ROM1 is redundant to PRPH2 as a molecular building block of photoreceptor disc rims
- Department of Ophthalmology, Duke University Medical Center, United States
- Department of Biomedical Engineering, University of Houston, United States
- College of Optometry, University of Houston, United States
- Department of Cell Biology, University of Oklahoma Health Sciences Center, United States
- Department of Pharmacology and Cancer Biology, Duke University Medical Center, United States
Figures
Figure 1
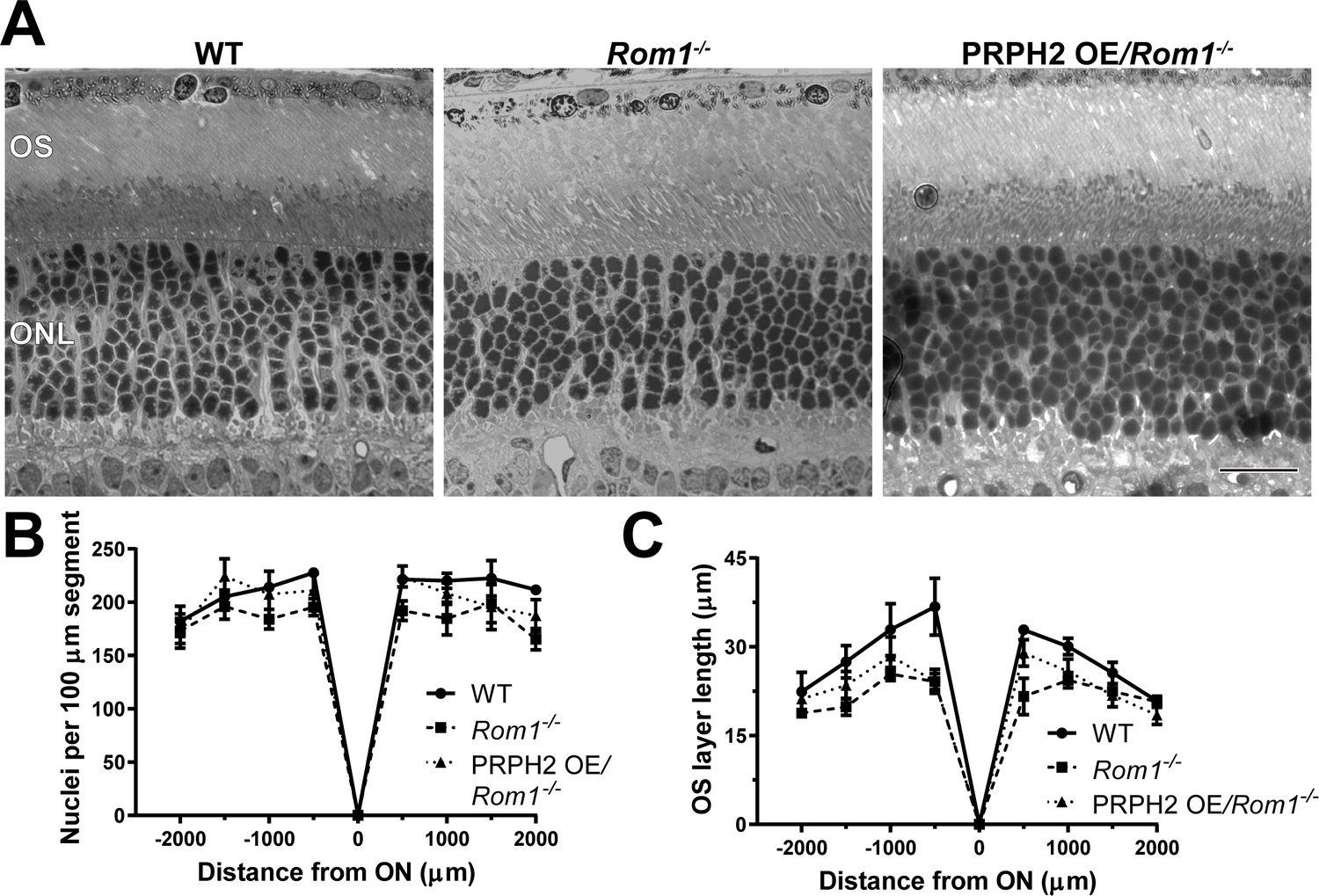
Light microscopy of retinas from WT, Rom1-/- and PRPH2 OE/Rom1-/- mice.
(A) Representative light microscopy images of WT, Rom1-/- and PRPH2 OE/Rom1-/- retinas analyzed at P30. OS: outer segment; ONL: outer nuclear layer. Scale bar: 20 µm. (B) Quantification of the number of photoreceptor nuclei in a 100 µm segment of the retina at 500 µm increments away from the optic nerve (ON). Three retinas were analyzed for each genotype. Two-way ANOVA revealed statistically significant differences in the nuclear counts across genotypes (p=0.0002). Sidak’s multiple comparisons post-hoc test revealed statistically significant differences between the total nuclear count between WT and Rom1-/- retinas (p=0.0001) and between Rom1-/- and PRPH2 OE/Rom1-/- retinas (p=0.0148), but not between WT and PRPH2 OE/Rom1-/- retinas (p=0.3564). (C) Quantification of the outer segment (OS) layer length at 500 µm increments away from the optic nerve. Three retinas were analyzed for each genotype. Two-way ANOVA revealed statistically significant differences in the OS layer lengths across genotypes (p<0.0001). Sidak’s multiple comparisons post-hoc test revealed statistically significant differences for the OS layer length between WT and Rom1-/- retinas (p<0.0001) and between WT and PRPH2 OE/Rom1-/- retinas (p=0.0006), but not between Rom1-/- and PRPH2 OE/Rom1-/- retinas (p=0.2924).
Figure 2 with 1 supplement
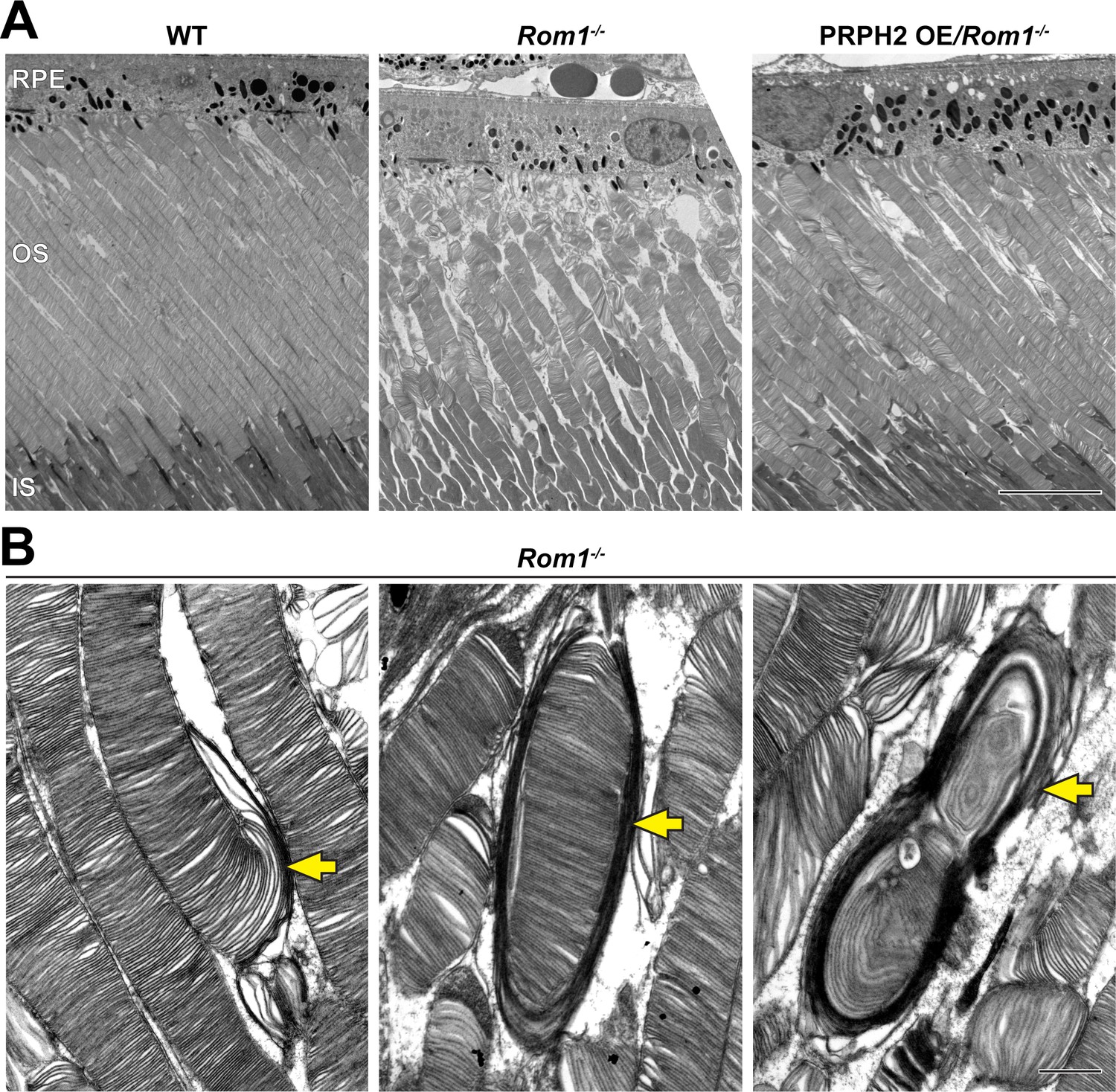
Ultrastructural analysis of retinas and rod outer segments from WT, Rom1-/- and PRPH2 OE/Rom1-/- mice.
(A) Representative low magnification TEM images of WT, Rom1-/- and PRPH2 OE/Rom1-/- retinas analyzed at P30. RPE: retinal pigment epithelium; OS: outer segment; IS: inner segment. Scale bar: 10 µm. (B) Representative high-magnification TEM images of Rom1-/- outer segments. Yellow arrows indicate outer segment structural defects that range from slightly overgrown open discs to membranous whorls. Scale bar: 1 µm.
Figure 2—figure supplement 1
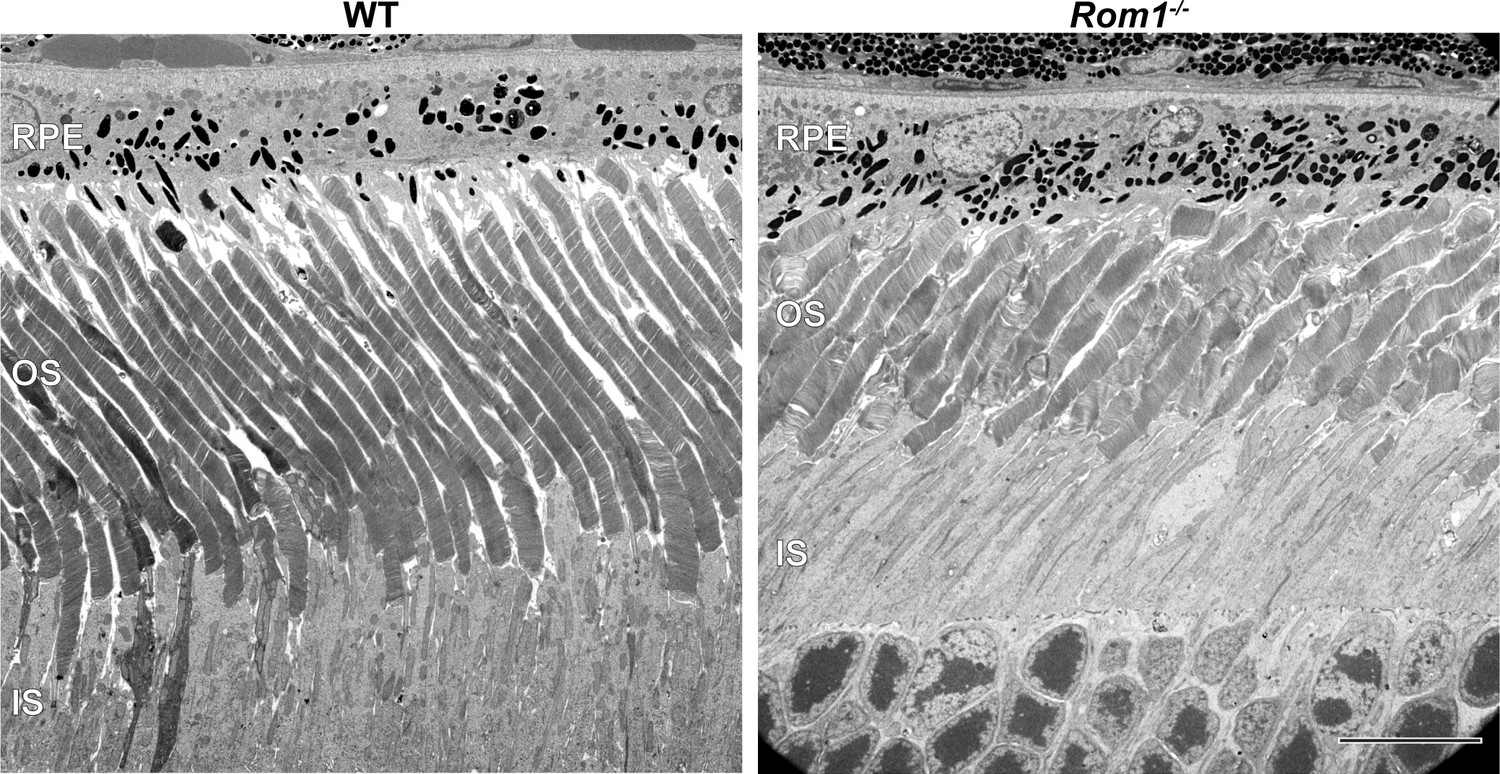
Ultrastructural analysis of WT and Rom1-/- retinas.
Representative low magnification TEM images of WT and Rom1-/- retinas contrasted with the conventionally used osmium tetroxide. RPE: retinal pigment epithelium; OS: outer segment; IS: inner segment. Scale bar: 10 µm.
Figure 3
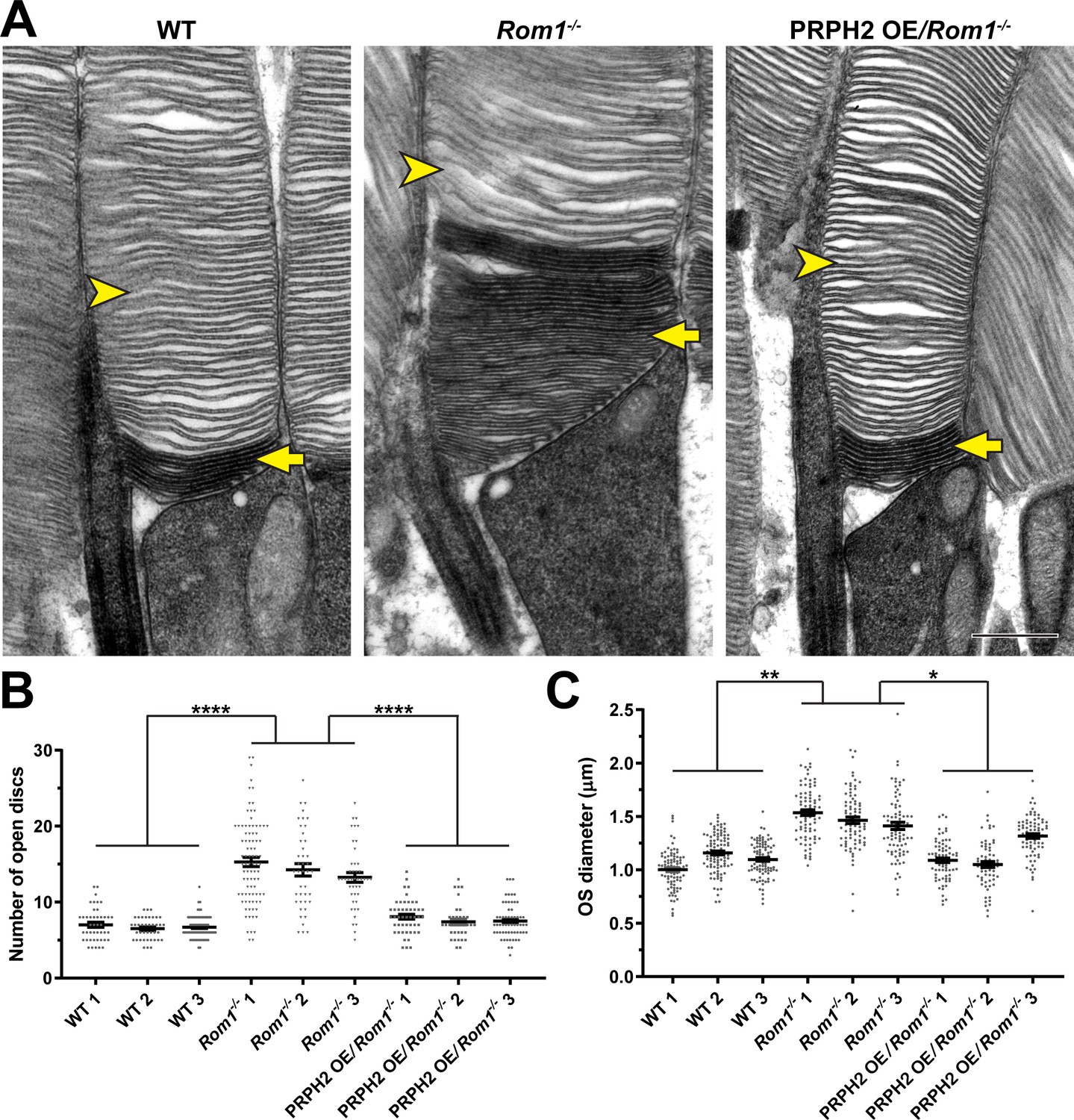
Analysis of disc enclosure in rod outer segments of WT, Rom1-/- and PRPH2 OE/Rom1-/- mice.
(A) Representative high magnification TEM images of tannic acid/uranyl acetate-stained retinas of WT, Rom1-/- and PRPH2 OE/Rom1-/- mice analyzed at P30. This approach stains newly forming ‘open’ discs more intensely than mature enclosed discs. Yellow arrows point to darkly stained, unenclosed discs; yellow arrowheads point to lightly stained, enclosed discs. Scale bar: 0.5 µm. (B) Quantification of the number of darkly stained open discs at the rod outer segment base. Each data point represents a single outer segment. For each genotype, three retinas were analyzed with at least 35 outer segments analyzed per retina. Data were plotted with samples separated, while statistical analysis was performed on the averages within each retina (n=3 for each genotype). One-way ANOVA revealed statistically significant differences in the number of open discs across genotypes (p<0.0001). Tukey’s multiple comparisons post-hoc test revealed statistically significant differences in the number of open discs between WT and Rom1-/- (p<0.0001) and Rom1-/- and PRPH2 OE/Rom1-/- (p<0.0001) mice, but not between WT and PRPH2 OE/Rom1-/- mice (p=0.2686). (C) Quantification of the outer segment (OS) diameter. For each genotype, three retinas were analyzed with at least 66 outer segments analyzed per retina. One-way ANOVA revealed statistically significant differences in the OS diameters across genotypes (p=0.0074). Tukey’s multiple comparisons post-hoc test revealed statistically significant differences in the OS diameters between WT and Rom1-/- (p=0.0083) and Rom1-/- and PRPH2 OE/Rom1-/- (p=0.0197) mice, but not between WT and PRPH2 OE/Rom1-/- mice (p=0.7198). Note that these diameters were measured in longitudinal sections, in which outer segment are not always sectioned across their widest part; therefore, these values are likely under-representations of the true OS diameters. However, this does not affect the comparison across genotypes.
Figure 4

Ultrastructural analysis of disc incisures in WT, Rom1-/- and PRPH2 OE/Rom1-/- mice.
Representative TEM images of retinas, tangentially sectioned through the outer segment layer, from WT, Rom1-/- and PRPH2 OE/Rom1-/- mice analyzed at P30. Yellow arrowheads indicate incisures observed in WT and PRPH2 OE/Rom1-/- discs, but not Rom1-/- discs. Scale bar: 1 µm.
Figure 5
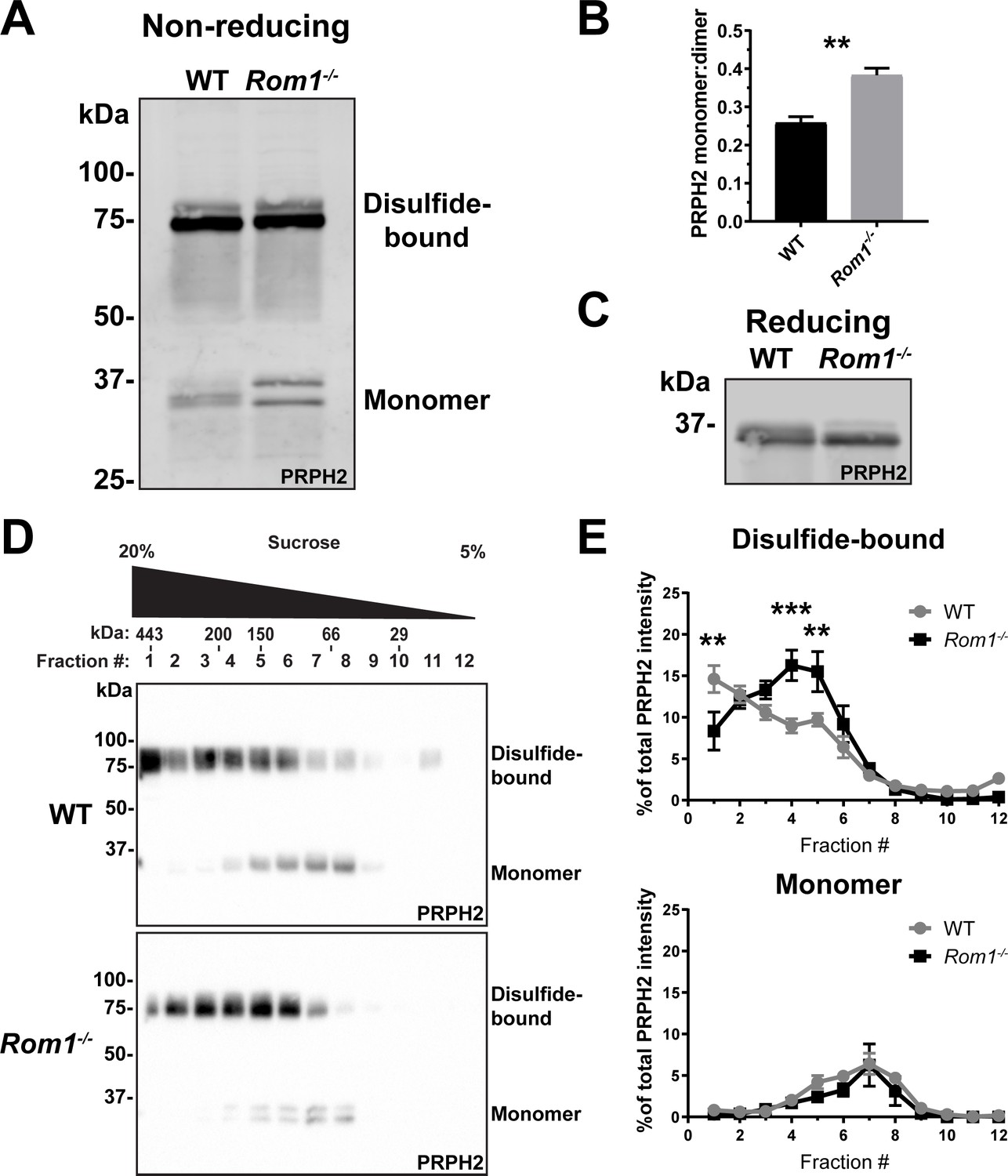
Loss of ROM1 alters the oligomerization status of PRPH2.
(A) Western blot probed for PRPH2 after protein separation by SDS-PAGE under non-reducing conditions. Each sample contained 10 µg of lysate obtained from eyecups of WT or Rom1-/- mice at P30. Under these conditions, PRPH2 runs as disulfide-bound dimers (~75 kDa) and monomers (~37 kDa). The monomer band of PRPH2 runs as a doublet in Rom1-/- but not WT eyecups. (B) Quantification of the ratio between monomer and disulfide-bound (dimer) bands of PRPH2 was performed using densitometry of three independent lysates. For the doublet of monomer bands in Rom1-/- lysates, both bands were used for quantification. Unpaired t-test revealed a statistically significant difference in the PRPH2 monomer:dimer ratio between WT and Rom1-/- retinas (p=0.0071). (C) Western blot probed for PRPH2 after protein separation by SDS-PAGE under reducing conditions. Each sample contained 10 µg of lysate obtained from eyecups of WT or Rom1-/- mice. (D) Lysates obtained under non-reducing conditions from eyecups of WT and Rom1-/- mice were subjected to velocity sedimentation on 5–20% sucrose gradients. Twelve fractions were collected with fraction #1 corresponding to 20% sucrose and fraction #12 to 5% sucrose. Proteins from each fraction were subjected to non-reducing SDS-PAGE and Western blotting for PRPH2. The distribution of molecular mass standards across fraction, as determined in Chakraborty et al., 2008, is shown above the panels. (E) Quantification of both the monomeric and disulfide-bound bands of PRPH2 in each fraction was performed using densitometry of at least four independent samples and normalized to the total PRPH2 content across all fractions. Two-way ANOVA revealed statistically significant differences in the PRPH2 content across genotypes and fractions (p<0.0001 for disulfide-bound, p=0.9489 for monomer). Sidak’s multiple comparisons post-hoc test revealed statistically significant differences between genotypes for the disulfide-bound form in fractions #1 (p=0.0022), #4 (p=0.0002), and #5 (p=0.0061).
-
Figure 5—source data 1
Full western blots associated with Figure 5A.
- https://cdn.elifesciences.org/articles/89444/elife-89444-fig5-data1-v1.zip
-
Figure 5—source data 2
Full western blots associated with Figure 5C.
- https://cdn.elifesciences.org/articles/89444/elife-89444-fig5-data2-v1.zip
-
Figure 5—source data 3
Full western blots associated with Figure 5D.
- https://cdn.elifesciences.org/articles/89444/elife-89444-fig5-data3-v1.zip
Figure 6
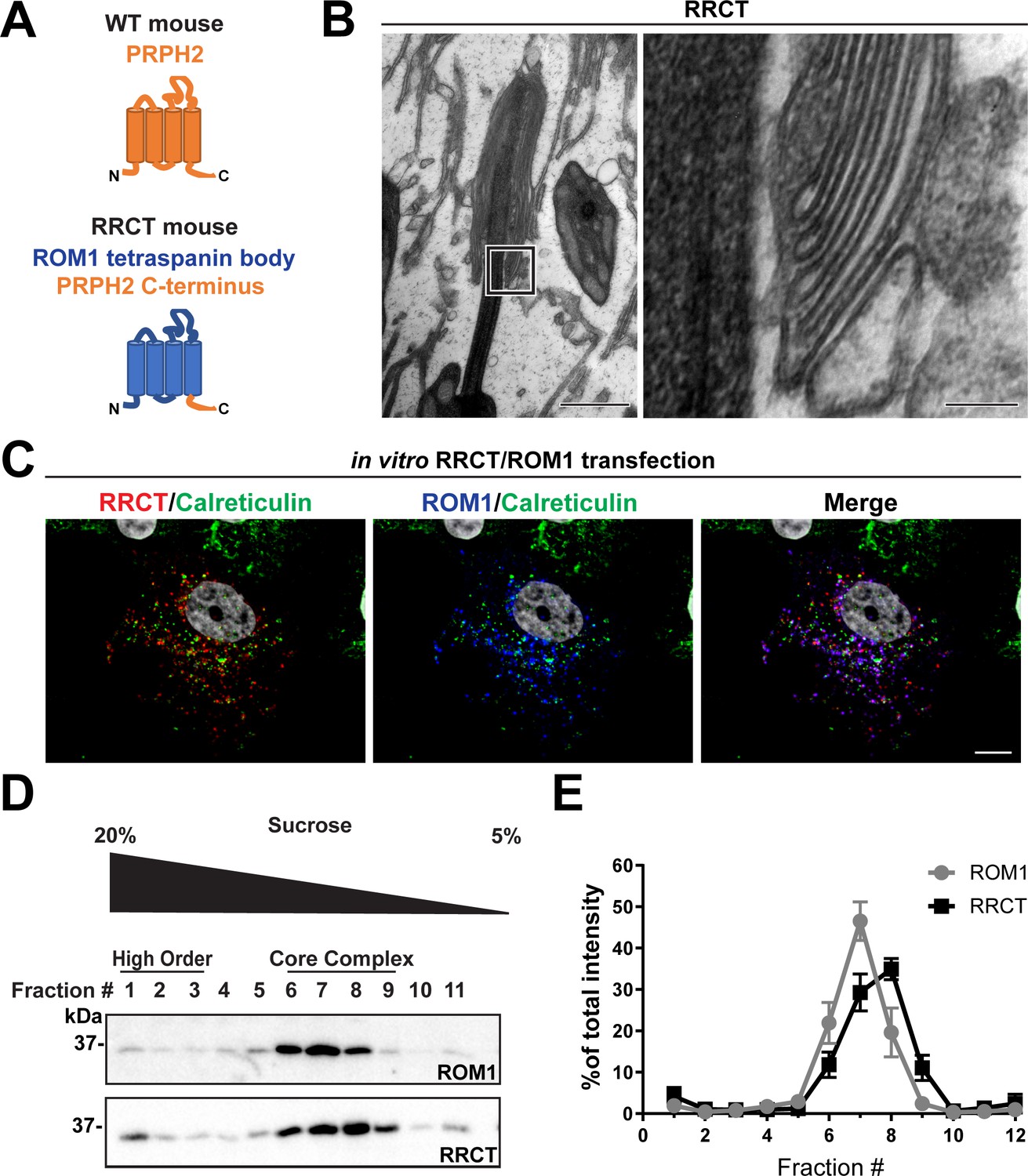
The tetraspanin body of PRPH2 can be replaced by that of ROM1 in disc rim formation.
(A) Cartoon schematic of the RRCT chimeric tetraspanin protein. In the RRCT mouse, the Prph2 gene has a knockin mutation that replaces it with a DNA sequence encoding a chimeric protein consisting of the tetraspanin body of ROM1 while retaining the C-terminal tail of PRPH2 that is essential for disc formation. (B) Representative TEM images of homozygous RRCT mice analyzed at P30. The boxed inset (left) is shown at a higher magnification (right) that reveals the presence of disc rims. Scale bars: 1 µm (left); 0.1 µm (right). (C) Immunofluorescent images of COS-7 cells co-transfected with RRCT-FLAG and ROM1 constructs. Cells were stained with antibodies against RRCT (red), ROM1 (blue) and calreticulin (green) to label ER membranes. Nuclei were counterstained with DAPI (grey). Scale bar: 10 µm. (D) Lysates obtained under non-reducing conditions from COS-7 cells co-transfected with RRCT-FLAG and ROM1 constructs were subjected to velocity sedimentation on 5–20% sucrose gradients. Twelve fractions were collected with fraction #1 corresponding to 20% sucrose and fraction #12 to 5% sucrose. Proteins from each fraction were subjected to reducing SDS-PAGE and Western blotting for ROM1 and RRCT. (E) Quantification of ROM1 and RRCT in each fraction was performed using densitometry of three independent lysates and normalized to the total content across all fractions.
-
Figure 6—source data 1
Full western blots associated with Figure 6D.
- https://cdn.elifesciences.org/articles/89444/elife-89444-fig6-data1-v1.zip
Figure 7
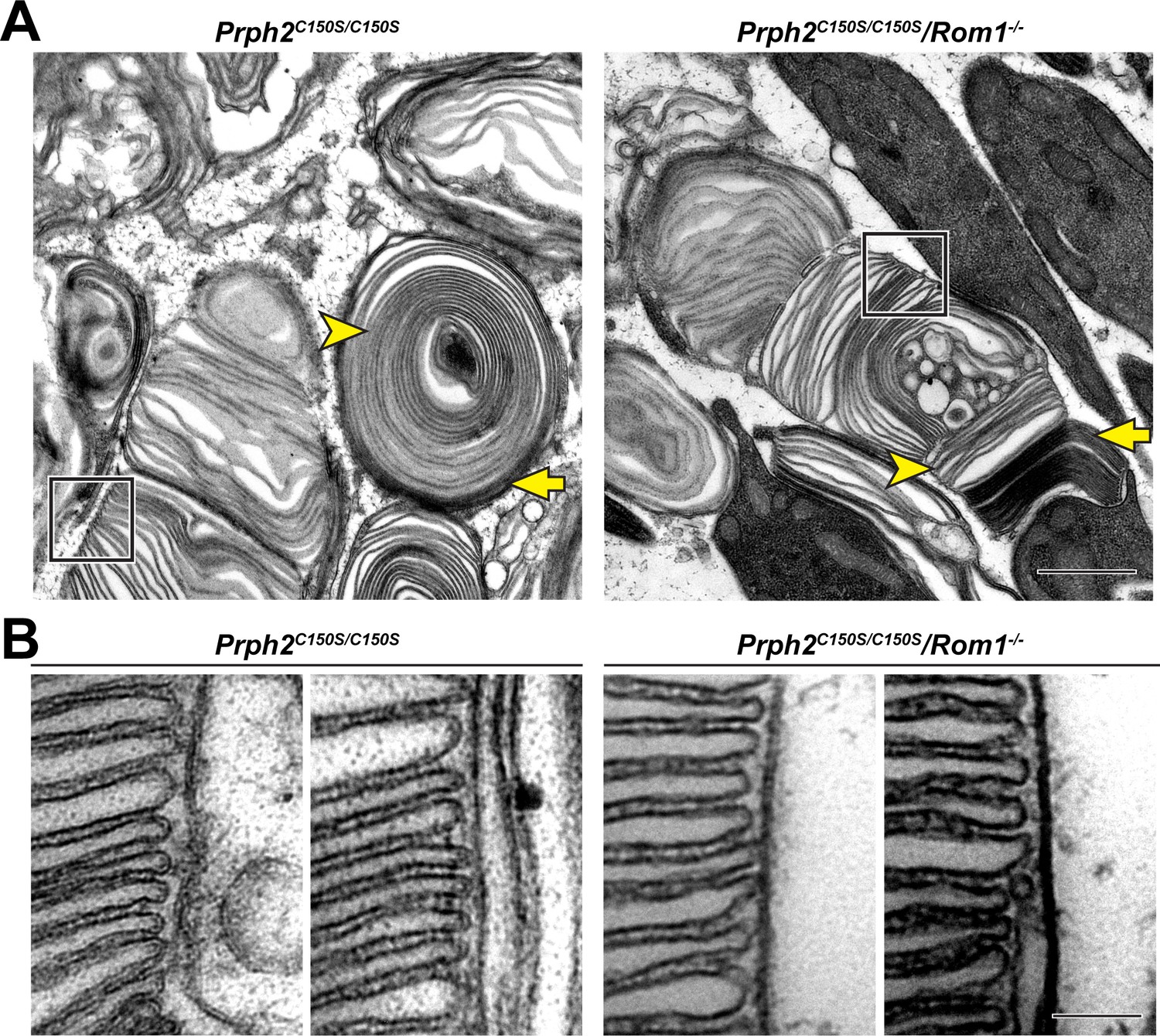
Disc rims can be formed in the absence of intermolecular disulfide bonds between PRPH2 and ROM1.
(A) Representative TEM images of tannic acid/uranyl acetate-stained retinas of Prph2C150S/C150S and compound Prph2C150S/C150S/Rom1-/- mice analyzed at P30. Photoreceptor outer segments of each mouse have severely perturbed outer segment structure. Yet, disc enclosure is not completely prevented as there are both darkly-stained, ‘open’ discs (yellow arrows) and lightly-stained, ‘enclosed’ discs (yellow arrowheads) in each genotype. The boxed region of each image depicts an area in which disc stacking appears relatively normal and permits the analysis of disc rim structure. Scale bar: 1 µm. (B) Higher magnification images of the regions in which disc stacking appears normal in both Prph2C150S/C150S and Prph2C150S/C150S/Rom1-/- mice. Disc rims are formed in each genotype. Scale bar: 0.1 µm.
Tables
Table 1
Quantification of molar ratios between PRPH2, ROM1 and rhodopsin in WT and Rom1-/- outer segments.
| Protein molar ratios* | WT | Rom1-/- |
|---|---|---|
| PRPH2:rhodopsin | 1:18.1±0.5 | 1:12.2±1.3 |
| ROM1:rhodopsin | 1:36.3±3.9 | n/a |
| PRPH2:ROM1 | 2.0:1±0.3 | n/a |
| (PRPH2 +ROM1):rhodopsin | 1:12.1±0.2 | 1:12.2±1.3 |
-
*
Values are shown as mean ± s.d. n/a: not applicable. Two outer segment preparations from mice of each genotype were analyzed.
-
Table 1—source data 1
Quantification of molar ratios between tetraspanins and rhodopsin in WT and Rom1-/- mice – raw data.
- https://cdn.elifesciences.org/articles/89444/elife-89444-table1-data1-v1.xlsx
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (Mus musculus) | C57BL/6 J | Jackson Labs | Jax#:000664 | |
| Genetic reagent (Mus musculus) | Prph2C150S | Stuck et al., 2014 | MGI:6367798 | |
| Genetic reagent (Mus musculus) | RRCT | Conley et al., 2019 | ||
| Genetic reagent (Mus musculus) | PRPH2 OE | Nour et al., 2004 | ||
| Genetic reagent (Mus musculus) | Rom1-/- | Clarke et al., 2000 | MGI:2181662 | |
| Cell line (Cercopithecus aethiops) | COS-7 | ATCC | CRL-1651 | ATCC provides authentication and confirmation that mycoplasma contamination was not detected |
| Recombinant DNA reagent | RRCT-FLAG | Conley et al., 2019 | pcDNA3.1 plasmid | |
| Recombinant DNA reagent | ROM1 | Conley et al., 2010 | pcDNA3.1 plasmid with murine Rom1 | |
| Antibody | Anti-PRPH2 (polyclonal rabbit) | Kedzierski et al., 1999 | WB: (1:1000) | |
| Antibody | Anti-ROM1 (polyclonal sheep) | Spencer et al., 2023 | WB: (1:5000) | |
| Antibody | Anti-RRCT (2B7; monoclonal mouse) | Conley et al., 2014 | WB: (1:500) | |
| Antibody | Anti-ROM1 (2H5; monoclonal mouse) | Conley et al., 2014 | WB: (1:500) | |
| Antibody | Anti-RRCT (2E7; monoclonal mouse) | Zulliger et al., 2015 | IF: (1:2) | |
| Antibody | Anti-ROM1 (2Rom1; polyclonal rabbit) | Ding et al., 2005 | IF: (1:400) | |
| Antibody | Anti-calreticulin (polyclonal chicken) | Abcam | ab2908 | IF: (1:250) |
Additional files
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
ROM1 is redundant to PRPH2 as a molecular building block of photoreceptor disc rims
eLife 12:RP89444.
https://doi.org/10.7554/eLife.89444.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}