Mitochondrial fusion but not fission regulates larval growth and synaptic development through steroid hormone production
- Baylor College of Medicine, United States
- Howard Hughes Medical Institute, Baylor College of Medicine, United States
- Texas Children's Hospital, United States
Figures
Figure 1 with 1 supplement
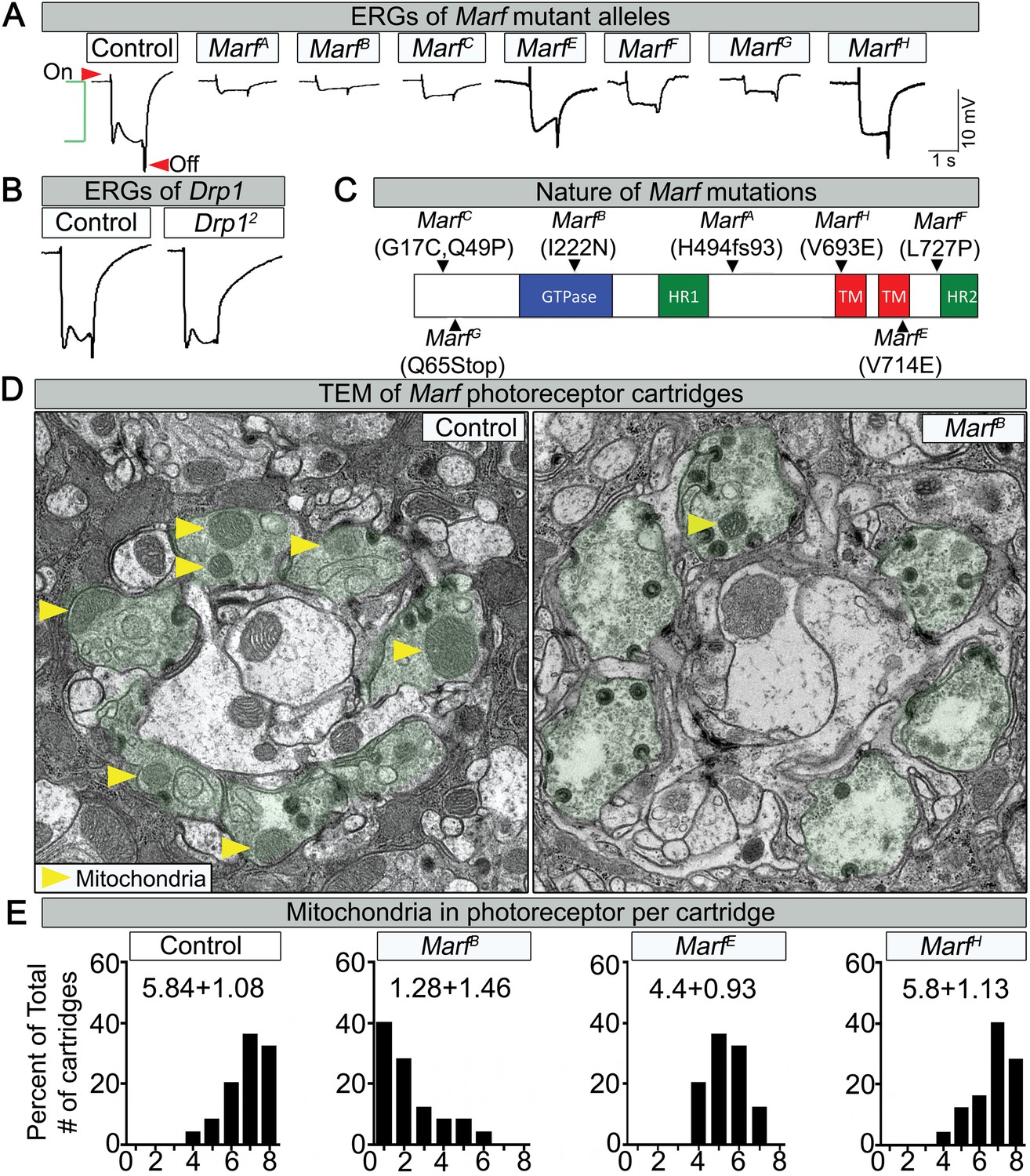
Loss of Marf impairs phototransduction and affects mitochondrial localization to photoreceptor terminals.
(A) Electroretinograms (ERGs) of 1 day old ey-FLP mutant clones of 7 different Marf mutants or isogenized wild type clones (Control). ERGs of Marf mutant alleles and control flies. A typical ERG trace is comprised of an on-transient (red arrow), a depolarization (green bracket) and an off-transient (red arrow). (B) ERGs of Drp1 mutants and control flies. (C) Marf protein domains and localization of EMS-induced mutations of the seven Marf mutant alleles identified by sequencing. H494fs93 = insertion of an A at nucleotide codon for amino acid H494 that generates 93 new amino acids followed by a premature stop codon. TM = transmembrane domain. HR = heptad repeat. (D) TEM sections of a cartridge containing fly photoreceptor terminals (green shading). Marf mutant photoreceptor terminals display reduced number and size of mitochondria (yellow arrow heads) compared to Marf-genomic rescue controls. (E) Quantification of total mitochondria number per cartridge in Marf mutants and Marf-genomic rescue photoreceptor terminals (Control). 50 cartridges per genotype.
Figure 1—figure supplement 1
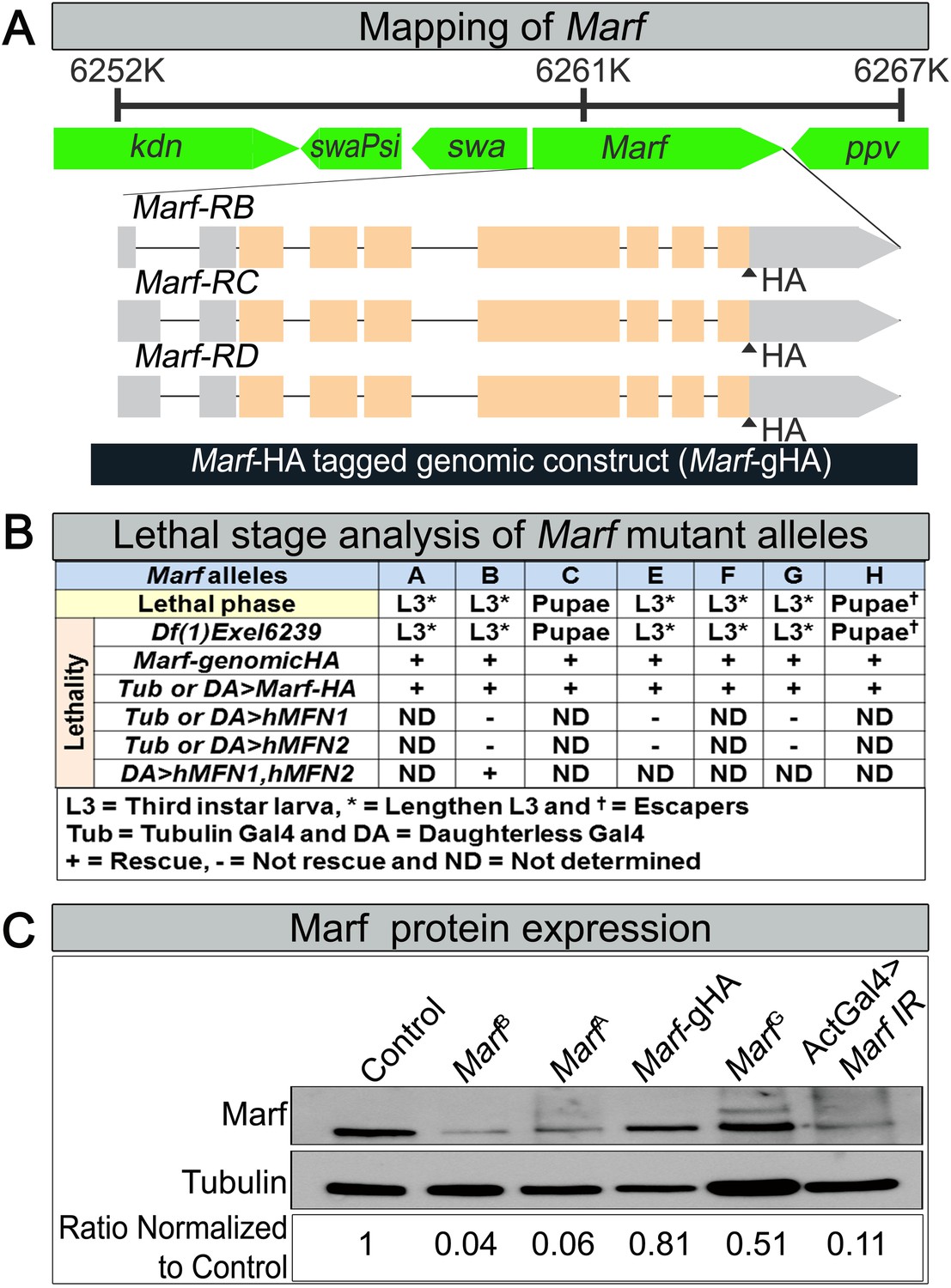
Mapping, lethal staging and Marf protein expression of Marf mutant alleles.
(A) For mapping of Marf, the lethality of all Marf alleles were rescued by large duplication Dp(1;Y)dx[+]5,y[+]/C(1)M5 (4C11;6D8 + 1A1;1B4) covering the Marf locus. A 6.1 kb genomic rescue fragment encompassing the Marf locus was used to generate a Marf-HA tagged genomic construct (Marf-gHA) to rescue the Marf alleles. (B) Lethal staging analysis of Marf mutant alleles and lethality rescue by Marf-gHA, UAS-Marf-HA, UAS-MFN1, UAS-MFN2 and UAS-MFN1/UAS-MFN2 cDNA constructs. (C) Marf Western blot (Ziviani et al., 2010) from MarfB, MarfA, MarfG, Marf-Genomic (Marf-gHA) and ubiquitous (Actin-Gal4) Marf knockdown in third instar larvae.
Figure 2
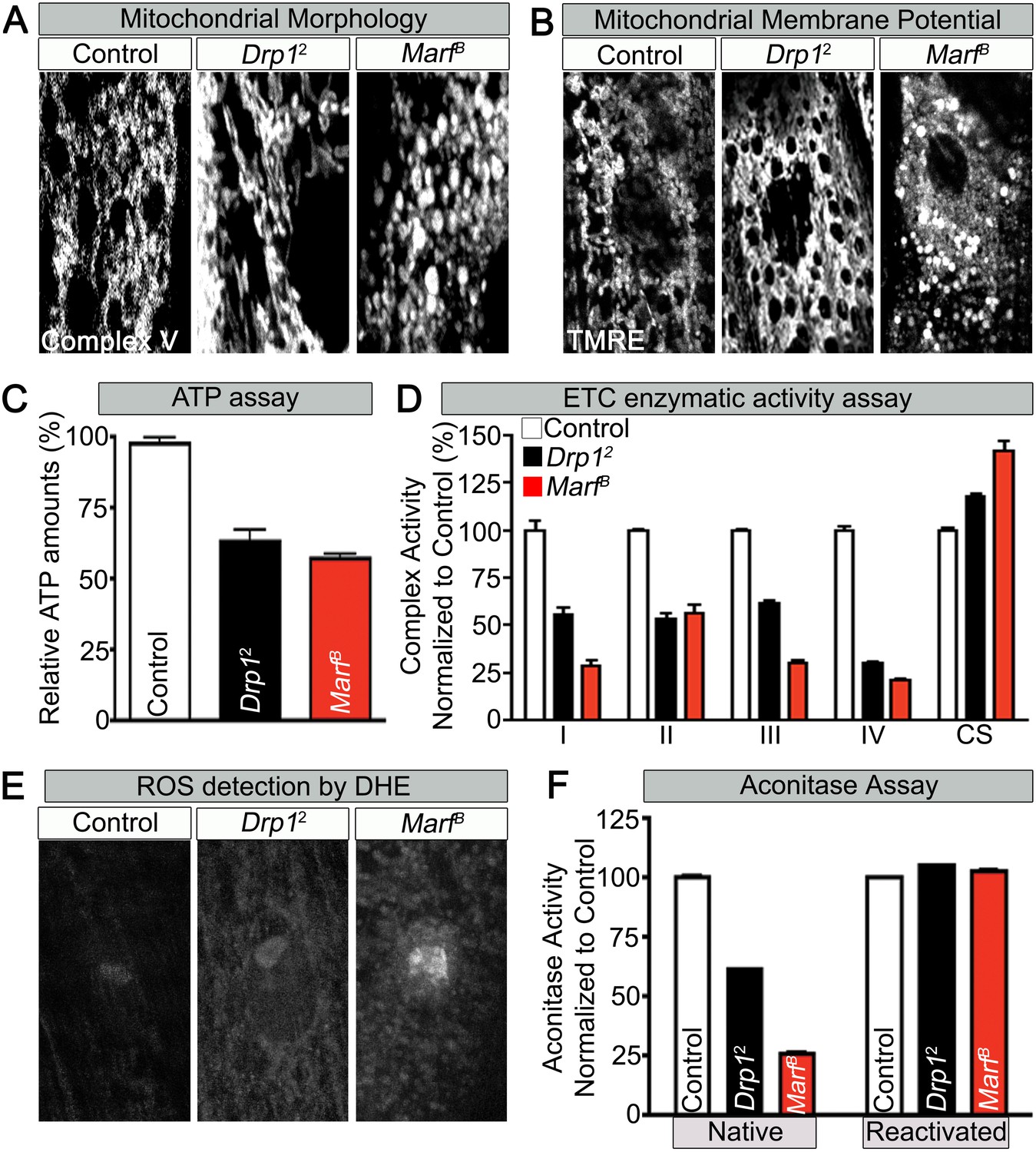
Mitochondrial morphology and function in Marf and Drp1 mutants.
(A) Mitochondrial morphology based on anti-Complex V antibody staining (Complex V) in larval muscles (Zoom in view around muscle nucleus). (B) Mitochondrial membrane potential as measured by the TMRE dye in larva muscle. (C) Relative ATP amounts. (D) Measurement of the enzymatic activity of electron transport chain (ETC) complexes (I–IV) from purified mitochondria from third instar larvae. All the ETC activities were normalized to citrate synthase (CS) activity of controls. (E and F) ROS is measured by two methods: (E) by DHE staining in larval muscles and (F) by measuring aconitase activity reduction from purified mitochondria. Reducing reagents reactivate native aconitase. Aconitase activities were normalized to controls. (C, D and F) error bars represent ± SEM.
-
Figure 2—source data 1
Lethal staging of Drp1 mutants.
Lethal staging of Drp1 transheterozygous combinations of Drp1KG38015, Drp1[T26] and Drp1 with Drp12 mutant alleles.
- https://doi.org/10.7554/eLife.03558.006
-
Figure 2—source data 2
Phenotypic comparison of Marf, Drp1 and Marf and Drp1 mutants.
Phenotypic comparison table of Marf, Drp1 and Marf and Drp1 mutants in mitochondria morphology, mitochondria membrane potential (MMP), ATP levels, ROS (DHE) intensity, bouton numbers and 20-hydroxyecdysone (20E) levels. Figure 2B MarfB panel has both puncta globular (P) and non-puncta (NP) staining that were both used to measure MMP. MMP, ATP levels and ROS intensity were normalized to controls and all columns are representative of three independent experiments ± SEM.
- https://doi.org/10.7554/eLife.03558.007
Figure 3 with 2 supplements
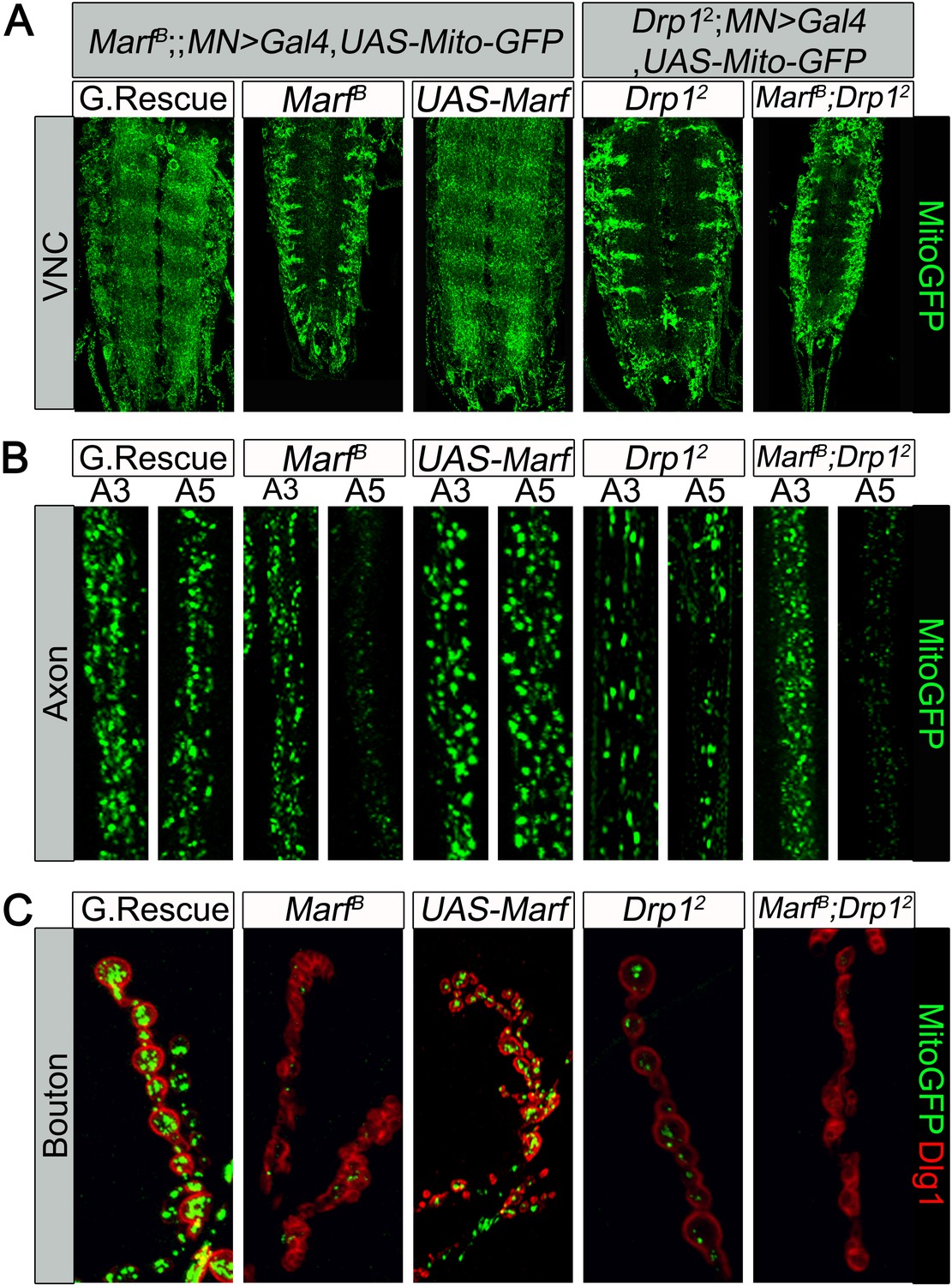
Mitochondrial trafficking defects in distal axons and boutons.
Mutations and controls were crossed to a motor neuron driver (D42-GAL4, UAS-MitoGFP) to label neuronal mitochondria. (A) Ventral nerve cord (VNC): Marf and Drp1 mutants exhibit clustered mitochondria in the soma. (B) Comparison of a proximal axonal segment in A3 and a distal segment in A5. Distal segments of A5 axons in Marf mutants contain many fewer mitochondria than proximal segments. (C) Marf mutants contain almost no mitochondria in boutons when co-stained with post-synaptic marker Discs Large 1 (Dlg1). Percentage of boutons with no mitochondria: Genomic rescue (0%), Marf B (89%), UAS-Marf (0%), Drp12 (36%) and Marf B;Drp12 (95%).
Figure 3—figure supplement 1
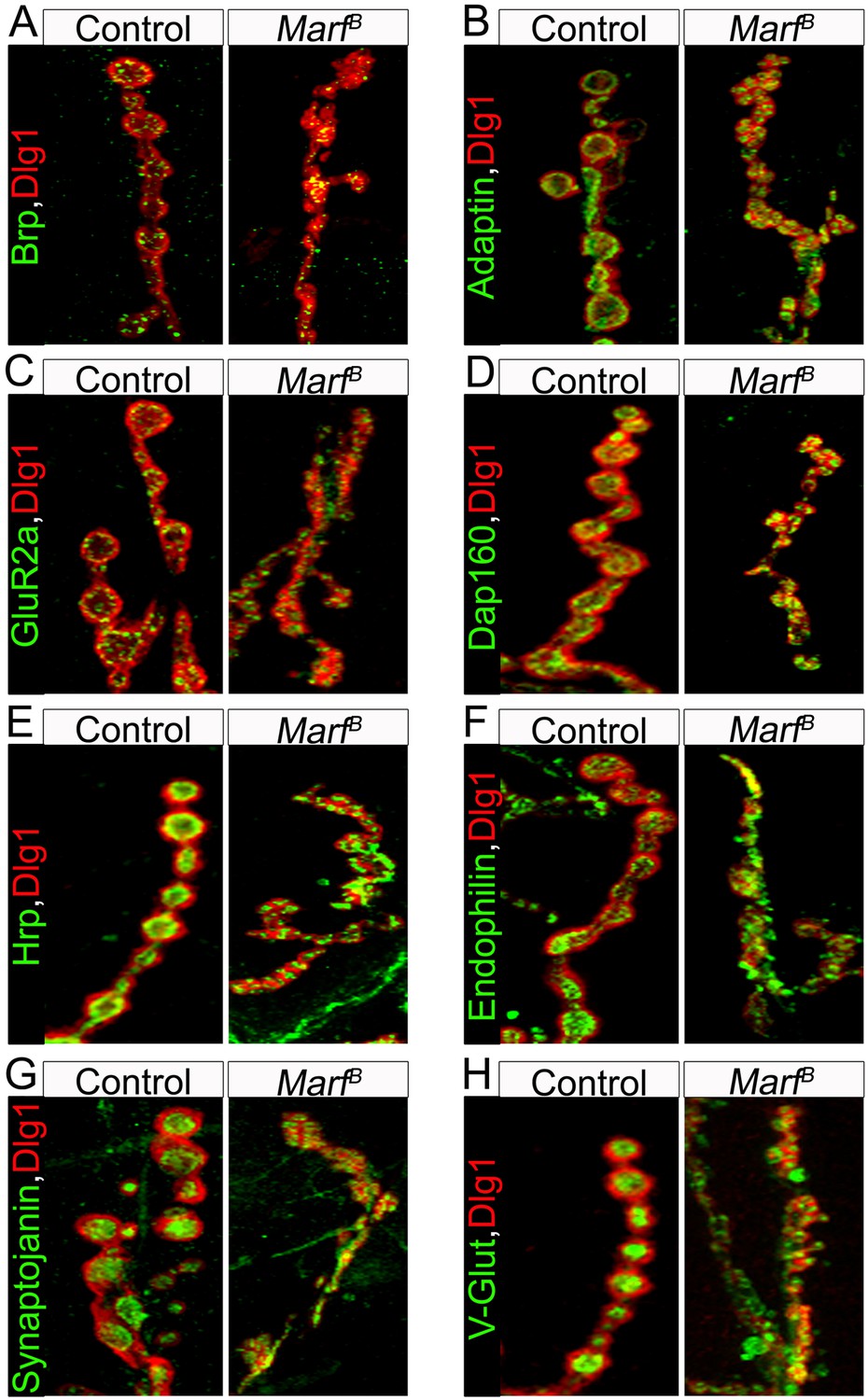
Pre-synaptic, endocytic and postsynaptic markers are present in Marf mutant boutons.
A panel of different NMJ markers co-stained with Dlg1: (A) Bruchpilot (Brp), (B) α-Adaptin, (C) Glutamate receptor IIa (GluRIIa), (D) Dap160, (E) Hrp, (F) Endophilin, (G) Synaptojanin and (H) Drosophila vesicular glutamate transporter (DV-Glut).
Figure 3—figure supplement 2
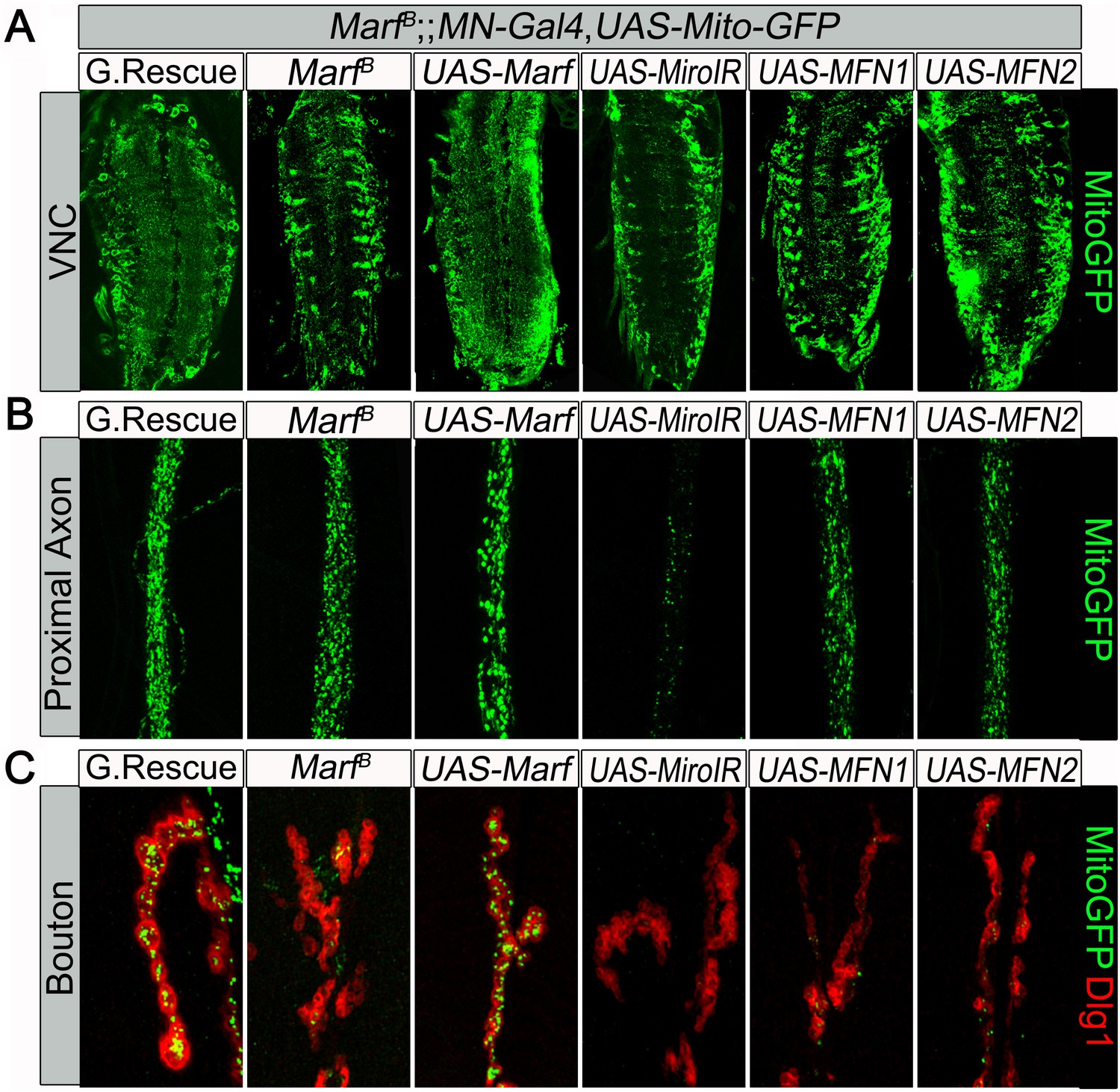
Mitochondrial trafficking defect in Marf mutants cannot be rescued by motor neuron expression of human MFN1 or MFN2.
Mutations and controls were crossed to a motor neuron (MN) driver (D42-GAL4, UAS-mitoGFP) to label neuronal mitochondria. (A) Ventral nerve cord (VNC), MN-knockdown of dmiro in Marf mutant exhibit more clustered mitochondria in the soma compared to Marf alone, while neither MN-expression of MFN1 or MFN2 rescued the VNC mitochondrial trafficking defect of Marf mutants. (B) At the proximal end of the A3 axon, MN-knockdown of dmiro in Marf mutants had severed reduction of mitochondrial trafficking compared to Marf alone. (C) Neither MN-expression of MFN1 or MFN2 rescued the mitochondrial trafficking defect of Marf mutants in boutons co-stained with post-synaptic marker Discs Large 1 (Dlg1).
Figure 4
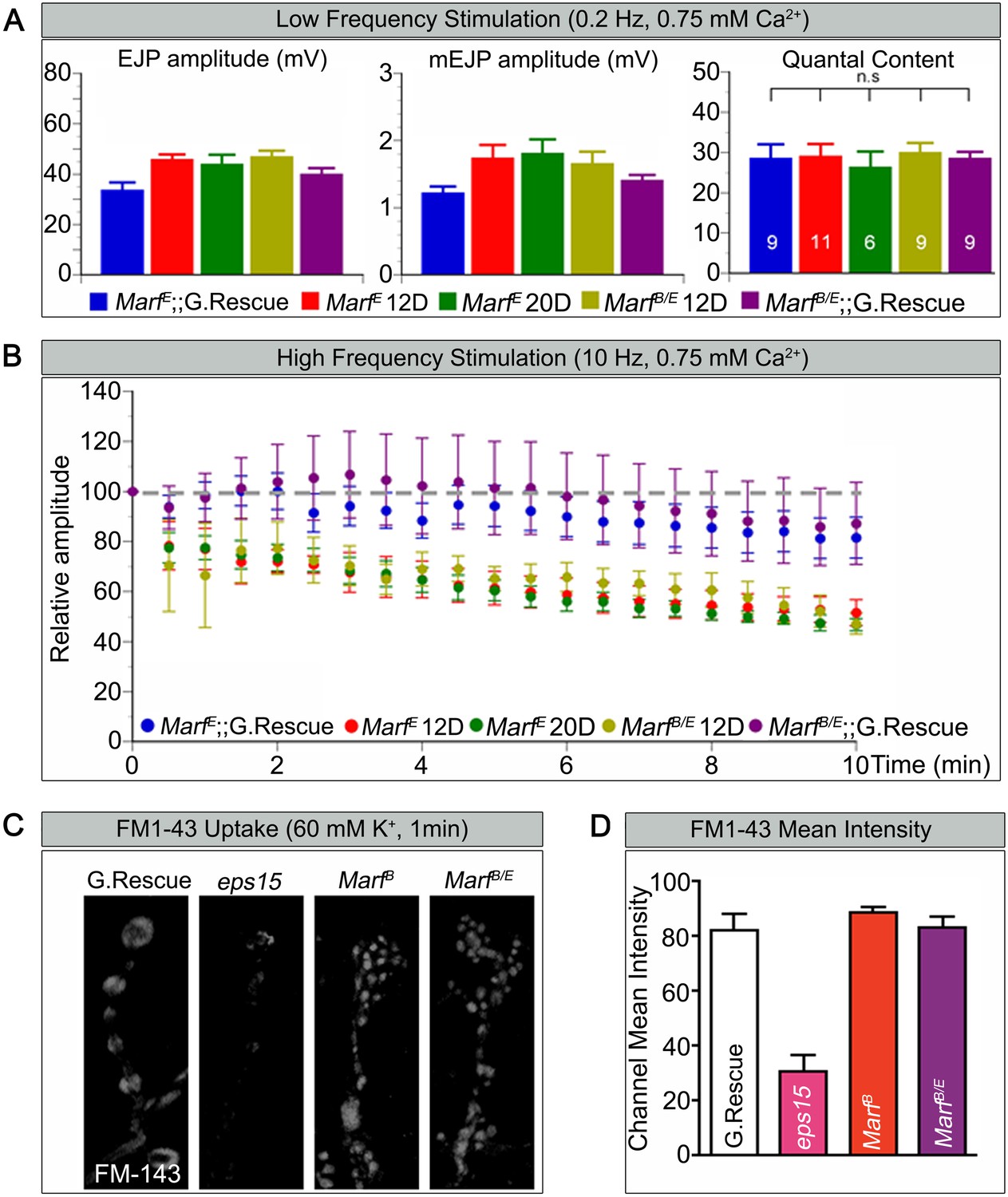
Marf is required to maintain synaptic transmission upon repetitive stimulation.
(A) Excitatory Junctional Potentials (EJP) and miniature EJPs (mEJP) measured at 0.2 Hz in 0.75 mM Ca2+ are similar in Marf mutants (day 12 or day 20 old larvae) and controls. Hence, quantal content in Marf mutants is also similar to controls (n = 6–11 larvae assayed). (B) Controls display facilitation whereas Marf mutants (day 12 or day 20 old larvae) show a rundown at 10 Hz in 0.75 mM Ca2+. (C) Assessing endocytosis using FM-143 dye uptake at 60 mM [K+] for 1 min shows no obvious differences between wild type controls and Marf mutants. (D) Quantification of FM-143 uptake. Error bars represent ± SEM.
Figure 5 with 1 supplement
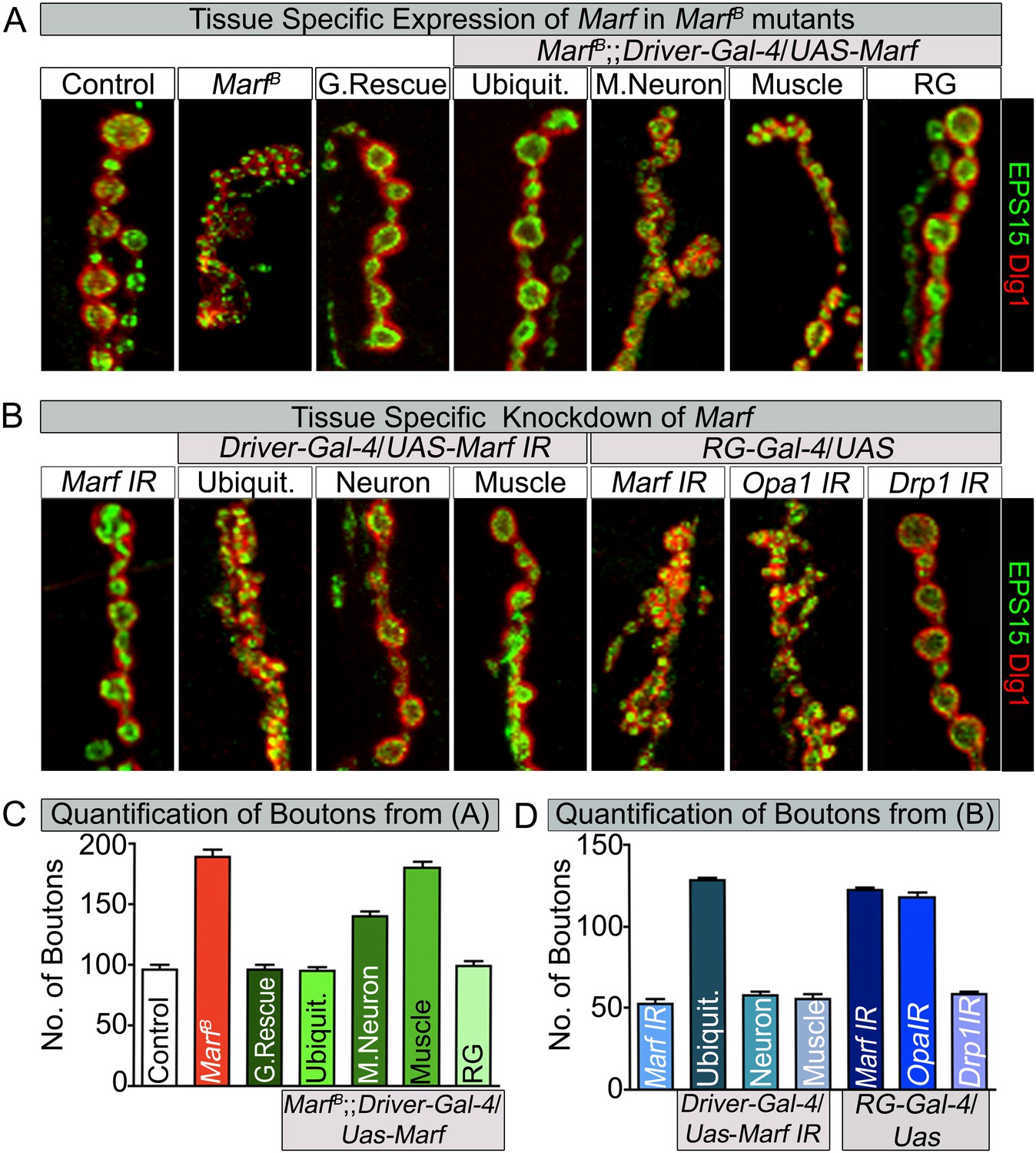
Loss of mitochondrial fusion but not fission in the ring gland results in altered bouton morphology.
Third instar larvae NMJs from muscles 6/7 segments A3 were stained with pre-synaptic (EPS15) and post-synaptic (Dlg1) markers. (A) Ubiquitous (Tubulin-Gal4) or ring gland (RG, Feb36-Gal4) expression of Marf rescue bouton morphology in Marf mutants, while motor neuron (D42-Gal4) or muscle (Mef-Gal4) Marf expression did not. (B) Ubiquitous or RG specific knockdown of Marf or Opa1 (Poole et al., 2010) phenocopy the bouton phenotype in Marf mutants while knockdown of Drp1 (Drp1 IR knockdown of Drp1 mRNA is 82% using ubiquitous driver Actin-Gal4) did not. (C and D) Quantification of bouton numbers from three independent experiments. Error bars represent ± SEM.
-
Figure 5—source data 1
Tissue specific Gal4 screen to assess rescue of lethality and bouton morphology by Marf expression.
Tissue specific Gal4 screen using UAS-Marf to assess rescuing ability of the Marf mutant lethal stage and bouton morphology phenotypes. Ubiquitous expression of Marf resulted in rescue of both lethality and bouton phenotype in Marf mutant, while RG specific expression of Marf rescues the Marf mutant bouton phenotype.
- https://doi.org/10.7554/eLife.03558.013
-
Figure 5—source data 2
Tissue specific Gal4 screen to assess lethality and alterations to bouton morphology by Marf knockdown.
Tissue specific Gal4 screen using Marf IR for phenocopying Marf mutant lethal stage and bouton morphology phenotypes. Ubiquitous knockdown of Marf resulted in both prolonged third instar larval stage and similar Marf mutant bouton phenotype, while RG specific knockdown of Marf phenocopied the Marf mutant bouton phenotype.
- https://doi.org/10.7554/eLife.03558.014
Figure 5—figure supplement 1
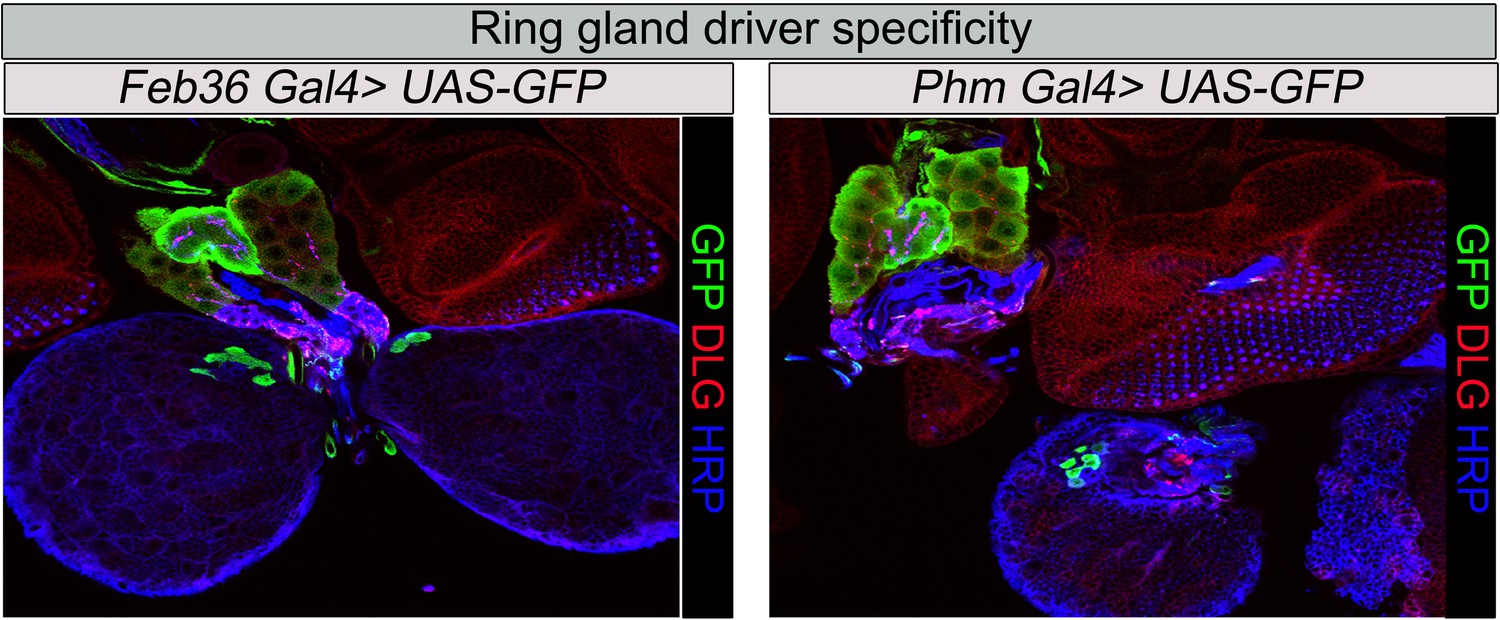
Ring gland drivers tissues specificity.
Specificity of RG driver expression used in this study: Feb36 or Phantom (Phm), (Mirth et al., 2005) Gal4 expression of UAS-GFP. Third instar larval RGs were stained with anti-GFP antibody, anti-HRP (presynaptic marker), and anti Dlg1 (post synaptic marker).
Figure 6 with 1 supplement
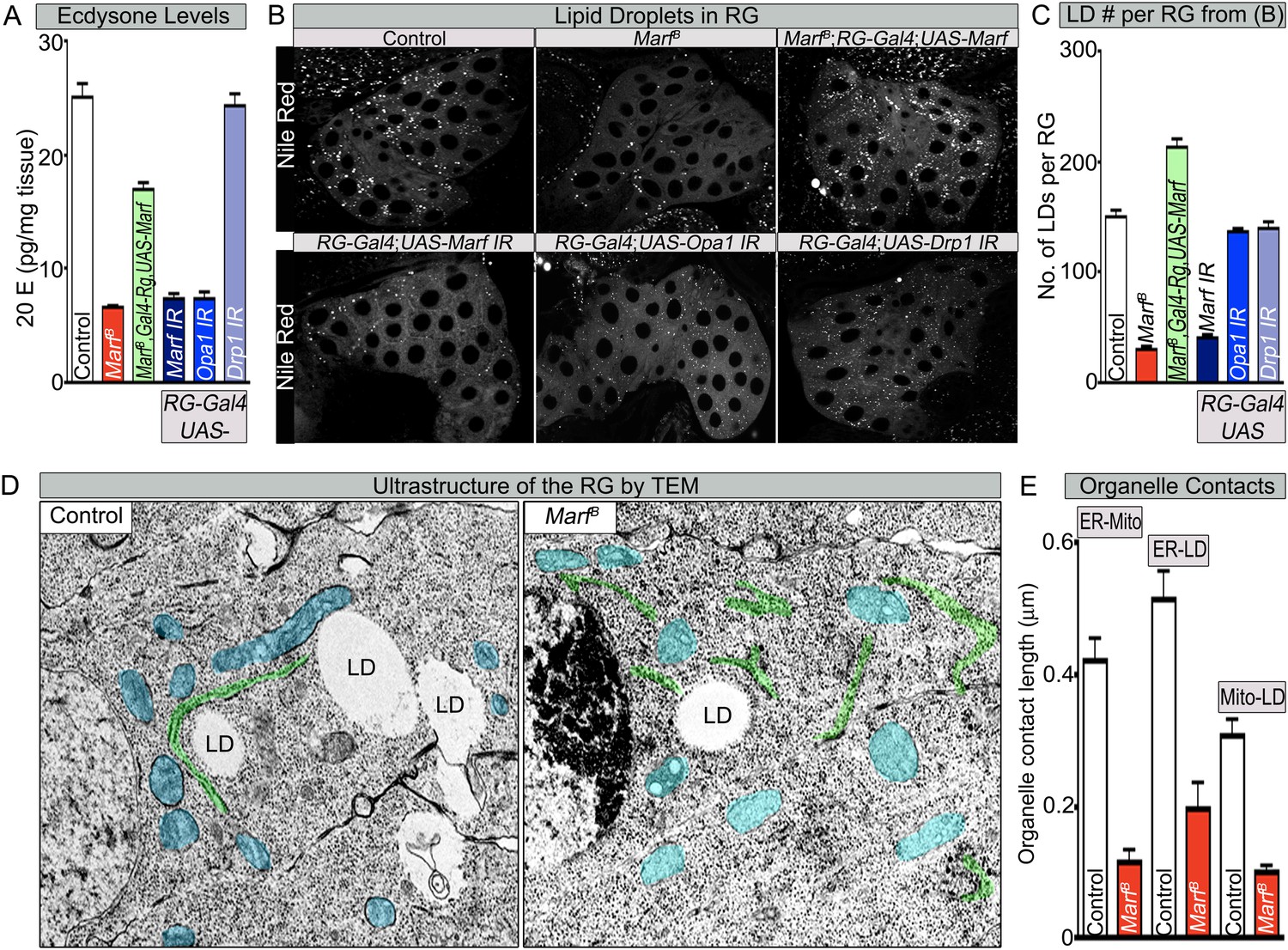
Both Marf and Opa1 regulate ecdysone synthesis in the ring gland, but only Marf promotes lipid droplet formation.
(A) Both loss of Marf and Opa1 in the RG have reduced 20-hydroxyecdysone (20E) levels when compared to loss of Drp1 and controls. 20E levels are determined and normalized by weight. (B) Only loss of Marf in the RG results in reduced lipid droplets (LDs) when stained by Nile Red compared to loss of Opa1 or Drp1. (C) Quantification of LDs in the ring gland (RG) from three independent experiments. (D) TEM sections of RG were the ER is labeled in green, mitochondria in blue and lipid droplets are labeled ‘LD’. Marf mutants display increased ER fragmentation and reduced numbers of LDs when compared to Marf-genomic rescue control animals. (E) Marf mutants have reduced contact length between mitochondria and ER, ER and LD, and mitochondria and LD when compared to controls. Error bars represent ± SEM.
Figure 6—figure supplement 1
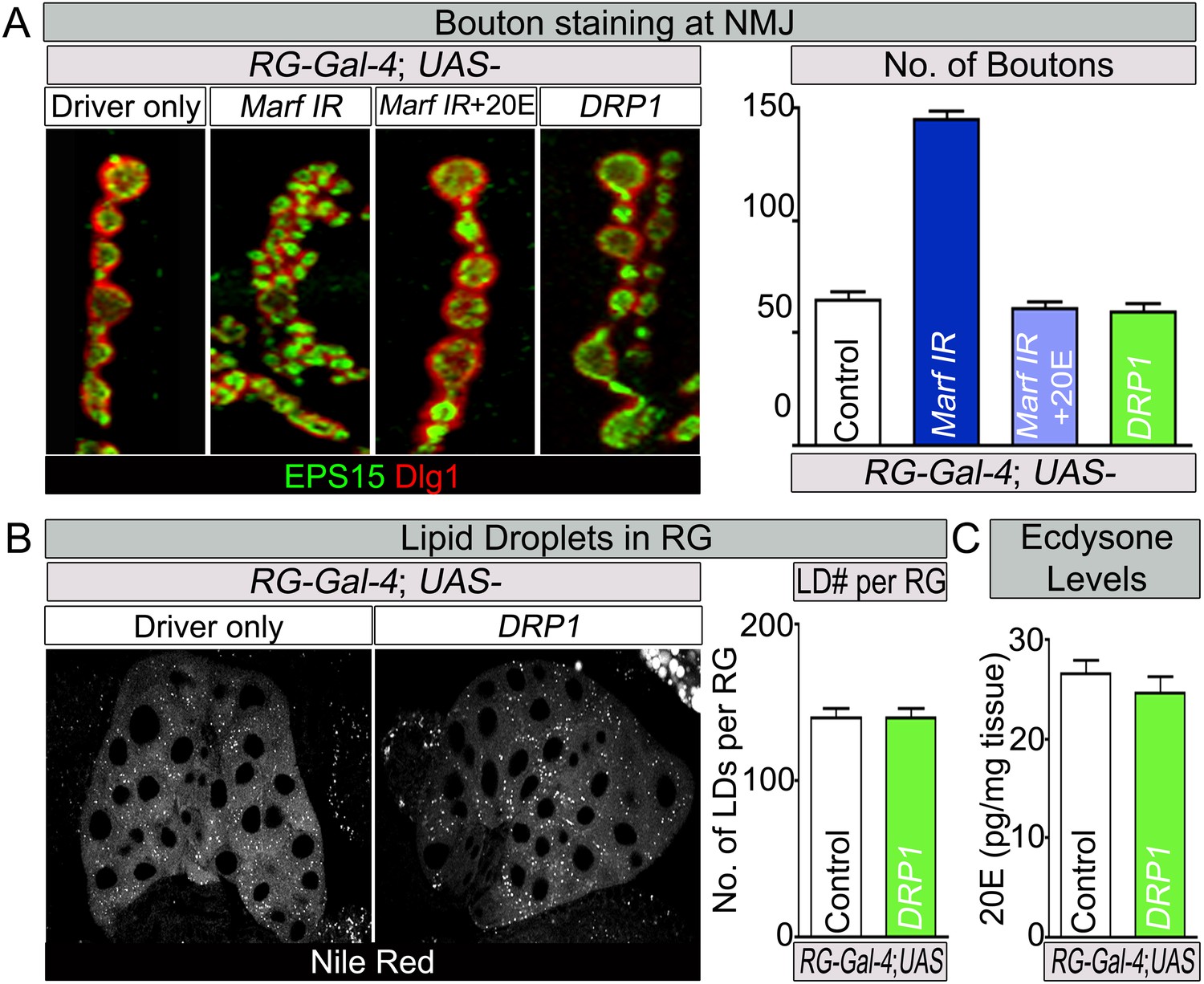
Feeding of 20E rescues the NMJ morphology of RG specific knockdown of Marf.
(A) Third instar larvae with a RG (Feb36-Gal4) specific knock down of Marf were fed either 20E (0.5 mM) or solvent (60% ethanol). NMJs from muscles 6/7 segments A3 were stained with pre-synaptic (EPS15) and post-synaptic (Dlg1) markers. Quantification of bouton numbers from three independent experiments. Expression of DRP1 in RGs (Deng et al., 2008) does not affect the NMJs. (B) Expression of DRP1 in RGs (Deng et al., 2008) does also not affect lipid droplets (LDs) numbers when stained by Nile Red and (C) 20E levels. Quantification of bouton numbers and 20E levels Error bars represent ± SEM.
Figure 7 with 2 supplements
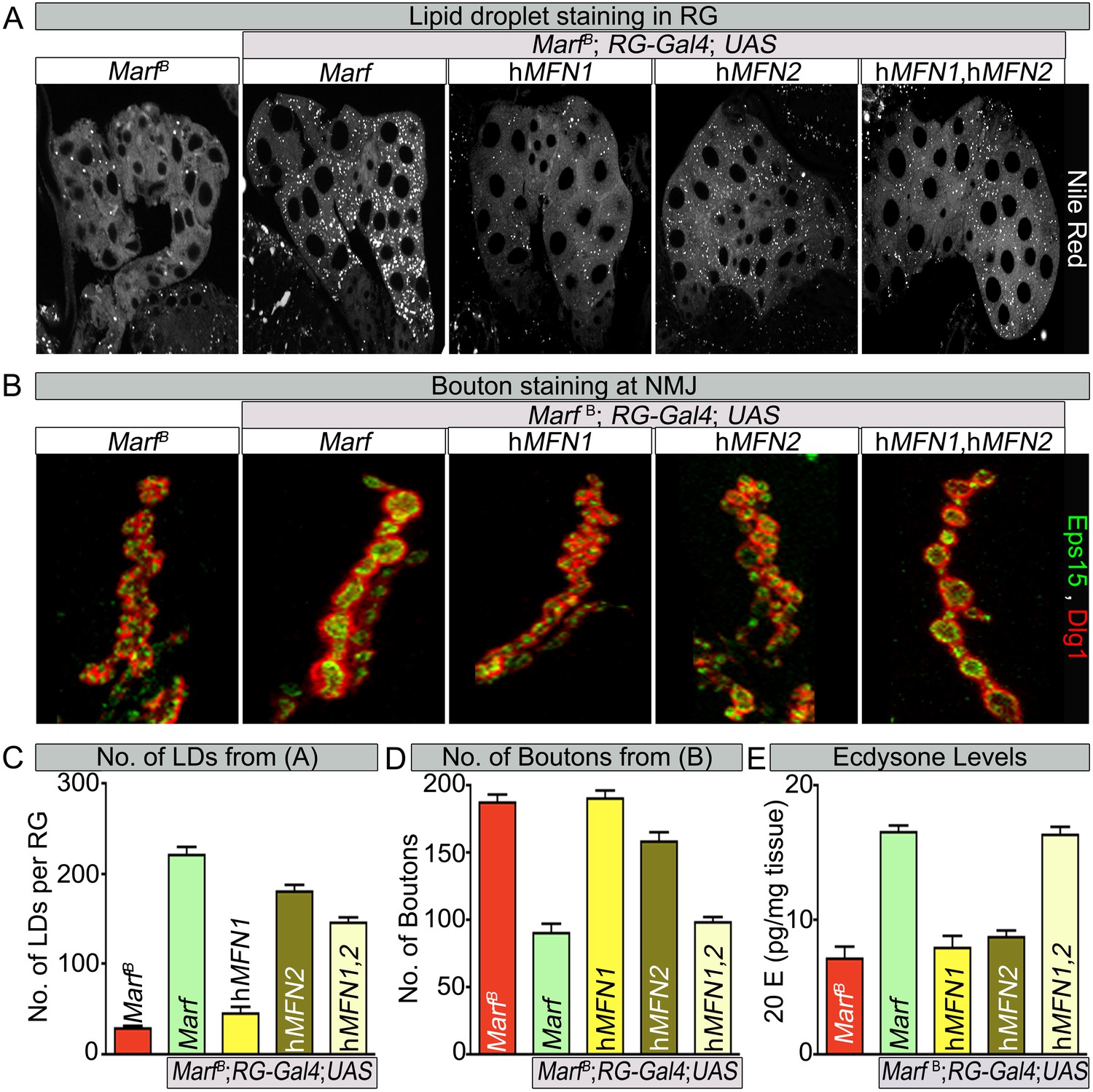
Human MFN2 restores LD numbers but both human MFN1 and MFN2 are required for steroid-hormone production in the ring glands.
(A) Rescue of lipid droplets numbers stained by Nile Red in Marf ring glands (RG) by MFN2 and MFN1/MFN2 co-expression, but not MFN1. (B) Rescue of Marf bouton morphology by expressing MFN1/MFN2 in RGs (Feb36-Gal4). Expression of MFN1 or MFN2 alone does not rescue the phenotype. (C–E) Quantification in control and Marf mutants for: (C) LDs (D) Boutons and (E) Ecdysone (20E levels) as described in Figures 5 and 6. Error bars represent ± SEM.
Figure 7—figure supplement 1
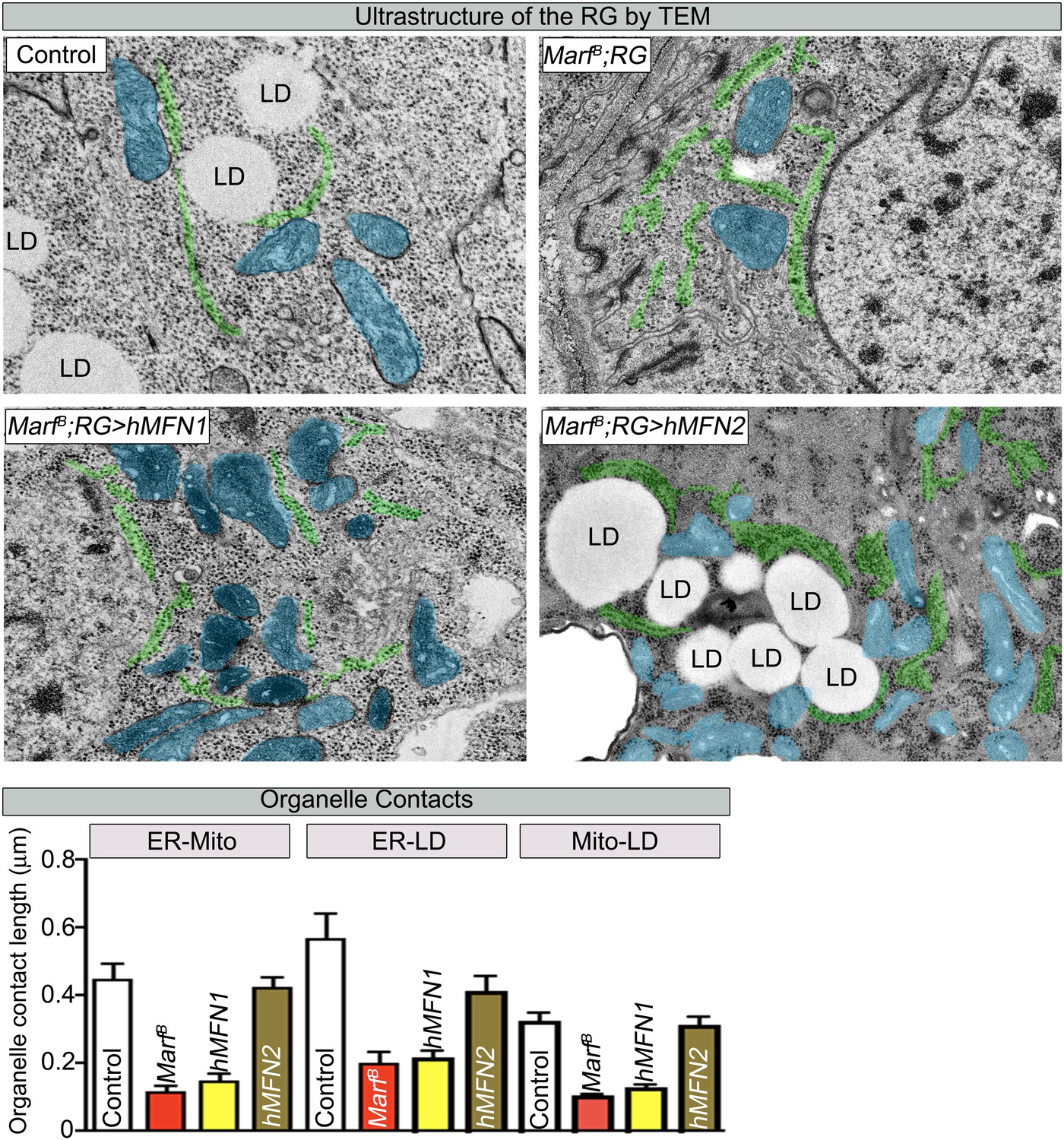
RG expression of human MFN2 restores organelle contact lengths in Marf mutants.
TEM sections of RGs that express human MFN1 or MFN2. The ER is labeled in green, mitochondria in blue and lipid droplets are labeled ‘LD’. Marf mutants with RG expression of human MFN2 display increased LD droplets and organelle contact lengths when compared to Marf mutants or Marf mutants with RG expression of human MFN1 animals. Error bars represent ± SEM.
Figure 7—figure supplement 2
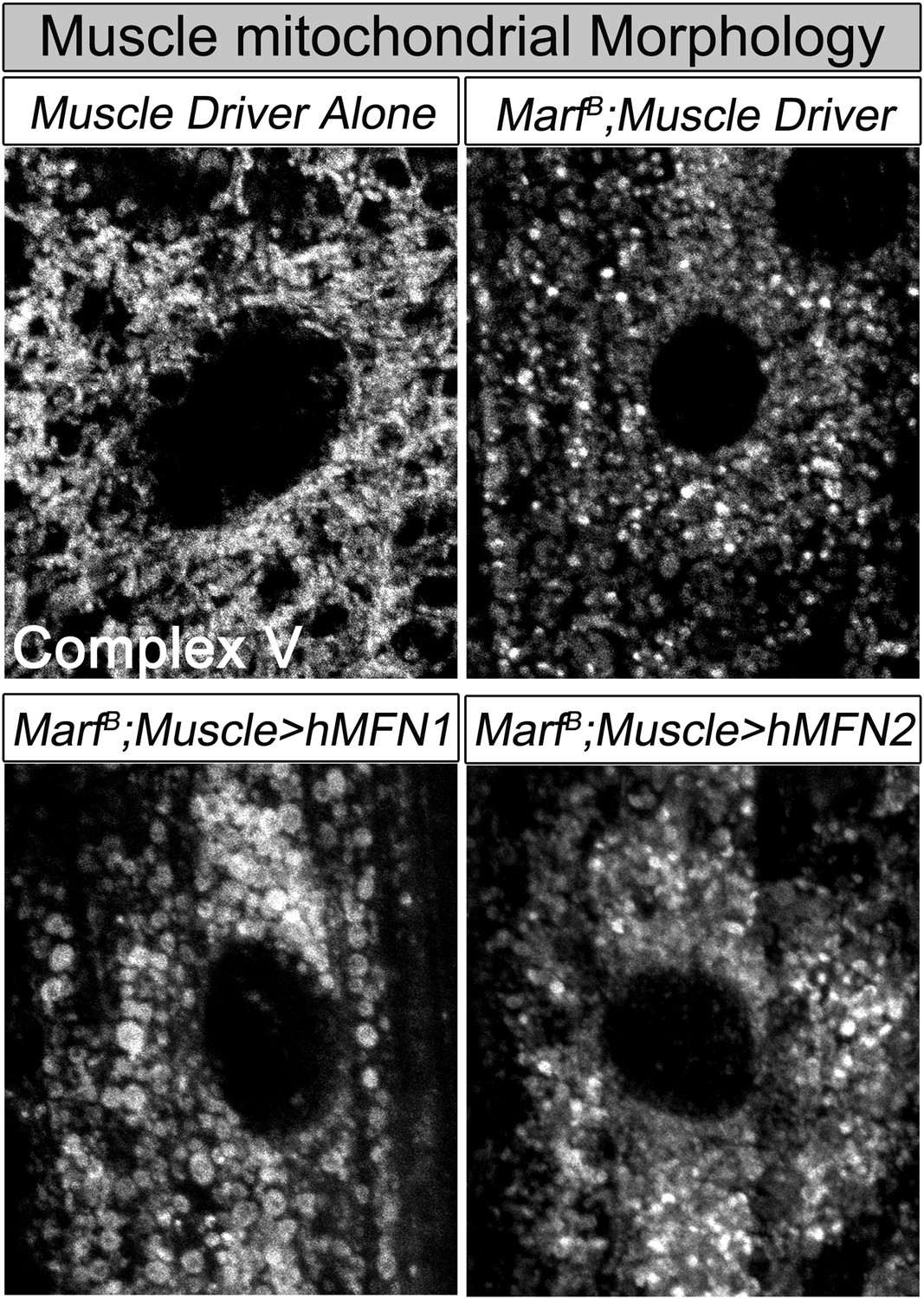
Muscle expression of either human MFN2 or MFN1 does not fully restores mitochondrial morphology in Marf mutants.
Mitochondrial morphology based on anti-Complex V antibody staining (Complex V) in larval muscles of Marf mutants with muscle expression of human MFN1 or MFN2.
Figure 8
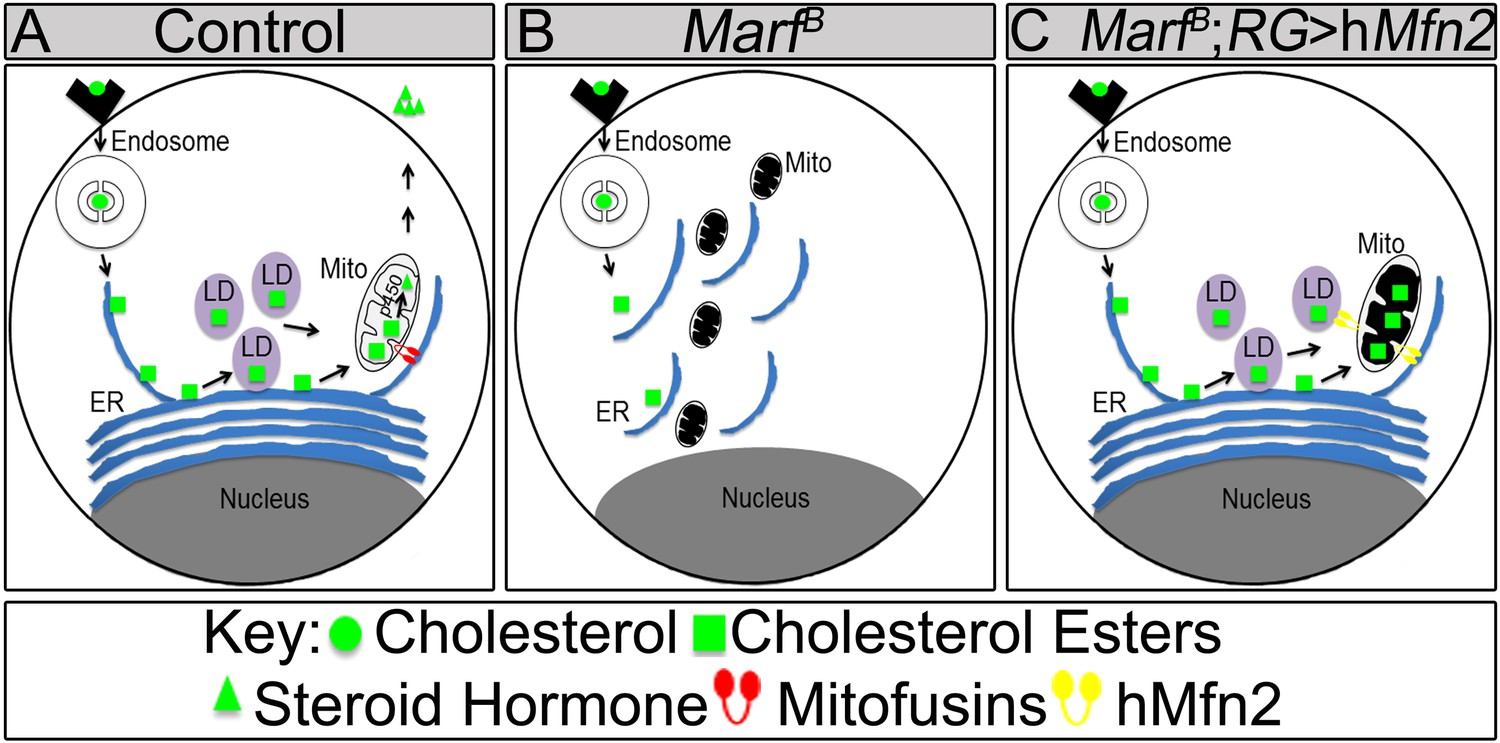
Model of Marf dual function in steroid synthesis in the ring glands.
(A) In wild type ring glands (RG), cholesterol must enter the cell first. Then, cholesterol undergoes a series of modifications in endosomes and along the ER to become free-cholesterol. Then, free-cholesterol is transferred into the mitochondrial inner matrix, where it is processed from free-cholesterol to steroid hormone by p450 enzymes. The steroid hormone is then secreted. As Drosophila larva develops it stores cholesterol in the form of cholesterol ester in lipid droplets (LDs) in order to accumulate a reserve of substrate so it can generate bursts of steroid hormone when needed. These LDs require the ER for synthesis. (B) In Marf mutants, the ER is fragmented and LD formation is severely reduced. (C) RG-specific expression of MFN2 in Marf mutant restores LD numbers but does not rescue hormone synthesis, suggesting that Marf has a second function within the mitochondria.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Mitochondrial fusion but not fission regulates larval growth and synaptic development through steroid hormone production
eLife 3:e03558.
https://doi.org/10.7554/eLife.03558




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}