The target of rapamycin signaling pathway regulates vegetative development, aflatoxin biosynthesis, and pathogenicity in Aspergillus flavus
- State Key Laboratory of Ecological Pest Control for Fujian and Taiwan Crops, Key Laboratory of Pathogenic, Fungi and Mycotoxins of Fujian Province, School of Life Sciences, Fujian Agriculture and Forestry University, China
eLife assessment
This manuscript provides important information about the influence of TOR signaling pathway on development and aflatoxin production in the plant and human fungal pathogen Aspergillus flavus. Compared to an earlier version, the authors have addressed most of the concerns of the reviewers, including the convincing demonstration of the essential TOR pathway in this fungus by constructing a xylose promoter mutant strain.
https://doi.org/10.7554/eLife.89478.5.sa0Significance of the findings:
Important: Findings that have theoretical or practical implications beyond a single subfield
- Landmark
- Fundamental
- Important
- Valuable
- Useful
Strength of evidence:
Convincing: Appropriate and validated methodology in line with current state-of-the-art
- Exceptional
- Compelling
- Convincing
- Solid
- Incomplete
- Inadequate
During the peer-review process the editor and reviewers write an eLife Assessment that summarises the significance of the findings reported in the article (on a scale ranging from landmark to useful) and the strength of the evidence (on a scale ranging from exceptional to inadequate). Learn more about eLife Assessments
Abstract
The target of rapamycin (TOR) signaling pathway is highly conserved and plays a crucial role in diverse biological processes in eukaryotes. Despite its significance, the underlying mechanism of the TOR pathway in Aspergillus flavus remains elusive. In this study, we comprehensively analyzed the TOR signaling pathway in A. flavus by identifying and characterizing nine genes that encode distinct components of this pathway. The FK506-binding protein Fkbp3 and its lysine succinylation are important for aflatoxin production and rapamycin resistance. The TorA kinase plays a pivotal role in the regulation of growth, spore production, aflatoxin biosynthesis, and responses to rapamycin and cell membrane stress. As a significant downstream effector molecule of the TorA kinase, the Sch9 kinase regulates aflatoxin B1 (AFB1) synthesis, osmotic and calcium stress response in A. flavus, and this regulation is mediated through its S_TKc, S_TK_X domains, and the ATP-binding site at K340. We also showed that the Sch9 kinase may have a regulatory impact on the high osmolarity glycerol (HOG) signaling pathway. TapA and TipA, the other downstream components of the TorA kinase, play a significant role in regulating cell wall stress response in A. flavus. Moreover, the members of the TapA-phosphatase complexes, SitA and Ppg1, are important for various biological processes in A. flavus, including vegetative growth, sclerotia formation, AFB1 biosynthesis, and pathogenicity. We also demonstrated that SitA and Ppg1 are involved in regulating lipid droplets (LDs) biogenesis and cell wall integrity (CWI) signaling pathways. In addition, another phosphatase complex, Nem1/Spo7, plays critical roles in hyphal development, conidiation, aflatoxin production, and LDs biogenesis. Collectively, our study has provided important insight into the regulatory network of the TOR signaling pathway and has elucidated the underlying molecular mechanisms of aflatoxin biosynthesis in A. flavus.
Introduction
Aspergillus flavus is a highly significant phytopathogenic fungus that frequently contaminates a diverse array of agricultural crops, leading to substantial economic losses in the food and agriculture industry and posing considerable health risks, particularly in developing countries (Klich, 2007). Moreover, A. flavus is recognized as an opportunistic human pathogenic fungus that can cause aspergillosis in immune-compromised patients (Hedayati et al., 2007). The fungus is notorious for its production of aflatoxins, which are potent secondary metabolites with toxic and carcinogenic properties and are classified among the most powerful carcinogens known in nature, with established links to liver cancer in both humans and animals (Khan et al., 2021). Aflatoxin biosynthesis is a multifaceted and intricate biochemical process encompassing a cascade of enzymatic reactions. Although the majority of the enzyme reactions and genes involved in aflatoxin biosynthesis have been elucidated (Yabe and Nakajima, 2004), the related signaling pathway network and specific regulatory mechanisms governing aflatoxin biosynthesis remain elusive. Therefore, exploring the regulatory mechanism of the aflatoxin biosynthesis signaling pathway would provide new insights for preventing A. flavus and aflatoxin contamination.
In eukaryotes, the target of rapamycin (TOR) signaling pathway is highly conserved and plays essential roles in various significant biological processes, including ribosome biosynthesis, cell growth, and autophagy (Wullschleger et al., 2006). The serine/threonine-protein kinase Tor serves as a crucial protein component in the TOR signaling pathway, which interacts with other proteins to form multiprotein complexes (De Virgilio and Loewith, 2006). In Saccharomyces cerevisiae, TOR exists in two distinct multiprotein functional complexes, namely target rapamycin complex 1 (TORC1) and TORC2 (Loewith et al., 2002). TORC1-mediated signaling governs cellular growth by modulating various growth-related mechanisms and exhibits sensitivity to rapamycin. Conversely, TORC2-mediated signaling primarily regulates cytoskeletal remodeling but remains unaffected by rapamycin (Loewith et al., 2002). In yeast, there are two tor genes, tor1 and tor2, whereas in higher eukaryotes such as plants, animals, and filamentous fungi, there is only one tor gene (Beauchamp and Platanias, 2013; Fu et al., 2021; Teichert et al., 2006).
In the fungal kingdom, the TOR signaling pathway has been extensively investigated in three prominent species: the budding yeast S. cerevisiae (Crespo and Hall, 2002; Rohde et al., 2008), the fission yeast Schizosaccharomyces pombe (Ishiguro et al., 2013; Otsubo and Yamamato, 2008; Petersen, 2009), and the human pathogen Candida albicans (Bastidas et al., 2009; Liu et al., 2017). In S. cerevisiae, the macrolide antibiotic rapamycin forms a complex with the cytosolic protein Fkbp12, which effectively inhibits the catalytic function of the Tor kinase by interacting with its FRB domain (Hu et al., 2016). The Tap42 phosphatase complex serves as the primary target of the Tor kinase and plays a crucial role in the regulation of downstream effectors. Additionally, the Tap42 protein interacts with the Tip41 protein to collaboratively modulate the activities of phosphatases, including PP2A and Sit4 (Wang et al., 2003; Yan et al., 2006). Among these, the Sit4 phosphatase is known to dephosphorylate the transcription factor Gln3, thereby regulating the nitrogen source metabolism pathway (Tate et al., 2019). In addition, the AGC kinase Sch9, an immediate effector of TORC1, is involved in regulating various cellular function processes, such as ribosome biosynthesis, translation initiation, and entry into the G0 phase (Urban et al., 2007). The TOR signaling pathway has been documented in various filamentous fungi, including Fusarium graminearum (Yu et al., 2014; Liu et al., 2019), Magnaporthe oryzae (Sun et al., 2018), Phanerochaete chrysosporium (Nguyen et al., 2020), Podospora anserine (Dementhon et al., 2003), Aspergillus nidulans (Fitzgibbon et al., 2005), Aspergillus fumigatus (Alves de Castro and Dos Reis, 2016; Baldin et al., 2015), Fusarium fujikuroi, Botrytis cinerea (Meléndez et al., 2009), Fusarium oxysporum (López-Berges et al., 2010), and Mucor circinelloides (Bastidas et al., 2012). However, the TOR signaling pathway in A. flavus remains unexplored.
In eukaryotic cells, the TOR signaling pathway and the mitogen-activated protein kinase (MAPK) signaling pathways play an important role in regulating adaptive responses to extra- and intracellular conditions (Madrid et al., 2016). Several studies have demonstrated that the TOR signaling pathway interacts with various other signaling pathways, such as MAPK and cell wall integrity (CWI) pathway. These pathways are recognized to participate in crosstalk, forming a complex metabolic network that regulates cellular processes (Torres et al., 2002; Chen et al., 2019; Gu et al., 2015; Qian et al., 2018; Qian et al., 2021). However, the underlying mechanisms of multiple crosstalks between the TOR and other signaling pathways in A. flavus remain largely unknown. Therefore, we intended to identify the genes associated with the TOR signaling pathway and elucidate their roles and contributions in regulating vegetative development and aflatoxin biosynthesis in A. flavus. Our results demonstrated that the TOR signaling pathway is involved in multiple cellular processes in A. flavus. Notably, our findings revealed a complex interplay between the TOR pathway and other signaling pathways (such as high osmolarity glycerol [HOG] and CWI) in A. flavus, which play a crucial role in regulating cellular growth and survival under environmental stress conditions.
Results
Rapamycin exhibits inhibitory effects on the growth, sporulation, sclerotia formation, and aflatoxin production in A. flavus
Rapamycin, a secondary metabolite synthesized by Streptomyces hygroscopicus, exhibits efficacy in suppressing specific filamentous fungi. In F. graminearum, rapamycin has a significant inhibitory impact on growth and asexual reproduction (Yu et al., 2014). To investigate the effects of rapamycin on vegetative development, sclerotia formation, and secondary metabolism in A. flavus, we conducted an assay to determine the sensitivity of the wild-type (WT) strain to rapamycin. We observed that the WT strain exhibited a high sensitivity to rapamycin. We found that 100 ng/mL of rapamycin significantly inhibited mycelial growth and conidia formation (Figure 1A, D, and E). More importantly, the WT strain exhibited a significant reduction in sclerotia formation and aflatoxin synthesis (Figure 1B, C, F, and G). Additionally, rapamycin resulted in a significant decrease in spore germination rate and sparser conidiophores (Figure 1—figure supplement 1A and B). These results illustrated that the TOR signaling pathway may be involved in regulating the growth, development, and secondary metabolism of A. flavus. Calcofluor white (CFW) is a chemifluorescent blue dye commonly employed for its nonspecific binding affinity to fungal chitin. Fluorescence microscopy analysis revealed a notable reduction in the blue fluorescence on the cell wall following treatment with rapamycin (Figure 1—figure supplement 1C). The intracellular lipid droplets (LDs) within the hyphae were labeled with BODIPY. We observed that the hyphae treated with rapamycin contained more LDs compared to the control group (Figure 1—figure supplement 1D). These findings suggested that the TOR pathway may play important roles in maintaining CWI and facilitating the biogenesis of LDs.
Figure 1 with 1 supplement see all
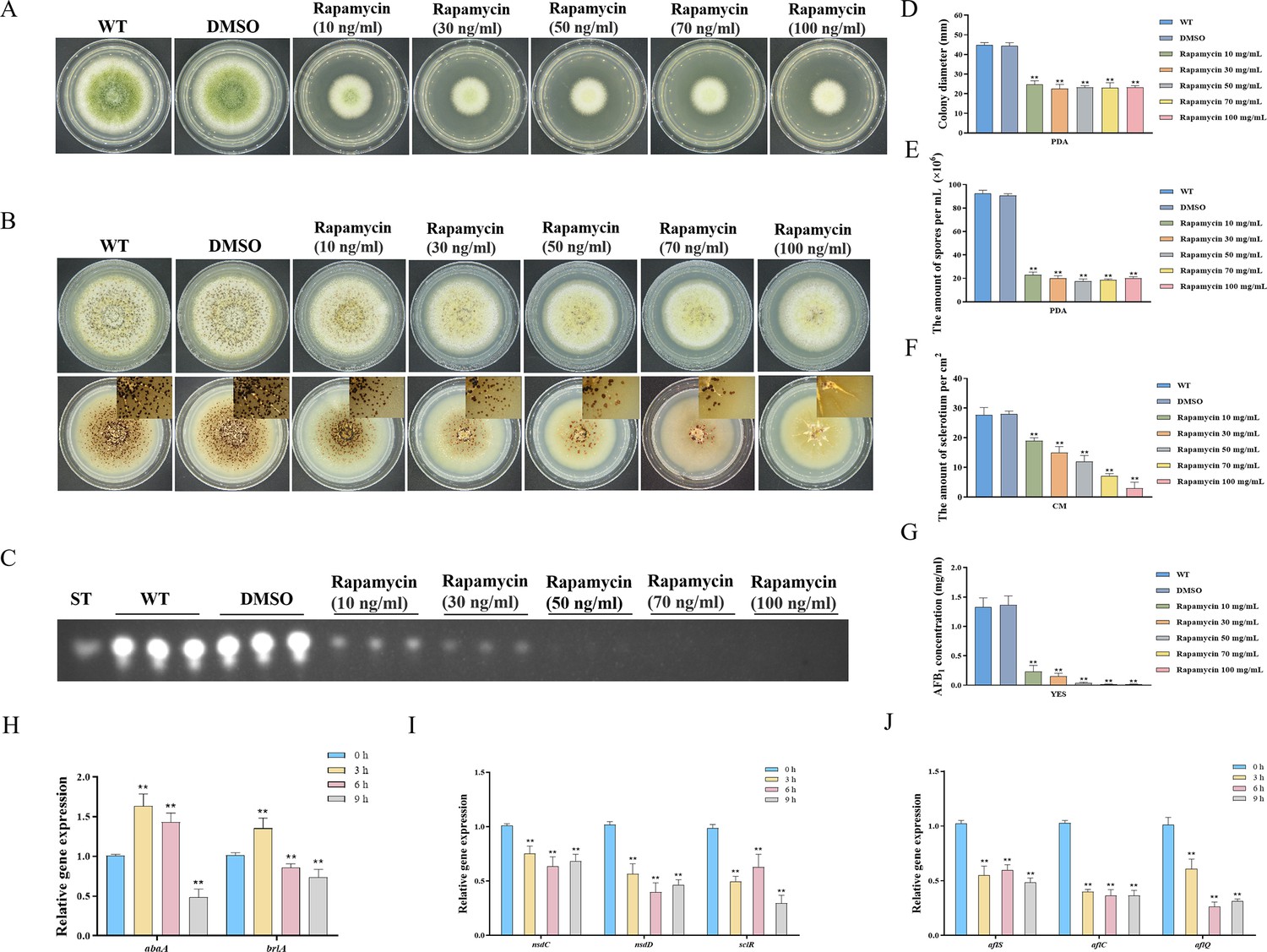
Impacts of rapamycin on the growth, sporulation, sclerotia formation, and aflatoxin production of A. flavus.
(A) Colony morphology of the wild-type (WT) strain cultured on potato dextrose agar (PDA) medium amended with different concentrations of rapamycin at 37°C for 5 days. (B) Colony phenotype of the WT strain grown on CM medium amended with different concentrations of rapamycin at 37°C for 7 days. (C) Thin layer chromatography (TLC) assay of aflatoxin B1 (AFB1) production by the WT strain cultured in yeast extract-sucrose agar (YES) liquid media amended with different concentrations of rapamycin at 29°C for 6 days. (D) Statistical analysis of the colony diameter by the WT strain treated with different concentrations of rapamycin described in (A). (E) Conidial quantification of the WT strain treated with different concentrations of rapamycin as mentioned in (A). (F) Quantitative analysis of sclerotium formation by the WT strain treated with different concentrations of rapamycin described in (B). (G) Relative quantification of AFB1 production by the WT strain treated with different concentrations of rapamycin as mentioned in (C). (H) Relative expression levels of conidia-related genes in WT strain treated with 100 ng/mL rapamycin after 3, 6, 9 hr. (I) Relative expression levels of sclerotia-related genes in WT strain treated with 100 ng/mL rapamycin after 3, 6, 9 hr. (J) Relative expression levels of aflatoxin biosynthesis regulatory and structural genes in WT strain treated with 100 ng/mL rapamycin after 3, 6, 9 hr. ** indicates that the significance level was p≤0.01, based on one-way ANOVA test with three replicates(n=3). Error bars represent the standard error of the mean (SEM).
To investigate the potential regulatory role of the TOR pathway in modulating the sporulation, sclerotia formation, and aflatoxin biosynthesis in A. flavus, we examined the expression levels of conidia-related, sclerotia formation-related, and aflatoxin-related genes after 3, 6, and 9 hr of treatment with rapamycin. To determine the effects of rapamycin on conidiation, qRT-PCR was conducted to assess the transcript levels of two conidia-related genes, abaA and brlA. We also conducted qRT-PCR to evaluate the expression levels of three regulators (nsdC, nsdD, and sclR) involved in sclerotia development, and genes involved in AFB1 biosynthesis, such as the regulatory gene aflS, as well as several structural genes (aflC and aflQ). The strains treated with rapamycin exhibited a significant reduction in the expression levels of genes related to sporulation, sclerotia formation, and aflatoxin synthesis (Figure 1H, I, and J). Overall, these results indicated that the TOR pathway plays a crucial role in regulating the growth, sporulation, sclerotia formation, and aflatoxin biosynthesis in A. flavus by modulating the expression of these genes.
The impact of Fkbp3 and its lysine succinylation on AFB1 biosynthesis and rapamycin resistance in A. flavus
FK506-binding proteins (Fkbps) belong to a highly conserved immunophilin family, which are involved in various cellular functions, such as protein folding, immunosuppression, signaling transduction, and transcription (Tong and Jiang, 2015). Fkbps function as molecular switches by binding to target proteins and inducing conformational changes (Tong and Jiang, 2015). In S. cerevisiae, Fkbp12 and rapamycin interact to form a complex that exerts a negative regulatory effect on the activity of the Tor kinase (Heitman et al., 1991). The fkbp12 gene homolog fprA in A. nidulans shares high levels of homology with that from S. cerevisiae (Fitzgibbon et al., 2005). According to the protein sequence of FprA from A. nidulans, four genes encoding putative proteins with FK506-binding domain, named fkbp1 (AFLA_126880), fkbp2 (AFLA_087480), fkbp3 (AFLA_010910), and fkbp4 (AFLA_128200), have been identified from the A. flavus genome. We generated four fkbp knockout strains using homologous recombination, and all fkbp disruption strains were confirmed by PCR and sequencing analysis (Figure 2—figure supplement 1A). Phenotypic analyses showed that the mycelial growth and conidia formation of all the fkbp mutants were similar to the WT strain on the potato dextrose agar (PDA) medium (Figure 2A), suggesting that Fkbps have minimal influence on the process of vegetative development. We evaluated AFB1 synthesis utilizing thin layer chromatography (TLC), and the findings demonstrated a noteworthy reduction in AFB1 synthesis upon deletion of Fkbp3, in comparison to the WT strain (Figure 2B and D). Additionally, the quantity of sclerotia exhibited a significant decrease in all fkbp mutants compared to the WT strain (Figure 2—figure supplement 2A and B). The aforementioned data indicated that Fkbp3 plays a crucial role in sclerotia formation and aflatoxin biosynthesis in A. flavus.
Figure 2 with 2 supplements see all
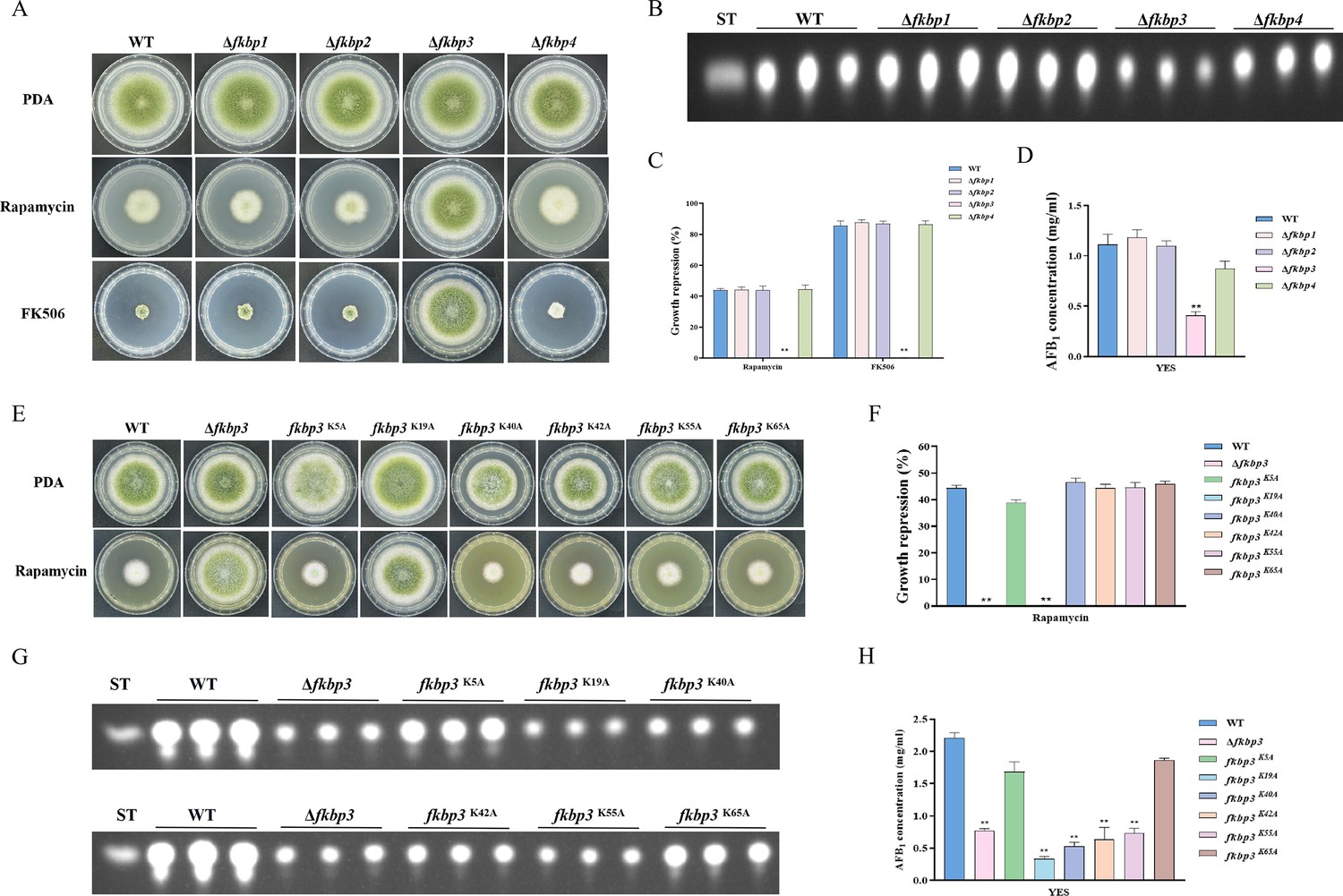
Disruption of fkbp3 significantly increases the resistance of A. flavus to rapamycin and FK506.
(A) Phenotype of the wild-type (WT) and all mutant strains (Δfkbp1, Δfkbp2, Δfkbp3, and Δfkbp4) grown on potato dextrose agar (PDA) amended with rapamycin and FK506 at 37°C for 5 days. (B) Thin layer chromatography (TLC) analysis of aflatoxin B1 (AFB1) production by the WT and all mutant strains (Δfkbp1, Δfkbp2, Δfkbp3, and Δfkbp4) cultured in yeast extract-sucrose agar (YES) liquid medium at 29°C for 6 days. (C) The growth inhibition rate of the WT and all mutant strains (Δfkbp1, Δfkbp2, Δfkbp3, and Δfkbp4) under rapamycin and FK506 stress. (D) AFB1 quantitative analysis of the WT and all mutant strains (Δfkbp1, Δfkbp2, Δfkbp3, and Δfkbp4) as described in (B). (E) Phenotype of the WT and all mutant strains (Δfkbp3, K5A, K19A, K40A, K42A, K55A, and K65A) grown on PDA amended with 100 ng/mL rapamycin at 37°C for 5 days. (F) The growth inhibition rate of the WT and all mutant strains (Δfkbp3, K5A, K19A, K40A, K42A, K55A, and K65A) under rapamycin stress. (G) TLC assay of AFB1 production by the WT and all mutant strains (Δfkbp3, K5A, K19A, K40A, K42A, K55A, and K65A) cultured in YES liquid medium at 29°C for 6 days. (H) Relative quantification of AFB1 production in the WT and all mutant strains (Δfkbp3, K5A, K19A, K40A, K42A, K55A, and K65A) as mentioned in (G). ** indicates that the significance level was p≤0.01, based on one-way ANOVA test with three replicates(n=3). Error bars represent the standard error of the mean (SEM).
To determine the specific Fkbp responsible for rapamycin sensitivity, all mutant strains were cultured on PDA solid plates amended with 100 ng/mL rapamycin. The strain with disrupted fkbp3 showed significant resistance to rapamycin, while the deletion mutants of fkbp1, fkbp2, and fkbp4 did not exhibit any alteration in resistance to rapamycin (Figure 2A and C). These results demonstrated that Fkbp3 serves as the principal target for rapamycin in A. flavus. In the FK506 sensitivity analysis, we observed a significant suppression of mycelial growth and conidia formation in WT strain with the addition of 10 ng/mL FK506 to the PDA medium. Additionally, the deletion of fkbp3 led to the heightened resistance to FK506 compared to the WT strain (Figure 2A and C). These findings suggested that Fkbp3 is mainly involved in the modulation of FK506 resistance in A. flavus.
Previous studies have shown that lysine succinylation plays a significant role in aflatoxin production and pathogenicity in A. flavus (Ren et al., 2018). Based on the analysis of our succinylome data, we have identified six reliable succinylation sites (K5, K19, K40, K42, K55, and K65) on the Fkbp3 protein. To identify the function of Fkbp3 succinylation sites, site-directed mutations were constructed according to the homologous recombination strategy. Compared to the WT strain, all point mutants exhibited a similar phenotype in vegetative growth and conidiation. Interestingly, we observed that the K19A mutation resulted in increased resistance to rapamycin (Figure 2E and F). Additionally, the K19A strain exhibited a significant reduction in the production of AFB1 compared to the WT strain (Figure 2G and H). These findings suggested that the succinylation of Fkbp3 at K19 plays a crucial role in rapamycin resistance and aflatoxin biosynthesis. Furthermore, we found that various point mutants, including K40A, K42A, and K55A, exhibited reduced levels of AFB1 production in comparison to the WT strain (Figure 2G and H), suggesting that the succinylation of Fkbp3 at other sites also contributes to AFB1 biosynthesis in A. flavus. The above results indicated that the K19 site likely plays a more prominent role in Fkbp3 succinylation compared to other sites.
The TorA kinase plays a critical role in A. flavus
In S. cerevisiae, the Tor kinase is known to interact with a diverse range of proteins, resulting in the formation of complex structures known as TORC1 and TORC2 (Weisman, 2016). TORC1 can detect various signals, including nutrients, growth factors, and environmental stress. It plays a crucial role in regulating cellular processes such as gene transcription, protein translation, ribosome synthesis, and autophagy (Loewith and Hall, 2011). To identify the ortholog of the Tor protein in A. flavus, the S. cerevisiae Tor protein was utilized as search reference in the A. flavus genome database using the Basic Local Alignment Search Tool (https://blast.ncbi.nlm.nih.gov/Blast.cgi). The genome of A. flavus contains a solitary ortholog of the torA gene (AFLA_044350), which encodes a protein exhibiting 48.91% similarity to S. cerevisiae Tor2. To elucidate the role of torA in A. flavus, we attempted to obtain torA deletion strains by homologous recombination, but not successful. As an alternative approach, a mutant strain with a xylose promoter mutation, referred as xylPtorA, was constructed to validate the function of torA. Subsequent confirmation via PCR analysis ensured the integrity of the mutant strain. Xylan and xylose are highly effective inducers, whereas glucose strongly represses the activity of the promoter. To ascertain the impact of torA on A. flavus, we conducted a comparative analysis of torA transcript levels between the xylPtorA and the WT strains. Our findings revealed that the torA transcript levels in the xylPtorA strain were markedly reduced compared to the WT strain (Figure 3—figure supplement 1). Both the WT and xylPtorA strains were cultured in yeast extract-xylose agar (YXT) medium supplemented with xylose and yeast extract-glucose agar (YGT) medium devoid of xylose for a duration of 5 days at 37°C under dark conditions. Remarkably, the xylPtorA strain exhibited complete inability to thrive in the absence of xylose, with only partial restoration of growth observed upon xylose supplementation (Figure 3A and D). Notably, the xylPtorA strain also demonstrated a markedly reduced growth rate on YXT compared to the WT strain (Figure 3A and C). TLC assays indicated a noteworthy decrease in AFB1 synthesis in the xylPtorA strain compared to the WT strain (Figure 3B and F). Additionally, the xylPtorA strain produced fewer conidia and sclerotium formation was absent on YXT medium (Figure 3E, Figure 3—figure supplement 2A and B). Interestingly, the xylPtorA strain exhibited a notable increase in sensitivity to rapamycin and SDS stress (Figure 3G and H). Collectively, these findings suggested that the TorA kinase plays a crucial role in various biological processes in A. flavus, including vegetative growth, asexual development, sclerotia formation, response to environmental stress, and biosynthesis of aflatoxins.
Figure 3 with 2 supplements see all
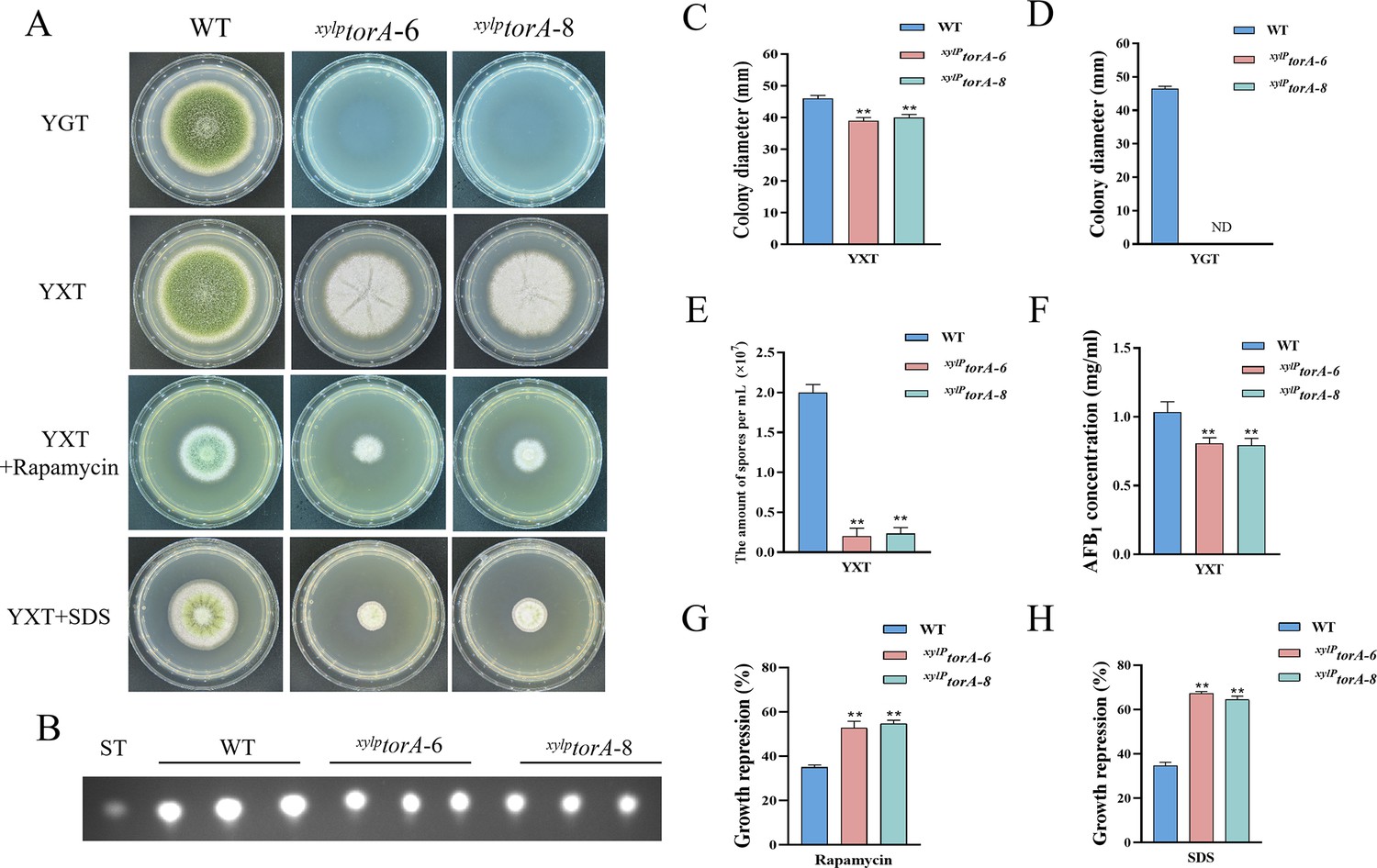
The TorA kinase plays a critical role in A. flavus.
(A) Phenotype of the wild-type (WT) and the xylPtorA strains in yeast extract-glucose agar (YGT) and yeast extract-xylose agar (YXT) medium amended with 100 ng/mL rapamycin and 300 mg/mL SDS at 37°C for 5 days. (B) Thin layer chromatography (TLC) assay of aflatoxin B1 (AFB1) production from the WT and the xylPtorA strains cultured in YXT medium containing 1 g/L MgSO4·7H2O at 29°C for 6 days. (C) Statistical analysis of the colony diameter by the WT and the xylPtorA strains in YXT medium. (D) Statistical analysis of the colony diameter by the WT and the xylPtorA strains in YGT medium. (E) Conidial quantification of the WT and the xylPtorA strains in YXT medium. (F) Quantitative analysis of AFB1 as shown in (B). (G) The growth inhibition rate of the WT and the xylPtorA strains under rapamycin. (H) The growth inhibition rate of the WT and the xylPtorA strains under SDS stress. ND indicates no detection. * indicates that the significance level was p≤0.05, ** indicates that the significance level was p≤0.01, based on one-way ANOVA test with three replicates(n=3). Error bars represent the standard error of the mean (SEM).
The Sch9 kinase is involved in aflatoxin biosynthesis and the HOG pathway
Sch9, a Ser/Thr kinase belonging to the AGC family, is directly phosphorylated by TORC1. The phosphorylation of Sch9 is inhibited by rapamycin, as well as by carbon or nitrogen starvation (Urban et al., 2007). The putative sch9 gene (AFLA_127440) in A. flavus encodes a protein consisting of 706 amino acids, exhibiting 80.31% identity to the Sch9 protein in A. nidulans. To investigate the role of Sch9 in A. flavus, we attempted to create a sch9 deletion mutant using a homologous recombination strategy. The ∆sch9 strain exhibited typical vegetative growth and conidiation similar to the WT strain (Figure 4B). TLC assay and quantitative analysis showed a significantly decreased aflatoxin production in the ∆sch9 strain compared to the WT strain (Figure 4C and G). These results indicated that Sch9 regulates aflatoxin biosynthesis in A. flavus.
Figure 4 with 1 supplement see all
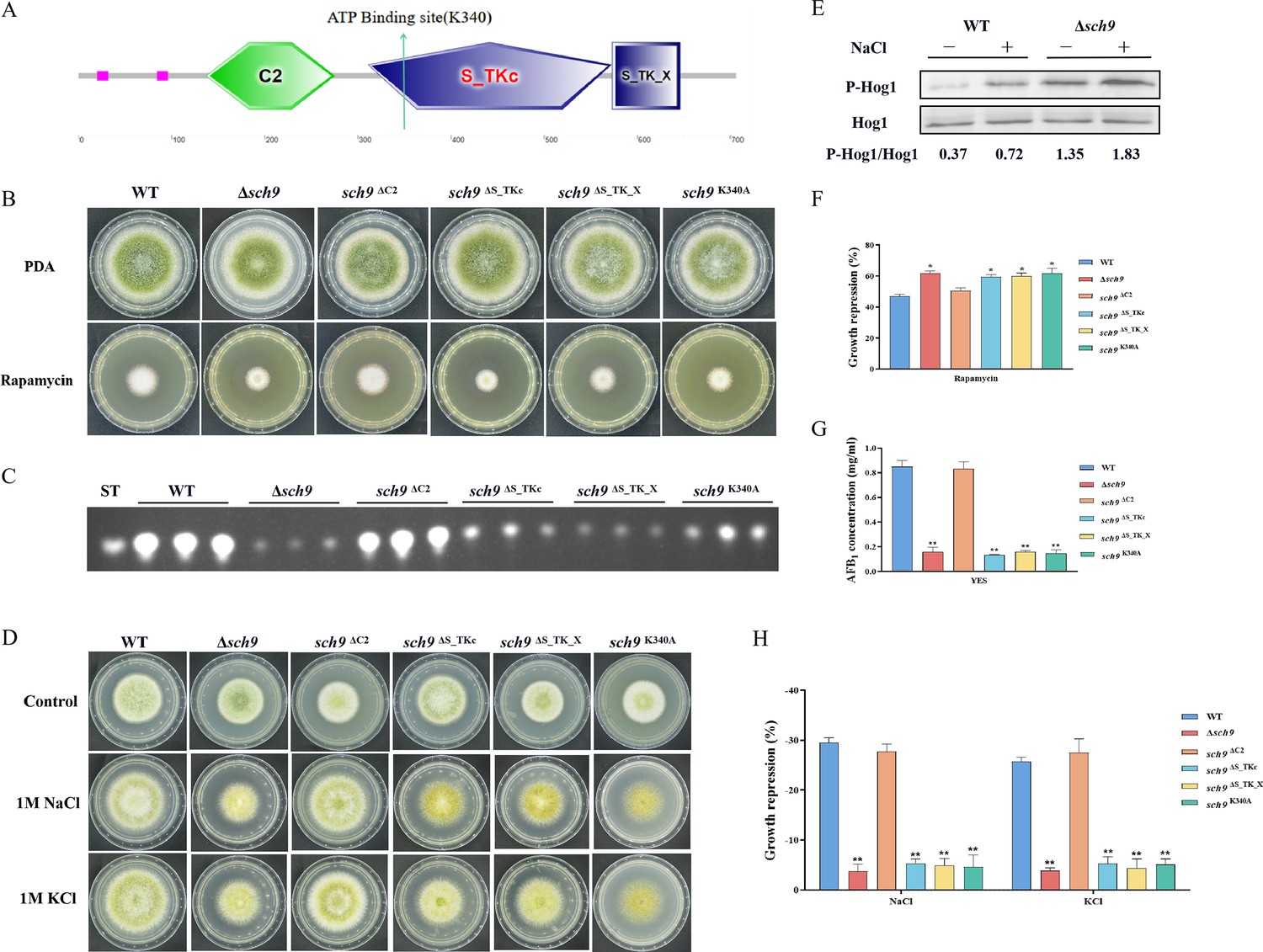
The Sch9 kinase participates in aflatoxin biosynthesis and the high osmolarity glycerol (HOG) pathway.
(A) The structure diagram of the Sch9 kinase. (B) Phenotype of the wild-type (WT), Δsch9, sch9ΔC2, sch9ΔS_TKc, sch9ΔS_TK_X, and sch9K340A strains grown on potato dextrose agar (PDA) medium amended with 100 ng/mL rapamycin at 37°C for 5 days. (C) Thin layer chromatography (TLC) assay of aflatoxin B1 (AFB1) production from the WT, Δsch9, sch9ΔC2, sch9ΔS_TKc, sch9ΔS_TK_X, and sch9K340A strains cultured in yeast extract-sucrose agar (YES) liquid medium at 29°C for 6 days. (D) The phenotypes of the WT, Δsch9, sch9ΔC2, sch9ΔS_TKc, sch9ΔS_TK_X, and sch9K340A strains on yeast extract-glucose agar (YGT) media amended with 1 M NaCl and 1 M KCl for 3 days. (E) The phosphorylation levels of Hog1 in the WT and Δsch9 strains were determined with or without osmotic stress. (F) The growth inhibition rate of the WT, Δsch9, sch9ΔC2, sch9ΔS_TKc, sch9ΔS_TK_X, and sch9K340A strains under rapamycin stress. (G) Quantitative analysis of AFB1 as shown in (C). (H) The growth inhibition rate of the WT, Δsch9, sch9ΔC2, sch9ΔS_TKc, sch9ΔS_TK_X, and sch9K340A strains under osmotic stress. * indicates that the significance level was p≤0.05, ** indicates that the significance level was p≤0.01, based on one-way ANOVA test with three replicates(n=3). Error bars represent the standard error of the mean (SEM).
-
Figure 4—source data 1
Original file for the western blot analysis in Figure 4E (anti-P-Hog1).
- https://cdn.elifesciences.org/articles/89478/elife-89478-fig4-data1-v1.zip
-
Figure 4—source data 2
Original file for the western blot analysis in Figure 4E (anti-Hog1).
- https://cdn.elifesciences.org/articles/89478/elife-89478-fig4-data2-v1.zip
-
Figure 4—source data 3
PDF containing Figure 4E and original scans of the relevant western blot analysis (anti-P-Hog1 and anti-Hog1) with highlighted bands and sample labels.
- https://cdn.elifesciences.org/articles/89478/elife-89478-fig4-data3-v1.zip
In yeast, Sch9 plays an important role in the transcriptional activation of osmostress-inducible genes (Pascual-Ahuir and Proft, 2007). To characterize the roles of Sch9 in stress response, we examined the sensitivity of the sch9 deletion strain to various stresses including osmotic stress and calcium stress. Compared with the WT strain, the ∆sch9 mutant significantly increased sensitivity to calcium stress (Figure 4—figure supplement 1A and B), and decreased sensitivity to osmotic stress induced by NaCl and KCl (Figure 4D and H). In A. flavus, the Sch9 kinase consists of several domains, including the protein kinase C conserved region 2 (C2), protein kinase (S_TKc), AGC-kinase C-terminal (S_TK_X) domain, and an ATP-binding site (Figure 4A). To explore the role of these domains and the ATP-binding site in Sch9 kinase, we constructed deletion strains for both domains and a sch9K340A mutant. The results revealed that the sch9ΔS_TKc strain, sch9ΔS_TK_X strain, and sch9K340A mutant exhibited similar phenotypes in osmotic stress and AFB1 production as the ∆sch9 mutant (Figure 4B–D). Therefore, we speculated that the protein kinase domain, kinase C-terminal domain, and the ATP-binding site at K340 play a crucial role in modulating the impact of Sch9 on aflatoxin biosynthesis and stress response in A. flavus.
To investigate the potential function of Sch9 in the HOG and TOR pathways in A. flavus, we conducted a sensitivity analysis of the Δsch9 strain to rapamycin. Compared to the WT strain, the sch9 deletion strain exhibited a slightly higher sensitivity to rapamycin (Figure 4B and F). The MAKP Hog1 is an element of the HOG pathway. To verify the connection between Sch9 and the HOG-MAPK pathway, the phosphorylation levels of Hog1 kinase were measured under osmotic stress (1 M NaCl). Western blot analysis showed that the phosphorylation level of Hog1 increased significantly in the ∆sch9 strain (Figure 4E). This finding suggested that Sch9 plays a pivotal role in modulating the response of Hog1 to osmotic stress. Taken together, these results reflected that Sch9 regulates osmotic stress response via the HOG pathway in A. flavus.
TapA and TipA participate in the regulation of cell wall stress response
Tap42, a protein that associates with phosphatase 2A, serves as the direct target of the Tor kinase in yeast. Tap42 plays a crucial role in various Tor functions, particularly in the regulation of transcriptional processes (Düvel and Broach, 2004). The genome of A. flavus contains a homolog of tap42 named tapA (AFLA_092770), which is predicted to encode a 355 amino acid protein (sharing 31.9% identity with S. cerevisiae Tap42). To further determine the function of the TapA regulator in the TOR signaling pathway of A. flavus, we used homologous recombination to replace the original promoter with the gpdA promoter, a well-characterized strong promoter, to construct an overexpression mutant (OE::tapA). The transformants of the mutant were verified using PCR. Quantitative expression analysis of the tapA gene in the overexpressed strains revealed a substantial upregulation compared to the WT strain (Figure 5—figure supplement 1). Our results showed that the colony diameter of the OE::tapA mutant was not significantly changed compared to the WT strain (Figure 5A). However, the rapamycin sensitivity assay indicated that the OE::tapA strain displayed increased sensitivity to rapamycin (Figure 5A and D). We found no significant difference in aflatoxin production between the OE::tapA and the WT strains (Figure 5C and E), suggesting that TapA may not play a direct role in the regulation of aflatoxin biosynthesis. In addition, the OE::tapA strain was more sensitive to cell wall and cell membrane stress, including SDS, CFW, and Congo red (CR) (Figure 5B and F). These findings have demonstrated the significantly positive impact of TapA on cell wall stress in A. flavus.
Figure 5 with 1 supplement see all
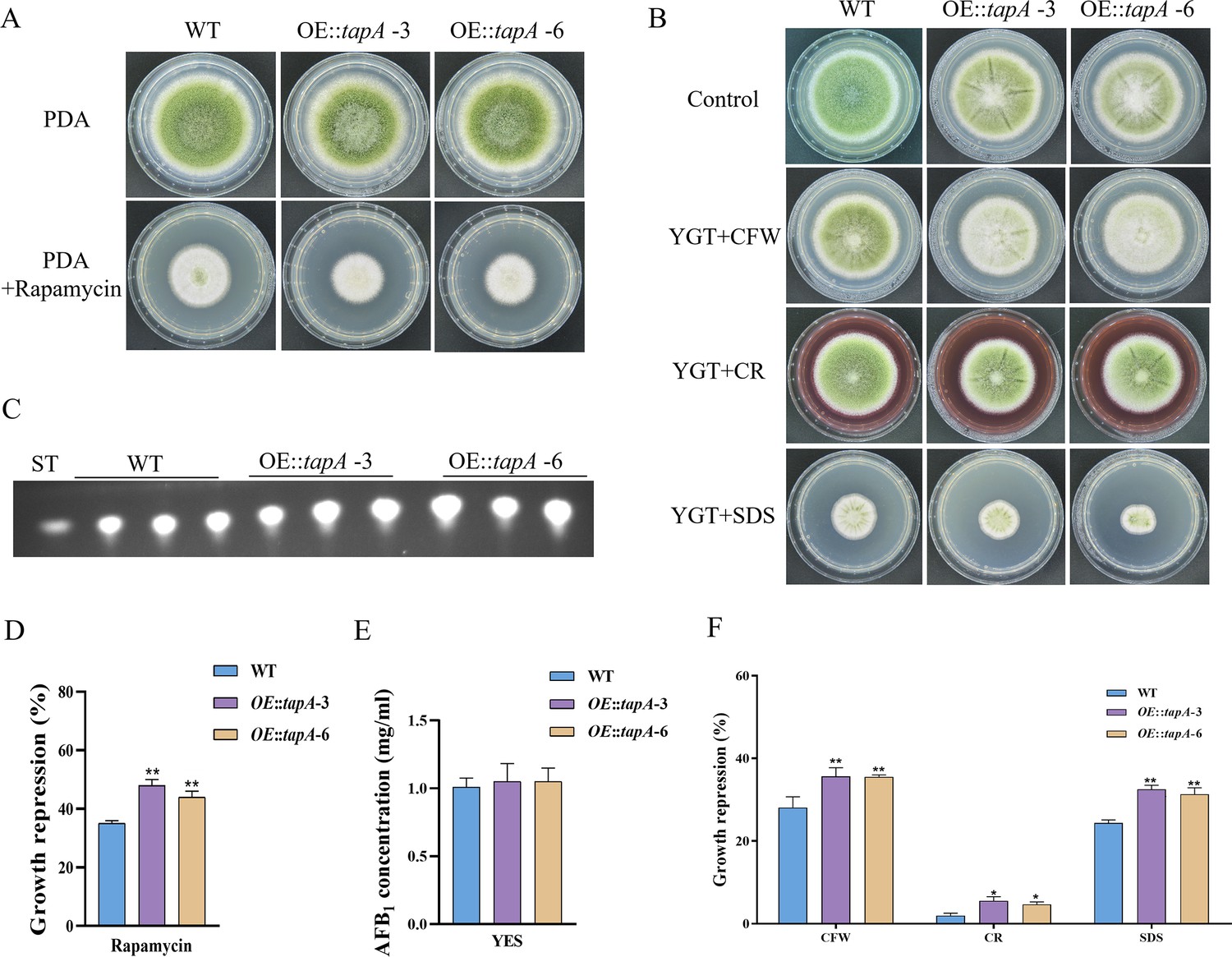
TapA regulates cell wall stress in A. flavus.
(A) Phenotype of the wild-type (WT) and OE::tapA strains grown on potato dextrose agar (PDA) amended with 100 ng/mL rapamycin at 37°C for 5 days. (B) Colony morphology of the WT and OE::tapA strains grown on PDA media supplemented with 200 µg/mL Congo red (CR), 200 µg/mL Calcofluor white (CFW), or 300 µg/mL SDS at 37°C for 5 days. (C) Thin layer chromatography (TLC) assay of aflatoxin B1 (AFB1) production by the WT and OE::tapA strains cultured in yeast extract-sucrose agar (YES) liquid medium at 29°C for 6 days. (D) The growth inhibition rate of the WT and OE::tapA strains under rapamycin. (E) Quantitative analysis of AFB1 as shown in (C). (F) The growth inhibition rate of the WT and OE::tapA strains under cell wall and cell membrane stress. * indicates that the significance level was p≤0.05, ** indicates that the significance level was p≤0.01, based on one-way ANOVA test with three replicates(n=3). Error bars represent the standard error of the mean (SEM).
In S. cerevisiae, Tip41 was identified as a Tap42-interacting protein and has been measured to function as a negative regulator of Tap42, which downregulates TORC1 signaling via activation of PP2A phosphatase (Jacinto et al., 2001). The putative ortholog of tip41 in A. flavus is named tipA (AFLA_047310). The amino acid sequence of TipA exhibits a similarity of 40.49% to Tip41 in S. cerevisiae. To elucidate the function of the TOR signaling pathway protein TipA in A. flavus, we generated the tipA deletion mutants. The ∆tipA strain exhibited a slightly reduced number of conidia produced on PDA compared to the WT strain (Figure 6A and C). Meanwhile, the ∆tipA strain exhibited decreased sensitivity to rapamycin (Figure 6A and E). The ∆tipA strain was unable to produce any sclerotia (Figure 6B and F). Whereas ∆tipA strain do not affect aflatoxin production (Figure 6D and G). In addition, the ∆tipA strain was more sensitive to the cell wall and cell membrane stress, including CR and SDS (Figure 6H and I). Additionally, the expression levels of nsdC, nsdD, and sclR exhibited a decrease in ∆tipA strain compared to the WT strain (Figure 6J). These results indicated that TipA is crucial for the sclerotial formation and cell wall stress in A. flavus.
Figure 6
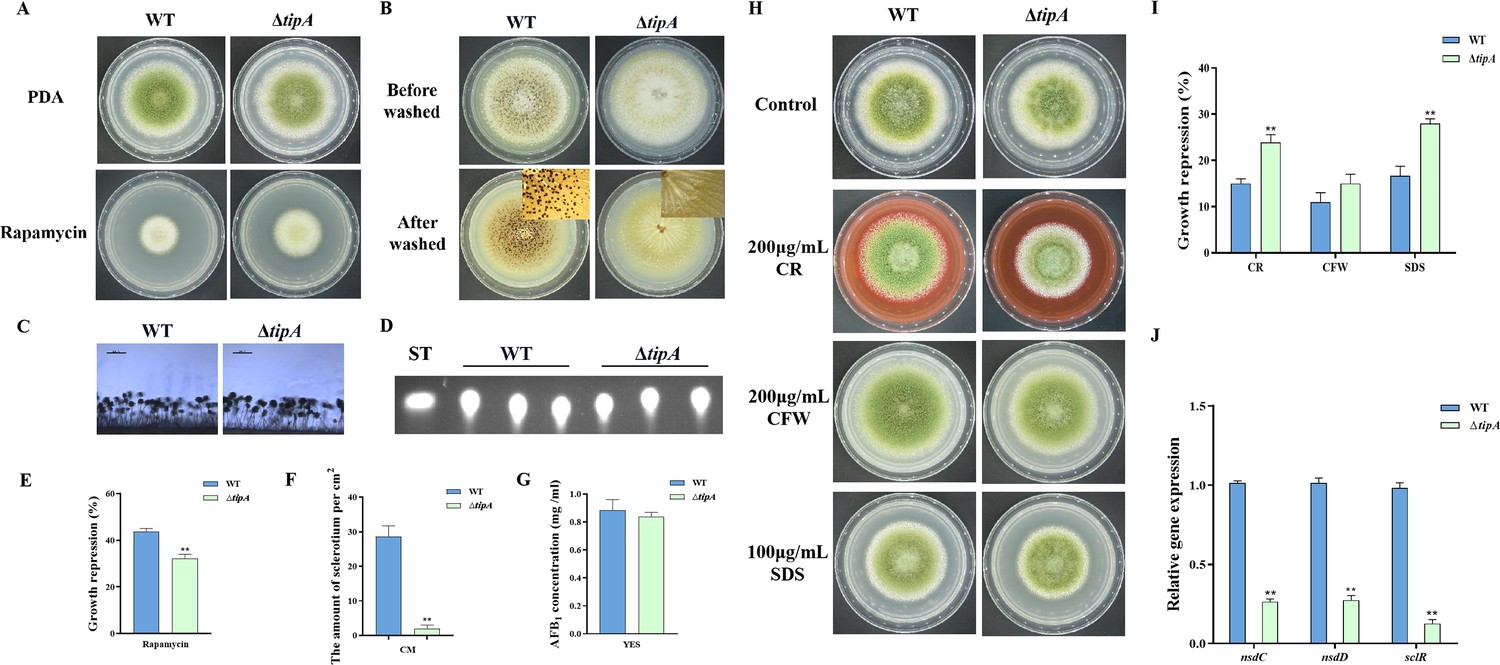
TipA regulates sclerotia development and cell wall stress in A. flavus.
(A) Phenotype of the wild-type (WT) and ∆tipA strains grown on potato dextrose agar (PDA) amended with 100 ng/mL rapamycin at 37°C for 5 days. (B) Phenotypic characterization of the WT and ∆tipA strains grown on CM medium at 37°C for 7 days. (C) Morphology of conidiophores in the WT and ∆tipA strains observed by microscope. (D) Thin layer chromatography (TLC) assay of aflatoxin B1 (AFB1) production by the WT and ∆tipA strains cultured in yeast extract-sucrose agar (YES) liquid medium at 29°C for 6 days. (E) The growth inhibition rate of the WT and ∆tipA strains under rapamycin stress. (F) Amount of sclerotia produced by the WT and ∆tipA strains. (G) AFB1 quantitative analysis of the WT and ∆tipA strains described in (D). (H) Colony morphology of the WT and ΔtipA strains grown on PDA media supplemented with 200 µg/mL Congo red (CR), 200 µg/mL Calcofluor white (CFW), and 100 µg/mL SDS at 37°C for 5 days. (I) The growth inhibition rate of the WT and ΔtipA strains under cell wall and cell membrane stress. (J) Relative expression levels of sclerotia formation genes in the WT and ΔtipA strains. ** indicates that the significance level was p≤0.01, based on one-way ANOVA test with three replicates(n=3). Error bars represent the standard error of the mean (SEM).
Phosphatase SitA and Ppg1 are involved in growth, conidiation, and sclerotial formation
In S. cerevisiae, Tap42 has been identified to interact with the catalytic subunit of type 2A protein phosphatases, including Pph3, Pph21, and Pph22, as well as the type 2A-like phosphatases Sit4 and Ppg1 (Wang et al., 2003). Phosphatase sitA (AFLA_108450) and ppg1 (AFLA_132610) were identified in A. flavus, encoding putative orthologs of sit4 and ppg1 in S. cerevisiae. To determine the biological functions of phosphatases SitA and Ppg1 in A. flavus, we disrupted the sitA and ppg1 by homologous recombination strategy. To further investigate the relationship between phosphatases SitA and Ppg1, we additionally generated complemented and double deletion mutants. The vegetative development phenotype analysis showed that the ∆sitA, ∆ppg1, and ∆sitA/∆ppg1 strains grew significantly more slowly than the WT strain on PDA (Figure 7A and D). Compared with the WT strain, the ∆sitA, ∆ppg1, and ∆sitA/∆ppg1 strains were more sensitive to rapamycin (Figure 7A). In addition, the ∆sitA, ∆ppg1, and ∆sitA/∆ppg1 strains formed lower number of conidiophores (Figure 7A). To gain further insight into the role of SitA and Ppg1 in the process of conidiation, qRT-PCR was employed to determine the transcript levels of conidiation-related genes (abaA and brlA). The expression levels of the abaA and brlA genes were significantly downregulated in the ∆sitA, ∆ppg1, and ∆sitA/∆ppg1 mutants when compared to the WT and complemented strains (Figure 7G). The analysis of sclerotial formation revealed that the ∆sitA, ∆ppg1, and ∆sitA/∆ppg1 mutants exhibited a complete inability to produce any sclerotia (Figure 7B). To further prove these findings, we conducted qRT-PCR to determine the expression levels of three regulators (nsdC, nsdD, and sclR) of sclerotia development. Our findings indicated that the expression levels of nsdC, nsdD, and sclR were all significantly reduced in the ∆sitA, ∆ppg1, and ∆sitA/∆ppg1 mutants (Figure 7H), which is consistent with the observed decrease in sclerotia production in the ∆sitA, ∆ppg1, and ∆sitA/∆ppg1 mutants. These results showed that SitA and Ppg1 are crucial for hyphal development, conidiation, and sclerotia formation in A. flavus.
Figure 7
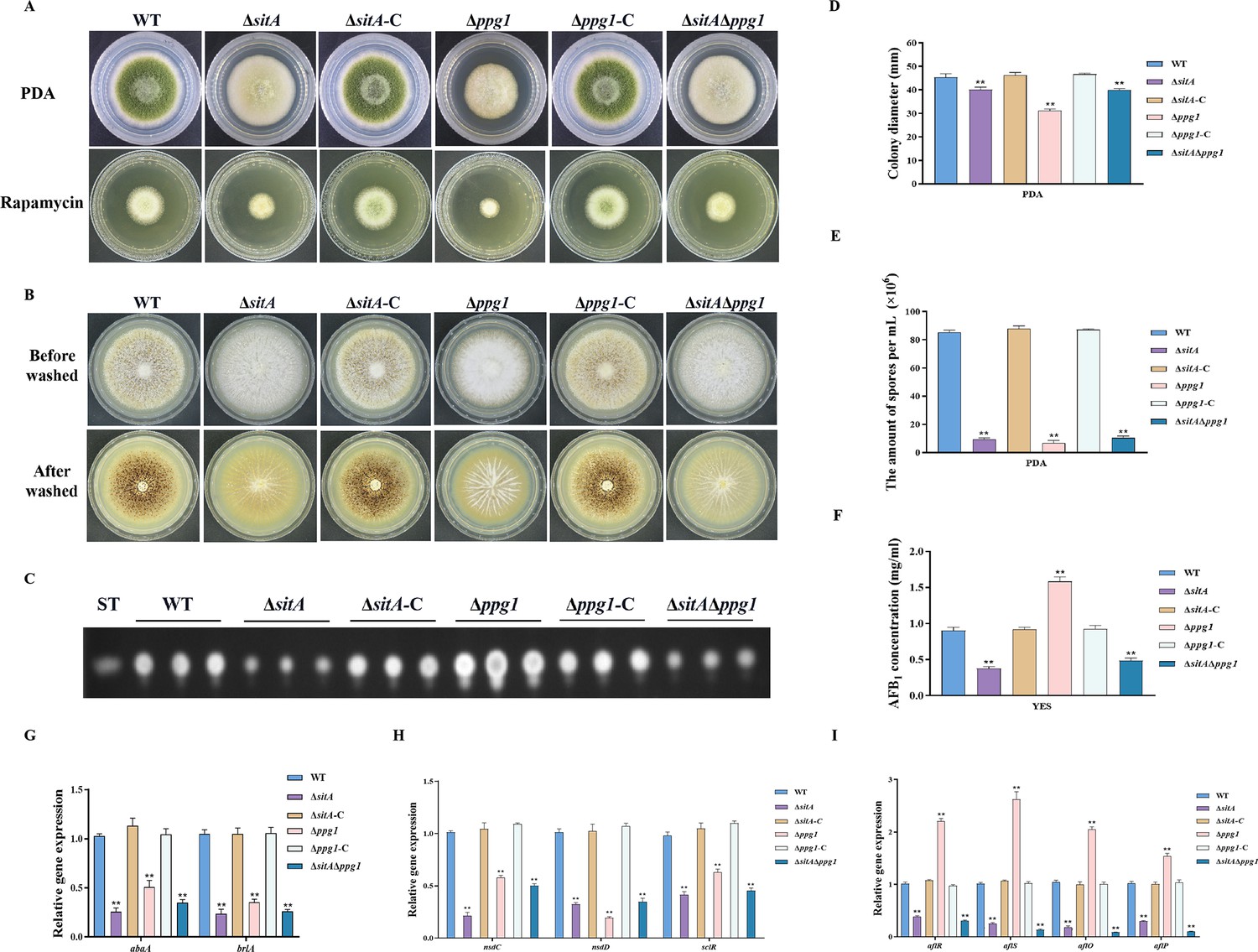
The impact of the phosphatases SitA and Ppg1 on the growth, conidiation, and aflatoxin biosynthesis in A. flavus.
(A) Colony morphology of the wild-type (WT), single knockout, and double knockout strains cultured on potato dextrose agar (PDA) medium amended with 100 ng/mL rapamycin at 37°C for 5 days. (B) Colony morphology of the WT, single knockout, and double knockout strains cultured on CM medium at 37°C for 7 days. (C) Thin layer chromatography (TLC) analysis of aflatoxin B1 (AFB1) production from the WT, single knockout, and double knockout strains cultured in yeast extract-sucrose agar (YES) liquid medium at 29°C for 6 days. (D) Growth diameter of the WT, single knockout, and double knockout strains on PDA media. (E) Conidial quantification of the WT, single knockout, and double knockout strains. (F) AFB1 quantitative analysis of the WT, single knockout, and double knockout strains. (G) Relative expression levels of conidia synthesis genes in the WT, single knockout, and double knockout strains. (H) Relative expression levels of sclerotia synthesis genes in the WT, single knockout, and double knockout strains. (I) Relative expression levels of aflatoxin biosynthesis genes in the WT, single knockout, and double knockout strains. ** indicates that the significance level was p≤0.01, based on one-way ANOVA test with three replicates(n=3). Error bars represent the standard error of the mean (SEM).
SitA and Ppg1 play crucial roles in aflatoxin biosynthesis and pathogenicity
We assayed AFB1 biosynthesis in the ∆sitA, ∆ppg1, and ∆sitA/∆ppg1 mutants. TLC analyses revealed that the ∆sitA and ∆sitA/∆ppg1 strains exhibited a decrease in AFB1 production, whereas the ∆ppg1 strain showed a significant increase in AFB1 production compared to the WT and complemented strains (Figure 7C and F). The expression levels of genes related to aflatoxin synthesis, including structural genes (aflO and aflP) and regulatory genes (aflR and aflS), were determined by qRT-PCR in the ∆sitA, ∆ppg1, and ∆sitA/∆ppg1 mutants. The results revealed a significant downregulation of these genes in the ∆sitA and ∆sitA/∆ppg1 mutants, while upregulated in the ∆ppg1 mutant, compared to the WT and complemented strains (Figure 7I). These results indicated that SitA and Ppg1 play different roles in regulating aflatoxin biosynthesis in A. flavus.
We conducted pathogenicity tests on peanuts and maize to examine the pathogenicity of the phosphatases SitA and Ppg1 on crops. The results showed that the WT and complemented strains demonstrated complete virulence on all peanut and maize seeds. In contrast, the ∆sitA, ∆ppg1, and ∆sitA/∆ppg1 mutants displayed compromised colonization on peanut and maize seeds (Figure 8A). In addition, the number of conidial productions in these mutants on the infected peanut seeds also dramatically decreased compared to those of the WT and complemented strains (Figure 8C). The TLC analyses revealed that the ∆ppg1 mutant exhibited increased production of AFB1 in peanuts and maize. Conversely, the levels of aflatoxin produced by the ∆sitA and ∆sitA/∆ppg1 mutants on peanut and maize were significantly reduced compared to the WT and complemented strains (Figure 8B and D). This observation aligns with the above results of aflatoxin biosynthesis from all mutant strains in the yeast extract-sucrose agar (YES) medium. All these data showed that SitA and Ppg1 are crucial for crop seeds’ pathogenicity.
Figure 8
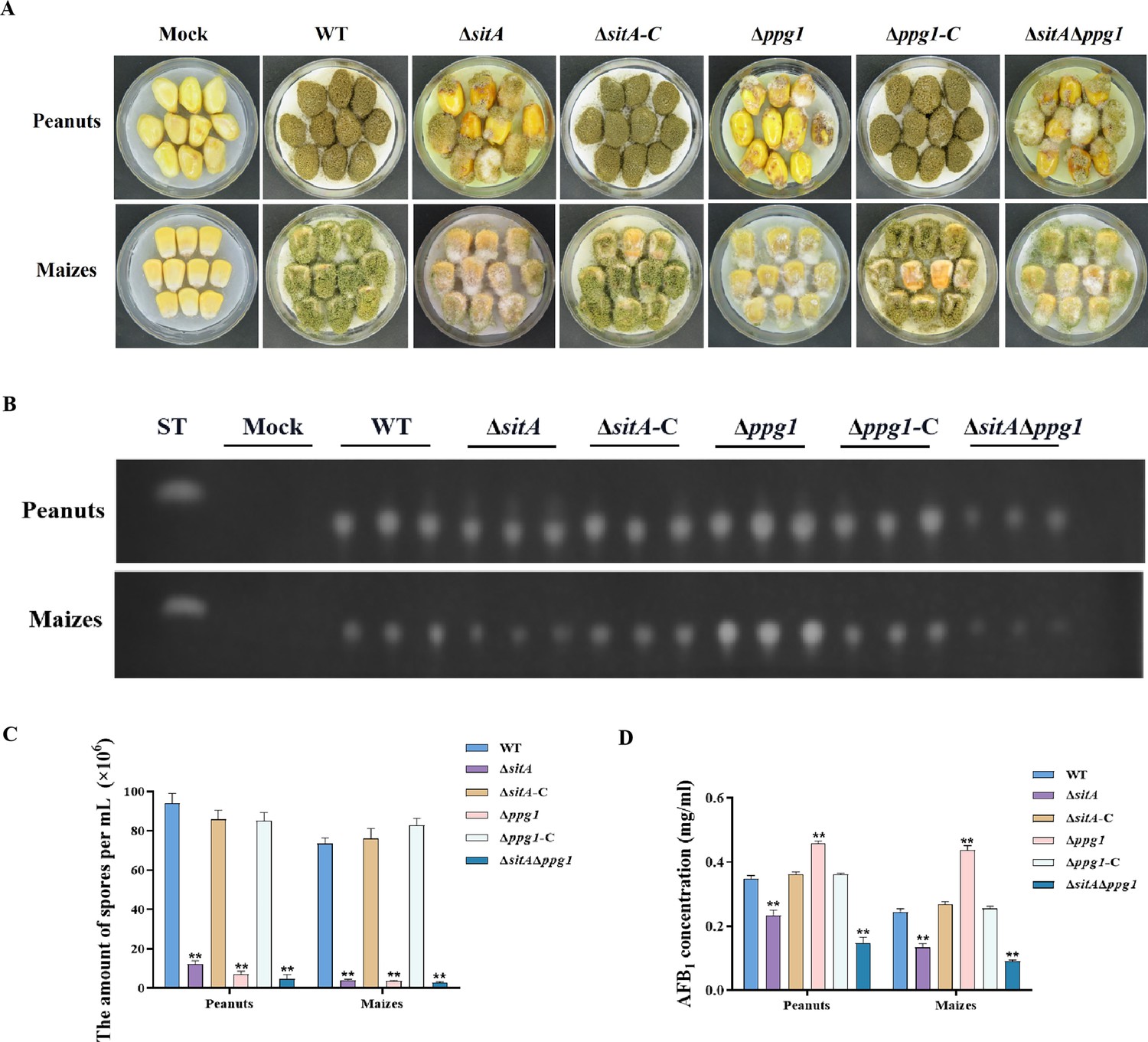
Pathogenicity analysis of the phosphatases SitA and Ppg1 in A. flavus.
(A) Phenotypes of peanut and maize infected by the wild-type (WT), single knockout, and double knockout strains. (B) Thin layer chromatography (TLC) was used to detect the aflatoxin B1 (AFB1) from peanut and maize infected by the WT, single knockout, and double knockout strains. (C) Quantitative analysis of conidia production from peanut and maize infected by the WT, single knockout, and double knockout strains. (D) Quantitative analysis of AFB1 from peanut and maize infected by the WT, single knockout, and double knockout strains. ** indicates that the significance level was p≤0.01, based on one-way ANOVA test with three replicates(n=3). Error bars represent the standard error of the mean (SEM).
SitA and Ppg1 are involved in regulating CWI
To explore the role of phosphatases SitA and Ppg1 in response to cell wall stress, all strains were cultivated on PDA amended with CFW, CR, and SDS. We found that the relative growth inhibition of ∆sitA, ∆ppg1, and ∆sitA/∆ppg1 mutants induced by cell wall stress was significantly increased compared to the WT and complemented strains (Figure 9A and B). Further investigations using qRT-PCR revealed a notable reduction in the expression levels of genes related to the synthesis of cell walls, specifically chitin synthase genes chsA, chsD, and chsE in the ∆sitA, ∆ppg1, and ∆sitA/∆ppg1 strains (Figure 9C). Western blot analysis also revealed that the phosphorylation level of Slt2 was significantly elevated in the ∆sitA, ∆ppg1, and ∆sitA/∆ppg1 strains under cell wall stress (200 μg/mL CFW) (Figure 9D). In conclusion, the phosphatases SitA and Ppg1 play a crucial role in regulating CWI.
Figure 9
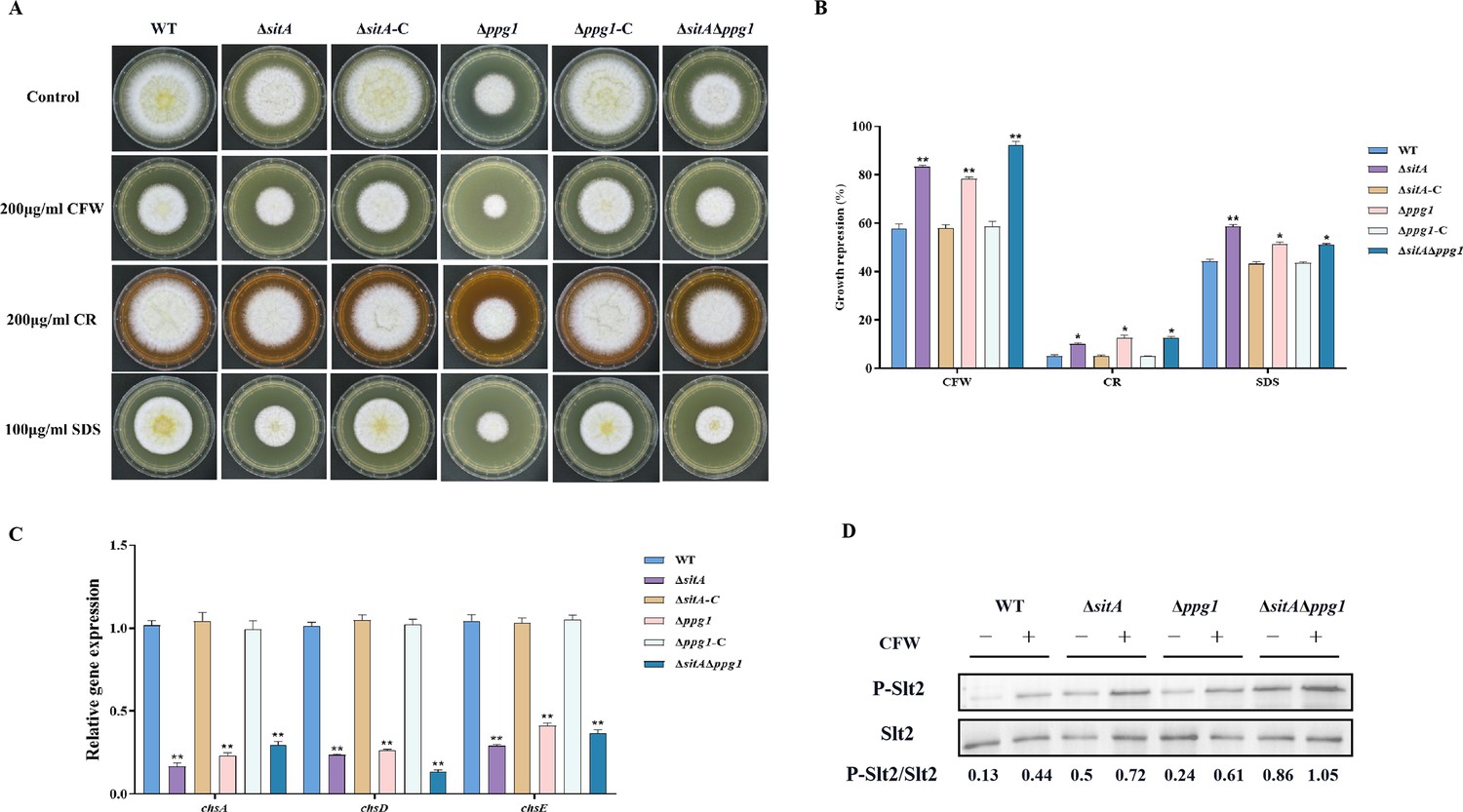
Sensitivity of phosphatases SitA and Ppg1 to cell wall damaging agents in A. flavus.
(A) Morphology of the wild-type (WT), single knockout, and double knockout strains grown on yeast extract-sucrose agar (YES) media supplemented with 200 µg/mL Congo red (CR), 200 µg/mL Calcofluor white (CFW), or 100 µg/mL SDS at 37°C for 5 days. (B) The growth inhibition rate of the WT, single knockout, and double knockout strains under cell wall and cell membrane stress. (C) Relative expression levels of cell wall synthesis genes (chsA, chsD, and chsE) in the WT, single knockout, and double knockout strains. (D) The phosphorylation level of Slt2 in the WT, single knockout, and double knockout strains was detected with or without cell wall stress. * indicates that the significance level was p≤0.05, ** indicates that the significance level was p≤0.01, based on one-way ANOVA test with three replicates(n=3). Error bars represent the standard error of the mean (SEM).
-
Figure 9—source data 1
Original file for the western blot analysis in Figure 9D (anti-P-Slt2).
- https://cdn.elifesciences.org/articles/89478/elife-89478-fig9-data1-v1.zip
-
Figure 9—source data 2
Original file for the western blot analysis in Figure 9D (anti-Slt2).
- https://cdn.elifesciences.org/articles/89478/elife-89478-fig9-data2-v1.zip
-
Figure 9—source data 3
PDF containing Figure 9D and original scans of the relevant western blot analysis (anti-P-Slt2 and anti-Slt2) with highlighted bands and sample labels.
- https://cdn.elifesciences.org/articles/89478/elife-89478-fig9-data3-v1.zip
The phosphatase complex Nem1/Spo7 is important for growth, aflatoxin biosynthesis, and LD biogenesis
LDs are highly dynamic spherical organelles, which appear in the cytosol of most eukaryotic cell types (Sánchez-Álvarez et al., 2022). Rapamycin treatment led to the accumulation of LDs in various filamentous fungi (Liu et al., 2019). The FgTOR signaling cascade is implicated in the regulation of FgNem1 phosphorylation profile, predominantly mediated by the sequential activation of the FgPpg1/Sit4 and FgCak1 kinase modules, along with additional kinases in F. graminearum (Liu et al., 2019). In this study, we observed that the treatment of hyphae with rapamycin resulted in the accumulation of LD biogenesis (Figure 10A). This finding suggested that the TOR pathway may regulate the LD biogenesis. However, the underlying regulatory mechanism of LD biosynthesis remains largely unknown in A. flavus. To investigate the regulatory mechanism of the TOR pathway in A. flavus that controls LD biogenesis, we examined LD accumulation in ∆sch9, ∆sitA, and ∆ppg1 mutants. BODIPY staining assays revealed that LD biogenesis was not observed in the ∆sitA and ∆ppg1 strains upon treatment with rapamycin, in contrast to the WT and ∆sch9 strains (Figure 10A–D). These results indicated that LD biogenesis induced by rapamycin is predominantly reliant on the phosphatases SitA and Ppg1 in A. flavus. In F. graminearum, the phosphatase FgNem1/FgSpo7 complex was involved in the induction of LD biogenesis by rapamycin (Liu et al., 2019). Phosphatase nem1 (AFLA_002649) and spo7 (AFLA_028590) were identified in A. flavus, which encode putative orthologs of nem1 and spo7 in F. graminearum. To examine the role of the nem1 and spo7 genes in A. flavus, we generated ∆nem1 and ∆spo7 mutants using homologous recombination. The ∆nem1 and ∆spo7 mutants displayed a notable reduction in both vegetative growth and conidiation compared to the WT strain (Figure 11A, C, and D). TLC assay revealed a significantly decreased aflatoxin production in the ∆nem1 and ∆spo7 compared to the WT strain (Figure 11B). In addition, BODIPY staining showed no visible LDs in the hyphae of ∆nem1 and ∆spo7 after rapamycin treatment (Figure 11E). Taken together, these results indicated that Nem1/Spo7 are involved in vegetative growth, conidiation, aflatoxin production, and LD biogenesis.
Figure 10
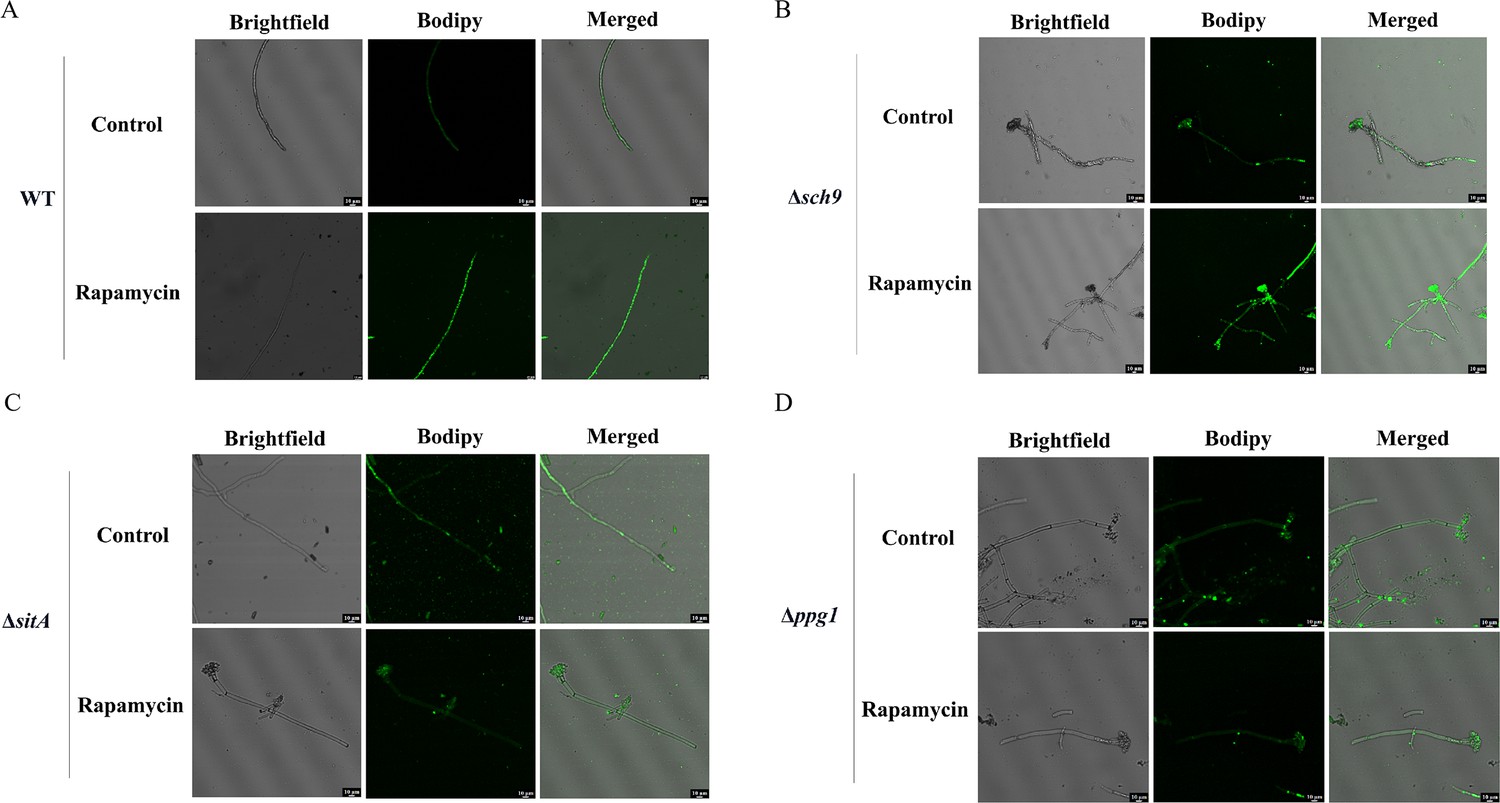
Phosphatases SitA and Ppg1 are involved in regulating lipid droplet biogenesis.
(A) The intracellular lipid droplets were stained by boron dipyrromethene difluoride (BODIPY) in hyphae of the wild-type (WT) strain and observed under fluorescence microscopy. The phenotype of lipid droplets accumulation in the mycelia of the WT strain treated with or without 100 ng/mL rapamycin for 6 hr (Scale bar, 10 μm). (B) Phenotype of lipid droplets accumulation in the mycelia of the Δsch9 strain treated with or without 100 ng/mL rapamycin for 6 hr (Scale bar, 10 μm). (C) Phenotype of lipid droplets accumulation in the mycelia of the ΔsitA strain treated with or without 100 ng/mL rapamycin for 6 hr (Scale bar, 10 μm). (D) Phenotype of lipid droplets accumulation in the mycelia of the Δppg1 strain treated with or without 100 ng/mL rapamycin for 6 hr (Scale bar, 10 μm).
Figure 11
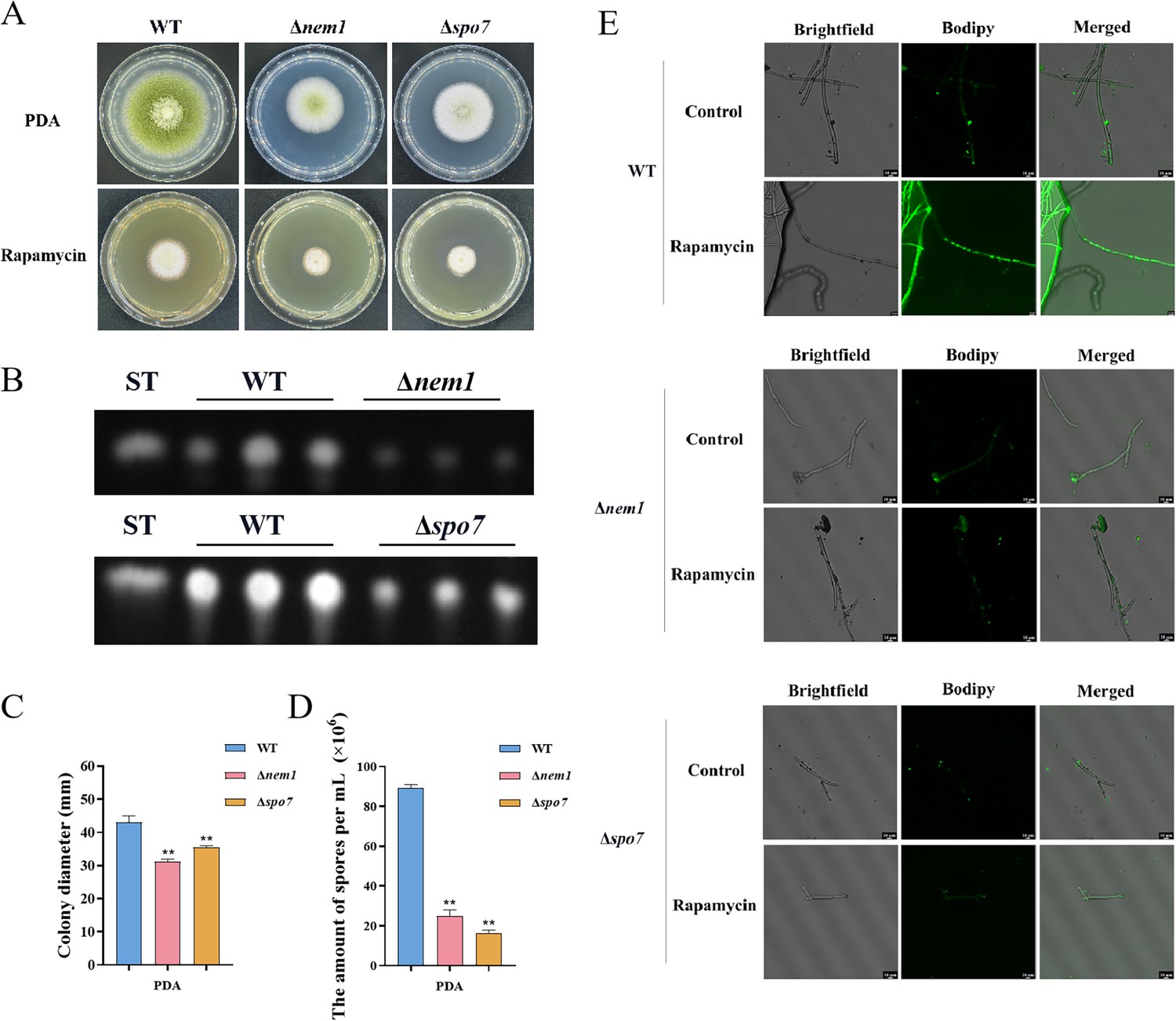
Phosphatase complex Nem1/Spo7 play significant role in growth, conidiation, aflatoxin, and lipid droplet biogenesis.
(A) Phenotype of the wild-type (WT), ∆nem1, and ∆spo7 strains grown on potato dextrose agar (PDA) amended with 100 ng/mL rapamycin at 37°C for 5 days. (B) Thin layer chromatography (TLC) assay of aflatoxin B1 (AFB1) production from the WT, ∆nem1, and ∆spo7 strains cultured in yeast extract-sucrose agar (YES) liquid medium at 29°C for 6 days. (C) Growth diameter of the WT, ∆nem1, and ∆spo7 strains on PDA media. (D) Statistical analysis of the sporulation by the WT, ∆nem1, and ∆spo7 strains. (E) Phenotype of lipid droplets accumulation in the mycelia of the WT, ∆nem1, and ∆spo7 strains treated with or without 100 ng/mL rapamycin for 6 hr (Scale bar, 10 μm). ** indicates that the significance level was p≤0.01, based on one-way ANOVA test with three replicates(n=3). Error bars represent the standard error of the mean (SEM).
Discussion
Aflatoxins are a class of toxic secondary metabolites synthesized by A. flavus, including aflatoxin B1, B2, G1, and G2, among which AFB1 has been regarded as the most pathogenic and toxic aflatoxin (Gizachew et al., 2019). Therefore, understanding the regulatory mechanisms involved in fungal secondary metabolites biosynthesis would be instrumental for effectively addressing aflatoxin control in A. flavus. In this study, we demonstrated that the TOR signaling pathway plays a pivotal role in regulating growth, sporulation, sclerotia formation, aflatoxin production, and various stress responses in A. flavus. Through conducting functional studies on genes within the TOR signaling pathway, we explored the conservation and complexity exhibited by the TOR signaling pathway in A. flavus. In addition, we constructed a crosstalk network between the TOR and other signaling pathways (HOG, CWI) and analyzed the specific regulatory process. We proposed that the TOR signaling pathway interacts with multiple signaling pathways, including HOG and CWI pathways, to modulate the cellular responses to diverse environmental stresses (Figure 12).
Figure 12
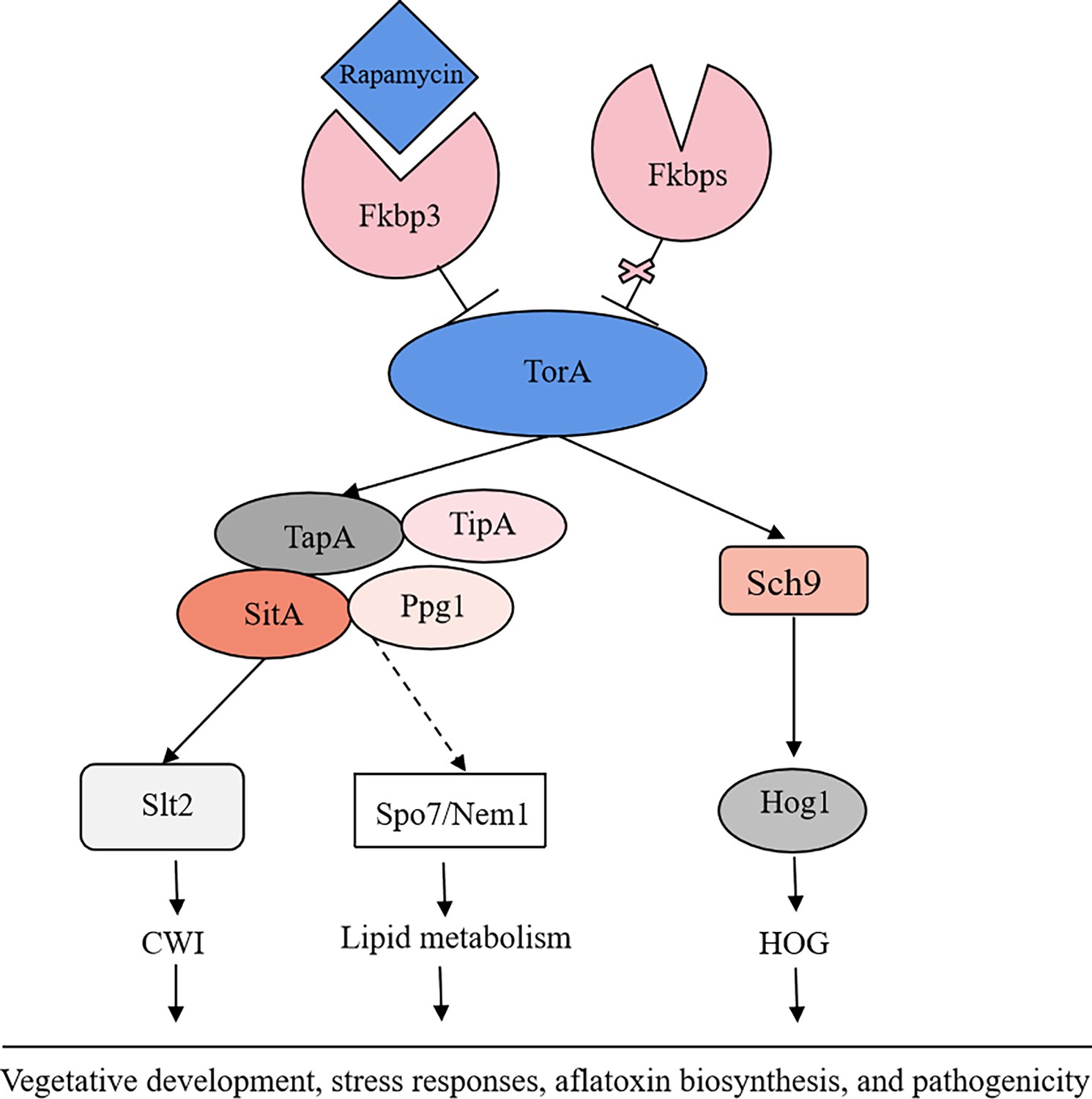
The proposed model of the target of rapamycin (TOR) pathway in A. flavus.
Based on the above results, we propose a hypothesis regarding the TOR signaling pathway model in A. flavus. Rapamycin forms a complex with Fkbp3, and this particular complex can bind to the TorA kinase, thereby impeding its regular functionality. Sch9, functioning as a downstream element of the TorA kinase, regulates aflatoxin biosynthesis and high osmolarity glycerol (HOG) signaling pathway. As another important target of the TorA kinase, TapA-phosphatase complex is involved in the regulation of growth, sporulation, sclerotia, aflatoxin production, cell wall integrity (CWI) signaling pathway, lipid droplet synthesis, and other processes. In conclusion, the TOR signaling pathway plays a crucial role in various aspects of A. flavus, including vegetative development, stress response, aflatoxin biosynthesis, and pathogenicity.
Our findings indicated that the TOR signaling pathway is pivotal in A. flavus, we have observed that rapamycin has a significant impact on the growth and sporulation of A. flavus (Figure 1A). Additionally, it exhibits a strong inhibitory effect on sclerotia production and aflatoxin biosynthesis (Figure 1B and C). The inhibitory effect has been documented in various fungal species, including C. albicans (Cruz et al., 2001), Cryptococcus neoformans (Cruz et al., 2001), P. anserine (Dementhon et al., 2003), A. nidulans (Fitzgibbon et al., 2005), M. oryzae (Qian et al., 2018), and F. graminearum (Yu et al., 2014). This suggested that the TOR signaling pathway may have a conserved function in filamentous fungi. The number of FK506-binding proteins varies among different species: there are four Fkbps proteins in S. cerevisiae and three Fkbps proteins in S. pombe (Galat, 2013). In S. cerevisiae, rapamycin can bind to Fkbp protein, resulting in the irreversible inhibition of the G1 phase of the cell cycle and the regulation of cell growth (Kasahara, 2021). In A. nidulans, the fkbp12 gene homolog fprA displays considerable sequence similarity with homologs identified in the S. cerevisiae (Fitzgibbon et al., 2005). In this study, based on the homologous alignment of the protein sequence of FprA, it was observed that four fkbp genes are present in A. flavus. Among these, deletion of fkbp3 exhibited enhanced tolerance to rapamycin (Figure 2A). So we speculated that rapamycin forms a complex with Fkbp3 in A. flavus, subsequently targeting downstream target genes to exert its biological function. We found that Fkbp3 is involved in sclerotia formation and aflatoxin biosynthesis in A. flavus (Figure 2A, Figure 2—figure supplement 2), and subsequent analysis revealed that the underlying mechanism may be associated with succinylation modification. The succinylation sites of Fkbp3 were identified through the utilization of succinylation proteomics data. By constructing point mutant strains and conducting phenotype experiments, we found that the succinylation of Fkbp3 at residue K19 is involved in rapamycin resistance and aflatoxin biosynthesis (Figure 2E and G). Our results provided new insights to the understanding of post-translational modifications in the regulation of TOR pathway components. Previous studies have demonstrated that Fkbp12 has been identified as the receptor for FK506 and rapamycin, which inhibits calcineurin and TORC1, respectively (Kasahara, 2021). The deletion of Fkbp12 confers the resistance to rapamycin in C. albicans (Cruz et al., 2001) and C. neoformans (Cruz et al., 2001). The entomopathogenic fungus Beauveria bassiana contains three putative fkbp genes, and disruption of Bbfkbp12 significantly increases the resistance to rapamycin and FK506 (Li et al., 2020). The plant pathogenic fungus F. graminearum harbors three Fkbps proteins, among which deletion of Fgfkbp12 leads to resistance to rapamycin (Yu et al., 2014). Thus, we proposed that these Fkbps may exhibit analogous functionalities in conferring resistance to rapamycin. In B. cinerea, deletion of Bcfkbp12 caused a reduction of the virulence of strain T4, while knockout of the fkbp12 gene did not affect the pathogenic development of strain B05.10 (Meléndez et al., 2009). These findings demonstrated that the fkbp12 homologous genes exhibited conservation in terms of rapamycin resistance. However, there is significant variation in other biological functions among fungal species, which may be attributed to species or strain evolutionary divergence. Our study indicated the pivotal role of Fkbp3 in modulating the effects of rapamycin on A. flavus, Fkbp3 is also crucial for aflatoxin biosynthesis. Targeted inhibition of Fkbp3 could potentially suppress aflatoxin production, given its significance, Fkbp3 can be developed as a fungicide target to reduce aflatoxin biosynthesis, thereby reducing the risk of aflatoxin contamination in agricultural products.
The Tor kinase functions as a fundamental component of the TOR signaling pathway, which exhibits evolutionary conservation across fungi, plants, and mammals (Xiong and Sheen, 2014). In S. cerevisiae, the Tor kinase can interact with a diverse range of proteins, resulting in the formation of complex structures known as TORC1 and TORC2 (Weisman, 2016). TORC1 can detect various signals, including nutrients, growth factors, and environmental stress. It plays a crucial role in regulating cellular processes such as gene transcription, protein translation, ribosome synthesis, and autophagy (Loewith and Hall, 2011). A characteristic that distinguishes TORC1 is its interaction with KOG1 (raptor in mammals) and its sensitivity to the rapamycin-FKBP12 complex (Adami et al., 2007). The Tor kinase is widely conserved both in terms of its structural characteristics and its role as the target of Fkbp-rapamycin (Tatebe and Shiozaki, 2017). Typically, the Tor kinase exhibits similar structural characteristics, including the HEAT-like, FAT, FRB, kinase, and FATC domains (Tatebe and Shiozaki, 2017). The FRB domain is the binding region of the Fkbp-rapamycin complex, and rapamycin resistance is usually caused by the disruption of the FRB domain (Loewith and Hall, 2011). Homology analysis and phylogenetic studies have identified a single predicted torA gene within the A. flavus genome. To elucidate the functional role of torA in A. flavus, our initial approach involved an attempt to generate an torA deletion strain through homologous recombination. However, these efforts were unsuccessful. We speculated that the deletion of the torA gene may be lethal in A. flavus. This observation aligns with the situation observed in F. graminearum (Yu et al., 2014). Therefore, to investigate the function of torA in A. flavus, we constructed a mutant strain (xylPtorA) using an xylose-inducible promoter, which allows for conditional induction with the addition of xylose. We observed that the xylPtorA mutant is completely unable to grow in YGT medium without xylose, whereas partial growth is restored in YXT medium with the addition of xylose (Figure 3A). These results indicated that TorA is essential for the growth of A. flavus. We also discovered that the TorA kinase is involved in the vegetative growth, asexual development, sclerotial, and aflatoxin production of A. flavus (Figure 3A and B, Figure 3—figure supplement 2). In addition, the the xylPtorA mutant shows a high sensitivity to rapamycin and SDS (Figure 3A). Our findings have demonstrated that the TorA kinase plays a crucial role in the processes of morphogenesis, aflatoxin biosynthesis, and multiple stress responses in A. flavus. Previous findings demonstrated that the Tor kinase could function as a molecular switch connecting an iron cue to defend against pathogen infection in C. elegans (Ma et al., 2021). In A. fumigatus, the Tor kinase represents a central regulatory node controlling genes and proteins involved in nutrient sensing, stress response, cell cycle progression, protein biosynthesis, and degradation, as well as adaptation to low iron conditions. In summary, the Tor kinase acts as a central regulator in the TOR signaling pathway, playing a crucial role in the regulation of the cell cycle, protein synthesis, and cellular energy metabolism. Its influence on key cellular processes makes it a promising target for future research to control fungal growth and aflatoxin production.
The serine/threonine protein kinase Sch9, a critical downstream effector of TORC1 in S. cerevisiae, has been extensively studied for its roles in stress resistance, longevity, and nutrient sensing (Pascual-Ahuir and Proft, 2007). In addition, the TORC1-Sch9 pathway is also recognized as crucial mediator of chronological lifespan (Pascual-Ahuir and Proft, 2007). In filamentous fungi, an intricate network of interactions exists between the TOR and HOG pathways. In F. graminearum (Gu et al., 2015), FgSch9 and FgHog1 mutants displayed increased susceptibility to osmotic and oxidative stresses. The phosphorylation level of FgHog1 increased significantly in FgSch9 mutant. Additionally, the affinity capture-MS assay showed that the Tor kinase was among the proteins co-purifying with FgSch9. These results suggested that FgSch9 serves as an intermediary for the TOR and HOG signaling pathways (Gu et al., 2015). Furthermore, FgSch9 plays a crucial role in the regulation of various biological processes such as hyphal differentiation, asexual development, virulence, and DON biosynthesis (Gu et al., 2015). In A. fumigatus, the SchA mutant was sensitive to rapamycin, high concentrations of calcium and hyperosmotic stress, and SchA was involved in iron metabolism. The SchA mutant exhibited enhanced phosphorylation of SakA in response to osmotic stress (Alves de Castro and Dos Reis, 2016). In this study, we discovered that Sch9 is involved in regulating aflatoxin biosynthesis (Figure 4C), and responses to osmotic and rapamycin stress (Figure 4B and D). The S_TKc domain deletion strain (sch9ΔS_TKc), S_TK_X domain deletion strain (sch9ΔS_TK_X), and sch9K340A mutant displayed a similar phenotype to the ∆sch9 strain (Figure 4B–D). These findings revealed that Sch9 plays a significant role in the biological synthesis of aflatoxin and in responding to various stresses. This role is primarily mediated through its kinase domain, kinase C domain, and ATP binding site K340. We also found that the phosphorylation level of Hog1 increased significantly in the ∆sch9 strain under osmotic stress (Figure 4E). Based on the above results, we hypothesized that Sch9 was involved in the osmotic stress response by modulating the HOG pathway in A. flavus.
The phosphorylation state of Tap42 is crucial for its interaction with PP2A subunits and is regulated by a balance between Tor-mediated phosphorylation and Cdc55/Tpd3-mediated dephosphorylation in yeast (Jiang and Broach, 1999). The genome of A. flavus contains a homolog of tap42 named tapA (AFLA_092770), which is predicted to encode a 355 amino acid protein (sharing 31.9% identity with S. cerevisiae Tap42). To further determine the function of the TapA regulator in the TOR signaling pathway of A. flavus, we conducted targeted gene deletion experiments on tapA. We failed to obtain a null mutant, we hypothesized that tapA knockout was lethal in A. flavus. To investigate the function of TapA, we employed homologous recombination, to replace the native promoter with the strong gpdA promoter, thereby constructing an overexpression strain (OE::tapA) with the enhanced promoter. Compared with WT strain, the OE::tapA strain were more sensitive to rapamycin and SDS stress, indicating that tapA gene plays an important role in regulating cell wall stress (Figure 5B and F). In S. pombe, the protein Tip41, which interacts with Tap42, plays a significant role in the cellular responses to nitrogen sources by regulating type 2A phosphatases (Fenyvuesvolgyi et al., 2005). In S. cerevisiae, Tap42-phosphatase complexes associate with TORC1, whereas Tip41 functions to attenuate TORC1 signaling by stimulating the PP2A phosphatase (Jacinto et al., 2001). Unlike its yeast counterpart Tip41, TIPRL, a mammalian Tip41-like protein, exerts a positive influence on mTORC1 signaling by interacting with PP2Ac (Nakashima et al., 2013). While some functions of Tap42 and its binding partners Tip41 may be conserved, the interaction between Tip41 and Tap42 was not observed in the filamentous fungi. The yeast two-hybrid assay showed that FgTip41 interacts with the phosphatase FgPpg1 rather than FgTap42, while the FgTap42 bind with phosphatase FgPp2A, implying that FgTap42, FgPpg1, and FgTip41 form a heterotrimer in F. graminearum (Yu et al., 2014). In the rice blast fungus M. oryzae, MoTip41 does not interact with MoTap42. However, MoTip41 can bind with the phosphatase MoPpe1, thereby facilitating crosstalk between the TOR pathway and the CWI pathway (Qian et al., 2021). In A. flavus, the OE::tapA and ∆tipA strains showed increased sensitivity to cell wall stress compared with the WT strain (Figures 5B and 6H). Based on the conserved roles of Tap42 and Tip41 in yeast and other filamentous fungi, we hypothesized that TapA and TipA in A. flavus also participate in CWI pathway. In summary, our study provided important insights into the role of TapA and TipA in the TOR signaling pathway of A. flavus, particularly in relation to cell wall stress. However, their interaction and implications for TOR and CWI signaling in A. flavus remain to be elucidated.
In S. cerevisiae, Tap42 can bind and regulate various PP2A phosphatases, including Pph3, Pph21, Pph22, Sit4, and Ppg1. This interaction plays a crucial role in the TOR signaling pathway, particularly in response to rapamycin treatment or nutrient deprivation (Jacinto et al., 2001). These phosphatases play important roles in other fungi and are involved in a variety of cellular processes, including growth, morphogenesis, and virulence. For example, the phosphatase Sit4 plays a crucial role in regulating cell growth, morphogenesis, and virulence in C. albicans (Han et al., 2019). Similarly, FgSit4 and FgPpg1 are associated with various cellular processes, including mycelial growth, conidiation, DON biosynthesis, and virulence in F. graminearum (Yu et al., 2014). These findings indicated the conserved role of PP2A phosphatases in fungal biology. In addition to their roles in growth and development, FgSit4 and FgPpg1 are involved in the regulation of various signaling pathways, such as the CWI pathway and LDs biogenesis regulation pathway (Yu et al., 2014). In this study, we found that the phosphatases SitA and Ppg1 play critical role in vegetative growth, conidiation, sclerotia formation, and aflatoxin biosynthesis in A. flavus (Figure 7A–C). Additionally, the phosphatases SitA and Ppg1 also affect pathogenicity and LD biogenesis (Figures 8A, 10C and D). This highlights the multifaceted influence of PP2A phosphatases on cellular signaling.
After the deletion of the sitA and ppg1 genes, the expression level of genes related to cell wall synthesis has decreased significantly, resulting in damage to the cell wall and showing a higher sensitivity to CWI stress (Figure 9C). Western blotting results showed that the phosphorylation level of MAPK kinase Slt2 has increased significantly in the ∆sitA and ∆ppg1 strains under cell wall stress, indicating that SitA and Ppg1 exert a negative regulatory effect on the CWI pathway (Figure 9D). In M. oryzae, MoPpe1 and MoSap1 serve as an adaptor complex that connects the CWI and TOR signaling pathways. The activation of the TOR pathway leads to the inhibition of the CWI pathway, highlighting a critical regulatory interplay between these two pathways in fungal biology (Qian et al., 2018). We speculated that SitA and Ppg1 might exhibit similar regulation within the CWI and TOR pathways across different fungal species. In addition, the phosphatase MoPpe1 interacts with the phosphatase-associated protein MoTip41, facilitating the exchange of information between the TOR and CWI signaling pathways (Qian et al., 2018). These results suggested that the phosphatase Ppe1 engages in interactions with various proteins to effectively coordinate the TOR and CWI signaling pathways, thereby regulating the growth and pathogenicity of the rice blast fungus M. oryzae (Qian et al., 2018). Given the conserved functions of PP2A phosphatases and the observed roles of SitA, Ppg1, TapA, and TipA in cell wall stress response, we hypothesized that TapA-phosphatase complexes may collaborate to regulate CWI processes in A. flavus. This hypothesis is supported by the similar functions exhibited by these proteins.
In F. graminearum, the phosphatase complex FgNem1/Spo7-FgPah1 cascade plays significant roles in fungal development, DON production, and LD biogenesis (Liu et al., 2019). Our study explored the conservation and functional significance of Nem1 and Spo7 homologs in A. flavus, revealing their pivotal roles in vegetative growth and secondary metabolism (Figure 11A and B). Our results indicated that LD biogenesis is facilitated by the Nem1/Spo7 and SitA/Ppg1 pathways in A. flavus (Figures 10C, D, 11E). These observations suggested a conserved and central role for PP2A phosphatases as core regulators within the LD biogenesis signaling network, highlighting their importance in cellular lipid metabolism and energy storage. While our findings underscored the significance of phosphatases in LD biogenesis, it is also plausible that other kinases contribute to the regulation of this process. To gain a comprehensive understanding of the precise molecular mechanism, it is essential to investigate the interplay between phosphatases and other kinases.
In conclusion, we identified the complex regulatory network that provides important insight into the interplay between the TOR, HOG, and CWI signaling pathways, and how they collectively regulate the development, aflatoxin biosynthesis, and pathogenicity of A. flavus. By elucidating the functions of genes associated with the TOR signaling pathway, our research could establish a solid foundation for comprehending the molecular mechanism underlying the aflatoxin biosynthesis of A. flavus. Our results could also contribute to the development of targeted strategies to manage aflatoxin contamination and mitigate the impact of A. flavus infections.
Materials and methods
Strains and cultural conditions
Request a detailed protocolThe A. flavus strains in this study were shown in Supplementary file 1a. All strains were cultivated on four different types of agar media: PDA, YES, YGT, and YXT. The growth and conidiation assays were conducted at 37°C. Additionally, aflatoxin production was assessed using YES and YXT liquid medium at 29°C.
Constructions of gene deletion and complemented mutants
Request a detailed protocolThe construction of gene deletion and complemented strains was carried out using the SOE-PCR strategy, following previously established protocols (Tan et al., 2013). To generate the xylPtorA mutant of A. flavus, an torA xylose promoter mutant cassette was synthesized via overlapping extension PCR. This cassette was designed to incorporate an upstream fragment of the torA gene, a selectable marker gene (pyrG from A. fumigatus), the xylP xylose-inducible promoter (Zadra et al., 2000), and a fragment of the torA coding sequence. Subsequently, this composite cassette was integrated into the A. flavus genome via homologous recombination, specifically replacing the native torA promoter with the inducible xylose promoter. To construct overexpression cassettes for the tapA gene, we utilized a molecular cloning strategy that entailed the fusion of PCR-amplified fragments from the pyrG-selectable marker and the gpdA promoter. Subsequently, employing a homologous recombination approach, we inserted this composite fragment upstream of the tapA gene, effectively replacing its native promoter with the stronger gpdA promoter. The primers utilized for the amplification of the flanking sequences of each gene can be found in Supplementary file 1b. The putative gene deletion mutants were validated through PCR assays using the appropriate primers (Figure 2—figure supplement 1), and their confirmation was subsequently obtained through sequencing analysis.
Growth, conidiation, and sclerotia analysis
Request a detailed protocolTo evaluate mycelial growth and conidia formation, 1 μL of 106 spores/mL conidia of all strains was point inoculated onto the center of the PDA, YGT, or YXT medium, and then cultured at 37°C for 5 days in the dark. The conidia were washed with 0.07% Tween-20 and counted using a hemocytometer and microscope (Ren et al., 2018). For sclerotia formation analysis, each strain was inoculated and cultivated on CM and YXT agar medium at 37°C in the dark for 7 days, and then 75% ethanol was used to wash away mycelia and conidia on the surface of the medium (Nie et al., 2016). These assays were repeated at least three times.
Extraction and quantitative analysis of aflatoxin
Request a detailed protocolTo extract and quantify aflatoxins production, 1 μL of 106 spores/mL conidia of all strains was point inoculated onto liquid YES medium, and cultures were then incubated at 29°C in the dark for 6 days. Aflatoxins were extracted from the media and detected using TLC (Nie et al., 2016). In addition, the standard AFB1 was employed for visual comparison, and aflatoxins were quantitatively analyzed using the ImageJ tool. The assays were repeated at least three times.
Stress assay
Request a detailed protocolTo determine the roles of the gene in A. flavus responding to various environmental stresses, 1 μL of 106 spores/mL conidia of all strains were point inoculated onto these PDA, YGT, YXT, or YES solid plates supplemented with various agents. These agents included 1 M NaCl, 1 M KCl, and 1 M CaCl2 for inducing osmotic stress, as well as 100 µg/mL or 300 µg/mL SDS, 200 µg/mL CFW, and 200 µg/mL CR for inducing cell wall stress. After 5 days of incubation at 37°C in the dark, the relative rates of inhibition were determined (Tan et al., 2021). The assays were repeated at least three times.
Seeds infection assays
Request a detailed protocolTo measure the pathogenicity of A. flavus, all peanut and maize seeds were washed in 0.05% sodium hypochlorite, 75% ethanol, and sterile water, respectively. Subsequently, all peanut and maize seeds were immersed in sterile water containing 106 spores/mL of all strains at 29°C in the dark for 60 min. After the conidia adhesion, the seeds were cultured at 29°C for 6 days in the dark (Tan et al., 2021).
Quantitative RT-PCR
Request a detailed protocolFor the real-time quantitative PCR (qPCR) assays, the A. flavus strains were cultivated in the PDA medium at 29°C for 3 days in the dark. Total RNA was extracted from the mycelia of all strains, and cDNA was synthesized with the First Strand cDNA Synthesis Kit, and then the cDNA was used as the template for qRT-PCR analysis with SYBR Green qPCR mix. The relative transcript level of each gene was calculated by the 2-ΔΔCT method (Livak and Schmittgen, 2001).
Western blot analysis
Request a detailed protocolTo extract the total protein, the mycelia of each strain were cultivated in YES liquid medium for 48 hr, then the sample was grinded under liquid nitrogen to a fine powder and transferred to a 1.5 mL microcentrifuge tube added with RIPA lysis buffer and 1 mM PMSF protease inhibitor. Proteins were separated by 12% SDS-PAGE and transferred onto a polyvinylidene fluoride membrane. Phosphorylation levels of Hog1 and Slt2 were detected by Phospho-p38 MAPK antibody (Cell Signaling Technology, Boston, MA, USA) and Phospho-p44/42 MAPK (Erk 1/2) antibody (Cell Signaling Technology, MA, USA), respectively. Monoclonal antibody Hog1 (Santa Cruz Biotechnology, Dallas, TX, USA) and Mpk1 antibody (Santa Cruz Biotechnology, Dallas, TX, USA) were used as loading controls. Total Slt2 levels were quantified with an anti-Mpk1 antibody. The experiment was conducted three times independently.
Statistical analysis
Request a detailed protocolAll data were measured with means ± standard deviation (SD) of three biological replicates. GraphPad Prism 7 software was used for conducting statistical and significance analysis, and p-values<0.05 represented statistical significance. Student’s t-test was used to compare the two means for differences, whereas Tukey’s multiple comparisons were used for testing multiple comparisons.
Data availability
The authors confirm that the data supporting the findings of this study are available within the article and supplementary file. Source data files have been provided for Figures 4 and 9 and Figure 2—figure supplement 1.
References
-
Structure of TOR and its complex with KOG1Molecular Cell 27:509–516.https://doi.org/10.1016/j.molcel.2007.05.040
-
Elucidating TOR signaling and rapamycin action: lessons from Saccharomyces cerevisiaeMicrobiology and Molecular Biology Reviews 66:579–591.https://doi.org/10.1128/MMBR.66.4.579-591.2002
-
Rapamycin and less immunosuppressive analogs are toxic to Candida albicans and Cryptococcus neoformans via FKBP12-dependent inhibition of TORAntimicrobial Agents and Chemotherapy 45:3162–3170.https://doi.org/10.1128/AAC.45.11.3162-3170.2001
-
The TOR signalling network from yeast to manThe International Journal of Biochemistry & Cell Biology 38:1476–1481.https://doi.org/10.1016/j.biocel.2006.02.013
-
The role of phosphatases in TOR signaling in yeastCurrent Topics in Microbiology and Immunology 279:19–38.https://doi.org/10.1007/978-3-642-18930-2_2
-
Fission yeast homologue of Tip41-like proteins regulates type 2A phosphatases and responses to nitrogen sourcesBiochimica et Biophysica Acta 1746:155–162.https://doi.org/10.1016/j.bbamcr.2005.09.006
-
Genetic analysis of the TOR pathway in Aspergillus nidulansEukaryotic Cell 4:1595–1598.https://doi.org/10.1128/EC.4.9.1595-1598.2005
-
Functional diversity and pharmacological profiles of the FKBPs and their complexes with small natural ligandsCellular and Molecular Life Sciences 70:3243–3275.https://doi.org/10.1007/s00018-012-1206-z
-
Aflatoxin B1 (AFB1) production by Aspergillus flavus and Aspergillus parasiticus on ground Nyjer seeds: The effect of water activity and temperatureInternational Journal of Food Microbiology 296:8–13.https://doi.org/10.1016/j.ijfoodmicro.2019.02.017
-
Ubiquitin regulates TORC1 in yeast Saccharomyces cerevisiaeMolecular Microbiology 100:303–314.https://doi.org/10.1111/mmi.13319
-
Aspergillus flavus: the major producer of aflatoxinMolecular Plant Pathology 8:713–722.https://doi.org/10.1111/j.1364-3703.2007.00436.x
-
Characterization of three FK506-binding proteins in the entomopathogenic fungus Beauveria bassianaJournal of Invertebrate Pathology 171:107334.https://doi.org/10.1016/j.jip.2020.107334
-
Role of the Botrytis cinerea FKBP12 ortholog in pathogenic development and in sulfur regulationFungal Genetics and Biology 46:308–320.https://doi.org/10.1016/j.fgb.2008.11.011
-
Aspergillus flavus SUMO contributes to fungal virulence and toxin attributesJournal of Agricultural and Food Chemistry 64:6772–6782.https://doi.org/10.1021/acs.jafc.6b02199
-
TOR signaling in fission yeastCritical Reviews in Biochemistry and Molecular Biology 43:277–283.https://doi.org/10.1080/10409230802254911
-
TOR signalling regulates mitotic commitment through stress-activated MAPK and polo kinase in response to nutrient stressBiochemical Society Transactions 37:273–277.https://doi.org/10.1042/BST0370273
-
Lysine succinylation contributes to aflatoxin production and pathogenicity in Aspergillus flavusMolecular & Cellular Proteomics 17:457–471.https://doi.org/10.1074/mcp.RA117.000393
-
Nutritional control via Tor signaling in Saccharomyces cerevisiaeCurrent Opinion in Microbiology 11:153–160.https://doi.org/10.1016/j.mib.2008.02.013
-
Insights into the biogenesis and emerging functions of lipid droplets from unbiased molecular profiling approachesFrontiers in Cell and Developmental Biology 10:901321.https://doi.org/10.3389/fcell.2022.901321
-
A SOE-PCR method of introducing multiple mutations into Mycoplasma gallisepticum neuraminidaseJournal of Microbiological Methods 94:117–120.https://doi.org/10.1016/j.mimet.2013.05.013
-
FK506-binding proteins and their diverse functionsCurrent Molecular Pharmacology 9:48–65.https://doi.org/10.2174/1874467208666150519113541
-
Regulation of the cell integrity pathway by rapamycin-sensitive TOR function in budding yeastThe Journal of Biological Chemistry 277:43495–43504.https://doi.org/10.1074/jbc.M205408200
-
Interaction with Tap42 is required for the essential function of Sit4 and type 2A phosphatasesMolecular Biology of the Cell 14:4342–4351.https://doi.org/10.1091/mbc.e03-02-0072
-
Enzyme reactions and genes in aflatoxin biosynthesisApplied Microbiology and Biotechnology 64:745–755.https://doi.org/10.1007/s00253-004-1566-x
-
xylP promoter-based expression system and its use for antisense downregulation of the Penicillium chrysogenum nitrogen regulator NREApplied and Environmental Microbiology 66:4810–4816.https://doi.org/10.1128/AEM.66.11.4810-4816.2000
Article and author information
Author details
Funding
National Natural Science Foundation of China (Grant No. 31972214)
- Shihua Wang
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
The authors also thank Zhenhong Zhuang, Jun Yuan, Yu Wang, Kunzhi Jia, Xiuna Wang, and Xinyi Nie for their help in the experiments.
Version history
Cite all versions
You can cite all versions using the DOI https://doi.org/10.7554/eLife.89478. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2023, Li et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 2,277
- views
-
- 213
- downloads
-
- 8
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 5
- citations for umbrella DOI https://doi.org/10.7554/eLife.89478
-
- 3
- citations for Version of Record https://doi.org/10.7554/eLife.89478.5
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The target of rapamycin signaling pathway regulates vegetative development, aflatoxin biosynthesis, and pathogenicity in Aspergillus flavus
eLife 12:RP89478.
https://doi.org/10.7554/eLife.89478.5




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}