Peer review process
Not revised: This Reviewed Preprint includes the authors’ original preprint (without revision), an eLife assessment, public reviews, and a provisional response from the authors.
Read more about eLife’s peer review process.Editors
- Reviewing EditorBabak MomeniBoston College, Chestnut Hill, United States of America
- Senior EditorWendy GarrettHarvard T.H. Chan School of Public Health, Boston, United States of America
Reviewer #1 (Public review):
Summary:
The authors claim that bacteria are guided by diffusiophoresis. They perform experiments of bacterial motility in microfluidic channels with salt gradients. The data show that P. pudita bacteria swim towards higher sodium chloride concentrations, but there is no evidence that this is due to a diffusiophoresis.
Weaknesses:
It is well known that bacteria perform chemotaxis in salt gradients (see e.g., PNAS 86, pp. 8358-8362, 1989). The underlying mechanism based on chemoreceptors is widely accepted, but the authors do not mention this possibility. I recommend a control experiment where the chemotaxis genes are knocked out. Even if this mechanism can be ruled out, the current data show no evidence for a mechanism based on diffusiophoresis.
Reviewer #2 (Public review):
Summary:
The authors investigate how salt gradients influence the transport of Pseudomonas putida in confined microfluidic environments. They report that salt gradients enhance directional migration, increase run persistence, and promote transport toward contaminant-rich regions. To explain these observations, the authors propose a physical steering mechanism in which differential diffusiophoretic mobilities of the cell body and flagellar bundle generate an aligning torque that reorients cells along the salt gradient.
Strengths:
The study addresses an interesting question at the interface of microbiology, complex fluids, and active matter. Their experiments suggest that salt gradients influence bacterial transport behavior and lead to more persistent, directional motion. Once confirmed, the proposed mechanism would broaden our understanding of how environmental gradients can shape microbial migration through physical interactions in addition to more traditional sensing-based pathways.
Weaknesses:
The main limitation of the current study is that the proposed steering mechanism is not directly demonstrated. The evidence for the diffusiophoretic torque is largely inferred from trajectory statistics and theoretical modeling. While the observed transport behavior is convincing, the causal link between the observed migration patterns and the proposed reorientation mechanism remains less well established. In particular, the manuscript focuses primarily on cell trajectories and transport properties, whereas the proposed mechanism fundamentally involves changes in cell orientation. Additional evidence connecting orientation dynamics to the proposed torque mechanism would strengthen the conclusions.
A related concern is whether alternative physical mechanisms associated with the imposed salt gradients have been fully excluded. For example, weak flow-mediated effects or other hydrodynamic influences could potentially contribute to the observed transport behavior. The manuscript would benefit from a more thorough discussion of such possibilities and a clearer justification for why the proposed diffusiophoretic mechanism should be regarded as the dominant explanation.
The manuscript would also benefit from a clearer positioning within the broader literature on physically induced microbial transport and swimmer reorientation. Previous studies have demonstrated directed migration arising from rheotaxis (Marcos et al., 2012, PNAS) and viscosity-gradient-induced steering (Stehnach et al., 2021, Nature Physics). While the mechanism proposed here appears distinct, a more explicit discussion of how the present work relates to these earlier studies would help readers better understand the specific conceptual advance being made.
Author response:
Public Reviews:
Reviewer #1 (Public review):
Summary:
The authors claim that bacteria are guided by diffusiophoresis. They perform experiments of bacterial motility in microfluidic channels with salt gradients. The data show that P. putida bacteria swim towards higher sodium chloride concentrations, but there is no evidence that this is due to diffusiophoresis.
Weaknesses:
It is well known that bacteria perform chemotaxis in salt gradients (see e.g., PNAS 86, pp. 8358-8362, 1989). The underlying mechanism based on chemoreceptors is widely accepted, but the authors do not mention this possibility. I recommend a control experiment where the chemotaxis genes are knocked out. Even if this mechanism can be ruled out, the current data show no evidence for a mechanism based on diffusiophoresis.
We thank the reviewer for raising this important comment. We agree that receptor-mediated salt taxis is a well-established mechanism in some bacteria, including the classic study by Qi and Adler (PNAS, 1989). We note, however, that the original manuscript did discuss this possibility and cited Qi and Adler in line 117: “We also note that we did not observe any significant difference in the tumble rates between the control and NaCl gradient cases (Figure 3h; Figure S2, SM), suggesting that NaCl gradients do not interfere with chemoreceptors (Qi and Adler, 1989).” In Figure S2, the run-time distributions show no significant difference between the no-gradient condition and the NaCl-gradient condition, with fitted tumble rates of λ = 0.41 s−1 and λ = 0.38 s−1, respectively. We also do not observe a directional bias in run duration, namely longer runs up the salt gradient and shorter runs down the gradient, which would be expected for canonical chemoreceptor-mediated taxis.
The basis for assigning the observed migration to diffusiophoresis is that the NaCl gradient produces a directional drift of the bacterial body without a measurable change in the run-and-tumble statistics. This behavior is consistent with our previous work [1], where non-motile bacteria were shown to undergo diffusiophoretic migration toward higher salt concentration. Because that migration occurred in non-motile cells and across different bacterial types and morphologies, it supports the interpretation that native bacterial surface charge can drive a non-specific diffusiophoretic response in salt gradients.
That said, we agree with the reviewer that a genetic control would provide a stronger test against receptor-mediated chemotaxis. We will therefore perform additional experiments using a ∆cheA strain. Because CheA is required for canonical chemotactic signal transduction, observing the same directional migration in the ∆cheA mutant would directly test whether the NaCl gradient response persists in the absence of receptor-mediated chemotaxis. We will include these new data and revise the manuscript to more explicitly distinguish diffusiophoretic drift from chemoreceptormediated salt taxis.
Reviewer #2 (Public review):
Summary:
The authors investigate how salt gradients influence the transport of Pseudomonas putida in confined microfluidic environments. They report that salt gradients enhance directional migration, increase run persistence, and promote transport toward contaminant-rich regions. To explain these observations, the authors propose a physical steering mechanism in which differential diffusiophoretic mobilities of the cell body and flagellar bundle generate an aligning torque that reorients cells along the salt gradient.
Strengths:
The study addresses an interesting question at the interface of microbiology, complex fluids, and active matter. Their experiments suggest that salt gradients influence bacterial transport behavior and lead to more persistent, directional motion. Once confirmed, the proposed mechanism would broaden our understanding of how environmental gradients can shape microbial migration through physical interactions in addition to more traditional sensing-based pathways.
Weaknesses:
The main limitation of the current study is that the proposed steering mechanism is not directly demonstrated. The evidence for the diffusiophoretic torque is largely inferred from trajectory statistics and theoretical modeling. While the observed transport behavior is convincing, the causal link between the observed migration patterns and the proposed reorientation mechanism remains less well established. In particular, the manuscript focuses primarily on cell trajectories and transport properties, whereas the proposed mechanism fundamentally involves changes in cell orientation. Additional evidence connecting orientation dynamics to the proposed torque mechanism would strengthen the conclusions.
We thank the reviewer for the constructive comments. We agree that the proposed steering mechanism should be supported by evidence that directly connects the salt gradient response to bacterial orientation dynamics, not only to trajectory-level transport statistics.
We would like to clarify that the original manuscript already includes an orientation-based analysis in Figure 4f,g in the main text. The corresponding methodology and results are described in lines 173–183 and in the Supporting Information. Specifically, we quantified cell steering by measuring the change in body angle, ∆θ, along individual run trajectories as a function of arc length, s, using the orientation correlation ⟨cos(∆θ)⟩s. In the absence of salt gradients, the orientation correlation decays slowly with arc length, indicating persistent swimming along the initial
Author response image 1.
Instantaneous angular velocity as a function of heading angle relative to the salt gradient orientation. (a) Experimental and (b) simulated mean angular velocity of cells as a function of heading angle θ (measured relative to the gradient direction; θ = 0° points toward the gel/high-salt side) in the absence (blue) and presence (red) of a NaCl gradient. Positive and negative values indicate counterclockwise and clockwise rotation, respectively, with arrows showing rotation direction. Under the gradient, both experiments and simulations show a signed, angle-dependent rotation rate that is largest near θ = ±90° and approaches zero near θ = 0° and 180°, consistent with a restoring torque that steers cell heading toward the gradient direction. Simulations reproduce this behavior with comparable magnitude to the experimental measurements, and in the absence of a gradient, angular velocity remains relatively small in the no-gradient case with no consistent directional bias across heading angles.
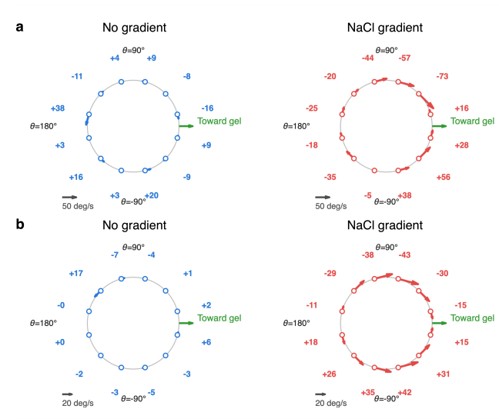
run direction. Under a salt gradient, the correlation decays more rapidly, indicating stronger directional reorientation during runs. Because the tumble statistics do not change significantly between the no-gradient and salt-gradient conditions, this enhanced orientational decorrelation is not attributed to increased tumbling or rotational noise. Instead, it is consistent with continuous deterministic steering during runs, as expected from a diffusiophoretic torque acting on the cell body–flagellar bundle system.
To further address the reviewer’s concern, we performed an additional orientation-dynamics analysis using the same dataset shown in Figure 4. Following the approach used by Stehnach et al. [2], we calculated the instantaneous angular velocity during individual runs as a function of the cell heading angle relative to the salt gradient direction. The cell orientation was obtained from the run trajectories, and tumble events were excluded because they produce large transient angular velocity spikes that are not representative of continuous steering during runs.
The new analysis is shown in Author response image 1. Under the no-gradient condition, the angular velocity remains small and nearly independent of heading angle. In contrast, under the salt gradient condition, the angular velocity becomes strongly heading-dependent. The angular velocity is largest when cells swim nearly perpendicular to the salt gradient, where a steering torque is expected to be maximal. Moreover, the sign of the angular velocity indicates rotation toward alignment with the gradient direction. This behavior is consistent with the proposed diffusiophoretic torque mechanism and provides a direct link between the observed transport behavior and salt-gradient-induced reorientation dynamics.
We plan to add this angular velocity analysis to the revised manuscript and revise the relevant text to make the connection between trajectory statistics, orientation dynamics, and the proposed torque mechanism clearer.
A related concern is whether alternative physical mechanisms associated with the imposed salt gradients have been fully excluded. For example, weak flow-mediated effects or other hydrodynamic influences could potentially contribute to the observed transport behavior. The manuscript would benefit from a more thorough discussion of such possibilities and a clearer justification for why the proposed diffusiophoretic mechanism should be regarded as the dominant explanation.
We thank the reviewer for raising this important point. We agree that alternative physical mechanisms associated with the imposed salt gradient should be considered explicitly. In the revised manuscript, we will add a more detailed discussion explaining why flow-mediated or other hydrodynamic mechanisms are unlikely to account for the observed steering behavior.
First, the characteristic diffusiophoretic drift velocity in our experiments is approximately ud ≈ 1 µm/s, corresponding to only about 1–5% of the typical swimming speed of P. putida. Thus, the proposed mechanism does not require externally driven advection of the cells. Instead, the salt gradient produces a weak but persistent differential diffusiophoretic slip on the cell body and flagellar bundle, which can generate a reorienting torque during active swimming.
We also considered whether diffusio-osmotic flow along the channel walls could generate sufficient shear to induce rheotaxis. In a dead-end channel, the diffusio-osmotic velocity profile can be estimated as [3]
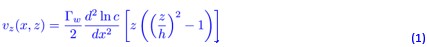
which gives a wall shear rate (at z = h) of
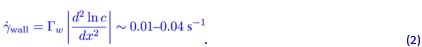
This value is below the shear rate threshold reported by Marcos et al. [2], where rheotactic drift becomes negligible for S < 0.1 s−1. Therefore, the shear generated by diffusio-osmotic wall flow in our experiments is too weak to explain the observed directional reorientation. This effect would be even smaller for P. putida, whose thin flagellar bundle is expected to experience weaker shear-induced alignment than organisms with larger flagellar structures.
We further considered viscotaxis as a possible mechanism. However, the viscosity difference between 1 mM and 100 mM NaCl solutions is marginal. This contrast is far smaller than the approximately 4–5-fold viscosity difference reported to induce strong viscophobic turning in bacteria [4]. Thus, salt-gradient-induced viscosity variations are insufficient to account for the measured steering response.
Taken together, these estimates indicate that hydrodynamic shear, rheotaxis, and viscotaxis are too weak under our experimental conditions to explain the observed migration and orientation dynamics. In contrast, the proposed nonuniform diffusiophoretic mechanism naturally accounts for the key observations: directional migration up the salt gradient, enhanced curvature during runs, heading-dependent angular velocity, and the absence of significant changes in tumble statistics. We will include this analysis in the revised manuscript to clarify why diffusiophoretic steering is the dominant mechanism under the present conditions.
The manuscript would also benefit from a clearer positioning within the broader literature on physically induced microbial transport and swimmer reorientation. Previous studies have demonstrated directed migration arising from rheotaxis (Marcos et al., 2012, PNAS) and viscosity-gradient-induced steering (Stehnach et al., 2021, Nature Physics). While the mechanism proposed here appears distinct, a more explicit discussion of how the present work relates to these earlier studies would help readers better understand the specific conceptual advance being made.
We thank the reviewer for this helpful suggestion. We agree that the manuscript should more clearly position the proposed mechanism within the broader literature on physically induced microbial transport and swimmer reorientation.
In the revised manuscript, we have added a discussion comparing our results with prior studies on rheotaxis and viscosity-gradient-induced steering. Specifically, we now discuss the work of Marcos et al. [4], which showed that shear flow can generate a torque on the helical flagellum of Bacillus subtilis, producing rheotactic alignment independent of chemical sensing. We also discuss the work of Stehnach et al. [2], which showed that viscosity gradients can steer Chlamydomonas reinhardtii through asymmetric viscous drag on its two flagella, producing viscophobic turning down the viscosity gradient.
The mechanism proposed in the present work is distinct from both of these cases. Unlike rheotaxis, it does not require externally imposed shear flow. Unlike viscophobic turning, it does not rely on a substantial viscosity contrast. Instead, we propose that a salt concentration gradient generates differential diffusiophoretic motion of the cell body and flagellar bundle, producing a torque that continuously reorients swimming cells along the gradient. This mechanism therefore identifies salt gradients as a distinct physical cue capable of steering bacteria through surface-mediated transport rather than through flow, viscosity contrast, or canonical chemoreceptor signaling.
We have added the following text to the revised manuscript: ”Our findings also relate to other physical mechanisms of microbial reorientation. Bacterial rheotaxis, arising from a torque generated by shear flow acting on the helical flagellum, steers Bacillus subtilis independent of any chemical gradient [4]. Viscosity gradients similarly drive a viscophobic turning in the alga Chlamydomonas reinhardtii, where uneven viscous drag on its two flagella produces a torque that reorients cells down the gradient [2], a behavior confirmed by measuring angular velocity as a function of heading angle and revealing a sinusoidal form, ω(θ) = −ω visc sin(θ), which resembles what we report here for diffusiophoresis in (Figure S4). This identifies salt gradients, independent of flow or viscosity, as a distinct physical route by which swimming cells can be steered and guided, broadening the set of known nonchemoreceptor mechanisms for directed microbial transport.” References
(1) V. S. Doan, P. Saingam, T. Yan, S. Shin, A trace amount of surfactants enables diffusiophoretic swimming of bacteria, ACS Nano 14 (10) (2020) 14219–14227.
(2) M. R. Stehnach, N. Waisbord, D. M. Walkama, J. S. Guasto, Viscophobic turning dictates microalgae transport in viscosity gradients, Nature Physics 17 (8) (2021) 926–930.
(3) S. Shin, E. Um, B. Sabass, J. T. Ault, M. Rahimi, P. B. Warren, H. A. Stone, Size-dependent control of colloid transport via solute gradients in dead-end channels, Proc. Natl. Acad. Sci. 113 (2) (2016) 257–261.
(4) Marcos, H. C. Fu, T. R. Powers, R. Stocker, Bacterial rheotaxis, Proceedings of the National Academy of Sciences 109 (13) (2012) 4780–4785.



