NOVA-dependent regulation of cryptic NMD exons controls synaptic protein levels after seizure
- Rockefeller University, United States
- Baylor College of Medicine, United States
- Howard Hughes Medical Institute, Rockefeller University, United States
Figures
Figure 1 with 2 supplements
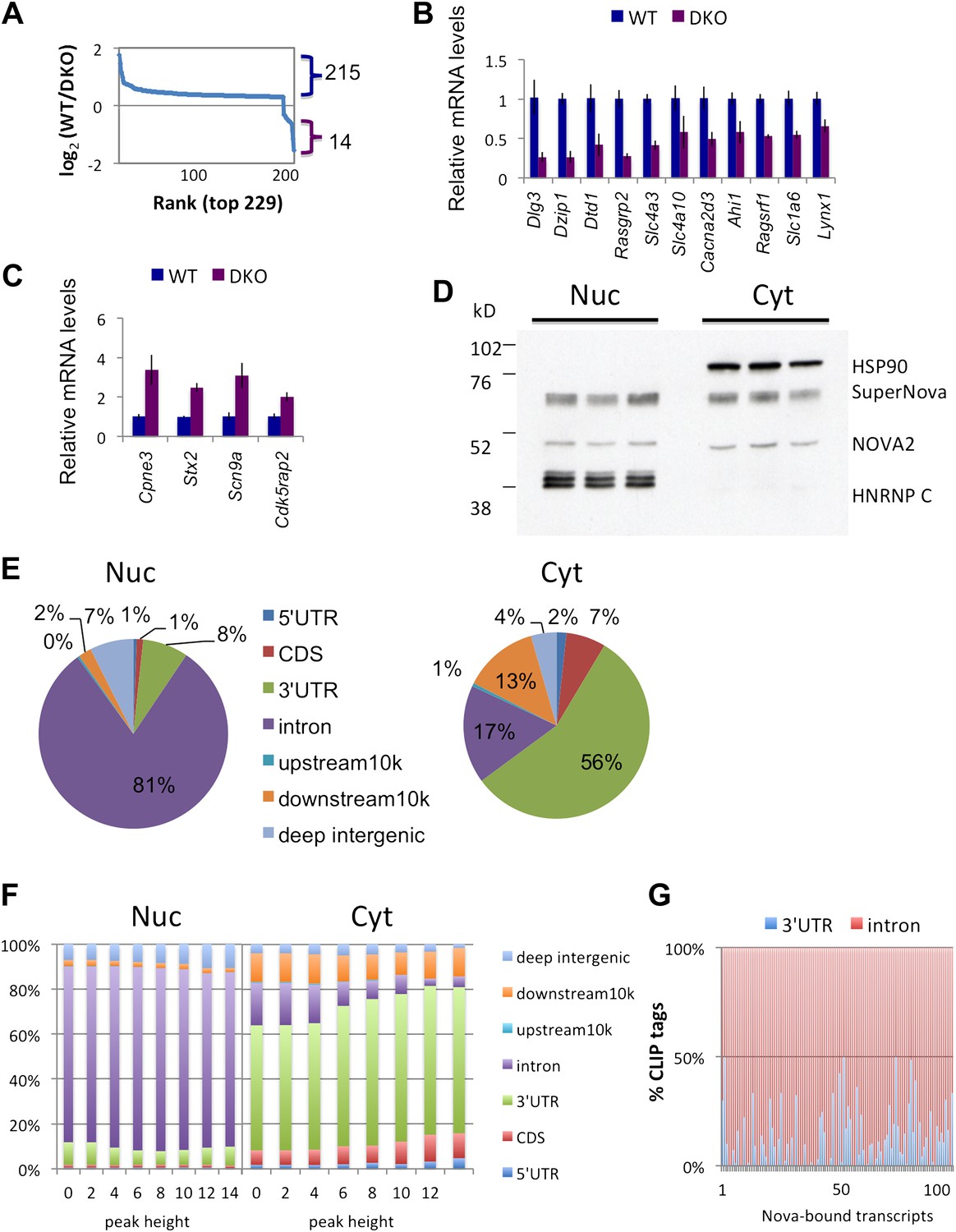
NOVA proteins up/down-regulate transcript levels.
(A) Affymetrix exon arrays were interrogated with RNA from WT vs DKO E18.5 mouse whole brains, and normalized transcript intensities were plotted in log2 scale (log2(WT/DKO) > 0.3 or −0.3 and p<0.05). The X-axis indicates ranks of transcripts from the top and Y-axis is the measure of relative transcript levels (WT/DKO) in log2 scale. Blue bracket represents transcripts whose levels are increased in WT relative to DKO brain (NOVA-dependent ‘upregulation’ of steady-state mRNA levels), and the purple bracket represents down-regulated transcripts in WT relative to DKO (NOVA-dependent repression of steady-state mRNA levels). (B),(C) qRT-PCR data of representative NOVA regulated transcripts. Y-axis represents the mRNA levels in which WT is normalized to 1.0. Data is from three biologic replicates (three animals) and three technical replicates (nine reactions per point); error bars represent standard deviation. For each point p<0.001; see Table 1 for additional data. NOVA up-regulated transcripts in WT versus DKO (B; corresponding to blue bracket in Figure 1A) and NOVA down-regulated transcripts (C; corresponding to purple bracket in Figure 1A) are shown. (D) Immunoblot analysis of NOVA distribution in nuclear and cytoplasmic fractions from mouse brain irradiated by UV. Each lane represents the different brain extracts as biological replicates. HSP90 is used as a cytoplasmic marker, and hnRNP-C1/C2 as a nuclear marker. The NOVA2 antibody detects both large and small NOVA2 isoforms (Yang et al., 1998). (E) Breakdown of BC = 4 clusters for nuclear and cytoplasmic Nova HITS-CLIP. Downstream 10K clusters are enriched in unannotated 3′ UTRs (Licatalosi et al., 2008); see also Table 2. (F) Distribution of BC = 4 clusters by peak height for both nuclear and cytoplasmic HITS-CLIP; more stringent cytoplasmic clusters show enrichment in 3′ UTR. (G) Distribution of CLIP tags (intronic, red; 3′ UTR, blue) from the list of NOVA up-regulated RNAs. Each point on the X-axis represents a Nova-dependent gene (in arbitrary order) and Y-axis represents the percentage of intronic/3′ UTR tags for transcripts with total tags >5.
Figure 1—figure supplement 1
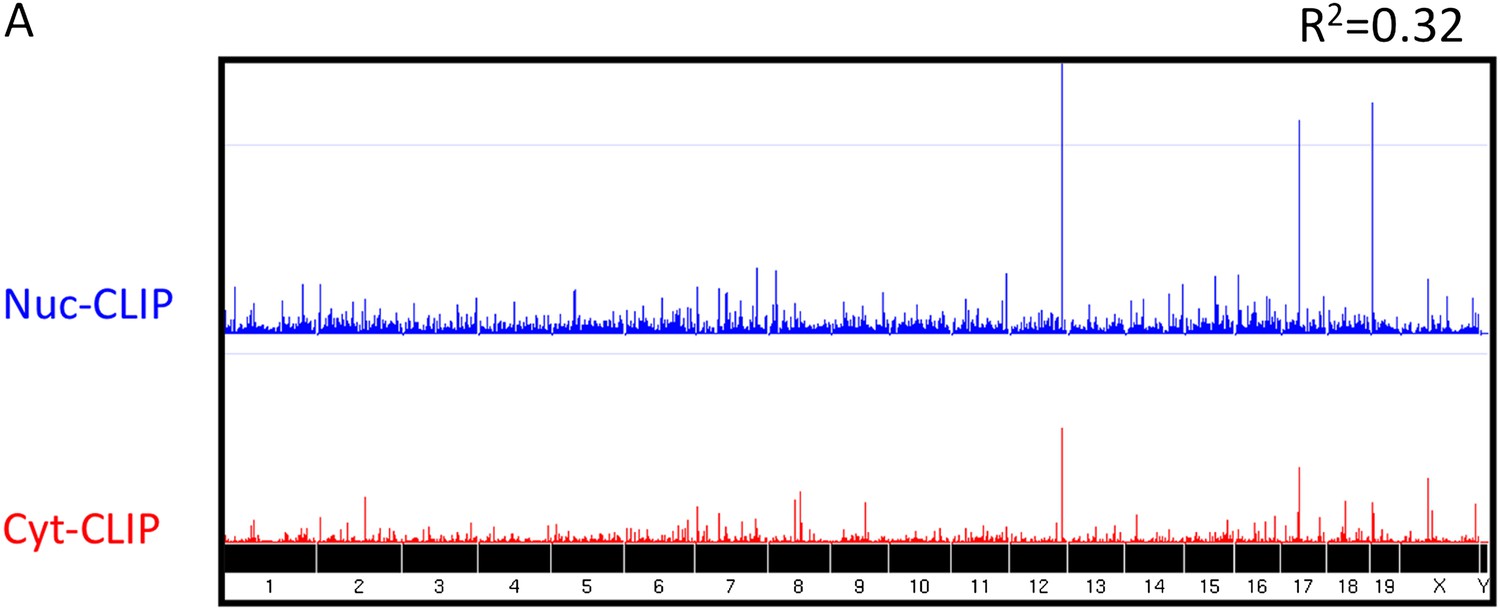
Nova CLIP results.
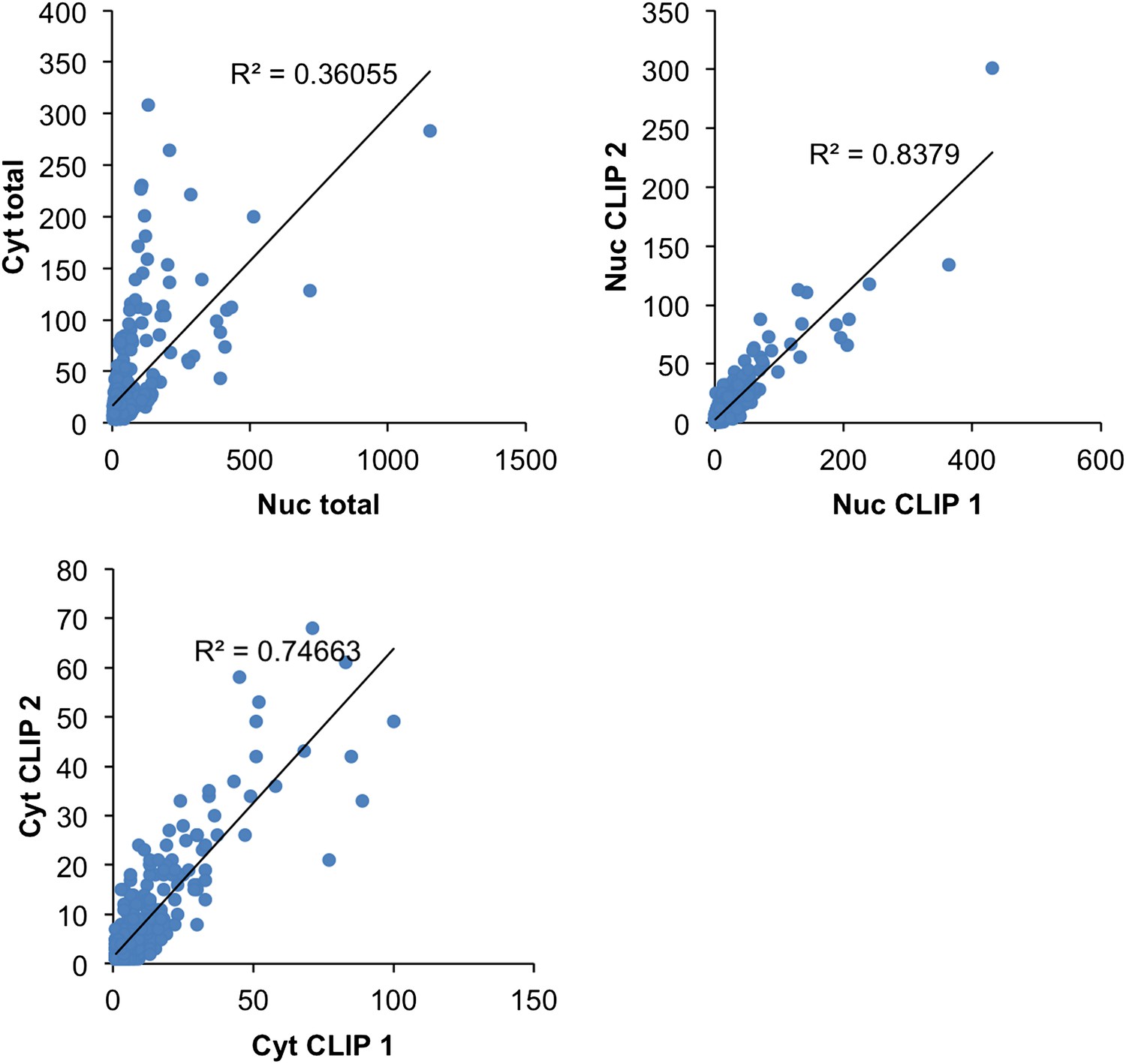
Nuclear (Nuc, blue; upper) and cytoplasmic (Cyt, red; lower) HITS-CLIP tags are shown. All unique tags from nuclear and cytoplasmic HITS-CLIP clusters were plotted onto the whole genome and showed a relatively low correlation (R2 = 0.32). By comparison, two independent cytoplasmic HITS-CLIP tags showed higher R2 values (0.765; data not shown). (B) BC4 clusters were grouped according to each gene, and CLIP tags from these clusters were summed up gene-by-gene for each individual experiment, to obtain the total number of BC4 tags for each gene. The first plot (Cyt total vs Nuc total) shows the correlation (R2 = 0.36) between the total number of nuclear CLIP tags per gene and the total number of cytoplasmic tags per gene; each point represents a gene. The other two plots are similar but the correlations were calculated using two Nuc CLIP experiments or two Cyt CLIP experiments.
Figure 1—figure supplement 2

Correlations: analysis of Nova CLIP results.
NOVA HITS-CLIP BC4 clusters were grouped according to each gene, and CLIP tags from these clusters were summed up gene-by-gene for each individual experiment, to obtain the total number of BC4 tags for each gene. The first plot (Cyt total vs Nuc total) shows the correlation (R2 = 0.36) between the total number of nuclear CLIP tags per gene and the total number of cytoplasmic tags per gene; each point represents a gene. The other two plots are similar but the correlations were calculated using two Nuc CLIP experiments or two Cyt CLIP experiments.
Figure 2 with 2 supplements
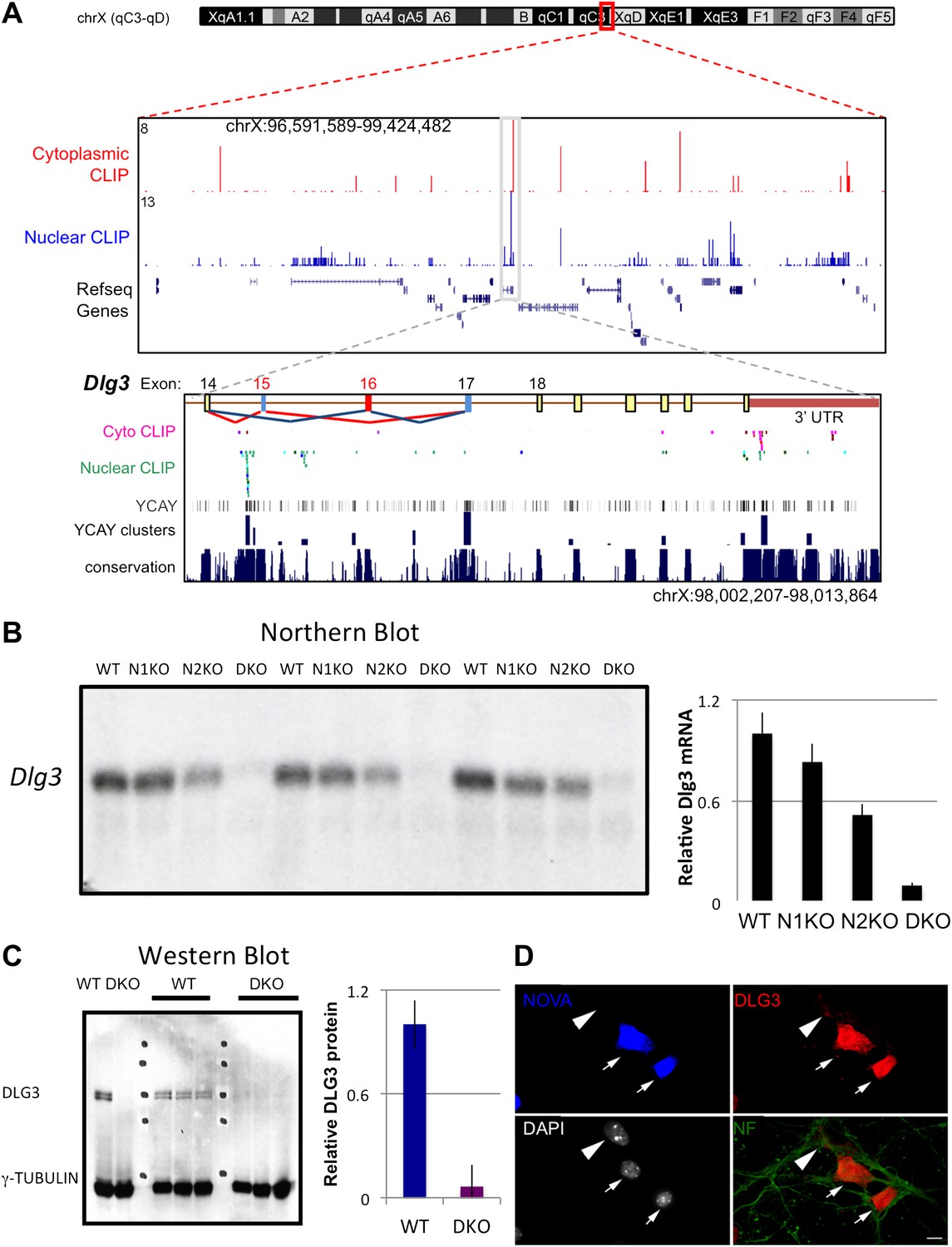
NOVA regulates the expression of Dlg3 mRNA and protein.
(A) Location of NOVA cytoplasmic and nuclear CLIP tags in chromosome X:96591589-99424482. Red and purple colors represent cytoplasmic CLIP tags and green and blue colors represent nuclear CLIP tags. The location of Dlg3 is boxed in black and magnified in the lower box (chromosomeX:98002207-98013864). This higher magnification illustrates the position of Dlg3 constitutive (yellow), alternative (colored) exons and 3′ UTR (brown) relative to CLIP tags, YCAY elements, and sequence conservation across species. More cytoplasmic tags were evident in the 3′ UTR and more nuclear tags in introns. Clusters of CLIP tags can be seen to overlap with the location of clusters of YCAY sequences (in grey) as well as bioinformatically predicated clusters of YCAY elements (in blue; see Zhang et al., 2010). (B) Northern blot analysis of Dlg3 mRNA from three biologic replicates of WT or Nova KO brain mRNA. Equal amount of RNA was loaded (see Figure 2—figure supplement 2). Quantitation of relative RNA intensity (WT/DKO) was plotted as a relative ratio of Dlg3 mRNA in WT, N1 KO, N2 KO or DKO brain as indicated; error bars represent standard deviation (p<0.05); about 90% of Dlg3 mRNA is absent in DKO brain. (C) Immunoblot analysis of DLG3 in WT vs DKO. Protein extracts from the four different WT or DKO mouse brains (as indicated; E18.5) were assessed, and γ-TUBULIN was used as a normalizing control. Quantitation of protein intensity is indicated in graph to the right, plotted as relative ratio of DLG3 in WT/DKO, indicate that ∼90% of DLG3 protein is absent in DKO brain; error bars represent standard deviation (p<0.05). (D) Immunofluorescence detection of DLG3 (red), NOVA (blue) and Neurofilament (NF) (green) proteins on WT/DKO mixed primary mouse neuronal cultures. DAPI and neurofilament stained all neuronal nuclei and processes, respectively, while NOVA staining differentiates WT and DKO neurons. The DLG3 signal was markedly reduced in DKO neurons. Scale bar: 10 µm.
Figure 2—figure supplement 1
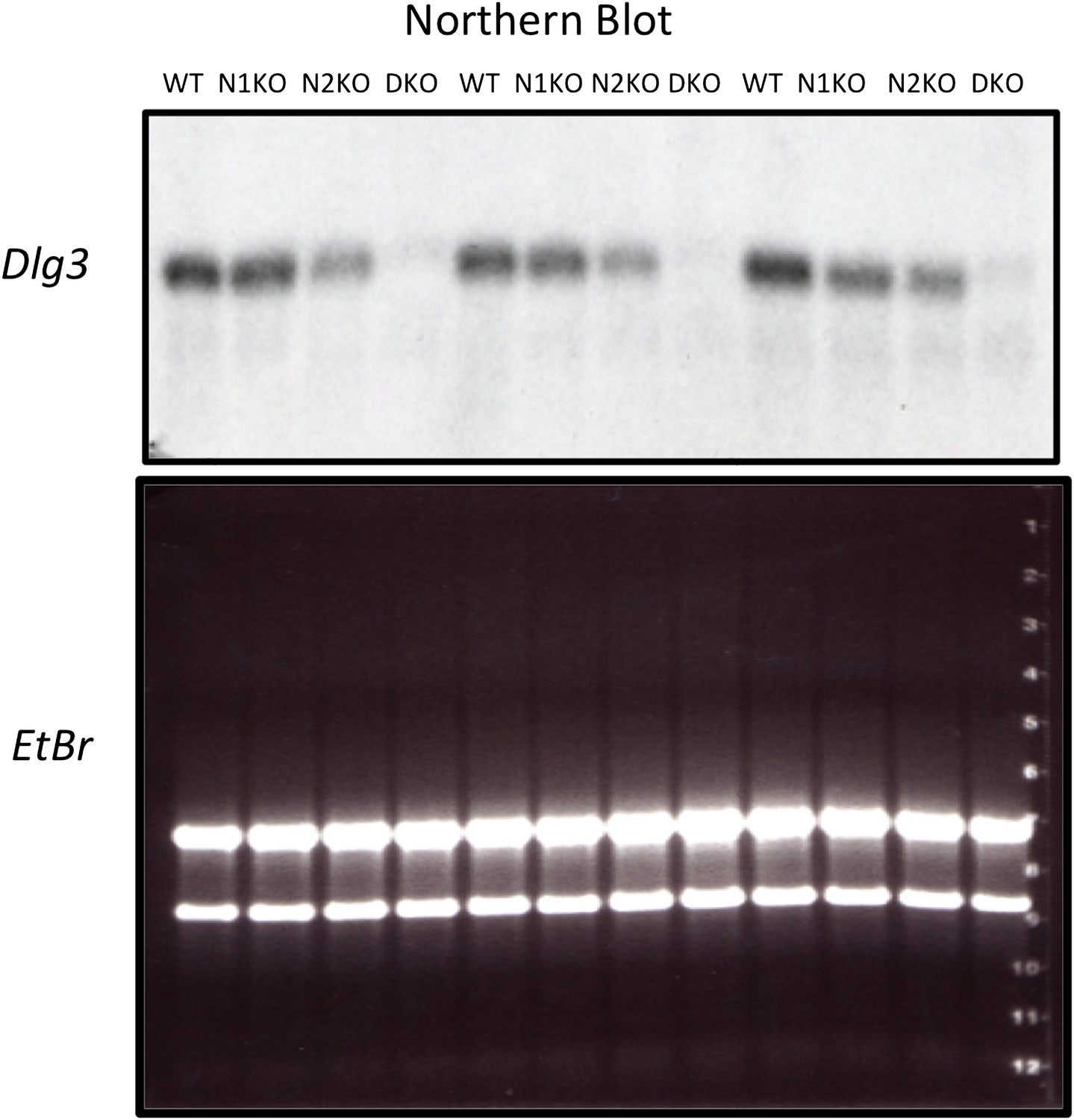
Dlg3 mRNA isoforms in Nova KO brain.
Northern blot analysis of Dlg3 mRNA in WT and Nova DKO brain. (A) Gapdh probe was used as a normalizing control. Panel to right: Quantitation of relative RNA intensity (WT/DKO) was plotted as a relative ratio of Dlg3 mRNA/GAPDH in WT/DKO; error bars represent standard deviation (p<0.05). About 75% was reduced in DKO.
Figure 2—figure supplement 2
Northern blot analysis of Dlg3 mRNA in Nova KO brain-reproducibility and control.
Reproduction of Northern blot for Dlg3 mRNA presented in Figure 2B, but also showing loading control below (Ethidium bromide (EtBr) stain of the gel), demonstrating equal loading of all RNAs.
Figure 3 with 2 supplements
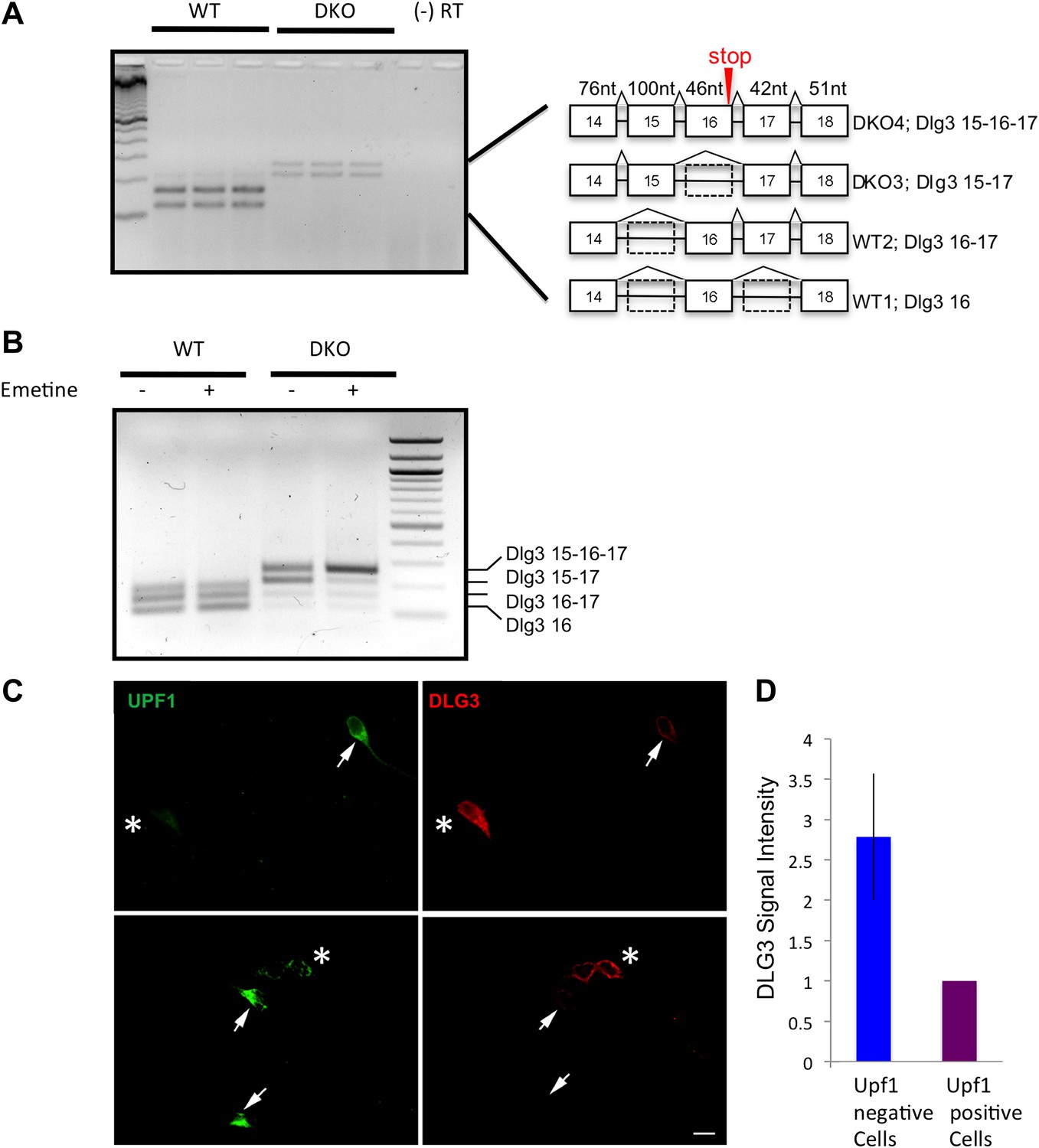
Dlg3 mRNA is decreased through inclusion of cryptic NMD exons in the absence of NOVA.
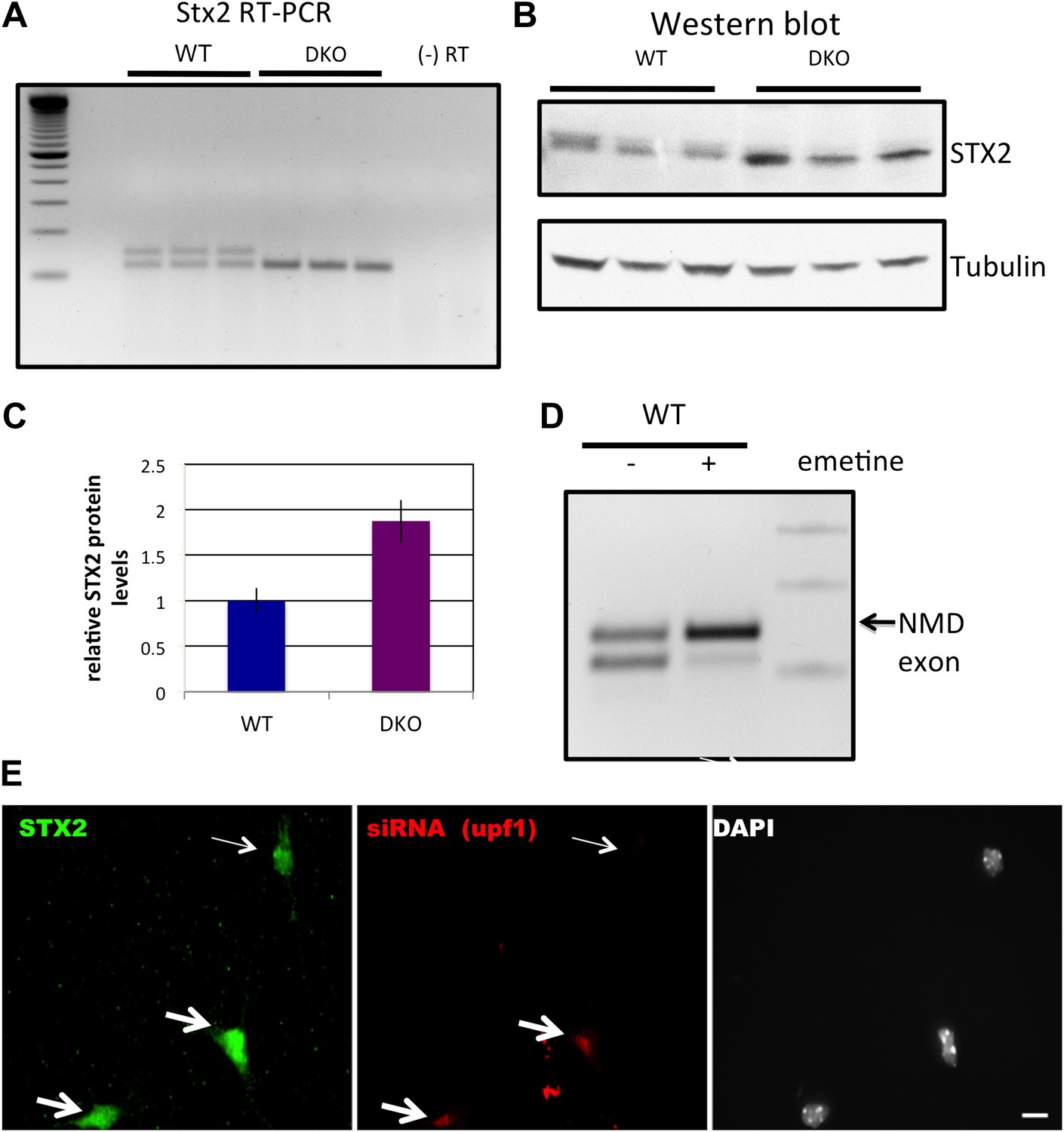
(A) RT-PCR and sequence analysis (Figure 3—source data 1) showed four Dlg3 isoforms which have different combinations of alternatively spliced exons. The lower two PCR products (WT1 and WT2), respectively, harbor alternative exon 16 (E16) with or without E17, and both encode in-frame protein variants. The upper two PCR products, evident in DKO brain (DKO3 and DKO4), include E15, with or without E16, and include E17. The DKO4 isoform was not annotated in Refseq, and the combination of E15 and E16 led to a frameshift and inclusion of a premature stop codon (TAA) in E16, as indicated in the schematic (also leading us to color E16 red in Figure 2A). The 15–17 containing product does not make a premature stop codon. (B) After 6 DIV primary mouse neuronal cultures were treated with 100 μg/ml emetine for 10 hr, RNA was harvested from triplicate samples and analyzed by RT-PCR. Emetine treatment had no effect cell viability nor on isoforms produced in WT cells, but led to accumulation of the NMD isoform DKO4 in Nova DKO neurons. Spliced isoforms are indicated. (C) siRNA to Upf1 was transfected into DKO mouse primary cells, and after 24 hr, DLG3 (red) and UPF1 (green) protein was detected by immunofluorescence microscopy. Arrows indicate cells that have relatively high UPF1 levels; these cells have low DLG3 levels. In contrast, cells transfected with Upf1 siRNA (asterisks) had markedly reduced Upf1 levels and had increased (rescued) DLG3 levels. Upper and lower rows represent independent experiments. Scale bar: 10 µm. (D) Quantitation of DLG3 signal in Upf1 positive and Upf1 knockdown cells in (C). Signal intensity was normalized to the signal in Upf1 positive cells. Error bar represents standard deviation (p<0.05).
-
Figure 3—source data 1
Sequence of RT-PCR products (from Figures 3 and 5).
- https://doi.org/10.7554/eLife.00178.012
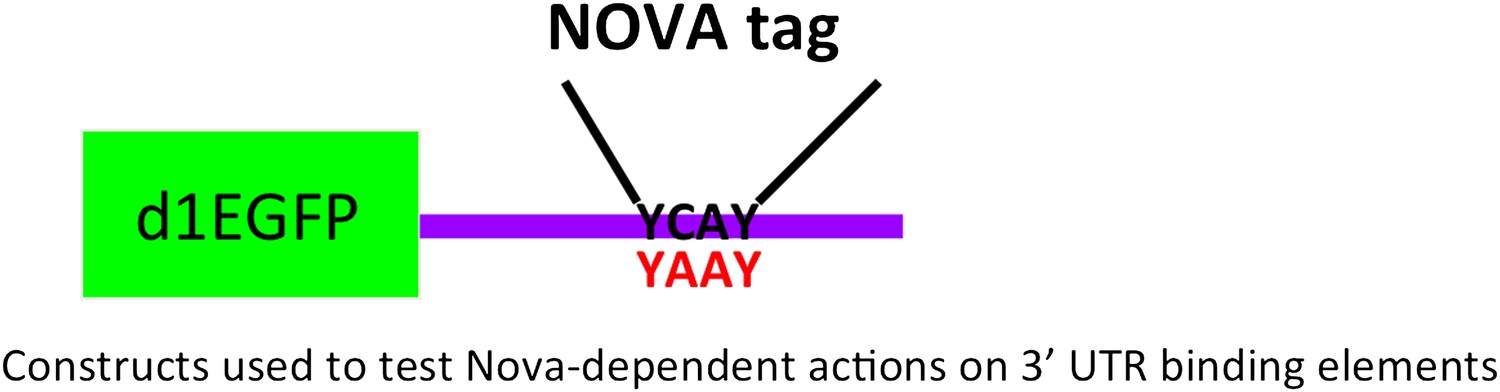
Figure 3—figure supplement 1

Reporter construct design for NOVA 3′ UTR actions.
Schematic of a reporter encoding destabilized d1EGFP and the 3′ UTR YCAY element of NOVA's target RNA. An identical construct in which the YCAY sequences were mutated to YAAY was also made.
Figure 3—figure supplement 2
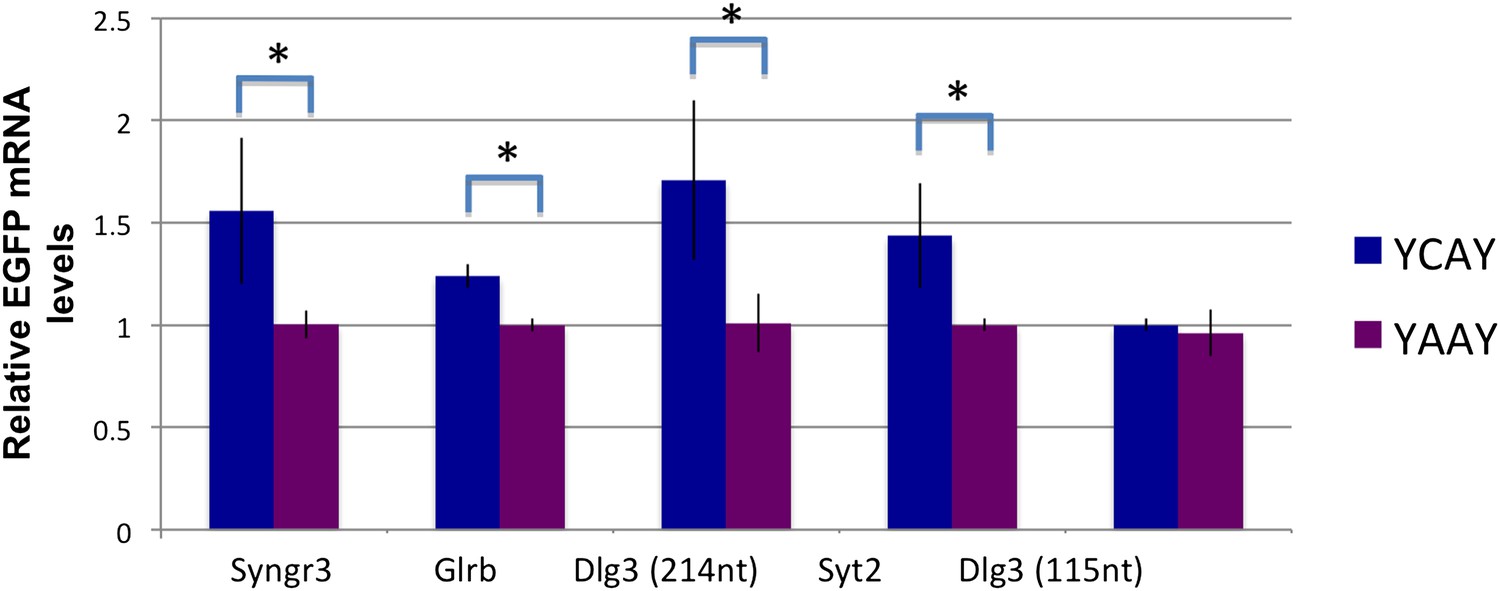
NOVA increases the stability of some RNAs through binding to 3′ UTR YCAY elements.
qRT-PCR was used to quantitate d1EGFP mRNA after transfection of various constructs (Syngr3 [synaptogyrin 3], Glrb [Glycine receptor b], Dlg3 and Syt2 [Synaptotagmin 2]). Quantitation of steady state changes in RNA levels (YAAY relative to YCAY constructs) were 50%, 25%, 65%, 48%, respectively, for Syngr3, Glrb, Dlg3 and Syt2. The Y-axis represents the relative mRNA levels (YCAY/YAAY). Interestingly, for Dlg3, a small 3′ UTR construct (115 nt) did not show any change, while a longer construct (214 nt) did, even though both harbored the YCAY element, suggesting that additional elements or RNA structures may be important for binding of NOVA to this 3′ UTR YCAY element. Asterisks indicate p<0.05.
Figure 4 with 2 supplements
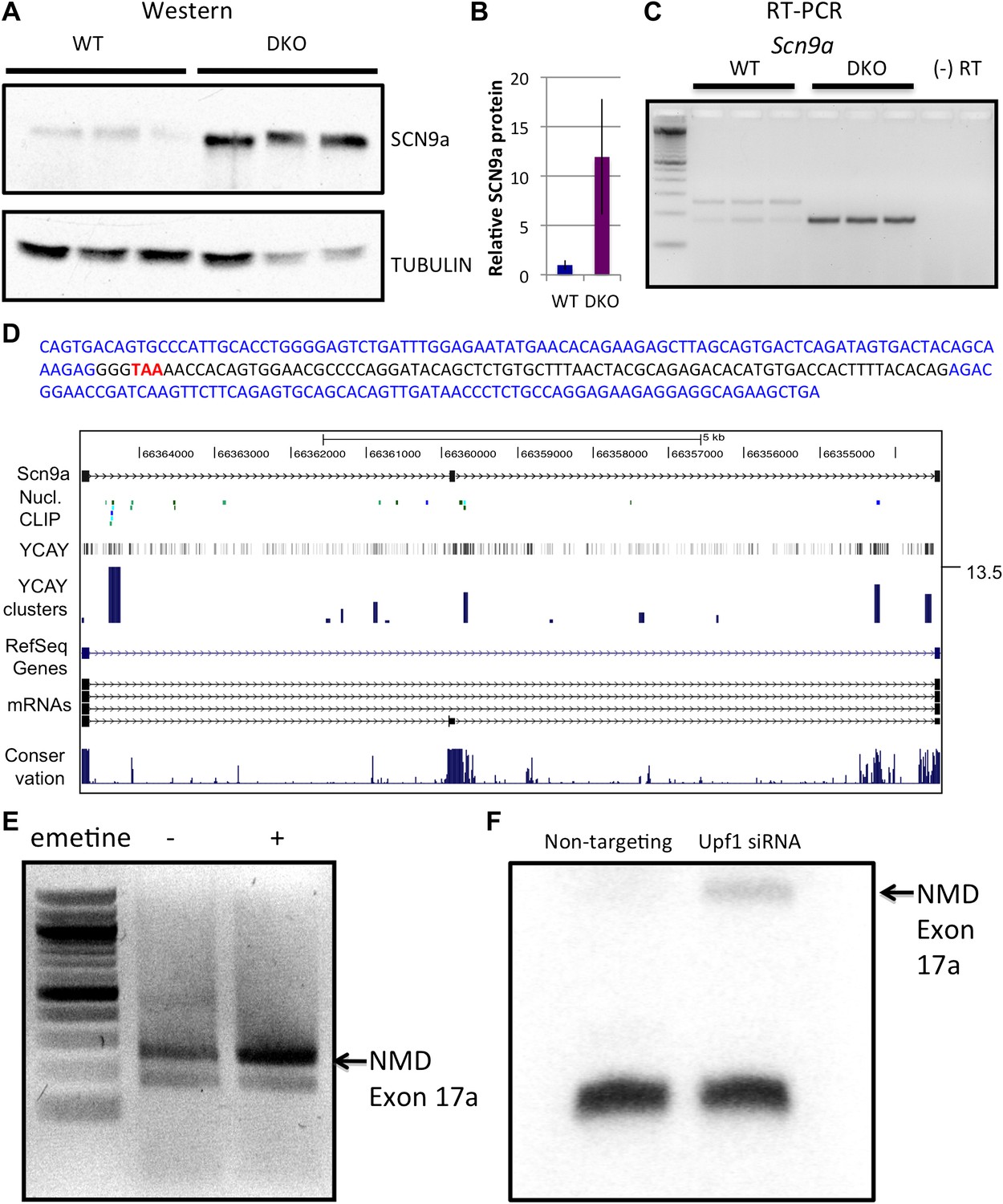
NOVA and NMD-mediated regulation of Scn9a mRNA and protein.
(A) Immunoblot analysis of SCN9a in WT vs DKO. Proteins extracts from WT (lane 1–3) vs DKO (lane 4–6) were loaded. γ-Tubulin was used as a normalizing control. (B) Quantitation of relative protein intensity (WT/DKO). The results were plotted as a relative ratio of SCN9a in WT/DKO; error bars represent standard deviation (p<0.05). About 90% signal was reduced in WT. (C) RT-PCR from WT and DKO (three biologic replicates) shows NOVA-dependent splicing of Scn9a. WT brains express two alternative splicing isoforms (lanes 1–3), while DKO brains express only the smaller isoform (lanes 4–6). A (−) RT control is indicated; primers are given in Supplementary file 2. (D) Sequence analysis and map of the spliced isoforms from Figure 4C showed that the larger band corresponds to a transcript in which an exon (17a) was included, introducing a premature stop codon. In the sequence shown, exon 17a is highlighted in black, and the TAA premature stop codon is indicated in red. (E) Six DIV WT primary mouse neuronal cultures were treated with emetine, as indicated, for 10 hr and RT-PCR was performed. The NMD exon was increased after emetine treatment. (F) siRNA to Upf1 or a non-targeting siRNA were transfected in WT mouse primary cells, and RT-PCR was performed with the same primers used in Figures 4C,E. The intensity of the NMD exon was increased specifically after Upf1 siRNA treatment.
Figure 4—figure supplement 1
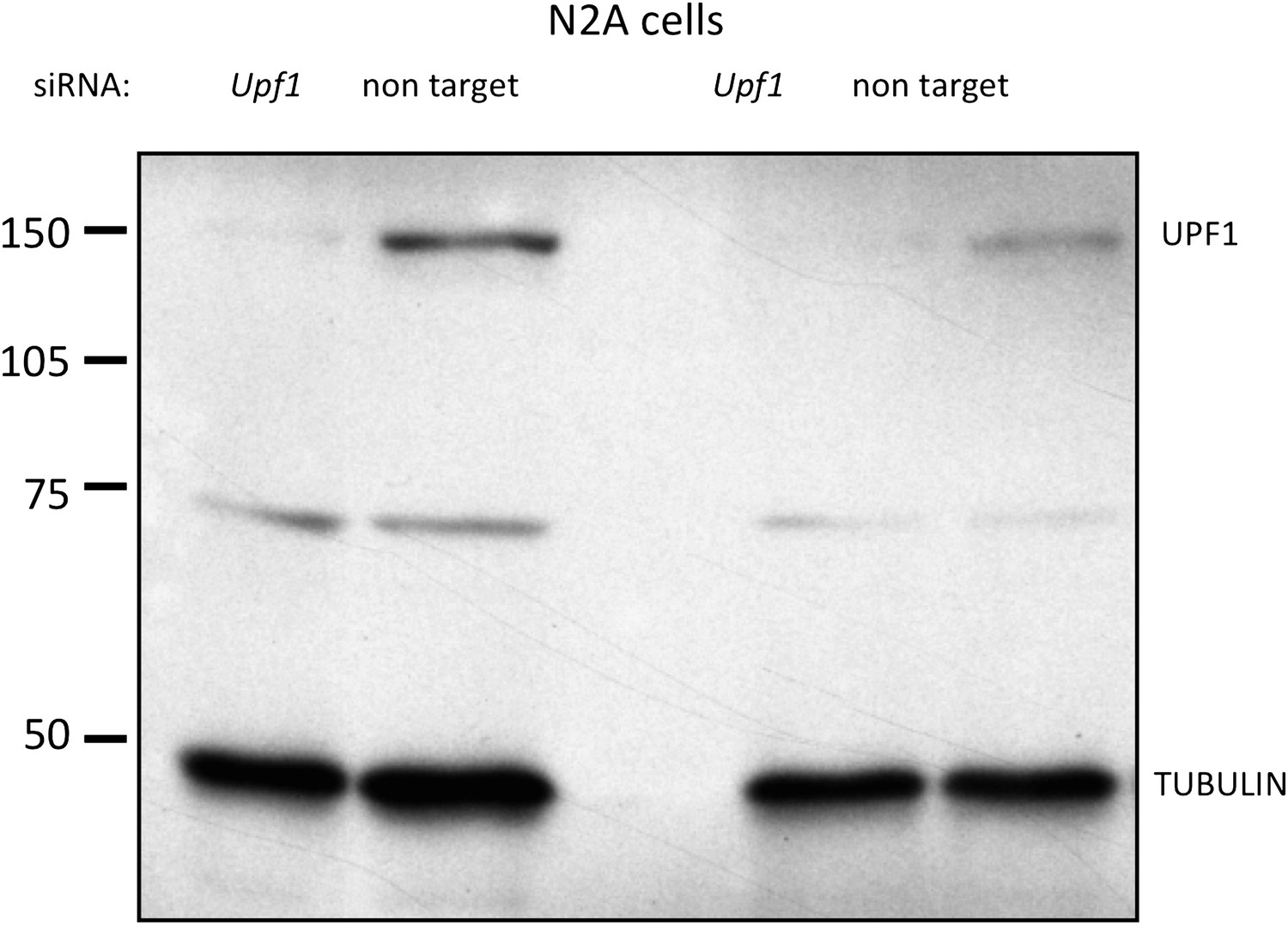
N2A cells: Upf1 siRNA efficiently reduces endogenous UPF1 protein levels.
siRNA targeting Upf1 was transfected into N2a cells and immunoblot analysis of UPF1 protein was performed. Results from two independent experiments are shown. siRNA to Upf1 showed the significant reduction of UPF1 protein expression compared with a control untargeting siRNA. γ-Tubulin was used as a normalization control.
Figure 4—figure supplement 2
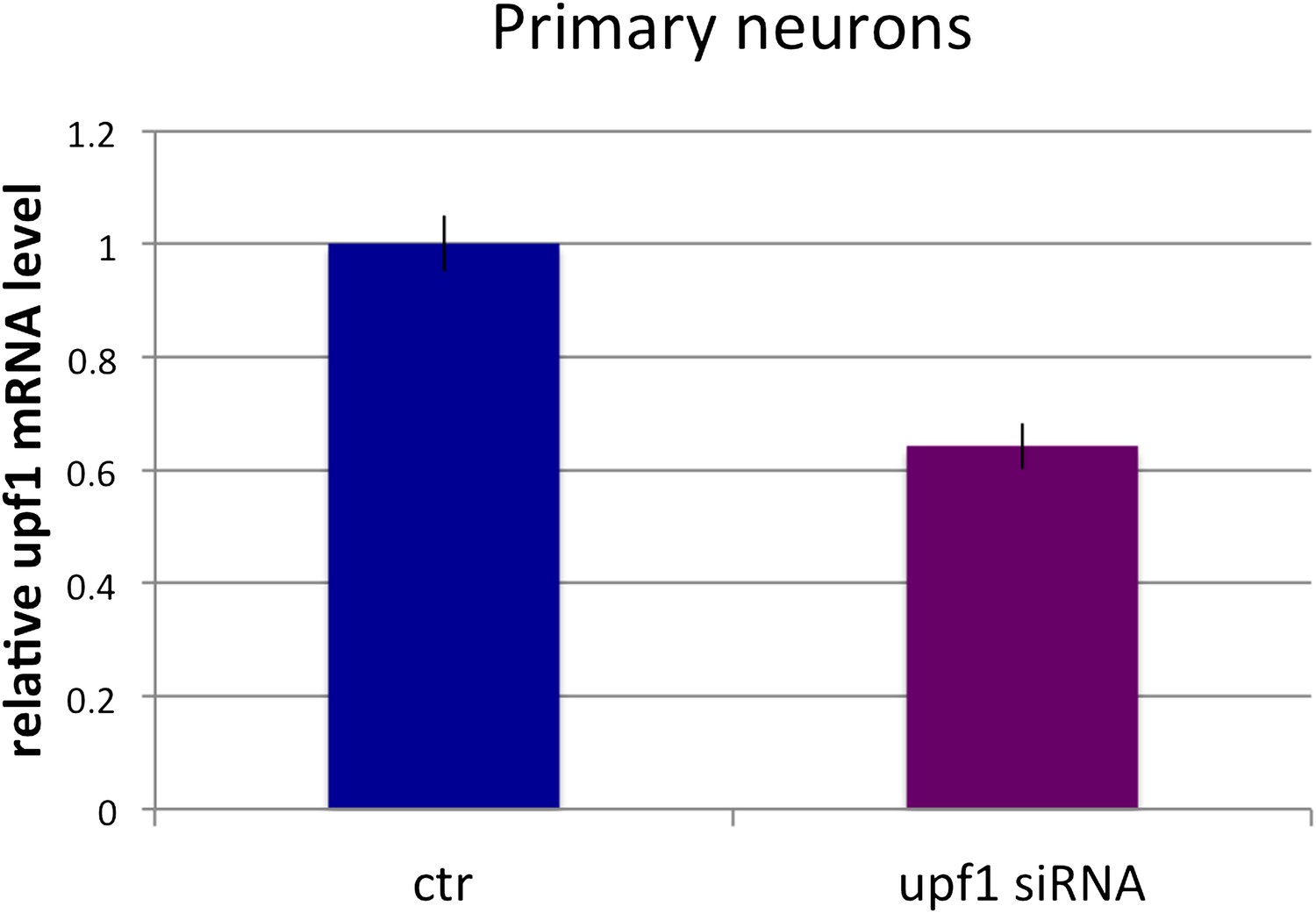
Primary neurons: Upf1 siRNA reduces endogenous UPF1 protein levels.
qRT-PCR quantitation of relative Upf1 mRNA levels in primary neurons with or without treatment with Upf1 siRNA. Error bars represents standard error of the mean; n = 3.
Figure 5 with 2 supplements

NOVA regulates cryptic NMD exons and transcript levels.
(A) Analysis of alternative spliced isoforms in transcripts chosen solely on exon array data showing NOVA-dependent steady-state mRNA changes and robust HITS-CLIP clusters in introns. Transcripts were then screened for the presence of cryptic NMD exons by RT-PCR using primers in exons bounding the intronic HITS-CLIP clusters. Data is divided into those transcripts down-regulated or up-regulated in Nova DKO, as indicated. Sequence analysis of RT-PCR products showed the presence of cryptic exons harboring premature stop codons (Figure 3—source data 1; Supplementary file 2). A diagram of the loci of each NMD exon present in Figure 5A is shown in Figure 5—figure supplement 1. For example, most transcripts down-regulated in Nova DKO brains show a larger, PTC containing exon in DKO; one exception is Actl6b, in which in the absence of NOVA there is a PTC, and in WT brain, an upper alternate isoform (exon) is present that corrects that frame-shift; (B) Effect of emetine on putative NOVA-regulated cryptic NMD exons. The steady-state level of six transcripts identified in Figures 3B, 4C and 5A were assessed by qRT-PCR in six DIV WT vs Nova DKO primary mouse neurons incubated for 10 hr in the presence or absence of emetine. The results were plotted with the Y-axis as a measure of the degree of putative NOVA-dependent NMD regulation (the fold change of transcript levels in DKO neurons in the presence or absence of emetine, divided by that of WT, in log2 scale). For example, for Dlg3 the log2 value is about 1.0 indicating that emetine treatment increased the Dlg3 NMD-isoform in DKO neurons relative to WT neurons by a factor of two, while emetine decreased the NMD isoform of Scn9a by ∼1.4-fold, leading to decrease or increase in the respective proteins in Nova DKO neurons (Figures 2 and 3 or Figure 4, respectively). Three independent experiments were performed and error bars represent standard deviation (p<0.05).
Figure 5—figure supplement 1
Diagrams of each of the NMD exons shown in Figure 5A
The position of the exon harboring a premature termination codon is labeled in red (PTC).
Figure 5—figure supplement 2
NOVA regulates the expression of Stx2 (Syntaxin 2) mRNA and protein.
(A) RT-PCR from WT and DKO brain showed NOVA-dependent alternative splicing of a previously unknown, higher molecular weight isoform of Stx2. A (−) RT negative control is shown. (B) Immunoblot analysis of STX2 protein in biologic triplicate samples of WT vs Nova DKO brain. γ-Tubulin was used as a normalization control. (C) Quantitation of relative protein intensity (WT/DKO) from (B), plotted as relative ratio of STX2 in WT/DKO; error bars represent standard deviation (p<0.05). (D) Six DIV WT primary mouse culture were treated with vehicle (−) or emetine (+) for 10 hr and RT-PCR was performed for Stx2 as in (A). Quantitation revealed that the NMD exon (labeled) was increased twofold after emetine treatment. (E) DY547 direct-labeled siRNA targeting Upf1 was transfected into WT mouse primary cells, and immunofluorescence microscopy used to detect STX2 (green) and siRNA to Upf1 (red). The thin arrow (top right corner) indicates a cell which was not transfected with siRNA, and which shows a baseline signal of STX2 protein. The two thicker arrows indicate cells which had a good red signal (siRNA to Upf1) and stronger green signals (indicating increased STX2 protein). Scale bar: 10 µm.
Figure 6 with 1 supplement
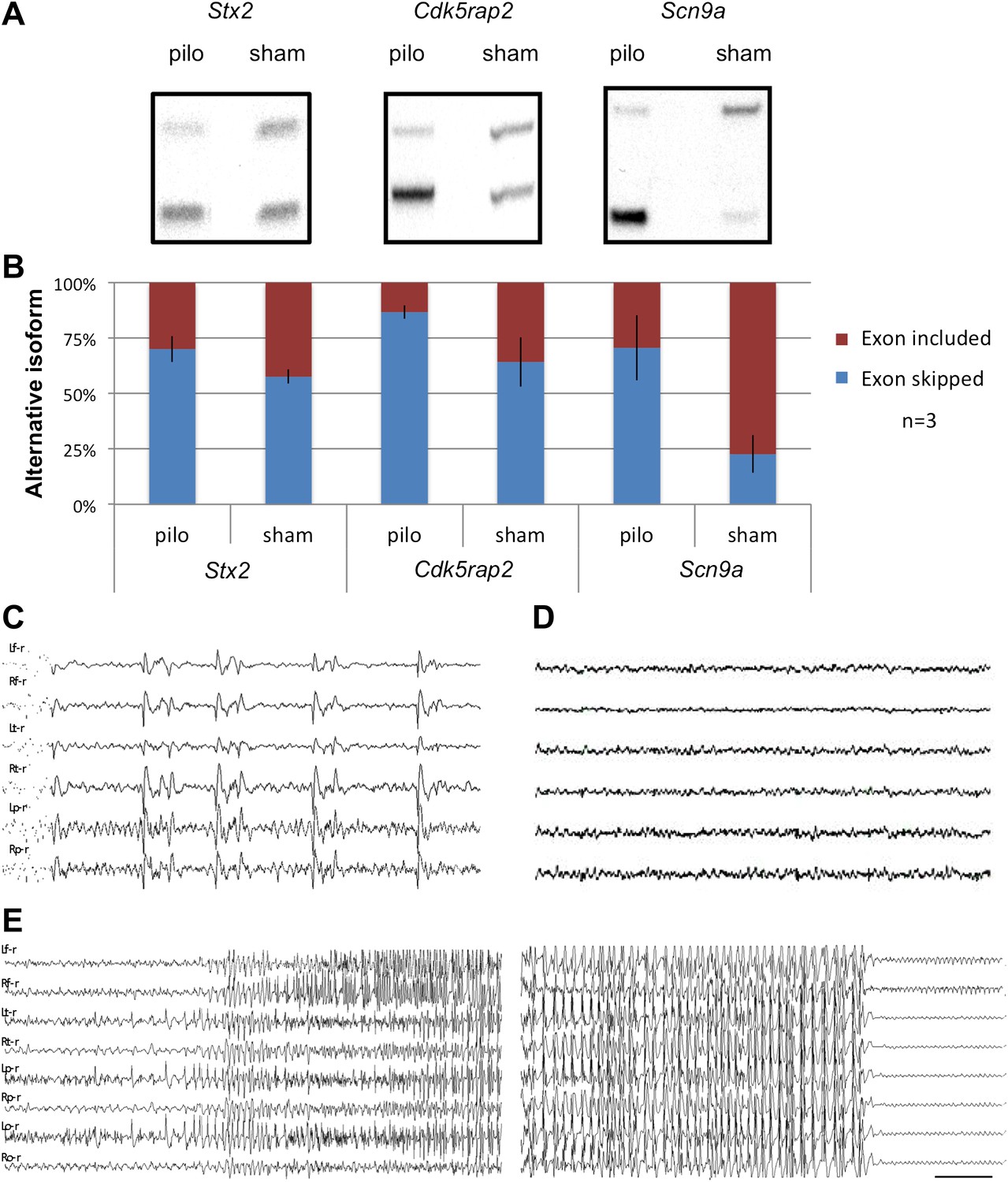
Seizure-induced inhibition of NOVA-regulated NMD exons.
(A) Mice were treated with pilocarpine (pilo) to induce seizures or were mock-treated (sham). 2 hr later brains were harvested and splicing of NMD exons assayed by RT-PCR. (B) Quantitation of experiments described in (A), from three biologic replicates. Error bars represent standard deviation (p<0.05; Student's t-test). (C),(D) EEG of freely moving Nova2+/− mutant displays frequent synchronous cortical interictal discharges (C) not detected in wild type mice (D). (E) Spontaneous generalized seizure discharge in adult Nova2+/− mutant. A 20 s gap of continuous hypersynchronized EEG pattern separates the beginning and end of the seizure discharge. Bilateral left and right frontal, temporal, parietal (C, D) and occipital (E) leads are shown. Time calibration 1 s (C, D), 2 s (E).
Figure 6—figure supplement 1
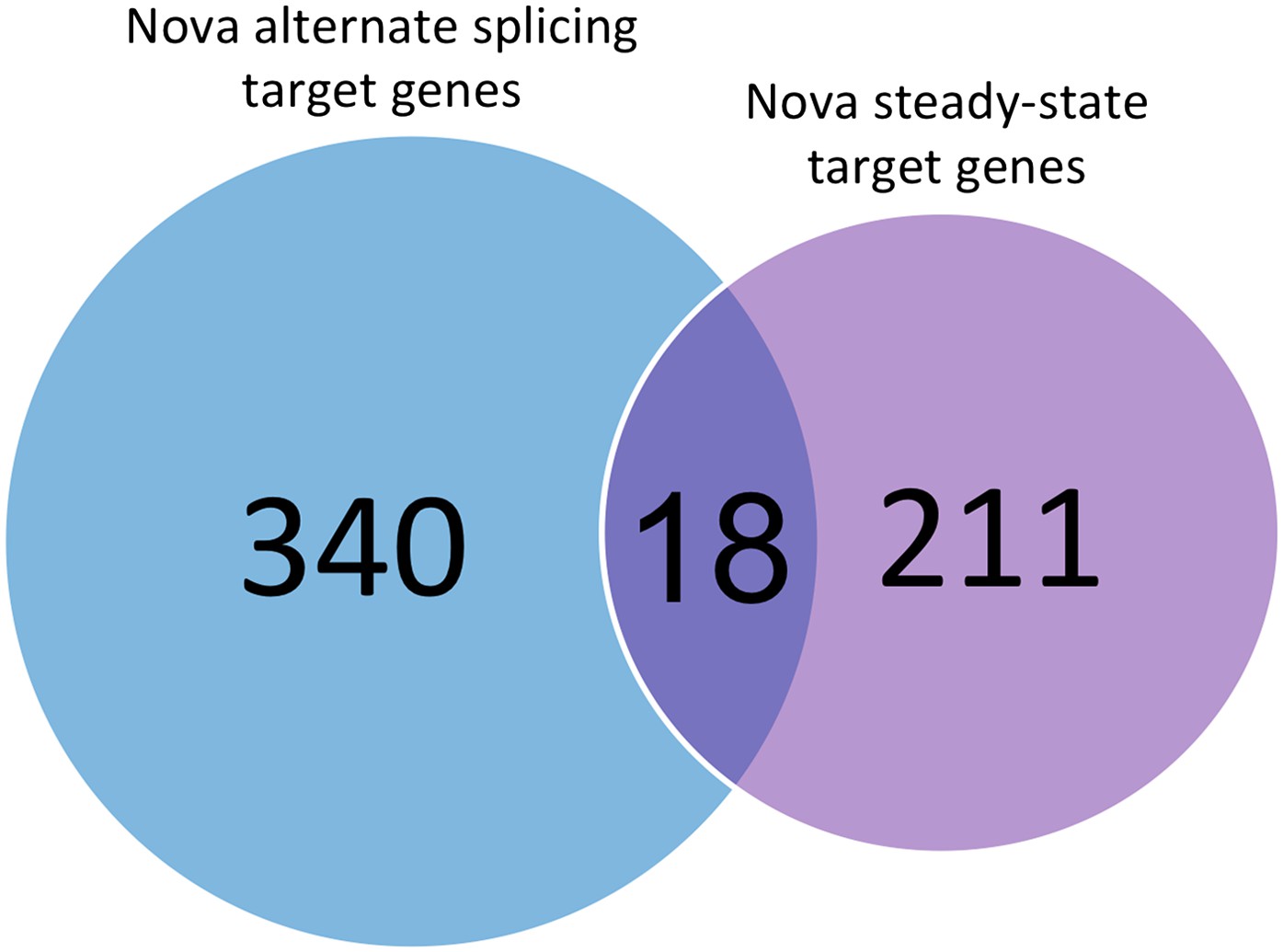
Comparison of different sets of transcripts regulated by NOVA.
The number of transcripts in which NOVA regulates alternative splicing without changing steady-state mRNA levels are shown in blue, and in purple transcripts showing Nova-dependent changes in steady-state mRNA levels (e.g. through NMD or transcript stability). 340 transcripts in which cassette exons are regulated by NOVA were compiled from previous studies (Ule et al., 2003; Licatalosi et al., 2008; Zhang et al., 2010), and compared with the 211 transcripts identified here by exon array. The datasets are largely mutually exclusive, with only 18 transcripts in common.
Figure 7
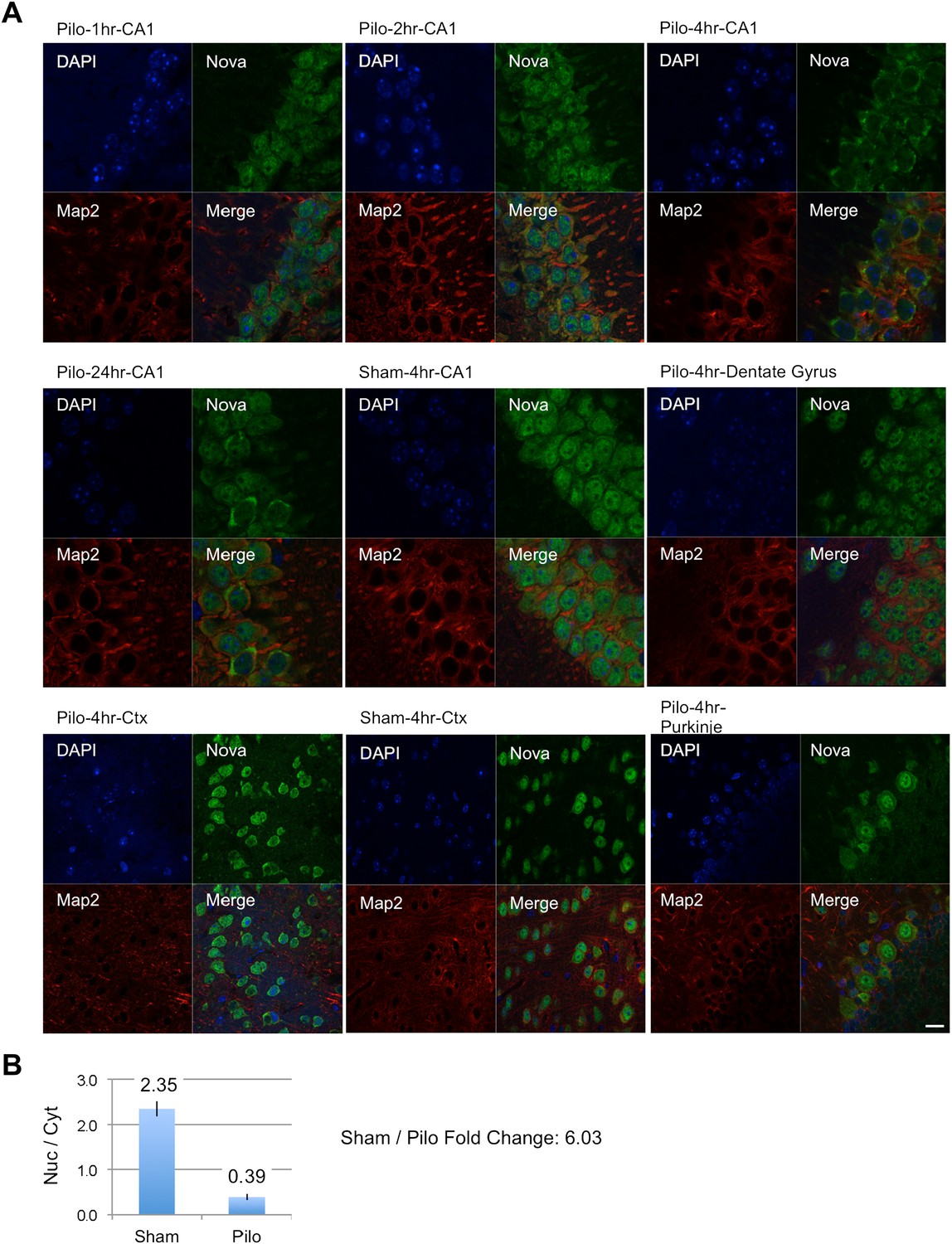
NOVA subcellular localization after pilocarpine-induced status epilepticus.
(A) NOVA and MAP2 protein localization were visualized by immunofluorescence (IF) in mouse brain sections from pilocarpine treated (pilo) or control animals (sham). Regions include CA1 and dentate gyrus (DG) from hippocampal area, cortex (Ctx) and cerebellar Purkinje neurons. Changes were clearly evident in CA1 neurons, were not evident in DG, and were variable in Ctx. No changes were expected nor observed in Purkinje neurons. (B) Quantification of IF signal intensity from nuclear area (Nuc) divided by signal intensity from cytoplasmic area (Cyt) in CA1 neurons. Nuc/Cyt ratios (sham divided by pilocarpine signal) were obtained from 22 cells in two sham animals and 23 cells in four pilocarpine animals at the 4 hr time point. Scale bar 20 μm.
Figure 8
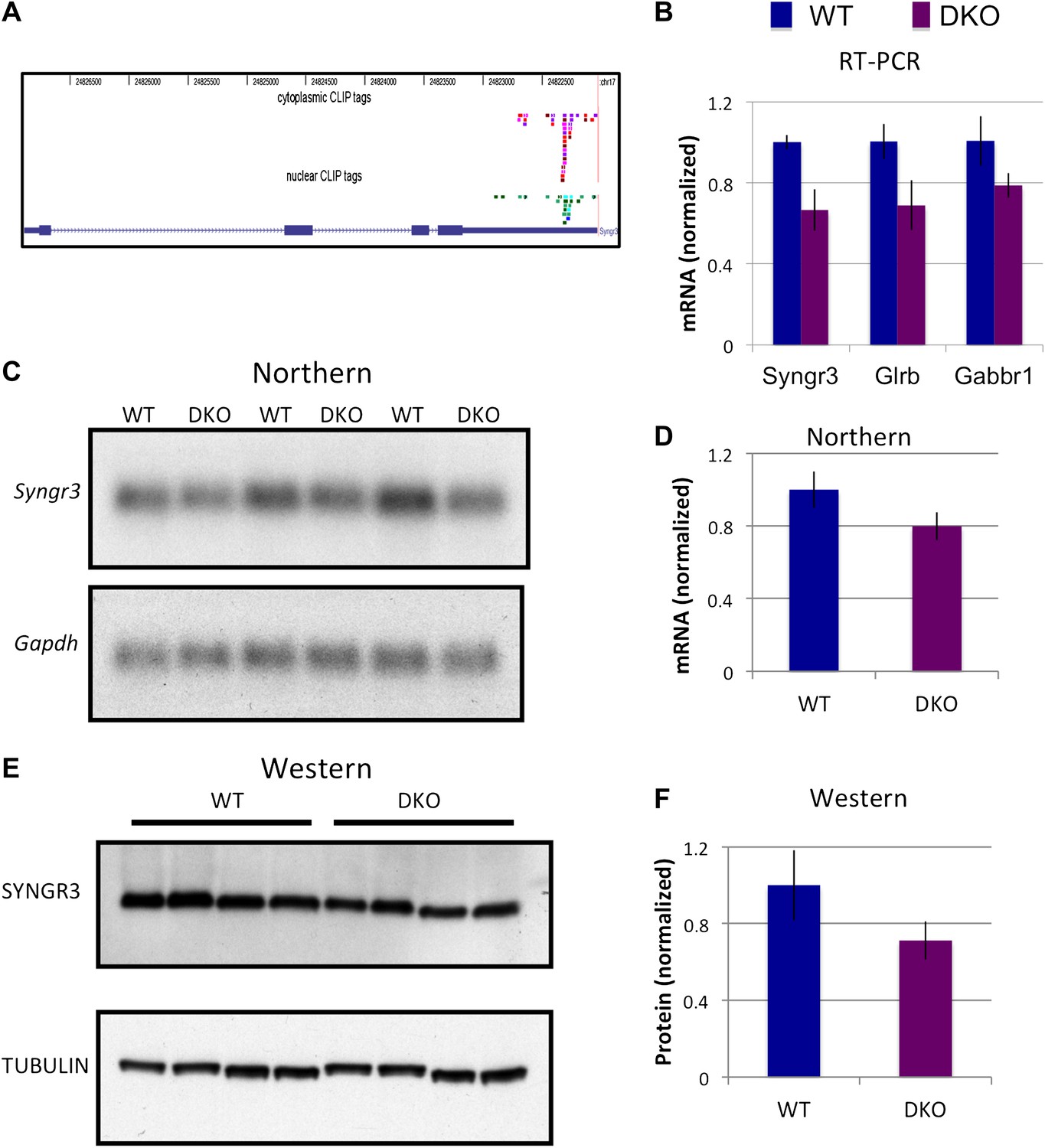
NOVA regulates the expression of synaptogyrin III (Syngr3) mRNA and protein.
(A) Location of cytoplasmic and nuclear CLIP for Syngr3. Red and purple colors represent Cytoplasmic CLIP tags and green and blue tags represent nuclear CLIP tags. All tags were located in 3′ UTR. (B) qRT-PCR data of Syngr3, Glrb (Glycine receptor b), and Gabbr1 (GABA B receptor1) showed mild reduction (about 30%) in Nova DKO brain. Y-axis represents the relative mRNA levels (WT/DKO). p<0.05. (C) Northern blot analysis of Syngr3 mRNA in WT (lane 1, 3, 5) vs DKO (lane 2, 4, 6). Gapdh probe was used as a normalizing control. (D) Quantitation of relative RNA intensity (WT/DKO) was plotted as a relative ratio of Syngr3 mRNA/Gapdh in Nova WT/DKO; error bars represent standard deviation (p<0.05); Syngr3 was reduced by about ∼ 20% in DKO brain. (E) Immunoblot analysis of SYNGR3 in WT vs DKO. Protein extracts from WT (lane 1–4) vs DKO (lane 5–8) were loaded. γ-TUBULIN is used as a normalizing control. (F) Quantitation of relative protein intensity (WT/DKO). The results were plotted as relative ratio of SYNGR3 in WT/DKO; error bars represent standard deviation (p<0.05); SYNGR3 protein was reduced ∼35% in DKO brain.
Figure 9
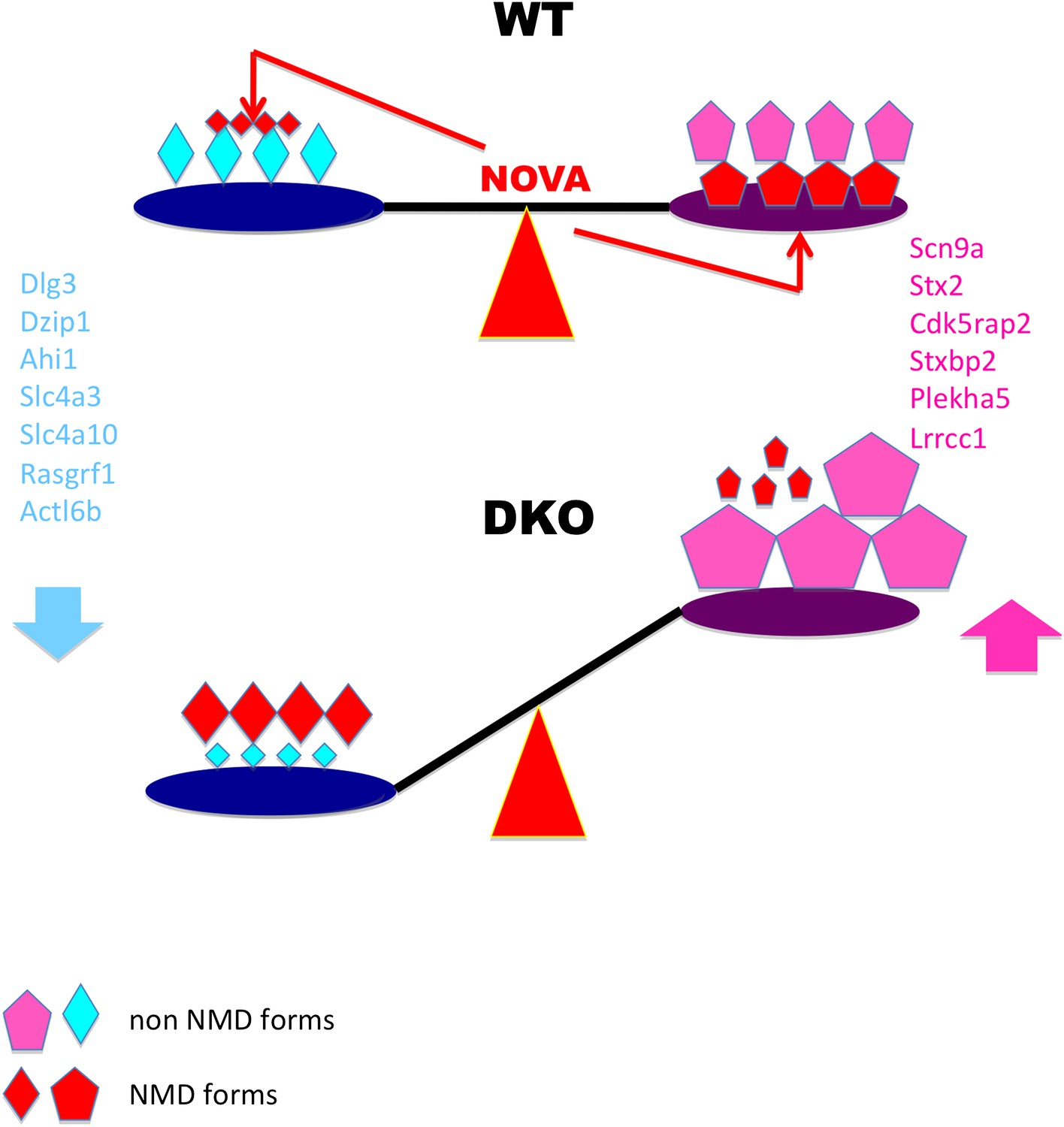
Model linking electrical activity with Nova-dependent splicing of cryptic NMD exons to maintain the balance of synaptic proteins.
In WT brain, NOVA represses some cryptic NMD isoforms (small red diamonds on left) while promoting others (large red pentagons on right), thereby maintaining the balance of protein levels. To a lesser degree, NOVA also stabilizes transcripts through 3′ UTR interactions (Figure 8). In DKO brain, the absence of NOVA disturbs this balance of protein in expression, contributing to aberrations in synaptic transmission. For example, Nova-regulation of cryptic NMD exons alters levels of the NMDA-receptor associated Dlg3 (Figures 2 and 3) and sodium channel Scn9a (Figure 4) proteins, which are implicated in familial epileptic disorders and are dynamically regulated after seizures in mice (Figure 6).
Tables
Table 1
Validation data for transcripts showing Nova-dependent changes in steady-state mRNA levels
| qRT-PCR fold | Microarray-predicted fold | p-value | Gene |
|---|---|---|---|
| Transcripts with larger predicted fold changes | |||
| 3.85 ± 0.84 | 3.39 | <0.001 | Dlg3 (Sap102) |
| 3.79 ± 0.29 | 2.73 | <0.001 | Dzip1 |
| 2.40 ± 0.43 | 2.25 | <0.001 | Dtd1 |
| 3.64 ± 0.39 | 2.23 | <0.001 | Rasgrp2 |
| 2.42 ± 0.15 | 1.97 | <0.001 | Slc4a3 |
| 1.74 ± 0.28 | 1.81 | <0.001 | Slc4a10 |
| 2.06 ± 0.3 | 1.72 | <0.001 | Cacna2d3 |
| 1.73 ± 0.14 | 1.71 | <0.001 | Ahi1 |
| 1.90 ± 0.12 | 1.68 | <0.001 | Rasgrf1 |
| 1.83 ± 0.18 | 1.67 | <0.001 | Slc1a6 |
| 1.53 ± 0.13 | 1.67 | <0.001 | Lynx1 |
| 0.50 ± 0.07 | 0.68 | <0.001 | Cdk5rap2 |
| 0.40 ± 0.02 | 0.54 | <0.001 | Stx2(syntaxin 2) |
| 0.30 ± 0.04 | 0.45 | <0.001 | Cpne3(copine III) |
| 0.33 ± 0.07 | 0.34 | <0.001 | Scn9a |
| Transcripts with smaller predicted fold changes | |||
| 1.98 ± 0.16 | 1.48 | <0.001 | Syt2 |
| 1.41 ± 0.1 | 1.47 | <0.001 | Actl6b |
| 1.74 ± 0.07 | 1.40 | <0.001 | Gria3 |
| 1.56 ± 0.09 | 1.30 | <0.001 | Syngr3* |
| 1.53 ± 0.13 | 1.29 | <0.001 | Glrb* |
| 1.33 ± 0.20 | 1.25 | <0.001 | Gabbr1* |
| 0.69 ± 0.07 | 0.77 | <0.001 | Plekha5 |
-
Summary of validation data for transcripts showing Nova-dependent change in steady-state mRNA levels. Upper table: validation data for transcripts showing larger fold predicted changes by microarray. Lower table: validation data for transcripts showing small predicted changes by microarray, to illustrate sensitivity of the data. In each table, the first column shows fold change from qRT-PCR, the second column shows fold change from Affymetrix-exon array data, the third column shows the p value and the fourth column shows the name of gene. Data for each gene is derived from three biologic replicates (three animals) and three technical replicates (nine reactions per point); these data are statistically significant (p<0.001; Student's t-test). Genes in bold are cryptic exons confirmed by sequencing (see Figure 3—source data 1 and Figure 4D). Genes with asterisk are those with robust 3′ UTR clusters (see Figure 8).
Table 2
Summary of CLIP tag mapping data
| Sample | Tag total | Tag mappable | Tag unique |
|---|---|---|---|
| Nuclear 1 | 1,504,345 | 1,031,115 | 429,527 |
| Nuclear 2 | 4,984,997 | 3,161,194 | 363,771 |
| Nuclear 3 | 7,184,066 | 3,944,912 | 214,982 |
| Nuclear 4 | 6,656,628 | 3,144,744 | 325,507 |
| Nuclear sum | 1,333,787 | ||
| Cytoplasm 1 | 1,454,697 | 975,222 | 89,463 |
| Cytoplasm 2 | 4,751,331 | 2,780,173 | 37,027 |
| Cytoplasm 3 | 3,653,600 | 2,005,202 | 36,228 |
| Cytoplasm 4 | 3,813,960 | 2,382,946 | 39,145 |
| Cytoplasmic sum | 201,863 |
Table 3
Splicing changes in brain after pilocarpine-induced seizures
| |ΔI t-test| ≥ 5.6 (p<0.005) | ||
|---|---|---|
| 4 hr | Total | Validated |
| |ΔI t-test| ≥ 0.4 | 7 | 2 |
| |ΔI t-test| ≥ 0.3 | 21 | 5 |
| |ΔI t-test| ≥ 0.2 | 117 | 22 |
| |ΔI t-test| ≥ 8.6 (p<0.001) | ||
|---|---|---|
| 4 hr | Total | Validated |
| |ΔI t-test| ≥ 0.4 | 7 | 2 |
| |ΔI t-test| ≥ 0.3 | 18 | 5 |
| |ΔI t-test| ≥ 0.2 | 96 | 22 |
| |ΔI| ≥ 0.15 | ||
|---|---|---|
| 24 hr | Total | Validated |
| |ΔI| ≥ 0.3 | 3 | 1 |
| |ΔI| ≥ 0.2 | 23 | 6 |
| |ΔI| ≥ 0.15 | 56 | 6 |
-
Exon junction array results assessing splicing changes in the hippocampus after pilocarpine seizure, compared to sham controls. The total number of alternative exons identified from the brains of pilocarpine treated animals from which RNA was analyzed by genome-wide exon junction arrays is shown. The data in is organized as a function of differing stringency thresholds (as previously described: ΔI, a measure of the inclusion of exons in sham relative to pilocarpine treated animals; ΔI was determined using the ASPIRE algorithm (Ule et al., 2005b) and ΔI t-test using ASPIRE2 (Licatalosi et al., 2008)). Each data point represents analysis of all results from three pairs of biological replicates. Validated refers to independent qRT-PCR validation (see Supplementary file 1) of selected transcripts from total.
Table 4
GO analysis
| Term | Count | % | p-Value | Fold | Benjamini |
|---|---|---|---|---|---|
| Biological process | |||||
| Transport | 57 | 27.67% | 1.40 × 10−08 | 2.10779601 | 7.27 × 10−05 |
| Establishment of localization | 57 | 27.67% | 3.74 × 10−08 | 2.0512741 | 9.71 × 10−05 |
| Localization | 60 | 29.13% | 1.46 × 10−07 | 1.91908255 | 2.53 × 10−04 |
| Ion transport | 25 | 12.14% | 1.50 × 10−06 | 3.08118005 | 0.001943 |
| Synaptic transmission | 12 | 5.83% | 1.16 × 10−05 | 5.49530653 | 0.011935 |
| Chloride transport | 7 | 3.40% | 4.92 × 10−05 | 10.5684476 | 0.041641 |
| Transmission of nerve impulse | 12 | 5.83% | 6.38 × 10−05 | 4.56499873 | 0.04624 |
| Exocytosis | 8 | 3.88% | 6.71 × 10−05 | 7.88782093 | 0.042589 |
| Cation transport | 17 | 8.25% | 1.41 × 10−04 | 3.04192354 | 0.07808 |
| Metal ion transport | 15 | 7.28% | 1.72 × 10−04 | 3.30910296 | 0.085535 |
| Cell–cell signaling | 13 | 6.31% | 4.67 × 10−04 | 3.37670829 | 0.197738 |
| Cellular component | |||||
| Synapse | 19 | 9.22% | 1.69 × 10−10 | 7.12724905 | 1.33 × 10−07 |
| Postsynaptic membrane | 12 | 5.83% | 3.96 × 10−08 | 9.73280098 | 1.55 × 10−05 |
| Synapse part | 12 | 5.83% | 1.25 × 10−07 | 8.71242669 | 3.27 × 10−05 |
| Cytoplasmic part | 66 | 32.04% | 1.11 × 10−06 | 1.75847144 | 2.17 × 10−04 |
| Cell junction | 18 | 8.74% | 2.18 × 10−06 | 4.05127841 | 3.42 × 10−04 |
| Cytoplasm | 91 | 44.17% | 1.52 × 10−05 | 1.44439091 | 0.00198 |
| Plasma membrane | 44 | 21.36% | 5.11 × 10−05 | 1.85625586 | 0.005706 |
| Molecular function | |||||
| Ion transmembrane transporter activity | 30 | 14.56% | 1.74 × 10−10 | 4.09234024 | 4.70 × 10−07 |
| Transporter activity | 42 | 20.39% | 4.86 × 10−10 | 2.91474846 | 6.58 × 10−07 |
| Substrate-specific transmembrane transporter | 31 | 15.05% | 1.15 × 10−09 | 3.65758191 | 1.04 × 10−06 |
| Transmembrane transporter activity | 33 | 16.02% | 2.22 × 10−09 | 3.36413152 | 1.50 × 10−06 |
| Substrate-specific transporter activity | 33 | 16.02% | 1.85 × 10−08 | 3.08048906 | 9.99 × 10−06 |
| Gated channel activity | 17 | 8.25% | 3.63 × 10−08 | 5.82250711 | 1.64 × 10−05 |
| Ligand-gated channel activity | 11 | 5.34% | 5.03 × 10−08 | 11.1421945 | 1.70 × 10−05 |
| Ligand-gated ion channel activity | 11 | 5.34% | 5.03 × 10−08 | 11.1421945 | 1.70 × 10−05 |
| Anion transmembrane transporter activity | 12 | 5.83% | 6.01 × 10−08 | 9.36542127 | 1.81 × 10−05 |
| Ion channel activity | 18 | 8.74% | 2.53 × 10−07 | 4.7344533 | 6.84 × 10−05 |
| Substrate specific channel activity | 18 | 8.74% | 3.85 × 10−07 | 4.59483135 | 9.47 × 10−05 |
| GO term analysis (top increase in DKO 14 genes) | |||||
| Biological process | |||||
| Vesicle-mediated transport | 3 | 21.43% | 0.02265 | 11.0206034 | 35.4494 |
| Molecular function | |||||
| Binding | 11 | 78.57% | 0.031126 | 1.41461519 | 1 |
| Transporter activity | 4 | 28.57% | 0.045036 | 4.34057779 | 1 |
| Protein binding | 9 | 64.29% | 0.005192 | 2.31262748 | 0.999999 |
-
GO (gene ontology) term analysis of transcripts up/down-regulated in WT vs DKO (see Figure 1A). Data of upper table are derived from top 215 down-regulated transcripts in DKO; (positive value in Figure 1A). Data of lower table are from top 14 up-regulated transcripts in DKO; (negative value in Figure 1A). Taken together, two tables suggested that NOVA regulates the levels of many transcripts relating to synaptic transmission (Dennis et al., 2003; Huang et al., 2009).
Additional files
-
Supplementary file 1
List of validated targets that were alternatively spliced after pilocarpine induced status epilepticus.
Targets were grouped according to their biological process and molecular function. Calculated delta I (ΔI) was listed for both the hippocampus (Hp) and cortex (Ctx) at 4 hr and 24 hr time points. ΔI from exon junction arrays are listed; other exons were empirically examined. Total fold change (TFC) of transcript level was calculated as the signal from status epilepticus divided by the signal of sham in the hippocampus. Alternative splice (AS) type is also listed, with abbreviations as follows: CASS: cassette exon; ALT5: alternative 5′ splice sites; ALT3: alternative 3′ splice sites; MUTX: mutually exclusive exons; IRET: retained intron; APA3: alternative polyA usage coupled with 3′ splice site selection; ALTP: alternative promoter; TACA: tandem cassette exons.
- https://doi.org/10.7554/eLife.00178.028
-
Supplementary file 2
PCR primers used in this work. All PCR primers are shown, oriented 5′ to 3′.
- https://doi.org/10.7554/eLife.00178.029
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
NOVA-dependent regulation of cryptic NMD exons controls synaptic protein levels after seizure
eLife 2:e00178.
https://doi.org/10.7554/eLife.00178




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}