Peer review process
Not revised: This Reviewed Preprint includes the authors’ original preprint (without revision), an eLife assessment, public reviews, and a provisional response from the authors.
Read more about eLife’s peer review process.Editors
- Reviewing EditorRoel NusseStanford University, Stanford, United States of America
- Senior EditorUtpal BanerjeeUniversity of California, Los Angeles, Los Angeles, United States of America
Reviewer #1 (Public Review):
This manuscript presents a model in which combined action of the transporter-like protein DISP and the sheddases ADAM10/17 promote shedding of a mono-cholesteroylated Sonic Hedgehog (SHH) species following cleavage of palmitate from the dually lipidated precursor ligand. The authors propose that this leads to transfer of the cholesterol-modified SHH to HDL for solubilization. The minimal requirement for SHH release by this mechanism is proposed to be the covalently linked cholesterol modification because DISP could promote transfer of a cholesteroylated mCherry reporter protein to serum HDL. The authors used an in vitro system to demonstrate dependency on DISP/SCUBE2 for release of the cholesterol modified ligand. These results confirm previously published results from other groups (PMC3387659 and PMC3682496). In vivo support for these activities is provided by data from previously published studies from this group. It is unclear whether new in vivo experiments were conducted for this study.
A strength of the work is the use of a bicistronic SHH-Hhat system to consistently generate dually-lipidated ligands to determine the quantity and lipidation status of SHH released into cell culture media.
A critical shortcoming of the study is that the experiments showing SHH secretion/export by western blot of media fractions do not include a SHH(-) control condition. This is an essential control because SHH media blots can be dirty. Without demonstration that the bands being analyzed are specific for SHH(+) conditions, these experiments cannot be appropriately evaluated. Further, it appears that SHH is transiently transfected/expressed for each experimental condition. A stably expressing SHH/HHAT cell line would reduce condition to condition and experiment to experiment variability. Unusual normalization strategies are used for many experiments, and quantification/statistical analyses are missing for several experiments. Due to these shortcomings, the data do not justify the conclusions. The significance of the data provided is overstated because many of the presented experiments confirm/support previously published work. The study provides a modest advance in the understanding of the complex issue of SHH membrane extraction.
Reviewer #2 (Public Review):
Ehring et al. analyze contributions of Dispatched, Scube2, serum lipoproteins and Sonic Hedgehog lipid modifications to the generation of different Shh release forms. Hedgehog proteins are anchored in cellular membranes by N-terminal palmitate and C-terminal cholesterol modifications, yet spread through tissues and are released into the circulation. How Hedgehog proteins can be released, and in which form, remains unclear. The authors systematically dissect contributions of several previously identified factors, and present evidence that Disp, Scube2 and lipoproteins concertedly act to release a novel Shh variant that is cholesterol-modified but not palmitoylated. The systematic analysis of key factors that control Shh release is a commendable effort and helps to reconcile apparently disparate models. However, the results concerning the roles of lipoproteins and Shh lipid modifications are largely confirmatory of previous results, and molecular identity/physiological relevance of the newly identified Shh variant remain unclear.
The authors conclude that an important result of the study is the identification of HDL as a previously overlooked serum factor for secretion of lipid-linked Shh (p15, l24-25). This statement should be removed. A detailed analysis of Shh release on human lipoproteins was reported previously, including contributions of the major lipoprotein classes, in cells that endogenously express Shh, in human plasma and for Shh variants lacking palmitate and/or cholesterol modifications (PMID 23554573). The involvement of Disp is also not unexpected: the importance of Dips for release of cholesterol-modified Shh is well established, as is the essential function of Drosophila Disp for formation of lipoprotein-associated hemolymph Hh. A similar argument can be made for the sufficiency of sterol modification for lipoprotein association. The authors point out that GFP insertion at the C-terminus of the N-terminal Shh domain does not abrogate function. Perhaps more relevant, an mCherry-sterol that was generated using a similar strategy as in the present study associates with Drosophila lipoproteins (PMID 20685986).
A novel and surprising finding of the present study is the differential removal of Shh N- or C-terminal lipid anchors depending on the presence of HDL and/or Disp. In particular, the identification of a non-palmitoylated but cholesterol-modified Shh variant that associates with lipoproteins is potentially important. However, the significance of this result could be substantially improved in two ways: 1) The molecular properties of the processed Shh variants are unclear - incorporation of palmitate/cholesterol and removal of peptides were not directly demonstrated. This is particularly relevant for the N-terminus, as the signaling activity of non-palmitoylated Hedgehog proteins is controversial. A decrease in hydrophobicity is no proof for cleavage of palmitate, this could also be due to addition of a shorter acyl group. 2) All experiments rely on over-expression of Shh in a single cell line. The authors point out that co-overexpression of Hhat is important to ensure Shh palmitoylation, but the same argument could be made for any other protein that acts in Shh release, such as Disp or a plasma membrane sheddase. The authors detect Shh variants that are released independently of Disp and Scube2 in secretion assays, which however are excluded from interpretation as experimental artifacts. Thus, it would be important to demonstrate key findings in cells that secrete Shh endogenously.
The co-fractionation of Shh and ApoA1 in serum-containing media is not convincing (Fig. 4C), as the two proteins peak at different molecular weights. To support their conclusion, the authors could use an orthogonal approach, optimally a demonstration of physical interaction, or at least fractionation by a different parameter (density). On a technical note, all chromatography results are presented as stylized graphs. Please include individual data points.
Author Response
Reviewer #1 (Public Review):
1) “It is unclear whether new in vivo experiments were conducted for this study”.
All in vivo experiments shown were conducted independently by new researchers in the lab, using the original fly stocks. This will be more clearly stated in the revised supplement. The aim of repeating the experiments was to directly compare the consequences of impaired N- and C-terminal shedding side-by-side in two Hh-dependent developmental systems.
2) “A critical shortcoming of the study is that experiments showing Shh secretion/export do not include a Shh(-) control condition. Without demonstration that the bands analyzed are specific for Shh(+) conditions, these experiments cannot be appropriately evaluated”.
C9C5 antibody reactivity and specificity is shown below, and this control will be added to the revised manuscript. We established the C9C5 immunoblotting protocol – and generated the blot shown in Author Response Image 1 - before any of the experiments in the manuscript were started. The immunoblot clearly shows Shh specificity similar to that of R&D AF464 anti-Shh antibodies that were previously used in the lab. The immunoblot also shows that both antibodies detect the same Shh signals in media, that C9C5 is more sensitive, and that AF464 and C9C5 detect 5E1-IP’d dual-lipidated and monolipidated soluble Shh equally well. Also note that, in our hands, C9C5 is highly specific: this antibody detects N-truncated C25S;Δ26-35Shh of increased electrophoretic mobility, but does not cause unspecific signals above or below, even if the blot is strongly overexposed (as shown here). Specific Shh detection by C9C5 is also discussed in our response to editor’s comments below.
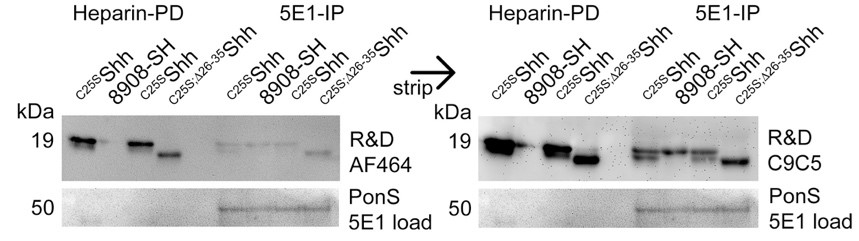
Cells were transfected with constructs encoding full-length C25SShh or truncated C25S;Δ26-35Shh, and proteins in serum-containing media were 5E1 immunoprecipitated or concentrated by heparin-sepharose pulldown. Dual-lipidated R&D 8908-SH was dissolved in the same medium and subjected to the same 5E1 immunoprecipitation or heparin pulldown. The blot was incubated with antibody AF464 and (after stripping) with antibody C9C5. Immunoblot analysis revealed high specificity of both antibodies and also revealed poor interactions of dual-lipidated 8908-SH with highly charged heparin.
3) “A stably expressing Shh/Hhat cell line would reduce condition to condition and experiment to experiment variability”.
We fully agree with this reviewer and therefore aimed to establish stable Hhat expressing cell lines several years ago. However, stable Hhat expression eliminated transfected cells after several passages, or cells gradually ceased to express Hhat, preventing us to establish a stable line despite several attempts and tried strategies. For this reason, we established transient co-expression of Shh/Hhat from the same mRNA to at least eliminate variability between relative Shh/Hhat expression levels and to assure complete Shh palmitoylation in our assays.
4) “Unusual normalization strategies are used for many experiments, and quantification/statistical analyses are missing for several experiments”.
This comment refers to data shown in Fig. 3 (here, no quantification of Scube2 function in Disp-/- cells had been conducted) and to qPCR data shown in Fig. 4 (here, Shh and C25AShh were compared only indirectly via dual-lipidated R&D 8908-SH, but not directly in a side-by-side experiment, and Shh variants with an N-terminal alanine or a serine were directly compared). We agree with the reviewer and therefore currently repeat qPCR assays and quantify blots to eliminate these technical shortcomings from the final manuscript.
5) “The study provides a modest advance in the understanding of the complex issue of Shh membrane extraction”
Our investigation identified unexpected links between Disp as a furin-activated Hh exporter, sheddase-mediated Shh release, Scube2-mediated Shh release and lipoprotein-mediated Hh transport – established modes indeed but with no previously established direct connections – that increase their relevance. We also identified a previously unknown N-processed Shh variant attached to lipoproteins and show that Disp/Scube2 function absolutely requires lipoproteins. Therefore, although we do agree that our findings are confirmatory for the above modes, they also provide new mechanistic insight and challenge the currently dominating model of Disp-mediated hand-over of dual-lipidated Hh to Scube2 chaperones (this model does not predict a role for lipoprotein particles but for both Shh lipids in signaling, for a recent discussion, see PMID 36932157). Our findings suggest an answer to the intensely debated question of whether Disp/Ptch extract cholesterol from the outer or inner plasma membrane leaflet, and suggest that N-palmitate is dispensable for signaling of lipoprotein-associated Shh to Ptch receptors. Finally, we note that previous in vivo studies in flies often relied on Hh overexpression in the fat body, raising questions on their physiological relevance. Our in vivo analyses of Hh function in wing- and eye discs are more physiologically relevant and can explain the previously reported presence of non-lipidated bioactive Hh in disc tissue (PMID: 23554573).
Reviewer #2 (Public Review):
1) “However, the results concerning the roles of lipoproteins and Shh lipid modifications are largely confirmatory of previous results, and molecular identity/physiological relevance of the newly identified Shh variant remain unclear”.
Regarding the confirmatory aspects of our work, please also refer to our response to reviewer 1. In addition, we would like to reply that our unbiased experimental approach was designed to challenge the model of Shh shedding by testing whether established Shh release regulators affect it (e.g. support it) or not. As described in our work, Disp, Scube2 and lipoproteins all contribute to increased shedding (which is new), that Disp function depends on lipoprotein presence (also new), and that lipoproteins modify the outcome of Shh shedding (dual Shh shedding versus N-shedding and lipoprotein association), which is also new.
Regarding physiological relevance, we would like to reply that our finding that artificially generated monolipidated variants (C25SShh and ShhN) solubilize in uncontrolled manner from producing cells can explain previously observed, highly variable gain-of-function or loss-of-function phenotypes upon their overexpression in vivo 1, 2, 3, 4, 5. Our data is also supported by the observed presence of variably lipidated Shh/Hh variants in vivo 6, and the in vivo observation that complete removal of Scube activity in zebrafish embryos phenocopies a complete loss of Hh function that is bypassed by increased ligand expression - and even results in wild-type-like ectopic Shh target gene expression 7. The in vivo observations are compatible with our data but are incompatible with proposed alternative models of Scube-mediated dual-lipidated Shh extraction and continued Shh/Scube association to allow for morphogen transport.
2) “Thus, it would be important to demonstrate key findings in cells that secrete Shh endogenously”.
Experimental data shown in Fig. S8B demonstrates that en-controlled expression of sheddase-resistant Hh variants blocks endogenous Hh function in the same wing disc compartment. To our knowledge, this assay is the most physiologically relevant test of the mechanism of Disp-mediated Hh release. Still, we have now started to analyze Hh from Drosophila disc tissue biochemically and hope that we can include our findings in the final manuscript.
3) “The authors could use an orthogonal approach, optimally a demonstration of physical interaction, or at least fractionation by a different parameter”.
We agree with this reviewer’s assessment and are currently in the process to establish co-IP and density gradient conditions to test physical HDL/Shh interactions. The results will be included in the final version of record.



