Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorJason LerchUniversity of Oxford, Oxford, United Kingdom
- Senior EditorMichael FrankBrown University, Providence, United States of America
Reviewer #1 (Public Review):
Valk and Engert et al. examined the potential relations between three different mental training modules, hippocampal structure and functional connectivity, and cortisol levels (stress) over a 9-month period. They found that among the three types of mental training: Presence (attention and introspective awareness), Affect (socio-emotional - compassion and prosocial motivation), and Perspective (socio-cognitive - metacognition and perspective taking) modules; Affect training most robustly related to changes in hippocampal structure and function - specifically, CA1-3 subfields of the hippocampus. Moreover, change in intrinsic functional connectivity related to changes in diurnal cortisol release and long-term cortisol exposure. These changes are proposed to result from a combination of factors, which is supported by multivariate analyses showing changes across subfields and training content relate to cortisol changes.
The authors demonstrate that mindfulness training programs are a potential avenue for stress interventions that impact hippocampal structure and cortisol, providing a promising approach to improve health. The data contribute to the literature on plasticity of hippocampal subfields during adulthood, the impact of mental training interventions on the brain, and the link between CA1-3 and both short- and long-term stress changes.
The authors thoughtfully approached the study of hippocampal subfields, utilizing a method designed for T1w images that outperformed Freesurfer 5.3 and that produced comparable results to an earlier version of ASHS. The authors note the limitations of their approaches and provide detailed information on the data used and analyses conducted. The results provide a strong basis from which future studies can expand using computational approaches or more fine-grained investigations of the impact of mindfulness training on cortisol levels and the hippocampus.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Valk and Engert et al. examined the potential relations between three different mental training modules, hippocampal structure and functional connectivity, and cortisol levels over a 9-month period. They found that among the three types of mental training: Presence (attention and introspective awareness), Affect (socio-emotional - compassion and prosocial motivation), and Perspective (socio-cognitive - metacognition and perspective taking) modules; Affect training most consistently related to changes in hippocampal structure and function - specifically, CA1-3 subfields of the hippocampus. Moreover, decreases in diurnal cortisol correlated to bilateral increases in volume, and decreases in diurnal and chronic cortisol left CA1-3 functional connectivity. Chronic cortisol levels also related to right CA4/DG volume and left subiculum function. The authors demonstrate that mindfulness training programs impact hippocampus and are a potential avenue for stress interventions, a potential avenue to improve health. The data contribute to the literature on plasticity of hippocampal subfields during adulthood, the impact of mental training interventions on the brain, and the link between CA1-3 and both short- and long-term stress changes. Additional clarification and extension of the methods is needed to strengthen the authors' conclusions.
We thank the Reviewer for their positive evaluation and summary of our findings and work. We made additional changes as suggested by the Reviewer and hope this clarified any open points.
(1) The authors thoughtfully approached the study of hippocampal subfields, utilizing a method designed for T1w images that outperformed Freesurfer 5.3 and that produced comparable results to an earlier version of ASHS. However, given the use of normalized T1-weighted images to delineate hippocampal subfield volume, some caution may be warranted (Wisse et al. 2020). While the authors note the assessment of quality control processes, the difficulty in ensuring valid measurement is an ongoing conversation in the literature. This also extends to the impact of functional co-registration using segmentations. I appreciate the inclusion of Table 5 in documenting reasons for missing data across subjects. Providing additional details on the distribution of quality ratings across subfields would help contextualize the results and ensure there is equal quality of segmentations across subfields.
We thank the Reviewer for bringing up this point. In the current work, we assessed the overall segmentation of all six subfields per individual. Thus, unfortunately, we have no data of quality of segmentation of individual subfields beyond our holistic assessment. Indeed, registration of hippocampal subfields remains a challenge and we have further highlighted this limitation in the Discussion of the current work.
“It is of note that the current work relies on a segmentation approach of hippocampal subfields including projection to MNI template space, an implicit correction for total brain volume through the use of a stereotaxic reference frame. Some caution for this method may be warranted, as complex hippocampal anatomy can in some cases lead to over- as well as underestimation of subfield volumes, as well as subfield boundaries may not always be clearly demarcated (1). Future work, studying the hippocampal surface at higher granularity, for example though unfolding the hippocampal sheet (2-5), may further help with both alignment and identification of not only subfield-specific change but also alterations as a function of the hippocampal long axis, a key dimension of hippocampal structural and functional variation that was not assessed in the current work (6, 7).”
(2) Given the consistent pattern of finding results with CA1-3, in contrast to other subfields, it would help to know if the effects of the different training modules on subfields differed from each other statistically (i.e., not just that one is significant, and one is not) to provide an additional context of the strength of results focused on Affect training and CA1-3 (for example, those shown in Figure 3).
Our work investigated i) whether the effects of the individual Training Modules differed from each other statistically. We found that the Affect Training Module showed increases in CA1-3 volume, and that these increases remained when testing effects relative to changes in this subfield following Perspective training and in retest controls. Moreover, in CA1-3 we found changes in functional connectivity when comparing the Affect to Perspective training Module. These changes were only present in this contrast, but not significant in each of the Training Modules per se. To test for specificity, we additionally evaluated whether subfield-specific changes were present above and beyond changes in the other ipsilateral hippocampal subfields. Relative to other subfields, right CA1-3 showed increases in the Affect vs Perspective contrast (left: t-value: 2.298, p=0.022, Q>0.1; right: t-value: 3.045, p=0.0025, Q=0.015). No other subfield showed significant changes. We now include this statement in the revised Results and Supplementary Tables.
“Moreover, associations between CA1-3 and Affect, relative to Perspective, seemed to go largely above and beyond changes in the other subfields (left: t-value: 2.298, p=0.022, Q>0.1; right: t-value: 3.045, p=0.0025, Q=0.015, see further Supplementary File 1h).”
Author response table 1.
Subfield-specific changes following the Training Modules, controlling for the other two ipsilateral subfields

Reviewer #1 (Recommendations For The Authors):
(1) In Figure 1, using different colors for subfields versus the modules (yellow, red, green) would help as it could lead the reader to try to draw connections between the two when it is namely a depiction of the delineations.
As suggested, we updated Figure 1 accordingly and present the subfields in different shades of purple for clarity. Please find the updated figure below.
Author response image 1.
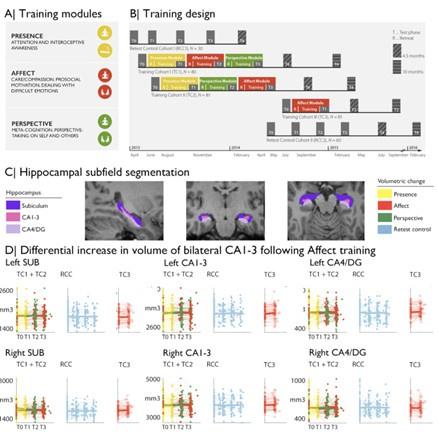
(2) In the Results, it was at times hard to follow when Affect off Perspective where the focus of the results. Perhaps the authors could restructure or add additional context for clarity.
We are happy to clarify. For the first analysis on Module-specific changes in hippocampal subfield volume, we compared effects across Training Modules. Here, main contrasts were ran between subjects: Presence vs active control and within subjects: Affect versus Perspective. In additional secondary contrasts, we studied training effects vs retest control. After observing consistent increases in bilateral CA1-3 following Affect, in the following analysis, we evaluated 1) intrinsic functional networks in main and supplementary contrasts and 2) diurnal cortisol measures within the Training modules only and all three Training Modules combined, and also adopted 3) a multivariate approach (PLS) (see comments Reviewer 2). We now also report effects of cortisol change on structural and functional subfield change in Presence and Perspective, for additional completeness and clarity.
“To study whether there was any training module-specific change in hippocampal subfield volumes following mental training, we compared training effects between all three Training Modules (Presence, Affect, and Perspective). Main contrasts were: Presence vs Active control (between subjects) and Affect vs Perspective (within subjects). Supplementary comparisons were made vs retest controls and within training groups.”
“Overall, for all hippocampal subfields, findings associated with volume increases in CA1-3 fol-lowing the Affect training were most consistent across timepoints and contrasts (Supplementary File 1a-f).”
“Subsequently, we studied whether hippocampal CA1-3 would show corresponding changes in intrinsic function following the Affect mental training.”
“In particular, the moderately consistent CA1-3 volume increases following Affect training were complemented with differential functional connectivity alterations of this subfield when comparing Affect to Perspective training”
“Last, we probed whether group-level changes in hippocampal subfield CA1-3 volume would correlate with individual-level changes in diurnal cortisol indices (Presence: n= 86; Affect: n=92; Perspective: n=81), given that the hippocampal formation is a nexus of the HPA-axis (8). We took a two-step approach. First, we studied associations between cortisol and subfield change, particularly focusing on the Affect module and CA1-3 volume based on increases in CA1-3 volume identified in our group-level analysis.”
“We observed that increases in bilateral CA1-3 following Affect showed a negative association with change in total diurnal cortisol output […]”
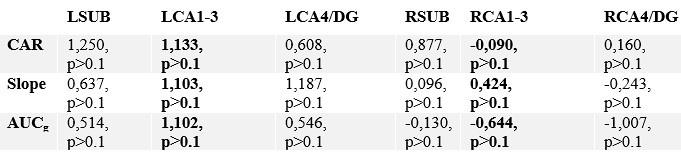
“We did not observe alterations in CA1-3 volume in relation to change in cortisol markers in Presence or Perspective. Yet, for Presence, we observed association between slope and LCA4/DG change (t=-2.89, p=0.005, q=0.03), (Supplementary File 1uv).”
“In case of intrinsic function, we also did not observe alterations in CA1-3 in relation to change in cortisol markers in Presence or Perspective, nor in other subfields (Supplementary File 1wx).”
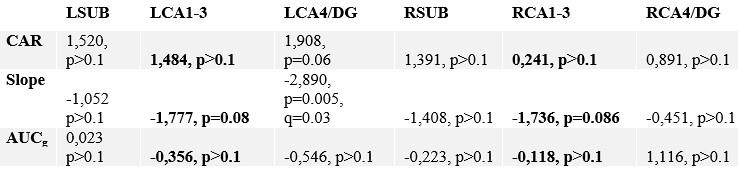
Author response table 2.
Correlating change in subfield volume and diurnal cortisol indices in Presence. Main focus was on CA1-3 based on volumetric observations and are highlighted in bold.
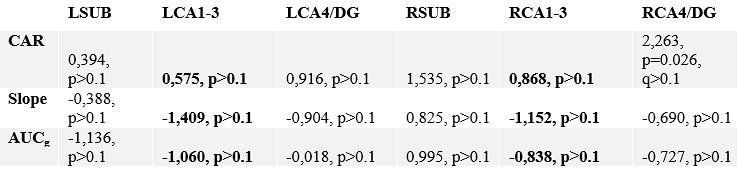
Author response table 3.
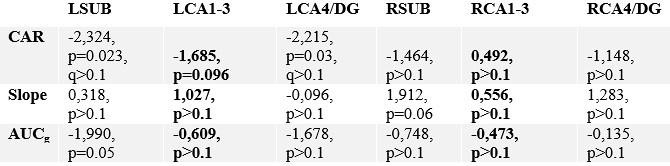
Correlating change in subfield volume and diurnal cortisol indices in Perspective. Main focus was on CA1-3 based on volumetric observations and are highlighted in bold.
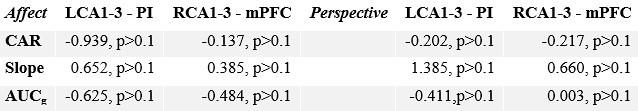
Author response table 4.
Association between stress-markers and within functional network sub-regions in Affect and Perspective.
Author response table 5.
Correlating change in subfield function and diurnal cortisol indices in Presence. Main focus was on CA1-3 based on volumetric observations and are highlighted in bold. For these multiple comparisons (FDRq, corrected for two subfields) values are reported if uncorrected p values are below p<.05.
Author response table 6.
Correlating change in subfield function and diurnal cortisol indices in Perspective. Main focus was on CA1-3 based on volumetric observations and are highlighted in bold. For these multiple comparisons (FDRq, corrected for two subfields) values are reported if uncorrected p values are below p<.05.
(3) In the Methods, the authors note that corrections for multiple comparisons were used where needed, throughout the manuscript there is some switching between corrected and uncorrected p-values. At times, this made it difficult to follow in terms of when these corrections were needed.
For clarity, we added explicit multiple comparisons information a) in main and supplementary results, and b) wherever extra information was needed. Also, we only included main contrasts in Table 1-3 to avoid confusion and moved the information on changes in SUB and CA4/DG to the Supplementary tables.
(4) Typically, when correcting for intracranial volume the purpose is the ensure that sexual dimorphism in the size of the brain is accounted for. I would recommend the authors assess whether sex differences are accounted for by the MNI normalization approach taken. In the reading of the original Methods paper for the patch-based algorithm used, ICV was used to transform to MNI152 space. It would help to have additional information on how the normalization was done in the current study in order to draw comparisons to other findings in the literature.
We are happy to further clarify. In the current work, we used the same approach as in the original paper. Volumes were linearly registered to the MNI template using FSL flirt. We now provided this additional information in the revised methods.
“Hippocampal volumes were estimated based on T1w data that were linearly registered to MNI152 using FSL flirt (http://www.fmrib.ox.ac.uk/fsl/), such that intracranial volume was implicitly controlled for.”
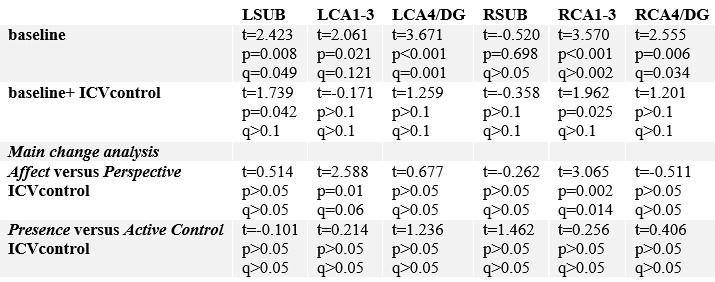
We agree with the Reviewer that sex differences may still be present, and investigated this. At baseline, sex differences were found in all subfields in the left hemisphere, and right CA4/DG (FDRq<0.05). Regressing out ICV resolved remaining sex differences. We then evaluated whether main results of volumetric subfield change were impacted by ICV differences. Differences between Affect and Perspective remained stable. We have now added this additional analysis in the Supplementary Materials.
“Although stereotaxic normalization to MNI space would in theory account for global sex differences in intra-cranial volume, we still observed sex differences in various subfield volumes at baseline. Yet, accounting for ICV did not impact our main results suggesting changes in CA1-3 following Affect were robust to sex differences in overall brain volume (Supplementary File1j).”
Author response table 7.
Sex differences (female versus male) in hippocampal subfield volumes.
Reviewer #2 (Public Review):
In this study, Valk, Engert et al. investigated effects of stress-reducing behavioral intervention on hippocampal structure and function across different conditions of mental training and in relation to diurnal and chronic cortisol levels. The authors provide convincing multimodal evidence of a link between hippocampal integrity and stress regulation, showing changes in both volume and intrinsic functional connectivity, as measured by resting-state fMRI, in hippocampal subfield CA1-3 after socio-affective training as compared to training in a socio-cognitive module. In particular, increased CA1-3 volume following socio-affective training overlapped with increased functional connectivity to medial prefrontal cortex, and reductions in cortisol. The conclusions of this paper are well supported by the data, although some aspects of the data analysis would benefit from being clarified and extended.
A main strength of the study is the rigorous design of the behavioral intervention, including test-retest cohorts, an active control group, and a previously established training paradigm, contributing to an overall high quality of included data. Similarly, systematic quality checking of hippocampal subfield segmentations contributes to a reliable foundation for structural and functional investigations.
We thank the Reviewer for the thoughtful summary and appreciation of our work, as well as requests for further clarification and analyses. We addressed each of them in a point by point fashion below.
Another strength of the study is the multimodal data, including both structural and functional markers of hippocampal integrity as well as both diurnal and chronic estimates of cortisol levels.
(1) However, the included analyses are not optimally suited for elucidating multivariate interrelationships between these measures. Instead, effects of training on structure and function, and their links to cortisol, are largely characterized separately from each other. This results in the overall interpretation of results, and conclusions, being dependent on a large number of separate associations. Adopting multivariate approaches would better target the question of whether there is cortisol-related structural and functional plasticity in the hippocampus after mental training aimed at reducing stress.
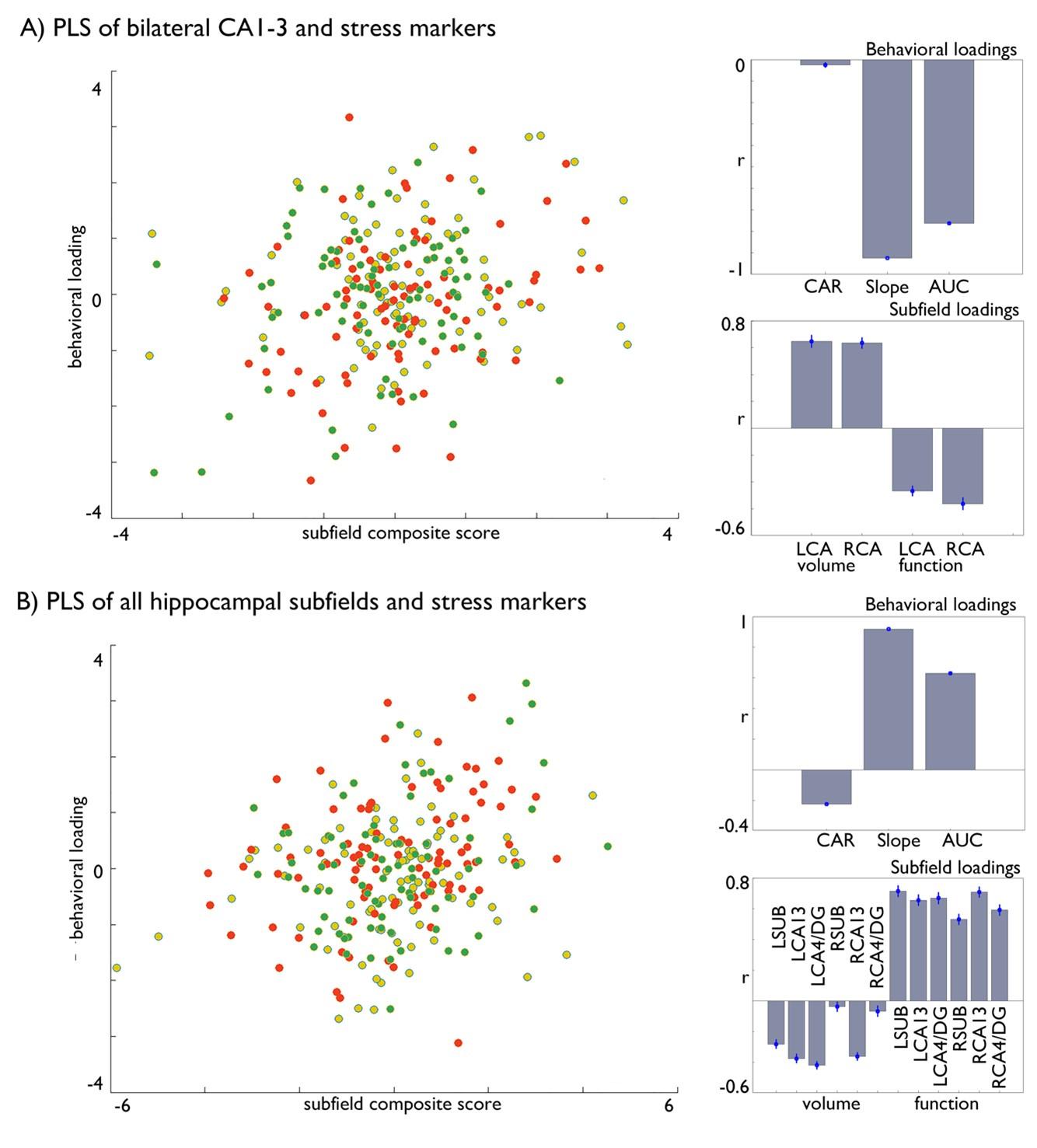
We thank the Reviewer for this suggestion. Indeed, our project combined different univariate analyses to uncover the association between hippocampal subfield structure, function, and cortisol markers. While systematic, a downside of this approach is indeed that interpretation of our results depend on a large number of analyses. To further explore the question whether there is cortisol-related structural and functional plasticity in the hippocampus, we followed the Reviewer’s suggestion and additionally adopted a multivariate partial least squares (PLS) model. We ran two complementary models. One focusing on the bilateral CA1-3, as this region showed increases in volume following Affect training and differential change between Affect and Perspective training in our resting state analyses and one model including all subfields. Both models included all stress markers. We found that both models could significantly relate stress markers to brain measures, and that in particular Affect showed strong associations with significant the latent markers. Both analyses showed inverse effects of structure and function in relation to stress markers and both slope and AUC changes showed strongest loadings. We now include these analyses the revised manuscript.
Abstract
“Of note, using a multivariate approach we found that other subfields, showing no group-level changes, also contributed to alterations in cortisol levels, suggesting circuit-level alterations within the hippocampal formation.”
Methods
“Partial least squares analysis
To assess potential relationships between cortisol change and hippocampal subfield volume and functional change, we performed a partial least squares analysis (PLS) (9, 10). PLS is a multivariate associative model that to optimizes the covariance between two matrices, by generating latent components (LCs), which are optimal linear combinations of the original matrices (9, 10). In our study, we utilized PLS to analyze the relationships between change in volume and intrinsic function of hippocampal subfields and diurnal cortisol measures. Here we included all Training Modules and regressed out effects of age, sex, and random effects of subject on the brain measures before conducting the PLS analysis. The PLS process involves data normalization within training groups, cross-covariance, and singular value decomposition. Subsequently, subfield and behavioral scores are computed, and permutation testing (1000 iterations) is conducted to evaluate the significance of each latent factor solution (FDR corrected). We report then the correlation of the individual hippocampal and cortisol markers with the latent factors. To estimate confidence intervals for these correlations, we applied a bootstrapping procedure that generated 100 samples with replacement from subjects’ RSFC and behavioral data.”
Results
“Last, to further explore the question whether there is concordant cortisol-related structural and functional plasticity in the hippocampus we adopted a multivariate partial least square approach, with 1000 permutations to account for stability (9, 10) and bootstrapping (100 times) with replacement. We ran two complementary models including all Training Modules whilst regressing out age, sex and random effects of subject. First, we focused on the bilateral CA1-3, as this region showed increases in volume following Affect training and differential change between Affect and Perspective training in our resting state analyses. In the second model included structural and functional data of all subfields. Both models included all stress markers. We found that both models could identify significant associations between cortisol stress markers and hippocampal plasticity (FDRq<0.05), and that in particular Affect showed strongest associations with the latent markers for CA1-3 (Table 5). Both analyses showed inverse effects of subfield structure and function in relation to stress markers and both slope and AUC changes showed strongest associations with the latent factor.”
Author response table 8.
Multivariate PLS analyses linking cortisol markers to hippocampal subfield volume and function.

Discussion
“Last, performing multivariate analysis, we again observed associations between CA1-3 volume and function plasticity and stress change, strongest in Affect. Yet combining all subfields in a single model indicated that other subfields also link to stress alterations, indicating that ultimately circuit-level alterations within the hippocampal formation relate to latent changes in diurnal stress markers across Training Modules.”
“This interpretation is also supported by our multivariate observations.”
“In line with our observations in univariate analysis, we found multivariate associations between hippocampal subfield volume, intrinsic function and cortisol markers. Again, the contribution of volume and intrinsic function was inverse. This may possibly relate to the averaging procedure of the functional networks. Combined, outcomes of our univariate and multivariate analyses point to an association between change in hippocampal subfields and stress markers, and that these changes, at the level of the individual, ultimately reflect complex interactions within and across hippocampal subfields and may capture different aspects of diurnal stress. Future work may more comprehensively study the plasticity of the hippocampal structure, and link this to intrinsic functional change and cortisol to gain full insights in the specificity and system-level interplay across subfields, for example using more detailed hippocampal models (3). Incorporating further multivariate, computational, models is needed to further unpack and investigate the complex and nuanced association between hippocampal structure and function, in particular in relation to subfield plasticity and short and long-term stress markers.”
“…based on univariate analysis. Our multivariate analysis further nuanced this observation, but again pointed to an overall association between hippocampal subfield changes and cortisol changes, but this time more at a systems level.”
“Lastly, our multivariate analyses also point to a circuit level understanding of latent diurnal stress scores.”
Author response image 2.
Multivariate associations between changes in structure and function of hippocampal subfield volume and markers of stress change in Affect. A) Multivariate associations between bilateral CA1-3 volume and intrinsic function and stress markers. Left: Scatter of loadings, colored by Training Module; Right upper: individual correlations of stress markers; Right lower: individual correlation of subfields; B). Multivariate associations between all subfields’ volume and intrinsic function and stress markers. Left: Scatter of loadings, colored by Training Module; Right upper: individual correlations of stress markers; Right lower: individual correlation of subfields.
(2) The authors emphasize a link between hippocampal subfield CA1-3 and stress regulation, and indeed, multiple lines of evidence converge to highlight a most consistent role of CA1-3. There are, however, some aspects of the results that limit the robustness of this conclusion. First, formal comparisons between subfields are incomplete, making it difficult to judge whether the CA1-3, to a greater degree than other subfields, display effects of training.
We thank the Reviewer for this comment. To further test for specificity, we additionally evaluated subfield-specific changes relative to other subfields for our main contrasts (Presence versus Active Control and Affect versus Perspective). Relative to other subfields, right CA1-3 showed increases in the Affect vs Perspective contrast (left: t-value: 2.298, p=0.022, Q>0.1; right: t-value: 3.045, p=0.0025, Q=0.015); no other subfield showed significant changes. We now include this statement in Results and Supplementary Tables.
“Moreover, associations between CA1-3 and Affect, relative to Perspective, seemed to go largely above and beyond changes in the other subfields (left: t-value: 2.298, p=0.022, Q>0.1; right: t-value: 3.045, p=0.0025, Q=0.015, see further Supplementary File 1h).”
Author response table 9.
Subfield-specific changes following the Training Modules, controlling for the other two ipsilateral subfields

(3) Relatedly, it would be of interest to assess whether changes in CA1-3 make a significant contribution to explaining the link between hippocampal integrity and cortisol, as compared to structure and functional connectivity of the whole hippocampus.
We thank the Reviewer for this comment. Please see the PLS analysis performed above (R2Q1). Indeed, not only CA1-3 but also other subfields seem to show a relationship with cortisol, in line with circuit level accounts on stress regulation and hippocampal circuit alterations (8, 11-15).
(4) Second, both structural and functional effects (although functional to a greater degree), were most pronounced in the specific comparison of "Affect" and "Perspective" training conditions, possibly limiting the study's ability to inform general principles of hippocampal stress-regulation.
We agree with the Reviewer that the association between stress and hippocampal plasticity, on the one hand, and mental training and hippocampal plasticity, on the other hand, make it not very straightforward to inform general principles on hippocampal stress regulation. However, as underscored in the discussion, in previous work we could also link mental training to stress reductions(16-18). We hope that the additional analyses and explanations further explain the multilevel insights of the current work, on the one hand using group-level analysis to investigate and illustrate the association between mental training and hippocampal subfield volume and intrinsic function, and on the other hand using individual level analysis to unpack the association between cortisol change and hippocampal subfield change.
Reviewer #2 (Recommendations For The Authors):
(1) In the Results, the description of how the hippocampal subfields' functional networks were defined would benefit from some clarification. It is also somewhat unclear what is meant by (on page 10): "Evaluating functional connectivity changes, we found that connectivity of the right CA1-3 functional network showed differential changes when comparing Affect training to Perspective training (2.420, p=0.016, FDRq=0.032, Cohens D =0.289), but not versus retest control (Table 1 and Supplementary Table 8-14)." Were there significant changes in CA1-3 FC following both training conditions (but these differed from each other)? A description of what this difference reflected would increase the reader's understanding.
We are happy to clarify. We included information of change of individual modules in the Supplementary materials, Supplementary Table 1 and 2, 9 and 10. Changes for functional connectivity were largely due to the differences in Modules, but did not show strong effects in one Module alone. We now include information on Affect and Perspective un-contrasted change in the main results text:
“… which could be attributed to decreases in right CA1-3 mean FC following Perspective (t=-2.012, p=0.045, M:-0.024, std: 0.081, CI [-0.041 -0.006]), but not Affect (t=1.691, p=0.092, M: 0.010, std: 0.098, CI [-0.01 0.031]); changes were not present when comparing Affect training versus retest control (Table 1 and Supplementary File 1k-q).”
(2) As described in the Public Review, the lack of multivariate assessments may risk selling the data short. Including analyses of concomitant functional and structural changes, in relation to cortisol, seems like an approach better adapted to characterize meaningful interrelationships between these measures.
We thank the Reviewer for suggesting multivariate assessments. To understand the interrelation between behavioral intervention, hippocampal plasticity, and cortisol changes, the current work first evaluates a simpler operationalization of the relationship between hippocampal subfield structure and volume, and cortisol as a function of mental training. Thus, given the complex nature of the study, we initially opted for a model where we assess structural and functional changes independently, with structural changes as the basis of our investigations. Now we have also included a multivariate approach (PLS) to further test the association between hippocampal subfields and cortisol markers, please see our additions to the manuscript above. We now highlighted multivariate associations in the Discussion as well, and suggest this as an important next step for more detailed, future investigations.
“Incorporating further multivariate, computational, models is needed to further unpack and investigate the complex and nuanced association between hippocampal structure and function, in particular in relation to subfield plasticity and short and long-term stress markers.”
(3) A minor comment regards the Figures. Some main effects should be visualized in a clearer manner. For instance, the scatterplots in Figure 1, panel D. Also, some of the current headings within the figures could be made more intuitive to the reader.
We thank the Reviewer for this comment. To improve clarity, we updated figure headings. For Figure 1D, the challenge is that the data are quite scattered and we aimed to visualize our observations in a naturalistic way. Therefore, we added additional y-axis information to further clarify the figures. Creating more overlap or differentiation would make other elements of the figure less clear, hence we remained with the current set-up detailing the intra- and inter-individual alterations of the current model.
(1) Wisse LEM, Chetelat G, Daugherty AM, de Flores R, la Joie R, Mueller SG, et al. (2021): Hippocampal subfield volumetry from structural isotropic 1 mm(3) MRI scans: A note of caution. Hum Brain Mapp. 42:539-550.
(2) DeKraker J, Kohler S, Khan AR (2021): Surface-based hippocampal subfield segmentation. Trends Neurosci. 44:856-863.
(3) DeKraker J, Haast RAM, Yousif MD, Karat B, Lau JC, Kohler S, et al. (2022): Automated hippocampal unfolding for morphometry and subfield segmentation with HippUnfold. Elife. 11.
(4) Vos de Wael R, Lariviere S, Caldairou B, Hong SJ, Margulies DS, Jefferies E, et al. (2018): Anatomical and microstructural determinants of hippocampal subfield functional connectome embedding. Proc Natl Acad Sci U S A. 115:10154-10159.
(5) Bernhardt BC, Bernasconi A, Liu M, Hong SJ, Caldairou B, Goubran M, et al. (2016): The spectrum of structural and functional imaging abnormalities in temporal lobe epilepsy. Ann Neurol. 80:142-153.
(6) Vogel JW, La Joie R, Grothe MJ, Diaz-Papkovich A, Doyle A, Vachon-Presseau E, et al. (2020): A molecular gradient along the longitudinal axis of the human hippocampus informs large-scale behavioral systems. Nat Commun. 11:960.
(7) Genon S, Bernhardt BC, La Joie R, Amunts K, Eickhoff SB (2021): The many dimensions of human hippocampal organization and (dys)function. Trends Neurosci. 44:977-989.
(8) McEwen BS (1999): Stress and hippocampal plasticity. Annu Rev Neurosci. 22:105-122.
(9) Kebets V, Holmes AJ, Orban C, Tang S, Li J, Sun N, et al. (2019): Somatosensory-Motor Dysconnectivity Spans Multiple Transdiagnostic Dimensions of Psychopathology. Biol Psychiatry. 86:779-791.
(10) McIntosh AR, Lobaugh NJ (2004): Partial least squares analysis of neuroimaging data: applications and advances. Neuroimage. 23 Suppl 1:S250-263.
(11) Paquola C, Benkarim O, DeKraker J, Lariviere S, Frassle S, Royer J, et al. (2020): Convergence of cortical types and functional motifs in the human mesiotemporal lobe. Elife. 9.
(12) DeKraker J, Ferko KM, Lau JC, Kohler S, Khan AR (2018): Unfolding the hippocampus: An intrinsic coordinate system for subfield segmentations and quantitative mapping. Neuroimage. 167:408-418.
(13) McEwen BS, Nasca C, Gray JD (2016): Stress Effects on Neuronal Structure: Hippocampus, Amygdala, and Prefrontal Cortex. Neuropsychopharmacology. 41:3-23.
(14) Sapolsky RM (2000): Glucocorticoids and hippocampal atrophy in neuropsychiatric disorders. Arch Gen Psychiatry. 57:925-935.
(15) Jacobson L, Sapolsky R (1991): The role of the hippocampus in feedback regulation of the hypothalamic-pituitary-adrenocortical axis. Endocr Rev. 12:118-134.
(16) Engert V, Hoehne K, Singer T (2023): Specific reduction in the cortisol awakening response after socio-affective mental training. Mindfulness.
(17) Puhlmann LMC, Vrticka P, Linz R, Stalder T, Kirschbaum C, Engert V, et al. (2021): Contemplative Mental Training Reduces Hair Glucocorticoid Levels in a Randomized Clinical Trial. Psychosom Med. 83:894-905.
(18) Engert V, Kok BE, Papassotiriou I, Chrousos GP, Singer T (2017): Specific reduction in cortisol stress reactivity after social but not attention-based mental training. Sci Adv. 3:e1700495.



