LRRC23 truncation impairs radial spoke 3 head assembly and sperm motility underlying male infertility
- Department of Cellular and Molecular Physiology, Yale School of Medicine, Yale University, United States
- Department of Molecular Biology, Pusan National University, Republic of Korea
- Department of Molecular Biophysics and Biochemistry, Yale School of Medicine, Yale University, United States
- Department of Biotechnology, Faculty of BiologicalSciences, Quaid-i-Azam University, Pakistan
- Department of Genetics, YaleSchool of Medicine, Yale University, United States
- Department of Biomedical Sciences, Korea University College of Medicine, Republic of Korea
- Yale Center forGenome Analysis, Yale University, United States
- Department of Biochemistry, Faculty of Biological Sciences, Quaid-i-Azam University, Pakistan
- Laboratory of Human Genetics and Genomics, The Rockefeller University, United States
- Department of Obstetrics, Gynecology and Reproductive Sciences, Yale School of Medicine, Yale University, United States
Figures
Figure 1 with 1 supplement
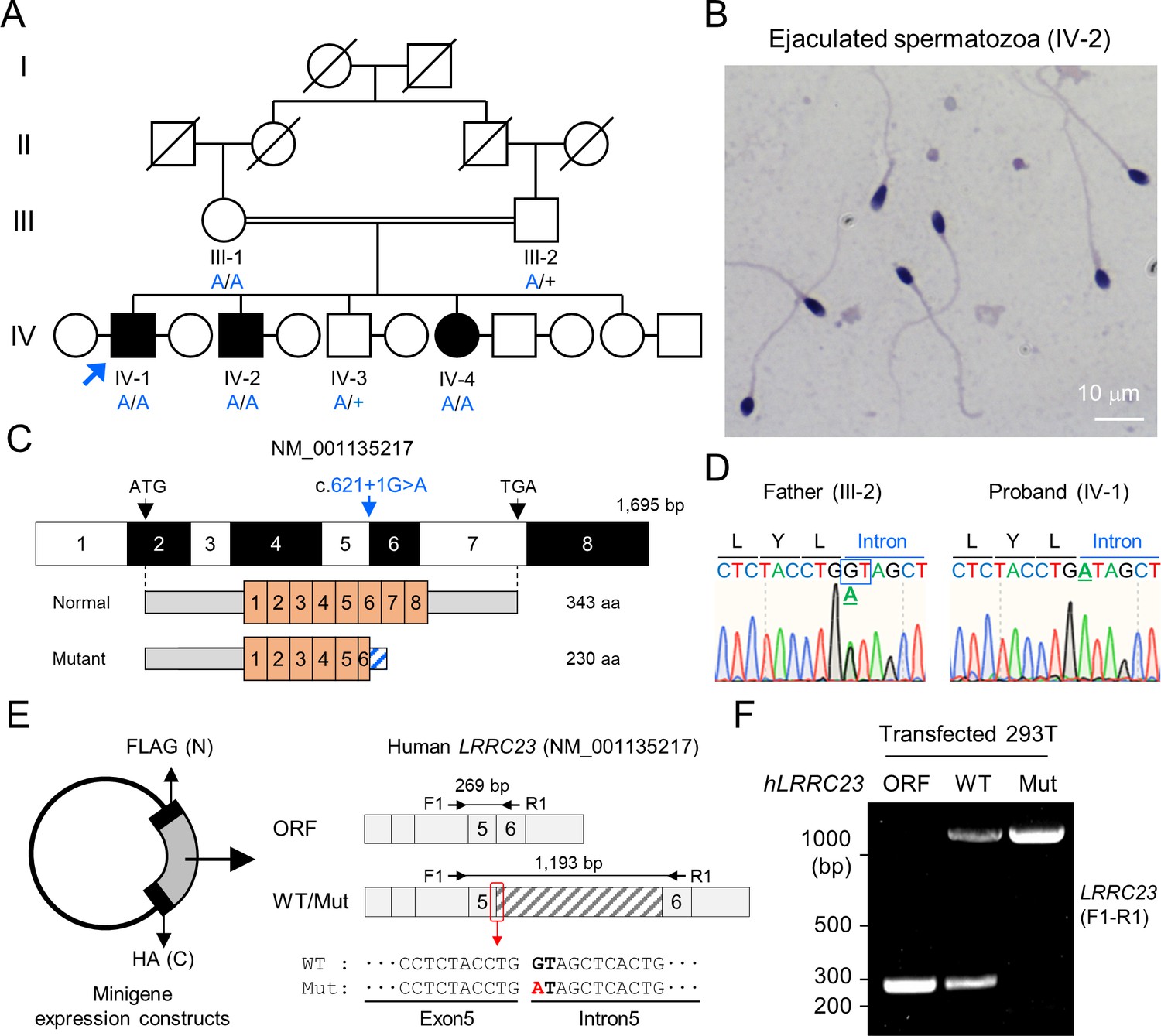
A bi-allelic splicing donor site variant in LRRC23 was identified from asthenozoospermia patients.
(A) A consanguineous pedigree with two infertile males (IV-1 and IV-2). IV-1 was subjected for WES (arrow). Genotypes of the variant (blue) in all family members included in this study (III-1, III-2, IV-1, IV-2, IV-3, and IV-4) are confirmed by Sanger sequencing. +, wild-type allele. An infertile female sibling (IV-4) is marked in black circle. (B) Papanicolaou-stained sperm from the infertile male (IV-2). (C) Mapping of the LRRC23 variant. Mutation of G to A at the splicing donor site in the 5th intron is predicted to prevent LRRC23 mRNA from splicing. (D) Sequencing chromatograms presenting the LRRC23 variant in the infertile male (IV-1) and his father (III-2). The variant is underlined and normal splicing donor site (GT) is boxed. (E, F) Minigene assay for testing altered splicing of LRRC23 by the variant. (E) Minigene constructs expressing LRRC23 ORF containing the 5th intron (sashed) with wild-type (WT) or mutant (Mut, red) splicing donor site were generated. The constructs are tagged with FLAG and HA at N- and C-termini, respectively. (F) RT-PCR of the 293T cells transfected with the minigene constructs reveals the 5th intron is not spliced out and retained by the variant. Intron-spanning primers, F1 and R1, are used. Repeated three times with biological replications.
Figure 1—figure supplement 1
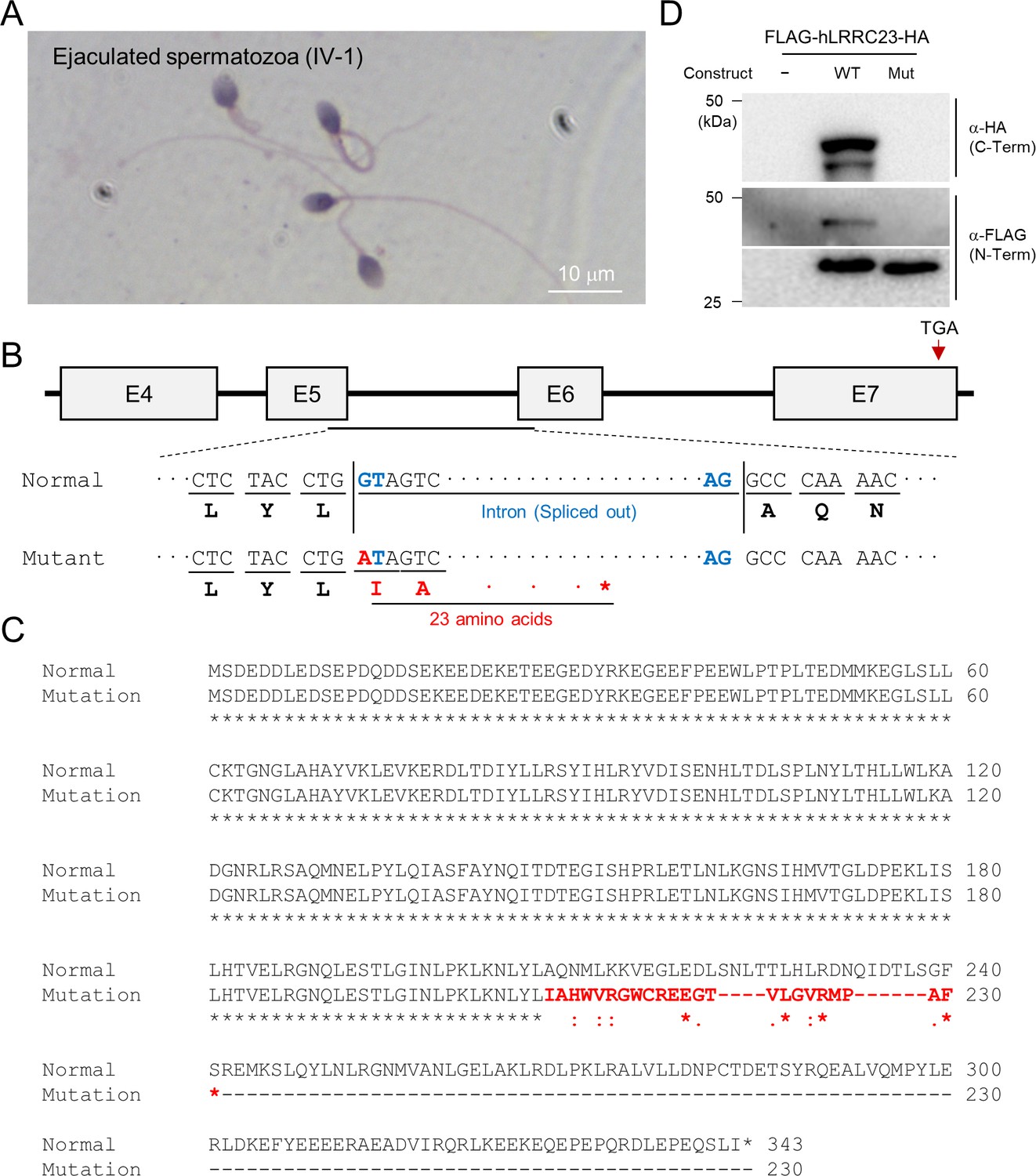
A LRRC23 splicing site variant identified from infertile male patients.
(A) Papanicolaou-stained sperm from the infertile male member, IV-2. (B) The splicing site variant in the 5th intron of LRRC23 gene. The variant is predicted to cause early termination of the protein translation. (C) Sequence alignment of the normal and predicted mutant LRRC23 by the splicing site mutation. Non-native amino acid sequences encoded by the mutant allele are colored in red. (D) Immunoblot analysis of 293T cells transfected with the human LRRC23 minigene constructs containing the 5th intron. The construct carrying the intron with the identified variant generate only a truncated LRRC23. Non-transfected cells (NTC) are used for a negative control. Three biological replications.
-
Figure 1—figure supplement 1—source data 1
Uncropped blot images for Figure 1—figure supplement 1.
- https://cdn.elifesciences.org/articles/90095/elife-90095-fig1-figsupp1-data1-v1.zip
Figure 2 with 2 supplements
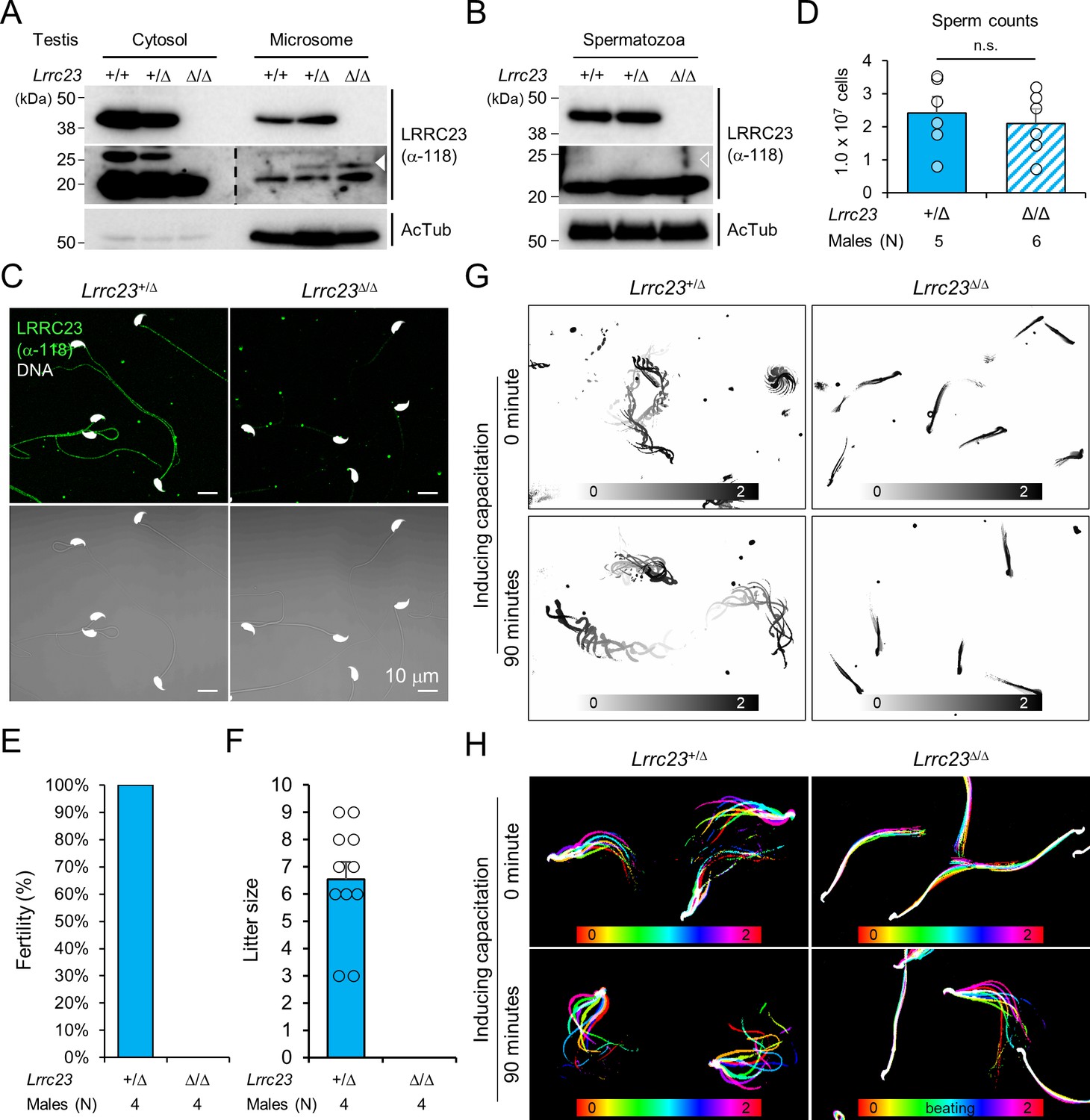
Lrrc23 mutant mice mimicking human splice variant phenocopy male infertility and reduced sperm motility.
(A, B) Immunoblotting of LRRC23 in testis (A) and epididymal sperm (B) from mutant male mice. Truncated LRRC23 (arrowheads) is detected from testis microsome fraction (filled), but not in mature sperm (empty), of heterozygous (+/Δ) and homozygous (Δ/Δ) males. Acetylated tubulin (AcTub) is a loading control. Experiments were performed with three biological replications. (C) Confocal images of immunostained LRRC23 in Lrrc23+/Δ and Lrrc23Δ/Δ epididymal sperm Experiments were repeated with three biological replications. (D) Epididymal sperm counts. n.s., not significant. (E) Pregnancy rate of Lrrc23+/Δ and Lrrc23Δ/Δ males. (F) Number of litters from fertile females mated with Lrrc23+/Δ and Lrrc23Δ/Δ males. (G) Swimming trajectory of Lrrc23+/Δ and Lrrc23Δ/Δ sperm in viscous media (0.3% methylcellulose). Swimming trajectory for 2 s is overlaid. Experiments were performed with three biological replications. See Video 1. (H) Flagellar waveforms of Lrrc23+/Δ and Lrrc23Δ/Δ sperm before (0 min) and after (90 min) inducing capacitation. Flagellar movements for two beat cycles are overlaid and color coded in time. Experiments were performed with three biological replications. See Video 2. In (A–C), samples from WT were used for positive or negative control of normal or truncated LRRC23. In (D, F), circles indicate sperm counts from individual males (D) and pup numbers from each litter (F), and data represented as mean ± SEM (D, Mann-whiteny U test; F, Student’s t-test). n.s., non-significant.
-
Figure 2—source data 1
Uncropped blot images for Figure 2.
- https://cdn.elifesciences.org/articles/90095/elife-90095-fig2-data1-v1.zip
Figure 2—figure supplement 1
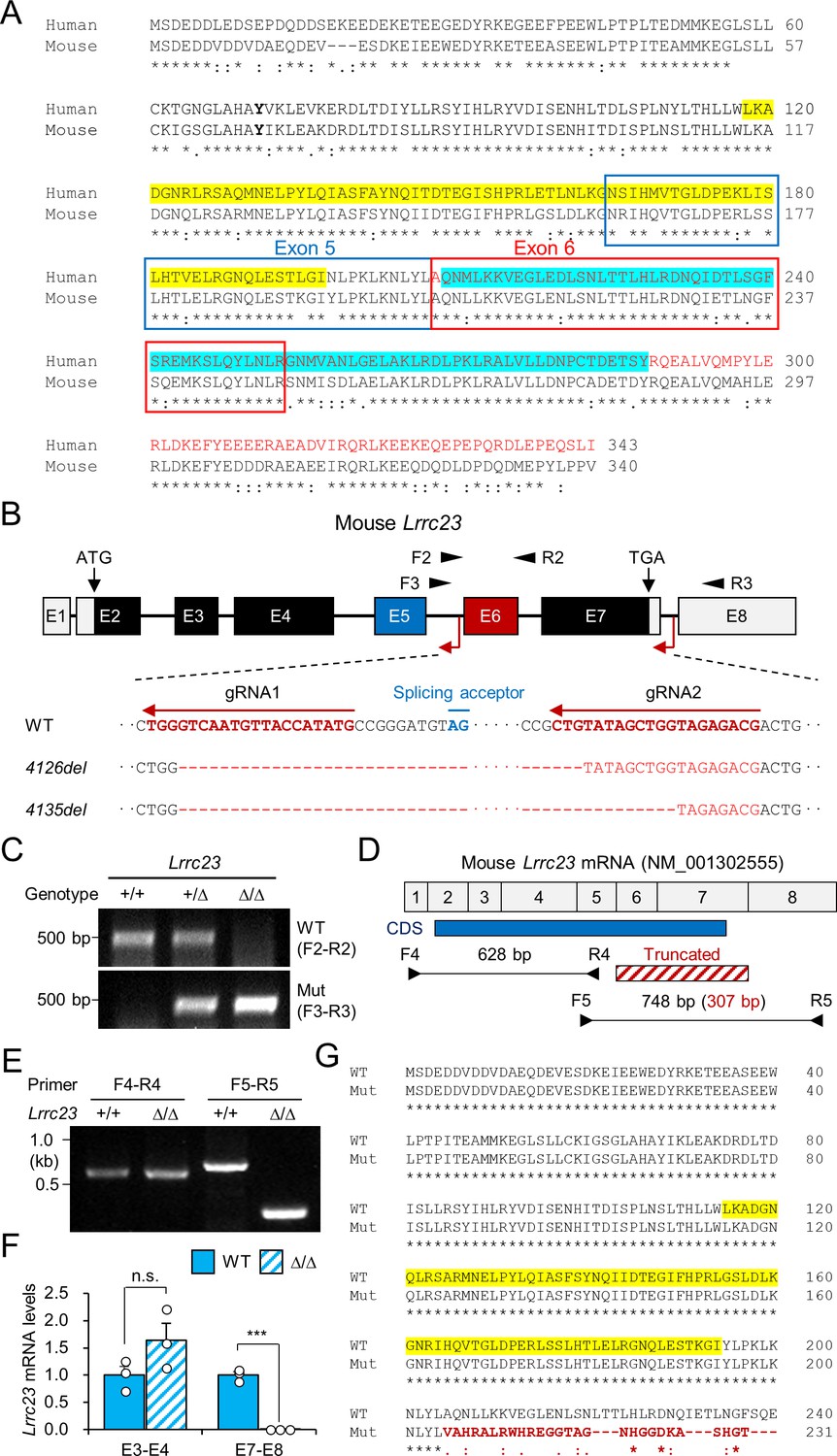
Generation of LRRC23 mutant mouse models.
(A) A pair-wise alignment of human and mouse LRRC23 protein sequences. Deleted amino acids by the splice site mutation (encoded by exon 6 and 7) are written in red. Epitopes for the LRRC23 antibodies against N- (118–197, yellow) and C-terminus (209–288, cyan) are highlighted. (B) Generation of LRRC23 mutant mouse models by CRISPR/Cas9 genome editing. Two guide RNAs targeting 5th and 7th intron were used to generate mouse models expressing LRRC23 lacking C-terminus like the predicted human mutant LRRC23. Two mutant alleles with 4126 (Lrrc23-4126del) or 4135 bp (Lrrc23-4135del) deletion were established. (C) Genotyping of the generated mouse models. Primer locations are marked in panel (B). Lrrc23-4126delΔ/Δ and Lrrc23-4135del Δ/Δ mice show the identical phenotypes. Lrrc23-4126Δ/Δ mice were used as Lrrc23Δ/Δ mice in this study unless indicated. (D–F) Lrrc23 mRNA expression in testis from Lrrc23 mutant males. (D) Shown is a diagram of exon composition of mouse Lrrc23 mRNA (NM_001302555). Protein coding region (CDS), truncated exons in the mutant allele, and primers for RT-PCR are marked. A primer pair spanning truncated region (F5–R5) amplifies 748 bp and 307 bp PCR product from WT and the mutant alleles, respectively. (E) Endpoint and (F) real-time RT-PCR analyses of Lrrc23 mRNA expression in Lrrc23Δ/Δ testis. Primers amplifying exon 3 and 4 (E3–E4) and exon 7 and 8 (E7–E8) were used for real-time RT-PCR. Circles indicate relative levels from individual animals. N=3 for real-time PCR. Statistical analysis was performed by Student’s t-test. n.s., not significant; p***<0.001. (G) A pair-wise sequence alignment of WT and the predicted mutant LRRC23 proteins generated from the truncated allele. Non-native sequences from the truncated Lrrc23 mRNA are colored in red.
-
Figure 2—figure supplement 1—source data 1
- https://cdn.elifesciences.org/articles/90095/elife-90095-fig2-figsupp1-data1-v1.zip
Figure 2—figure supplement 2
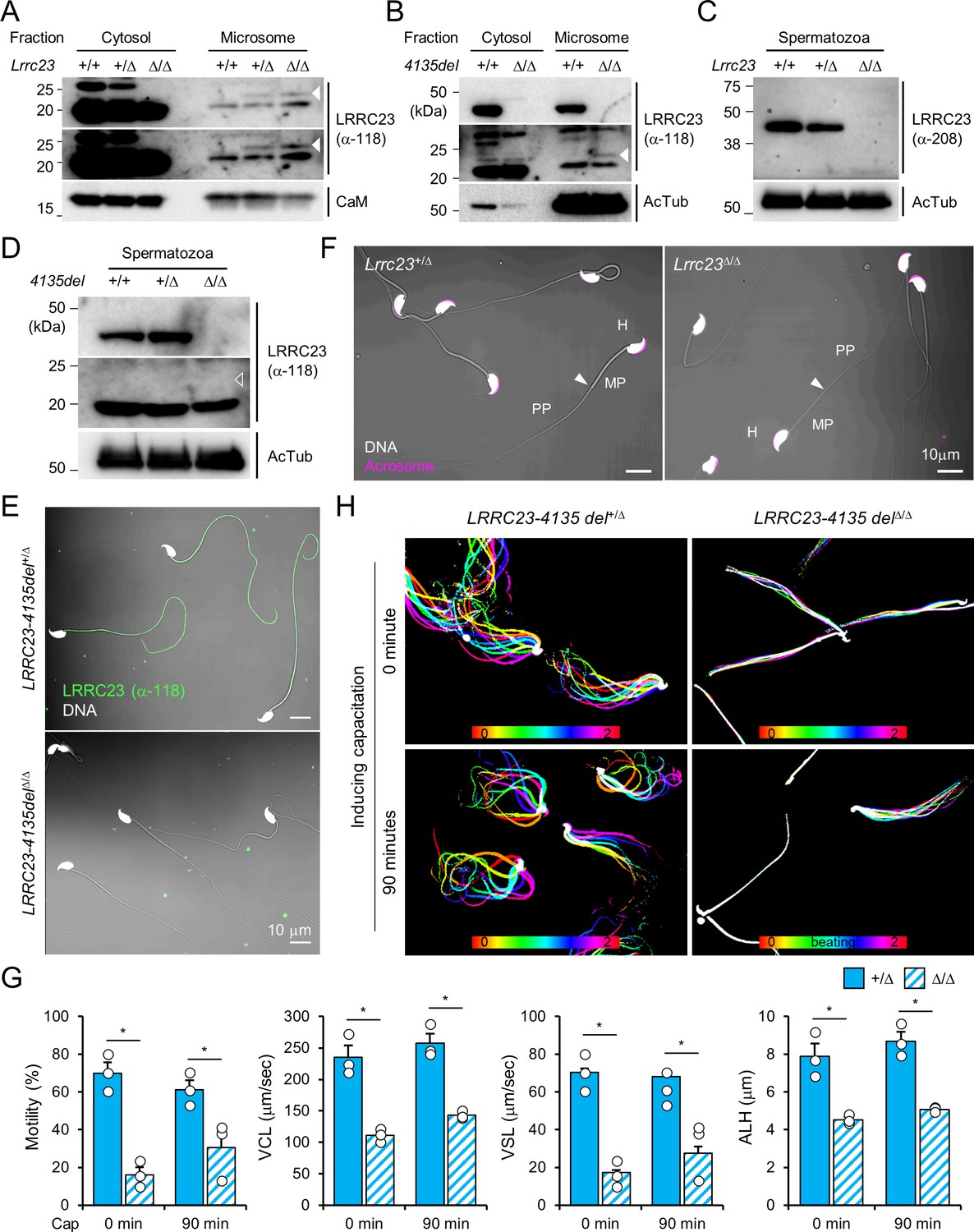
Characterization of the Lrrc23 lossof-function male mice.
(A–D) Protein expression of LRRC23 in wild type (WT) and Lrrc23 mutant males. (A) Original images of truncated LRRC23 immunoblotting in testis shown in Figure 2A. Short (top) and long (middle) exposure images are shown. Calmodulin (CaM) is a loading control. (B) Immunobloting of LRRC23 in testis from Lrrc23-4135del mutant males. (C) LRRC23 immunoblotting in epididymal sperm from WT, Lrrc23+/Δ, and Lrrc23Δ/Δ males. (D) Immunoblotting of LRRC23 in WT and Lrrc23-4135delΔ/Δ sperm. Truncated LRRC23 (arrowheads) is detected from testis (filled, (A and B)), but not from epididymal sperm (empty, (D)) of Lrrc23 mutant males. Acetylated tubulin (AcTub) is a loading control. Samples from WT males were used for either positive or negative controls of normal and truncated LRRC23, respectively (A, B, C, and D). (E) Confocal images of immunostained LRRC23 in Lrrc23-4135del+/Δ and Lrrc23-4135delΔ/Δ sperm. (F) Confocal images of epididymal sperm from Lrrc23+/Δ and Lrrc23Δ/Δ males. Acrosome is stained with wheat-germ agglutinin. Arrowheads indicate the annulus. (G) Computer assisted sperm analysis (CASA) to measure motility parameters of Lrrc23+/Δ and Lrrc23Δ/Δ sperm. Sperm motility parameters were measured before (0 min) and after (90 min) inducing capacitation. VCL, curvilinear velocity; VSL, straight line velocity; ALH, amplitude of lateral head. Mann-Whitney U test was used for statistical comparison. *p≤0.05. N=3. Circles indicate values from individual animals. Data is represented as mean ± SEM. (H) Flagellar movement of the Lrrc23-4135del+/Δ (left) and Lrrc23-4135delΔ/Δ (right). Shown are overlays of sperm tail movement over two beat cycles before (0 min, top) and after (90 min, bottom) inducing capacitation. Used are LRRC23 antibodies recognizing N- (α–118) or C- (α–208) terminal region (A, B, C, D, and E). Hoechst is used for counterstaining DNA and fluorescence and DIC images are merged (E and F). All experiments were carried out with over three biological replications.
-
Figure 2—figure supplement 2—source data 1
Uncropped blot images for Figure 2—figure supplement 1.
- https://cdn.elifesciences.org/articles/90095/elife-90095-fig2-figsupp2-data1-v1.zip
Figure 3 with 1 supplement
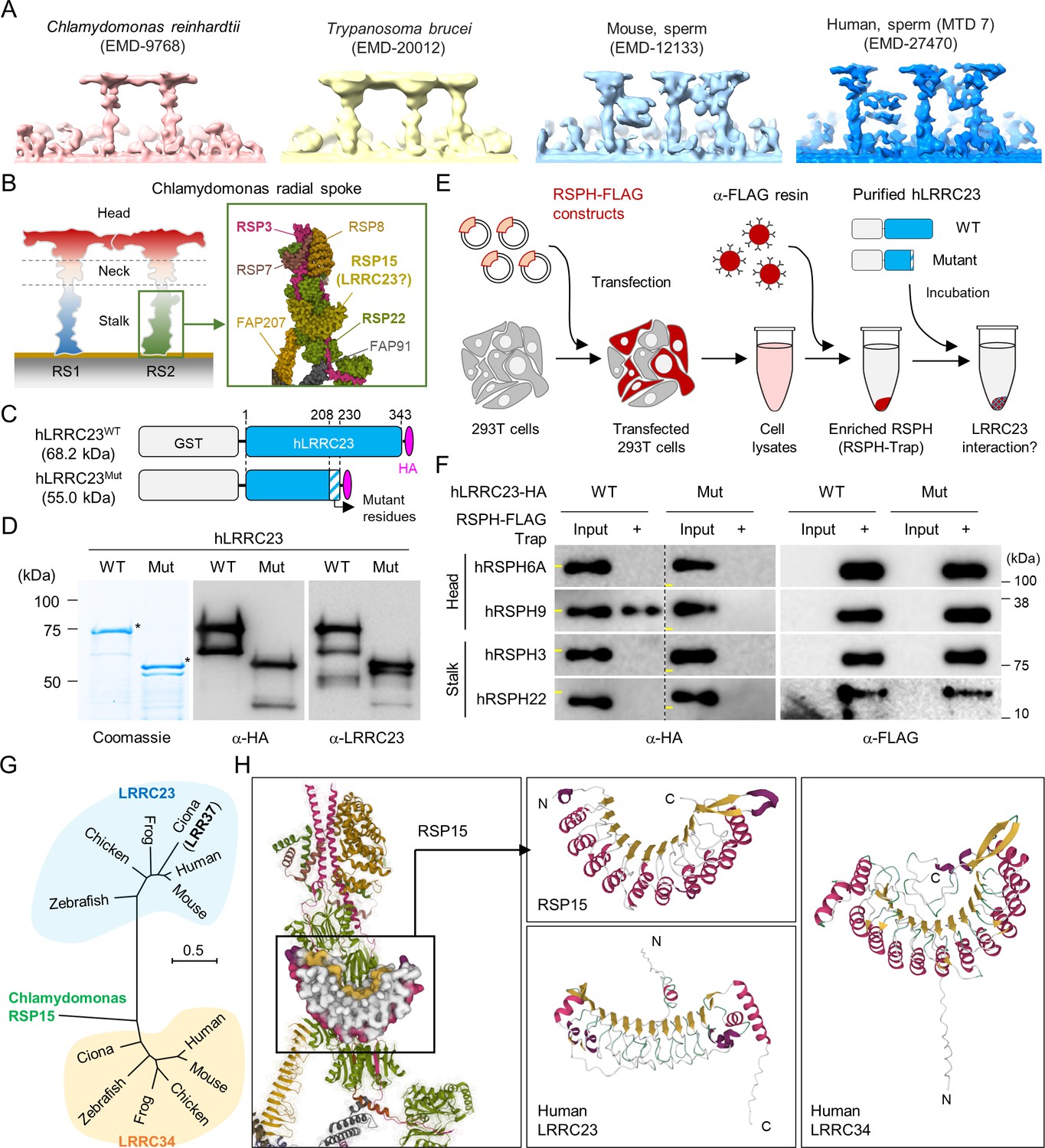
C-terminal truncation of human LRRC23 by the splicing site mutation prevents its interaction with radial spoke (RS) head.
(A) Sub-tomogram averaging images of RSs from Chlamydomonas reinhardtii (red), Trypanosoma brucei (yellow), mouse sperm (sky blue), and human sperm (blue). RSs at 7th microtubule doublet (MTD) are shown for human sperm. Original data from Electron Microscopy Data Bank was rendered. (B) Structure of RS in C. reinhardtii. A schematic cartoon shows the RS1 and 2. The structure of RS2 stalk is shown in inset (PDB Id: 7JRJ). (C, D) Purification of normal (hLRRC23WT) and the mutant human LRRC23 (hLRRC23Mut) by the splicing site mutation (c.621+1 G>A) in this study. (C) Diagrams for the purified recombinant normal and mutant proteins tagged with tagged with GST and HA at N- and C-termini, respectively. (D) Purified proteins by Coomassie blue staining (left) and immunoblotting with α-HA (middle) and a-LRRC23 (right). Proteins matched to the predicted size were marked with asterisks. (E) A cartoon of the RSPH-trap approach to test LRRC23 interaction with RS proteins. Individual human RS proteins tagged with FLAG (RSPH-FLAG) are expressed in 293T cells and enriched by α-FLAG resin from cell lysates. The recombinant RSPH proteins were incubated with the purified hLRRC23WT or hLRRC23Mut and subjected to immunoblotting. (F) Interaction of hLRRC23 to a RS head component, RSPH9. The purified hLRRC23 were incubated with the RSPH-Trap (RS head, RSPH6A and RSPH9; stalk, RSPH3 and RSPH22) and subjected to immunoblotting. 5% amount of the hLRRC23s used for the trap assay were loaded as inputs. Yellow lines in individual α-HA blot images indicate marker information (75 kDa, left; 50 kDa, right). Experiments were repeated four times. Purified GST was used for negative control (Figure 3—figure supplement 1B). Experiments were repeated three times with biological replications. (G) A phylogenetic tree constructed by Maximum-likelihood analysis of the protein sequences of the C. reinhardtii RSP15 and the orthologs of LRRC23 and LRRC34. LRR37, the first LRRC23 ortholog identified in Ciona intestinalis is marked in bold. (H) Comparison of the reported RSP15 from C. reinhardtii and the predicted structure of LRRC23 and LRRC34 from human. Atomic structure of the C. reinhardtii RS2 containing RSP15 are represented by ribbon (RS2) and surface (RSP15) diagram (left, PDB Id: 7JU4). Ribbon diagrams of C. reinhardtii RSP15 and AlphaFold-predicted human LRRC23 (middle) and LRRC34 (right) are shown for structural comparison. Secondary structures are color-coded. Different from C. reinhardtii RSP15 and LRRC34, LRRC23 does not display repeated α-helix (magenta) between β-sheets (gold).
-
Figure 3—source data 1
Uncropped gel and blot images for Figure 3.
- https://cdn.elifesciences.org/articles/90095/elife-90095-fig3-data1-v1.zip
Figure 3—figure supplement 1
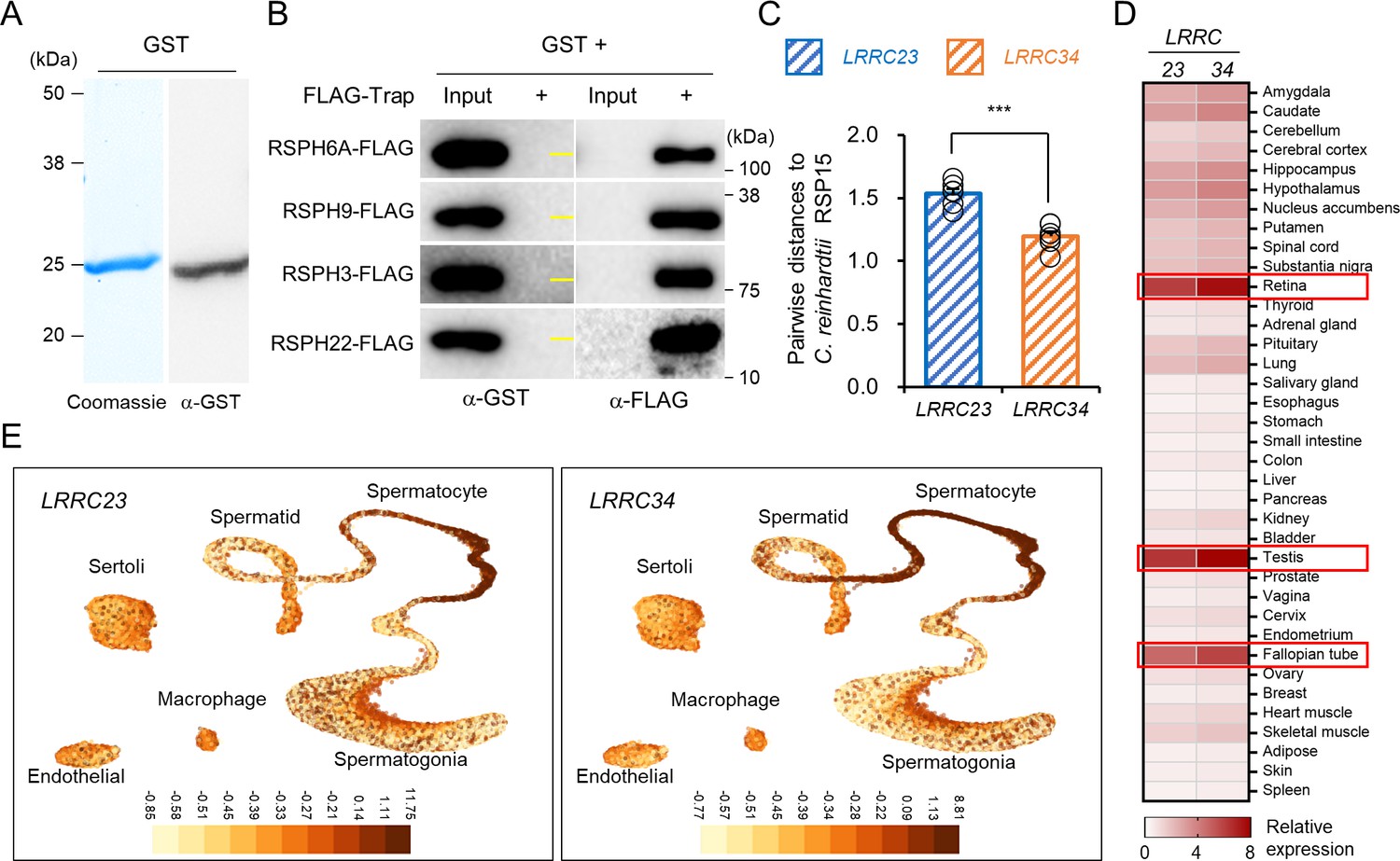
A predicted RSP15 ortholog, LRRC34, in metazoan species.
(A) Purified recombinant GST confirmed by Coomassie blue staining (left) and immunoblotting (right). (B) Interaction test of GST to human radial spoke proteins (RSPH) by trap-assay as a negative control. Yellow lines in α-GST blot images indicate molecular weight (25 kDa). (C) Comparison of the pairwise distances of Chlamydomonas reinhardtii RSP15 to the LRRC23 or LRRC34 orthologs. Protein sequences of C. reinhardtii RSP15 and the metazoan LRRC23 and LRRC34 orthologs were aligned and pairwise distances were calculated. Protein sequences of the C. reinhardtii RSP15 are closer to LRRC34 orthologs evolutionarily than LRRC23 orthologs in metazoan. Statistical analysis was performed by Student’s t-test. ***p<0.001, Data is represented by mean ± SEM. (D) A heatmap to represent human tissue mRNA expression of LRRC23 and LRRC34. Relative mRNA expression levels were calculated by normalizing tissue nTPM values with the median values. Three tissues with the highest LRRC23 and LRRC34 mRNA levels are highlighted. RNA GTEx tissue gene data from The Human Cell Atlas was used. (E) LRRC23 (left) and LRRC34 (right) mRNA expression in human testicular cells. mRNA levels in individual cells are represented by UMAP plots downloaded from UCSC cell browser.
-
Figure 3—figure supplement 1—source data 1
Uncropped blot images for Figure 3—figure supplement 1.
- https://cdn.elifesciences.org/articles/90095/elife-90095-fig3-figsupp1-data1-v1.zip
Figure 4 with 1 supplement
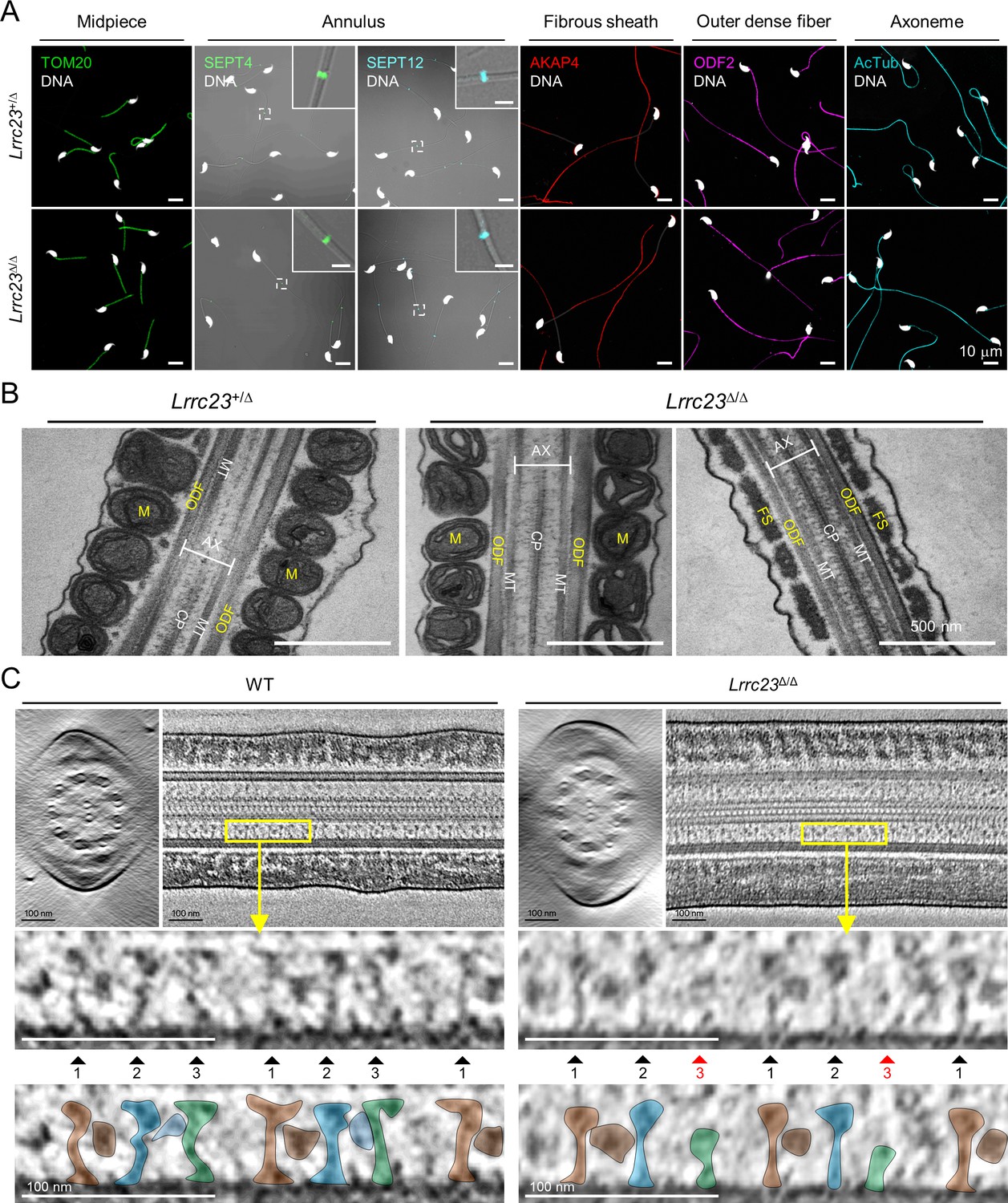
LRRC23 mutation disrupts the third radial spoke (RS) in sperm flagellum.
(A) Immunostaining of flagellar proteins in different compartments. Shown are midpiece (TOM20), annulus (SEPT4 and SEPT12), fibrous sheath (AKAP4), outer dense fiber (ODF2), and axoneme (acetylated tubulin, AcTub) in Lrrc23+/Δ (top) and Lrrc23Δ/Δ (bottom) sperm. Magnified insets are represented for annulus proteins (scale bars in insets = 2 μm). Fluorescence and corresponding DIC images are merged. Sperm heads were counter stained with Hoechst. Lrrc23+/Δ sperm were used for positive control. Experiments were performed with three biological replications. (B) Transmission electron microscopy images of Lrrc23+/Δ (left) and Lrrc23Δ/Δ (right) sperm. Shown are longitudinal section of sperm flagella. M, mitochondria; ODF, outer dense fiber; AX, axoneme; CP, central pair; MT, microtubule; FS, fibrous sheath. Lrrc23+/Δ sperm were used for positive control. (C) Cryo-electron tomography (cryo-ET) of WT and Lrrc23Δ/Δ sperm flagella. Shown are representative tomographic slices from WT (left) and Lrrc23Δ/Δ sperm (right). The 9+2 axonemal structure are shown in both WT and Lrrc23Δ/Δ in cross-sectional view (left). Axonemal structures are shown with proximal side of the flagellum on the left in longitudinal view (right; see Video 3). Magnified insets (bottom) reveal that RS1, 2, and 3 are shown in WT sperm (left, filled arrowheads) but RS3, especially head part, is not clearly visible (right, red arrowheads) in Lrrc23Δ/Δ sperm. RS1, 2, and 3 are distinguished by the interval between each set of RS1, 2, and 3, and the electron dense area corresponding to the barrel (RS1) and bridge (RS2-3) structures. WT sperm were used for positive control.
Figure 4—figure supplement 1
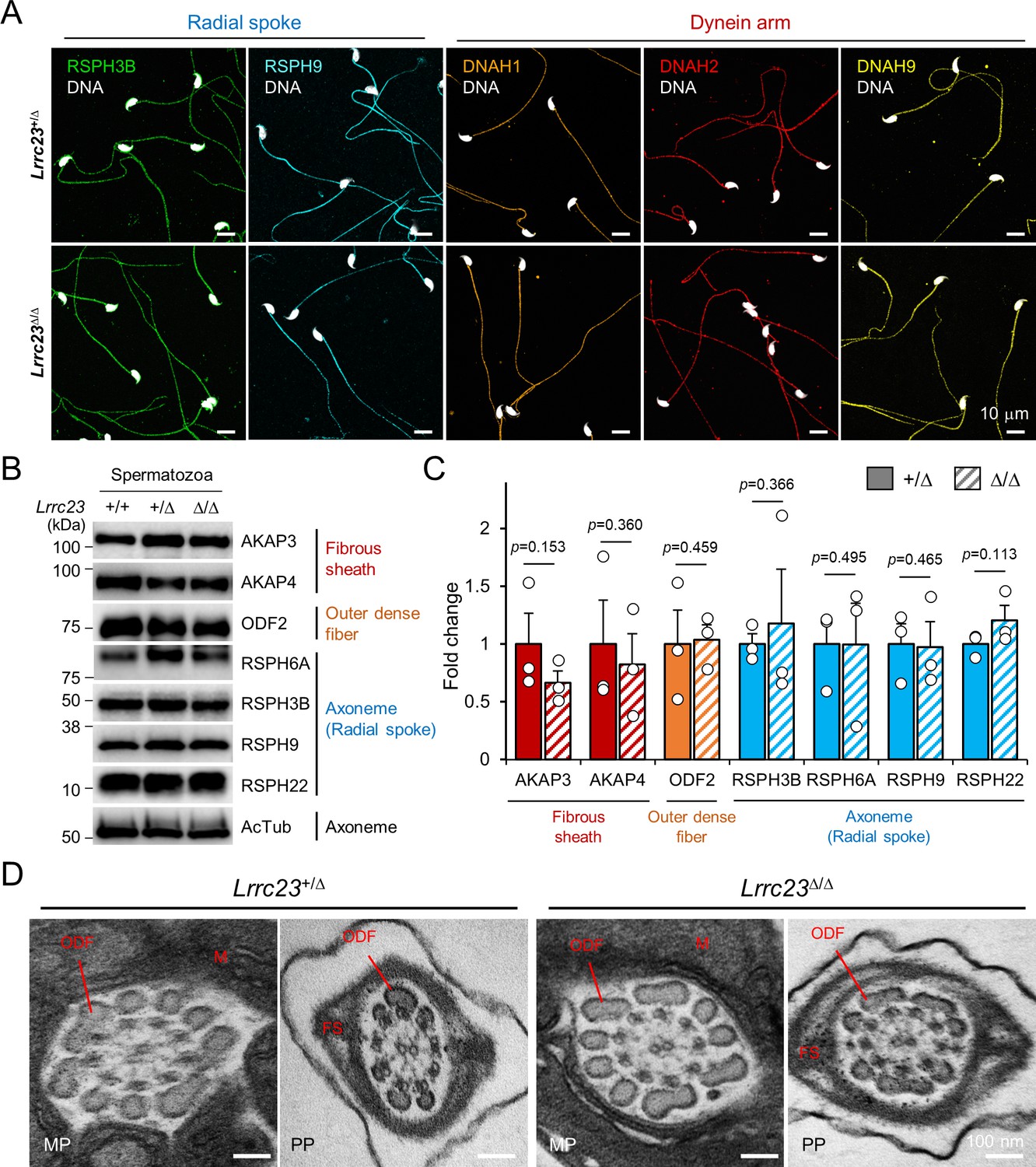
Flagellar compartmentalization in Lrrc23-mutant sperm.
(A) Distribution of the axonemal components in Lrrc23Δ/Δ sperm. Immunostained radial spoke (RSPH3B and RSPH9) and dynein arm (DNAH1, DNAH2, and DNAH9) in Lrrc23+/Δ and Lrrc23Δ/Δ sperm are shown by confocal images. Hoechst is used for counterstaining the sperm head. Experiment was performed with three biological replications. (B, C) Unaffected protein levels of the representative components of fibrous sheath, outer dense fiber, and axoneme in Lrrc23Δ/Δ sperm. (B) Immunoblotting of the flagellar components in cauda sperm. Acetylated tubulin (AcTub) is a loading control. (C) Fold-changes of the flagellar components levels in Lrrc23+/Δ (filled) and Lrrc23Δ/Δ (sash) sperm. Relative protein levels were quantified by measuring the band intensity and normalized by the AcTub intensity. The average level of each protein in Lrrc23+/Δ sperm is set to onefold. Circles represents fold changes of each sperm protein from individual males. Data represented as mean ± SEM. Statistical analysis was performed by Student’s t-test. N=3. (D) Transmission electron microscopy (TEM) images of epididymal sperm from Lrrc23+/Δ and Lrrc23Δ/Δ males. Shown are representative cross section TEM images of Lrrc23+/Δ (left) and Lrrc23Δ/Δ (right) sperm. No obvious structural defects at the midpiece (MP) and principal piece (PP) were observed from and Lrrc23Δ/Δ sperm. M, mitochondria; FS, fibrous sheath; ODF, outer dense fiber; AX, axoneme. WT and/or Lrrc23+/Δ sperm were used for positive control (A, B, C and D).
-
Figure 4—figure supplement 1—source data 1
Uncropped blot images for Figure 4—figure supplement 1.
- https://cdn.elifesciences.org/articles/90095/elife-90095-fig4-figsupp1-data1-v1.zip
Figure 5 with 1 supplement
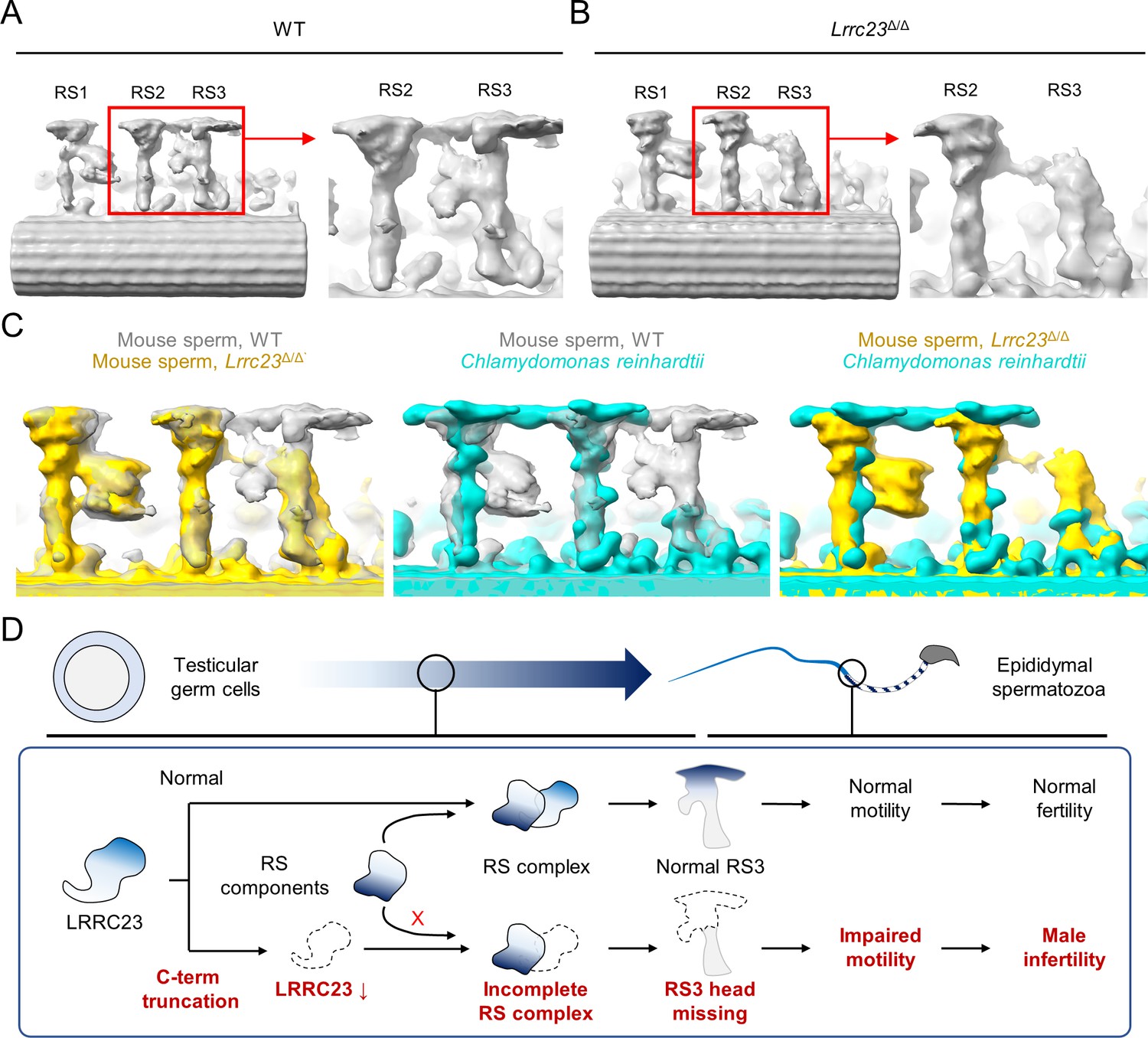
Head of the third radial spoke is absent in Lrrc23Δ/Δ sperm flagella.
(A, B) Sub-tomogram averaging (STA) to analyze structural defects at radial spoke (RS) of WT (A) and Lrrc23Δ/Δ sperm (B). Shown are STA images resulted from 96 nm doublet repeats from WT and Lrrc23Δ/Δ sperm. RS2 and 3 are magnified and density to represent RS3 head and the bridge between RS2 and RS3 (red circle) is missed in Lrrc23Δ/Δ sperm specifically. (C) Overwrapped STA images from 96 nm-doublet repeats from WT (gray) and Lrrc23Δ/Δ (gold) sperm, and Chlamydomonas reinhardtii (cyan). (D) A proposed model of impaired sperm motility and male infertility by the LRRC23 loss of function.
Figure 5—figure supplement 1
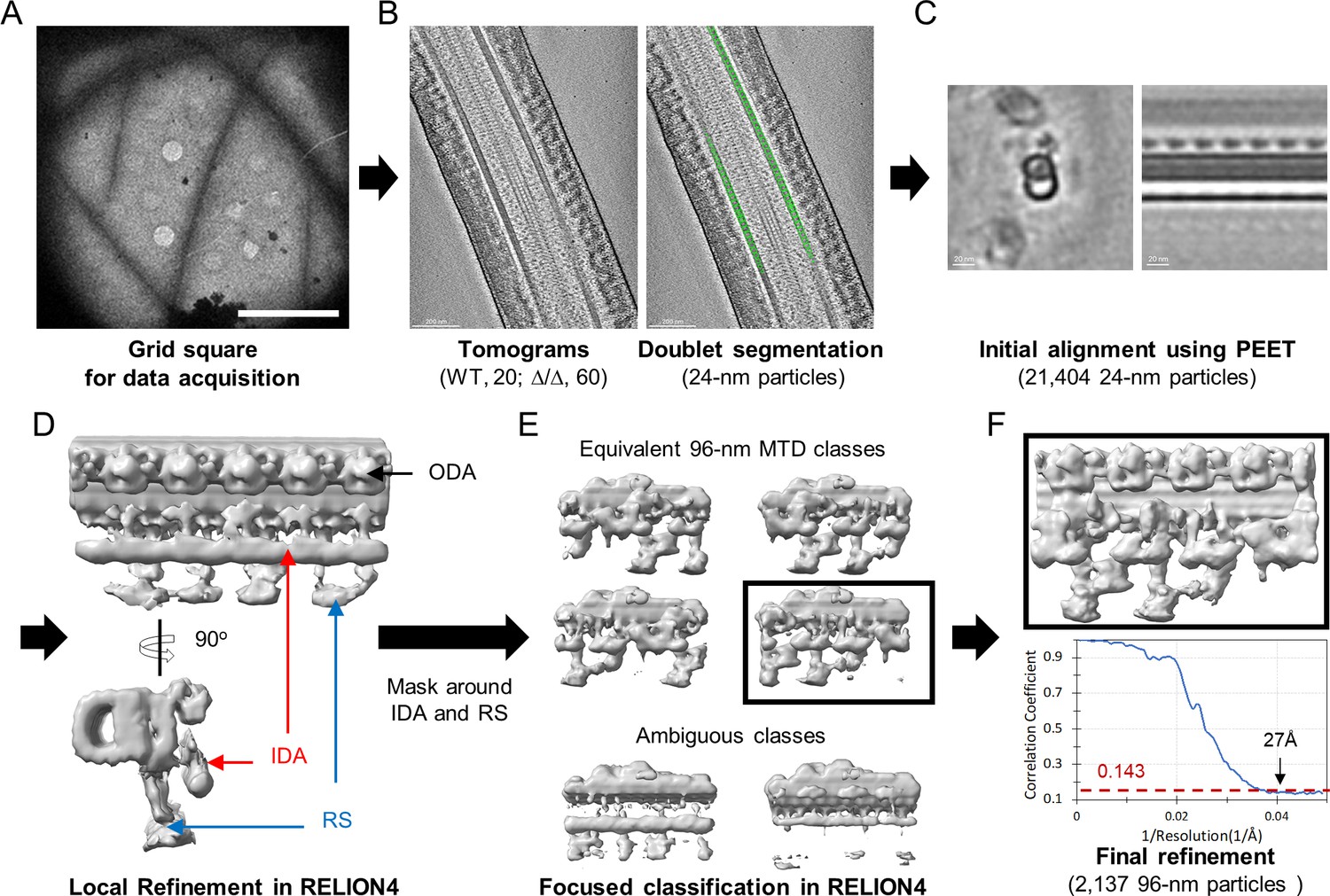
Workflow of cryo-electron tomography (cryo-ET) and sub-tomogram averaging (STA) processing of 96 nm microtubule doublet repeat from mouse sperm.
(A) An example of low magnification map of Lrrc23Δ/Δ mouse sperm on grid square. Scale bar, 10 µm. (B) An example tomographic slice obtained from sperm on the grid and segmented particles with 24 nm interval (green circles, right). (C) Initial alignment using PEET. Cross-sectional (left) and longitudinal (right) views are shown. (D) Local refinement of 24 nm particles in RELION4 after coordination and Euler angles transform. Density for outer dynein arm (ODA, black arrow) are visualized. Densities to represent inner dynein arm (IDA, red arrows) and radial spoke (RS, blue arrows) were averaged due to symmetry mismatch. (E) A soft-edge mask around IDA and RS. 96 nm repeat of microtubule doublet (MTD) classes with different translational symmetries (top) were obtained using focused classification in RELION4. The ambiguous classes (bottom) were discarded. (F) Final local refinement using one 96 nm repeat class. The gold-standard Fourier Shell Correlation (FSC) curve is shown at bottom.
Videos
Video 1
Lrrc23+/Δ and Lrrc23Δ/Δ sperm swimming freely in a viscous environment.
Free-swimming Lrrc23+/Δ and Lrrc23Δ/Δ sperm in the viscous condition containing 0.3% methylcellulose were recorded for 2 s before and after inducing capacitation for 90 min. Individual videos are played at 50 fps (1/2 speed).
Video 2
Flagellar waveform of Lrrc23+/Δ and Lrrc23Δ/Δ sperm before and after inducing capacitation.
Tail movements of head-tethered sperm from Lrrc23+/Δ and Lrrc23Δ/Δ males are recorded for 2 s before and after incubation under capacitating conditions for 90 min. Each video is played at 100 fps (1/2 speed).
Video 3
Tilted series of cryo-electron tomogram slices from WT (left) and Lrrc23Δ/Δ (right) spermatozoa.
Tomogram images were acquired of WT and Lrrc23Δ/Δ sperm were acquired on the grid and image slices were rendered to show the axonemal structure in a tilt series.
Additional files
-
Supplementary file 1
Clinical diagnosis of the infertile patients.
- https://cdn.elifesciences.org/articles/90095/elife-90095-supp1-v1.docx
-
Supplementary file 2
Variant Detail in the infertility Family.
- https://cdn.elifesciences.org/articles/90095/elife-90095-supp2-v1.docx
-
Supplementary file 3
Summarized imaging acquisition parameters and 3D refinement statistics.
- https://cdn.elifesciences.org/articles/90095/elife-90095-supp3-v1.docx
-
MDAR checklist
- https://cdn.elifesciences.org/articles/90095/elife-90095-mdarchecklist1-v1.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
LRRC23 truncation impairs radial spoke 3 head assembly and sperm motility underlying male infertility
eLife 12:RP90095.
https://doi.org/10.7554/eLife.90095.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}