Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorJungmin ChoiKorea University, Seoul, Republic of Korea
- Senior EditorMurim ChoiSeoul National University, Seoul, Republic of Korea
Reviewer #1 (Public Review):
In this study, the researchers aimed to investigate the cellular landscape and cell-cell interactions in cavernous tissues under diabetic conditions, specifically focusing on erectile dysfunction (ED). They employed single-cell RNA sequencing to analyze gene expression patterns in various cell types within the cavernous tissues of diabetic individuals. The researchers identified decreased expression of genes associated with collagen or extracellular matrix organization and angiogenesis in several cell types, including fibroblasts, chondrocytes, myofibroblasts, valve-related lymphatic endothelial cells, and pericytes. They also discovered a newly identified marker, LBH, that distinguishes pericytes from smooth muscle cells in mouse and human cavernous tissues. Furthermore, the study revealed that pericytes play a role in angiogenesis, adhesion, and migration by communicating with other cell types within the corpus cavernosum. However, these interactions were found to be significantly reduced under diabetic conditions. The study also investigated the role of LBH and its interactions with other proteins (CRYAB and VIM) in maintaining pericyte function and highlighted their potential involvement in regulating neurovascular regeneration. Overall, the manuscript is well-written and the study provides novel insights into the pathogenesis of ED in patients with diabetes and identifies potential therapeutic targets for further investigation.
Comments on revised version:
For Figure 4, immunofluorecent staining of LBH following intracavernous injections with lentiviruses is required to justify overexpression and tissue specificity.
Reviewer #3 (Public Review):
Bae et al. described the key roles of pericytes in cavernous tissues in diabetic erectile dysfunction using both mouse and human single-cell transcriptomic analysis. Erectile dysfunction (ED) is caused by dysfunction of the cavernous tissue and affects a significant proportion of men aged 40-70. The most common treatment for ED is phosphodiesterase 5 inhibitors; however, these are less effective in patients with diabetic ED. Therefore, there is an unmet need for a better understanding of the cavernous microenvironment, cell-cell communications in patients with diabetic ED, and the development of new therapeutic treatments to improve the quality of life.
Pericytes are mesenchymal-derived mural cells that directly interact with capillary endothelial cells (ECs). They play a vital role in the pathogenesis of erectile function as their interactions with ECs are essential for penile erection. Loss of pericytes has been associated with diabetic retinopathy, cancer, and Alzheimer's disease and has been investigated in relation to the permeability of cavernous blood vessels and neurovascular regeneration in the authors' previous studies. This manuscript explores the mechanisms underlying the effect of diabetes on pericyte dysfunction in ED. Additionally, the cellular landscape of cavernous tissues and cell type-specific transcriptional changes were carefully examined using both mouse and human single-cell RNA sequencing in diabetic ED. The novelty of this work lies in the identification of a newly identified pericyte (PC)-specific marker, LBH, in mouse and human cavernous tissues, which distinguishes pericytes from smooth muscle cells. LBH not only serves as a cavernous pericyte marker, but its expression level is also reduced in diabetic conditions. The LBH-interacting proteins (Cryab and Vim) were further identified in mouse cavernous pericytes, indicating that these signaling interactions are critical for maintaining normal pericyte function. Overall, this study demonstrates the novel marker of pericytes and highlights the critical role of pericytes in diabetic ED.
Comments on revised version:
Bae and colleagues substantially improved the data quality and revised their manuscript "Pericytes contribute to pulmonary vascular remodeling via HIF2a signaling". While these revisions clarify some of the concerns raised, others remain. In my view, the following question must be addressed.
In my prior question on #3, I completely disagree with the statement that "identified cells with pericyte-like characteristics in the walls of large blood vessels". The staining that authors provided for LBH, was clearly stained for SMCs, not pericytes. Per Fig 2E, the authors are correct that LBH is colocalized with SMA+ cells( SMCs). However, the red signal from LBH clearly stains endothelial cells. In the rest of 2E and 2D, LBH is CD31- and their location suggests LBH stained for SMCs in the Aorta, Kidney vasculature, Dorsal vein, and Dorsal Artery.
Author Response
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
In this study, the researchers aimed to investigate the cellular landscape and cell-cell interactions in cavernous tissues under diabetic conditions, specifically focusing on erectile dysfunction (ED). They employed single-cell RNA sequencing to analyze gene expression patterns in various cell types within the cavernous tissues of diabetic individuals. The researchers identified decreased expression of genes associated with collagen or extracellular matrix organization and angiogenesis in several cell types, including fibroblasts, chondrocytes, myofibroblasts, valve-related lymphatic endothelial cells, and pericytes. They also discovered a newly identified marker, LBH, that distinguishes pericytes from smooth muscle cells in mouse and human cavernous tissues. Furthermore, the study revealed that pericytes play a role in angiogenesis, adhesion, and migration by communicating with other cell types within the corpus cavernosum. However, these interactions were found to be significantly reduced under diabetic conditions. The study also investigated the role of LBH and its interactions with other proteins (CRYAB and VIM) in maintaining pericyte function and highlighted their potential involvement in regulating neurovascular regeneration. Overall, the manuscript is well-written and the study provides novel insights into the pathogenesis of ED in patients with diabetes and identifies potential therapeutic targets for further investigation.
Reviewer #2 (Public Review):
Summary: In this manuscript, the authors performed single cell RNA-sequencing of cells from the penises of healthy and diabetes mellitus model (STZ injection-based) mice, identified Lbh as a marker of penis pericytes, and report that penis-specific overexpression of Lbh is sufficient to rescue erectile function in diabetic animals. In public human single cell RNA-sea datasets, the authors report that LBH is similarly specific to pericytes and down regulated in diabetic patients. Additionally, the authors report discovery of CRYAB and VIM1 as protein interacting partners with LBH.
The authors contributions are of interest to the erectile dysfunction community and their Lbh overexpression experiments are especially interesting and well-conducted. However, claims in the manuscript regarding the specificity of Lbh as a pericyte marker, the mechanism by which Lbh overexpression rescues erectile function, cell-cell interactions impaired by diabetes, and protein-interaction partners require qualification or further evidence to justify.
Major claims and evidence:
1) Marker gene specificity and quantification: One of the authors' major contributions is the identification of Lbh as a marker of pericytes in their data. The authors present qualitative evidence for this marker gene relationship, but it is unclear from the data presented if Lbh is truly a specific marker gene for the pericyte lineage (either based on gene expression or IF presented in Fig. 2D, E). Prior results (see Tabula Muris Consortium, 2018) suggest that Lbh is widely expressed in non-pericyte cell types, so the claims presented in the manuscript may be overly broad. Even if Lbh is not a globally specific marker, the authors' subsequent intervention experiments argue that it is still an important gene worth studying.
Answer: We appreciate this comment. In our scRNAseq data for the mouse cavernosum tissues, previously known markers such as Rgs5, Pdgfrb, Cspg4, Kcnj8, Higd1b, and Cox4i2 were found to be expressed not exclusively in pericytes, while Lbh exhibited specific expression patterns in pericytes (Fig. 2 and Supplementary Fig. 5). LBH expression was easily distinguishable from α-SMA, not only in mouse cavernosum but also in dorsal artery and dorsal vein tissues within penile tissues. This distinctive expression pattern of LBH was also observed in the human cavernous pericytes (Fig. 5). Then, we examined Lbh expression patterns in various mouse tissues using the mouse single-cell atlas (Tabula Muris), although endothelial and pericyte clusters were not subclustered in most tissues from Tabula Muris. To identify pericytes, we relied on the expression pattern of known marker genes (Pecam1 for endothelial cells, Rgs5, Pdgfrb, and Cspg4 for pericytes). Lbh was expressed in pericytes of the bladder, heart and aorta, kidney, and trachea but not as specifically in penile pericytes (Supplementary Fig. 6A-D). However, it is worth noting that other known pericyte markers were also did not exhibit exclusive expression in pericytes across all the tissues we analyzed. Therefore, in certain tissues, particularly in mouse penile tissues, Lbh may be a valuable marker in conjunction with other established pericyte marker genes for distinguishing pericytes.
2) Cell-cell communication and regulon activity changes in the diabetic penis: The authors present cell-cell communication analysis and TF regulon analysis in Fig 3 and report differential activities in healthy and DM mice. These results are certainly interesting, however, no statistical analyses are performed to justify claimed changes in the disease state and no validations are performed. It is therefore challenging to interpret these results, and the relevant claims do not seem well supported.
Answer: In response to these helpful suggestions, we calculated statistical significance and performed experimental validation. CellphoneDB permutes the cluster labels of all cells 1000 times and calculates the mean(mean(molecule 1 in cluster X), mean(molecule 2 in cluster Y)) at each time for each interaction pair, for each pairwise comparison between two cell types. We only considered interactions in which the difference in means calculated by these permutations were greater than 0.25-fold between diabetes and normal. Also, we considered that the interactions with P-value < 0.05 were significant.
To assess differential regulon activities of transcription factor (SCENIC) between diabetic and normal pericytes, we utilized a generalized linear model with scaled activity scores for each cell as input. These scaled regulon activity values for angiogenesis-related TFs exhibited differences between diabetic and normal pericytes. The results of the generalized linear model revealed that Klf5, Egr1, and Junb were TFs with significantly altered regulon activities in diabetic pericytes. Experimental data indicated that the expression level of Lmo2, Junb, Elk1, and Hoxd10 was higher (Hoxd10) or lower (Lmo2, Junb, Elk1) in diabetic pericytes compared to normal pericytes (Supplementary Fig. 9). We have added the scaled regulon activity values and statistical significance in Fig. 3E.
3) Rescue of ED by Lbh overexpression: This is a striking and very interesting result that warrants attention. By simple overexpression of the pericyte marker gene Lbh, the authors report rescue of erectile function in diabetic animals. While mechanistic details are lacking, the phenomenon appears to have a large effect size and the experiments appear sophisticated and well conducted. If anything, the authors appear to underplay the magnitude of this result.
Answer: We appreciate this comment. Therefore, we have added relevant clarification in the revised manuscript discussion section to emphasize the importance of LBH overexpression on rescuing ED as follows: “To test our hypothesis, we utilized the diabetes-induced ED mouse model, commonly employed in various studies focusing on microvascular complications associated with type 1 diabetes. We observed that the overexpression of LBH in diabetic mice led to the restoration of reduced erectile function by enhancing neurovascular regeneration. However, this study primarily demonstrated the observed phenomenon without delving into the detailed mechanisms. Nonetheless, these results of LBH on erections provide us with new strategies for treating ED and should be of considerable concern.” (Please see revised ‘Discussion’)
4) Mechanistic claims for rescue of ED by Lbh overexpression: The authors claim that cell type-specific effects on MPCs are responsible for the rescue of erectile function induced by Lbh overexpression. This causal claim is unsupported by the data, which only show that Lbh overexpression influences MPC performance. In vivo, it's likely that Lbh is being over expressed by diverse cell types, any of which could be the causal driver of ED rescue. In fact, the authors report rescue of cell type abundance in endothelial cells and neuronal cells. Therefore, it cannot be concluded that MPC effects alone or in principal are responsible for ED rescue.
Answer: We agree with these claims. Therefore, we have added relevant clarifications in the discussion section of the revised manuscript. Our findings suggest that LBH can affect the function of cavernous pericytes, although we cannot definitively specify which particular cavernous cell types are affected by the overexpressed LBH, whether it be cavernous endothelial cells, smooth muscle cells, or others. Subsequent research will be required to conduct more comprehensive mechanistic investigations, such as in vitro studies using cavernous endothelial cells, smooth muscle cells, and fibroblasts to address these knowledge gaps. (Please see revised ‘Discussion’)
5) Protein interaction data: The authors claim that CRYAB and VIM1 are novel interacting partners of LBH. However, the evidence presented (2 blots in Fig. 6A,B) lack the relevant controls. It is possible that CRYAB and VIM1 are cross-reactive with the anti-LBH antibody or were not washed out completely. The abundance of bands on the Coomassie stain in Fig. 6A suggests that either event is plausible. Therefore, the evidence presented is insufficient to support the claim that CRYAB and VIM1 are protein interacting partners of LBH.
Answer: We agree with these claims. Therefore, we have added the relevant controls(Input) and performed Co-IP (IP: CRYAB or VIM, WB: LBH) to demonstrate CRYAB and VIM1 are not simply cross-reactive antigens to their LBH antibody. Our results show that we can detect the expression of CRYAB and VIM after LBH IP, and we also detect the expression of LBH after CRYAB and VIM IP. In addition, it can be seen from our results that the binding of LBH to VIM is higher than that of CRYAB. Regardless, these results indicate that the binding of CRYAB or VIM to LBH is not a random phenomenon. (Please see revised ‘Result’ and ‘Figure 6B’)
Impact: These data will trigger interest in Lbh as a target gene within the erectile dysfunction community.
Reviewer #3 (Public Review):
Bae et al. described the key roles of pericytes in cavernous tissues in diabetic erectile dysfunction using both mouse and human single-cell transcriptomic analysis. Erectile dysfunction (ED) is caused by dysfunction of the cavernous tissue and affects a significant proportion of men aged 40-70. The most common treatment for ED is phosphodiesterase 5 inhibitors; however, these are less effective in patients with diabetic ED. Therefore, there is an unmet need for a better understanding of the cavernous microenvironment, cell-cell communications in patients with diabetic ED, and the development of new therapeutic treatments to improve the quality of life.
Pericytes are mesenchymal-derived mural cells that directly interact with capillary endothelial cells (ECs). They play a vital role in the pathogenesis of erectile function as their interactions with ECs are essential for penile erection. Loss of pericytes has been associated with diabetic retinopathy, cancer, and Alzheimer's disease and has been investigated in relation to the permeability of cavernous blood vessels and neurovascular regeneration in the authors' previous studies. This manuscript explores the mechanisms underlying the effect of diabetes on pericyte dysfunction in ED. Additionally, the cellular landscape of cavernous tissues and cell type-specific transcriptional changes were carefully examined using both mouse and human single-cell RNA sequencing in diabetic ED. The novelty of this work lies in the identification of a newly identified pericyte (PC)-specific marker, LBH, in mouse and human cavernous tissues, which distinguishes pericytes from smooth muscle cells. LBH not only serves as a cavernous pericyte marker, but its expression level is also reduced in diabetic conditions. The LBH-interacting proteins (Cryab and Vim) were further identified in mouse cavernous pericytes, indicating that these signaling interactions are critical for maintaining normal pericyte function. Overall, this study demonstrates the novel marker of pericytes and highlights the critical role of pericytes in diabetic ED.
Reviewer #1 (Recommendations For The Authors):
1) The methods are poorly written. It lacks specific information on the sample size, experimental design, and data analysis methods employed. The absence of these crucial details makes it difficult to evaluate the robustness and reliability of the findings.
Answer: We agree with the reviewer’s suggestion, now we revised the methods of our manuscript, and added detailed information or references. For sample size we have added detailed information in Figure legend (Please see revised ‘Method’ , Figure Legend, and Supplementary information.)
2) The cell number in the scRNA-seq analysis is small (~12000) and some minor cell types are probably underrepresented. It is not clear whether the authors pooled the cells from different mice as one sample, or replicates in different groups have been included. It will be helpful to label different samples in the UMAP. The authors should repeat the experiments with more replicates to increase the cell number and validate the findings.
Answer: We understand the reviewer's concern, but due to the small size of mouse penile tissue, we had to pool 5 corpus cavernosum tissues for each group (using pooled samples) for scRNA-seq analysis. Moreover, owing to the unique nature of mouse penile tissue, which is highly resistant, it posed challenges for the dissolution and isolation of single cells using conventional single-cell separation methods. Consequently, we had to increase the concentration of the enzyme to finally obtain 12,894 cells. Rather than conducting a repetitive scRNAseq analysis on the same mouse model, we validated our findings in human cavernous single-cell transcriptome data. This analysis allowed us to confirm the presence of pericyte in human corpus cavernosum, specific expression of LBH in human cavernous pericytes, and the identification of relevant GO terms associated with pericyte functions (Figure 5). We have add these information in ‘Method’ (Please see revised ‘Method’).
3) Functional studies are lacking to justify how manipulating LBH expression or its interacting proteins might lead to effective therapeutic approaches for diabetic ED.
Answer: We have performed the functional study to evaluate LBH expression might lead to effective therapeutic approaches for diabetic ED as showed in Figure 4G. Assessment of intracavernous pressure (ICP) is the most representative test for evaluating erectile function. Therefore, we modulated LBH expression in the penis of diabetic mice and assessed the erectile function of the mice by intracavernous pressure. However, we have not performed ICP studies and relative in vitro studies (migration, survival experiment) to assess whether LBH-interacting proteins have the same effect.
4) Although the abstract identifies novel targets for potential interventions, such as LBH and its interacting proteins, the clinical relevance of these findings remains uncertain. The authors should include a discussion regarding the translation of these discoveries into therapeutic strategies or their potential impact on patients with diabetes and ED.
Answer: We appreciate the reviewer's suggestion and have added a discussion as per the reviewer’s recommendation (Please see revised ‘Discussion’).
5) While the study highlights the importance of pericytes in penile erection, it fails to mention the broader context of other cell types involved in the pathogenesis of ED. Neglecting to discuss potential contributions from endothelial cells, smooth muscle cells, or neural elements limits the comprehensive understanding of the cellular interactions underlying diabetic ED.
Answer: We agree with the reviewer's suggestion and have added a discussion regarding the significance of other cell populations in penile tissues, such as endothelial cells, smooth muscle cells fibroblasts, and neural elements, along with the rationale for our focus on pericytes. (Please see revised ‘Discussion’).
Reviewer #2 (Recommendations For The Authors):
We congratulate the authors on an interesting study. We were especially excited to see their Lbh overexpression results. However, we felt other claims in the paper could benefit from additional investigation, analysis, and statistical rigor. We have provided a set of suggestions for improvement below.
Major points:
1) Pericyte marker gene proposal: See public review for commentary on the following suggested experiments. The authors should perform binary classification analysis using Lbh and report the performance of this gene as a marker (e.g. using the area under the receiver operating characteristic, accuracy, precision and recall). Further, they should consider performing this analysis for all other genes in their data to determine whether Lbh is the best marker gene.
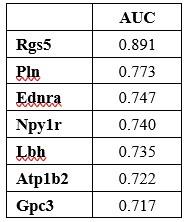
Answer: We appreciate this comment. AUC scores of Rgs5, Pln, Ednra, Npylr, Atp1b2, and Gpc3 for ability of a binary classifier to distinguish between pericyte and the other cell types in mouse penile tissues were measured by using FindMarkers function. Rgs5 had the highest AUC, but Rgs5 was also expressed in SMCs in our data. Pln, Ednra, Gpc3, and Npy1r also seemed to be candidate markers, but the literature search excluded these genes as they are also expressed in the SMCs of other tissues or different cell types. The AUC score of Lbh was over 0.7, and expression in SMC was not identified in previous studies, and ultimately, we experimentally identified that Lbh is penis pericyte specific. We have added this to the manuscript.
Author response table 1.
Robust differential expression analysis should also be performed for this gene (if not all) and the statistics should be reported, given known issues with the statistical approach used by the authors for differential expression (see: Squair 2021, 10.1038/s41467-021-25960-2). The authors' should also report the number of cells involved in these comparisons, as the number of pericytes in the data (Fig 1B) appears quite small.
Answer: We appreciate this comment. We used “MAST” to identify differentially expressed genes. This test is often used to find DEGs in single-cell RNA data. However, because the pseudobulk method has advantages over the single cell DEG method (Squair 2021, 10.1038/s41467-021-25960-2), we additionally performed DEG analysis with DESeq2 to confirm whether Lbh can distinguish pericytes from other cell types in the penile. As a result, even when tested with DESeq2, Lbh expression was significantly higher in pericytes than in other cell types in penile (adjusted p-value = 2.694475e-07 in Pericyte vs SMC, adjusted P-value = 3.700118e-58 in Pericyte vs the other cell types). Mouse penile tissue is small in size, and the number of pericytes in mouse penile tissue is relatively smaller compared to fibroblasts and chondrocytes. In our mouse penile scRNAseq data, the number of pericytes is as follows: normal: 58, diabetes: 116. Despite the limited number of cells, we were able to establish statistical significance in our analyses.
Immunostaining results in Fig. 2D, E should likewise be quantified. At present, it's unclear that LBH and aSMA are mutually exclusive as claimed. The authors should also investigate Lbh expression in public single cell genomics data, rather than performing candidate gene literature searches. For example, the Tabula Muris suggests Lbh is expressed widely outside pericytes.
Answer: For Figure 2D and E, the aim of these analyses was to assess the distribution of LBH and other cellular markers to see if they overlap and if they can be distinguished. We think that some of the overlapping staining in the tissue may be caused by multilayered cellular structures, so staining within cells would be more convincing. Therefore, we quantified the percentage of LBH- or α-SMA-expressed pericytes and relative expression in smooth muscle cells in cell staining (Supplementary Fig. 5E). We found that only 3% of smooth muscle cells expressed LBH, 67% of mouse cavernous pericytes (MCPs) expressed α-SMA, and more than 97% of MCPs expressed LBH. Therefore, these results may illustrate the specific expression of LBH in MCPs. These information was added as ‘Supplementary Fig. 5E’ (Please see revised ‘Supplementary information’). We also examined Lbh expression patterns in various mouse tissues using the public mouse single-cell atlas (Tabula Muris), and provided a detailed response in reviewer 2’s public review 1.
Even if Lbh is not the best marker, the authors' intervention experiment still motivates study of the gene, but these analyses would help contextualize the result for readers.
2) Statistical anslyses for cell-cell communication and TF regulon analysis: See public review for context on these comments. The authors should perform statistical tests to evaluate the significance of differences detected for each of these analysis. For example, generalized linear models can be used to assess the significance of TF regulon activity scores from SCENIC, and permutation tests can be used to measure the significance of cell-cell interaction score changes. Without these statistical tests, it's challenging for a reader to interpret whether the results reported are meaningful or within the realm of experimental noise.
Answer: We appreciate this comment. We calculated statistical significance TF regulon analyses as suggested by the reviewer and described a detailed statistical calculation method for cell-cell communication. We provided a detailed response in reviewer 2’s public review 2.
3) Mechanism of ED rescue by Lbh overexpression: To support this claim, the authors would need to perform an experiment where Lbh is over expressed specifically in MPCs (using e.g. a specific promoter on their LTV construct, or a transgenic line with a cell type-specific Cre-Lox system). Absent these data, the claim should be removed.
Answer: We agree with the reviewer's suggestion and we have reworked the claim that ‘LBH overexpression is affected by pericytes during ED recovery’ and have added relevant clarification in the Discussion section to clearly state that LBH overexpression may affect many cavernosum cells, such as cavernous endothelial cells, smooth muscle cells, fibroblasts, and pericytes (Please see revised ‘Result’ and ‘Discussion’)
4) Protein interaction claims: This experiment would require that the authors perform a similar pull-down with LBH KO cells and or a reciprocal Co-IP (e.g. IP: CRYAB or VIM1, WB: LBH) to demonstrate CRYAB and VIM1 are not simply cross-reactive antigens to their LBH antibody. Further, these experiments appear to only have a single replicate for each condition. The authors should either remove associated claims, or perform a Co-IP experiment with the relevant controls with sufficient replication.
Answer: We agree with the claims. Therefore, we have included the necessary controls (Input) and performed Co-IP (IP: CRYAB or VIM1, WB: LBH) to demonstrate that CRYAB and VIM1 are not simply cross-reactive antigens to their LBH antibody. Our results show that we can detect the expression of CRYAB and VIM after LBH IP, and we also detect the expression of LBH after CRYAB and VIM IP. In addition, it can be seen from our results that the binding of LBH to VIM is higher than that of CRYAB. Regardless, these results indicate that the binding of CRYAB or VIM to LBH is not a random phenomenon. Additionally, all IP experiments were replicated at least three times. (Please see revised ‘Result’ and ‘Figure 6B’)
Minor Points:
- The reference "especially in men" on line 56 seems odd given that only males can experience penile erectile dysfunction.
Answer: We agree with the reviewer's suggestion and have removed the description 'especially male' (Please see revised ‘Introduction’)
- Line 109, it's unclear what genes showed altered expression in Schwann cells.
Answer: We apologize for the confusion. There was no significant differentially expressed genes between normal and diabetes in Schwann cells. We revised this part in the manuscript. (Schwann cells showed an increased expression compared to normal cells in diabetes, though not significant. In Schwann cells, there were no significant DEGs between diabetic and normal cells.)
- It would be helpful for readers to see an analysis of the cell types that are transduced in the Lbh overexpression experiment in vivo. At present, some pericyte specificity is implied, but not demonstrated.
Answer: We appreciate this comment. Our findings suggest that LBH can affect the function of cavernous pericytes, although we cannot definitively conclude which specific-cavernous cell types are affected by the overexpressed LBH, whether it be cavernous endothelial cells, smooth muscle cells, or others. Subsequent research will be required to conduct more comprehensive mechanistic investigations, such as in vitro studies using cavernous endothelial cells, smooth muscle cells, and fibroblasts to address these knowledge gaps. These were also mentioned in the manuscript.
- To improve clarity and enhance readability, define abbreviations before their initial usage in the text. For instance, in the second paragraph of the Introduction, the abbreviation 'ECs' is used without prior definition. It can be inferred that it is referring to endothelial cells, mentioned in parentheses in the subsequent sentence.
Answer: We agree with the reviewer's suggestion to expand acronyms and ensure that all acronyms are defined in the revised manuscript before they are used for the first time in the text (Please see revised Manuscript).
- It is important to include relevant references that align with the content being discussed. For example, in the Introduction, pericytes are described as being involved in various processes such as angiogenesis, vasoconstriction, and permeability. The text refers to a single reverence, a review by Gerhardt and Besholtz, which primarily focuses on pericyte's role in regulating angiogenesis. Adding additional sources, such as the review by Bergers and Song (Neuro Oncol., 2005) is recommended.
Answer: We agree with the reviewer's suggestion, and have added the reference as reviewer recommended (Please see revised Manuscript and reference).
- Figure 3E: it is stated that a panel of 53 angiogenesis factors were tested, it is stated that only MMP3 showed increased expression. However, various unlabeled spots appear to show changed expression patterns. It would be helpful to show a summary graph with the relative intensities of the full array of factors tested.
Answer: We agree with the reviewer’s suggestion, now we showed all spots density in angiogenesis array as Supplementary Table 1. The condition of the spots we selected was that the expression density was at least above 1500, and the change ratio was greater than 1.2. (Please see revised ‘Supplementary information’)
Reviewer #3 (Recommendations For The Authors):
Detailed statistical power calculation
Data availability statement( were both mouse and human scRNA deposited in GEO with a taken and when will they be released to the public?)
Answer: Human scRNA data have been deposited in GEO under accession number GSE206528. Our mouse scRNA dataset has been uploaded to KoNA and is available for download (https://www.kobic.re.kr/kona/review?encrypt_url=amlod2FucGFya3xLQUQyMzAxMDEz)
Major concerns about this work
1) The single cell RNAseq data collected for mouse diabetic ED(Fig 1B), FB are the most abundant cell population compared to PC, EC, SMC and other clusters. The rationale for studying FB clusters (in Figure 1, D-F) instead of PC cluster is unclear. Which cluster DEG did the authors annotate for Fig 1G-H?
Answer: We understand the reviewer's suggestion and confusion. Although other major cell populations in penile tissue such as smooth muscle cells, endothelial cell, and fibroblasts have been extensively studied, pericytes have mainly been investigated in the context of the central nervous system (CNS). For example, in the CNS, pericytes are involved in maintaining the integrity of the brain's blood-brain barrier (BBB) [PMID: 27916653], regulating blood flow at capillary junctions [PMID: 33051294], and promoting neuroinflammatory processes [PMID: 31316352], whose dysfunction is considered an important factor in the progression of vascular diseases such as Alzheimer's disease [PMID: 24946075]. But little is known about the role of pericytes in penile tissue [PMID: 35865945; PMID: 36009395; PMID: 26044953]. In order to explore the role of pericytes in repairing the corpus cavernosum vascular and neural tissues damaged by DM, we focused on pericytes, which are multipotent perivascular cells that contribute to the generation and repair of various tissues in response to injury. Although recent studies have shown that pericytes are involved in physiological mechanisms of erection, little is known about their detailed mechanisms. We have also added this rationale in discussion.
Single cell level study has not been conducted in mouse penile tissues. Therefore, before delving into pericytes, we aimed to identify overall transcriptome differences between normal and diabetic conditions in mouse penile tissues. We presented the analyses of FB, which make up the largest proportion among the cell types in the mouse penis, in Fig. 1D-F. The analysis of other cell types is provided in Supplementary Fig. 1-4. Fig. 1G-H are GO terms for Fibroblasts clusters. We added this information in the figure.
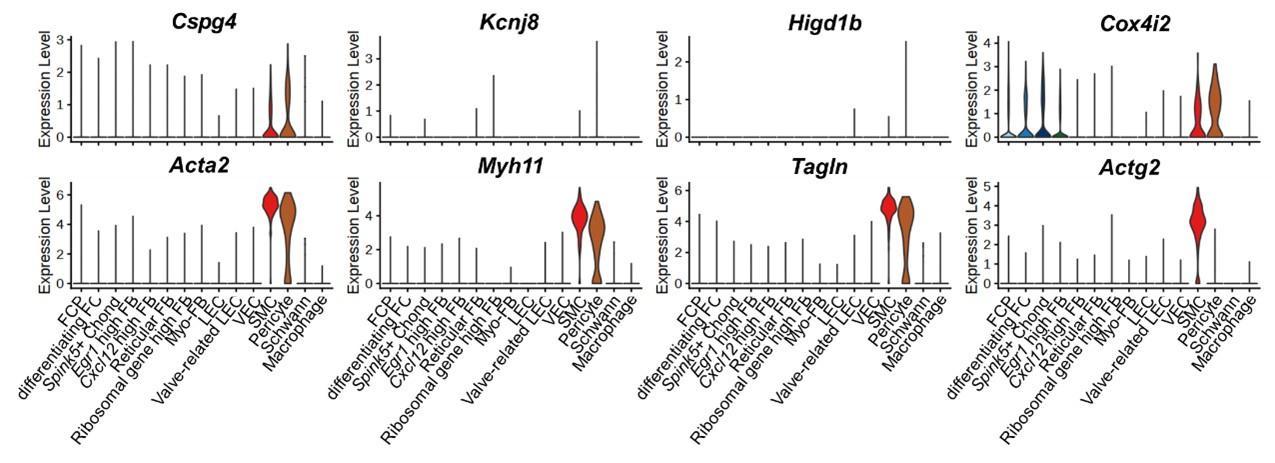
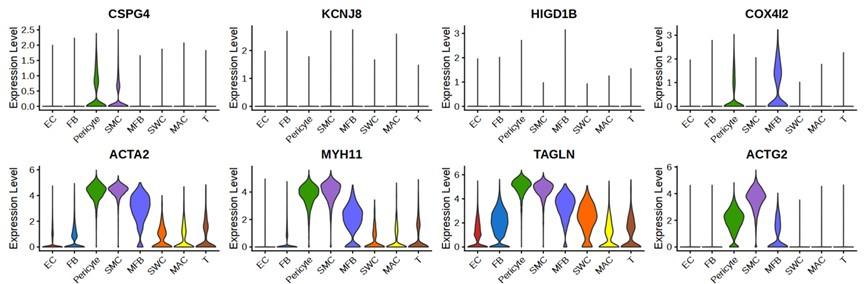
2) Fig 2 is the critical data to show Lbh is a cavernous PC specific marker. More PC violin plots to identify PC cluster such as Cspg4, Kcnj8, Higd1b, Cox4i2 and more SMC violin plots to identify SMC cluster such as Acta2, Myh11, Tagln, Actg2 should be used for inclusion and exclusion of PC( the same concern applied to human scRNAseq in Fig 5B).
Answer: We appreciate this comment. We examined the expression of other marker genes of pericytes and SMCs. Although some marker genes were rarely expressed in the mouse penis data (Kcnj8, Higd1b), the expression of marker genes tended to be relatively high in each cluster. The expression of Cspg4 and Cox4i2 was higher in pericytes than in SMCs, while the expression of Acta2, Myh11,and Tagln was higher in SMCs than in pericytes. Actag2 was specifically expressed in SMCs. Through the gene set enrichment test as well as the expression of known cell type marker genes, we identified that the annotation of pericyte and SMC was appropriate (Fig. 2B and Fig. 5C). We added the violin plots of these marker genes in Supplementary Fig. 5.
Author response image 1.
(Mouse)
In human penis data, ACTA2 and MYH11 were expressed in SMCs, pericytes, and myofibroblasts, as in the previous paper [PMID: 35879305]. Among pericyte markers, the number of cells expressing KCNJ8 and HIGD1B was small. The cluster we annotated as pericyte was double positive for pericyte markers CSPG4 and COX4I2. ACTG2, a marker for SMC, was expressed more highly in SMC than in pericytes and myofibroblasts. As in the mouse penis data, we identified that the annotation of each cell type was appropriate through the gene set enrichment test in the human penis data. We added the violin plots of CSPG4, COX4I2, and ACTG2 in Supplementary Fig. 11.
Author response image 2.
(Human)
When exploring Lbh expression levels in "Database of gene expression in adult mouse brain and lung vascular and perivascular cells" from https://betsholtzlab.org/VascularSingleCells/database.html, Lbh is not uniquely expressed in PC, suggesting its tissue-specific expression level. This difference should be discussed in the Discussion section.
Answer: We appreciate this valuable comment. For the answer to this comment, we extensively analyzed Lbh expression patterns in various mouse tissues using the public mouse single-cell atlas (Tabula Muris) as also suggested by Reviewer 2. Please see our detailed response in reviewer 2’s public review 1.
3) In prior studies on PC morphology and location (PMID: 21839917), they reside in capillaries (diameter less than 10um) or distal vessels (diameter less than 25um) and have oval cell body and long processes. Due to the non-specificity of Pdgfrb, SMC are positive for Pdgfrb staining (this has been shown in many publications that SMC are Pdgfrb+; unfortunately, NG2 antibody also stains for both PC and SMC). Therefore, the LBH immunostaining (in Fig 2D and 2E of large-sized vessels) are very likely for SMC identity, not PC. PC should be in close contact with CD31+ ECs in healthy conditions. The LBH immunostaining of PC in both mouse and human tissues (Fig 4) must be replaced and better characterized.
Answer: We agree with the reviewer's suggestion. As it is widely known, peicytes are primarily located in capillaries, where they surround endothelial cells of blood vessels. However, recent discoveries have identified cells with pericyte-like characteristics in the walls of large blood vessels, challenging the traditional concept [PMID: 27268036]. In our study, we observed minimal overlap in staining between LBH and α-SMA, suggesting that the cells expressing LBH were not smooth muscle cells but possibly pericyte-like cells in large vessels. In small vessels within the bladder, kidney, and even the aorta, we found LBH-expressing cells surrounding CD31-expressing vessels, consistent with the known characteristics of pericytes. Further research is needed to comprehend the differences in LBH expression and its characteristics in both large and small blood vessels. We have added discussions and references for this issue (Please see revised ‘Discussion’ and ‘Reference’)
4) How do mouse cavernous pericytes isolate? How is purity?
Answer: As the reviewer points out, we isolated mouse spongiform pericytes following our and other previously published methods. We used pigment epithelium-derived factor (PEDF), which removes non-pericytic cells [PMID: 30929324, 23493068]. Although there are no purity study results such as FACS, other staining results thoroughly support the notion that this method yields pericytes with a notably high level of purity. (Please see ‘Method’ section).
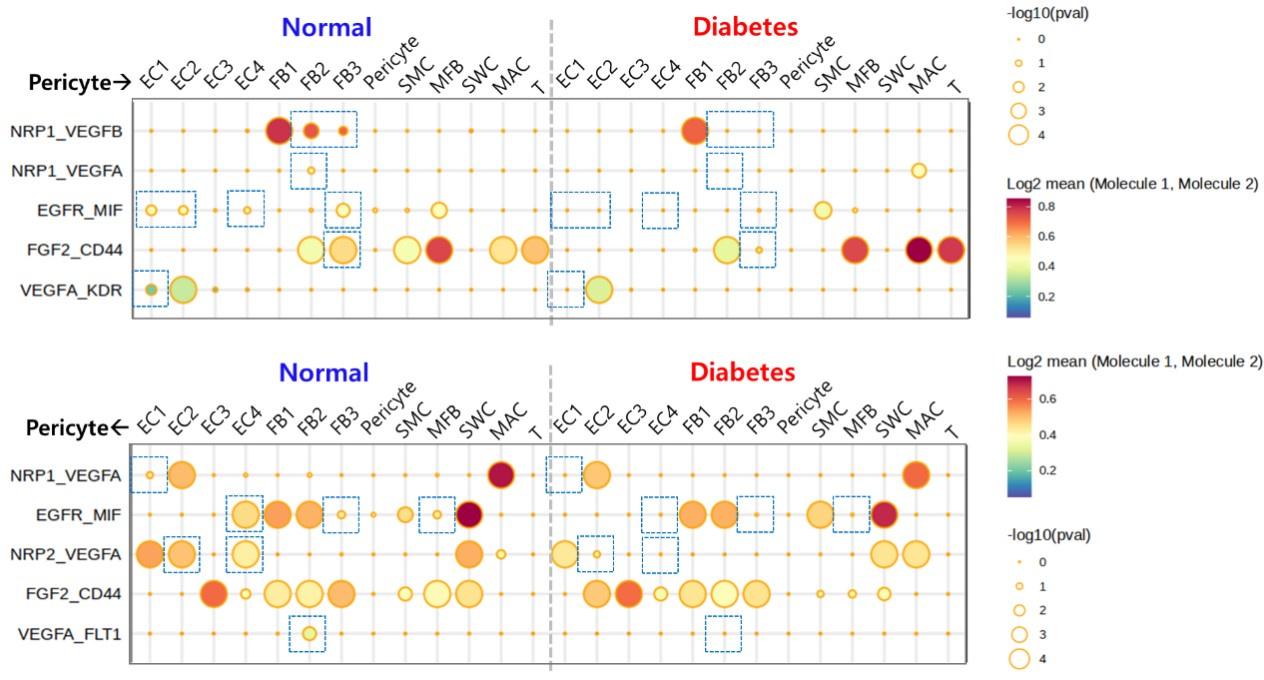
5) Can mouse scRNAseq cell-cell communication in Fig 3 be reproducible in human scRNAseq cell-cell communication? The results in human ED are more clinically significant than in mouse data.
Answer: In human scRNAseq data, the difference between angiogenesis-related interactions between normal and diabetes was not as significant as that in mouse data. Because the cell type composition of the human and mouse penis is not completely identical, there are limitations in comparing cell-cell interactions. However, in the human penis data, some interactions related to angiogenesis between pericytes and other cell types were decreased in diabetes compared to normal (boxed parts).
Author response image 3.
6) Fibroblasts also express Vim. Murine PC VIM/CRYAB( should be written as Vim/Cryab as mouse proteins) direct interaction with Lbh is unclear from Lbh IP as Fig 6A red boxes showed a wide range of sizes. Where is the band for Lbh? Do human PC LBH interact with VIM/CRYAB?
Answer: We agree with the reviewer's comment. VIM is a type III intermediate filament protein expressed in many cell types. We have added the relevant controls (Input) and performed Co-IP (IP: CRYAB or VIM, WB: LBH) to demonstrate CRYAB and VIM are not simply cross-reactive antigens to their LBH antibody. In western blot study, the LBH band was expressed between 35 kDa-48 kDa. From Figure 6A, we detected CRYAB in band 1 and VIM in bands 2 and 3. This may be due to the formation of dimers or multimers by VIM. We did not use human PCs for IP studies because IP requires large amounts of protein, making IP studies using human pericyte challenging. Nevertheless, the interaction between LBH and CRYAB in humans has been reported through fluorescent resonance energy transfer assay and affinity chromatography technology assay [PMID:34000384, PMID:20587334].
7) In Fig 6H and I, why does CRYAB expression significantly reduce in vitro and in vivo under diabetic conditions, whereas VIM expression significantly increases?
Answer: As the reviewer pointed out, and we have discussed on this issue in the manuscript, CRYAB is known to promote angiogenesis. Diabetes reduces CRYAB expression, so angiogenesis may be impaired. Furthermore, since VIM is a multifunctional protein, it interacts with several other proteins with multiple functions under various pathophysiological conditions. There are many relevant literatures showing that VIM expression is increased under diabetic conditions [PMID: 28348116 and PMID: 32557212]. And VIM deficiency protects against obesity and insulin resistance in patients with type 2 diabetes. Therefore, we hypothesize that exogenous LBH may have the ability to bind to the increased VIM in diabetic conditions and inactivate the effects of VIM. Thereby achieving the protective effect. This needs to be proved in further studies.
8) The therapeutic strategies targeting (Lbh-Cryab-Vim) on mouse diabetic ED model is not investigated and need to be further validated and discussed.
Answer: As the reviewers pointed out, in this study, we did not evaluate the targeted therapeutic strategy for LBH-CRYAB-VIM in a mouse diabetic ED model. We only identified the binding potential of these three proteins. Evaluation of this treatment strategy requires further study. For example, we can employ shRNA lentivirus, either alone or in combination, to downregulate CRYABexpression [PMID: 31612679] in normal mice, utilize a lentiviral vector CMV-GFP-puro-vimentin to overexpress Vimentin [PMID: 36912679], and then treat it with LBH to evaluate whether the LBH effect still exists (in vivo erectile function study and in vitro angiogenesis assay). We include this information in the Discussion section as a limitation of this study (Please see revised ‘Discussion’).
9) The Discussion of current knowledge of pericytes in diabetic ED and other diseases and the significance of this study as well as clinical implications, should be expanded.
Answer: As the reviewers pointed out, we have expanded the current knowledge of pericytes in diabetic ED and other diseases (CNS disease) and clinical implications as follows: “Although other major cell populations in penile tissue such as smooth muscle cells, endothelial cell, and fibroblasts have been extensively studied, pericytes have mainly been investigated in the context of the central nervous system (CNS). For example, in the CNS, pericytes are involved in maintaining the integrity of the brain's blood-brain barrier (BBB), regulating blood flow at capillary junctions, and promoting neuroinflammatory processes, whose dysfunction is considered an important factor in the progression of vascular diseases such as Alzheimer's disease. But little is known about the role of pericytes in penile tissue.” (Please see revised ‘Discussion’).
10) How many clinical samples were used? How many times did each experiment repeat?
Answer: As the reviewers pointed out, the clinical samples’ information was added in ‘method’ section. A total four human samples were used in this study (‘human corpus cavernosum tissues were obtained from two patients with congenital penile curvature (59-year-old and 47-year-old) who had normal erectile function during reconstructive penile surgery and two patients with diabetic ED (69-year-old and 56-year-old) during penile prosthesis implantation.’). For in vivo study, we quantified four different fields from human samples.
Minor concerns
1) Fig 1A, why normal mouse's body size is the same as DM?
Answer: As the reviewer pointed out, in Figure 1A, while the size of normal mice and DM mice may not appear significantly different, there are indeed notable difference in body weight and size. The normal mice body weigh we used was about 30 grams, while DM mice body weigh was generally less than 24 grams. We found that we missed information on physiological and metabolic parameters from in vivo studies (ICP function study). Therefore, we have added it in Supplementary Table 2 (Please see revised ‘Supplementary information’)
2) The label and negative, and positive controls for Fig 6B are missing.
Answer: We thank for pointing out this. We have added the relevant controls (Input) and performed Co-IP (IP: CRYAB or VIM1, WB: LBH) to demonstrate CRYAB and VIM1 are not simply cross-reactive antigens to their LBH antibody and all IP was replicated for at least 3 times. (Please see revised ‘Result’ and ‘Figure 6B’)
3) The limitation of this study and future work should be discussed.
Answer: As the reviewer pointed out, we have added the limitation of this study and future direction in the discussion section (Please see revised ‘Discussion’).



