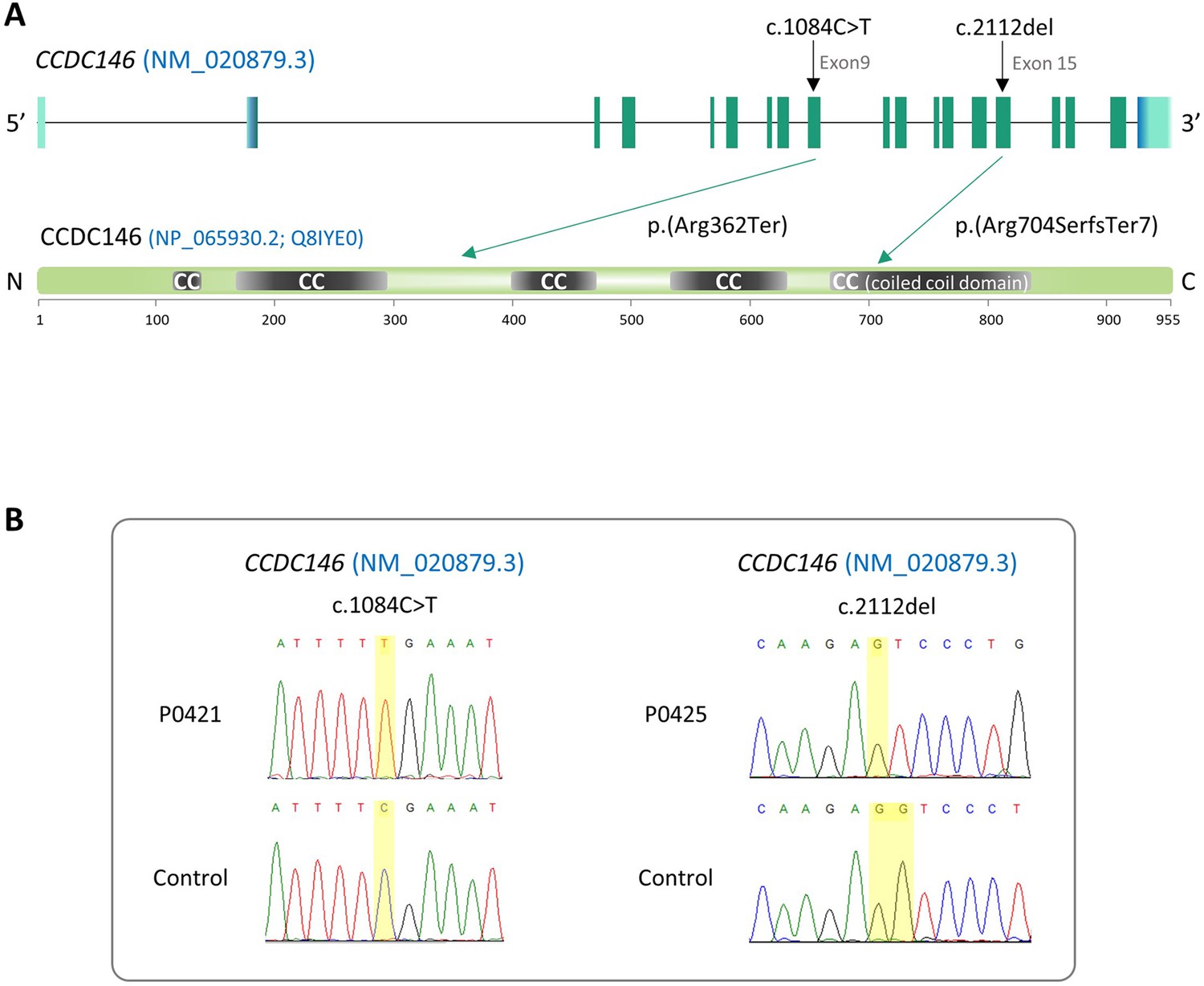
Identification of two CCDC146 gene variants in multiple morphological abnormalities of the flagellum (MMAF) patients.
(A) Structure of the canonical CCDC146 gene transcript showing the position of the variants and their impact on translation. Variants are annotated according to HGVS recommendations. Position of the observed variants in both probands. (B) Electropherograms indicating the homozygous state of the identified variant: variant c.1084C>T is a nonsense mutation, and c.2112Del is a single-nucleotide deletion predicted to induce a translational frameshift.
Figure 1—figure supplement 1
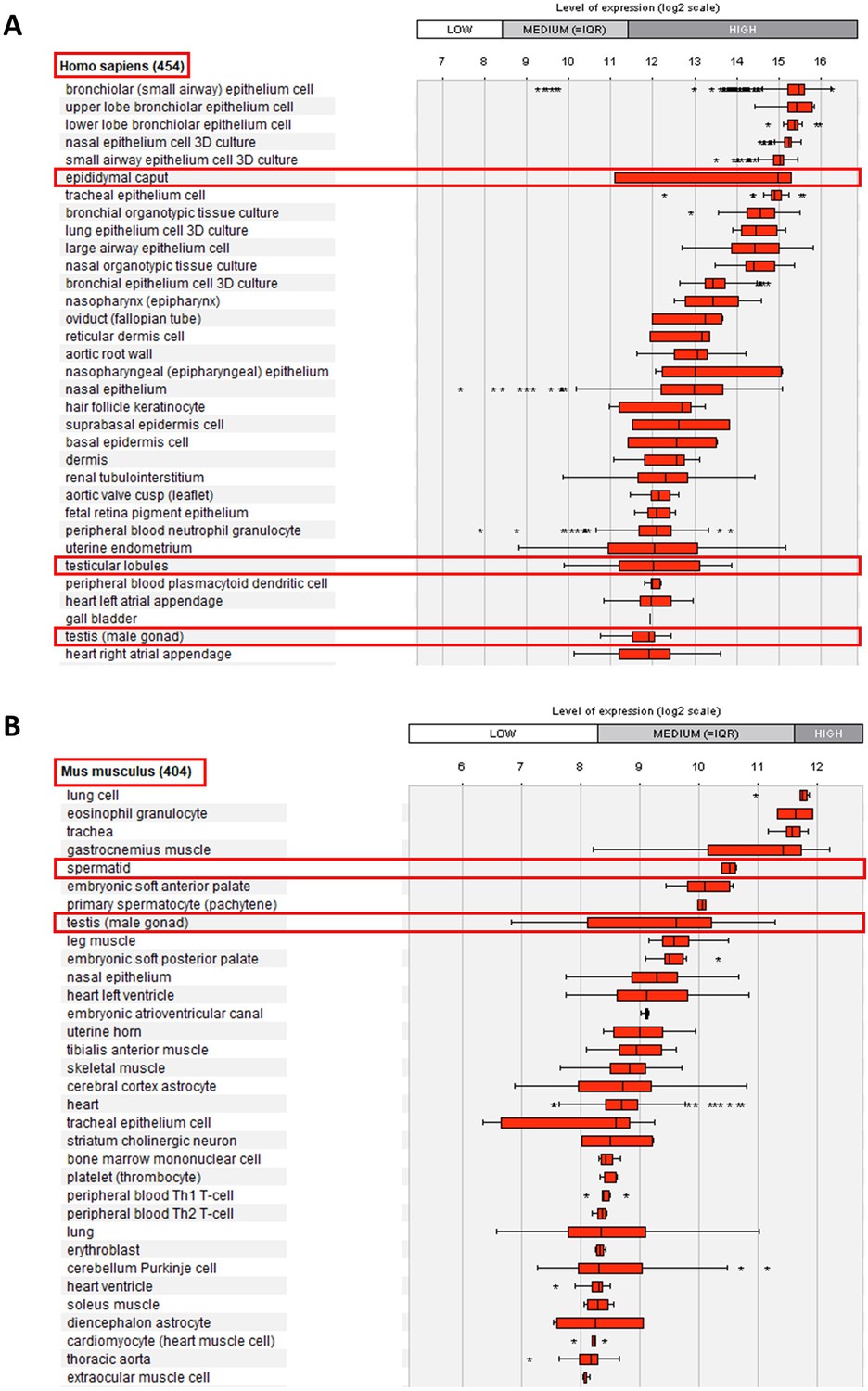
Relative mRNA expression levels for human and mouse Ccdc146 transcripts.
(A) Ccdc146 mRNA levels measured in different tissues/cells in humans using Affymetrix microarrays (data available from the Genevestigator database, https://genevestigator.com). Red rectangles highlight the high expression level in male reproductive organs. (B) Similar data for mice. The level of expression is medium in mice. Data were generated with Genevestigator (Hruz et al., 2008).
Figure 2 with 2 supplements

Ccdc146 KO mice are infertile and KO sperm exhibit a typical multiple morphological abnormalities of the flagellum (MMAF) phenotype.
(A) Number of pups produced by wild-type (+/+, WT), CCDC146 heterozygote (+/-), and Ccdc146 knock-out (-/-, KO) males (three males per genotype) after mating with fertile WT females (two females per male) over a period of 3 mo. (B) Sperm concentration and (C) comparison of testis weights (mg). (D) Illustration of WT and KO sperm morphologies stained with Papanicolaou and observed under optic microscopy. Scale bars of images represent 10 µm. (E) Histograms showing proportions of total, head and flagella morphological anomalies (mean ± SD) for each Ccdc146 genotype (n = 4). (F) sperm mobility. (A–C, F) Statistical comparisons were based on ordinary one-way ANOVA tests and (E) Statistical significance was assessed by applying an unpaired t-test;p-value of 0.05 or lower was considered statistically significant. ****p<0.0001; ***p<0.001, **p<0.01, *p<0.05. Error bars show standard deviation.
Figure 2—figure supplement 1
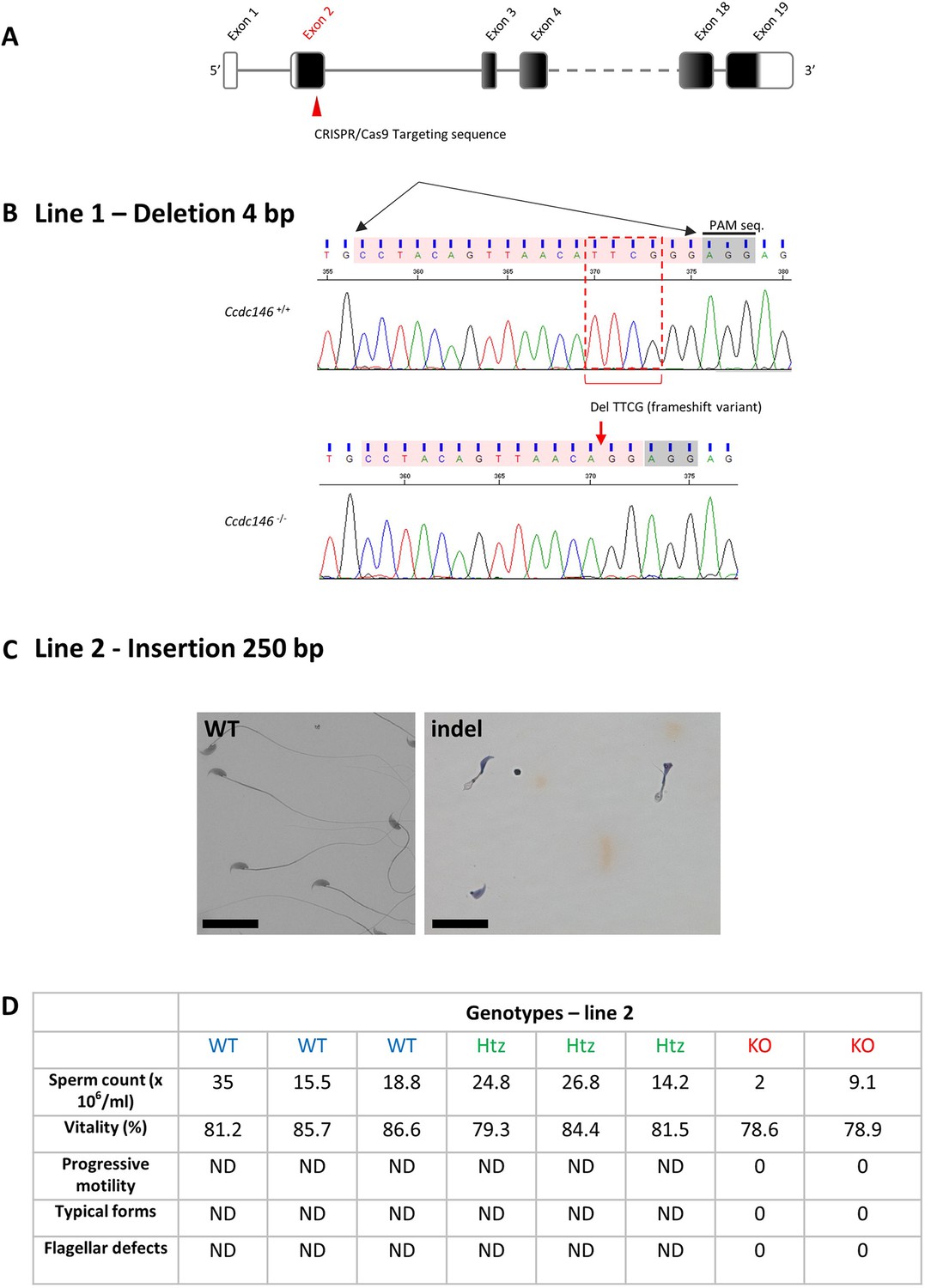
Molecular strategy used to generate Ccdc146 KO mice by CRISPR/Cas9.
(A) The exonic structure of mouse Ccdc146 is shown and the coding sequence indicated in black. (B) Exon 2, the first coding sequence, was targeted by an RNA guide (5’-CCT ACA GTT AAC ATT CGG G-3) and the Cas9 induced in line 1a deletion of four nucleotides upstream the PAM sequence, as indicated by the red box. Electropherogram presenting the WT and the homozygote deletion is shown. (C) In line 2, the Cas9 induced an insertion of 250 nucleotides. Sperm morphology of F2 males from line 2 was severely damaged. Scale bars 30 µm. (D) Comparison of sperm parameters in WT, heterozygote, and KO animals from line 2; ND not determined.
Figure 2—figure supplement 2
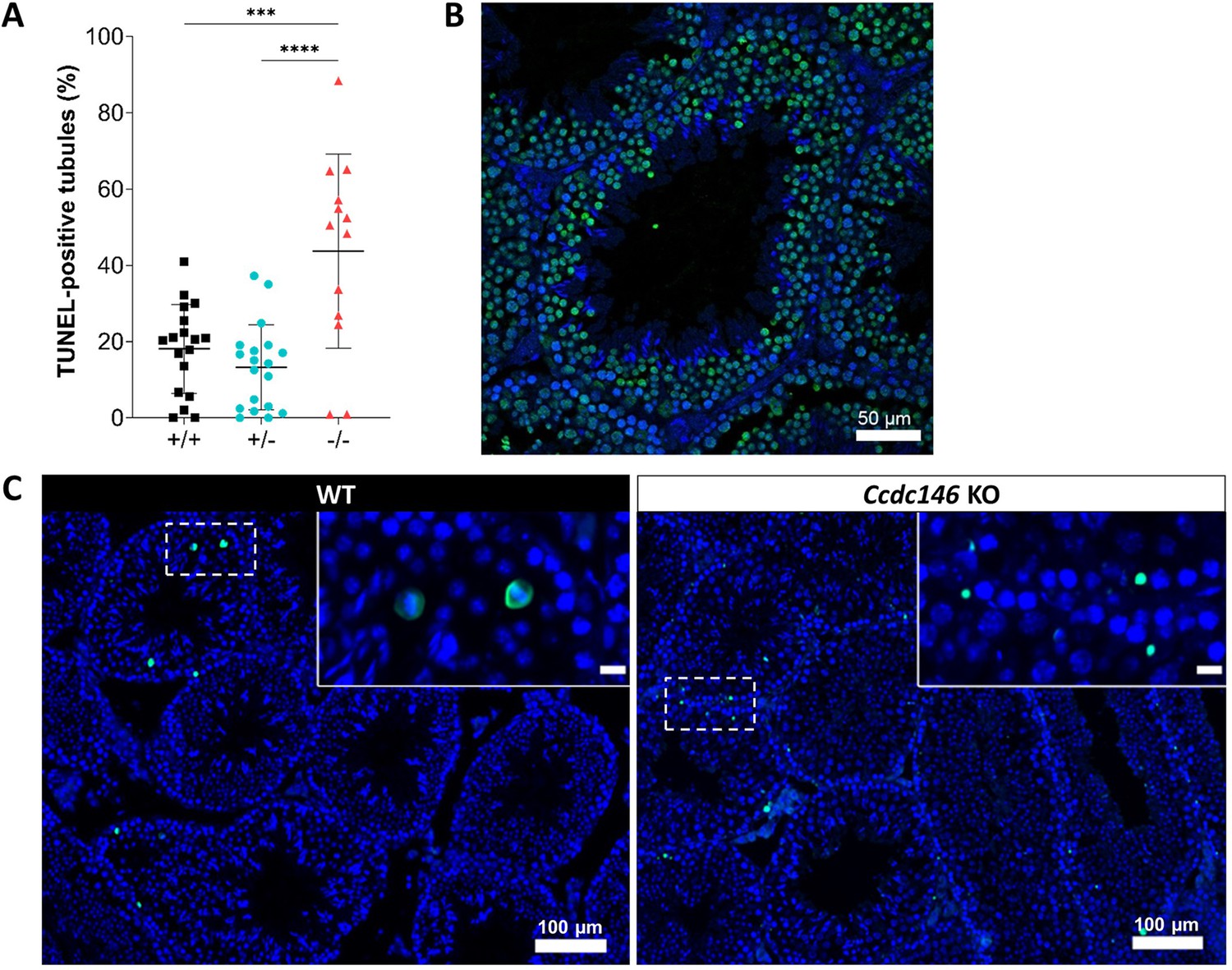
Increased levels of apoptosis in testes from Ccdc146 KO mice.
The TUNEL assay was used to visualize double-strand DNA breaks as an indication of the level of apoptosis during WT and KO spermatogenesis. (A) Comparison of the % of tubules per testis cross-section containing at least one fluorescent (TUNEL-positive) cell in WT, heterozygote, and Ccdc146 KO animals. Number of sections counted per genotype n = 13–20, three different mice per genotype. (B) Control testis section treated with H2O2. (C) The majority of TUNEL-positive cells in the WT corresponded to pachytene cells undergoing meiosis (C, WT zoomed image), whereas the localization of TUNEL-positive cells in KO was more scattered. Statistical comparisons according to ordinary one-way ANOVA test. p-value of 0.05 or lower was considered statistically significant. ****p<0.0001; ***p<0.001, **p<0.01, *p<0.05. Error bars show standard deviation. Scale bars of zoomed images 50 µm.
Figure 3
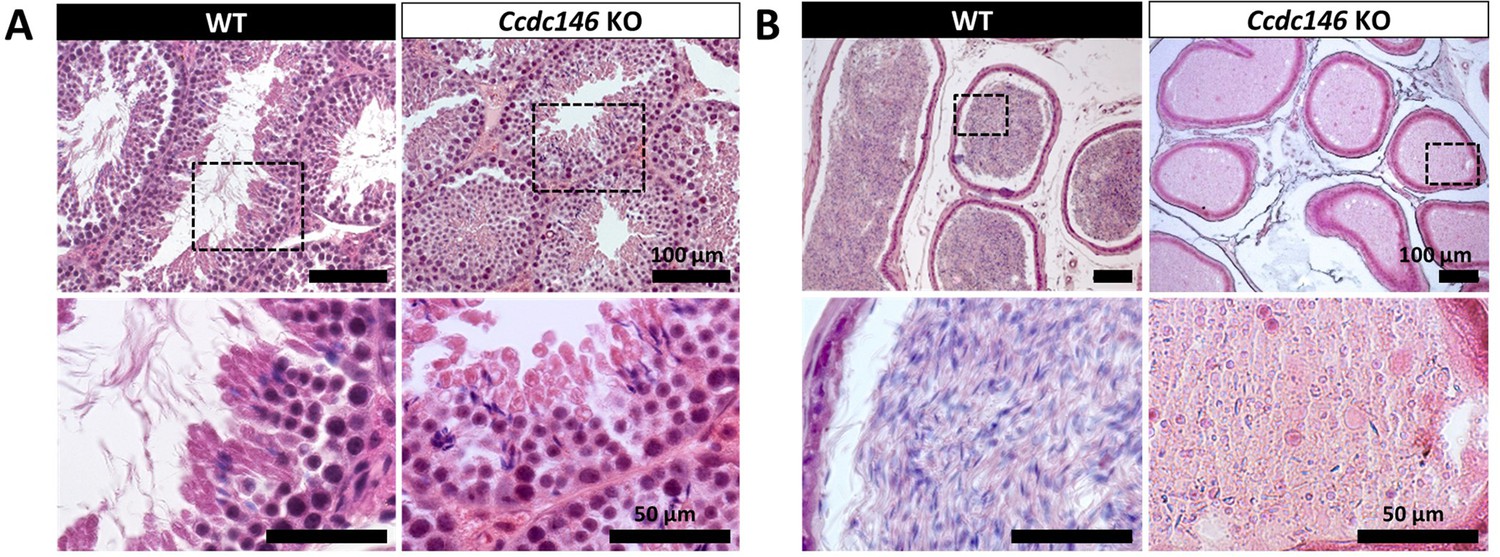
Histological evidence that spermiogenesis is disrupted in Ccdc146 KO males and leads to a strong decrease in sperm concentration in the epididymis.
(A) Transversal sections of WT and KO testes stained with hematoxylin and eosin. The upper images show the sections at low magnification (scale bars 100 µm) and the lower images are an enlargement of the dotted square (scale bars 50 µm). In the KO, spermatid nuclei were very elongated and remarkably thin (green arrow heads) and no flagella were visible within the seminiferous tubule lumen. (B) Transversal sections of WT and KO epididymides stained with hematoxylin and eosin. Despite similar epididymis section diameters in WT and KO testes, KO lumen were filled with round cells and contained few spermatozoa with abnormally shaped heads and flagella. The upper images show the sections at low magnification (scale bars 100 µm) and the lower images are an enlargement of the dotted square (scale bars 50 µm).
Figure 4 with 2 supplements
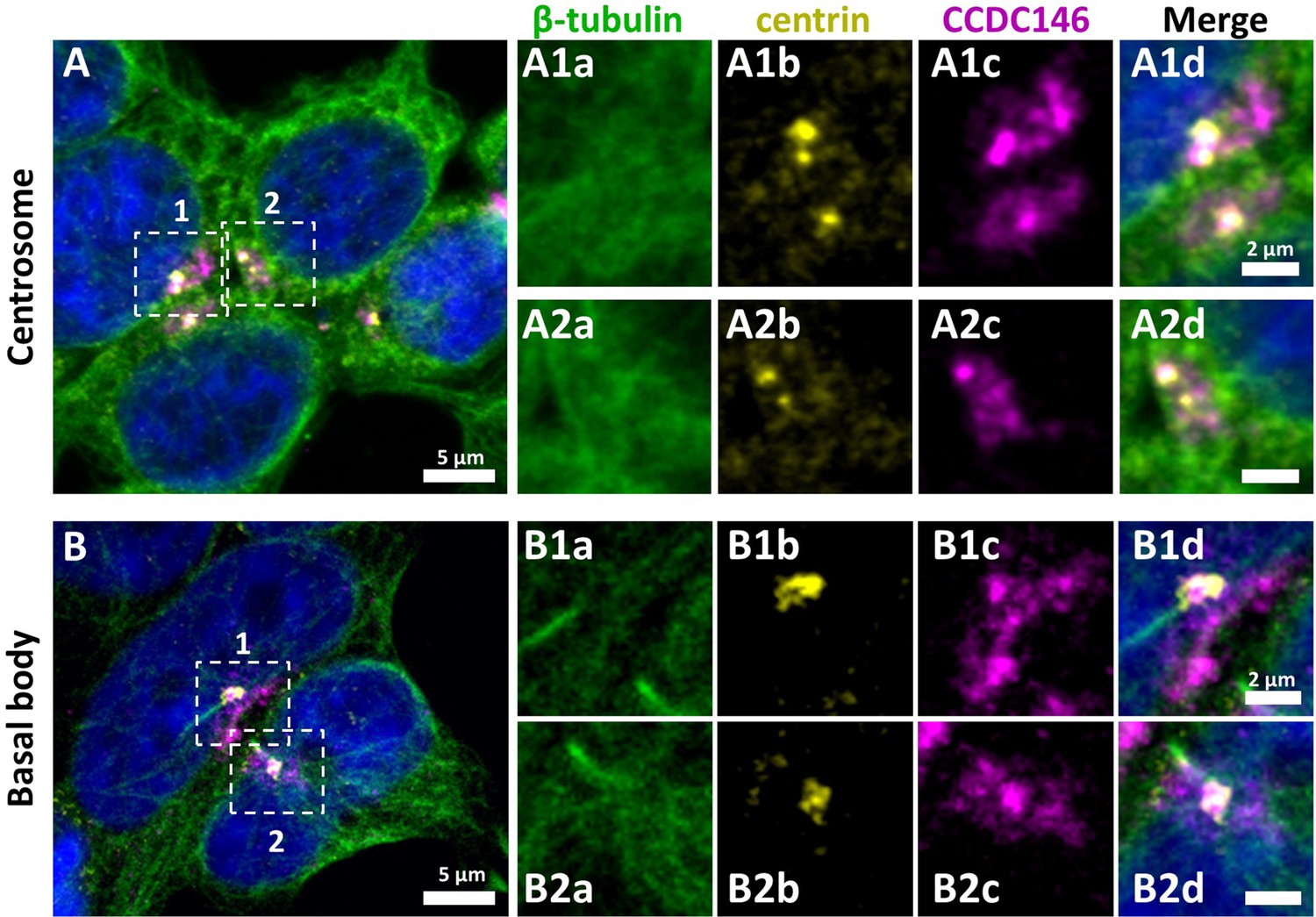
CCDC146 has a centriolar and pericentriolar localization in interphase somatic HEK-293T cells.
HEK-293T cells were immunolabeled for β-tubulin (green), centrin (yellow), and CCDC146 (magenta). DNA was stained with Hoechst (blue). (A) CCDC146 localized to centrioles and/or to the pericentriolar material in interphase cells. The centrosome area of two cells is shown enlarged in (A1) and (A2). (B) In serum-starved cells with primary cilia, CCDC146 localized to the basal body of primary cilia. The basal body of two cells is shown enlarged in (B1) and (B2). CCDC146 was also present as dotted signal resembling the pattern for centriolar satellite proteins. Scale bars 5 µm and 2 µm on zoomed images.
Figure 4—figure supplement 1
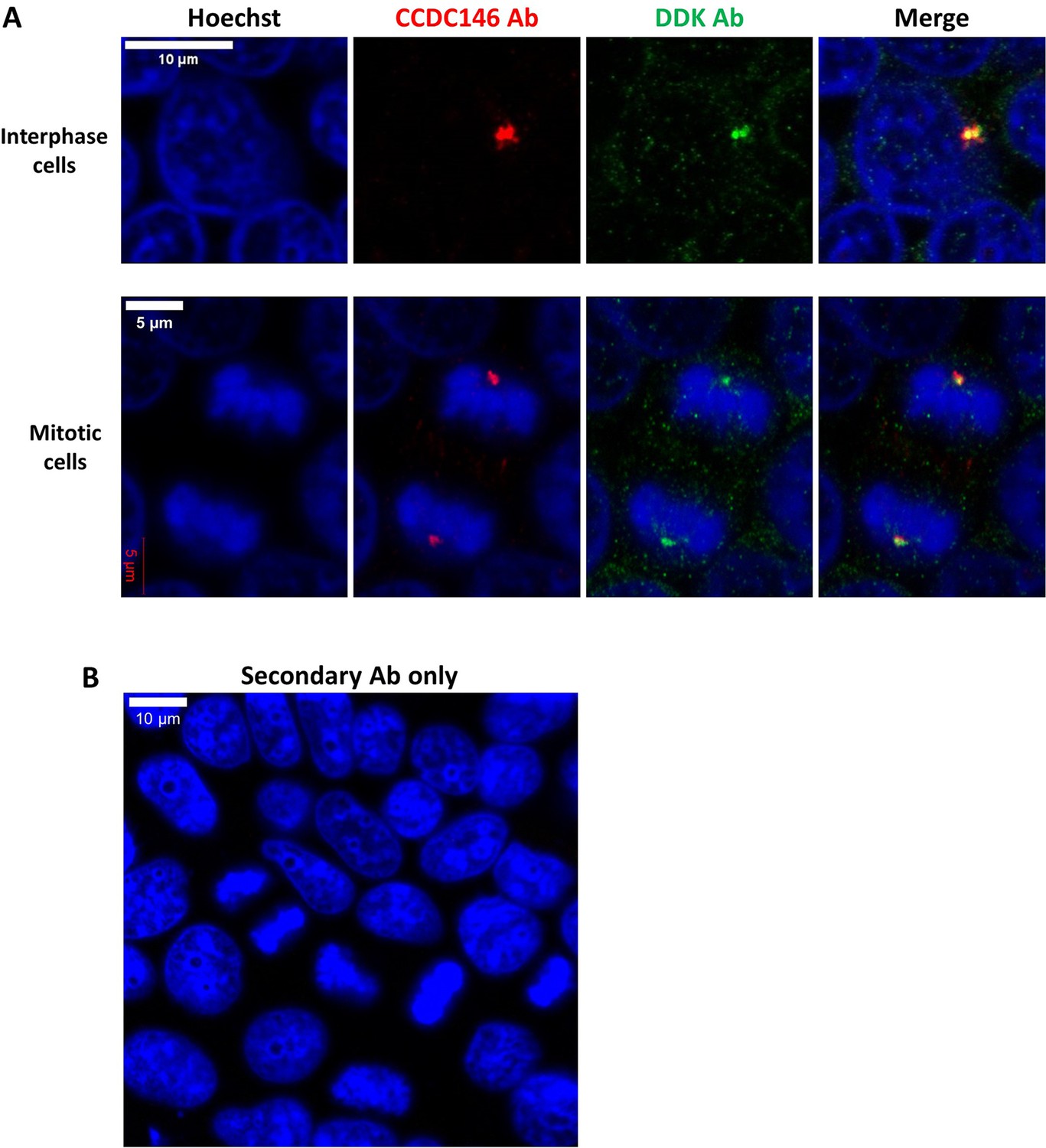
DDK and CCDC146 Abs immunodecorate the same cellular components.
HEK-293T cells were transfected with a plasmid containing the sequence of DDK-CCDC146 (human). After cell fixation, cells were double immunolabeled for CCDC146 (red) and DDK (green) with the corresponding Abs. (A) Immunostaining of interphase and mitotic cells. Both antibodies stain the centrosome. (B) Same experiment with secondary Ab only. Scale bars as indicated.
Figure 4—figure supplement 2
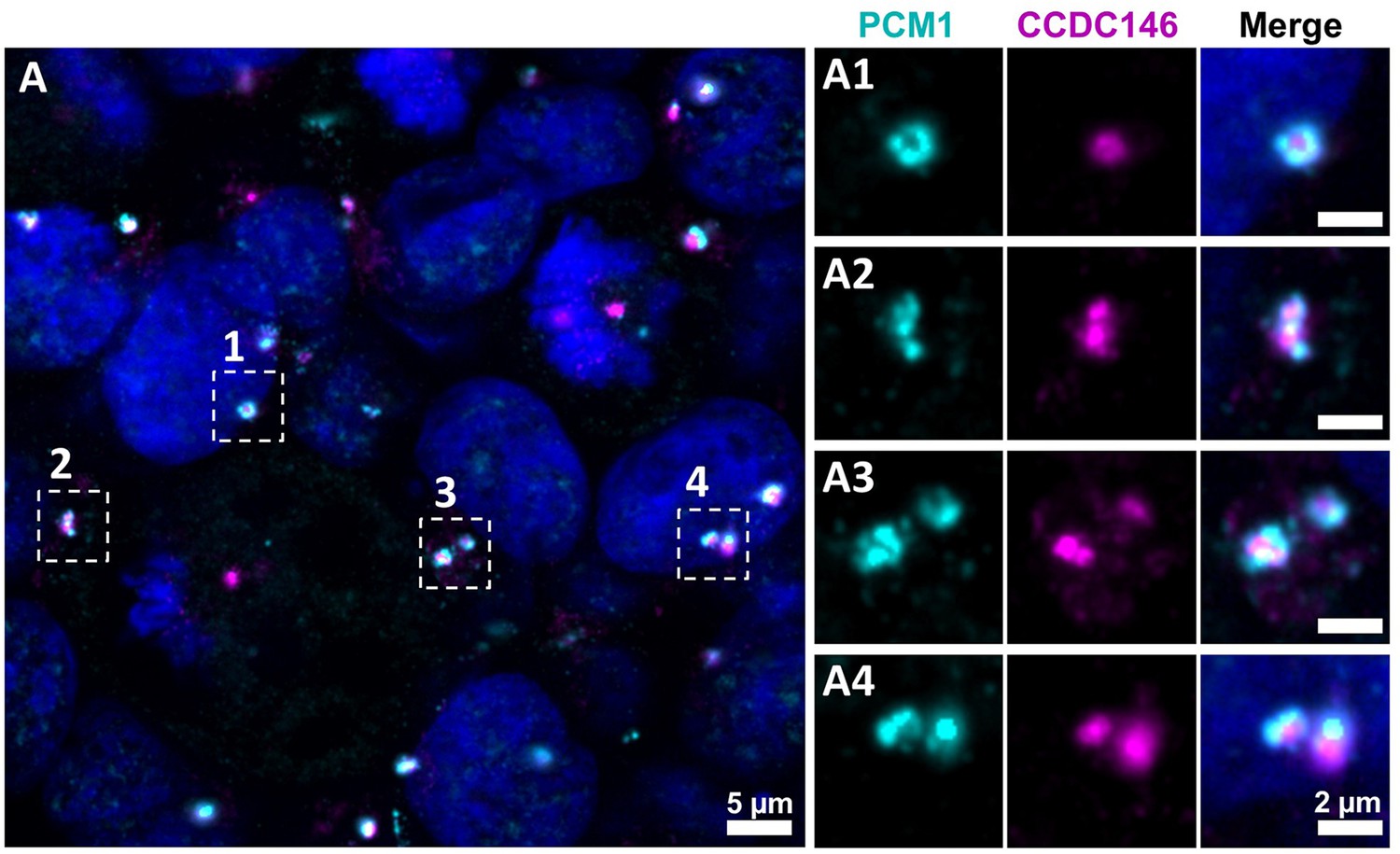
CCDC146 does not colocalize with the centriolar satellite marker PCM1.
(A) HEK-293T cells were double immunolabeled for PCM1 (cyan) and CCDC146 (magenta). (A1–A4) Images on the right show the enlargement of the dotted squares in the left image. PCM1 surrounds the CCDC146 signal, but no colocalization is observed, suggesting that CCDC146 is not a centriolar satellite protein. DNA was stained with Hoechst (blue). Scale bars as indicated and on zoomed images 2 µm.
Figure 5 with 1 supplement
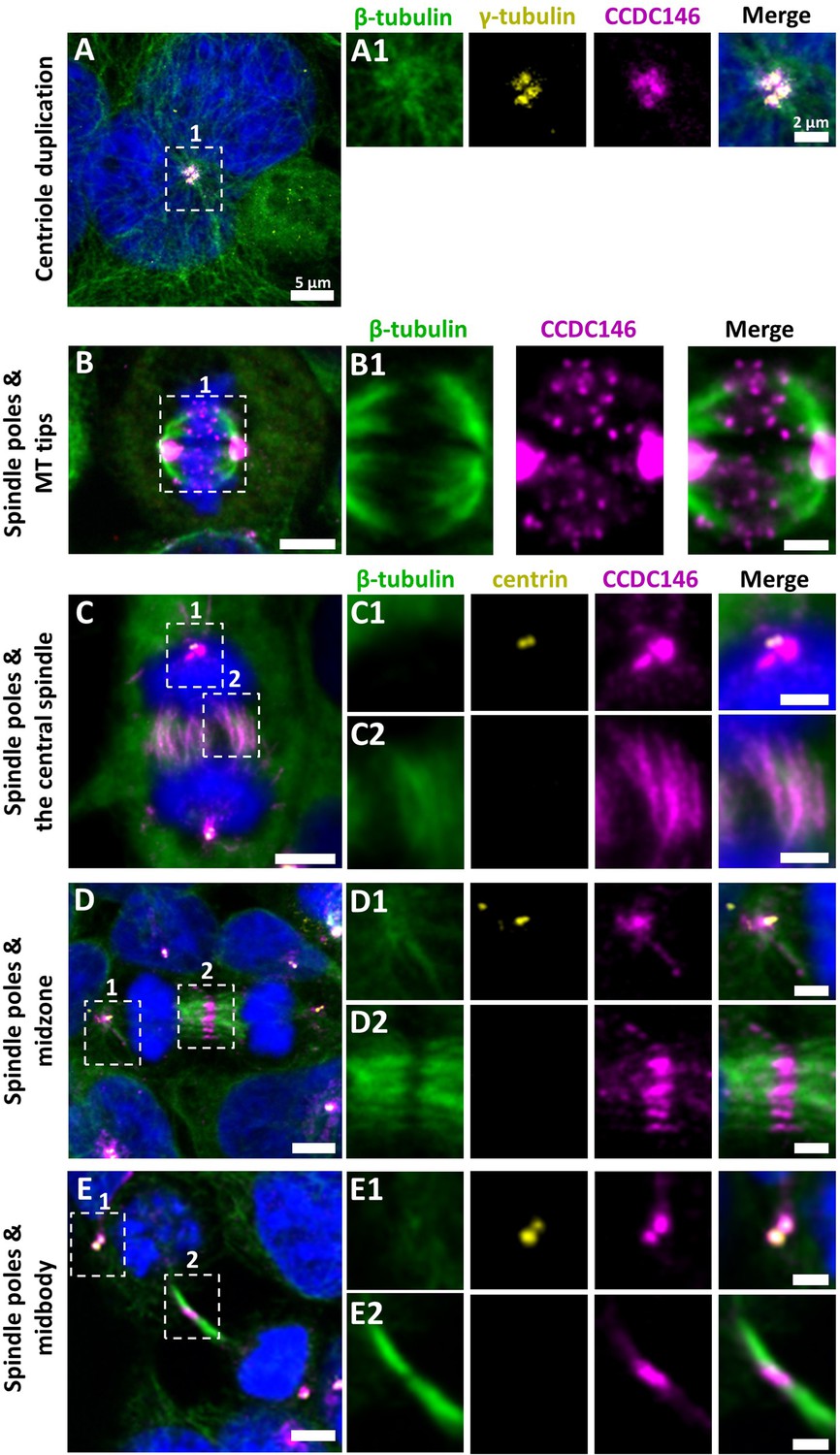
CCDC146 is a microtubule-associated protein (MAP) associating with microtubule-based structures throughout the cell cycle.
HEK-293T cells were immunolabeled with anti-β-tubulin (panels A–E, green), anti-γ tubulin (yellow, panel A), anti-centrin (yellow panels C–E), and anti-CCDC146 (panels A–E, magenta) Abs. DNA was stained with Hoechst (blue). (A) In synchronized HEK-293T cells, CCDC146 is observed associated with mother centrioles and their corresponding procentrioles during centriole duplication. (B–E) In non-synchronized cells, CCDC146 is observed associated with spindle poles (B–E) and with microtubule (MT) tips during metaphase (B), with the central spindle during anaphase (C), with the midzone during telophase (D), and with the midbody during cytokinesis (E). Images on the right show the enlargement of the dotted square in the left image. Scale bars 5 µm and scale bars of zoomed insets 2 µm.
Figure 5—figure supplement 1
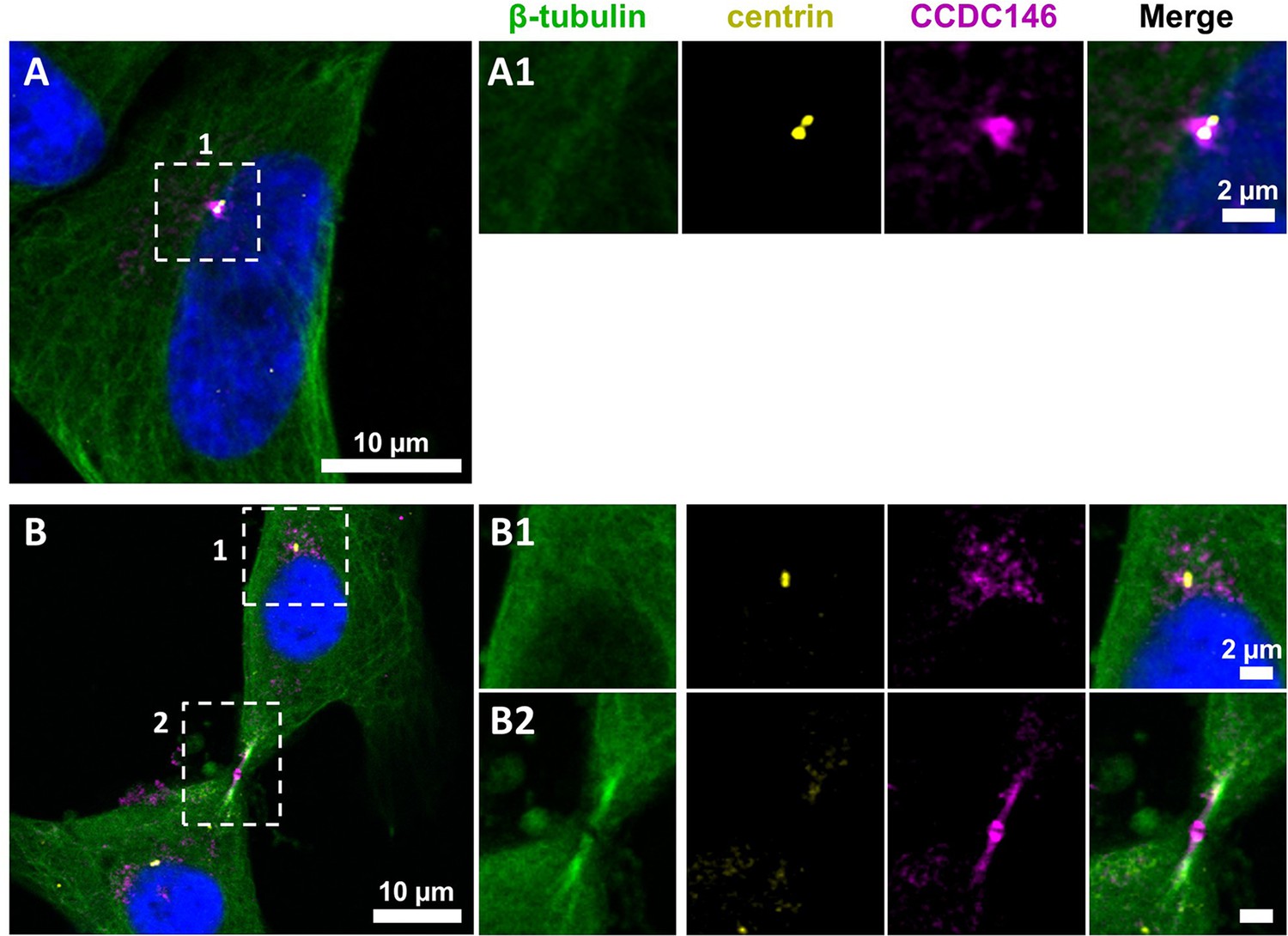
CCDC146 shows a similar localization to the centrosome and to the midbody in primary human foreskin fibroblast (HFF) cells.
(A) Primary HFF cells were triple immunolabeled with anti-β-tubulin (green), anti-centrin (yellow, showing the centrioles), and anti-CCDC146 (magenta). (A1) The right-hand images show the enlargement of the dotted squares in the left-hand image. CCDC146 localized to and around the centrioles. (B) Staining of the centrosome area (B1) and (B2) shows co-localization of the CCDC146 staining with the midbody during cytokinesis. Scale bars 10 µm and scale bars of zoomed insets 2 µm.
Figure 6 with 1 supplement
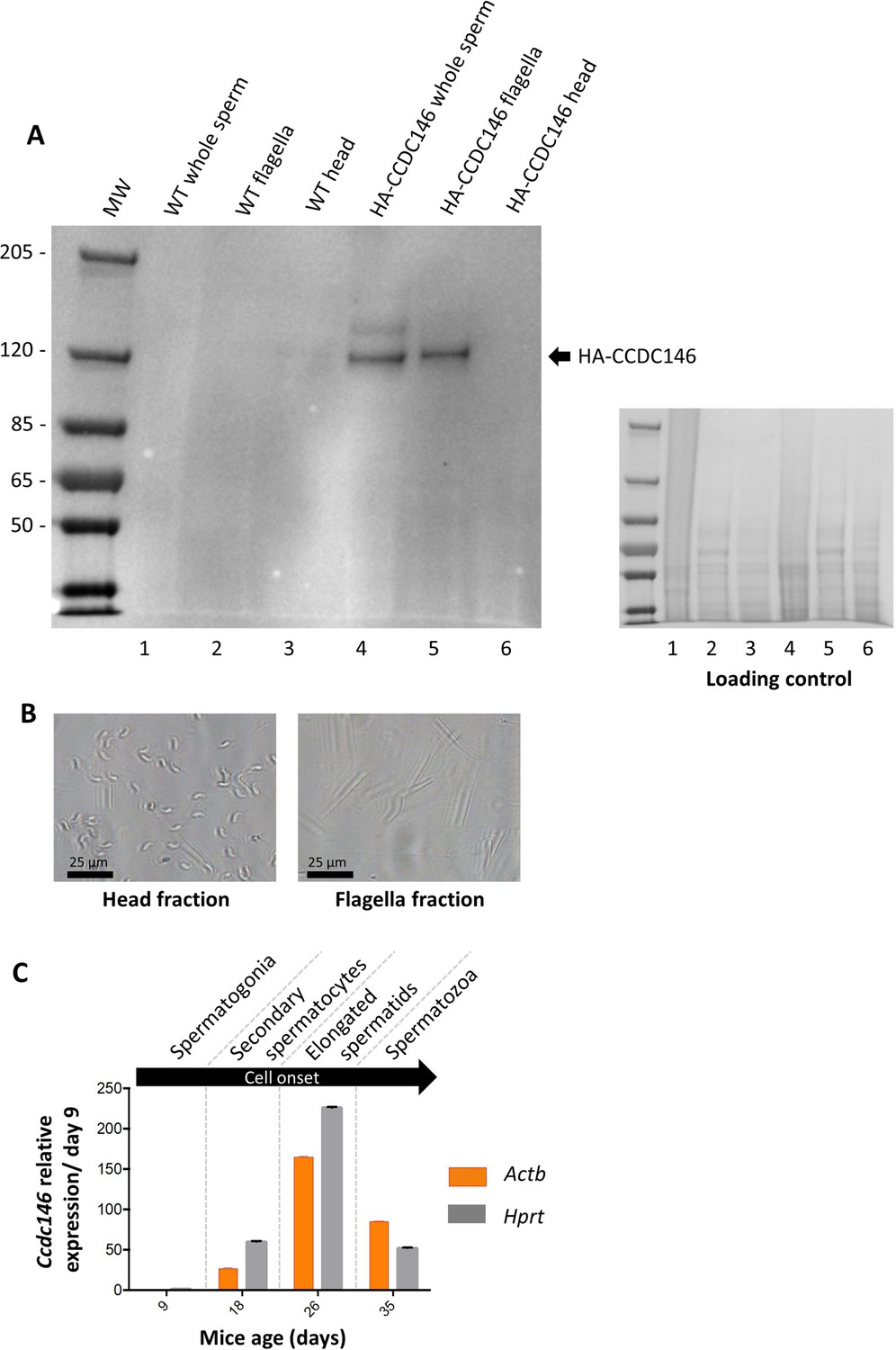
CCDC146 protein is present in epididymal spermatozoa in mouse and Ccdc146 mRNA is expressed in late spermatocyte and in spermatids.
(A) CCDC146 is retained in epididymal sperm. Western blot of HA-tagged epididymal whole sperm shows a band (arrow) corresponding to HA-tagged CCDC146 (HA-CCDC146), whereas no band was observed with WT epididymal sperm, demonstrating the specificity of the band observed in HA-CCDC146 sperm lane. Sperm extracts obtained from flagella and head fractions of WT and HA-CCDC146 were also analyzed and the presence of HA-CCDC146 was revealed by an anti-HA Ab in flagella fraction from HA-CCDC146 sperm only. Loading control (ponceau staining) of the gel is shown on the right. (B) Image of heads and flagella fractions. Scale bars as indicated (C) mRNA expression levels of Ccdc146 relative to Actb and Hprt in HA-CCDC146 mouse pups’ testes during the first wave of spermatogenesis (n = 3). Extremely low Ccdc146 expression was detected at day 9, corresponding to testes containing spermatogonia and Sertoli cells only. Ccdc146 expression was observed from postnatal day 18 (formation of secondary spermatocytes), peaked at day 26 (formation of elongated spermatids), and subsequently decreased from day 35 (formation of spermatozoa), suggesting that Ccdc146 is particularly expressed in elongated spermatids during spermatogenesis. One-way ANOVA test was used for statistical comparisons. p-value of 0.05 or lower was considered statistically significant. ****p<0.0001; ***p<0.001, **p<0.01, *p<0.05. Error bars show standard deviation.
-
Figure 6—source data 1
Uncropped gel of Figure 6A: whole sperm, flagella fraction and head fraction.
- https://cdn.elifesciences.org/articles/86845/elife-86845-fig6-data1-v1.zip
-
Figure 6—source data 2
Uncropped gel of Figure 6A: loading control.
- https://cdn.elifesciences.org/articles/86845/elife-86845-fig6-data2-v1.zip
Figure 6—figure supplement 1
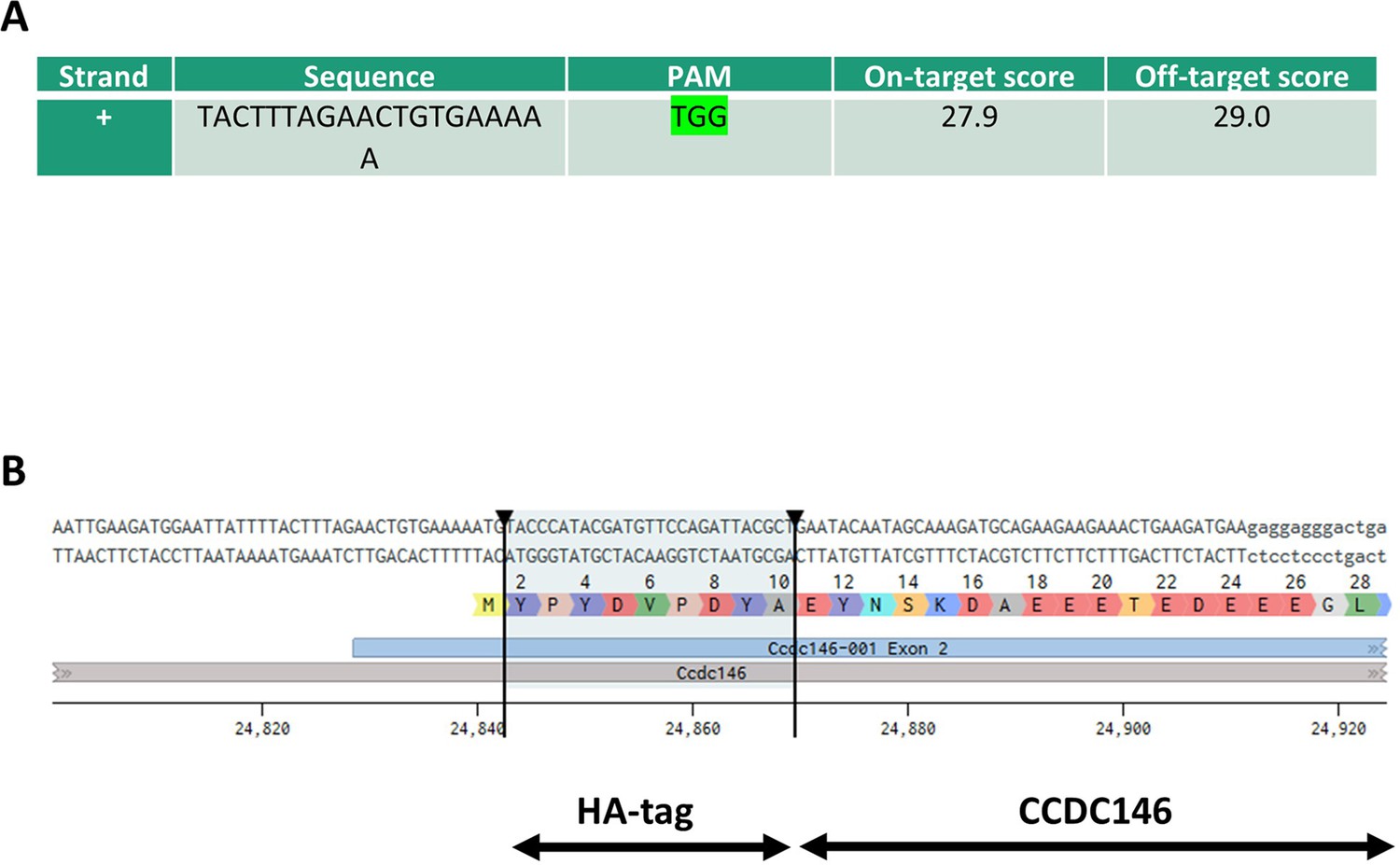
Molecular strategy used to generate HA-tagged CCDC146 mice by CRISPR/Cas9.
(A) Exon 2, the first coding sequence, was targeted by an RNA guide (5’-TAC TTT AGA ACT GTG AAA AAT GG-3’). (B) Using a single-stranded DNA (ssDNA) template, the HA sequence (5’-TAC CCA TAC GAT GTT CCA GAT TAC GCT-3’) was inserted upstream of the PAM sequence.
Figure 7 with 1 supplement
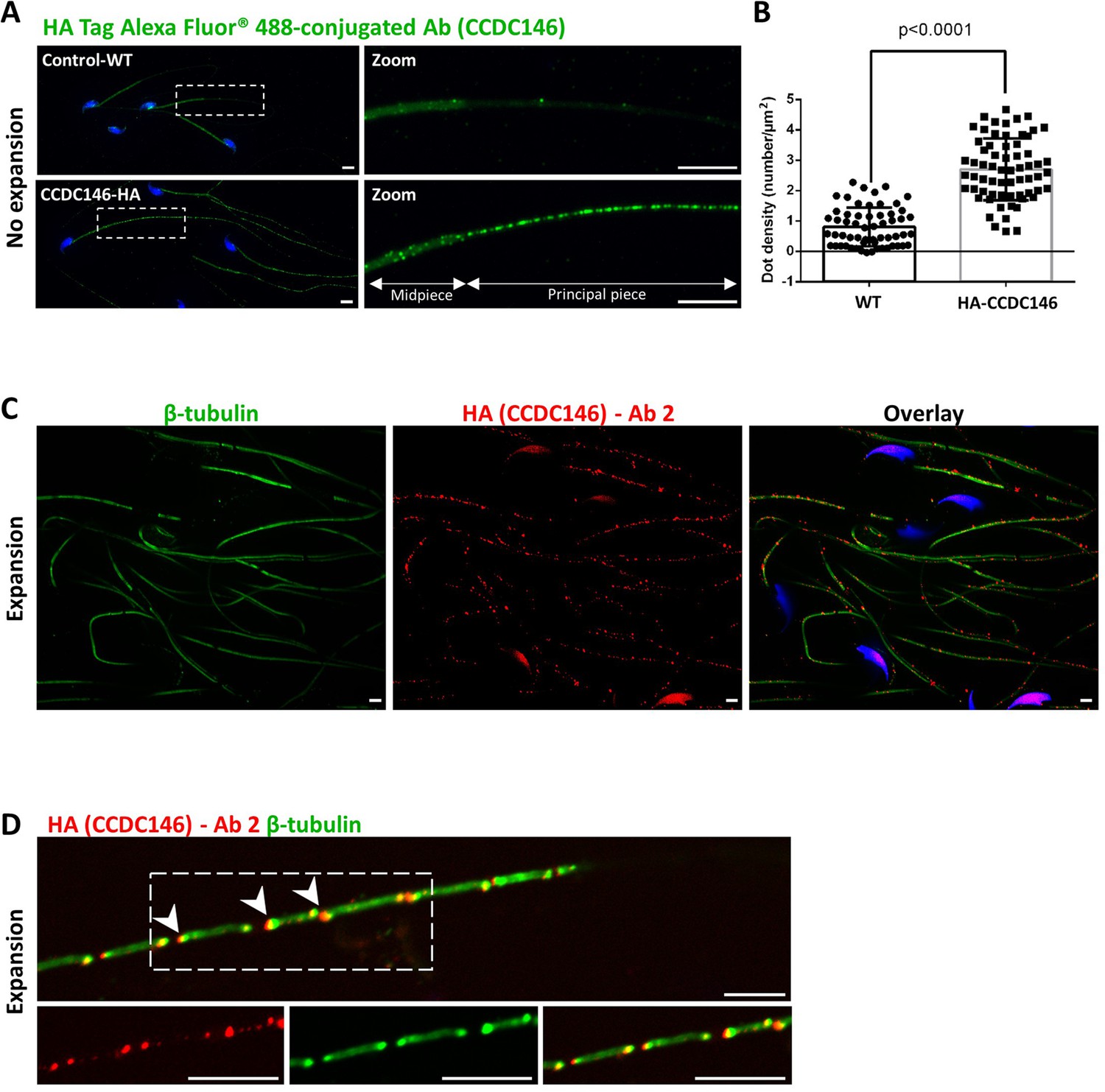
CCDC146 localizes to the flagellum of mouse epididymal spermatozoa.
(A) Mouse epididymal spermatozoa observed with conventional IF. WT and HA-CCDC146 sperm were labeled with anti-HA Tag Alexa Fluor 488-conjugated 3 (green) Abs (anti-HA #1). DNA was stained with Hoechst (blue). The upper image shows the staining of a WT sperm and the lower images, the staining of a HA-CCDC146 sperm. A punctuate signal is observed in HA-CCDC146 sperm (scale bars 5 µm). (B) Quantification of the density of dots per square µm (µm2) and statistical significance between WT and HA-CCDC146 sperm was assessed by Mann–Whitney test (n=58 for WT and 65 for HA_CCDC146), p-value as indicated.p-value of 0.05 or lower was considered statistically significant. Error bars show standard deviation. (C) Mouse epididymal spermatozoa observed with expansion microscopy. HA-CCDC146 sperm were immunolabeled with anti-β-tubulin (red) and anti-HA #2 (green) Abs, and DNA was stained with Hoechst (blue). The right image shows the sperm with merged immunostaining. Scale bars 5 µm. (D) Mouse epididymal spermatozoa observed with expansion microscopy. The lower images show the staining, HA-CCDC146 (red), tubulin (green), and merge observed in the principal piece of the flagellum. Strong red punctiform signals were observed at the level of axonemal breakages induced by the expansion process. White arrows indicate the zones of the micro breaks. Scale bars 10 μm.
Figure 7—figure supplement 1
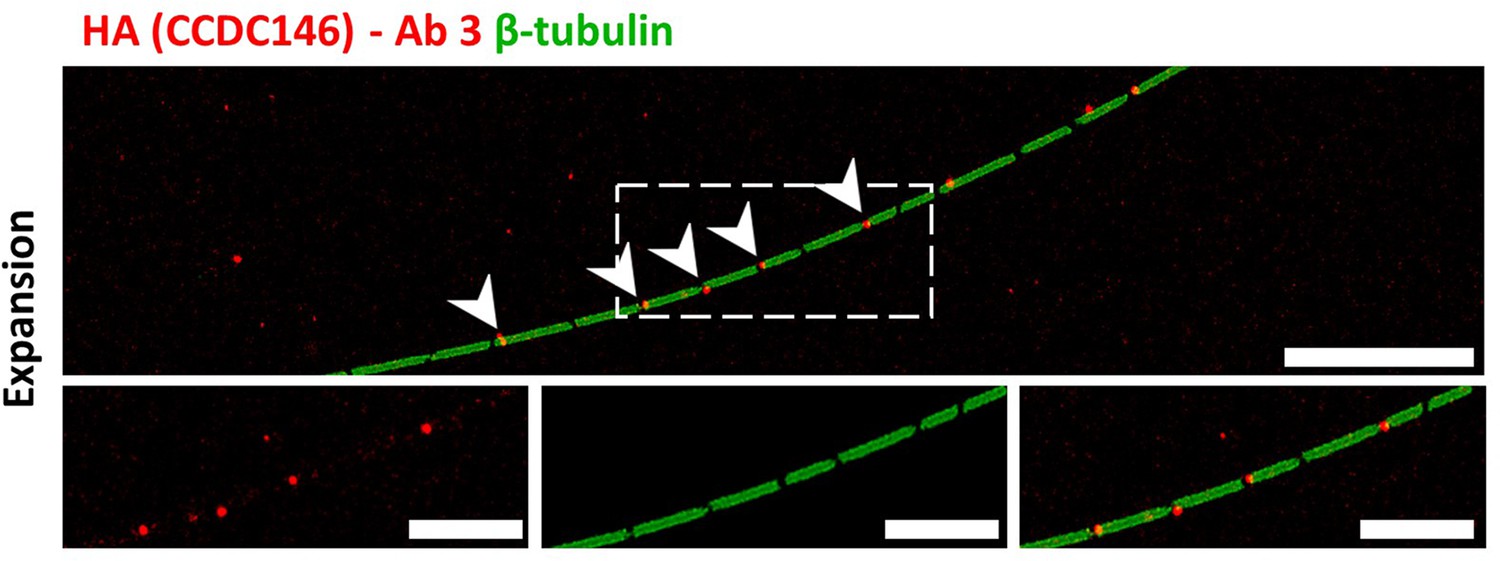
Staining by anti-HA Abs of axonemal breaks induced by expansion.
Mouse epididymal spermatozoa observed with expansion microscopy. HA-CCDC146 sperm were immunolabeled with anti-β-tubulin (green) and anti-HA #3 (red) Abs. The right image shows the sperm with merged immunostaining. Expansion induced axonemal breaks associated with strong HA-CCDC146 staining (red). Scale bars correspond to 10 µm.
Figure 8 with 1 supplement

CCDC146 localizes to the flagellum but not to the centrioles of ejaculated human spermatozoa.
(A) Human ejaculated sperm were immunolabeled with Abs recognizing centrin (yellow), CCDC146 (magenta), and β-tubulin (green). DNA was stained with Hoechst (blue). (A1–A3) Enlargement of dotted square focused on sperm neck: no colocalization between CCDC146 and centrin. (A4) A faint signal for CCDC146 is present along the length of the sperm flagellum. Scale bar of zoomed images: 1 µm. (B) Human ejaculated sperm observed by expansion microscopy. Sperm were immunolabeled with anti-CCDC146 (magenta) and anti-β-tubulin (green) abs, and DNA was stained with Hoechst (blue). (B1 and B5) show strong staining for CCDC146 in the axoneme, (B3) shows the localization of the axoneme through tubulin staining. (B2, B4, and B6) show enlargements of the dotted squares focused on the sperm neck. CCDC146 did not colocalize with the centrioles at the base of the axoneme. The CCDC146 staining observed probably corresponds to non-specific labeling of mitochondria, as suggested by Figure 8—figure supplement 1. Scale bars 5 µm.
Figure 8—figure supplement 1
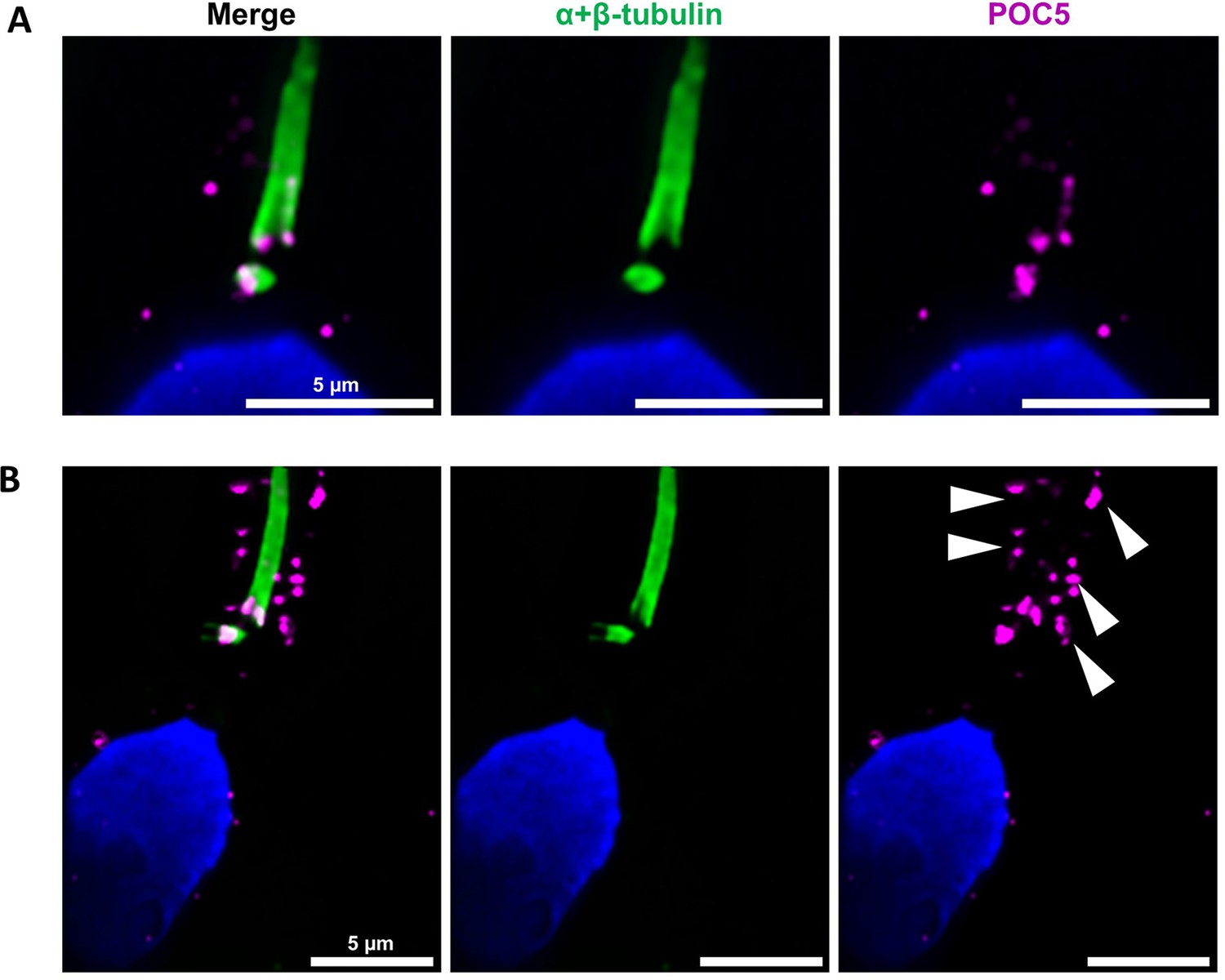
Centrioles are identified by anti-POC5 Abs in expanded human ejaculated spermatozoa.
Human control sperm were co-stained, after expansion, with anti- α+β-tubulin (green) and anti-POC5 (magenta) Abs. (A) The centrosomal protein POC5 locates to centrioles at the base of the axoneme. (B) Apart the centriole staining, a scattered staining was also observed in the midpiece around the axoneme (white arrows heads). Scale bars 5 μm.
Figure 9

CCDC146 localizes to the microtubule doublets of the axoneme in human and mouse.
(A) Sperm was double-stained with anti-tubulin (green) and anti-CCDC146 (magenta) Abs and observed by expansion microscopy. Scale bar 10 µm. (B) Measurement of the green-tubulin signal intensity perpendicularly to the axoneme is quite characteristic with two peaks corresponding to the left and right microtubule doublet. It identifies five axonemal compartments (left outer, left doublet [Tub1], central pair [CP], right doublet [Tub2], and right outer). To determine the CP area, the distance (d) between the center of the flagella and the peak of Tub2 was measured, and all fluorescent dots located in between -d/4 to +d/4 from the center were counted as CP dots. (C) Example of the measurement of the tubulin (green) and the CCDC146 signal intensities measured at the white rectangle (2). The image corresponds to the orange rectangle in (A). In this example, the CCDC146 signal is localized in the right doublet of microtubules (Tub2). (D) The position of the CCDC146 signal with respect to the tubulin signal was measured along the entire flagellum. Each CCDC146 signal was assigned to a different compartment of the axoneme, allowing to obtain an histogram showing the distribution of CCDC146 labeling in ejaculated human sperm (n = 2, 38 dots analyzed). (E) Flagellum of a mouse epididymal spermatozoa observed with expansion microscopy. HA-CCDC146 sperm were immunolabeled with anti-HA #2 (red) and anti-β-tubulin (green). The upper image shows the sperm with merged immunostaining, and the lower images, the staining (red, green, and merge) observed in the principal piece of the flagellum. Scale bars 10 µm.
Figure 10 with 1 supplement
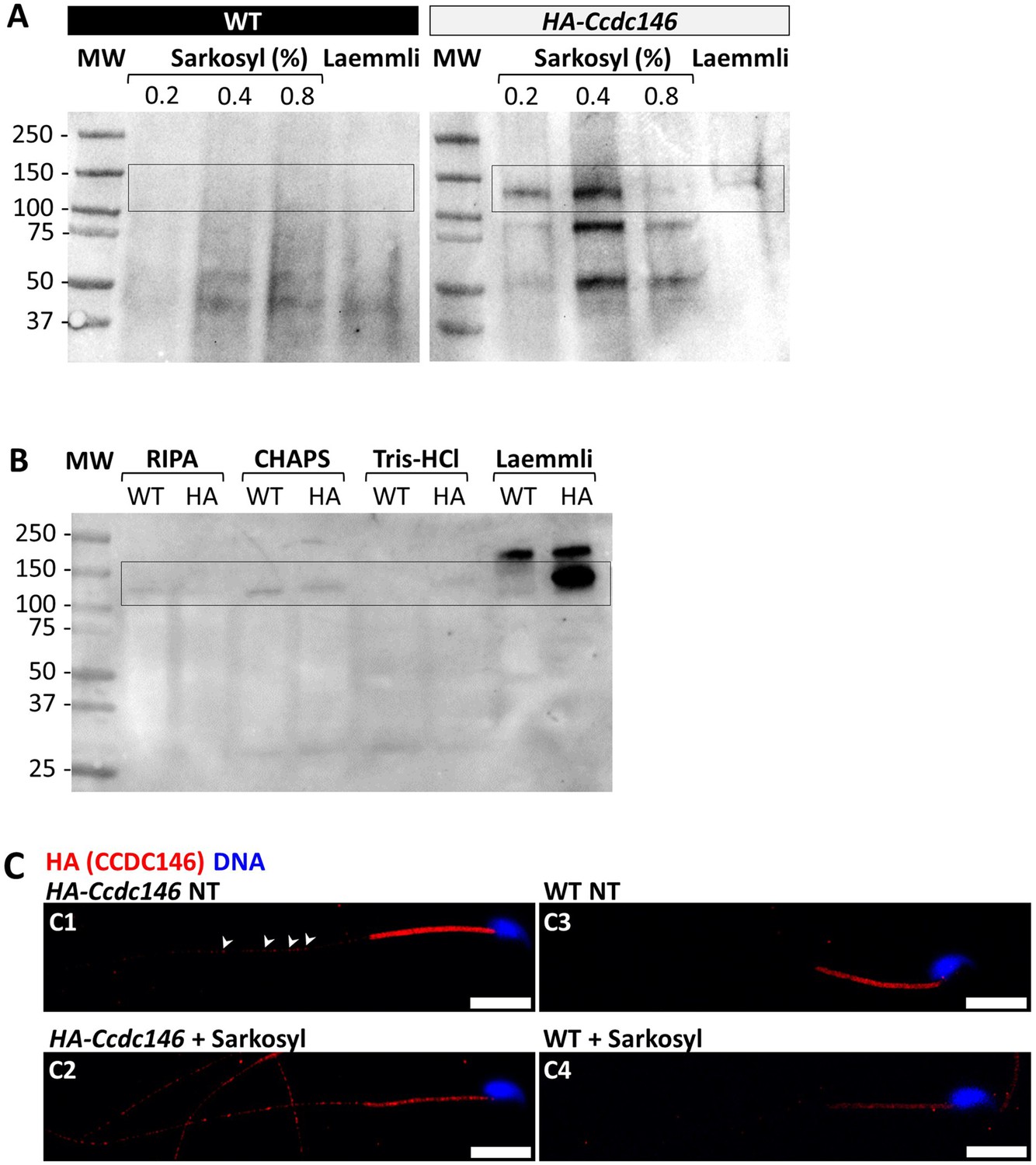
CCDC146 is solubilized by sarkosyl and sarkosyl treatment enhances IF signal.
(A) Western blot of WT and HA-CCDC146 sperm extract solubilized with N-lauroylsarcosine (sarkosyl), an anionic detergent. Sarkosyl was used at increasing concentrations (0.2 and 0.4%). The presence of HA-CCDC146 was detected by an anti-HA Ab. (B) Western blot of WT and HA-CCDC146 sperm extracts solubilized with alternative detergents (RIPA, CHAPS, Tris-HCl) and whole sperm extract solubilized in Laemmli. The presence of HA-CCDC146 was revealed by an anti-HA Ab. (C) Epididymal HA-CCDC146 sperm (C1–C2) and WT sperm (C3–C4), treated with sarkosyl (5 min, 0.2% sarkosyl) or not (NT), were immunostained to reveal the HA-tag (red) and counterstained with Hoechst (blue). (C1) Without treatment, a faint CCDC146 signal (white arrowheads) is observed along the flagellum from HA-CCDC146 sperm. (C2) Treatment with sarkosyl enhanced the HA-CCDC146 signal along the sperm flagellum. (C3) The HA signal present the midpiece is likely non-specific since it is present in WT non-treated (NT) sperm. See also Figure 10—figure supplement 1B, suggesting that this signal is due to secondary Abs. (C4) The HA signal in WT sperm is not enhanced by sarkosyl treatment. Scale bars 10 μm.
-
Figure 10—source data 1
Uncropped gel of Figure 10A: tagged sperm.
- https://cdn.elifesciences.org/articles/86845/elife-86845-fig10-data1-v1.zip
-
Figure 10—source data 2
Uncropped gel of Figure 10A: WT sperm.
- https://cdn.elifesciences.org/articles/86845/elife-86845-fig10-data2-v1.zip
-
Figure 10—source data 3
Uncropped gel of Figure 10B.
- https://cdn.elifesciences.org/articles/86845/elife-86845-fig10-data3-v1.zip
Figure 10—figure supplement 1
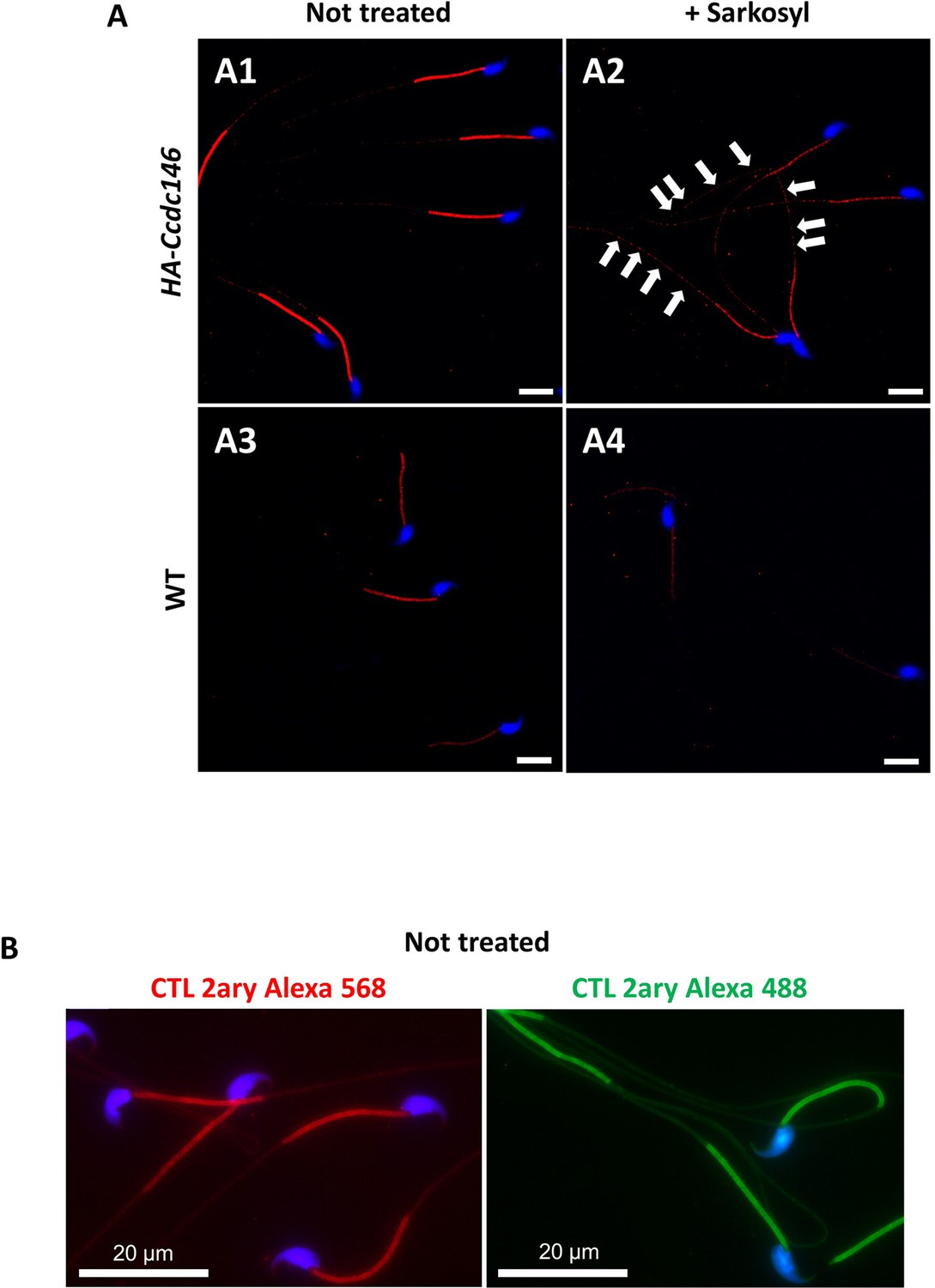
Sperm sarkosyl treatment corroborates the presence of CCDC146 along the mouse flagellum.
HA-CCDC146 sperm (A1–A2) and epididymal WT (A3–A4) not treated or treated (5 min, 0.2% sarkosyl) were immunostained with anti-HA Ab (red) and counterstained with Hoechst (blue). (A1) Without treatment, a faint HA-CCDC146 signal was observed along the HA-CCDC146 principal piece. The strong staining in the midpiece is not specific (see panel B). (A2) After treatment with sarkosyl, the HA-CCDC146 signal along the sperm principal piece was enhanced (white arrows), whereas the signal in the midpiece decreased. (A3) WT untreated (NT) sperm exhibited almost no HA-CCDC146 signal in the principal piece. The midpiece is stained but it is likely not specific (see panel B). (A4) The HA signal is not enhanced in WT principal piece by sarkosyl treatment, suggesting that the enhanced signal observed with sarkosyl on HA-CCDC146 sperm is specific. Scale bars 10 μm. (B) HA-CCDC146 sperm were only immunolabeled with secondary antibodies used to reveal HA staining. Strong staining is observed on the midpiece, confirming its non-specific nature. Scale bars 20 μm.
Figure 11
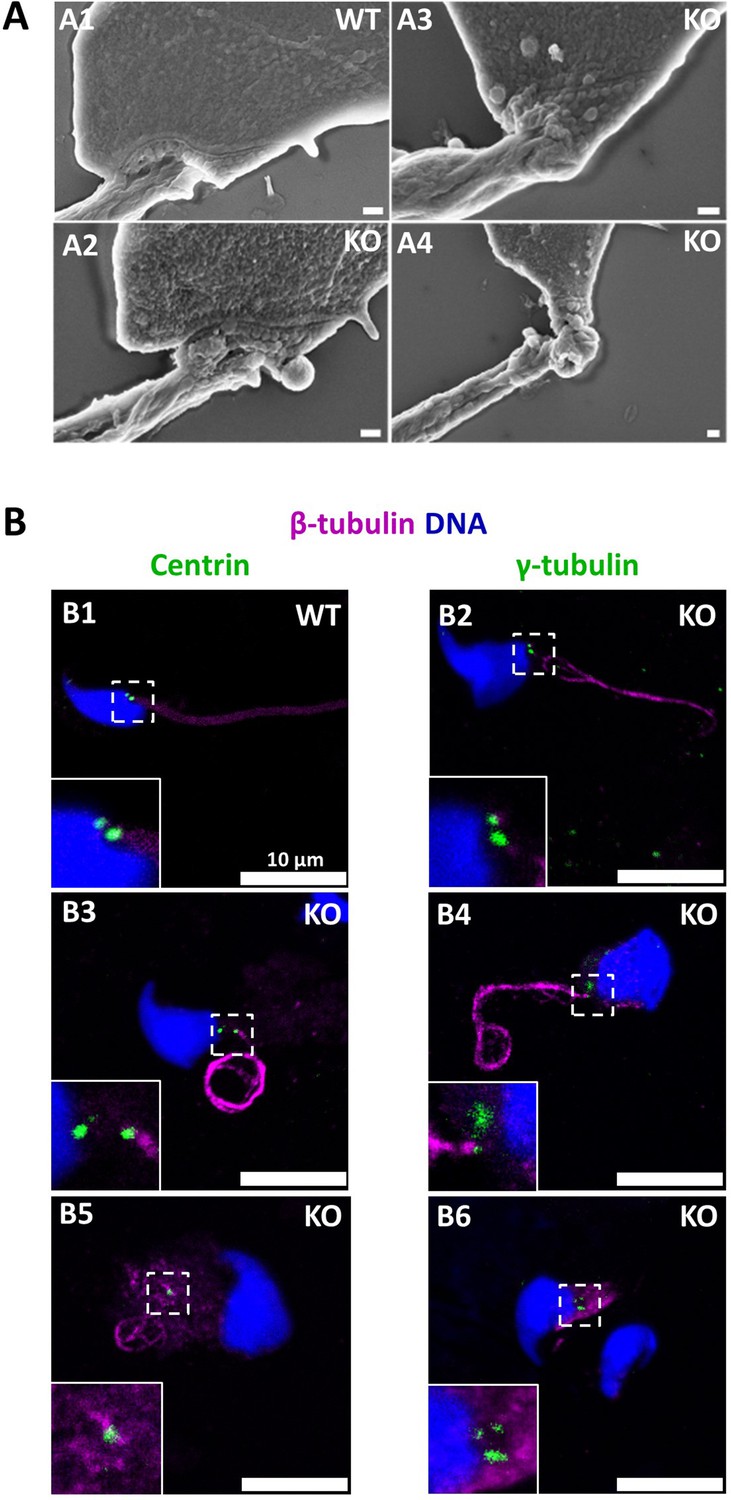
The absence of CCDC146 causes defects of the head-tail coupling apparatus in epididymal spermatozoa and duplication and mislocalization of centriole in testicular sperm.
(A) Scanning electron microscopy of WT and Ccdc146 KO epididymal spermatozoa shows aberrant head morphologies and irregular head-tail coupling apparatus (HTCA) linking the sperm head with the flagellum. There is a great variability in the morphological damage, with sperm presenting almost intact HTCA (A2), whereas other were strongly impacted by the absence of the protein (A3–4). Scale bars 200 nm. (B) Testicular spermatozoa from WT (B1) and Ccdc146 KO (B2–B6) mice immunolabeled with anti-β-tubulin (magenta) and anti-centrin (B1–B3) or anti-γ-tubulin (B4–B6) (green) Abs. Centrioles appeared to be normal (B2) in some spermatozoa, separated but partially attached to the head (B3, B4), completely detached from the sperm head (B5) or duplicated (B6). Scale bar 10 µm.
Figure 12 with 2 supplements
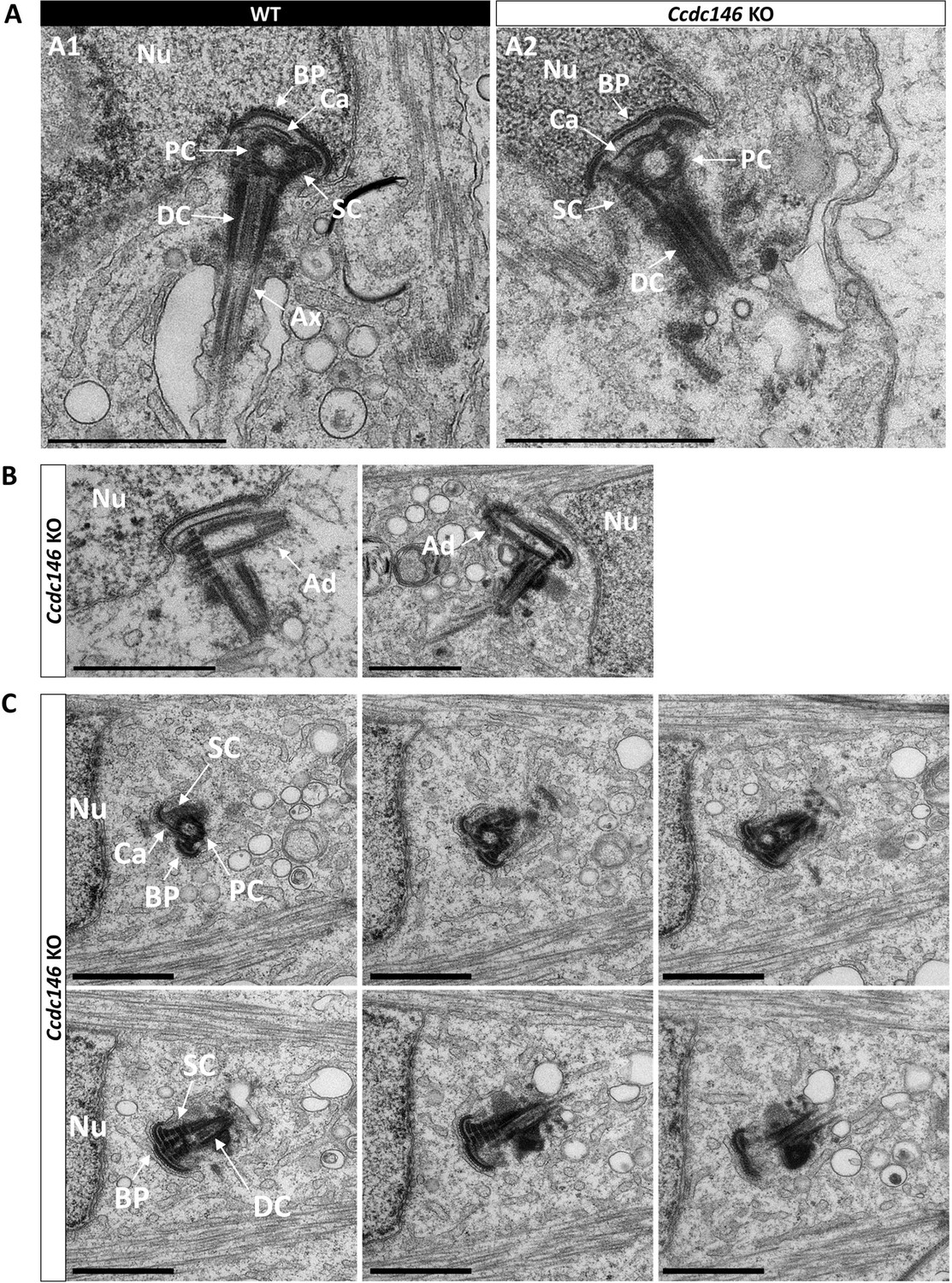
Absence of elongation of axonemal microtubules at the base of the distal centriole.
(A) In WT spermatids (A1), the proximal centriole (PC) is linked to the base of the compacting nucleus (Nu) through the basal plate (BP) and the capitulum (Ca), and the distal centriole (DC) is embedded in the segmented column (SC). All these sperm-specific cytoskeletal structures make up the head-tail coupling apparatus (HTCA). At the base of the distal centriole, axonemal microtubules (Ax) grow. In Ccdc146 KO elongating spermatids (A2), the overall structure of the HTCA is conserved, with the presence of the centrioles and the accessory cytoskeletal structures. However, no axonemal microtubules are visible, emerging from the DC. (B) The adjunct (Ad) of the proximal centriole is also preserved in Ccdc146 KO spermatids. (C) Serial sections of the HTCA of a Ccdc146 KO spermatid confirm the absence of axonemal microtubules at the base of the DC during spermatid elongation. Scale bars 1 µm.
Figure 12—figure supplement 1

Ultrastructure of WT sperm showing the head to tail coupling apparatus (HTCA) in elongating spermatids.
The adjunct is connected to the proximal centriole (PC). The axoneme (Ax) is elongated from the distal centriole (DC). Scale bars 500 nm.
Figure 12—figure supplement 2
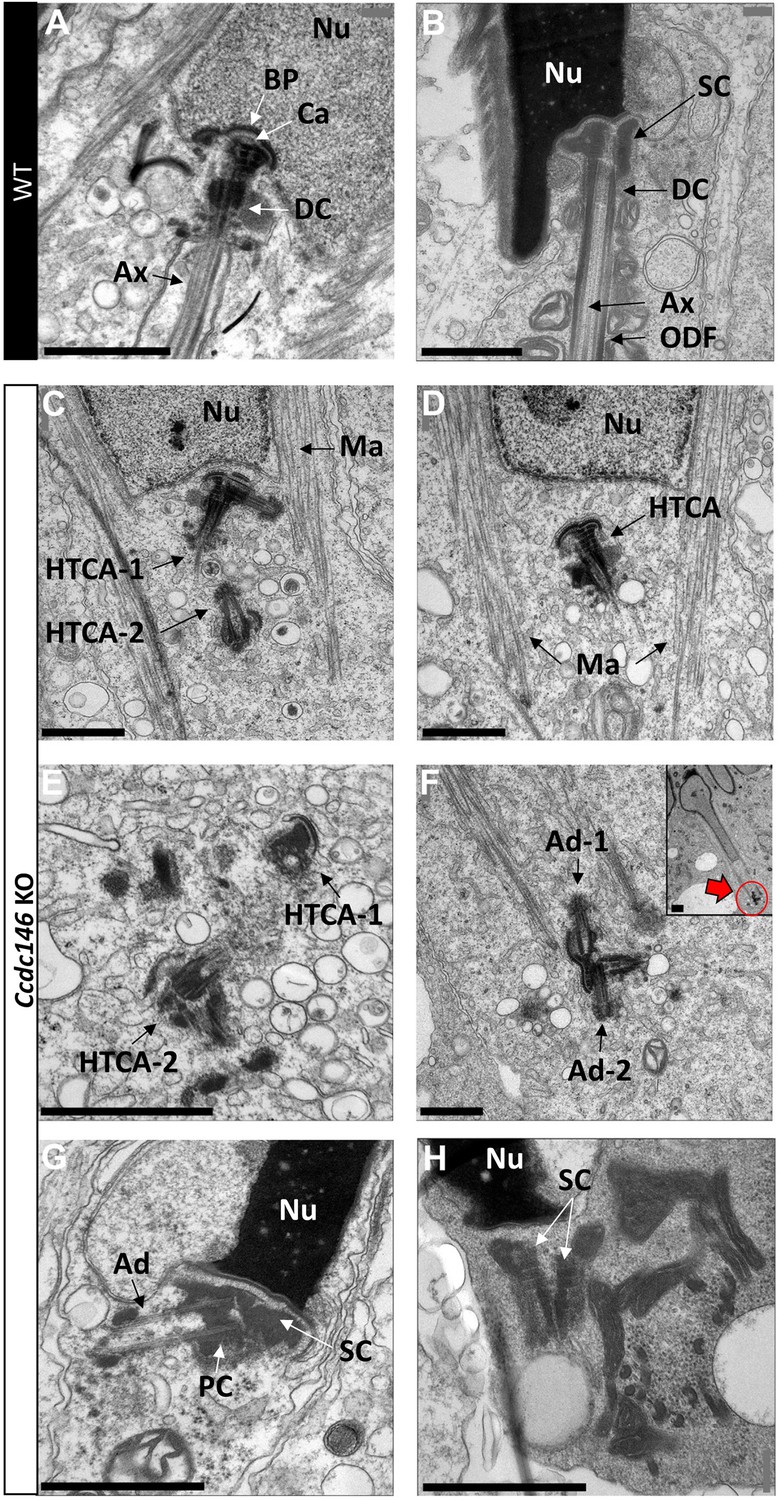
Lack of CCDC146 causes centriole duplication and mislocalization in Ccdc146 KO spermatids.
Ultrastructural analysis of centrioles in adult mouse WT (A, B) and Ccdc146 KO (C–H) testis sections. (A) In WT spermatids, the axoneme is attached to the base of the compacting nucleus (Nu) through the basal plate (BP) and the capitulum (Ca), and the distal centriole (DC) is embedded in the segmented column (SC). These sperm-specific cytoskeletal structures make up the head-to-tail coupling apparatus (HTCA). (B) In WT elongated spermatids, the different components of the axonemal structures (Ax) and outer dense fibers (ODF) were visible downstream the distal centriole. (C) In Ccdc146 KO elongating spermatids, the overall structure of the HTCA was conserved, with the presence of the centrioles and the accessory cytoskeletal structures. However, the HTCAs were often duplicated (C, E, F) and separated from their usual nuclear attachment site (C–F, H), and sometimes misplaced far away from the nucleus (F), the red arrow in (F) indicating the misplaced centrioles at the end of the manchette. The axoneme is missing (C–F). In elongated spermatids with condensed nucleus, malformed and detached centrioles with poorly assembled or missing flagella compared to the WT (B) can be seen. Manchette (Ma), adjunct (Ad), proximal centriole (PC). Scale bars 1 µm.
Figure 13 with 1 supplement
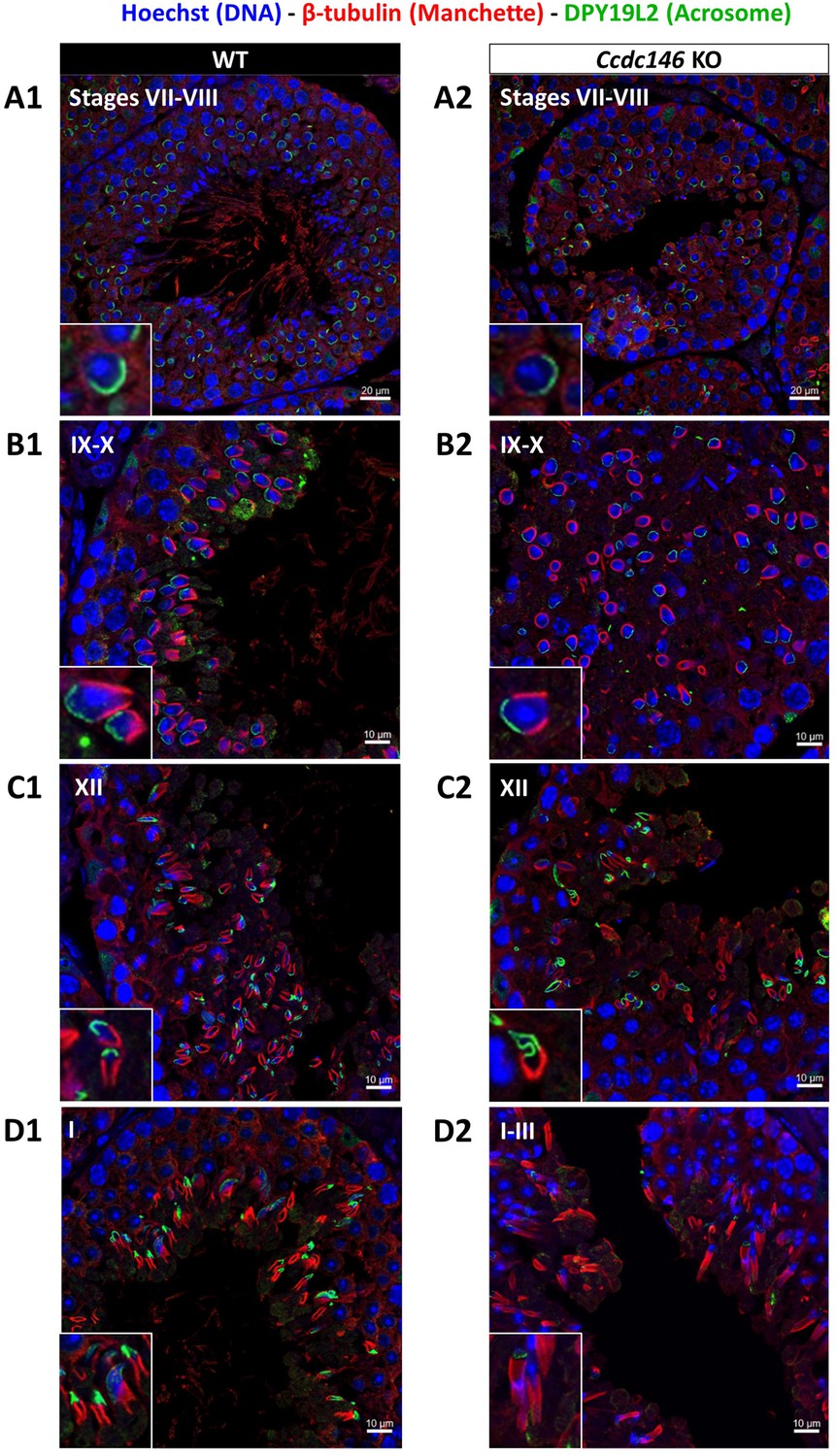
Analysis of stages of spermatogenesis by IF reveals acrosome formation and manchette elongation defects in Ccdc146 KO spermatids.
Cross-sections of WT (A1–D1) and Ccdc146 KO (A2–D2) testes showing different stages of mouse spermatogenesis (I, VII–XII). Stages were determined by double immunostaining for β-tubulin (red; manchette elongation) and DPY19L2 (green; acrosome localization), and DNA was stained with Hoechst (blue). (A1/A2) At stage VII–VIII, acrosome spreading on round spermatids appeared similar between WT and KO. However, very few mature spermatozoa lined the lumen in the KO. (B1/B2) Cell orientations appeared random from stage IX–X in the KO and the tubules contained more advanced spermatid stages. Random orientations of cells are evidenced by manchette cross-sections in various planes. (C2) Abnormal acrosomes of elongating KO spermatids are observed by stage XII. (D1/D2) The manchette of elongated spermatids at stage I–III was longer in the KO compared with the WT. Insets in A1–D1 and A2–D2 show typical spermatids from stages I–XII from WT and KO males, respectively, showing details of acrosome formation (green) and manchette elongation (red). Scale bars stages VII–VIII 20 µm, scale bars IX–X, XII and I–III 10 µm.
Figure 13—figure supplement 1
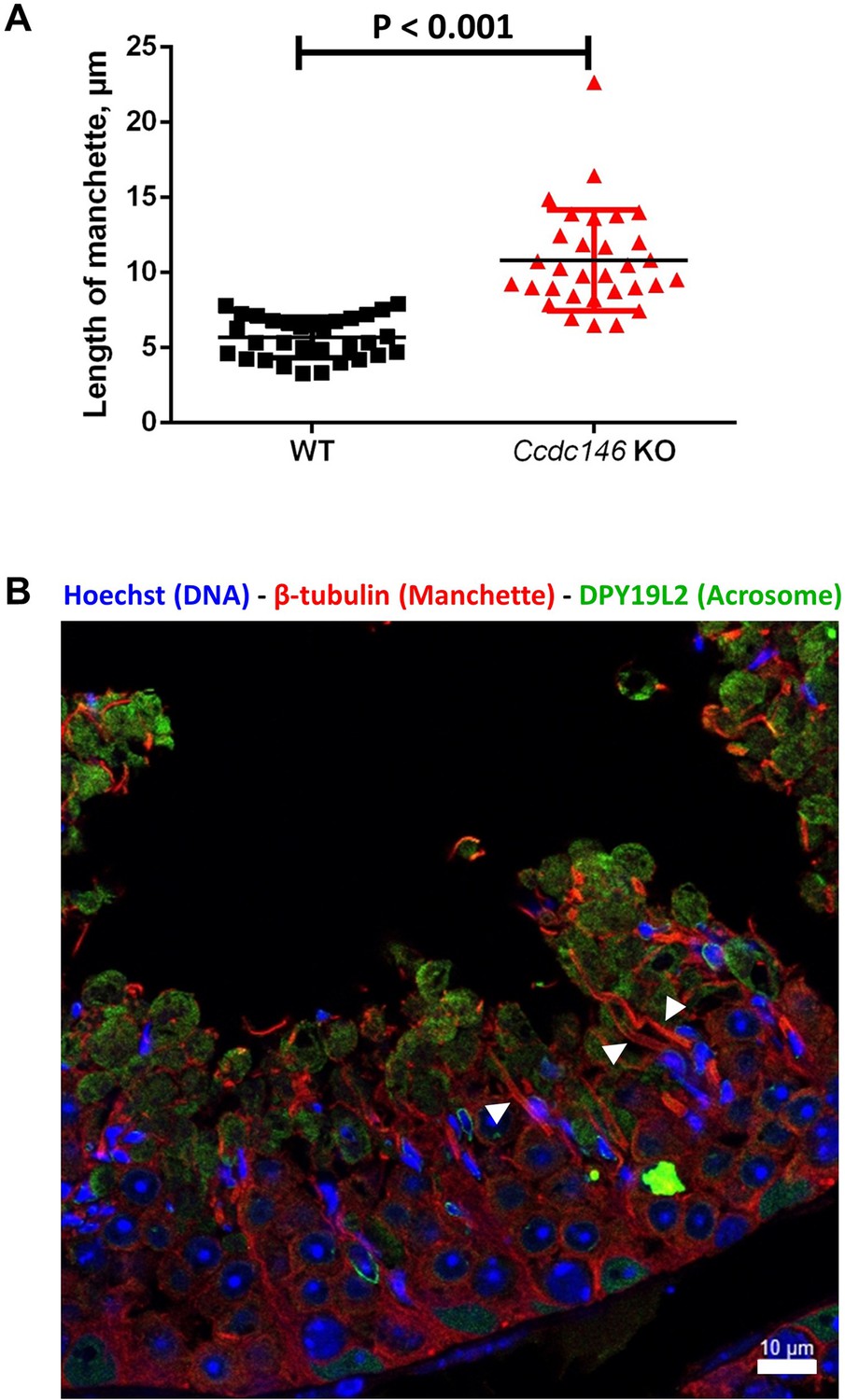
The manchette of elongating spermatids from Ccdc146 KO male is longer than those from WT males.
(A) Comparison of the length of the manchette of elongating spermatids measured in cross-sections of seminiferous tubules at stage I for WT and stages I–III for Ccdc146 KO spermatids (n=32 and 31 for WT and Ccdc146 KO, respectively). Statistical significance between WT and Ccdc146-KO spermatids was assessed by Mann–Whitney test, p-value of 0.05 or lower was considered statistically significant. Error bars show standard deviation. p-value as indicated. (B) Examples of very long manchettes (white arrowheads) in Ccdc146 KO cross-sections of seminiferous tubules at stages I–III. WT cross-section is presented Figure 13D1. Scale bars as indicated.
Figure 14
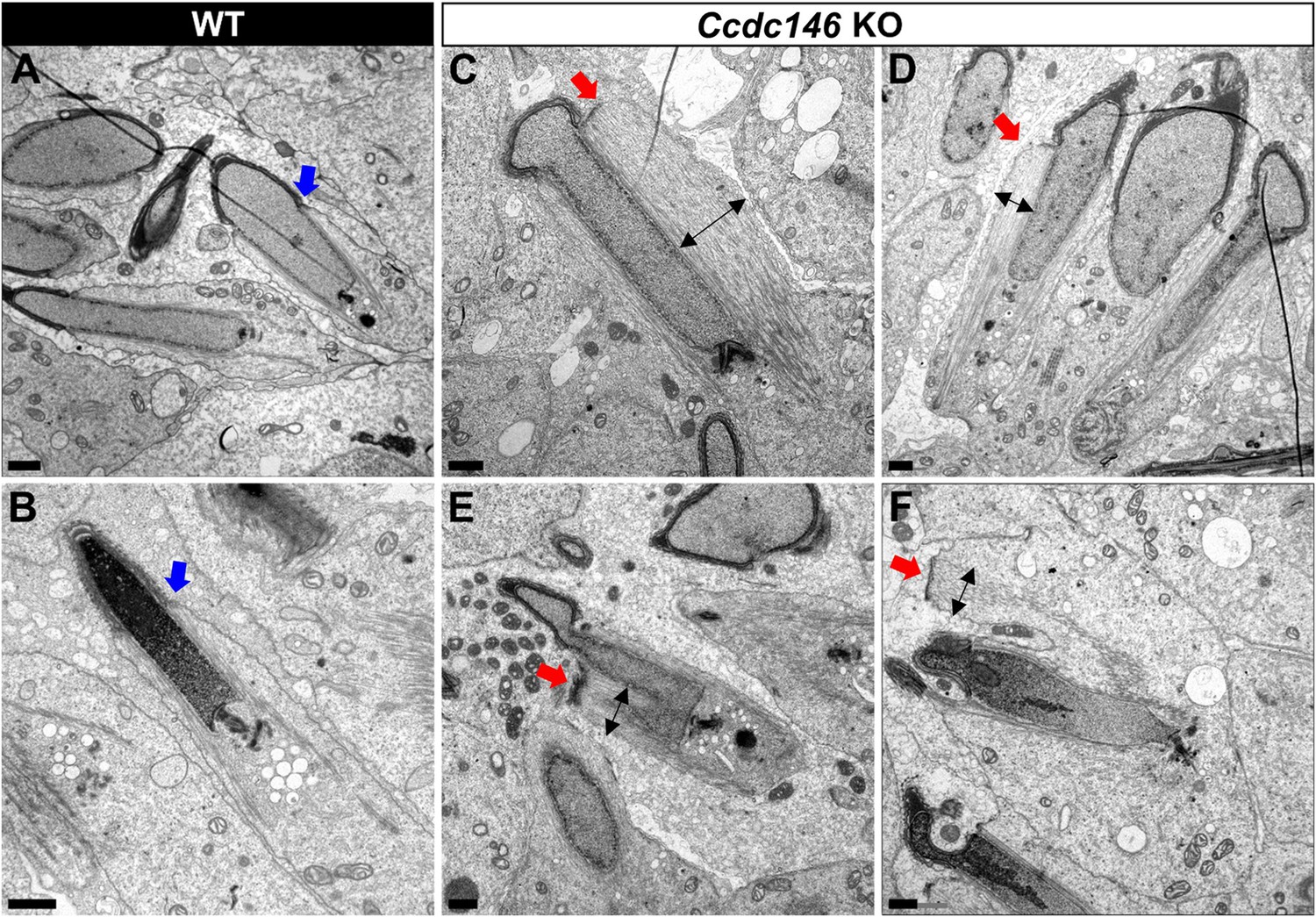
TEM of elongating spermatids from Ccdc146 KO male shows ultrastructural defects of the manchette.
(A, B) Ultrastructural analysis of the manchette in WT elongating spermatids shows the normal thin perinuclear ring, anchored below the acrosome (blue arrows) and allowing a narrow array of microtubules to anchor. (C–F) In elongating spermatids from Ccdc146 KO animals, the perinuclear ring was abnormally broad, usually located on one side of the spermatid (red arrows), creating an asymmetric and wide bundle of microtubules. The resulting manchette was wider and often longer than in WT animals (black double arrows). (F) The tubulin nucleation location was sometimes ectopic in the KO (red arrow) and coincided with irregularly shaped sperm heads. Scale bars 1 µm.
Figure 15 with 1 supplement

The axonemes of Ccdc146 KO spermatids present multiple defects visible under TEM.
(A1) A longitudinal section of a WT flagellum shows a typical structure of the principal piece, with outer dense fibers (ODF) at the periphery, microtubules (MT) in the center, and mitochondria (Mi) aligned along the flagellum. (A2) A longitudinal section of a Ccdc146 KO flagellum shows a disorganized midpiece, with altered mitochondria, the presence of an amorphous fibrous sheath (FS) and altered microtubules. Scale bars 0.5 µm (B) Longitudinal section of a Ccdc146 KO sperm showing dispersed and non-assembled flagellar material in a cytoplasmic mass. The right-hand image is the enlargement of the red square, showing the presence of an external ring of mitochondria surrounding an ODF ring devoid of microtubular material. (C) Longitudinal section of another Ccdc146 KO sperm showing a similar abnormal midpiece structure. The right-hand image is the enlargement of the red square, showing the presence of an external ring of mitochondria surrounding an ODF ring devoid of microtubular material. Scale bars as indicated.
Figure 15—figure supplement 1
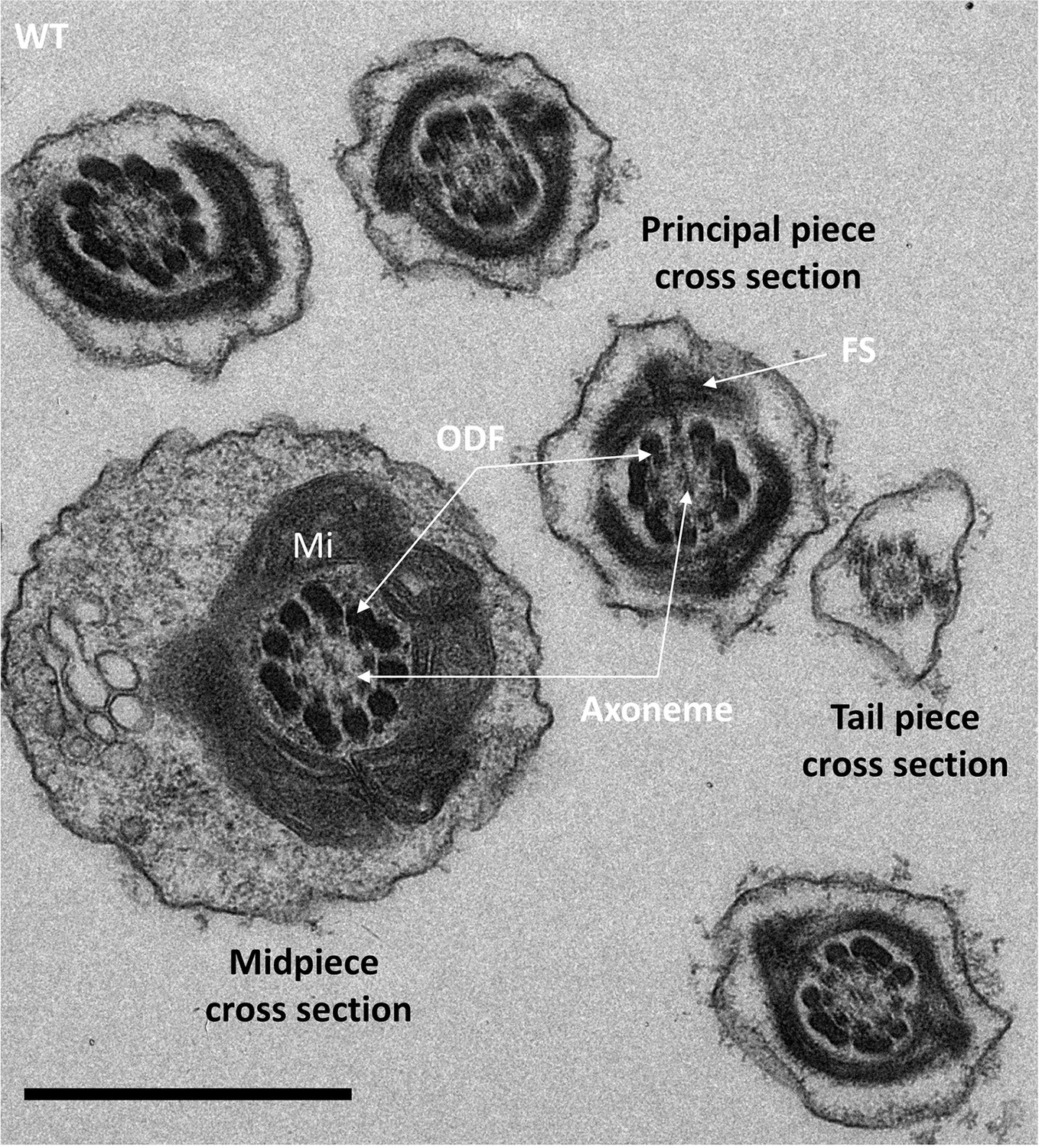
Cross-sections of the midpiece, principal piece, and tail piece of WT elongating spermatids showing the ultrastructure of the different pieces.
In the midpiece cross section, the axoneme is surrounded by nine outer dense fibers (ODF) and an external layer of mitochondria (Mi). In the principal piece, the ODF are surrounded by the fibrous sheet (FS). Scale bar 1 µm.
Figure 16
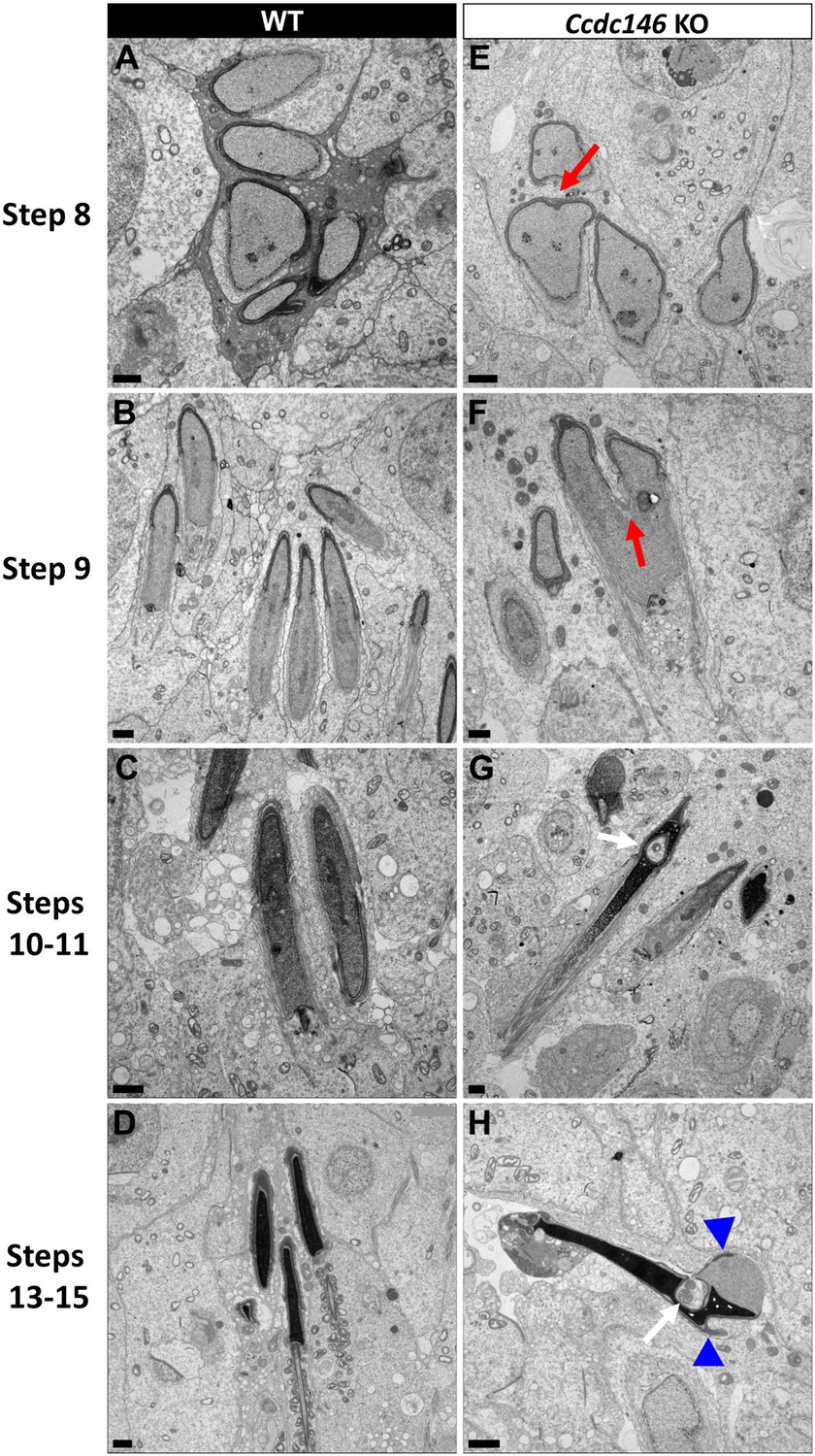
Spermatid head shape is aberrant in the absence of CCDC146.
Comparative ultrastructural analysis of the spermatid head in WT (A–D) and Ccdc146 KO (E–H) testis sections. (A, E) Spermatid nuclei at the beginning of elongation. KO spermatid nuclei showing nuclear membrane invaginations and irregular shape that were not present in the WT (red arrow). The acrosome of KO spermatids appeared intact. (B, F) Morphology of nuclei in elongating spermatids. Whereas nucleus elongation is symmetric in the WT, in the Ccdc146 KO, more pronounced head invaginations are observed (red arrow). (C, G) Elongated spermatids. Although nuclear condensation appeared normal in both WT and KO nuclei, vacuolization is observed in KO nucleus (white arrow). (D, H) Elongated KO spermatids showed malformed elongated nuclear shapes with frequent invaginations (white arrow) and absence of flagella (H) compared to the WT (D). The acrosome of KO spermatids showed numerous defects such as detachment induced by swelling/bubbling of the plasma membrane (blue arrowheads). Scale bars 1 µm.
Tables
Table 1
Detailed semen parameters of both multiple morphological abnormalities of the flagellum (MMAF) individuals harboring a CCDC146 variant.
Values are percentages unless specified otherwise. NA: not available, Reference limits (5th centiles and their 95% confidence intervals) according to World Health Organization (WHO) standards (Cooper et al., 2010). Patient 08IF39 variant c.1084C>T and patient O9IF26 variant c.2112Del. Green and red colors indicate normal and abnormal values, respectively.
| Patient# 08IF039 | Patient# 09IF026 | Lower reference limits (WHO) | |
|---|---|---|---|
| Age (years) | 48 | 52 | |
| Semen volume (mL) | 5.5 | 2 | 1.5 |
| pH | NA | 7.7 | ≥7.2 |
| Viscosity (normal: 1; abnormal: 2) | 2 | 1 | |
| Sperm concentration (million/mL) | 35 | 42 | 15 |
| Total sperm number (million/ejaculate) | 192.5 | 84 | 39 |
| Non sperm cells/round cells (million/mL) | 3.2 | 1 | |
| Polynuclear neutrophils | NA | 0 | |
| Total motility (a + b + c) after 1 hr (%) | 15 | 10 | 40 |
| Progressive motility (PR; a + b) | 10 | 7 | 32 |
| Vitality (%) | 42 | 62 | 58 |
| Sperm morphology: normal forms (%) | 1 | 0 | 4 |
| Multiple anomalies index (MAI) | NA | 2.4 | − |
| Tapered head | NA | 5 | − |
| Thin head | NA | 2 | − |
| Microcephalic | NA | 6 | − |
| Macrocephalic | NA | 2 | − |
| Multiple heads | NA | 0 | − |
| Abnormal base (abnormal post-acrosomal region) | NA | 6 | − |
| Abnormal acrosomal region | NA | 34 | − |
| Excess residual cytoplasm | NA | 11 | − |
| Thin midpiece | NA | 3 | − |
| Bent or misaligned tail | NA | NA | |
| No tail | >10% | 35 | − |
| Short tail | >10% | 37 | − |
| Irregularly shaped tail | >10% | 67 | − |
| Coiled tail | >10% | 18 | − |
| Multiple tails | >10% | 3 | − |
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene (human) | CCD146 | GenBANK | Gene ID: 57639 | |
| Gene (Mus musculus) | Ccdc146 | GenBANK | Gene ID: 75172 | |
| Cell line (human) | HEK293T | ATCC | ||
| Transfected construct (M. musculus) | Myc-DDK-tagged Ccdc146 plasmid DNA | OriGene Technologies, Inc | NM_029195 | Validation of anti-CCDC146-human |
| Biological sample (human) | Human Foreskin Fibroblasts | This paper | Primary cell line | |
| Antibody | Anti-human CCDC146 (rabbit polyclonal) | Atlas Antibodies | HPA020082 | IF: 1/200 |
| Antibody | Anti-centrin- clone 20H5 (mouse monoclonal) | Merck | 04-1624 | IF: 1/200 |
| Antibody | Anti-γ-tubulin (mouse monoclonal) | Santa Cruz Biotechnology | sc-17787 | IF: 1/500 |
| Antibody | Anti-β-tubulin (guinea pig polyclonal) | Geneva Antibody Facility | AA344-GP | IF: 1/500 |
| Antibody | Anti-β-tubulin (mouse monoclonal) | Cell Signaling Technology | 2128 | IF: 1/100 |
| Antibody | Anti-PCM1 (G-6) (mouse monoclonal) | Santa Cruz Biotechnology | sc-398365 | IF: 1/200 |
| Antibody | Anti-α-tubulin (recombinant antibody- isotype mouse monoclonal) | Geneva Antibody Facility | AA-344 | U-ExM: 1/250 IF: 1/500 https://doi.org/10.24450/journals/abrep.2019.e108 |
| Antibody | Anti-β-tubulin (recombinant antibody- isotype mouse monoclonal) | Geneva Antibody Facility | AA-345 | U-ExM: 1/250 IF: 1/500 https://doi.org/10.24450/journals/abrep.2019.e108 |
| Antibody | Anti-POC5 (rabbit polyclonal) | Bethyl | A303-341A | IF: 1/250 U-ExM: 1/200 |
| Antibody | Anti-high affinity (HA) (rat monoclonal) | Roche | 11867423001 | IF: 1/400 U-ExM: 1/400 WB: 1/2500 |
| Antibody | Anti-high affinity (HA) (recombinant antibody isotype rabbit monoclonal) | Cell Signaling Technology | 3724 | U-ExM: 1/100 |
| Antibody | Anti-high affinity (HA) (rabbit polyclonal) | Sigma-Aldrich | H6908 | U-ExM: 1/200 |
| Antibody | HA Tag Alexa Fluor 488-conjugated antibody (recombinant Ab, rabbit monoclonal) | Cell Signaling | 28427 | IF:1/400 |
| Antibody | Anti- Dpy19L2 (rabbit polyclonal) | Home made as described in Pierre et al., 2012 | IF: 1/100 | |
| Sequence-based reagent | gRNA-Ex2 | GenScript | gRNA for exon 2 of Ccdc146 KO | 5′-CCT ACA GTT AAC ATT CGG G-3′ |
| Sequence-based reagent | gRNA-EX4 | GenScript | gRNA for exon 4 of Ccdc146 KO | 5′-GGG AGT ACA ATA TTC AGT AC-3′ |
| Sequence-based reagent | gRNA-Ex2 for HA-tag | GenScript | gRNA for exon 2 of Ccdc146 tag | 5′-TAC TTT AGA ACT GTG AAA AA-3′ |
| Sequence-based reagent | Ccdc146-F | Eurogentec | Forward primer for RT-PCR experiment | 5′-TGCTGCATGACGCCGTGATG-3′ |
| Sequence-based reagent | Ccdc146-R | Eurogentec | Reverse primer for RT-PCR experiment | 5′-GGAGACCTCCGTGGAGAATGCTTC-3′ |
| Sequence-based reagent | Hprt-F | Eurogentec | Forward primer for RT-PCR experiment | 5′-CCTAATCATTATGCCGAGGATTTGG-3′ |
| Sequence-based reagent | Hprt-R | Eurogentec | Reverse primer for RT-PCR experiment | 5′-TCCCATCTCCTTCATGACATCTCG-3′ |
| Sequence-based reagent | Actb-F | Eurogentec | Forward primer for RT-PCR experiment | 5′-CTTCTTTGCAGCTCCTTCGTTGC-3′ |
| Sequence-based reagent | Actb-R | Eurogentec | Reverse primer for RT-PCR experiment | 5′-AGCCGTTGTCGACGACCAGC-3′ |
| Commercial assay or kit | Click-iT Plus TUNEL Assay kit | Invitrogen | C10617 |
Table 2
List of primers used for RT-PCR experiments.
| Forward primer | Reverse primer | |||
|---|---|---|---|---|
| Genes | Sequence | Concentration (nM) | Sequence | Concentration (nM) |
| Ccdc146 | 5′-TGCTGCATGACGCCGTGATG-3′ | 750 | 5′-GGAGACCTCCGTGGAGAATGCTTC-3′ | 500 |
| Hprt | 5′-CCTAATCATTATGCCGAGGATTTGG-3′ | 500 | 5′-TCCCATCTCCTTCATGACATCTCG-3′ | 250 |
| Actb | 5′-CTTCTTTGCAGCTCCTTCGTTGC-3′ | 250 | 5′-AGCCGTTGTCGACGACCAGC-3′ | 250 |
Additional files
-
Supplementary file 1
Amino acid conservation of CCDC146 orthologs from man to X. tropicalis.
The amino acids altered and missing from the presence of the variant p.Arg704serfsTer7 are highlighted in yellow.
- https://cdn.elifesciences.org/articles/86845/elife-86845-supp1-v1.pdf
-
Supplementary file 2
Lists of primers for genotyping.
(a) List of primers used for Sanger verification of the identified variants by WES. (b) List of primers used for knock-out mice genotyping. (c) List of primers used for knock-in mice (HA-Tag) genotyping.
- https://cdn.elifesciences.org/articles/86845/elife-86845-supp2-v1.docx
-
MDAR checklist
- https://cdn.elifesciences.org/articles/86845/elife-86845-mdarchecklist1-v1.pdf
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Lack of CCDC146, a ubiquitous centriole and microtubule-associated protein, leads to non-syndromic male infertility in human and mouse
eLife 12:RP86845.
https://doi.org/10.7554/eLife.86845.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}