Isoleucine gate blocks K+ conduction in C-type inactivation
- Laboratório de Biologia Teórica e Computacional (LBTC), Universidade de Brasília, Brazil
- Department of Biochemistry and Molecular Biology, The University of Chicago, United States
- Department of Neurobiology, The University of Chicago, United States
- Laboratoire International Associé Centre National de la Recherche Scientifique et University of Illinois at Urbana−Champaign, Unité Mixte de Recherche No. 7019, Université de Lorraine, Université de Lorraine, France
- NIH Center for Macromolecular Modeling and Bioinformatics, Beckman Institute for Advanced Science and Technology, and Department of Physics, University of Illinois at Urbana−Champaign, United States
- Centro Interdisciplinario de Neurociencia de Valparaíso, Facultad de Ciencias, Universidad de Valparaíso, Chile
Figures
Figure 1
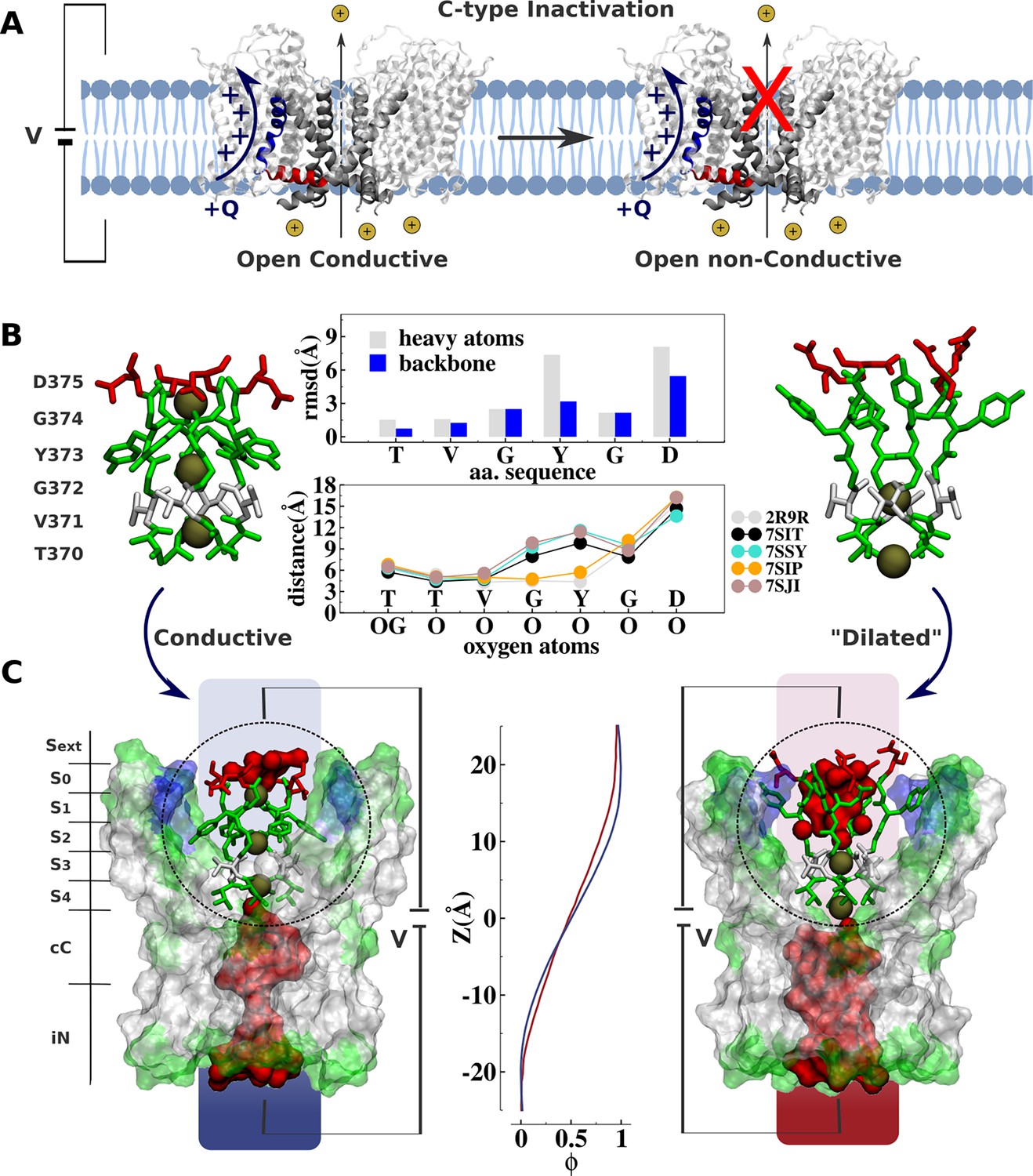
Comparative analysis of Kv channel structures.
(A) Schematic representation of a voltage-gated K+ channel undergoing C-type inactivation, whereby prolonged activation by an external voltage V leads to blockage of ionic conduction across the selectivity filter of the open channel. The voltage-sensor positively charged S4 helix (blue), the S4S5 linker (red), and main-pore S6 helix (silver) are highlighted. (B) Structural models of the selectivity filter in the conductive and dilated conformations. The conductive and dilated conformations derive respectively from the high-resolution X-ray structures of the wild-type (Long et al., 2007) and triple-mutant (Reddi et al., 2022) kv1.2-kv2.1 channel (PDB codes 2R9R and 7SIT). Major structural deviations (root-mean-square deviation [RMSD]) between the selectivity filter conformations are primarily accounted for side-chain rearrangements of Y373 and D375. Despite the side-chain rearrangements of Y373 and D375, the profile of oxygen-oxygen distances between opposing subunits of the selectivity filter indicates that the geometry of sites S4 and S3 in the dilated conformation closely resembles that of the conductive state. For comparison purposes, the profile of oxygen-oxygen distances is also shown along the selectivity filter of the experimental structures of the conductive state of Shaker B (Tan et al., 2022) (PDB code 7SIP) and the dilated conformations of Shaker-W434F (Tan et al., 2022) (PDB code 7SJI) and Kv1.3 (Selvakumar et al., 2022) (PDB code 7SJ1). (C) Electrostatic properties of the conductive and dilated conformations of the selectivity filter. Shown are molecular representations of the main-pore S6 segments of the channel, highlighting the permeation pathway along the intracellular entrance (iN), central cavity (cC), and selectivity-filter sites (S4, S3, S2, S1, S0, Sext). Hydrated cavities (red) give ionic access to the selectivity filter from the intracellular and extracellular milieu. The dielectric morphology of the protein and waters accounts for a significant voltage drop (ɸ) across the selectivity filter. The voltage-drop profile along the permeation pathway was computed as described elsewhere (Souza et al., 2014), following the charge-imbalance protocol which is a variant of the linear field method (Roux, 2008).
Figure 2 with 2 supplements
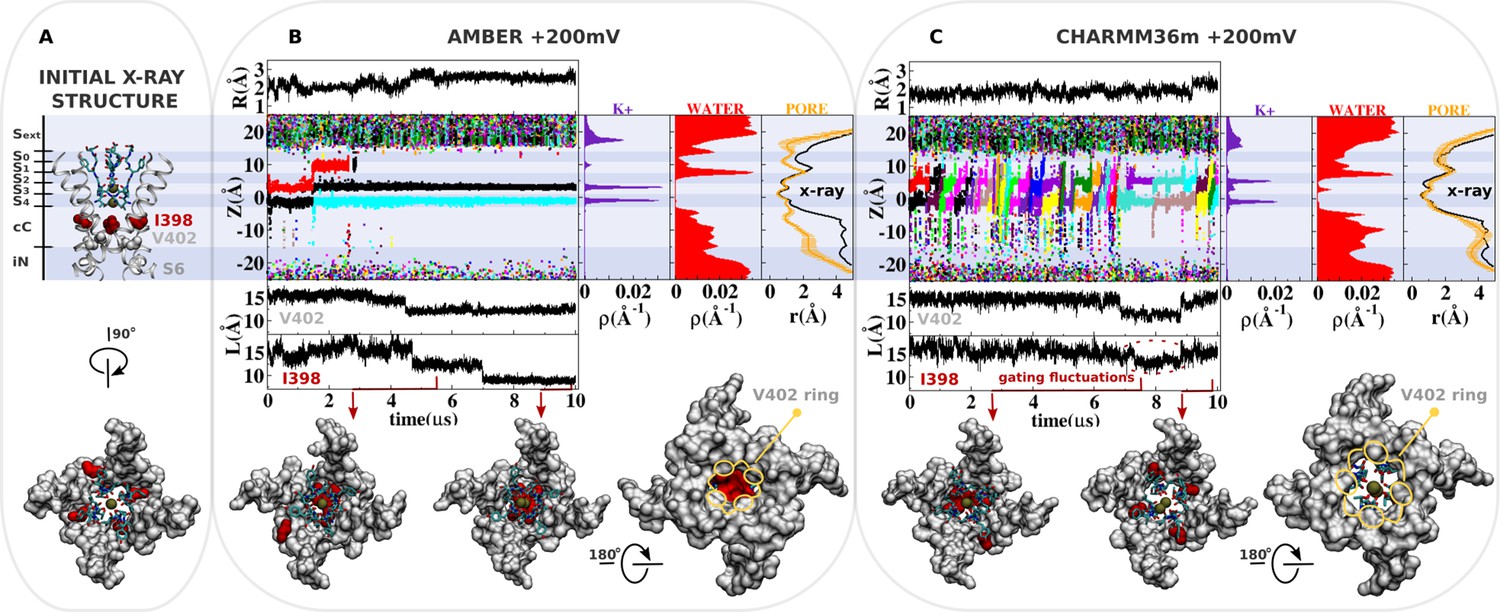
Molecular dynamics (MD) simulation of kv1.2-kv2.1-3m at +200 mV.
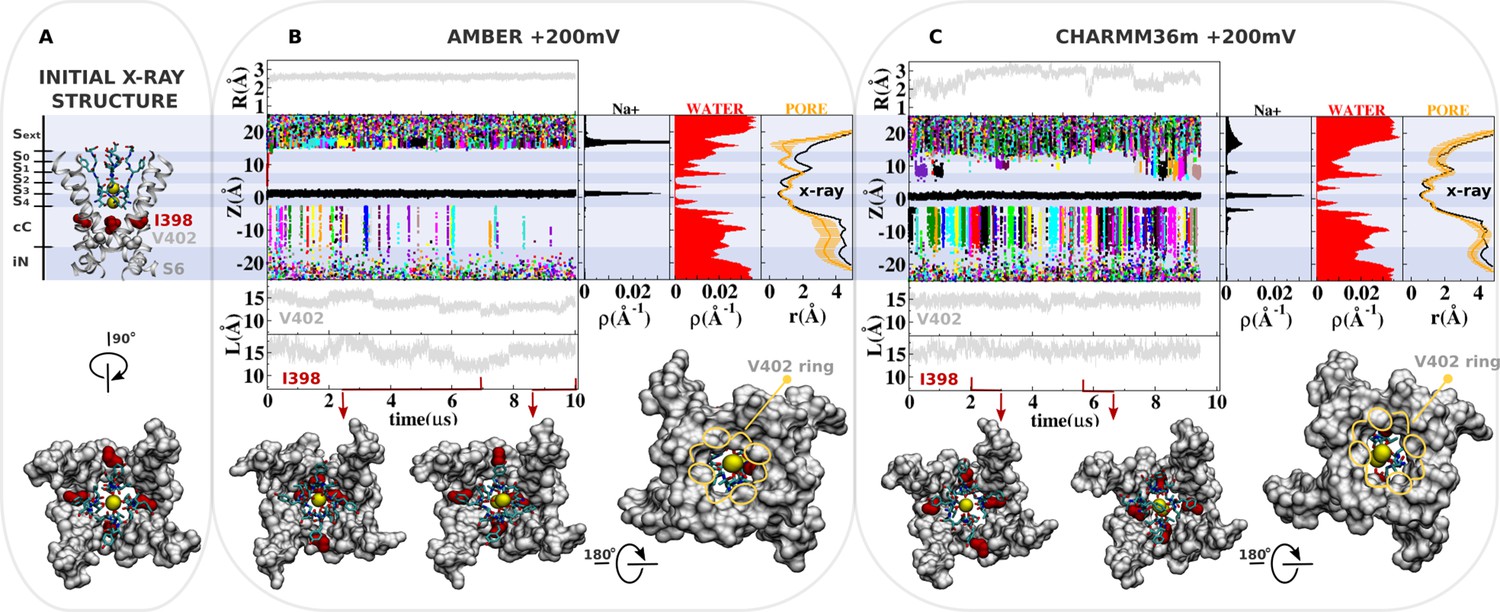
(A) Molecular representation of the main pore of the channel, highlighting the initial configuration of the selectivity filter, I398 (red) and V402 (light gray). (B and C) Analysis of AMBER and CHARMM36m trajectories. Shown is the structural deviation of the selectivity filter (R), the trajectory of K+ ions along the permeation pathway (Z), and the intersubunit Cβ-Cβ separation distance of I398 and V402 (L) as a function of simulation time. Inset shows instantaneous configurations of I398 (red arrows). Time averages are the linear density of K+ ions, the linear density of water oxygen, and the pore radius profile (r) along the permeation pathway Z.
Figure 2—figure supplement 1
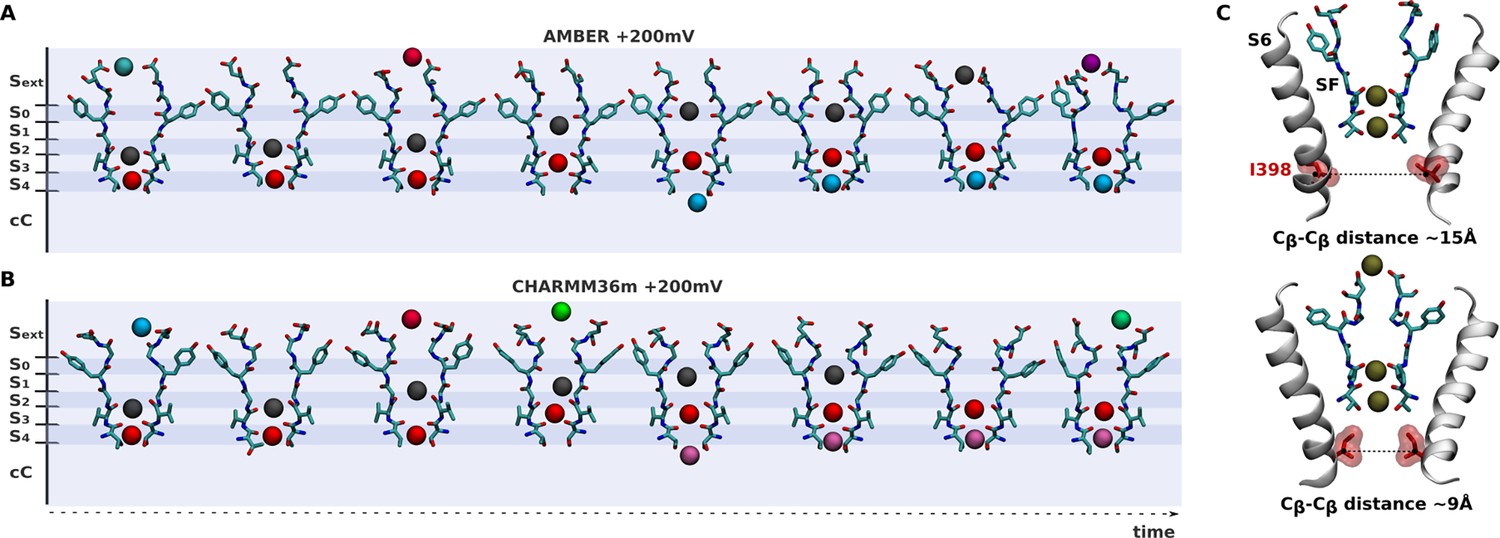
Ion conduction across the dilated conformation of the selectivity filter of kv1.2-kv2.1-3m.
(A, B) Shown are representative single-ion conduction events across the selectivity filter of the triple-mutant channel along AMBER and CHARMM36m simulations. (C) Closure of the isoleucine gate blocks ion conduction across the the selectivity filter.
Figure 2—figure supplement 2

Molecular dynamics (MD) simulation of kv1.2-kv2.1-3m at +200mV.
(A) Molecular representation of the main pore of the channel, highlighting the initial configuration of the selectivity filter, I398 (red) and V402 (light gray). (B and C) Analysis of AMBER* (restrained to keep I398 open) and CHARMM36m-NBFIX trajectories. Shown are the structural deviation of the selectivity filter (R), trajectory of K+ along the permeation pathway (Z), and the intersubunit Cβ-Cβ separation distance of I398 and V402 (L) as a function of simulation time. Inset shows instantaneous configurations of I398 (red arrows). Time averages are the linear density of K+ ions, the linear density of waters, and the pore radius profile (r) along the permeation pathway Z.
Figure 3 with 5 supplements
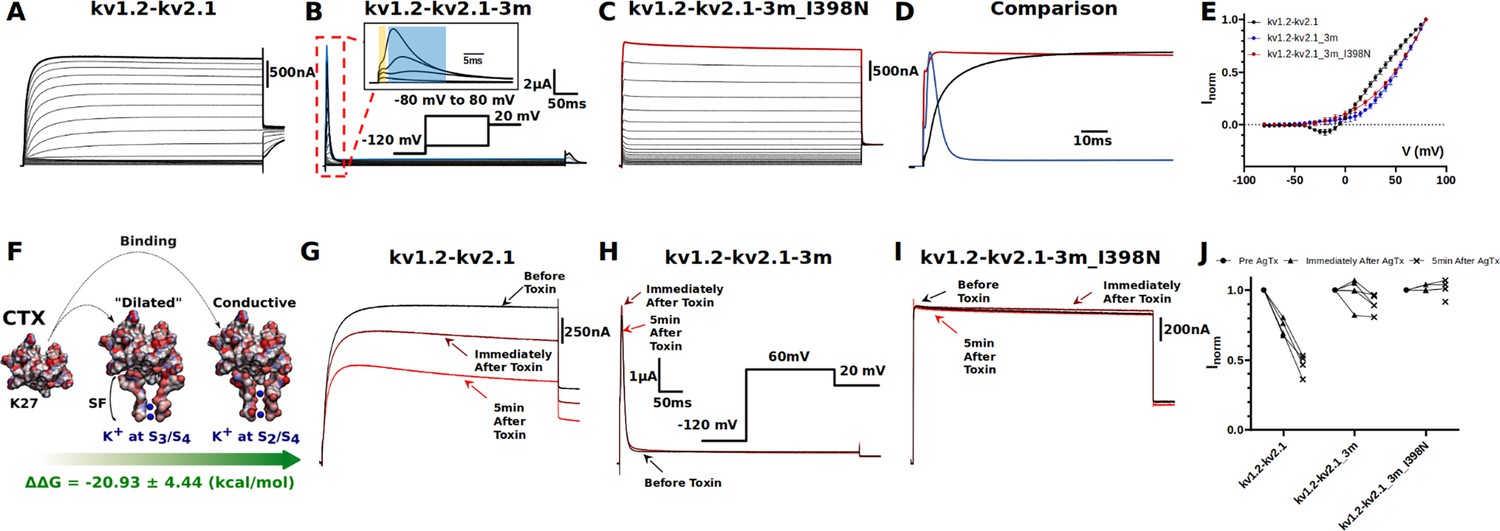
Electrophysiology measurements of I398N substitution.
(A, B, C, and D) Macroscopic current recorded from: (A) kv1.2-kv2.1 chimera, (B) triple-mutant kv1.2-kv2.1-3m, (C) triple-mutant kv1.2-kv2.1-3m with I398N substitution, and (D) their respective comparison at +80 mV (the line colors correspond to the colors indicated in A, B, and C of the three mutants at +80 mV). The triple mutation W362F, S367T, and V377T in kv1.2-kv2.1-3m significantly speed up the inactivation process and the gating current could be seen simultaneously with ionic current (shown in inset, with gating current highlighted in yellow and ionic current highlighted in blue). Note the effect of the triple mutation cancel out with the I398N substitution. (E) Current-voltage relationship (IV curve). Due to the mixture of gating current and ionic current in kv1.2-kv2.1-3m, only curves shown are taken from the peak current. Voltage-dependent activation relationship for kv1.2-kv2.1, kv1.2-2.1-2m, and kv1.2–2.1-2m_I398N are shown in Figure 3—figure supplement 3. (F) Net free-energy difference (ᐃᐃG) involved in the binding of the charybdotoxin (CTX) to the conductive (Banerjee et al., 2013) and dilated conformations of the selectivity filter of the kv1.2-kv2.1 chimera channel. ᐃᐃG indicates a strong preference of CTX to the conductive conformation of the selectivity filter. The same binding preference is found between CTX and kv1.2-kv2.1-3m (Supplementary file 5). (G, H, I, and J) Effects of agitoxin-II, a more potent CTX analogous (Takacs et al., 2009), on kv1.2-kv2.1 chimera, triple-mutant kv1.2-kv2.1-3m, triple-mutant kv1.2-kv2.1-3m with I398N substitution. Clearly, agitoxin binds and blocks the chimera channel while shows minimal influence on triple and triple_I398N, suggesting the selectivity filter in triple_I398N likely also adopts a dilated conformation.
Figure 3—figure supplement 1
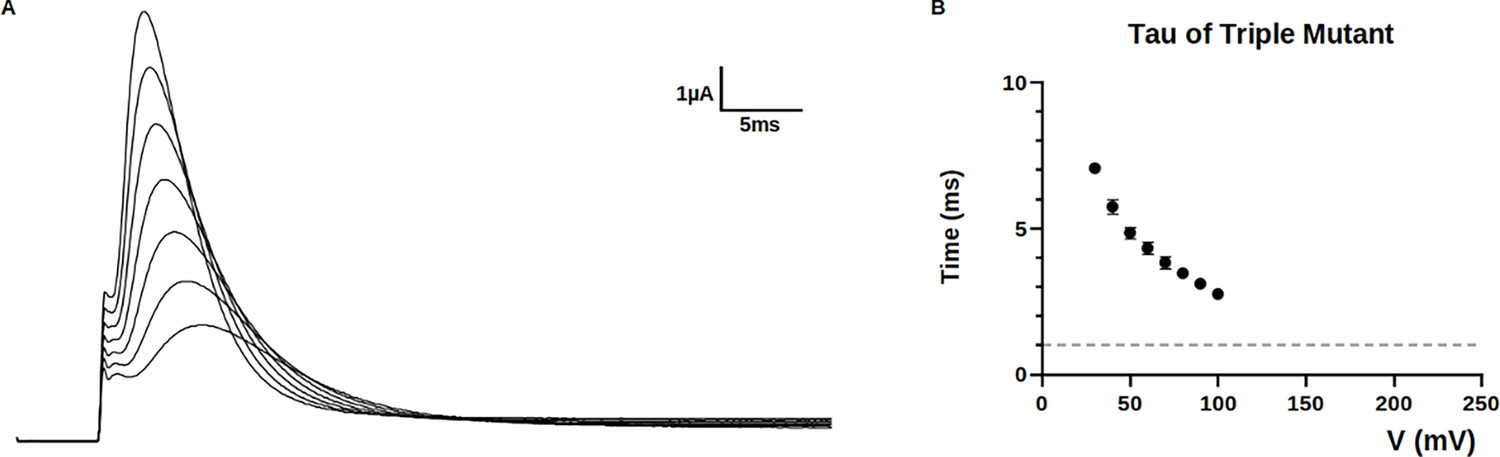
Decay of the ionic current of the triple-mutant kv1.2-kv2.1-3m channel.
(A) Ionic traces from the triple-mutant channel and (B) its time constants fitted with a one exponential decay. When extrapolated to +200 mV where the simulation was performed, the time constant (tau) was in the microsecond range (gray dashed line indicates 1 ms), suggesting the timescale of the simulation and the conformational changes seen were relevant in physiological terms. Data plotted with mean and SEM.
Figure 3—figure supplement 2
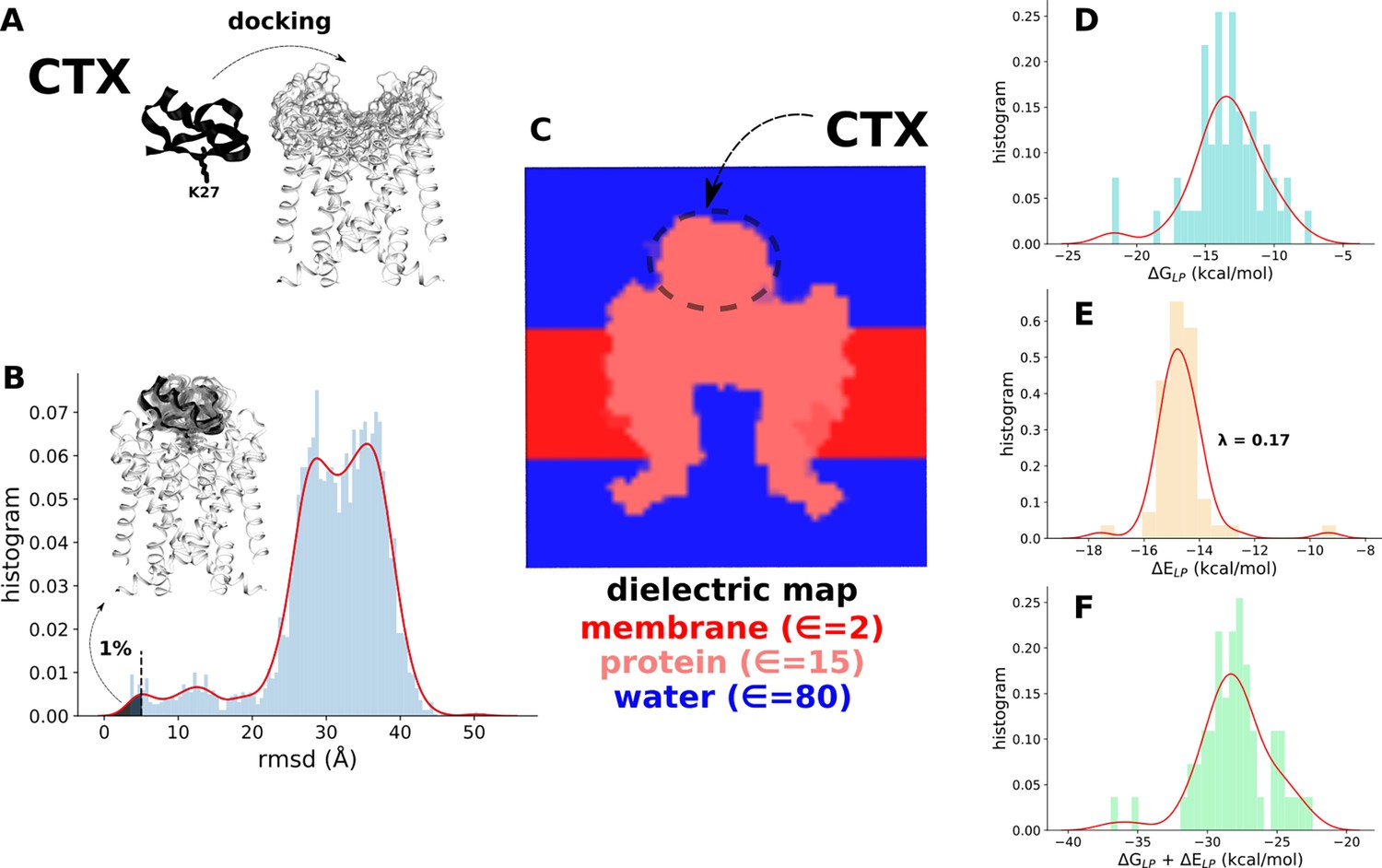
Estimation of the binding free energy of charybdotoxin (CTX).
(A) Docking of CTX to the selectivity filter of the kv1.2-kv2.1 chimera channel. (B) Docking solutions (light black traces) best reproducing the experimentally resolved bound state of the toxin (tick black trace, PDB 4JTD). (C) Dielectric (∈) map of the toxin-protein complex considered in the Poisson-Boltzmann calculation. (D, E, and F) Energy distribution of best docking solutions shown in (B). Poisson-Boltzmann (ᐃGLP) and van der Waals (ᐃELP) electrostatic contributions to the binding free energy of CTX are shown. The van der Waals component was scaled by an empirical factor λ=0.17, intended to resolve the protein-solvent interaction absent in the implicit solvent representation. The same procedure was adopted to estimate the binding energy of CTX to all channel constructs and states (Supplementary file 5).
Figure 3—figure supplement 3
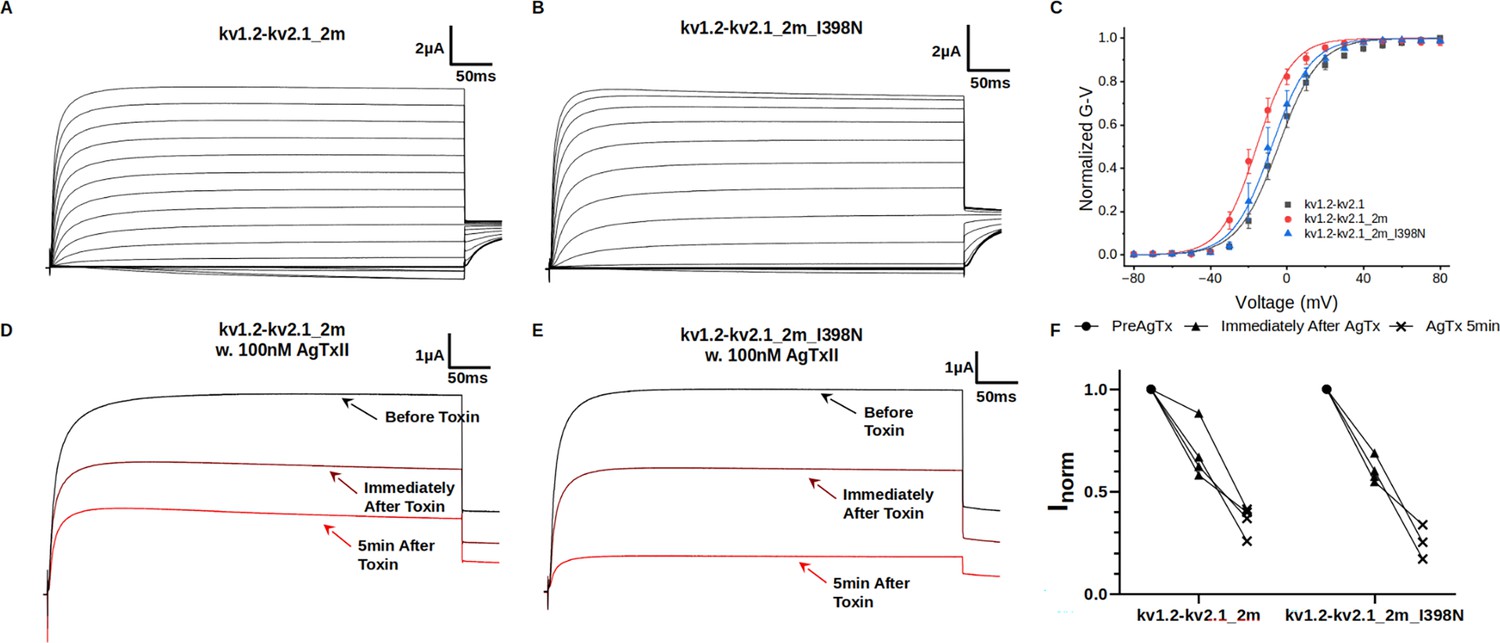
State dependence of I398N effects.
(A) Macroscopic current from the double-mutant chimera channel kv1.2-kv2.1-2m (S367T/V377T). (B) Macroscopic current from kv1.2-kv2.1-2m with I398N substitution. Mutation I398N in the absence of W362F does not significantly alter the phenotype of the channel. (C) G-V curves for kv1.2-kv2.1, kv1.2–2.1-2m, and kv1.2–2.1-2m_I398N. The G-V curves were measured from the tail currents and fitted using a two-state model given by: , where z is the apparent charge expressed in units of elementary charge , V is the voltage and is the voltage of half maximal conductance. R, T, and F have their usual meanings. All three mutants share similar voltage dependency in activation: V1/2 (mV): kv1.2-kv2.1 (–4.6±0.7), kv1.2-2.1-2m (–15.9±0.6) and kv1.2-2.1-2m_I398N (–8.1±0.6) and z: kv1.2-kv2.1 (2.4±0.1), kv1.2-2.1-2m (2.7±0.2) and kv1.2-2.1-2m_I398N (2.5±0.1). All three mutants share similar voltage dependency in activation. (D, E, and F) Effects of 100 nM agitoxin-II (AgTxII) on the double-mutant kv1.2-kv2.1-2m and double-mutant kv1.2-kv2.1-2m with I398N substitution. AgTxII binds and blocks efficiently both channel constructs, suggesting the effect of I398N depends on the C-type inactivation induced by the W362F mutation.
Figure 3—figure supplement 4
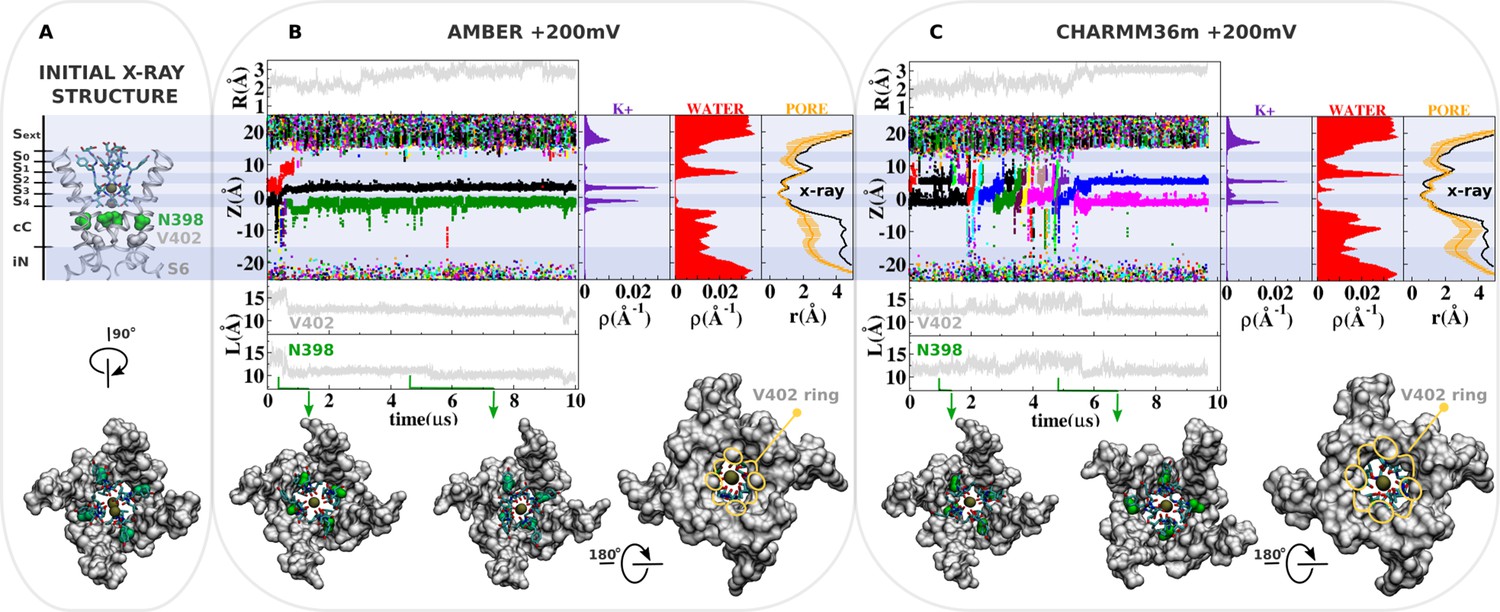
Molecular dynamics (MD) simulation of kv1.2-kv2.1-3m with I398N at +200mV.
(A) Molecular representation of the main pore of the channel, highlighting the initial configuration of the selectivity filter, N398 (green) and V402 (light gray). (B and C) Analysis of AMBER and CHARMM36m trajectories. Shown are the structural deviation of the selectivity filter (R), trajectory of K+ along the permeation pathway (Z), and the intersubunit Cβ-Cβ separation distance of N398 and V402 (L) as a function of simulation time. Inset shows instantaneous configurations of N398 (green arrows). Time averages are the linear density of K+ ions, the linear density of waters, and the pore radius profile (r) along the permeation pathway Z.
Figure 3—figure supplement 5

Primary-sequence conservation throughout the main-pore segments PH, SF, and S6.
(A) Logos conservation across K+ channels. (B) Multiple sequence alignment of most studied K+ channels.
Figure 4 with 3 supplements
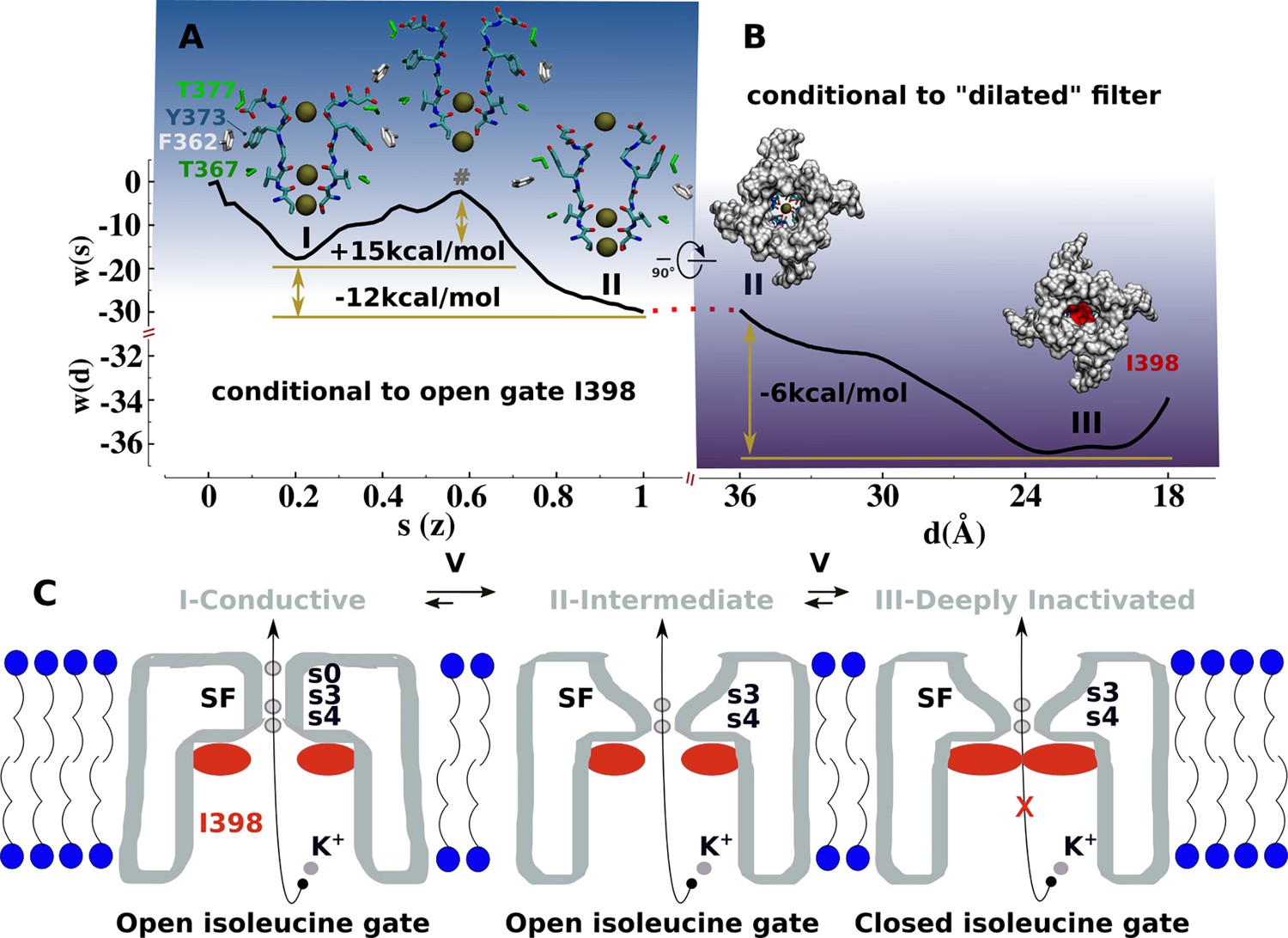
Mechanism of C-type inactivation of the triple-mutant channel kv1.2-kv2.1-3m.
(A) Free-energy profile w(s) along the conformational transition path s connecting the conductive (s=0.2) and dilated (s=1) states of the selectivity filter. The free-energy profile is conditional to the open configuration of the isoleucine gate. (B) Free-energy profile w(d) associated to closure of the isoleucine gate. The reaction coordinate d corresponds to the inter-subunit separation distance between β-carbon atoms of I398. The free-energy profile is conditional to the dilated conformation of the selectivity filter. (C) C-type inactivation mechanism of the triple-mutant channel inferred from the representative structures of the selectivity filter and isoleucine gate along the free-energy profiles (A) and (B).
Figure 4—figure supplement 1
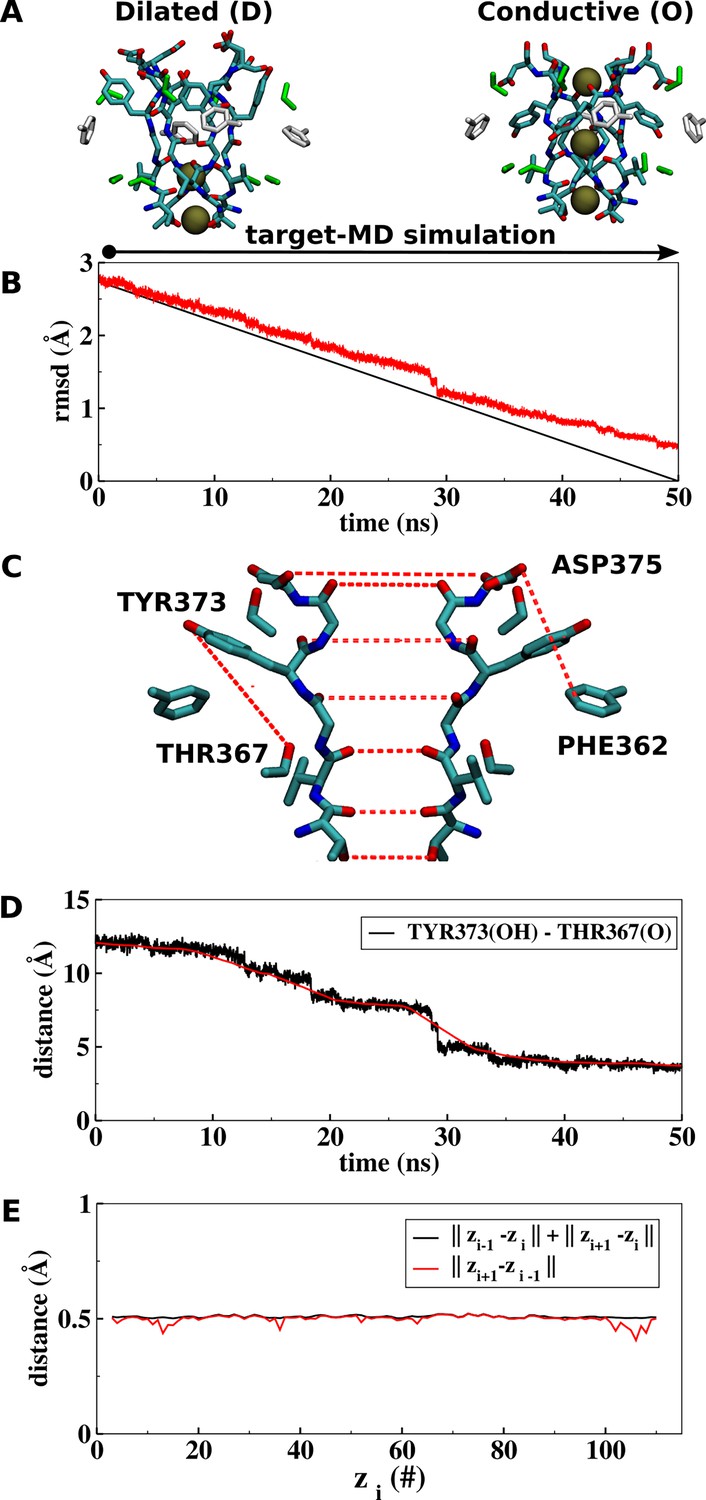
Conformational path between the conductive (O) and dilated (D) states of the selectivity filter.
(A, B) Time evolution of the instantaneous (red) and target (black) root-mean-square deviation (RMSD) of the selectivity filter between states O and D. Reference structures of the dilated and conductive states of the selectivity filter were respectively defined on basis of the high-resolution X-ray structures of the triple-mutant kv1.2-kv2.1-3m and kv1.2-kv2.1 (PDB codes 7SIT and 2R9R). Targeted molecular dynamics (TMD) was carried out for 50 ns with an applied constant force of 400 kcal/mol/Å2, corresponding to a per atom constant force of 2.38 kcal/mol/Å2. (C) A total of 22 atomic distances were used for definition of the conformational path between states D and O (red dashes): 14 main-chain carbonyl distances between two opposing subunits of the channel, 4 side-chain distances between TYR373(OH) and THR367(O), and 4 side-chain distances between ASP375(OD2) and PHE362(CD1). Atomic distances were symmetrized across the channel to support convergence of the calculation. (D) Time evolution of a representative coordinate of the transformation path. The coordinate was smoothed out following a running average procedure of the data. The running average length was 10% of the total number of data points, i.e., 500 (red). (E) The resulting string z consisted of 111 equidistant points i satisfying the monotonic condition at a distance interval of 0.25 Å.
Figure 4—figure supplement 2
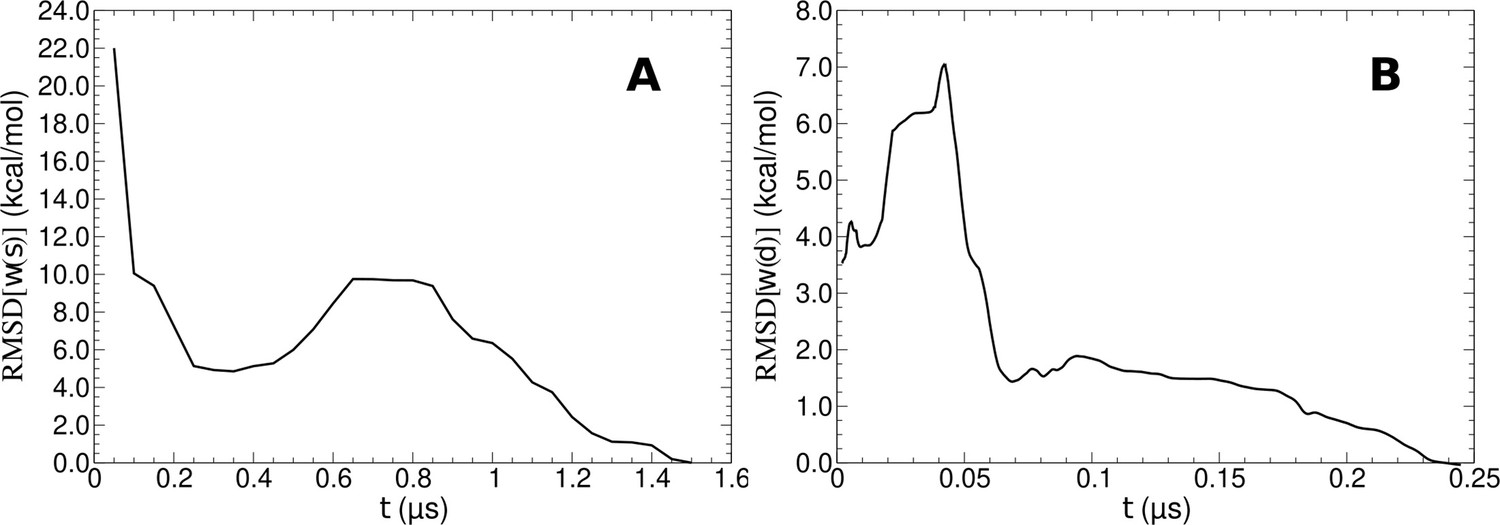
Convergence analysis of free-energy calculations.
(A, B) Respectively shown is the root-mean-square deviation (RMSD) of the free-energy profiles w(s) and w(d) as a function of simulation time t.
Figure 4—figure supplement 3
Molecular dynamics (MD) simulation of kv1.2-kv2.1-3m at +200 mV and 150 mM NaCl.
(A) Molecular representation of the main pore of the channel, highlighting the initial configuration of the selectivity filter, I398 (red) and V402 (light gray). Two sodium (Na+) ions (yellow) are shown in the selectivity filter at the ion binding sites S3 and S4. (B and C) Analysis of AMBER and CHARMM36m trajectories. Shown are the structural deviation of the selectivity filter (R), trajectory of Na+ along the permeation pathway (Z), and the intersubunit Cβ-Cβ separation distance of I398 and V402 (L) as a function of simulation time. Inset shows instantaneous configurations of I398 (red arrows). Time averages are the linear density of Na+ ions, the linear density of waters, and the pore radius profile (r) along the permeation pathway Z.
Additional files
-
Supplementary file 1
Molecular dynamics (MD) simulations of triple-mutant channel kv1.2-kv2.1-3m.
- https://cdn.elifesciences.org/articles/97696/elife-97696-supp1-v3.docx
-
Supplementary file 2
NBFixes for potassium, carbonyl, and water interactions.
- https://cdn.elifesciences.org/articles/97696/elife-97696-supp2-v3.docx
-
Supplementary file 3
Number of conduction events along simulations of the open-conductive MthK.
- https://cdn.elifesciences.org/articles/97696/elife-97696-supp3-v3.docx
-
Supplementary file 4
Number of conduction events along molecular dynamics (MD) simulations in which the isoleucine gate is open.
- https://cdn.elifesciences.org/articles/97696/elife-97696-supp4-v3.docx
-
Supplementary file 5
Binding free-energy difference of charybdotoxin (CTX).
- https://cdn.elifesciences.org/articles/97696/elife-97696-supp5-v3.docx
-
Supplementary file 6
Comparative analysis of the average properties of the isoleucine gate with and without mutation (I398N).
- https://cdn.elifesciences.org/articles/97696/elife-97696-supp6-v3.docx
-
MDAR checklist
- https://cdn.elifesciences.org/articles/97696/elife-97696-mdarchecklist1-v3.pdf
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Isoleucine gate blocks K+ conduction in C-type inactivation
eLife 13:e97696.
https://doi.org/10.7554/eLife.97696




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}