Deep learning insights into the architecture of the mammalian egg-sperm fusion synapse
- Science for Life Laboratory and Department of Biochemistry and Biophysics, Stockholm University, Sweden
- Department of Biosciences and Nutrition, Karolinska Institutet, Sweden
- Department of Biology, Hull York Medical School, York Biomedical Research Institute, University of York, United Kingdom
Figures
Figure 1 with 1 supplement
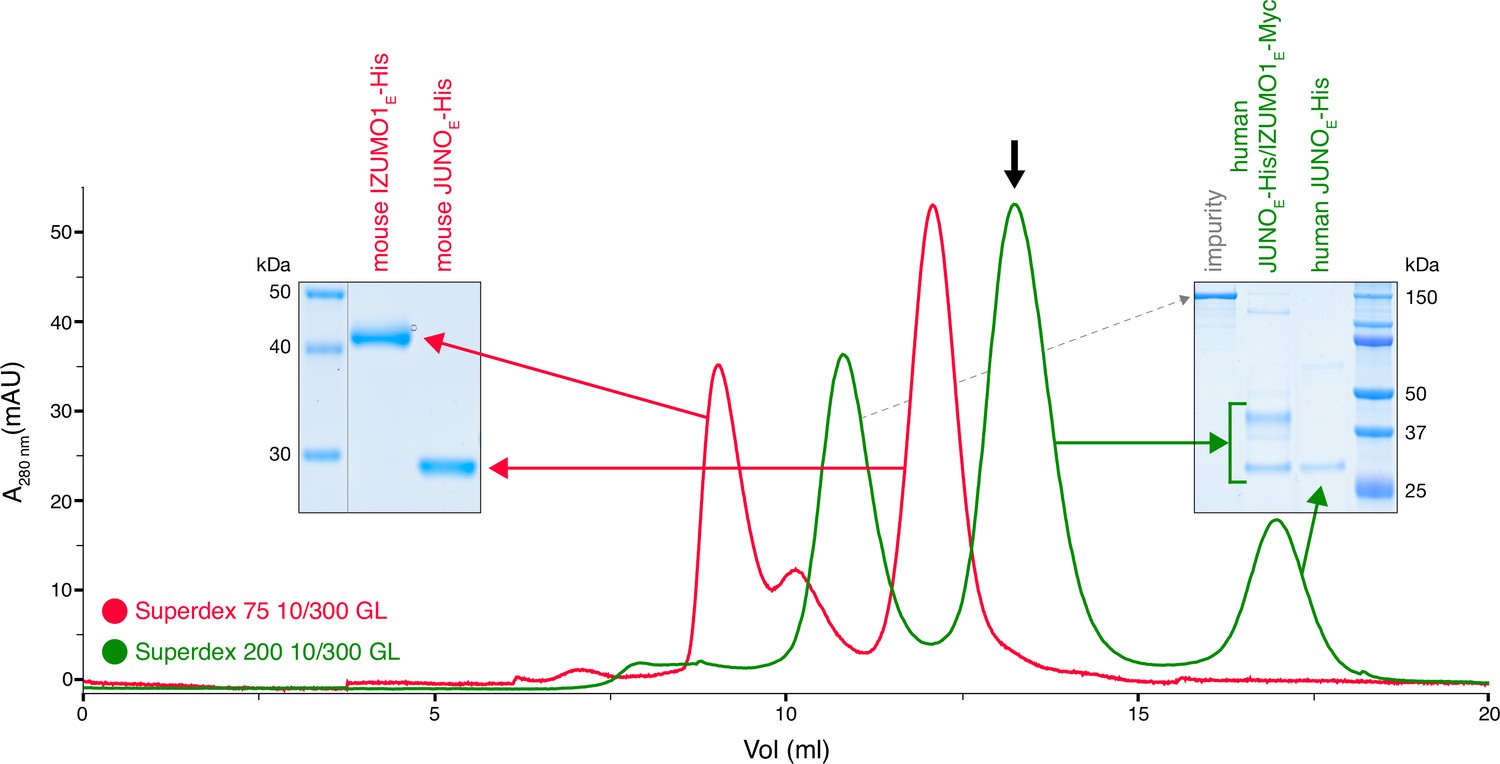
Human but not mouse JUNO and IZUMO1 ectodomains form a stable complex in solution.
The SEC elution profile of immobilized metal affinity chromatography (IMAC)-purified human JUNOE-His/IZUMO1E-Myc (green trace) shows a major peak that contains both proteins (black arrow), as well as a peak corresponding to unbound JUNOE-His (SDS-PAGE analysis on the right). On the contrary, IMAC-purified mouse JUNOE-His and mouse IZUMO1E-His elute separately on gel filtration (red trace and SDS-PAGE analysis on the left). See also Figure 1—figure supplement 1.
-
Figure 1—source data 1
Uncropped gel scans for Figure 1.
- https://cdn.elifesciences.org/articles/93131/elife-93131-fig1-data1-v1.zip
Figure 1—figure supplement 1

Immunoblot analysis of the SEC peak corresponding to the human JUNO/IZUMO1 ectodomain complex.
The peak indicated by a black arrow in Figure 1 was separated in parallel on two separate SDS-PAGE gels under reducing conditions and then probed by immunoblot using the indicated monoclonal antibodies (mAbs).
-
Figure 1—figure supplement 1—source data 1
Uncropped blot scans for Figure 1—figure supplement 1.
- https://cdn.elifesciences.org/articles/93131/elife-93131-fig1-figsupp1-data1-v1.zip
Figure 2
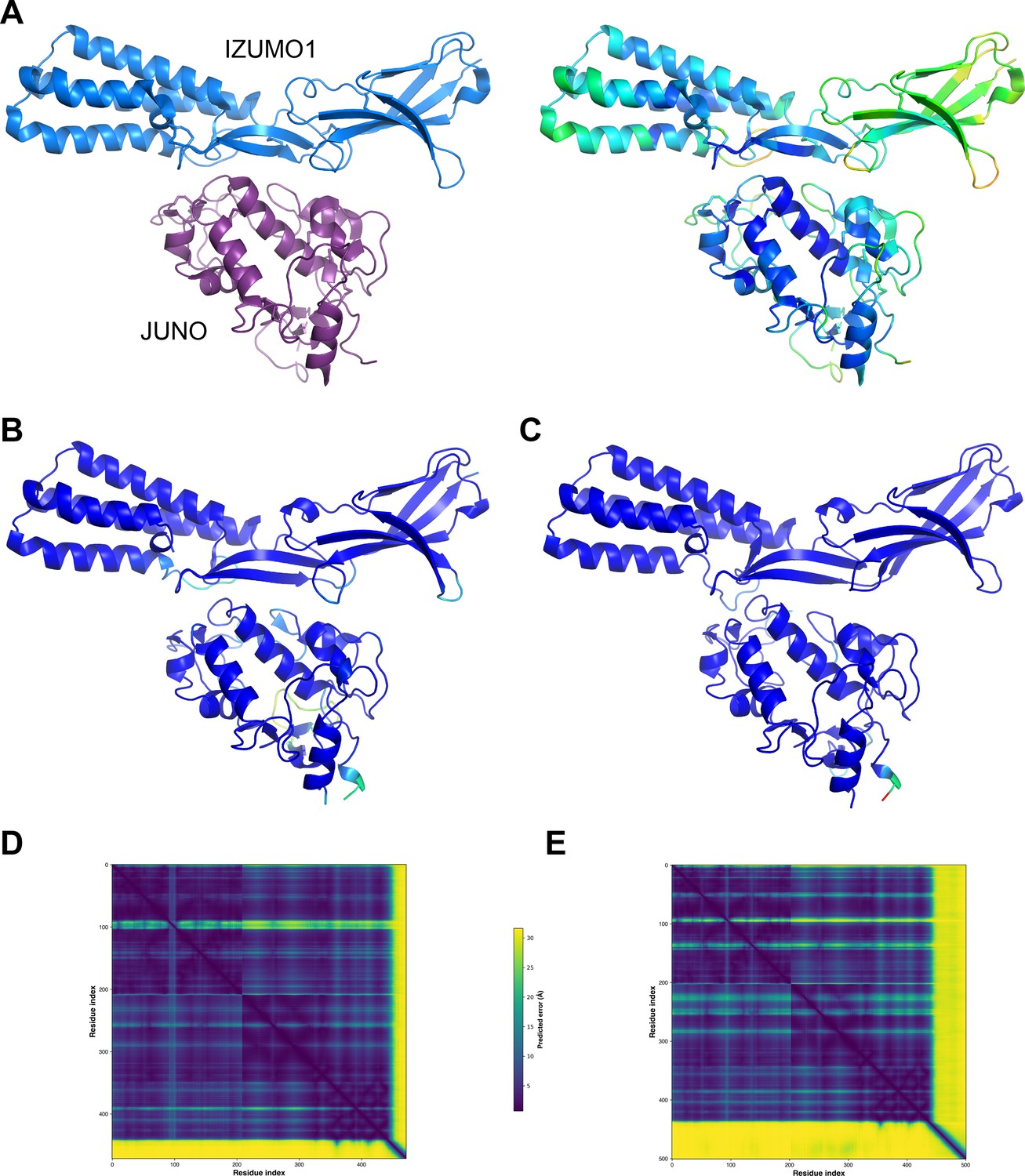
Mouse JUNO-IZUMO1 complex structure prediction.
(A) The crystal structure of the human JUNOE/IZUMO1E complex (PDB 5F4E; Aydin et al., 2016), shown in cartoon representation and colored by chain (left) or by B-factor (right). (B) AlphaFold-Multimer template-free prediction of the structure of the human JUNOE/IZUMO1E complex. The top-ranked model has a ranking confidence (rc = 0.8*predicted interface Template Modeling score (ipTM) +0.2*predicted Template Modeling score (pTM)) of 0.87, and an average root mean square deviation (RMSD) from PDB 5F4E of 2.34 Å over 437 Cα (0.88 Å over 380 Cα after outlier rejection). Only the residues that match those resolved in the crystal structure are shown; the model is colored by prediction confidence from blue to red, according to a 100‐(per‐residue confidence (predicted local distance difference test, pLDDT) Jumper et al., 2021) scale that ranges from 0 (blue; maximum confidence) to 100 (red; minimum confidence), respectively. (C) AlphaFold-Multimer top-ranked template-free prediction of the structure of the mouse JUNOE/IZUMO1E complex (rc = 0.85; RMSD vs. 5F4E=2.53 Å over 435 Cα (1.73 Å over 389 Cα after outlier rejection)), depicted and colored as in panel B. (D) Predicted Aligned Error (PAE) plot for the human complex model shown in panel B. Residue indexes refer to the sequence of JUNO (amino acids G20-S228) followed by that of IZUMO1 (amino acids C22-Q284). The high PAE regions correspond to loop 2 of JUNO (residues V110-G123) and the C-terminal tail of IZUMO1E (residues K255-Q284), both of which have low pLDDT scores and are far away from the interface between the two proteins. (E) PAE plot of the mouse complex shown in panel C, with residue indexes referring to JUNO (amino acids G20-G222) followed by IZUMO1 (amino acids C22-R319).
-
Figure 2—source data 1
Input and output files for the AlphaFold-Multimer predictions shown in panels B-E.
- https://cdn.elifesciences.org/articles/93131/elife-93131-fig2-data1-v1.zip
Figure 3
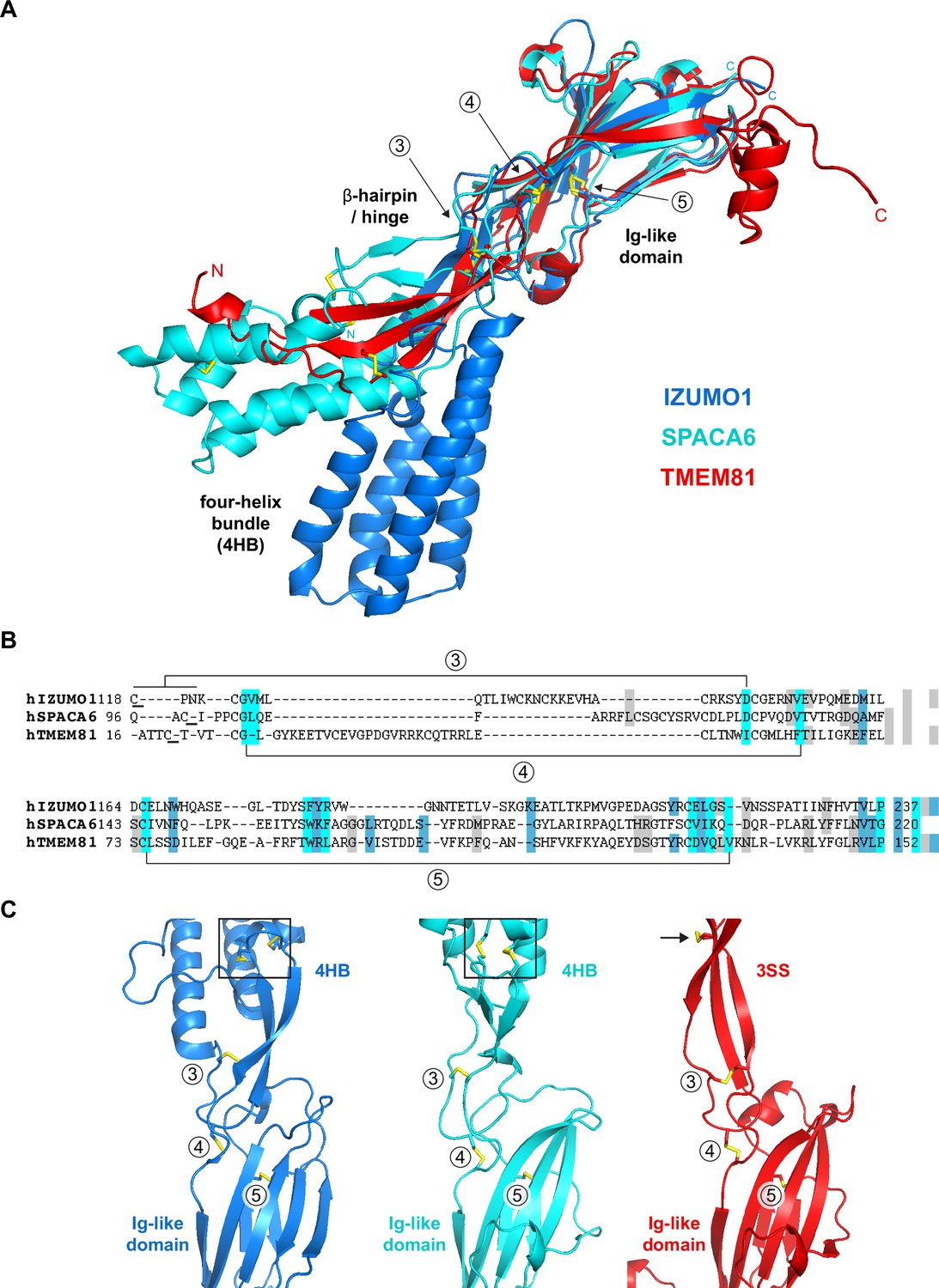
Structural homology between IZUMO1, SPACA6 and TMEM81.
(A) Structural superposition of the ectodomains of human IZUMO1 (residues C22-K255 of PDB 5JK9 chain A; Aydin et al., 2016), human SPACA6 (residues C27-G246 of PDB 7TA2; Vance et al., 2022) and an AlphaFold model of the ectodomain of human TMEM81 (corresponding to residues I31-P218 of UniProt entry Q6P7N7, extracted from https://alphafold.ebi.ac.uk/files/AF-Q6P7N7-F1-model_v4.pdb). The three different regions of IZUMO1 and SPACA6 are indicated in black. Disulfide bonds are shown as yellow sticks, with arrows indicating disulfides 3–5 of IZUMO1 that are conserved in both SPACA6 and TMEM81. N- and C-termini are marked. (B) Structure-based alignment of the sequence regions includes conserved disulfides 3 and 4, followed by the Ig-like domain harboring conserved disulfide 5. (C) Partial grid view of the superposition shown in panel A, centered around the junction between the three molecules' variable (top) and conserved (bottom) domains. Note the strikingly similar relative arrangement of invariant disulfides 3, 4, and 5, and how an additional disulfide within the three-stranded sheet (3SS) of TMEM81 (black arrow) roughly matches the position of the double CXXC motifs of IZUMO1 and SPACA6 (black boxes).
Figure 4
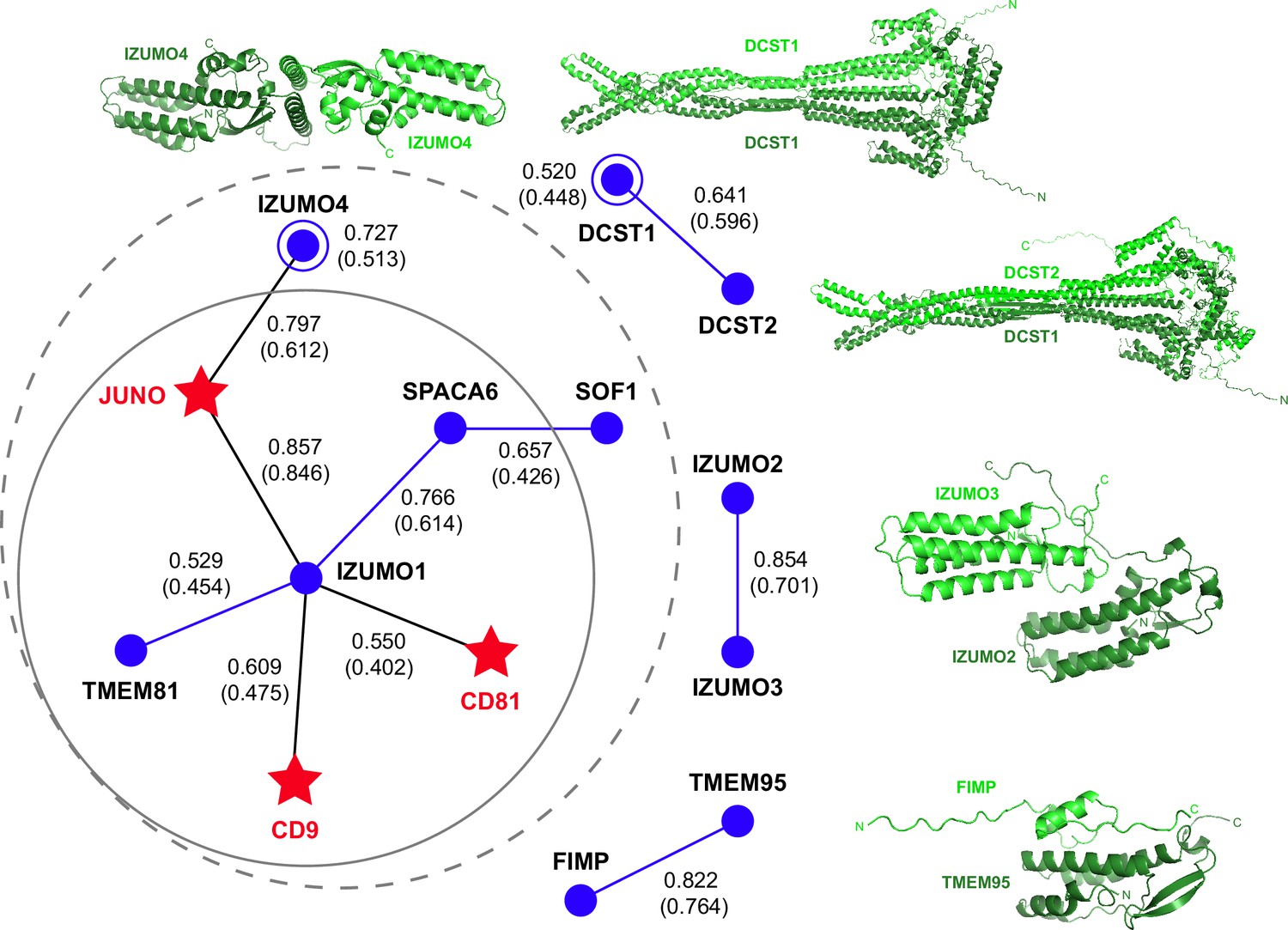
AlphaFold-Multimer prediction of interactions between fusion-associated human gamete proteins.
Egg and sperm proteins are indicated by red star and blue circle symbols, respectively. Interactions between egg and sperm proteins are shown as black lines connecting the respective symbols; homomeric and heteromeric interactions between sperm proteins are depicted as blue lines and open circles, respectively. For every interaction, the top-ranking model rc is reported, with the corresponding mean rc in parenthesis (for additional metrics, see Supplementary file 1). The gray dashed circle indicates a network of seven interactions, identified using a mean rc cutoff of 0.4; the inner continuous circle highlights the five interactions within the network that involve sperm IZUMO1. Top-ranked predictions for the isolated binary interactions of other sperm subunits are shown in cartoon representation, with the two moieties of each complex colored dark and light green and the N- and C-termini of each chain indicated when possible.
-
Figure 4—source data 1
Full metrics file and PDB files for all displayed AlphaFold-Multimer predictions.
- https://cdn.elifesciences.org/articles/93131/elife-93131-fig4-data1-v1.zip
Figure 5 with 1 supplement
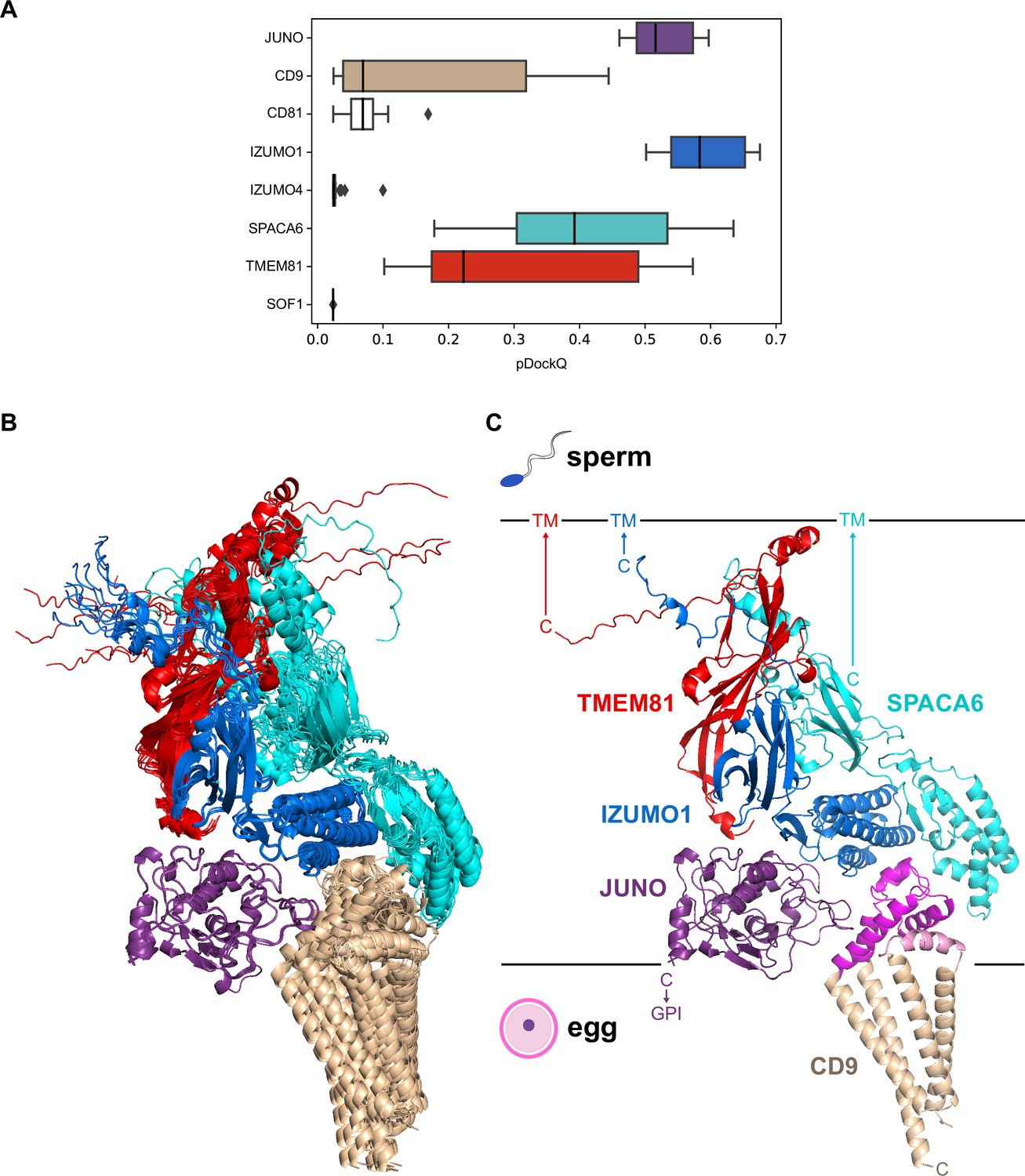
A predicted five-subunit complex at the egg/sperm plasma membrane interface.
(A) pDockQ analysis of 25 AlphaFold-Multimer predictions for a complex consisting of the 8 proteins enclosed by the dashed gray circle in Figure 4. The pDockQ score for each component of every prediction was calculated with respect to the rest of the corresponding complex, and the 25 scores for each chain were then plotted as a box plot. (B) Superposition of the ten top-ranked AlphaFold predictions for a five-subunit complex consisting of egg CD9 and the ectodomains of egg JUNO and sperm IZUMO1, SPACA6 and TMEM81 (mean rc = 0.67, mean ipTM = 0.66). Proteins are shown in cartoon representation and colored by chain according to panel A. (C) Top-ranked model from the ensemble in panel B (rc = 0.74, ipTM = 0.73). Subunits are colored as in the previous panels, except for CD9 whose short extracellular loop (SEL) and long extracellular loop (LEL) are highlighted in pink and magenta, respectively. Ectodomain (JUNO, IZUMO1, SPACA6, TMEM81) or protein (CD9) C-termini are marked, with horizontal lines representing the approximate surfaces of the gamete plasma membranes. TM, transmembrane domain; GPI, glycosylphosphatidylinositol anchor.
-
Figure 5—source data 1
Box plot data and input and output files for the AlphaFold-Multimer predictions shown in panels B and C.
- https://cdn.elifesciences.org/articles/93131/elife-93131-fig5-data1-v1.zip
Figure 5—figure supplement 1
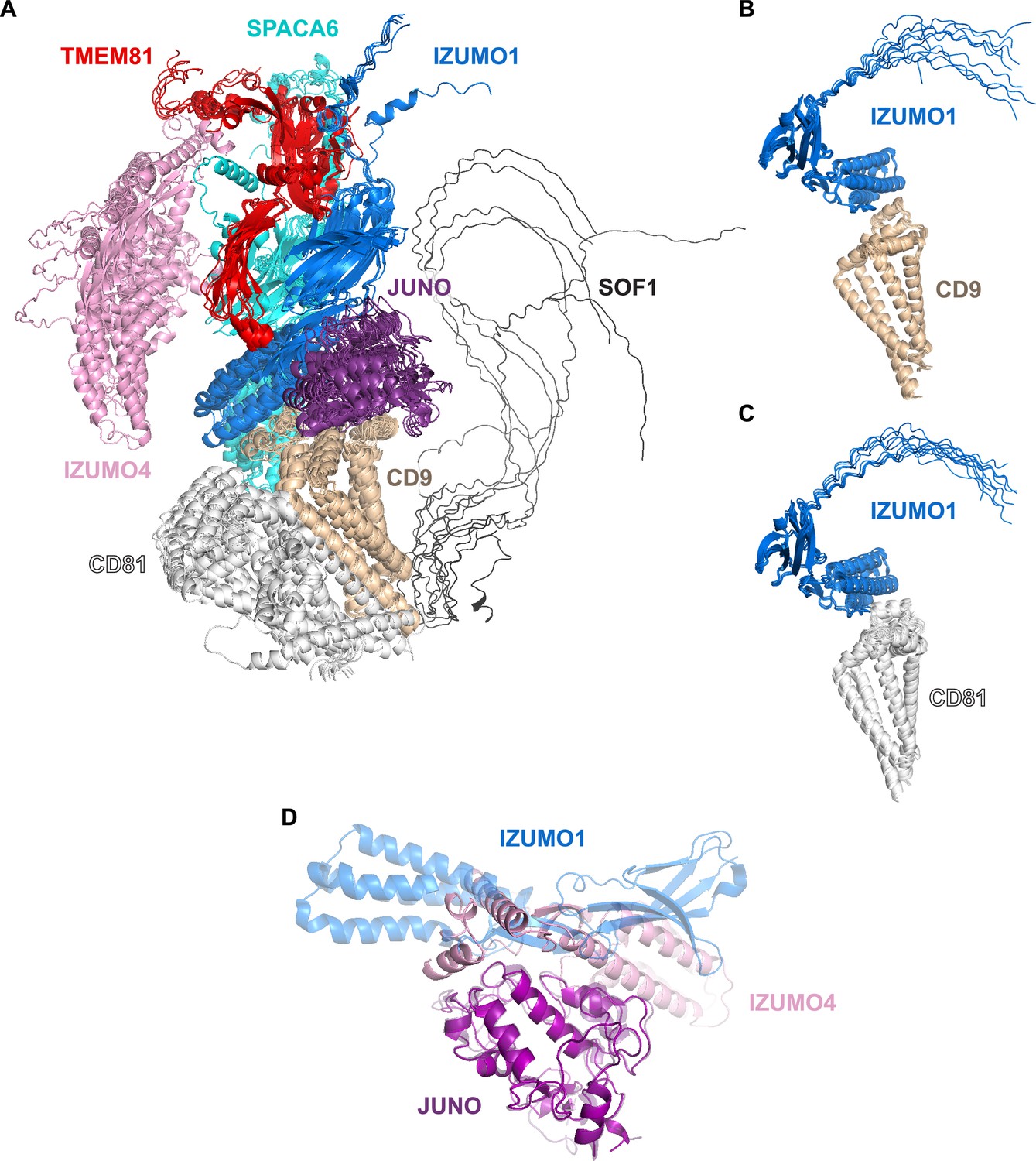
Modeling of the 8-protein network and binary subcomplexes thereof.
(A) Highest-scoring cluster of predictions for a complex that includes the 8 proteins enclosed by the dashed gray circle in Figure 4 (mean rc = 0.49, top rc = 0.51). (B) Superposition of the ten top-ranked predictions for the IZUMO1/CD9 binary interaction (mean rc = 0.57, top rc = 0.61). (C) Superposition of the ten top-ranked predictions for the IZUMO1/CD81 binary interaction (mean rc = 0.54, top rc = 0.55). (D) Comparison of the human JUNOE/IZUMO1E complex crystal structure (PDB 5F4E; Aydin et al., 2016) and the top-ranked prediction for a binary complex consisting of JUNOE and IZUMO4 (rc = 0.80). The models have been superimposed over JUNOE (RMSD 2.5 Å over 204 Cα atoms, or 0.6 Å over 166 Cα atoms after outlier rejection) and colored as in Figure 2A and A, respectively, with the experimental structure cartoon shown semi-transparent.
-
Figure 5—figure supplement 1—source data 1
PDB files for all displayed AlphaFold-Multimer predictions.
- https://cdn.elifesciences.org/articles/93131/elife-93131-fig5-figsupp1-data1-v1.zip
Figure 6
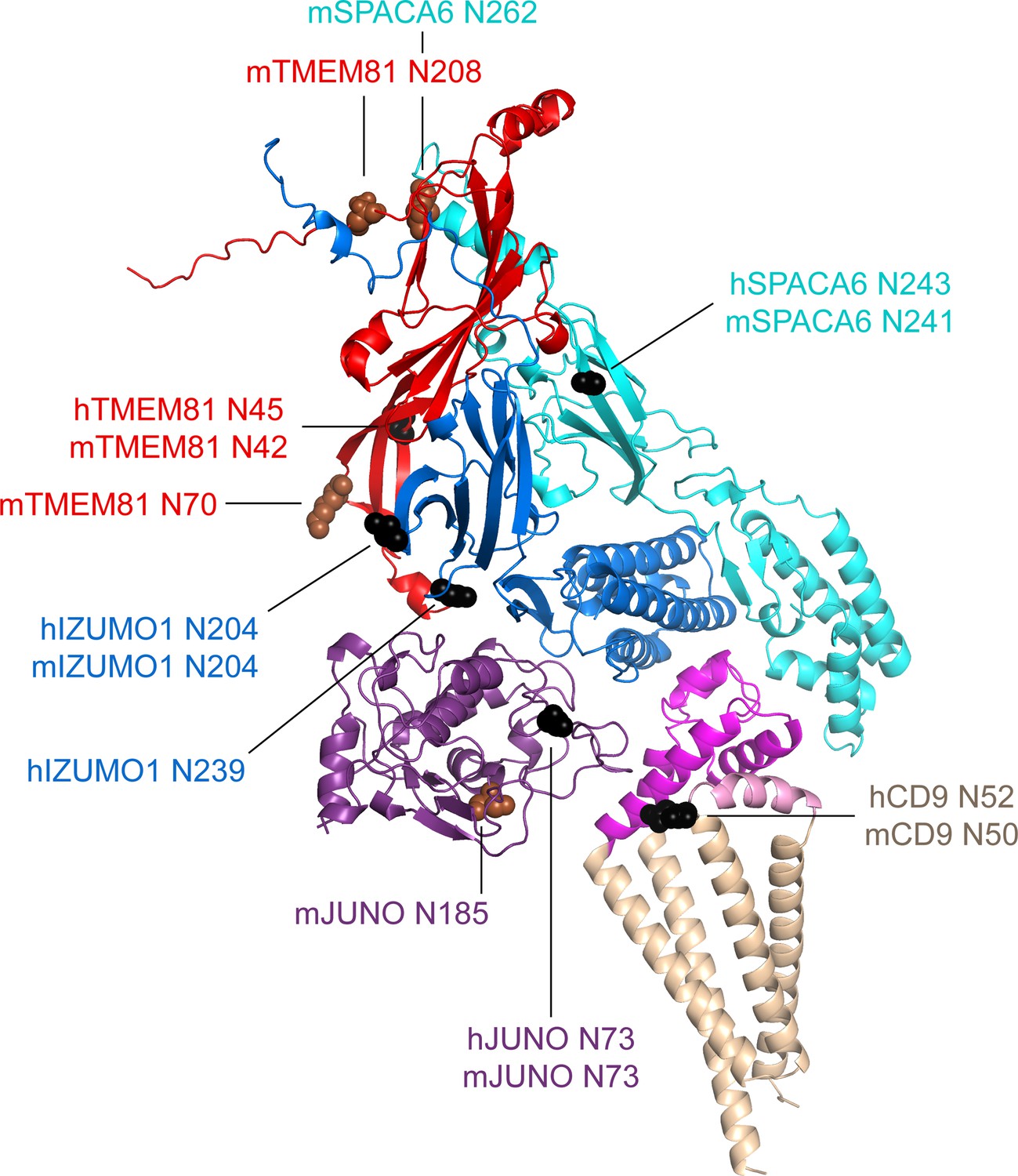
Mapping of sequon positions onto the 5-subunit complex prediction.
The positions of possible glycosylation sites are mapped onto the model of the 5-subunit assembly (depicted as in Figure 5C) by showing the corresponding Asn residues in sphere representation. Sequons found in human (h prefix) or in both human and mouse proteins are colored black, whereas sequons only found in mouse (m prefix) proteins are brown.
-
Figure 6—source data 1
Input and output files for the displayed AlphaFold-Multimer prediction.
- https://cdn.elifesciences.org/articles/93131/elife-93131-fig6-data1-v1.zip
Figure 7
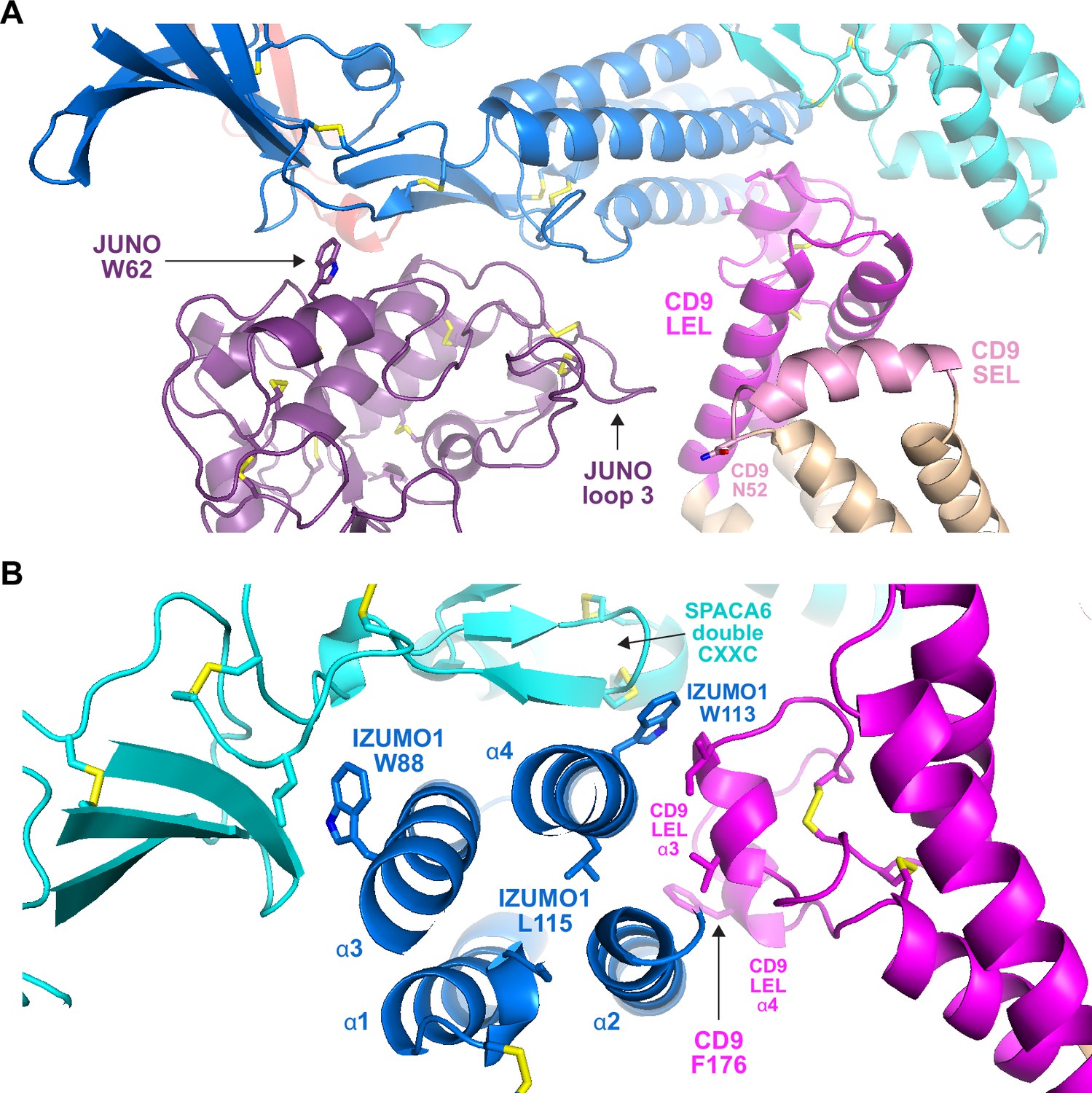
Subunit interfaces of the predicted complex involve protein elements previously implicated in fusion.
(A) Detail of the prediction shown in Figure 5C, highlighting functionally important regions of JUNO and CD9, as well putatively N-glycosylated CD9 N52. (B) Different view of the same complex prediction, centered around the IZUMO1 4HB.
-
Figure 7—source data 1
Input and output files for the displayed AlphaFold-Multimer prediction.
- https://cdn.elifesciences.org/articles/93131/elife-93131-fig7-data1-v1.zip
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Cell line (Homo sapiens) | HEK293T | Dr. Radu Aricescu and Dr. Yuguang Zhao (University of Oxford, UK) | ||
| Cell line (Homo sapiens) | HEK293S GnTI- | ATCC | CRL-3022; RRID:CVCL_A785 | |
| Antibody | Mouse IgG1 monoclonal Penta·His | QIAGEN | 34660; RRID:AB_2619735 | (1:1000) |
| Antibody | Mouse IgG1 monoclonal Anti-c-Myc (clone 9E10) | Sigma-Aldrich | M4439; RRID:AB_439694 | (1:5000) |
| Antibody | Peroxidase AffiniPure Goat Anti-Mouse IgG (H+L) | Jackson ImmunoResearch Laboratories | 115-035-003; RRID:AB_10015289 | (1:10000) |
| Transfected construct (Homo sapiens) | pHLsec3-hJUNO-(GGGS)2H8 | This publication | Methods section “DNA constructs” | |
| Transfected construct (Homo sapiens) | pHLsec3-hIZUMO1-Myc | This publication | Methods section “DNA constructs” | |
| Transfected construct (Homo sapiens) | pHLsec3-mJuno-H8 | Han et al., 2016 | ||
| Transfected construct (Homo sapiens) | pHLsec3-mIzumo1-LEH6 | Nishimura et al., 2016 | ||
| Chemical compound, drug | 25 kDa branched polyethyleneimine | Sigma-Aldrich | 408727 | |
| Chemical compound, drug | SimplyBlue SafeStain | Thermo Fisher Scientific | LC6060 | |
| Commercial assay or kit | PCR Mycoplasma Test Kit II | Applichem | A8994 | |
| Software, algorithm | AlphaFold2 | Jumper et al., 2021; Evans et al., 2021 | ||
| Software, algorithm | Belvu | Barson and Griffiths, 2016 | ||
| Software, algorithm | Dali | Holm, 2020 | ||
| Software, algorithm | Foldseek | van Kempen et al., 2024 | ||
| Software, algorithm | pDockQ | Bryant et al., 2022 | ||
| Software, algorithm | PyMOL | Schrödinger, LLC | ||
| Software, algorithm | UCSF Chimera | Meng et al., 2006 |
Additional files
-
Supplementary file 1
Evaluation metrics for protein complex predictions.
- https://cdn.elifesciences.org/articles/93131/elife-93131-supp1-v1.csv
-
MDAR checklist
- https://cdn.elifesciences.org/articles/93131/elife-93131-mdarchecklist1-v1.pdf
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Deep learning insights into the architecture of the mammalian egg-sperm fusion synapse
eLife 13:RP93131.
https://doi.org/10.7554/eLife.93131.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}