Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorTobias DonnerUniversity Medical Center Hamburg-Eppendorf, Hamburg, Germany
- Senior EditorJoshua GoldUniversity of Pennsylvania, Philadelphia, United States of America
Reviewer #1 (Public review):
Summary:
This paper proposes a neural mechanism underlying the perception of ambiguous images: neuromodulation changes the gain of neural circuits promoting a switch between two possible percepts. Converging evidence for this is provided by indirect measurements of neuromodulatory activity and large-scale brain dynamics which are linked by a neural network model. However, both the data analysis as well as the computational modeling are incomplete and would benefit from a more rigorous approach.
This is a revised version of the manuscript which, in my view, is a considerable step forward compared to the original submission.
In particular, the authors now model phasic gain changes in the RNN, based on the network's uncertainty. This is original and much closer to what is suggested by the phasic pupil responses. They also show that switching is actually a network effect because switching times depend on network configuration (Fig 2). This resolves my main comments 1 and 2 about the model.
The mechanism, as I understand it, is different from what the authors described before in the RNN with tonic gain changes. As uncertainty increases, the network enters a regime in which the two excitatory populations start to oscillate. My intuition is that this oscillation arises from the feedback loop created by the new gain control mechanism. If my intuition is correct, I think it would be worth to explain this mechanism in the paper more explicitly.
Overall, the modeling part of the paper has changed quite a lot and I think it is now more solid which is why I have updated my "strength of evidence" rating.
Reviewer #2 (Public review):
This paper tests the hypothesis that perceptual switches during the presentation of ambiguous stimuli are accompanied by changes in neuromodulation that alter neural gain and trigger abrupt changes in brain activity. To test this hypothesis, the study combines pupillometry, artificial recurrent network (RNN) analysis and fMRI recording. In particular, the study uses methods of energy landscape analysis inspired by physics, which is particularly interesting.
Strengths
- The authors should be commended for combining different methods (pupillometry, RNNs, fMRI) to test their hypothesis. This combination provides a mechanistic insight into perceptual switches in the brain and artificial neural networks.
- The study combines different viewpoints and fields of scientific literature, including neuroscience, psychology, physics, and dynamical systems. In order to make this combination more accessible to the reader, the different aspects are presented in a pedagogical way to be accessible to all fields.
- This combination of methods and viewpoints is rarely done, so it is very useful.
- The authors introduce dynamic gain modulation in their recurrent neural network, which is novel. They devote a section of the paper to studying the dynamics, fixed points and convergence of this type of network.
Weaknesses
- The study may not be specific to perceptual switches. This is because the study relies on a paradigm in which participants report when they identify a switch in the item category. Therefore, it is unclear whether the effects reported in the paper are related to the perceptual switch itself, to attention, or to the detection of behaviourally relevant events. The authors are cautious and explicitly acknowledge this point in their study.
- The demonstration of the causal role of gain modulation in perceptual switches is partial. This causality is clearly demonstrated in the simulation work with the RNN. However, it is not fully demonstrated in the pupil analysis and the fMRI analysis. One reason is that this work is correlative (which is already very informative). An analysis of the timing of the effect might have overcome this limitation. For example, in a previous study, the same group showed that fMRI activity in the LC region precedes changes in the energy landscape of fMRI dynamics, which is a step towards investigating causal links between gain modulation, changes in the energy landscape and perceptual switches.
- Some effects may reflect the expectation of a perceptual switch rather than the perceptual switch itself. To mitigate this risk, the design of the fMRI task included catch trials, in which no switch occurs, to reduce the expectation of a switch. The pupil study, however, did not include such catch trials.
- The paper uses RNN-based modelling to provide mechanistic insight into the role of gain modulation in perceptual switches. However, the RNN solves a task that differs markedly from that performed by human participants, which may limit the explanatory value of the model. The RNN is provided with two inputs characterising the sensory evidence supporting the first and last image category in the sequence (e.g. plane and shark). In contrast, observers in the task were naïve as to the identity of the last image at the beginning of the sequence. The brain first receives sensory evidence about the image category (e.g. plane) with which the sequence begins, which is very easy to recognise, then it sees a sequence of morphed images and has to discover what the final image category will be. To discover the final image category, the brain has to search a vast space of possible second images (it is a shark?, a frog?, a bird?, etc.), rather than comparing the likelihood of just two categories. This search process and the perceptual switch in the task appear to be mechanistically different from the competition between two inputs in the RNN.
- Another aspect of the motivation for the RNN model remains unclear. The authors introduce dynamic gain modulation in the RNN, but it is not clear what the added value of dynamic gain modulation is. Both static (Fig. S1) and dynamic (Fig. 2F) gain modulation lead to the predicted effect: faster switching when the gain is larger.
- The authors are to be commended for addressing their research questions with multiple tools and approaches. There are links between the different parts of the study. The RNN and the pupil are linked by the notion of gain modulation, the RNN and the fMRI analysis are linked by the study of the energy landscape, the fMRI study and the pupil study are indirectly linked by previous work for this group showing that the peak in LC fMRI activity precedes a flattening of the energy landscape. These links are very interesting but could have been stronger and more complete.
Author response:
The following is the authors’ response to the original reviews.
Public Reviews:
Reviewer #1 (Public Review):
Summary:
This paper investigates the neural mechanisms underlying the change in perception when viewing ambiguous figures. Each possible percept is related to an attractor-like brain state and a perceptual switch corresponds to a transition between these states. The hypothesis is that these switches are promoted by bursts of noradrenaline that change the gain of neural circuits. The authors present several lines of evidence consistent with this view: pupil diameter changes during the time point of the perceptual change; a gain change in neural network models promotes a state transition; and large-scale fMRI dynamics in a different experiment suggests a lower barrier between brain states at the change point. However, some assumptions of the computational model seem not well justified and the theoretical analysis is incomplete. The paper would also benefit from a more in-depth analysis of the experimental data.
Strengths:
The main strength of the paper is that it attempts to combine experimental measurements - from psychophysics, pupil measurements, and fMRI dynamics - and computational modeling to provide an emerging picture of how a perceptual switch emerges. This integrative approach is highly useful because the model has the potential to make the underlying mechanisms explicit and to make concrete predictions.
Weaknesses:
A general weakness is that the link between the three parts of the paper is not very strong. Pupil and fMRI measurements come from different experiments and additional analysis showing that the two experiments are comparable should be included. Crucially, the assumptions underlying the RNN modeling are unclear and the conclusions drawn from the simulation may depend on those assumptions.
With this comment in mind we have made substantial effort to better integrate the three different aspects of our paper. On the pupillometry side, we now show that the dynamic uncertainty associated with perceptual categorisation shares a similar waveform with the observed fluctuations in pupil diameter around the switch point (Fig 2B). To better link the modelling to the behaviour we have also made the gain of the activation function of each sigmoidal unit change dynamically as a function of the uncertainty (i.e. the entropy) of the network’s classification generating phasic changes in gain that mimic the observed phasic changes in pupil dilation explicitly linking the dynamics of gain in the RNN to the observed dynamics of pupil diameter (our non-invasive proxy for neuromodulatory tone). Finally we note that the predictions of the RNN (flattened egocentric landscape and peaks in low-dimensional brain state velocity at the time point of the perceptual switch) were tested directly in the whole-brain BOLD data, which links the modelling and BOLD analysis. Finally we note that whilst we agree that an experiment in which pupilometry and BOLD data were collected simultaneously would be ideal, these data were not available to us at the time of this study.
Main points:
Perceptual tasks in pupil and fMRI experiments: how comparable are these two tasks? It seems that the timing is very different, with long stimulus presentations and breaks in the fMRI task and a rapid sequence in the pupil task. Detailed information about the task timing in the pupil task is missing. What evidence is there that the same mechanisms underlie perceptual switches at these different timescales? Quantification of the distributions of switching times/switching points in both tasks is missing. Do the subjects in the fMRI task show the same overall behavior as in the pupil task? More information is needed to clarify these points.
We recognize the need for a more detailed and comparative analysis of the perceptual tasks used in our pupil and fMRI experiments, particularly regarding differences in timing, task structure, and instructions. The fMRI task incorporates jittered inter-trial intervals (ITIs) of 2, 4, 6, and 8 seconds, designed to enable effective deconvolution of the BOLD response (Stottinger et al., 2018). In contrast, the pupil task presents a more rapid sequence of stimuli without ITIs. These timing differences are reflected in the mean perceptual switch points: the 8th image in the fMRI task and the 9th image in the pupil task. This small yet consistent difference suggests subtle influences of task design on behavior.
Despite these structural and instructional differences, our analyses indicate that overall behavioral patterns remain consistent across the two modalities. The distributions of switching times align closely, and no significant behavioral deviations were observed that might suggest a fundamental difference in the underlying mechanisms driving perceptual switches. These findings suggest that the additional time and structural differences in the fMRI task do not significantly alter the behavioral outcomes compared to the pupil task.
To address these issues, we have added paragraphs in the Results, Methods, and Limitations sections of the manuscript. In the Results section, we provide a detailed comparison of switching point distributions across the two tasks, emphasizing behavioral consistencies and any observed variations. In the Methods section, we include an expanded description of task timing, instructions, and the presence or absence of catch trials to ensure clarity regarding the experimental setups. Finally, in the Limitations section, we acknowledge the structural differences between the tasks, particularly the lack of catch trials and rapid stimulus presentation in the pupil task, and discuss how these differences may influence perceptual dynamics.
These additions aim to clarify how task-specific factors, such as timing, instructions, and catch trials, influence perceptual dynamics while highlighting the consistency in behavioral outcomes across both experimental setups. We believe these revisions address the concerns raised and enhance the manuscript’s transparency and rigor.
Computational model:
(1) Modeling noradrenaline effects in the RNN: The pupil data suggests phasic bursts of NA would promote perceptual switches. But as I understand, in the RNN neuromodulation is modeled as different levels of gain throughout the trial. Making the neural gain time-dependent would allow investigation of whether a phasic gain change can explain the experimentally observed distribution of switching times.
We thank the reviewer for this very helpful suggestion. We updated the RNN so that, post-training, gain changes dynamically as a function of the network's classification uncertainty (i.e. the entropy of the network's output). Specifically, the gain dynamics of each unit in the neural network are governed by a linear ODE with a forcing function given by the entropy of the network’s classification (i.e. the uncertainty of the classification). This explicitly tests the hypothesis that uncertainty driven increases in gain near the perceptual switch (when the input is maximally ambiguous) speeds perceptual switches, and allows us to distinguish between tonic and phasic increases in gain (in the absence of uncertainty forcing gain decays exponentially to a tonic value of 1). Importantly, in line with our hypothesis, we found that switch times decreased as we increased the impact of uncertainty on gain (i.e. switch times decreased as the magnitude of uncertainty forcing increased). Finally, we wish to note that although making gain dynamical is relatively simple conceptually, actually implementing it and then analysing the dynamics turned out to be highly non-trivial. To our knowledge our model is the first RNN of reasonable size to implement dynamical gain requiring us to push the RNN modelling beyond the current state of the art (see Fig 2 - 4).
(2) Modeling perceptual switches: in the results, it is described that the networks were trained to output a categorical response, but the firing rates in Fig 2B do not seem categorical but rather seem to follow the input stimulus. The output signals of the network are not shown. If I understand correctly, a trivial network that would just represent the two input signals without any internal computation and relay them to the output would do the task correctly (because "the network's choice at each time point was the maximum of the two-dimensional output", p. 22). This seems like cheating: the very operation that the model should perform is to signal the change, in a categorical manner, not to represent the gradually changing input signals.
The output of the network was indeed trained to be categorical via a cross entropy loss function with the output defined by the max of the projection of the excitatory hidden units onto the output weights which is boilerplate RNN modelling practice. As requested we now show the output in Fig 2B. On the broader question of whether a trivially small network could solve the task we are in total agreement that with the right set of hand-crafted weights a two neuron sigmoidal network with winner-take-all readout could solve the task. We disagree, however, that using an RNN is cheating in any way. Many tasks in neuroscience can be trivially solved with a very small number of recurrent units (e.g. basically all 2AF tasks). The question we were interested in is how the brain might solve the task, and more specifically how neuromodulator control of gain changes the dynamics of our admittedly very simple task. We could have done this by hand crafting a small network to solve the task but we wanted to use the RNN modelling as a means of both hypothesis testing and hypothesis generation. We now expand on and justify this modelling choice in the second paragraph of the discussion:
“We chose to use an RNN, instead of a simpler (more transparent) model as we wanted to use the RNN as a means of both hypothesis generation and hypothesis testing. Specifically, unlike more standard neuronal models which are handcrafted to reproduce a specific effect, when building an RNN the modeller only specifies the network inputs, labels, and the parameter constraints (e.g. Dale’s law) in advance. The dynamics of the RNN are entirely determined by optimisation. Post-training manipulations of the RNN are not built in, or in any way guaranteed to work, making them more analogous to experimental manipulations of an approximately task-optimal brain-like system. Confirmatory results are arguably, therefore, a first steps towards an in vitro experimental test.”
(3) The mechanism of how increased gain leads to faster switches remains unclear to me. My first intuition was that increasing the gain of excitatory populations (the situation shown in Fig. 2E) in discrete attractor models would lead to deeper attractor wells and this would make it more difficult to switch. That is, a higher gain should lead to slower decisions in this case. However, here the switching time remains constant for a gain between 1 and 1.5. Lowering the gain, on the other hand, leads to slower switching. It is, of course, possible that the RNN behaves differently than classical point attractor models or that my intuition is incorrect (though I believe it is consistent with previous literature, e.g. Niyogi & Wong-Lin 2013 (doi:10.1371/journal.pcbi.1003099) who show higher firing rates - more stable attractors - for increased excitatory gain).
We thank the reviewer for the astute observation, which we entirely agree with. The energy landscape analysis is a method still under active development within our group and we are still learning how to best explain it and its relationship to more traditional ways of quantifying potential-like energy functions of dynamical systems which we think the reviewer has in mind. We have now included a second type of energy landscape analysis which gives a complementary perspective on the RNN dynamics and is more straightforwardly comparable to typical potential functions. We describe the new analysis in the section “Large-scale neural predictions of recurrent neural network model” as follows:
“Crucially, there are two complementary viewpoints from which we can construct an energy landscape; the first allocentric (i.e., third-person view) perspective quantifies the energy associated with each position in state space, whereas the second egocentric (i.e., first person view) perspective quantifies the energy associated relative changes independent of the direction of movement or the location in state space. The allocentric perspective is straightforwardly comparable to the potential function of a dynamical system but can only be applied to low dimensional data in settings where a position-like quantity is meaningfully defined. The egocentric perspective is analogous to taking the point of view of a single particle in a physical setting and quantifying the energy associated with movement relative to the particles initial location. An egocentric framework is thus more applicable, when signal magnitude is relative rather than absolute. See materials and methods, and (see Fig S4 for an intuitive explanation of the allocentric and egocentric energy landscape analysis on a toy dynamical system).”
From the allocentric perspective it is entirely true that increasing gain increases the depth of the landscape, equivalent to increasing the depth of the attractor. However, because the input to the network changes dynamically the location of the approximate fixed-point attractor changes and the network state “chases” this attractor over the course of the trial. Importantly, the location of the energy minima changes more rapidly as gain increases, effectively forcing the network to rapidly change course at the point of the perceptual switch (see Fig 4). To quantify this effect we constructed a new measure - neural work - which describes the amount of “force” exerted on the low-dimensional neural trajectory by the vector field quantified by the allocentric landscape. Specifically we treat the allocentric landscape as analogous to a potential function and then leverage the fact that force is equal to the negative gradient of potential energy to calculate the work (force x displacement) done on the low dimensional trajectory at each time point. This showed that as gain increases the amount of work done on the neuronal trajectory at turning points increases analogous to the application of an external force transiently increasing the kinetic energy of an object. From the perspective of the egocentric landscape this results in a flattening of the landscape as there is a lower energy (i.e. higher probability) assigned to large deviations in the neuronal trajectory around the perceptual switch.
Because of the novelty of the analyses we went to great lengths to carefully explain the methods in the updated manuscript. In addition we wrote a short tutorial style MATLAB script implementing both the allocentric and egocentric landscape analysis on a toy dynamical system with a known potential function (a supercritical pitchfork bifurcation).
(4) From the RNN model it is not clear how changes in excitatory and inhibitory gain lead to slower/faster switching. In order to better understand the role of inhibitory and excitatory gain on switching, I would suggest studying a simple discrete attractor model (a rate model, for example as in Wong and Wang 2006 or Roxin and Ledberg, Plos Comp. Bio 2008) which will allow to study these effects in terms of a very few model parameters. The Roxin paper also shows how to map rate models onto simplified one-dimensional systems such as the one in Fig S3. Setting up the model using this framework would allow for making much stronger, principled statements about how gain changes affect the energy landscape, and under which conditions increased inhibitory gain leads to faster switching.
One possibility is that increasing the excitatory gain in the RNN leads to saturated firing rates. If this is the reason for the different effects of excitatory and inhibitory gain changes, it should be properly explained. Moreover, the biological relevance of this effect should be discussed (assuming that saturation is indeed the explanation).
We thank the reviewer for this excellent suggestion. After some consideration we decided that studying a reduced model would likely not do justice to the dynamical mechanisms of RNN especially after making gain dynamical rather than stationary. Still we very much share the reviewer’s concern that we need a stronger link between the (now dynamical) gain alterations and energy landscape dynamics. To this end we now describe and interrogate the dynamics of the RNN at a circuit level through selectivity and lesion based analyses, at a population level through analysis of the dynamical regime traversed by the network, and finally, through an extended energy landscape framework which has far stronger links to traditional potential based descriptions of low-dimensional dynamical systems (also see to comment 3. above).
At a circuit level the speeding of perceptual switches is mediated by inhibition of the initially dominant population we describe in paragraphs 7 and 8 of the section “Computational evidence for neuromodulatory-mediated perceptual switches in a recurrent neural network” as follows:
“Having confirmed our hypothesis that increasing gain as a function of the network uncertainty increased the speed of perceptual switches, we next sought to understand the mechanisms governing this effect starting with the circuit level and working our way up to the population level (c.f. Sheringtonian and Hopfieldian modes of analysis(66)). Because of the constraint that the input and output weights are strictly positive, we could use their (normalised) value as a measure of stimulus selectivity. Inspection of the firing rates sorted by input weights revealed that the networks had learned to complete the task by segregating both excitatory and inhibitory units into two stimulus-selective clusters (Fig 2C). As the inhibitory units could not contribute to the networks read out, we hypothesised that they likely played an indirect role in perceptual switching by inhibiting the population of excitatory neurons selective for the currently dominant stimulus allowing the competing population to take over and a perceptual switch to occur.
To test this hypothesis, we sorted the inhibitory units by the selectivity of the excitatory units they inhibit (i.e. by the normalised value of the readout weights). Inspecting the histogram of this selectivity metric revealed a bimodal distribution with peaks at each extreme strongly inhibiting a stimulus selective excitatory population at the exclusion of the other (Fig S2). Based on the fact that leading up to the perceptual switch point both the input and firing rate of the dominant population are higher than the competing population, we hypothesized that gain likely speeds perceptual switches by actively inhibiting the currently dominant population rather than exciting/disinhibiting the competing population. We predicted, therefore, that lesioning the inhibitory units selective for the stimulus that is initially dominant would dramatically slow perceptual switches, whilst lesioning the inhibitory units selective for the stimulus the input is morphing into would have a comparatively minor slowing effect on switch times since the population is not receiving sufficient input to take over until approximately half way through the trial irrespective of the inhibition it receives. As selectivity is not entirely one-to-one, we expect both lesions to slow perceptual switches but differ in magnitude. In line with our prediction, lesioning the inhibitory units strongly selective for the initially dominant population greatly slowed perceptual switches (Fig 3F upper), whereas lesioning the population selective for the stimulus the input morphs into removed the speeding effect of gain but had a comparatively small slowing effect on perceptual switches (Fig 3F lower).”
At the population level we characterised the dynamics of the 2D parameter space (defined by gain and the difference between the input dimensions) traversed by the network over the course of a trial as input and gain dynamically change. We describe this paragraphs 9-14 of the section “Computational evidence for neuromodulatory-mediated perceptual switches in a recurrent neural network” which we reprint below for the reviewers convenience :
“Based on the selectivity of the network firing rates we hypothesised that the dynamics were shaped by a fixed-point attractor whose location and existence were determined by gain and and thus changed dynamically over the course of a single trial(67-70). Because of the large size of the network, we could not solve for the fixed points or study their stability analytically. Instead we opted for a numerical approach and characterised the dynamical regime (i.e. the location and existence of approximate fixed-point attractors) across all combinations of gain and visited by the network. Specifically, for each combination of elements in the parameter space we ran 100 simulations with initial conditions (firing rates) drawn from a uniform distribution between [0,1], and let the dynamics run for 10 seconds of simulation time (10 times the length of the task - longer simulation times did not qualitatively change the results) without noise. As we were interested in the existence of fixed-point attractors rather than their precise location, at each time point we computed the difference in firing rate between successive time points across the network. For each simulation we computed both the proportion of trials that converged to a value below 10^-2 giving us proxy for the presence of fixed points, and the time to convergence, giving us a measure of the “strength” of the attractor.
Across gain values when input had unambiguous values, the network rapidly converged across all initialisations (Fig 3A & 3C-H). When input became ambiguous, however, the dynamics acquired a decaying oscillation and did not converge within the time frame of the simulation. As gain increased, the range of values characterised by oscillatory dynamics broadened. Crucially, for sufficiently high values of gain, ambiguous values transitioned the network into a regime characterised by high amplitude inhibition-driven oscillations (Fig 3D & 3G). Each trial can, therefore, be characterised by a trajectory through this 2-dimensional parameter space, with dynamics shaped by the dynamical regimes of each location visited (Fig 3A-B).
When uncertainty has a small impact on gain the network has a trajectory through an initial regime characterised by the rapid convergence to a fixed point where the population representing the initial stimulus dominated whilst the other was silent (Fig 3C), an uncertain regime characterised by oscillations with all neurons partially activated (Fig 3D), and after passing through the oscillatory regime, the network once again enters a new fixed-point regime where the population representing the initial stimulus is now silent and the other is dominant (Fig 3E).
For high gain trails, the network again started and finished in states characterised by a rapid convergence to a fixed point representing the dominant input dimension (Fig 3F-H), but differed in how it transitioned between these states. Uncertain inputs now generated high amplitude oscillations with the network flip-flopping between active and silent states (Fig 3G). We hypothesised that, within the task, this has the effect of silencing the initially dominant population, and boosting the competing population. To test this we initialised each network with parameter values well inside the oscillatory regime (u = [ .5, .5] , gain = 1.5) with initial conditions determined by the selectivity of each unit. Excitatory units selective for input dimension 1, as well as the associated inhibitory units projecting to this population, were fully activated, whilst the excitatory units selective for input dimension 2 and the associated inhibitory units were silenced. As we predicted, when initialised in this state the network dynamics displayed an out of phase oscillation where the initially dominant population was rapidly silenced and the competing population was boosted after a brief delay (219 (ms), +/-114 Fig S3).”
From this we concluded that at a population level, heightened gain leading up to the perceptual switch speeds the switch by transiently pushing the dynamics into an unstable dynamical regime replacing the fixed-point attractor representing the input with an oscillatory regime that actively inhibits the currently dominant population and boosts the competing population before transitioning back into a regime with a stable (approximate) fixed-point attractor representing the new stimulus (Fig 3F-H & Fig S3).
As we describe in the our response to comment 3 above our extended energy-landscape analysis framework now includes an explicit link between the potential of the dynamical system and allocentric landscape, whilst also explaining how a transient deepening of the allocentric landscape (which can be essentially thought of analogous to a traditional potential function) relates to the flattening of the egocentric landscape.
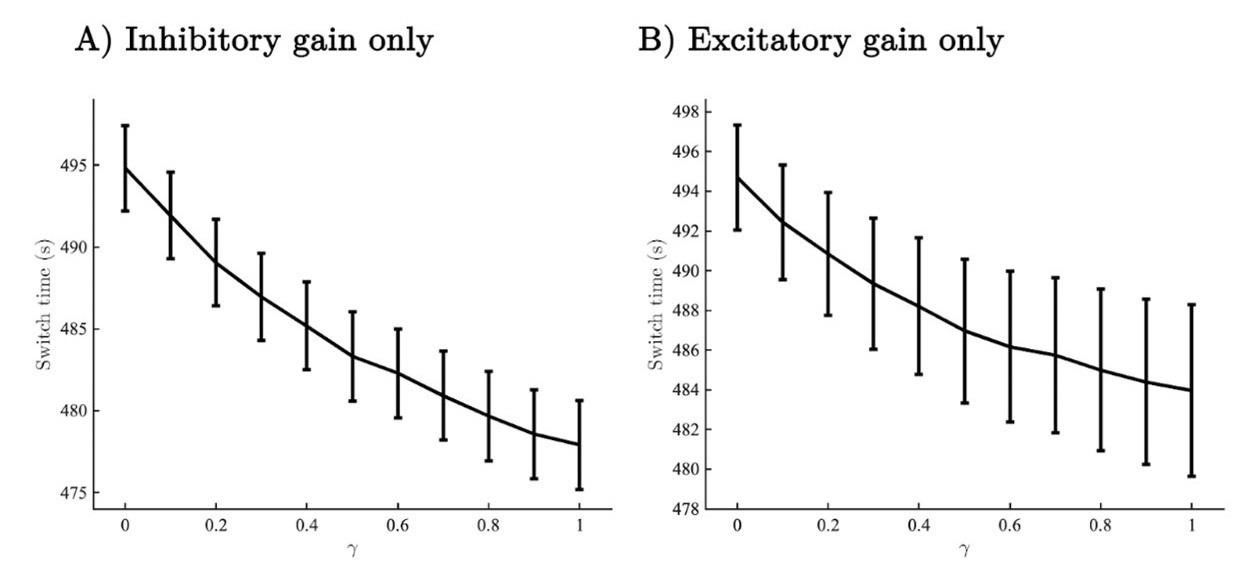
Finally, whilst we appreciate the interest in further characterising the effect of inhibitory gain compared with excitatory gain the topic is is largely orthogonal the aims of our paper so we have removed the discussion of inhibitory vs excitatory gain. Still, we understand that we need to do our due diligence and check that our results do not break down when we manipulate either inhibitory or excitatory gain in isolation. To this end we checked that dynamical gain still speeded perceptual switches when the effect was isolated to inhibitory or excitatory cells in isolation. We show the behavioural plots below for the reviewer’s interest.
Author response image 1.
Switch time as a function of uncertainty forcing
Alternative mechanisms:
It is mentioned in the introduction that changes in attention could drive perceptual switches. A priori, attention signals originating in the frontal cortex may be plausible mechanisms for perceptual switches, as an alternative to LC-controlled gain modulation. Does the observed fMRI dynamics allow us to distinguish these two hypotheses? In any case, I would suggest including alternative scenarios that may be compatible with the observed findings in the discussion.
We agree with the reviewer, in that attention is itself a confound and a process that is challenging to disentangle from the perceptual switching process in the current task. Importantly, we were not arguing for exclusivity in our manuscript, but merely testing the veracity of the hypothesis that the ascending arousal system may play a causal role in mediating and/or speeding perceptual switches. Future work with experiments that more specifically aim to dissociate these different features will be required to tease apart these different possibilities.
Reviewer #2 (Public Review):
Strengths
- the study combines different methods (pupillometry, RNNs, fMRI).
- the study combines different viewpoints and fields of the scientific literature, including neuroscience, psychology, physics, dynamical systems.
- This combination of methods and viewpoints is rarely done, it is thus very useful.
- Overall well-written.
Weaknesses
- The study relies on a report paradigm: participants report when they identify a switch in the item category. The sequence corresponds to the drawing of an object being gradually morphed into another object. Perceptual switches are therefore behaviorally relevant, and it is not clear whether the effect reported correspond to the perceptual switch per se, or the detection of an event that should change behavior (participant press a button indicating the perceived category, and thus switch buttons when they identify a perceptual change). The text mentions that motor actions are controlled for, but this fact only indicates that a motor action is performed on each trial (not only on the switch trial); there is still a motor change confounded with the switch. As a result, it is not clear whether the effect reported in pupil size, brain dynamics, and brain states is related to a perceptual change, or a decision process (to report this change).
We agree with the reviewer that the coupling of the motor change with the perceptual switch is confounded to some degree, but since motor preparation occurs on every trial we suspect that it is more accurate to describe it as confounded with task-relevance more than motor preparation per se. While it is possible that pupil diameter, network topology and energy landscape features are all related to motor change rather than the perceptual switch, we note that the weight of evidence is against this interpretation, given the simple mechanistic explanation created by the coupling of perceptual uncertainty to network gain.
- The study presents events that co-occur (perceptual switch, change in pupil size, energy landscape of brain dynamics) but we cannot identify the causes and consequences. Yet, the paper makes several claims about causality (e.g. in the abstract "neuromodulatory tone ... causally mediates perceptual switches", in the results "the system flattening the energy landscape ... facilitated an updating of the content of perception").
We have made an effort to soften the causal language, where appropriate. In addition, we note that we have changed the title to “Gain neuromodulation mediates task-relevant perceptual switches: evidence from pupillometry, fMRI, and RNN Modelling” to reflect the fact that our claims do not extent to cases of perceptual switches where the stimulus is only passively observed.
- Some effects may reflect the expectation of a perceptual switch, rather than the perceptual switch per se. Given the structure of the task, participants know that there will be a perceptual switch occurring once during a sequence of morphed drawings. This change is expected to occur roughly in the middle of the sequence, making early switches more surprising, and later switches less surprising. Differences in pupil response to early, medium, and late switches could reflect this expectation. The authors interpret this effect very differently ("the speed of a perceptual switch should be dependent on LC activity").
The task includes catch trials designed to reduce the expectation of a perceptual switch. In these trials, a perceptual switch occurs either earlier or later than usual. While these trials are valuable for mitigating predictability, we did not focus extensively on them, as they were thoroughly discussed in the original paper. Additionally, due to the limited number of catch trials, it is difficult—if not impossible—to calculate a reliable mean surprise per image set.
It is also worth noting that the pupil study does not include catch trials, which could contribute to differences in how perceptual switches are processed and interpreted between the fMRI and pupil experiments.
- The RNN is far more complex than needed for the task. It has two input units that indicate the level of evidence for the two categories being morphed, and it is trained to output the dominant category. A (non-recurrent) network with only these two units and an output unit whose activity is a sigmoid transform of the difference in the inputs can solve the task perfectly. The RNN activity is almost 1-dimensional probably for this reason. In addition, the difficult part of the computation done by the human brain in this task is already solved in the input that is provided to the network (the brain is not provided with the evidence level for each category, and in fact, it does not know in advance what the second category will be).
We agree that a simpler model could perform the task. We opted to use an RNN rather than hand craft a simpler model as we wanted to use the model as both a method of hypothesis testing and hypothesis generation. We now expand on and justify this modelling choice in the second paragraph of the discussion (also see our response to Reviewer 1 comment 4):
“We chose to use an RNN, instead of a simpler (more transparent) model as we wanted to use the RNN as a means of both hypothesis generation and hypothesis testing. Specifically, unlike more standard neuronal models which are handcrafted to reproduce a specific effect, when building an RNN the modeller only specifies the network inputs, labels, and the parameter constraints (e.g. Dale’s law) in advance. The dynamics of the RNN are entirely determined by optimisation. Post-training manipulations of the RNN are not built in, or in any way guaranteed to work, making them more analogous to experimental manipulations of an approximately task-optimal brain-like system. Confirmatory results are arguably, therefore, a first steps towards an in vitro experimental test.”
In other words, a simpler model would not have been appropriate to the aims. In addition we note that low dimensional dynamics are extremely common in the RNN literature and are in no way unique to our model.
- Basic fMRI results are missing and would be useful, before using elaborate analyses. For instance, what are the regions that are more active when a switch is detected?
We explicitly chose to not run a standard voxelwise statistical parametric approach on these data, as the results were reported extensively in the original study (Stottinger et al., 2018).
- The use of methods from physics may obscure some simple facts and simpler explanations. For instance, does the flatter energy landscape in the higher gain condition reflect a smaller number of states visited in the state space of the RNN because the activity of each unit gets in the saturation range? If correct, then it may be a more straightforward way of explaining the results.
We appreciate the reviewer's concern as this would indeed be a problem. However, this is not the case for our network. At the time point of the perceptual switch where the egocentric landscape dynamics are at their flattest the RNN firing rates are approximately 50% activated nowhere near the saturation point. In addition, a flatter landscape in the egocentric and allocentric landscape analyses only occurs - mathematically speaking - when there are more states visited not less.
In addition, we note that we are very sympathetic to the complexity of our physics based analyses and have gone to great lengths to describe them in an accessible manner in both the main text and methods. We have also included tutorial style code demonstrating how the analysis can be used on a toy dynamical system in the supplementary material.
- Some results are not as expected as the authors claim, at least in the current form of the paper. For instance, they show that, when trained to identify which of two inputs u1 and u2 is the largest (with u2=1-u1, starting with u1=1 and gradually decreasing u1), a higher gain results in the RNN reporting a switch in dominance before the true switch (e.g. when u1=0.6 and u2=0.4), and vice et versa with a lower gain. In other words, it seems to correspond to a change in criterion or bias in the RNN's decision. The authors should discuss more specifically how this result is related to previous studies and models on gain modulation. An alternative finding could have been that the network output is a more (or less) deterministic function of its inputs, but this aspect is not reported.
We appreciate this comment but it is simply not applicable to our network. There is no criterion in the RNN. We could certainly add one but this would be a significant departure from how decisions are typically modelled in RNNs. The (deterministic) readout is the max of the projection of the (instantaneous) excitatory firing rate onto the readout weights. A shift in criterion would imply that the dynamics are unaffected and the effect can be explained by a shift in the readout weights; this cannot be the case because the readout weights are stationary the change occurs at the level of the activation function.
We are aware that there is a large literature in decision making and psychophysics that uses the term gain in a slightly different way. Here we are strictly referring to the gain of the activation function. Although we agree that it would be interesting and important to discuss the differing uses of the term gain, this is beyond the scope of the present paper.



