Two-way Dispatched function in Sonic hedgehog shedding and transfer to high-density lipoproteins
- Institute of Physiological Chemistry and Pathobiochemistry, University of Münster, Germany
Figures
Figure 1 with 2 supplements
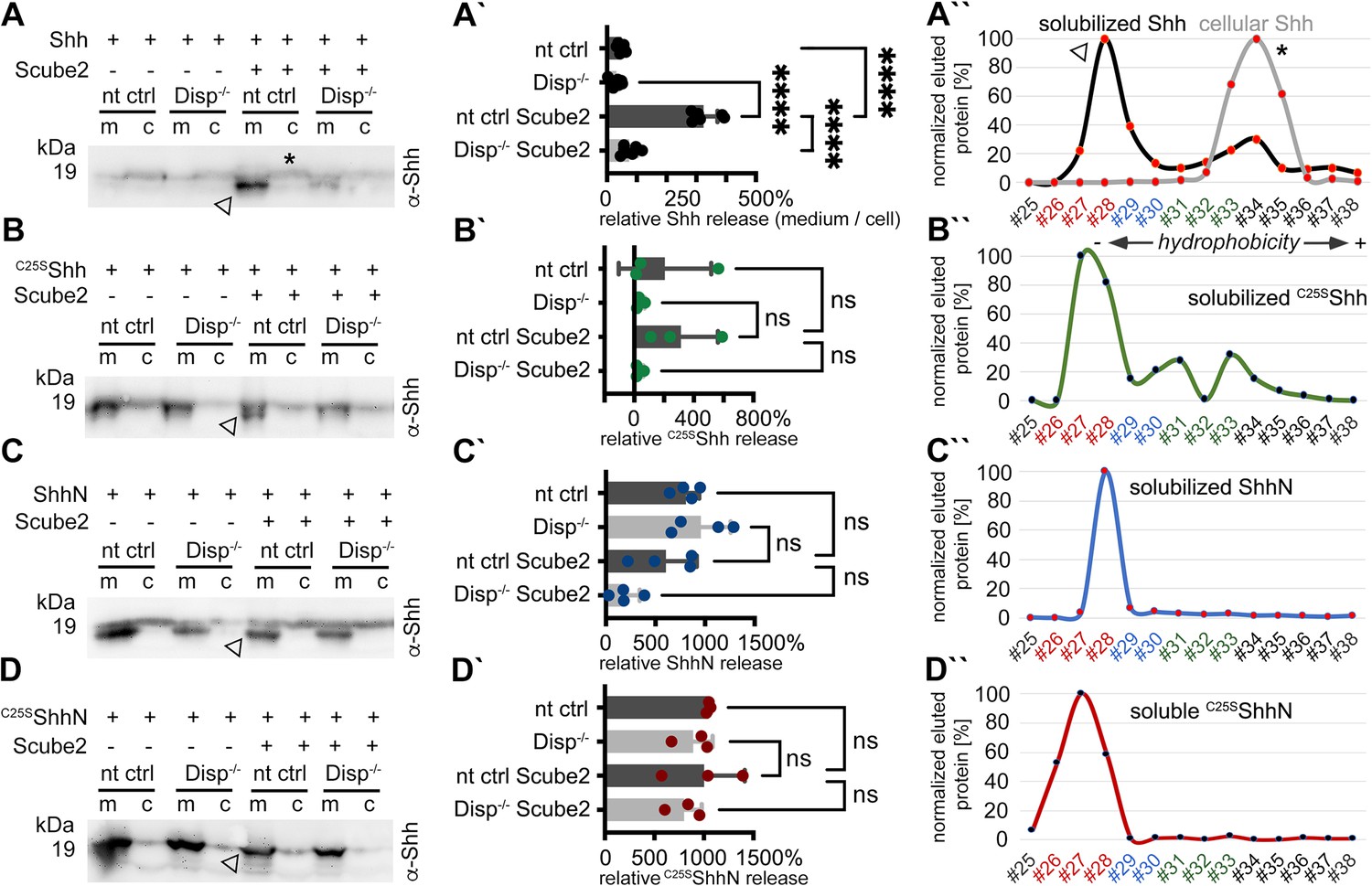
Disp and Scube2 enhance the shedding of cell surface-associated dual-lipidated Shh into delipidated soluble forms.
Media containing 10% serum was changed to serum-free Dulbecco’s Modified Eagle’s Medium (DMEM) 36 hr post-transfection and proteins were solubilized for 6 hr. Cells were not washed between media changes to leave residual serum traces in the assay. We refer to this experimental condition as ‘serum-depleted’ throughout this article. (A) Cap-dependent Shh translation and cap-independent Hhat translation from bicistronic mRNA ensured the generation of dual-lipidated, plasma membrane-associated proteins (asterisk in the cellular fraction) in all transfected cells. Disp and Scube2 synergistically and specifically enhance the conversion of dual-lipidated Shh precursors into truncated soluble variants during release (arrowhead). m: media; c: cell lysate. (A’) Quantification of relative Shh release from non-targeting control (nt ctrl) and Disp-/- cells in the presence or absence of Scube2. Amounts of solubilized Shh with higher electrophoretic mobility (the lower bands) were quantified and expressed as % relative to the respective cellular Shh, which was always set to 100%. One-way ANOVA, Dunnett’s multiple-comparisons test. ****p<0.0001. See Supplementary file 1 for detailed statistical information. (A’’) Reverse-phase high-performance liquid chromatography (RP-HPLC) analyses revealed that Shh solubilized by Disp and Scube2 (the same fraction indicated by the arrowhead in A, black line) was less hydrophobic than its cell surface-associated precursor (gray line; the asterisk indicates analysis of the same cellular fraction as shown in A). RP-HPLC calibration and color coding of Shh fractions are shown in Figure 1—figure supplement 2B–G. (B–D) Solubilization of non-palmitoylated C25SShh (in this artificial variant, the N-terminal palmitate acceptor cysteine is replaced by a non-accepting serine; functionally equivalent constructs with the cysteine exchanged for a non-accepting alanine [C25AShh] were also used in our study), non-cholesteroylated but palmitoylated ShhN and lipid-free control C25SShhN under the same serum-depleted conditions. Arrowheads indicate Shh variants that were solubilized in Disp- and Scube2 presence. (B’–D’) Processed protein quantifications from (B–D), again from nt ctrl and Disp-/- cells in the presence or absence of Scube2. One-way ANOVA, Dunnett’s multiple-comparisons test. ns: p>0.05. See Supplementary file 1 for detailed statistical information. (B’’–D’’) RP-HPLC shows similar elution of C25SShh, ShhN, and non-lipidated C25SShhN. This indicates that terminal lipids were removed during the release of C25SShh and ShhN (as well as Shh, A’’).
-
Figure 1—source data 1
Raw data and statistical analyses of Figure 1.
A–D contain uncropped western blots shown in Figure 1A–D. Folders A’–D’ contain biological replicates of the respective experiments. Prizm files A’–D’ quantify relative Shh release rates based on the data shown in folders A’–D’.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig1-data1-v1.zip
Figure 1—figure supplement 1
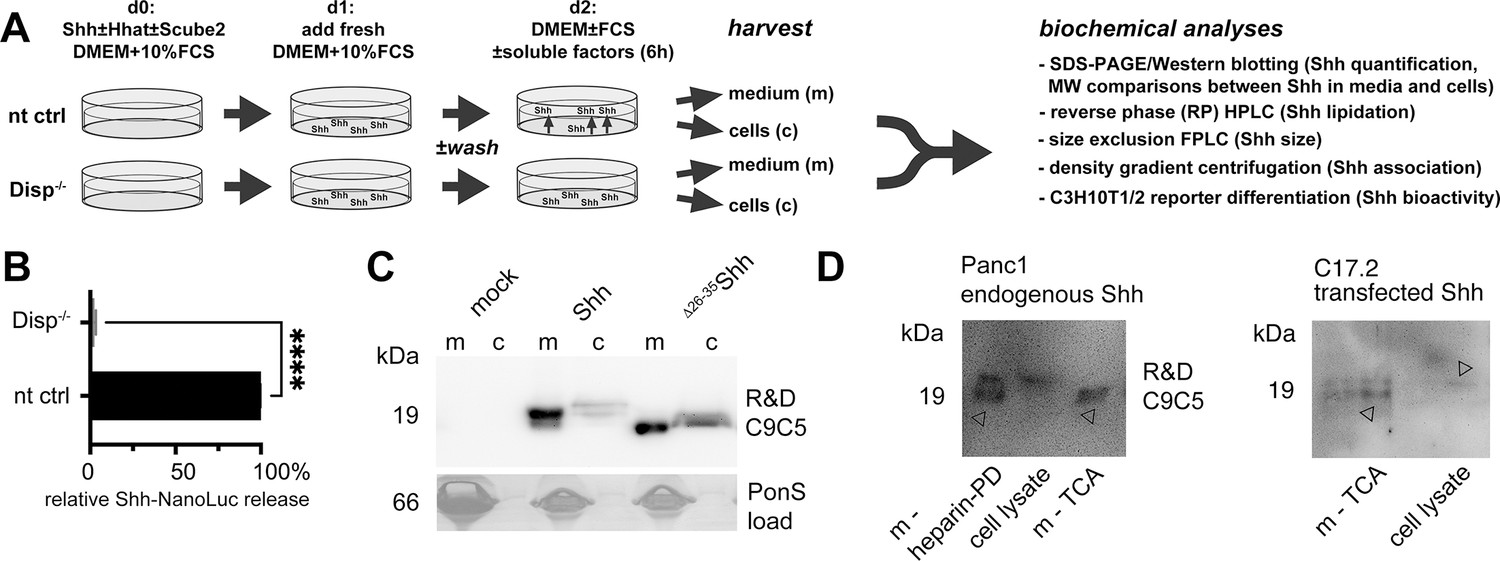
Experimental design and controls.
(A) nt Bosc23 ctrl cells that express Disp (top) or Disp-/- cells (bottom) were transfected on day 0 with Shh constructs together with Scube2 or empty control cDNA3.1. Fresh medium was added 1 day after transfection and cells were grown for 36 hr (100 µg/mL) in Dulbecco’s Modified Eagle’s Medium (DMEM) containing 10% fetal calf serum (FCS) and penicillin-streptomycin. For serum-depleted solubilization, on day 2, serum-containing media were aspirated and serum-free DMEM (with or without chloromethylketone [CMK] furin inhibitor, serum, or high-density lipoprotein [HDL]/low-density lipoprotein [lDL]) was added for 6 hr. Media were harvested and centrifuged at 300 × g for 10 min to remove debris. Supernatants containing FCS were subjected to heparin-pulldown, and serum-free or serum-depleted supernatants were incubated with 10% trichloroacetic acid for 30 min on ice, followed by centrifugation at 13,000 × g for 20 min to precipitate the proteins. Cells were lysed directly on the plate for subsequent reverse-phase high-performance liquid chromatography (RP-HPLC) or SDS-PAGE analysis. Size-exclusion chromatography (SEC), density gradient centrifugation and cell-based Shh activity/qPCR assays were also performed. (B) We confirmed impaired Shh solubilization from our Disp-/- cell line by using a Shh variant internally tagged with NanoLuciferase (Wang et al., 2021) and co-expressed with Hhat from bicistronic mRNA. Strongly reduced luminescence was detected in media from Shh-NanoLuc expressing Disp-/- cells. Unpaired t-test, two-tailed. *p=0.017, n = 3. Note that quantification of Shh-NanoLuc in media does not provide any information beyond increased/decreased protein release. In particular, it does not provide direct information on how Disp drives the release of dual-lipidated Hh from the plasma membrane, whether Disp acts directly or indirectly in this process, whether Shh remains fully lipidated during release, and to which carrier – if any – Hh is transferred. In contrast, the methods used in our work provide this information. (C) Cells were transfected with full-length Shh or N-truncated Δ26-35Shh, and proteins were concentrated by trichloroacetic acid (TCA) precipitation. The C9C5 antibody used for Shh detection throughout this work proved to be highly specific. (D) Reduced electrophoretic mobility of endogenous Shh from pancreatic ductal carcinoma (Panc1) cells (cell lysate) if compared to soluble proteins (m, arrowheads) indicates proteolytic processing of the endogenous protein during release. m-heparin-PD: Shh pulldown from serum-containing medium with heparin; m-TCA: TCA precipitation of Shh secreted into serum-depleted media. Shh was also released in processed form from transfected C17.2 cells, an immortalized mouse neural progenitor cell line derived from neonatal mouse cerebellum. This is consistent with N-truncated Shh release from cholesterol-depleted HeLa cells, Panc1 cells, and MIA PaCa-2 cells (Manikowski et al., 2019) and demonstrates that proteolytic Shh processing during release is not limited to our experimental Bosc23 system.
-
Figure 1—figure supplement 1—source data 1
Raw data of Figure 1—figure supplement 1.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig1-figsupp1-data1-v1.zip
Figure 1—figure supplement 2
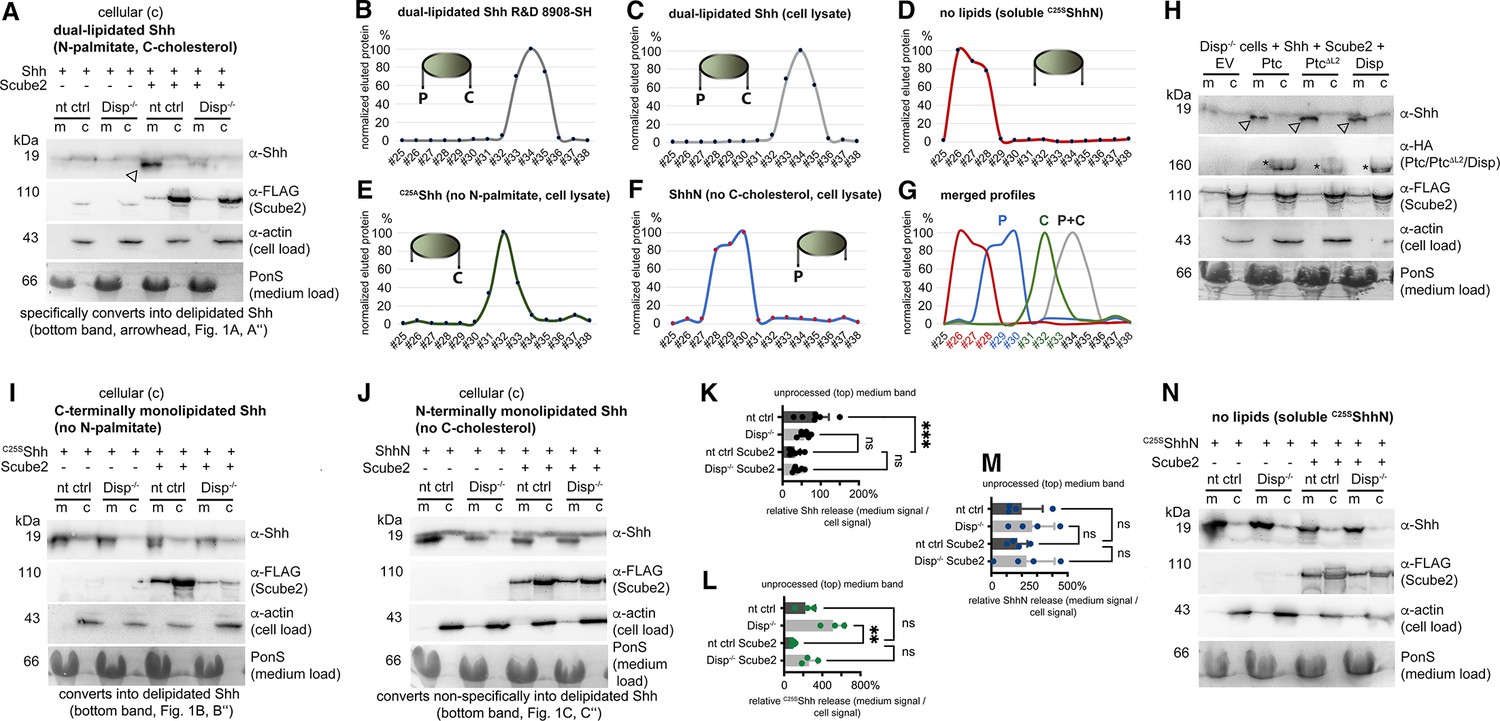
Loading controls, activity controls, and standards on the same stripped blots.
(A) Actin served as a loading control for cell lysates and PonceauS as a loading control in media, and α-FLAG antibodies detected FLAG-tagged Scube2 on the same (stripped) blot. The arrowhead indicates Shh specifically solubilized by Scube2 and Disp. (B–G) Reverse-phase high-performance liquid chromatography (RP-HPLC) of Shh variant standards. (B, C) Elution profiles of commercial, dual-lipidated R&D 8908-SH Shh (B) and of dual-lipidated Shh from Bosc23 cell lysates (C) are similar. Cartoons of Shh proteins indicate the presence or absence of N-terminal palmitate (P) and C-terminal cholesterol (C). (D) Bosc23-expressed non-lipidated control C25AShhN elutes in early fractions from the column. (E, F) The elution profile of monolipidated C25AShh (E) partially overlaps with that of dual-lipidated Shh, and the elution profile of palmitoylated ShhN (F) partially overlaps with that of C25AShhN. The increased hydrophobicity of cholesteroylated C25AShh over palmitoylated ShhN is consistent with the finding that cholesterol membrane anchoring strength is similar to dual lipidation motifs such as double geranylgeranylation or palmitoylation plus farnesylation in other lipidated proteins (Peters et al., 2004). (G) Merged profiles. Elution profiles are expressed relative to the highest protein amount in a given fraction (set to 100%). fr#: fraction numbers are colored according to Shh lipidation status in this figure and in other figures. (H) Co-expressed transgenic Ptch1, transgenic Ptch1ΔL2 lacking most of the second extracellular loop, and Disp-enhanced processed Shh release from Disp-/- cells into the media. Arrowheads indicate Shh specifically solubilized by Scube2 and Disp or Ptch1. Empty vector (EV)-transfected Disp-/- cells served as negative controls. (I, J) Actin served as a loading control for cell lysates and PonceauS as a loading control in media, and α-FLAG antibodies detected FLAG-tagged Scube2. (K–M) Quantification of the lower electrophoretic mobility band (the top band) of Shh (A, K), C25SShh (I, L), and ShhN (J, M) released from non-targeting control (nt ctrl) and Disp-/- cells. Amounts of solubilized and cellular proteins on immunoblots were determined and expressed as % increase or decrease relative to the respective cellular fraction, which was always set to 100%. ***p=0.0005, **p=0.0014, n.s.: p>0.05. (N) Actin served as a loading control for cell lysates and PonceauS as a loading control in media, and α-FLAG antibodies detected FLAG-tagged Scube2, as always on the same (stripped) blot.
-
Figure 1—figure supplement 2—source data 1
Uncropped western blots of Figure 1—figure supplement 2.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig1-figsupp2-data1-v1.zip
Figure 2 with 1 supplement
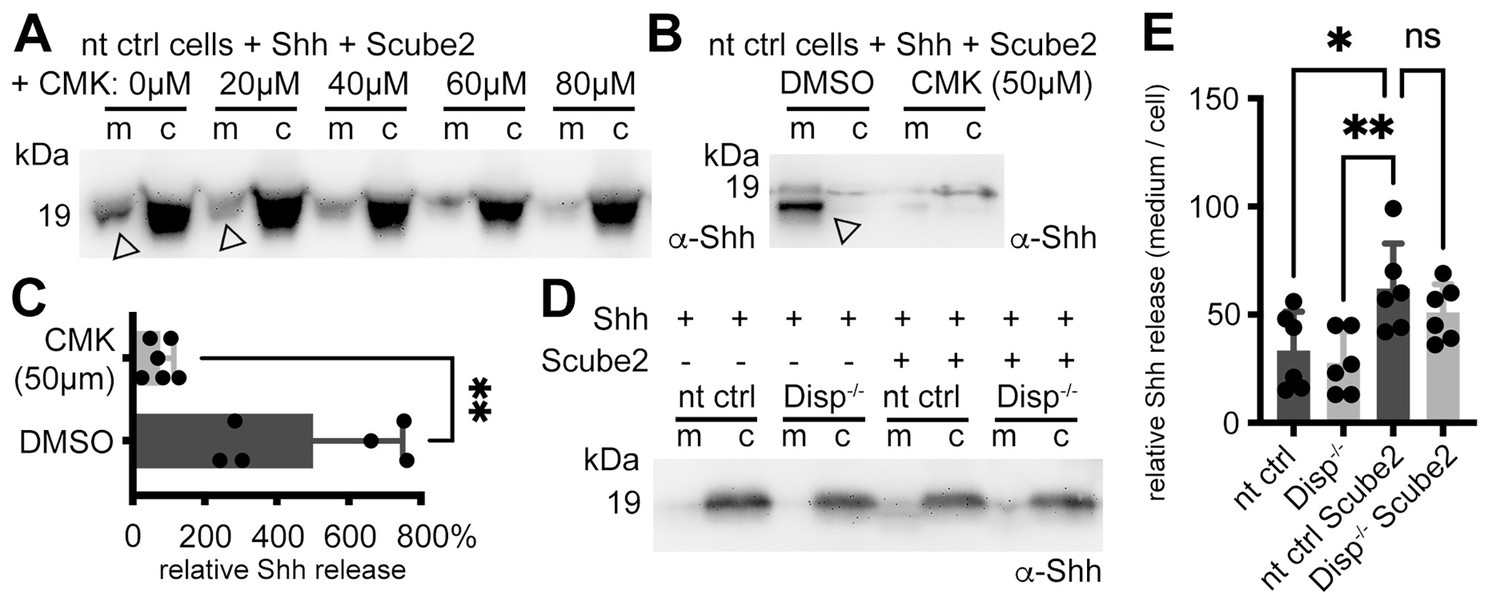
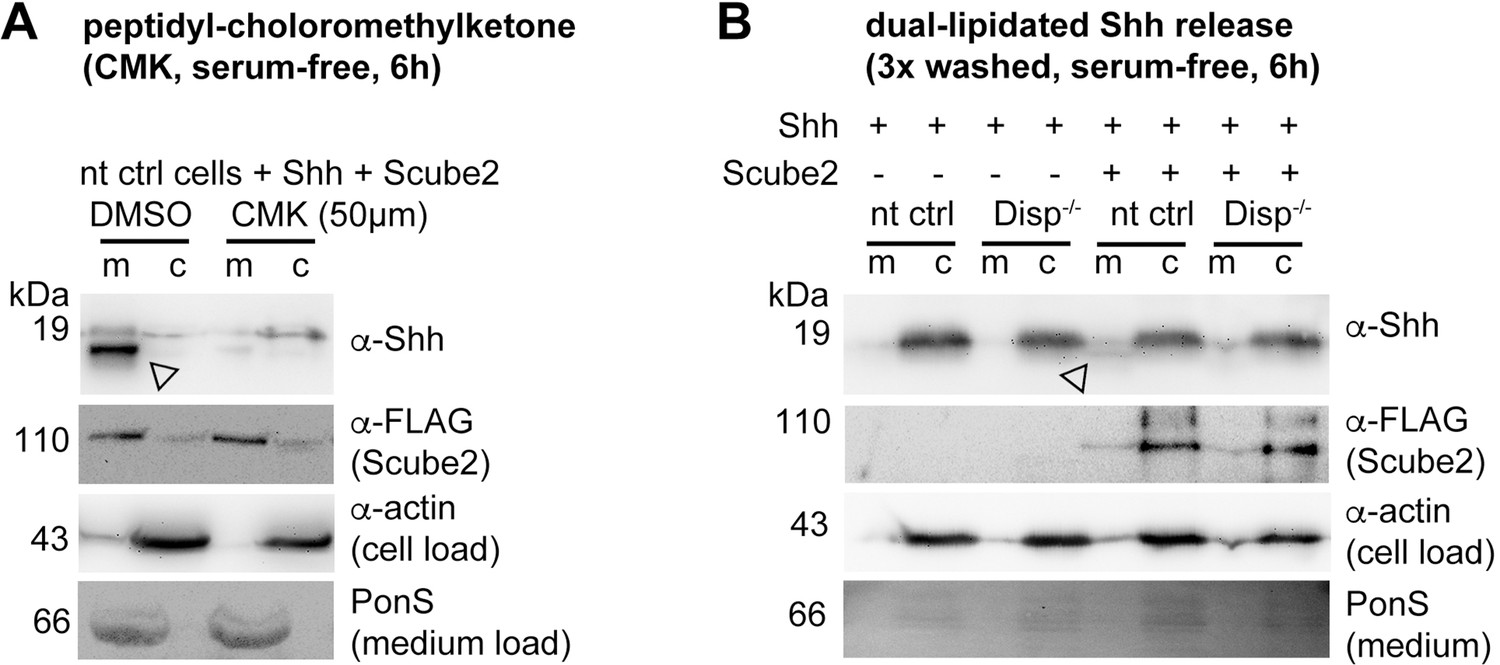
Shh shedding depends on Disp activation by furin and the presence of serum traces.
(A) Non-targeting control (nt ctrl) cells were transfected with Shh and Scube2, and Shh solubilization was monitored in the presence or absence of the furin inhibitor chloromethylketone (CMK). CMK impaired proteolytic processing and release of truncated (arrowheads) soluble Shh in a concentration-dependent manner. (B) Truncated (arrowhead) Shh release in the presence or absence of 50 μM CMK furin inhibitor. (C) Quantification of CMK-inhibited Shh shedding. Ratios of solubilized versus cellular Shh (set to 100%) in the presence of 50 μM CMK inhibitor were determined and expressed relative to Shh solubilization in the absence of inhibitor (DMSO). Unpaired t-test, two-tailed. **p=0.0021, n = 6. See Figure 2—figure supplement 1A for loading controls and Supplementary file 1 for additional statistical information. (D) Immunoblotted cellular (c) and medium (m) fractions of Shh expressing nt ctrl and Disp-/- cells in the complete absence of serum (referred to as ‘serum-free’ conditions throughout this article). Note that Shh solubilization is greatly reduced under serum-free conditions. (E) Processed Shh quantifications after secretion from nt ctrl and Disp-/- cells in the presence or absence of Scube2 into serum-free medium. One-way ANOVA, Dunnett’s multiple-comparisons test. **p=0.0059, *p=0.02, ns: p=0.54. See Supplementary file 1 for detailed statistical information.
-
Figure 2—source data 1
Raw data and statistical analyses of Figure 2.
A, B, D contain uncropped western blots of data shown in Figure 2A, B and D. C shows three additional representative examples of CMK-inhibited Shh release. E shows six biological replicates of impaired Shh release in the absence of serum that were quantified and displayed in Figure 2E. Prizm files C and E quantify relative Shh release rates based on the data shown in tiff-files B + C and E.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig2-data1-v1.zip
Figure 2—figure supplement 1
Loading controls.
(A, B) Actin, PonceauS, and FLAG-tagged Scube2 loading controls for (the same stripped) blots shown in Figure 2.
-
Figure 2—figure supplement 1—source data 1
Uncropped western blots of Figure 2—figure supplement 1.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig2-figsupp1-data1-v1.zip
Figure 3 with 2 supplements
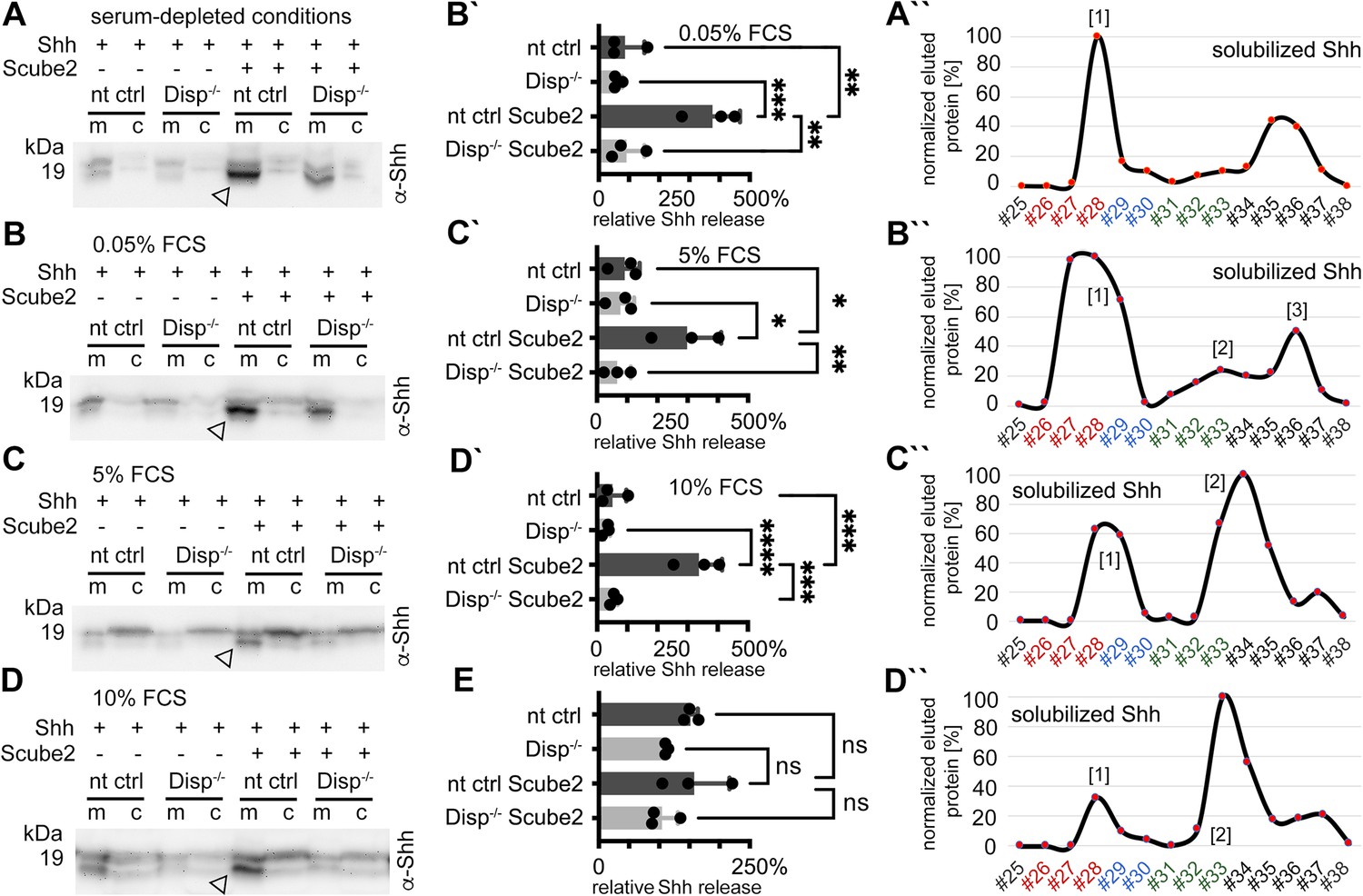
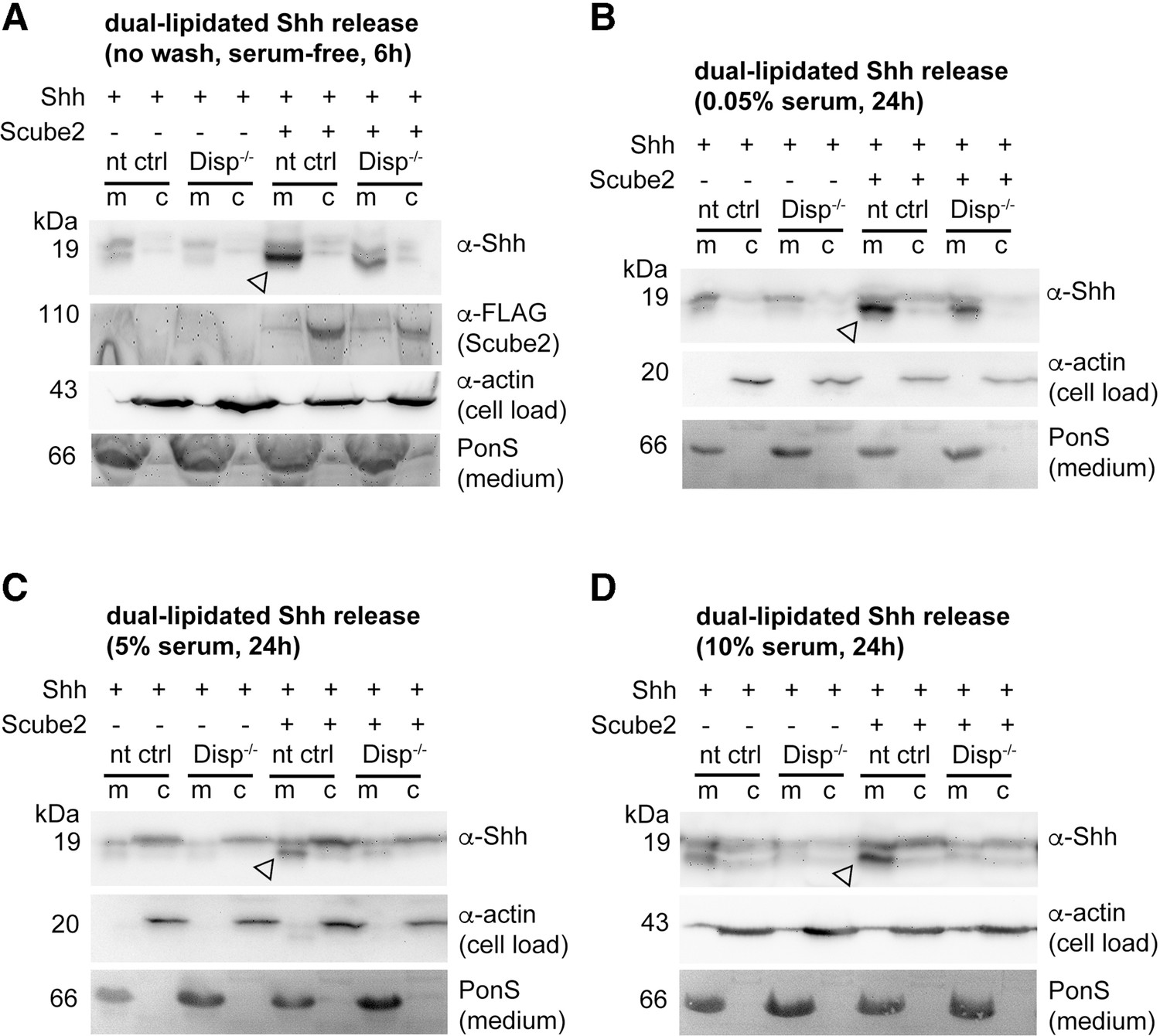
Dually lipidated cell surface Shh converts to delipidated soluble forms under low-serum and high-serum conditions.
Media were changed to serum-free Dulbecco’s Modified Eagle’s Medium (DMEM) (cells were not washed) or DMEM containing the indicated amounts of serum 36 hr post-transfection, and proteins were solubilized for 6 hr (serum-depleted) or for 24 hr (with 0.05, 5, and 10% serum). (A) Under serum-depleted conditions, Disp and Scube2 increase the conversion of dual-lipidated Shh into truncated soluble forms (arrowhead). m: media; c: cell lysate. (A’’) Reverse-phase high-performance liquid chromatography (RP-HPLC) confirmed the loss of both terminal lipidated Shh peptides during Disp- and Scube2-regulated shedding, as shown previously (Figure 1A’’). (B–D) The appearance of truncated Shh in serum-containing media remained dependent on Disp and, to a lesser extent, Scube2 (arrowheads). (B’–D’) Quantifications of (B–D). One-way ANOVA, Dunnett’s multiple-comparisons test. ****p<0.0001, ***p<0.001, **p<0.01, *p<0.05. See Supplementary file 1 for detailed statistical information. (B’’–D’’) RP-HPLC revealed that increased serum levels shift dual Shh shedding [1] toward N-terminally restricted shedding and release of a cholesteroylated Shh form [2]. Low levels of dually lipidated Shh are also detected [3]. (E) Quantification of unprocessed Shh release in the presence of 10% fetal calf serum (FCS) (top band in D). One-way ANOVA, Dunnett’s multiple-comparisons test. ns: p>0.05. See Figure 3—figure supplements 1 and 2 for additional information.
-
Figure 3—source data 1
Raw data and statistical analyses for Figure 3.
A–D contain uncropped western blots shown in Figure 3A–D. Prizm files contain all raw data and statistical analysis to quantify serum-dependent Shh release. B’–E contain uncropped western blots used for the quantification. D` quantifies truncated (proteolytically processed) solubilized Shh, E quantifies relative amounts of unprocessed Shh in media. A’–D’’ Excel file containing raw Shh RP-HPLC elution data as shown in Figure 3A’’–D’’.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig3-data1-v1.zip
Figure 3—figure supplement 1
Loading controls.
(A–D) Actin, PonceauS, and FLAG-tagged Scube2 loading controls for blots shown in Figure 3. Arrowheads indicate Shh specifically solubilized by Disp and Scube2.
-
Figure 3—figure supplement 1—source data 1
Uncropped western blots for Figure 3—figure supplement 1.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig3-figsupp1-data1-v1.zip
Figure 3—figure supplement 2
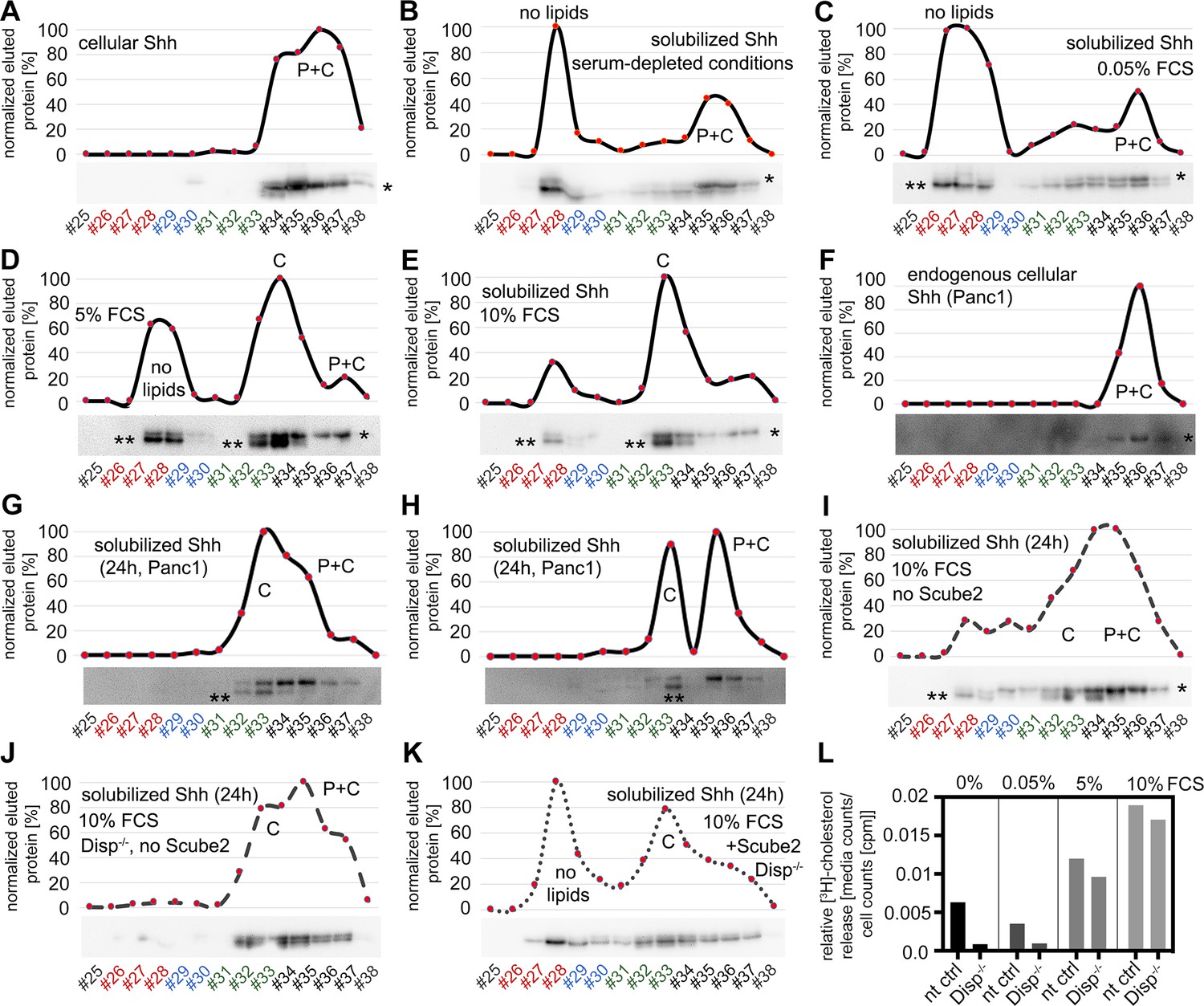
Source data and reverse-phase high-performance liquid chromatography (RP-HPLC) profiles.
(A) Dual-lipidated control Shh in cell lysates (‘top’ band, indicated by *). (B–E) Serum modulates the conversion of dual-lipidated Shh into variably lipidated soluble forms. Shh was enriched by heparin-agarose pulldown before analysis. Bottom bands (** in the blot image) represent processed proteins derived from the dually delipidated Shh precursors (corresponding to the non-lipidated control C25AShhN, Figure 1—figure supplement 2D) or the cholesteroylated N-truncated form (fractions #32–34). Variable amounts of dual-lipidated proteins are also detected (top-right bands [*], fractions #35–37). P: bound palmitate; C: bound cholesterol; P+C: dual-lipidated proteins. (F–H) Serum modulates the conversion of endogenous dual-lipidated Shh (F) into mono-lipidated soluble forms (lower bands, marked with ** in the blot images) in two independent analyses (G, H). (I–K) Increased relative amounts of dual-lipidated Shh released under physiologically irrelevant conditions. Proteins were released for 24 hr in Dulbecco’s Modified Eagle’s Medium (DMEM) +10% fetal calf serum (FCS) to compensate for impaired Shh solubilizations due to the absence of Scube2 and Disp. (I) Lack of Scube2 in Disp-expressing cells decreases delipidated Shh solubilization (**) and monolipidated Shh release into the medium and increases the relative amounts of dual-lipidated Shh (*). (J) Scube2 and Disp deficiency reduces delipidated and monolipidated Shh release, making dual-lipidated Shh the major soluble fraction detectable after 24 hr release. (K) Scube2 restores some delipidated Shh release from Disp-/- cells. (L) Bosc23 cells were loaded with [3H]-cholesterol for 48 hr and washed, and cholesterol release was measured for 3 hr into serum-depleted media or medium with 0.05, 5, or 10% FCS. [3H]-cholesterol signals in media were normalized with cellular counts. Note that cholesterol export from Disp-/- cells was always reduced if compared to non-targeting control (nt ctrl) cells under the same experimental conditions. n = 2 for all measurements.
-
Figure 3—figure supplement 2—source data 1
Uncropped western blots for Figure 3—figure supplement 2.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig3-figsupp2-data1-v1.zip
Figure 4 with 1 supplement
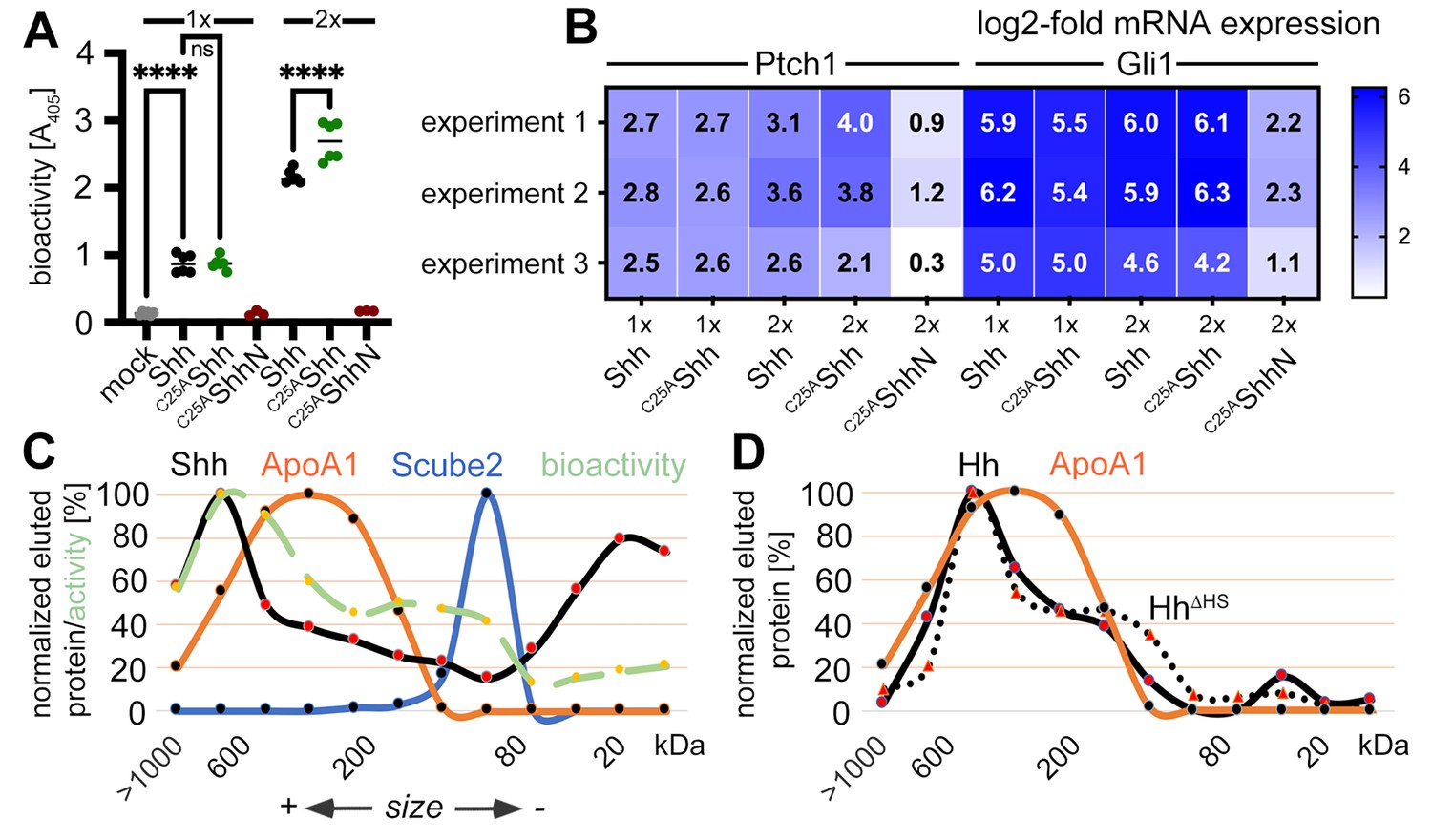
Activities and size-exclusion chromatography (SEC) of dual-lipidated Shh/Hh and depalmitoylated Shh variants solubilized into serum-containing media.
(A) Shh, C25AShh (this artificial variant has the cysteine palmitate acceptor changed to a non-accepting alanine), and non-lipidated C25AShhN were expressed in media containing 10% fetal calf serum (FCS); their protein levels were determined by immunoblotting and normalized; and the conditioned media were added to C3H10T1/2 reporter cells to induce their Hh-dependent differentiation into alkaline phosphatase (Alp)-producing osteoblasts. Mock-treated C3H10T1/2 cells served as non-differentiating controls. At lower concentrations (1×), Shh and C25AShh induced C3H10T1/2 differentiation in a similar manner, as determined by Alp activity measured at 405 nm. At higher concentrations (2×), the bioactivity of C25AShh was increased over that of Shh. C25AShhN was inactive. One-way ANOVA, Sidak’s multiple-comparisons test. ****p<0.0001, ns = 0.99, n = 3–9. See Supplementary file 1 for additional statistical information. (B) Similar transcription of the Hh target genes Ptch1 and Gli1 by Shh and C25AShh in three independent experiments. C3H10T1/2 reporter cells were stimulated with similar amounts of Shh, C25AShh, and C25AShhN (Figure 4—figure supplement 1E) at high (2×) and low (1×) concentrations. (C) SEC shows significant amounts of Shh of increased molecular weight in media containing 10% serum (black line). The increased molecular weight Shh eluted together with ApoA1 (orange line). In contrast, Scube2 was largely monomeric in solution (blue line). The level of Shh-induced Alp activity in C3H10T1/2 cells was measured as absorbance at 405 nm, showing the strongest C3H10T1/2 differentiation by eluted fractions containing large Shh aggregates. (D) SEC of Drosophila Hh (black line) and of a variant lacking its HS binding site (HhΔHS, black dotted line). Both proteins were expressed from S2 insect cells under actin-Gal4/UAS-control and solubilized into media containing 10% FCS.
-
Figure 4—source data 1
Raw data of Figure 4.
A, B contains files demonstrating that palmitoylated Shh and non-palmitoylated C25SShh are both biologically active. C, D Excel file containing raw Shh gel filtration and bioactivity data (as shown in Excel file C, a duplicate bioactivity analysis of eluted fractions is shown) as shown in Figure 4C. D shows an uncropped western blot of Drosophila Hh expressed from S2 cells into media containing 10%FCS subsequent gel filtration analysis. Hh proteins are marked. C, D Quantification of Hh detected in gel filtration analysis.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig4-data1-v1.zip
Figure 4—figure supplement 1
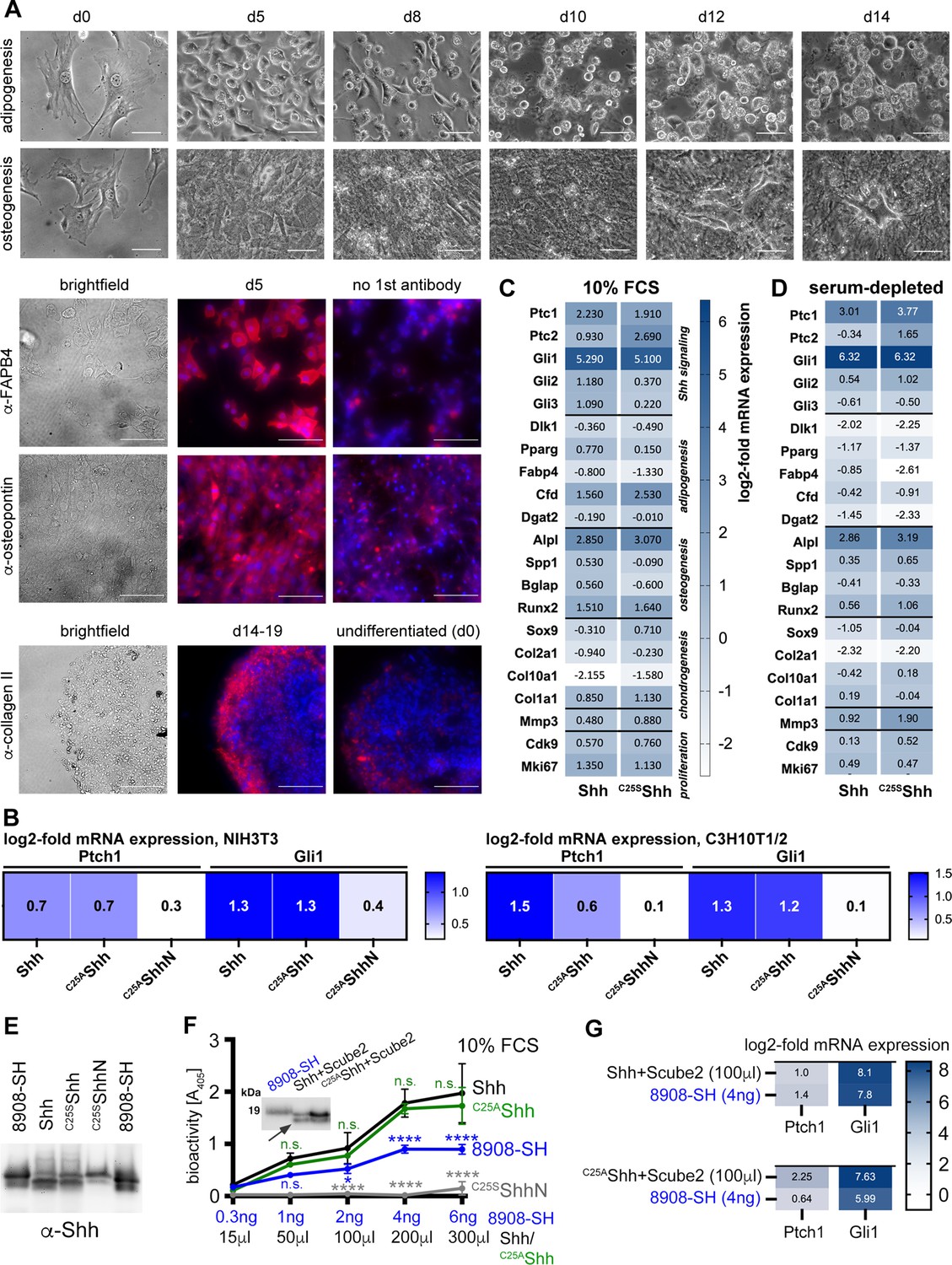
Confirmed multipotency of C3H10T1/2 cells and similar activities of palmitoylated and non-palmitoylated Shh released into serum containing media.
(A) C3H10T1/2 cells were cultured for the indicated time periods and their specific differentiation into adipocytes, osteoblasts, or chondrocytes was induced. Differentiation was confirmed by morphology (top) in combination with lipid droplet staining with OilRed to confirm adipogenesis (not shown), alizarin red staining to identify Ca2+-containing osteocytes (not shown), by anti-FABP-4 antibody binding that confirms adipogenic differentiation l, anti-collagen II antibody binding to confirm chondrogenic differentiation, and anti-osteopontin antibody binding to confirm osteogenesis. Cell nuclei were counterstained with DAPI (blue). Scale bars: 100 μm. (B) N-processed soluble Shh and non-palmitoylated C25AShh induced similar increases in Ptch1 and Gli1 mRNA expression in stimulated NIH3T3 cells and C3H10T1/2 osteoblast progenitor cells. Non-lipidated C25AShhN served as an inactive control. Gene expression was normalized to mock-treated cells. (C, D) N-processed Shh and partially N-processed C25SShh induced similar gene expression profiles when released into 10% fetal calf serum (FCS)-containing Dulbecco’s Modified Eagle’s Medium (DMEM) (C) or into serum-depleted DMEM supplemented with 300 μg/mL MβCD (D). MβCD served as an alternative chaperone for the Shh C-cholesterol to show that associated serum factors (such as lipoprotein particles [LPPs]) do not alter the Shh activity profiles. See Supplementary files 2 and 3 for qPCR target gene information. (E) Normalization of soluble proteins used for the assays shown in Figure 4B. (F, G) Activities of dual-lipidated Shh and depalmitoylated Shh variants solubilized into serum-containing media. (F) Shh, C25AShh and C25SShhN were solubilized from Scube2- and Disp expressing cells into media containing 10% FCS. Inset: immunoblotted proteins were used for quantification and visualization of dually lipidated R&D 8908-SH, Shh, C25AShh, and unlipidated C25SShhN. The arrow indicates proteolytic processing of the palmitoylated N-terminal peptide from Shh and C25AShh, but not from R&D 8908-SH. Graph: 0–4 ng of R&D 8908-SH dissolved into DMEM + 10% FCS induced C3H10T1/2 reporter differentiation in a concentration-dependent manner, as determined by Alp activity measured at 405 nm (blue line). Similar amounts of solubilized Shh, as determined by immunoblotting, increased C3H10T1/2 osteogenesis over that of the dual-lipidated R&D 8908-SH standard (black line). C25AShh showed similarly high bioactivity after release into serum-containing medium (green line). Artificial unlipidated soluble C25SShhN showed low activity (gray line). Data were normalized to mock-treated C3H10T1/2 cells. One-way ANOVA, Sidak’s multiple-comparisons test. ****p>0.0001, *p=0.03, n.s.: p>0.05. (G) Similar induction of transcription of the Hh target genes Ptch1 and Gli1 in C3H10T1/2 reporter cells. Cells were stimulated with 4 ng R&D 8908-SH or similar amounts of Shh and C25AShh. Shh and C25AShh were both solubilized from Bosc23 cells into serum containing media in the presence of Scube2.
-
Figure 4—figure supplement 1—source data 1
Raw data for bioassays shown in Figure 4—figure supplement 1.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig4-figsupp1-data1-v1.zip
Figure 5 with 1 supplement
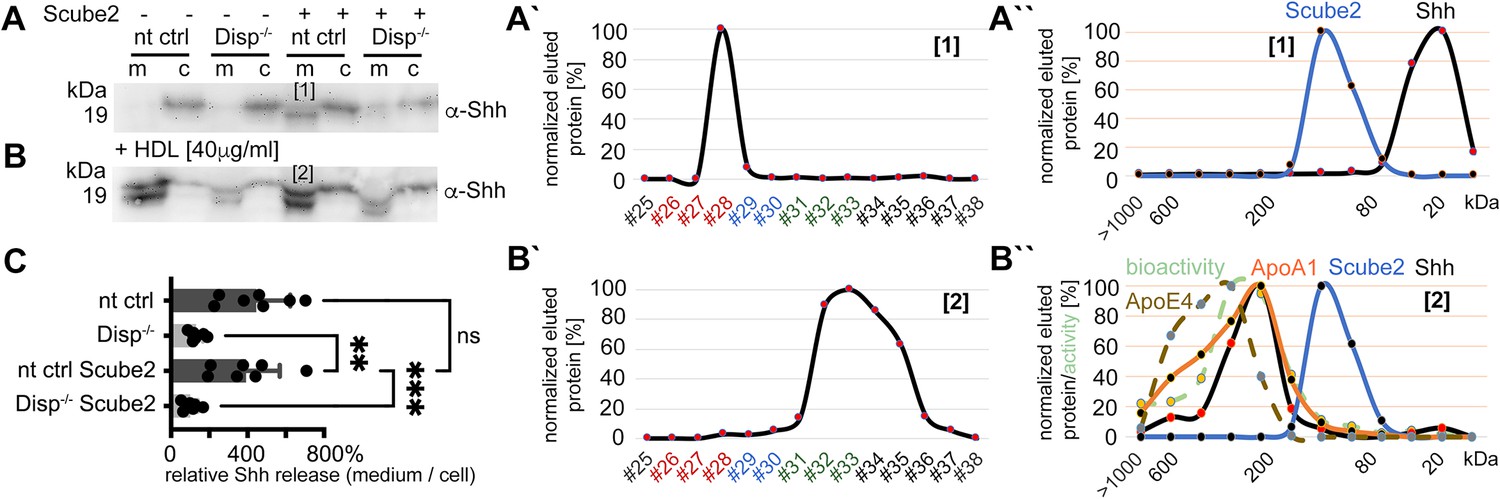
High-density lipoprotein (HDL) enhancess N-processed Shh solubilization by Disp.
Media were changed to serum-free Dulbecco’s Modified Eagle’s Medium (DMEM) (and cells washed three times) or serum-free DMEM supplemented with 40 µg/mL HDL 36 hr post-transfection before protein solubilization for 6 hr. (A) Immunoblotted cellular (c) and medium (m) fractions of Shh expressing nt ctrl and Disp-/- cells in the complete absence of serum. Note that only little Shh is released under this condition [1]. (A’) Reverse-phase high-performance liquid chromatography (RP-HPLC) of the material labeled [1] in (A) showed complete delipidation during release. (A’’) Size-exclusion chromatography (SEC) of the same delipidated material shows that it is readily soluble and not associated with Scube2. (B) Immunoblotted cellular (c) and medium (m) fractions of Shh expressing nt ctrl and Disp-/- cells in the presence of 40 μg/mL HDL. Shh shedding and solubilization are strongly increased by Disp [2] but not by Scube2. (B’) RP-HPLC of the material labeled [2] in (B) showed that HDL shifts Shh shedding from dual processing (A’, [1]) to release of cholesteroylated Shh. (B’’) SEC of the same material [2] (black line) shows an increase in molecular weight corresponding to the molecular weight range of HDL, as indicated by the marker apolipoproteins ApoA1 (orange line) and mobile ApoE4. The former provides structural stability to the particle and stimulates cholesterol efflux to HDL; the latter facilitates cholesterol storage and core expansion and is therefore a marker of larger mature HDL particles (brown dashed line). Again, the soluble Shh elution profile does not overlap with that of Scube2 (blue line). (C) Quantification of HDL-induced Shh solubilization from nt ctrl cells and Disp-/- cells. One-way ANOVA, Dunnett’s multiple-comparisons test. ***p=0.0008, **p=0.0023, ns: p=0.77, n = 7. Additional statistical information is provided in Supplementary file 1.
-
Figure 5—source data 1
Raw data of Figure 5.
A, B contain uncropped western blots shown in Figure 5A and B. C contains uncropped western blots of seven biological replicates showing that HDL presence renders Scube2 function obsolete. Prizm file C quantifies similar relative Shh release from nt Ctrl cells in the presence of HDL, irrespective of Scube2 presence or absence (as shown in Figure 5C). The Excel file contains raw data of Figure 5A’, A’’, B’ and B’’.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig5-data1-v1.zip
Figure 5—figure supplement 1
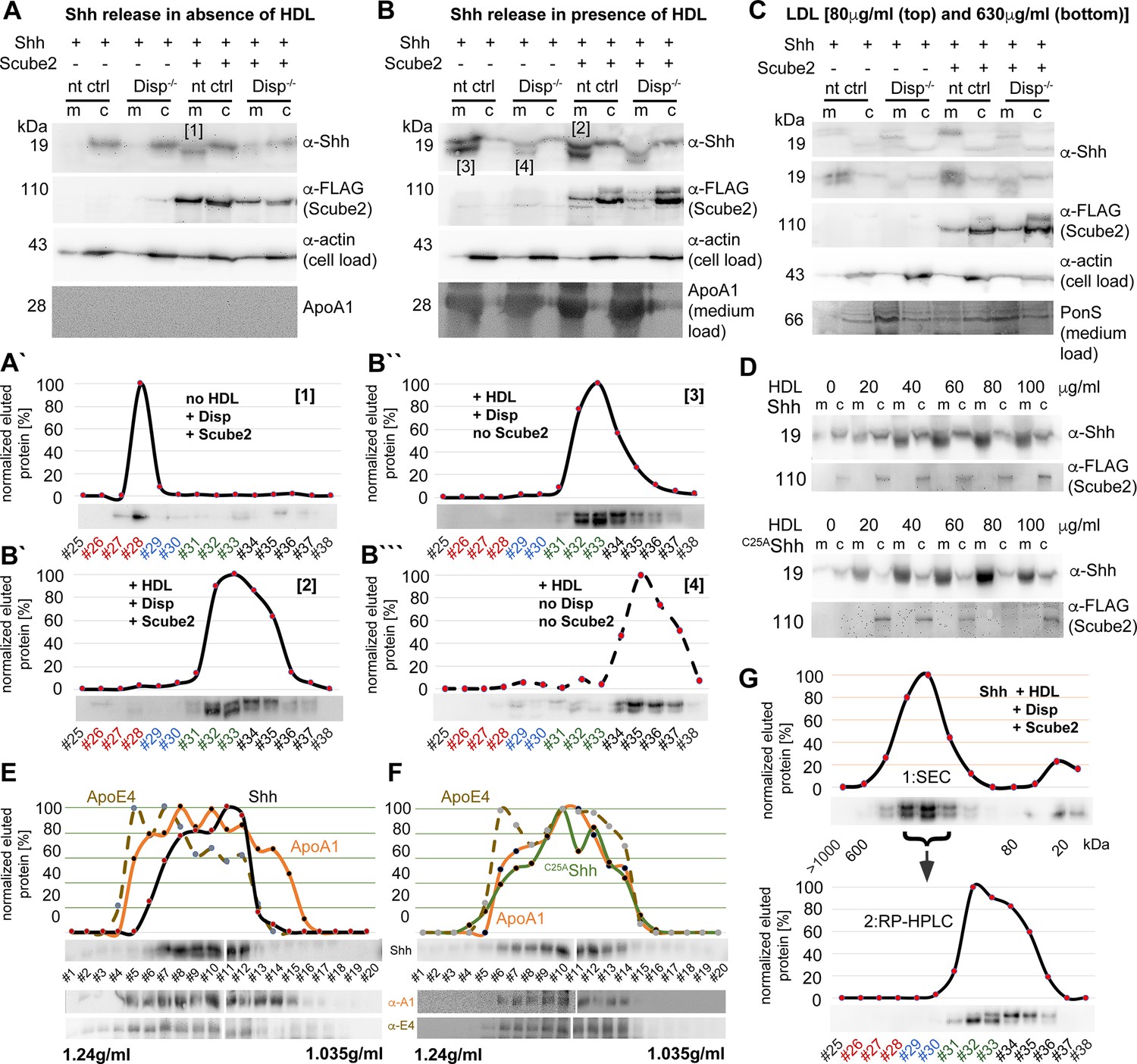
Loading controls.
(A, B) Actin and FLAG-tagged Scube2 loading controls for the blots shown in Figure 2. No residual BSA was detected in these media as a result of repeated careful washing of the cells prior to the addition of Dulbecco’s Modified Eagle’s Medium (DMEM) + high-density lipoprotein (HDL). ApoA1 was therefore used as the loading control. (A’–B’’’) Reverse-phase high-performance liquid chromatography (RP-HPLC) profiles and source data of aliquots of samples [1-4] as shown in (A, B). Note that Disp-/- cells release high relative amounts of dual-lipidated Shh in the absence of Scube2 (B’’’). This suggests that the transfer of dual-lipidated Shh to HDL is unlikely to be physiologically relevant. In contrast, Disp and Scube2 specifically enhance the release of Shh in delipidated (A’) or monolipidated (B’, B’’) form, depending on the presence or absence of HDL. (C) Low-density lipoprotein (LDL) does not increase Shh solubilization from Disp-expressing cells. Transfection and loading controls are shown for the stripped top blot (80 μg/mL LDL). (D) In contrast, HDL increases the release of Shh and of cholesteroylated, non-palmitoylated C25AShh in a concentration-dependent manner. (E, F) Shh (E) and C25AShh (F) were transfected into Disp-expressing Bosc23 cells and released overnight in the presence of 40 mg/mL HDL in 1.2 mL DMEM. Density gradient centrifugation of the solubilized material supports the physical interaction of both proteins with ApoA1 and ApoE4-bearing HDL of known corresponding density (1.1–1.2 g/mL). (G) Shh released from transfected Disp-expressing cells was released in the presence of HDL and subjected to size-exclusion chromatography (SEC). High molecular weight SEC fractions were subsequently analyzed by RP-HPLC, demonstrating that all HDL-associated proteins are cholesterylated.
-
Figure 5—figure supplement 1—source data 1
Raw data of Figure 5—figure supplement 1.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig5-figsupp1-data1-v1.zip
Figure 6
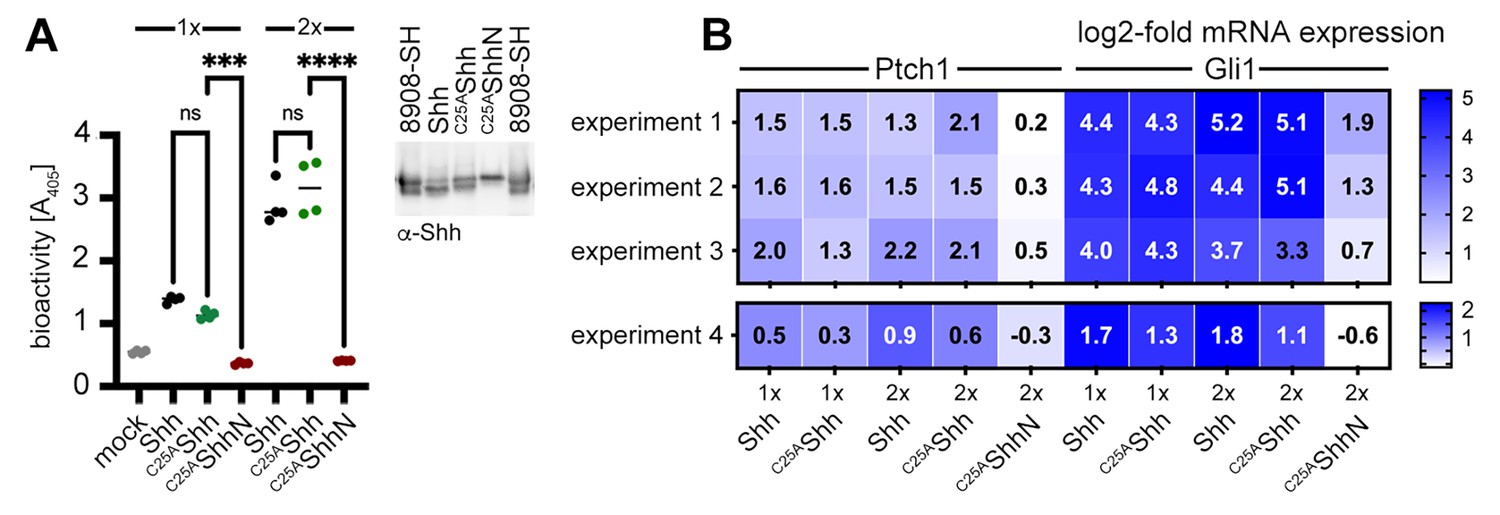
Activities of high-density lipoprotein (HDL)-associated Shh and non-palmitoylated variants.
(A) Shh, non-palmitoylated C25AShh, and non-lipidated C25AShhN were released into media containing 80 µg/mL HDL, their protein levels were determined by immunoblotting and normalized (inset), and conditioned media were added to C3H10T1/2 reporter cells to induce their differentiation into Alp-producing osteoblasts. Mock-treated C3H10T1/2 cells served as non-differentiating controls. At low (1×) and high (2×) concentrations, Shh and C25AShh induced C3H10T1/2 differentiation in a similar manner, as determined by Alp activity measured at 405 nm. Again, C25AShhN was completely inactive, in contrast to the bioactive HDL-associated non-palmitoylated C25SShh. One-way ANOVA, Sidak’s multiple-comparisons test. ****p<0.0001, ***p<0.001, ns > 0.1, n = 4. Additional statistical information is provided in Supplementary file 1. (B) Similar transcription of Hh target genes Ptch1 and Gli1 by HDL-associated Shh and C25AShh in C3H10T1/2 cells three independent experiments. Experiment 4 confirms similar Shh and C25AShh activities in NIH3T3 cells. Reporter cells were stimulated with similar amounts of Shh, C25AShh, and C25AShhN at high (2×) and low (1×) concentrations as determined by immunoblotting (A, inset).
-
Figure 6—source data 1
Statistical analyses of Figure 6.
A, B contain files demonstrating that palmitoylated Shh and non-palmitoylated C25SShh are both biologically active in C3H10T1/2 cells and NIH3T3 cells when released in HDL presence.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig6-data1-v1.zip
Figure 7 with 1 supplement
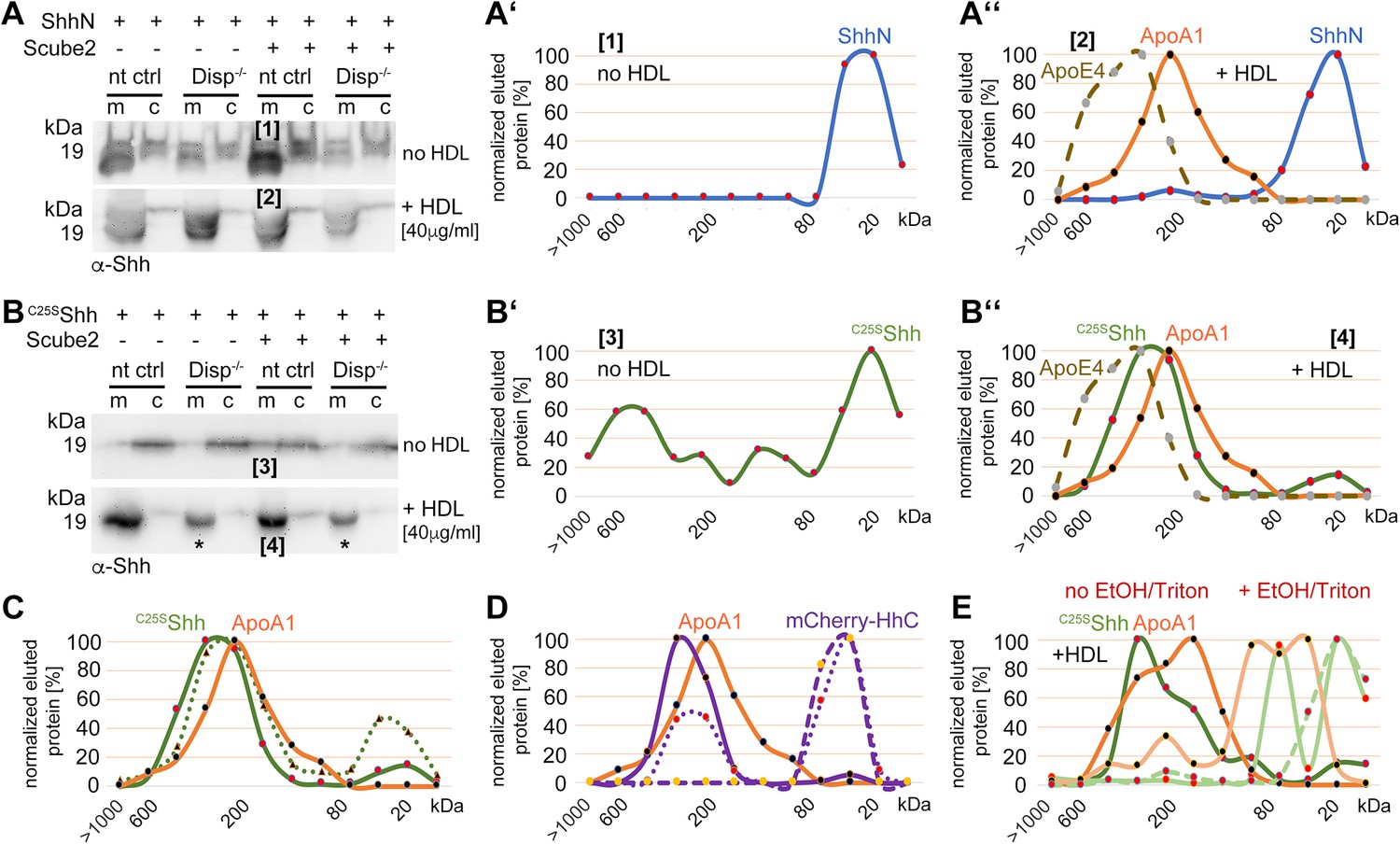
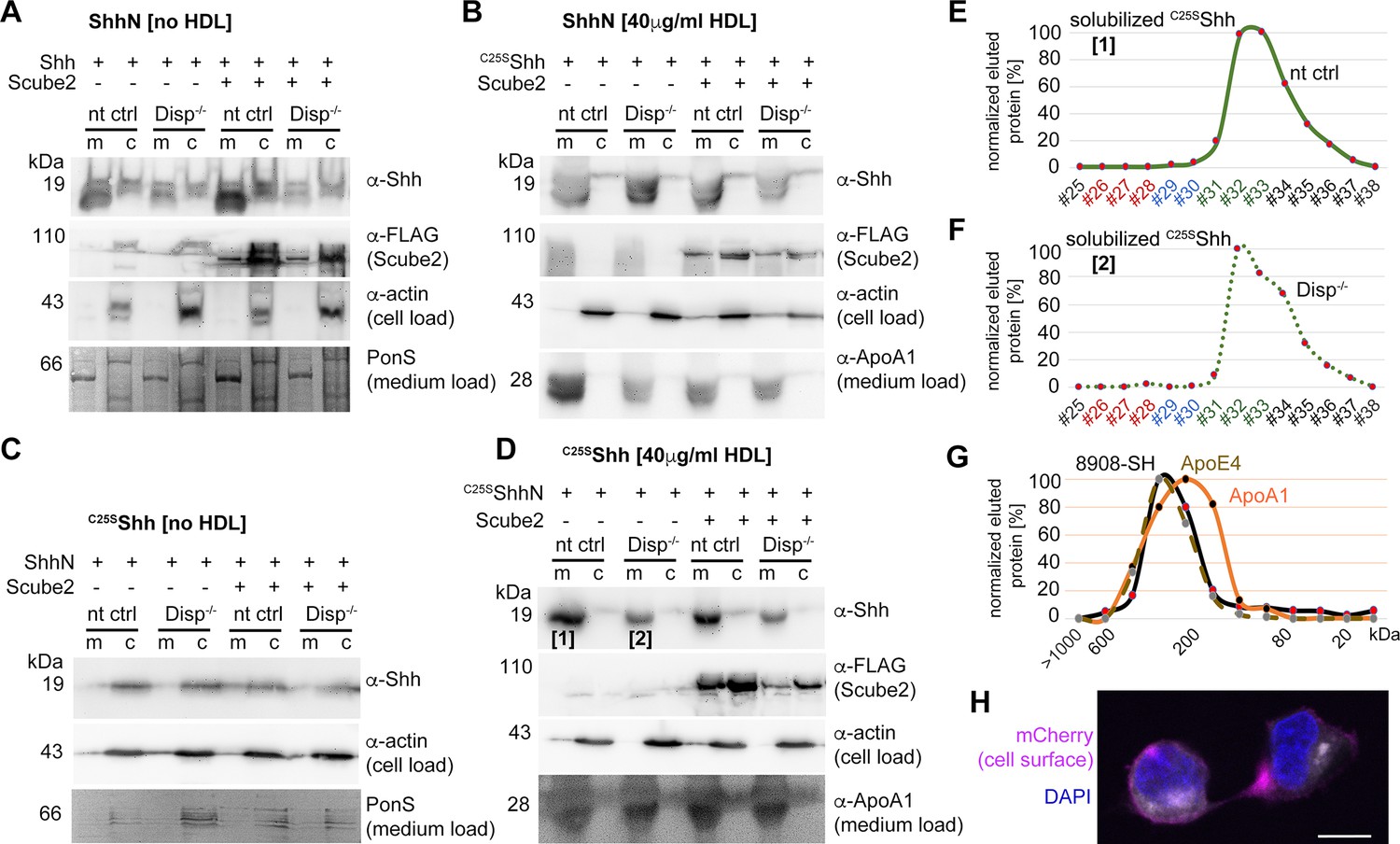
Cholesterylated C-terminal peptides are necessary and sufficient for Disp-mediated protein transfer to high-density lipoprotein (HDL).
(A) Immunoblotted cellular (c) and medium (m) fractions of ShhN-expressing nt ctrl and Disp-/- cells. Shown is unspecific ShhN solubilization into serum-free media (upper blot, labeled [1]) or into serum-free Dulbecco’s Modified Eagle’s Medium (DMEM) supplemented with HDL (lower blot, labeled [2]). (A’) ShhN [1] expressed under serum-free conditions is solubilized in a monomeric state. (A’’) ShhN [2] expressed in the presence of HDL remained monomeric (i.e., not HDL-associated). (B) Release of cholesterylated C25SShh into serum-free medium is very low (upper blot, labeled [3]), but increases in a Disp-dependent manner in the presence of HDL (lower blot, labeled [4]). Asterisks denote C25SShh solubilized independently of Disp function. (B’) Most C25SShh [3] in serum-free media is monomeric. (B’’) C25SShh [4] expressed in the presence of HDL increases in molecular weight to match the molecular weight range of HDL (orange line, dotted brown line). (C) Size-exclusion chromatography (SEC) of C25SShh solubilized from Disp-expressing cells (solid green line) or from Disp-/- cells (dotted line) shows Disp-independent physical desorption and unregulated HDL association of the monolipidated protein. (D) SEC of cholesteroylated mCherry solubilized from nt ctrl cells (solid lines) or from Disp-/- cells (dotted lines) under the same conditions. Dashed lines indicate proteins solubilized under serum-free conditions. Note that most of the mCherry associates with HDL in a Disp-mediated manner. (E) C25SShh (green line) dissociates from HDL in 50% ethanol (bright green line) or in 0.1% Triton X-100 (bright green dashed line). The disassembly of HDL (orange line) under the same conditions is confirmed by the size shift of ApoA1 toward the monomeric 32 kDa protein (light orange line).
-
Figure 7—source data 1
Raw data of Figure 7.
A, B contain uncropped western blots shown in Figure 7A and B. The Excel file contains raw data used in Figure 7A’–E.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig7-data1-v1.zip
Figure 7—figure supplement 1
Loading controls.
(A–D) Actin, PonceauS, and FLAG-tagged Scube2 loading controls for the blots shown in Figure 6A and B. (E, F) Reverse-phase high-performance liquid chromatography (RP-HPLC) profiles of samples [1] and [2] shown in (D). Note that proteins released from non-targeting control (nt ctrl) cells and Disp-/- cells are both cholesterylated. This suggests that overexpressed monolipidated Shh is only loosely associated with the plasma membrane and can ‘leak out’, or desorb, and thereby associate with high-density lipoprotein (HDL) in a non-enzymatic manner. (G) Purified commercial 8908-SH Shh and HDL were mixed, incubated for 10 min, and analyzed by size-exclusion chromatography (SEC). This revealed rapid spontaneous association and explains the background desorption of dual-lipidated Shh in our cellular assays. Note the excellent 8908-SH Shh co-elution with the ApoE4-bearing HDL fraction, suggesting that this mobile apolipoprotein may have facilitated spontaneous 8908-SH Shh association, or that other properties of ApoE-bearing ‘late’ HDL (Sacks and Jensen, 2018) somehow facilitate Shh association. (H) Dual-lipidated mCherry is efficiently secreted to the outer plasma membrane leaflet of producing Bosc23 cells. Intracellular mCherry fluorescence is shown in white, α-mCherry antibody binding to the surface of non-permeabilized cells is shown in magenta, and DAPI staining of the nucleus is shown in blue. Scale bar: 10 μm.
-
Figure 7—figure supplement 1—source data 1
Uncropped western blots of Figure 7—figure supplement 1.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig7-figsupp1-data1-v1.zip
Figure 8 with 1 supplement
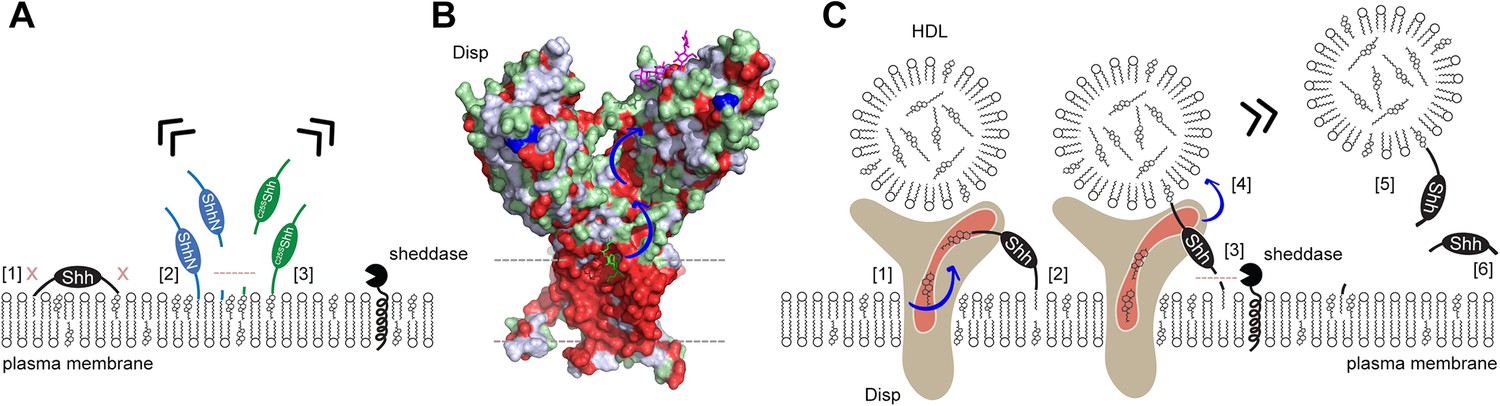
Model of two-way Disp-mediated Shh solubilization.
(A) Dual lipidation protects Shh from unregulated cell surface shedding by tight plasma membrane association of both lipids (blocked shedding is indicated by an x in [1]). In contrast, monolipidated ShhN [2] and C25SShh [3] are prone to unregulated membrane proximal shedding (indicated by the dashed line) or non-enzymatic desorption. (B) Depiction of the surface hydrophobicity of Disp (pdb:7RPH; Wang et al., 2021) suggests an extended hydrophobic surface channel (hydrophobic residues are shown in red) that may function as a ‘slide’ for lipophiles extending from the plasma membrane (dashed lines) to a cavity of the second extracellular domain (blue arrows). A sterol lifted upward (green stick representation) at the starting point of the hydrophobic track may represent an intermediate state of sterol extraction from the membrane, and a lipidic group modeled as the amphiphilic detergent lauryl maltose neopentyl glycol (violet stick structure) may represent the end point of the transfer (Wang et al., 2021) prior to delivery to high-density lipoprotein (HDL). (C) We propose two sequences of Shh transfer events. In the first event [1], plasma membrane sterol is transferred through the hydrophobic Disp surface channel to HDL acceptors. This process is similar to the established reverse cholesterol transport. In the second event, if present, C-terminal cholesterol moieties of Shh can also be transferred [2]. This partial Shh extraction exposes the N-terminal cleavage site [3] and makes it susceptible to proteolytic processing (similar to ShhN as shown in A). N-terminal Shh shedding can then release the protein from the plasma membrane [4] to complete the transfer [5]. In addition to, or competing with this process, cholesterol depletion of the plasma membrane (representing the first event, [1]) may indirectly trigger shedding of both terminal Shh peptides and the solubilization of monomeric proteins [6], possibly as a consequence of the disruption of lipid rafts. See ‘Discussion’ for details.
Figure 8—figure supplement 1
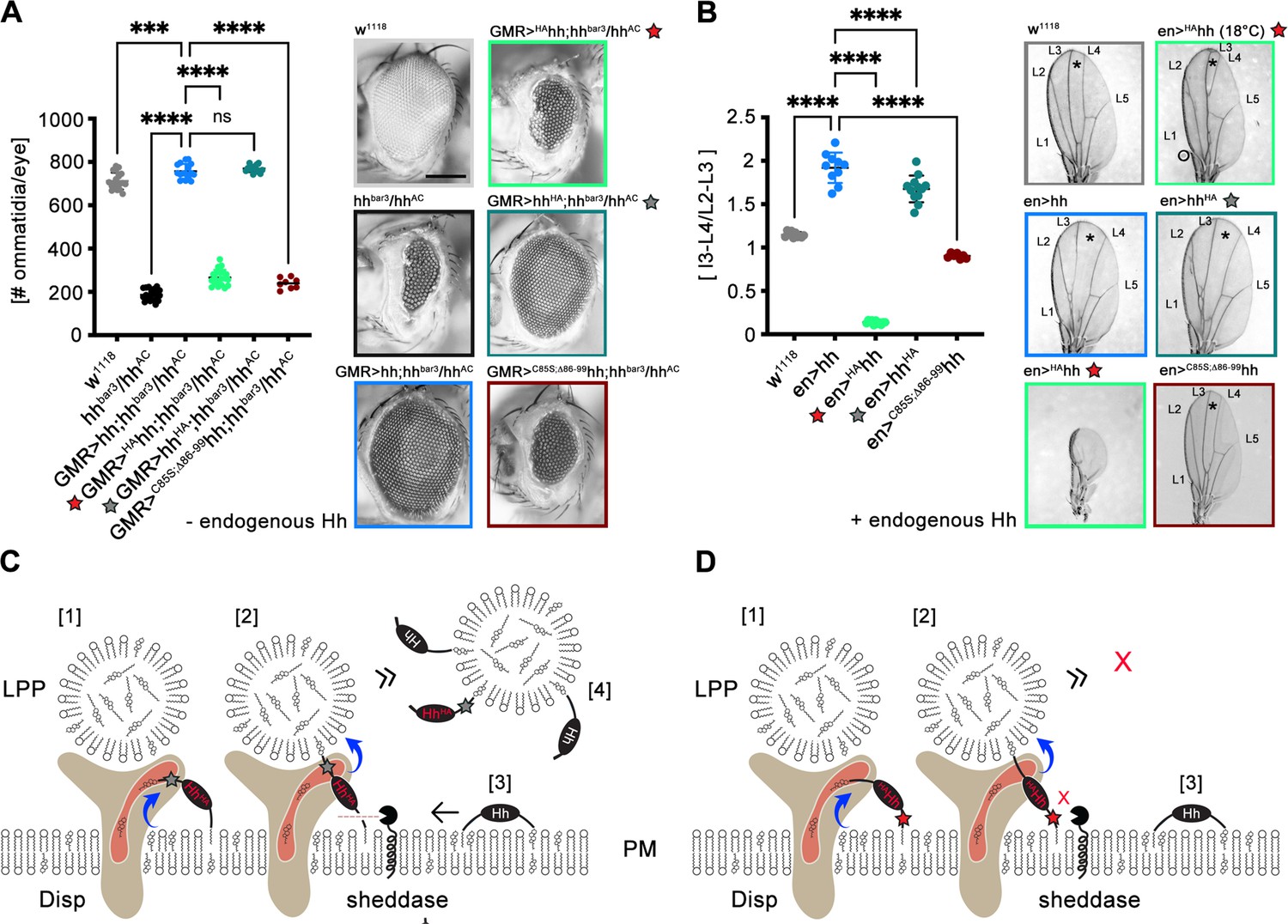
In vivo support for N-terminal shedding during Disp-mediated Hh export.
Experiments were independently repeated from established fly stocks and from published protocols to demonstrate that N-terminal, but not C-terminal proteolytic Hh processing is required during release (Schürmann et al., 2018; Kastl et al., 2018; Manikowski et al., 2020). (A) Wild-type flies (w1118) were used as positive controls. Eye discs lacking most Hh expression (in an hhbar3/hhAC background) develop into small eyes, and eye-disc-specific GMR-GAL4-controlled hh or hhHA (here, a protease-resistant hemagglutinin tag was inserted into the cholesteroylated C-terminal Hh peptide, gray star) restore eye development in the hhbar3/hhAC background. In contrast, GMR-GAL4-controlled HAhh expression (where the hemagglutinin tag was inserted into the N-palmitoylated Hh peptide to block sheddase function, red star) in the same background does not restore eye development. A non-palmitoylated N-truncated control variant (GMR>C85S;ΔN86-99hh) is also inactive. Left: quantification of phenotypes, right: representative examples of adult compound eyes. Scale bar: 100 μm. One-way ANOVA, Dunnett’s multiple-comparisons test. ****p<0.0001, ***p<0.001, n.s.: not significant (p>0.05). (B) The same Hh transgenes were produced in the posterior compartment of the wing disc pouch at 25°C under the control of en-GAL4. Cells in this compartment also produce endogenous Hh under the control of en, resulting in the simultaneous presence of endogenous and transgenic Hh at the cell surface. In adult wild-type flies (w1118) expressing only endogenous Hh, the wing disc develops into a wing blade with five longitudinal veins (L1-5) (Hartl and Scott, 2014). The central L3-L4 intervein tissue (asterisk) is derived from cells that receive the highest dose of Hh and thus serves as a readout for Hh function in the developing disc. As a consequence, en-GAL4-controlled overexpression of hh or hhHA leads to an expansion of the L3-L4 intervein area. In striking contrast, wing formation is abolished by overexpression of transgenic HAhh at 25°C under the control of en-GAL4, despite continued expression of endogenous hh. Reduced Gal4 promotor function at 18°C largely restores wing development with the exception of the most Hh-dependent L3-L4 domain. This indicates that overexpressed HAhh, but not hhHA, dominant-negatively inhibits endogenous Hh signaling in the developing wing, most likely by blocking N-terminal shedding. The inactive control mutant C85S;ΔN86-99hh does not suppress endogenous Hh function. One-way ANOVA, Dunnett’s multiple-comparisons test. ****p<0.0001. See Supplementary file 1 for source data. (C) We suggest that C-terminal protein tags (indicated by the gray star) do not interfere with Disp- and cholesterol-dependent protein extraction [1] and transfer [2] to lipoprotein particles (LPPs). As a consequence, extraction and transfer of endogenous Hh [3] to the same LPPs [4] is not impaired. (D) In contrast, impaired N-processing by the tag (red star) may stall Disp-mediated protein transfer [1,2] and create a bottleneck for endogenous Hh export [3], which in turn also reduces endogenous Hh signaling in the disc as well.
-
Figure 8—figure supplement 1—source data 1
Statistical raw data of Figure 8—figure supplement 1.
- https://cdn.elifesciences.org/articles/86920/elife-86920-fig8-figsupp1-data1-v1.zip
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene, fruit fly (Drosophila melanogaster) | Hedgehog; Hh | PMID:8252628 | NCBI ID:NM_001038976.1 | |
| Gene, fruit fly (D. melanogaster) | En-Gal4e16E (En>) | FlyBase; Öztürk-Çolak et al., 2024 | FlyBase ID FBrf0098595 | P(en2.4-GAL4)e16E |
| Gene, fruit fly (D. melanogaster) | GMR-Gal4 (GMR>) | Bloomington Drosophila Stock Center (Indiana University) | Bloomington stock #45433 | P(y[+t7.7]w[+mC]=GMR17G12-GAL4)attP2 |
| Gene, fruit fly (D. melanogaster) | UAS-hh, UAS-HAhh, UAS-hhHA | Grobe Lab Schürmann et al., 2018; Manikowski et al., 2020 | Hh or Hh variant expression under UAS-control | |
| Gene, fruit fly (D. melanogaster) | Low Hh expression in eye disc | Grobe Lab; Kastl et al., 2018 | w-;+/+; hhbar3/hhAC | Lacks Hh expression specifically in the eye disc |
| Cell line, human (Homo sapiens) | Bosc23 | Grobe Lab, provided by Dr. D. Robbins; Zeng et al., 2001 | PMID:11395778 | HEK293 derivative |
| Cell line, human (H. sapiens) | Disp1 knockout cells (Disp-/-) | Grobe Lab; Ehring et al., 2022 | Bosc23 clone #17.10 | Lacks Disp expression |
| Cell line, human (H. sapiens) | nt ctrl cells | Grobe Lab; Ehring et al., 2022 | Bosc23 clone #10.5 | Disp-expressing control cells |
| Cell line, human (H. sapiens) | Panc | American Type Culture Collection (ATCC) | ATCC CRL-2553 RRID:CVCL_0480 | Shh-expressing cancer cell line (pancreatic duct) |
| Cell line, murine (Mus musculus) | C17.2 | Merck | 07062902-1VL RRID:CVCL_4511 | Mouse multipotent neural progenitor or stem-like cells |
| Cell line, murine (M. musculus) | C3H10T1/2 | Grobe Lab (provided by Dr. Andrea Hoffmann, GBF Braunschweig, Germany) | Multipotent Shh reporter cell line | |
| Cell line, murine (M. musculus) | NIH3T3 | Leibniz-Institut DSMZ-Deutsche Sammlung von Mikroorganismen und Zellkulturen | DSMZ ACC 59 | Mouse fibroblast Shh reporter cell line |
| Transfected construct, murine (M. musculus) | pIRES | Jakobs et al., 2014 | Clontech 631605 | Bicistronic expression vector |
| Biological sample, human (H. sapiens) | HDL | Merck Millipore, Burlington, NH | Millipore, LP3, MW 175,000-360,000 | |
| Biological sample, human (H. sapiens) | LDL | Merck Millipore, Burlington, NH | Millipore, LP2, MW 2,300,000 Da | |
| Antibody | α-Shh (rabbit monoclonal) | Cell Signaling, Danvers, MA | C9C5 RRID:AB_2188191 | Used to detect cellular and solubilized Shh (1:5000) |
| Antibody | α-GAPDH (rabbit polyclonal) | Cell Signaling, Danvers, MA | 14C10, #2118 RRID:AB_1903993 | Loading control (1:2000) |
| Antibody | α-β-actin HRP conjugated (mouse monoclonal) | Sigma-Aldrich, St. Louis, MO | A3854 RRID:AB_262011 | Loading control (1:10,000) |
| Antibody | α-FLAG (rabbit polyclonal) | Sigma-Aldrich, St. Louis, MO | F7425 RRID:AB_439687 | Used to detect FLAG-tagged Scube2 (1:5000) |
| Antibody | α-mCherry antibodies (rabbit polyclonal) | Thermo Fisher Scientific, Rockford, IL | PA5-34974 RRID:AB_2552323 | (1:2000) |
| Antibody | α-Shh antibody 5E1 (mouse monoclonal) | Developmental Studies Hybridoma Bank (DSHB), Iowa City, IA | DSHB 5E1 RRID:AB_528466 | Binds to Shh pseudoactive site that also binds Ptch1 (1:1000) |
| Antibody | α-ApoA1 (mouse monoclonal) | NovusBio, Wiesbaden, Germany | NB400-147 RRID:AB_10001123 | Detects HDL (integral protein) (1:1000) |
| Antibody | α-ApoE4 (mouse monoclonal) | Cell Signaling, Danvers, MA | (4E4) #2208 RRID:AB_2238543 | Detects HDL (mobile protein) (1:1000) |
| Recombinant DNA reagent | Shh (murine) | Grobe Lab | OriGene NM_009170 | Dual-lipidated Shh |
| Recombinant DNA reagent | ShhN/ C25A/SShh | Grobe Lab | OriGene NM_009170 | Monolipidated Shh |
| Recombinant DNA reagent | Hhat | Grobe Lab | OriGene NM_018194 | |
| Recombinant DNA reagent | V5-tagged Disptg | Stacey Ogden (St. Jude Children’s Research Hospital, Memphis, USA) | ||
| Recombinant DNA reagent | Ptch1tg and Ptch1ΔL2 | Ehring et al., 2022 | Addgene_#120889 | |
| Recombinant DNA reagent | pNH-NanoLuc | Addgene, Watertown, MA | #173075 | |
| Peptide, recombinant protein | HEK293-derived human Shh | R&D Systems, Minneapolis, MN | 8908-SH | Dual-lipidated Shh extracted from transfected HEK cells |
| Commercial assay or kit | Gibson assembly | New England Biolabs, Frankfurt, Germany | HiFi Assembly Kit, NEB # E5520S | |
| Commercial assay or kit | Mouse mesenchyme stem cell functional id kit | R&D Systems, Minneapolis, MN | R&D Systems, SC010 | |
| Chemical compound, drug | [3H]-cholesterol | PerkinElmer, Waltham, MA | NET139250UC | |
| Software, algorithm | ImageJ | Schroeder et al., 2021 | Version 1.54g | Immunoblot quantification |
| Software, algorithm | GraphPad Prism | GraphPad Software, Boston, MA , https://graphad.com | Version 10.2.3 (347) | |
| Other | Protein A beads | Sigma-Aldrich, St. Louis, MO | P1406 | ProteinA derived from Staphylococcus aureus coupled to agarose beads for IgG antibody binding and subsequent immunoprecipitation |
Additional files
-
Supplementary file 1
Raw data used for the figures in this study.
- https://cdn.elifesciences.org/articles/86920/elife-86920-supp1-v1.docx
-
Supplementary file 2
qPCR target genes analyzed in this work.
- https://cdn.elifesciences.org/articles/86920/elife-86920-supp2-v1.docx
-
Supplementary file 3
Information regarding amplicons and primers.
- https://cdn.elifesciences.org/articles/86920/elife-86920-supp3-v1.docx
-
MDAR checklist
- https://cdn.elifesciences.org/articles/86920/elife-86920-mdarchecklist1-v1.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Two-way Dispatched function in Sonic hedgehog shedding and transfer to high-density lipoproteins
eLife 12:RP86920.
https://doi.org/10.7554/eLife.86920.4




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}