Establishment of regions of genomic activity during the Drosophila maternal to zygotic transition
- Howard Hughes Medical Institute, University of California Berkeley, United States
- University of Wisconsin, United States
- The Hebrew University of Jerusalem, Israel
- University of California, Berkeley, United States
Figures
Figure 1 with 1 supplement
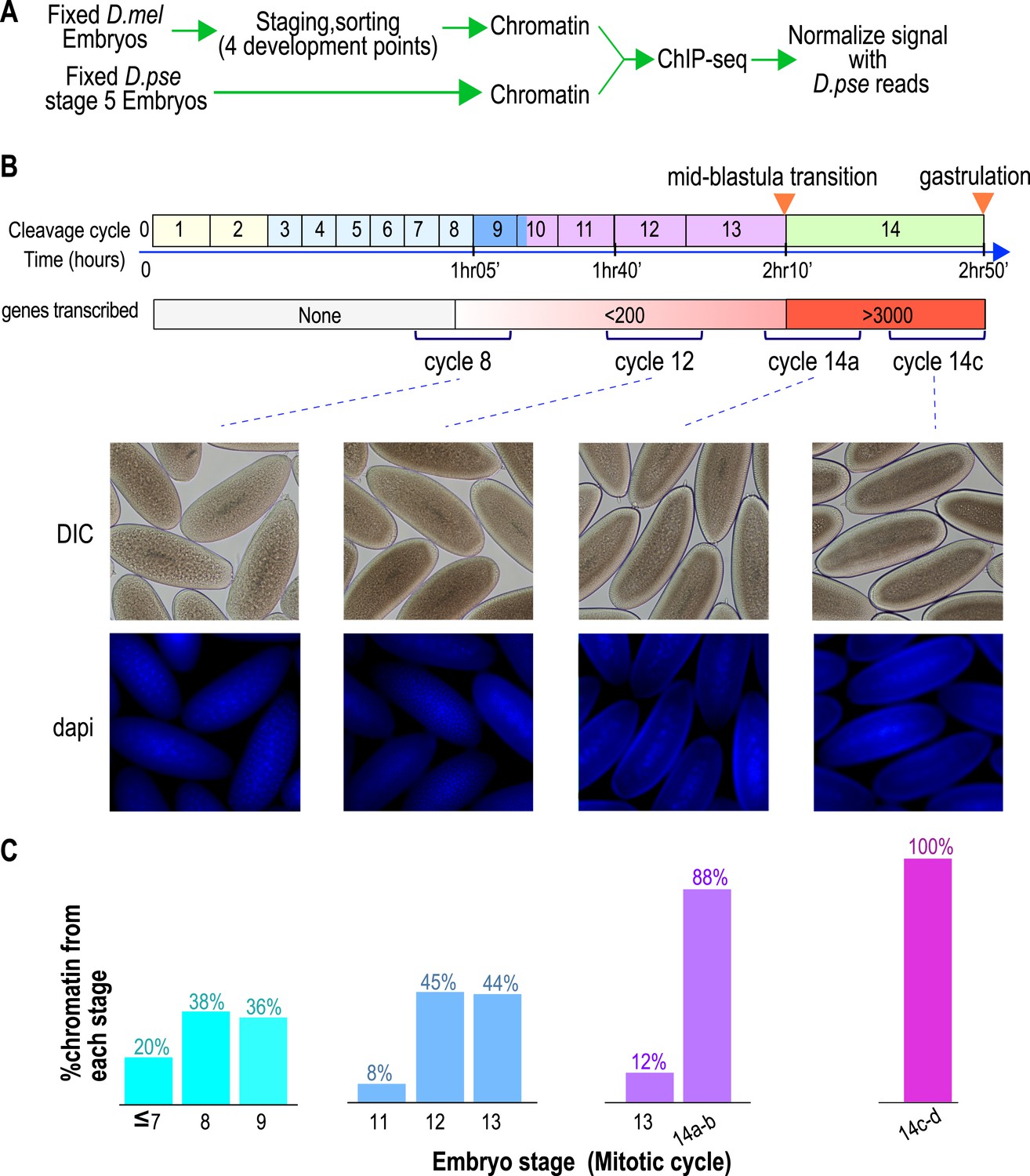
Hand sorting based on morphology results in tightly staged embryos.
(A) Experimental scheme. D. melanogaster embryos were collected and allowed to develop before being fixed with formaldehyde. Fixed embryos were hand sorted to obtain pools of embryos within a relatively narrow age distributions between mitotic cycle 8 and the end of cycle 14. To serve as carrier and normalization standard, chromatin from fixed stage 5 (cycle 14) D. pseudoobscura embryos was prepared and added to the chromatin from the sorted embryos prior to chromatin immunoprecipitation. In ChIP-seq data analysis, the sequencing reads for D. pseudoobscura were used to normalize the D. melanogaster ChIP-seq signals. (B) Embryo collection and sorting. The timeline of the early embryogenesis is depicted on top with the relative lengths of each mitotic cycle approximated by the size of the box. The developmental stages (from 1–5) are indicated by different colors. The earliest sustained transcription is detected is at cycle 7, and the mid-blastula transition (MBT) occurs when a large number of genes are transcriptionally activated at approximately the end of cycle 13. We generated four pools of sorted embryos with developmental stages centered around cycles 8, 12, 14a, or 14c as shown by differential interference contrast (DIC) and DAPI. (C) We determined the distribution of the developmental cycle of the embryos in each pool as shown by counting the number of nuclei in DAPI-stained embryos or by examining the extent of membrane envagination during cycle 14.
Figure 1—Figure supplement 1
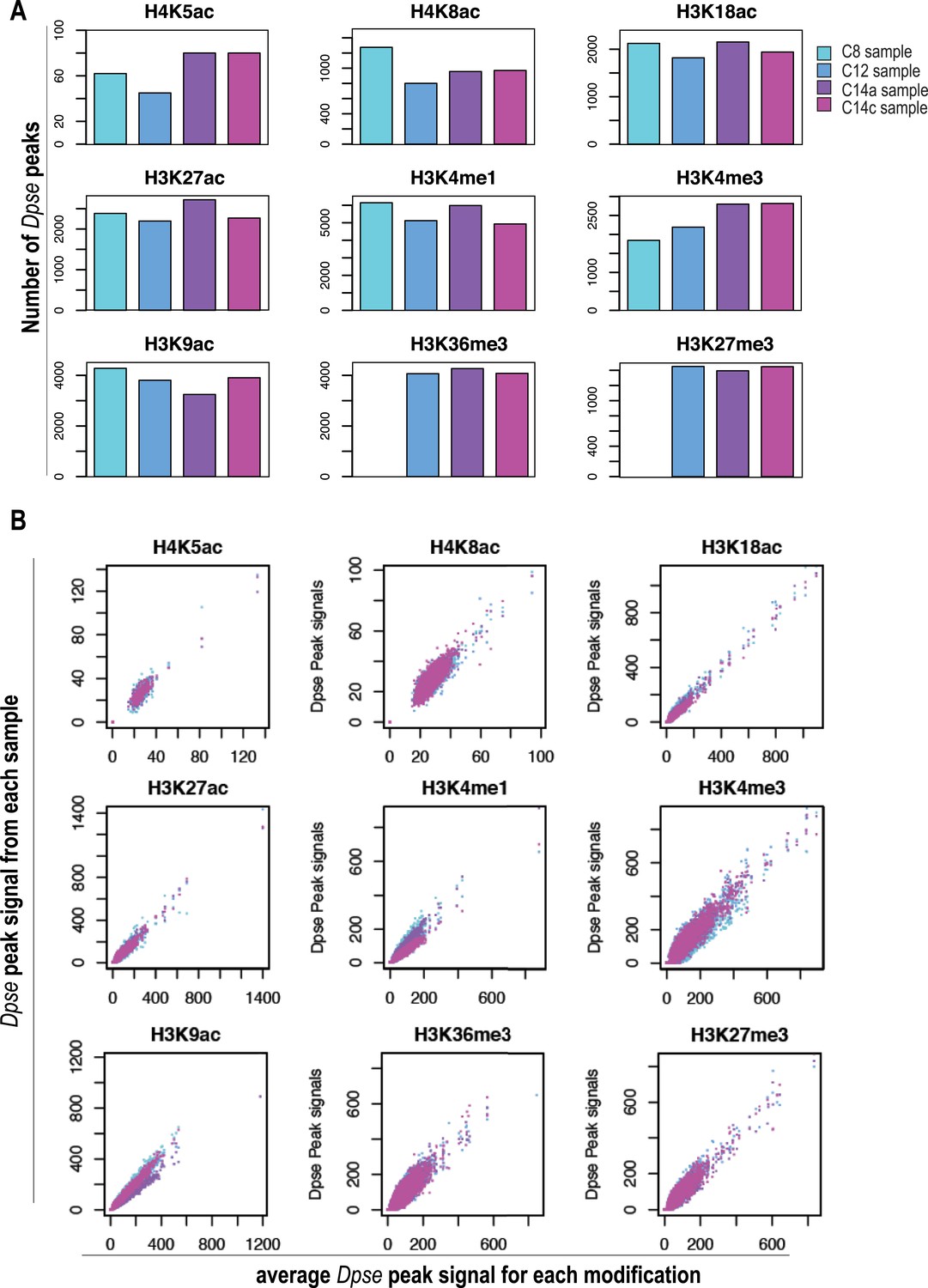
Normalization using D. pseudoobscura.
Analysis of the D. pseudoobscura chromatin used to normalize ChIP signals between different stages. Here we show the variance among the four replicates of D. pseudoobscura used to normalize the four D. melanogaster datasets (for cycles 8, 12, 14a and 14c). (A) Number of D. pseudoobscura peaks, as in Figure 2B (for D. melanogaster reads). (B) Comparison of each D. pse replicate (Y-axis) to their average (X-axis). Dpse. = D. pseudoobscura.
Figure 2

Global levels of histone marks change over early development.
(A) The number of aligned reads (after normalization to D. pseudoobscura) for the four developmental time points are indicated for each histone mark and histone H3. (B) The number of peaks detected using the peak calling program, MACS (Zhang Y et al., 2008), for each histone mark at each stage are shown. (C) Box plots show the trend of average ChIP-seq signals over ±500 bp around the peaks detected across all stages for each histone mark. The dark line in the middle of the plot represents the median, the edges of the box represent the first and third quartiles.
Figure 3

Dynamics of H3 and histone marks around selected genes.
The normalized ChIP-seq signal profiles, for histone H3 and nine different histone marks at four development time points at selected genomic loci. Shown are the early onset genes, sna and amos, and late onset genes, btsz and lea. The peak regions of histone acetylation marks detectable prior to MBT are highlighted with cyan-colored boxes. The peak regions for histone marks detected only after the MZT are highlighted by yellow-colored boxes. Below are the ZLD ChIP-seq profile (Harrison et al., 2011) from c8, 13, and 14 embryos, as well as RNA-seq signals (Lott et al., 2011) at c11, c13, c14b.
Figure 4
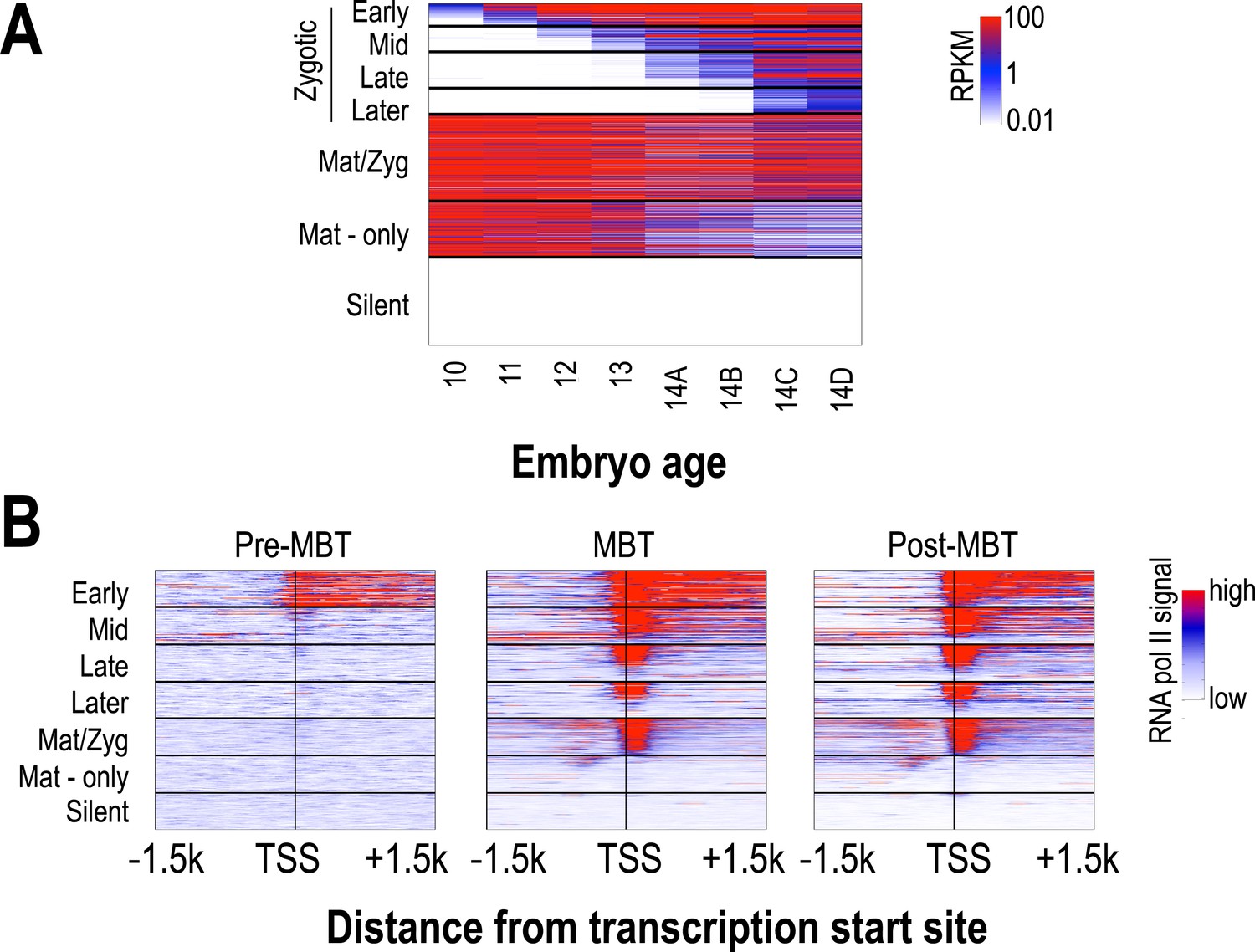
Classification of genes based on timing of transcriptional initiation during early embryogenesis.
Using single-embryo RNA-seq data from our group (Lott et al., 2011), we identified three broad classes of genes: those at high levels in the earliest embryos (‘maternal’ genes), those not present in the earliest embryos, but transcribed prior to or during mitotic cycle 14 (‘zygotic’), and those not present through cycle 14 (‘silent’). We further divided the zygotic genes into four different groups based on their onset of zygotic expression—‘Early’ genes with onset of expression around mitotic cycles 10–11, ‘Mid’ genes at cycles 12–13, ‘Late’ genes at early cycle 14, and ‘Later’ zygotic genes whose onset of expression was during late cycle 14. Post-MBT polII ChIP data (Chen et al., 2013) was used to define two maternal groups of genes—those bound by polII in the embryo (‘Mat/Zyg’ genes), and those that are strictly maternally deposited (‘Mat-only’ genes). (A) Heatmap showing the expression levels for all groups at 8 timepoints (from cycle 10 through 14D) across the MZT (Lott et al., 2011). (B) Heatmaps showing RNA polymerase II ChIP-seq signals (Chen et al., 2013) around the transcription start sites (±1.5 kb) of the genes in each category for three developmental time points, pre-MBT (left), MBT (middle), and post-MBT (right) embryos. Genes within each group were ordered based on cycle 14 RNA polymerase II signals (genes with the highest signal are on top).
Figure 5
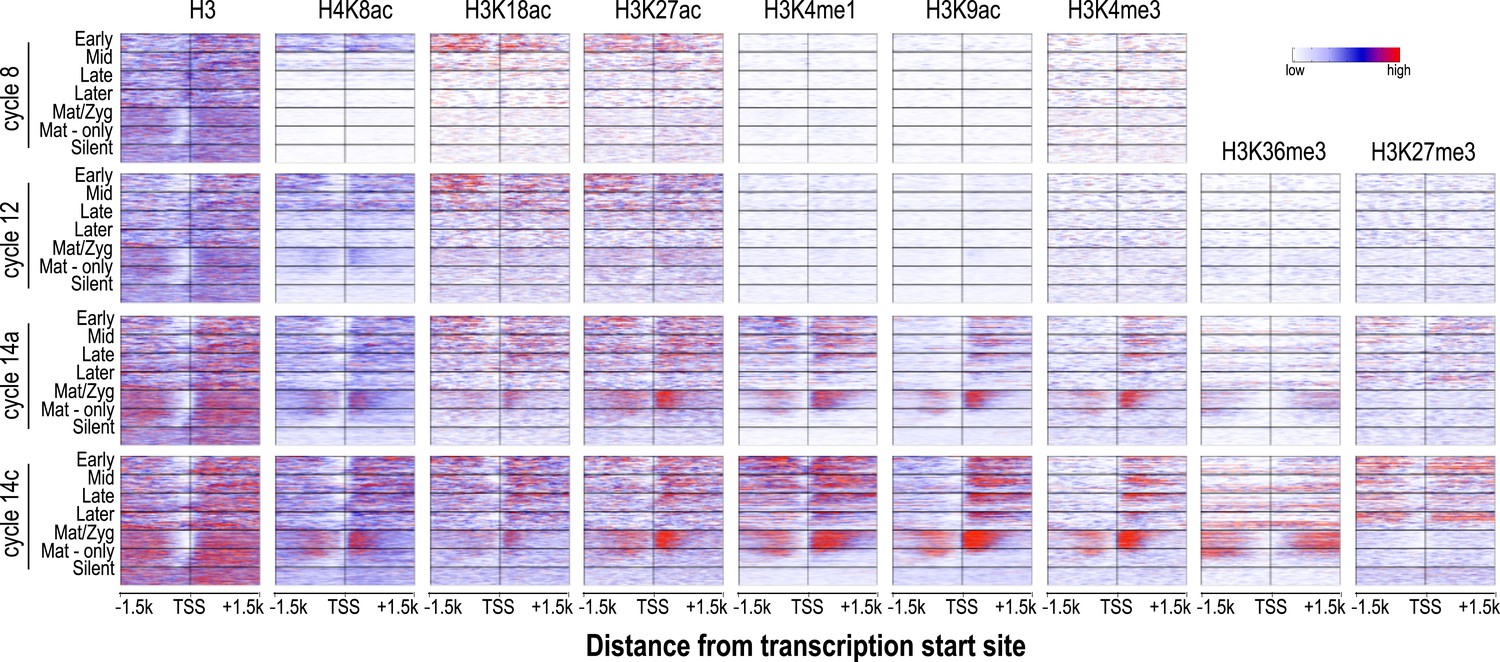
Relationship between H3 depletion, histone modifications and transcription dynamics.
Heatmaps show ChIP-seq signals for histone H3 and different histone modification marks at each stage centered around the transcription start sites (±1.5 kb). Genes are groups and ordered as described in Figure 4. For each histone mark and for histone H3 the same color scaling was used for heatmaps across all four developmental time points. TSS = transcriptional start site.
Figure 6
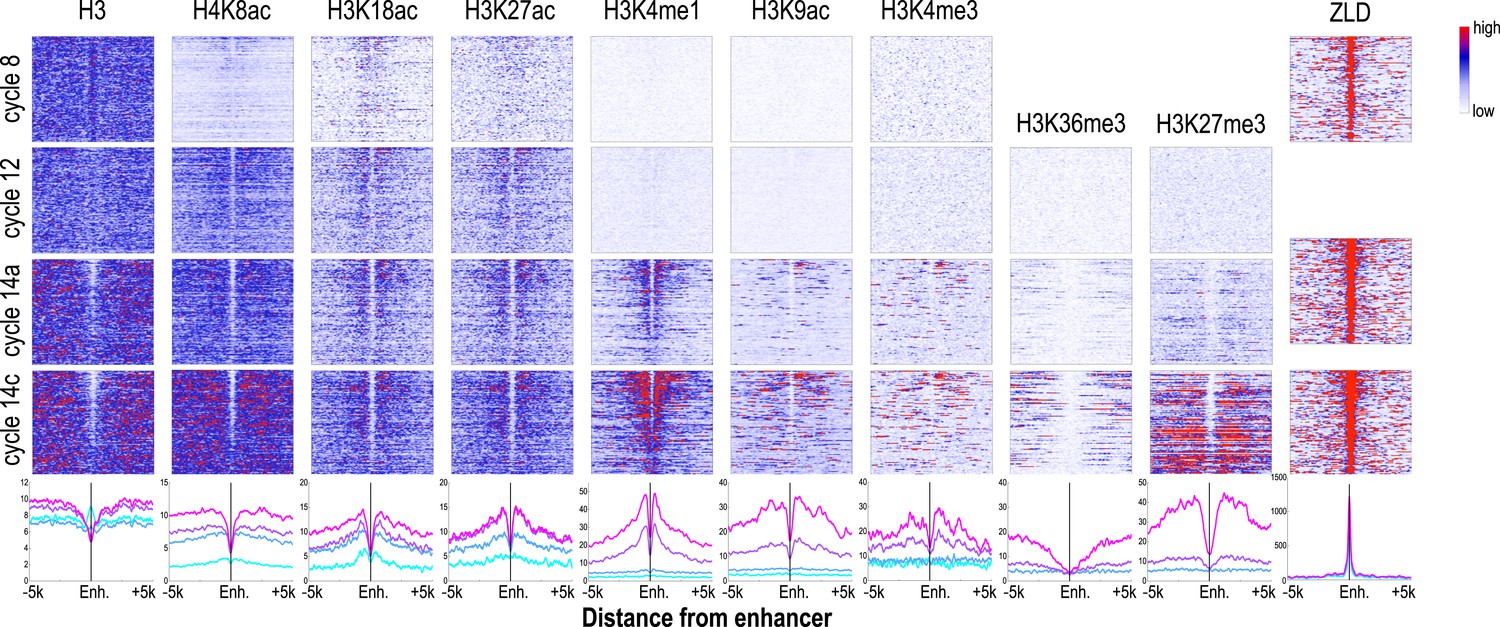
Dynamics of histone H3 depletion and histone modifications around blastoderm embryo enhancers.
Heatmaps show ChIP-seq signals for histone H3 and different histone modification marks at each stage centered around putative enhancers (as described in text). Enhancers are ordered by chromatin accessibility, as measured by DNaseI–seq signals from cycle 14 embryos (Thomas et al., 2011) from high (top) to low (bottom). On the right, the heatmaps show the ChIP-seq signals for ZLD binding around these enhancers at c8, c13, and c14 (Harrison et al., 2011). Line plots at the bottom show the average ChIP-seq for histone H3, histone modifications, and ZLD at each stage around the enhancers. Enh. = enhancer.
Figure 7
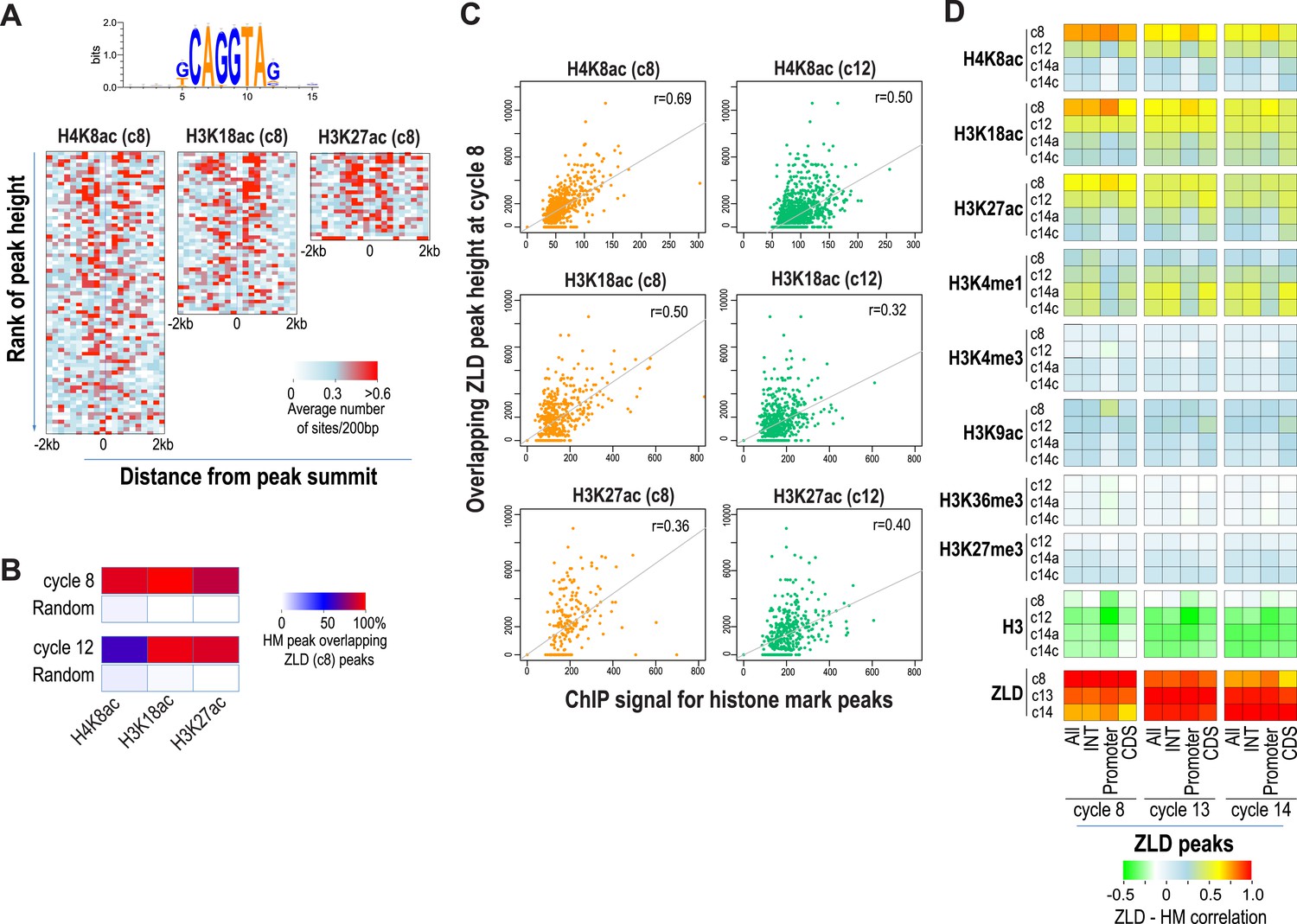
Relationship between histone occupancy, histone modification pattern and ZLD binding.
(A) ZLD DNA binding motif enrichment around cycle 8 peaks for H4K8ac, H3K18ac, and H3K27ac. Peaks were ranked based on peak height, divided into bins of 100, and analyzed. Heatmaps show, for each location (column) and set of peaks (row), the average number of ZLD putative sites at each position/set. (B) Heatmap showing the overlap between histone acetylation peaks detected at cycle 8 and 12, a.nd ZLD peaks detected at cycle 8 (top 2000 ranked peaks). As a control, overlaps between histone mark peaks with random set of genomic positions that matched the number of ZLD peaks are shown. (C) Scatter plots showing the correlation between the signals around the peaks for the histone acetylation marks at cycle 8 and cycle 12 (X-axis) and the heights of the associated ZLD peaks within 1 kb of the histone mark peaks (Y-axis). The signal for each peak was the average over the ±1 kb region surrounding the peak. The correlation coefficient (r) for each plot is shown. (D) Heatmaps showing Pearson correlation coefficients between ChIP signal of top 5000 ZLD peaks and histone marks at same locations. ChIP-seq signals for histone H3 was averaged over a ±200 bp region around each ZLD peak. Histone marks were averaged over ±1 kb around ZLD peaks. The correlation coefficients were calculated individually for all the ZLD peaks (‘All’), for intergenic and intronic ZLD peaks (‘INT’), for promoter peaks (‘Promoter’), and for ZLD peaks within coding sequences (CDS).
Figure 8
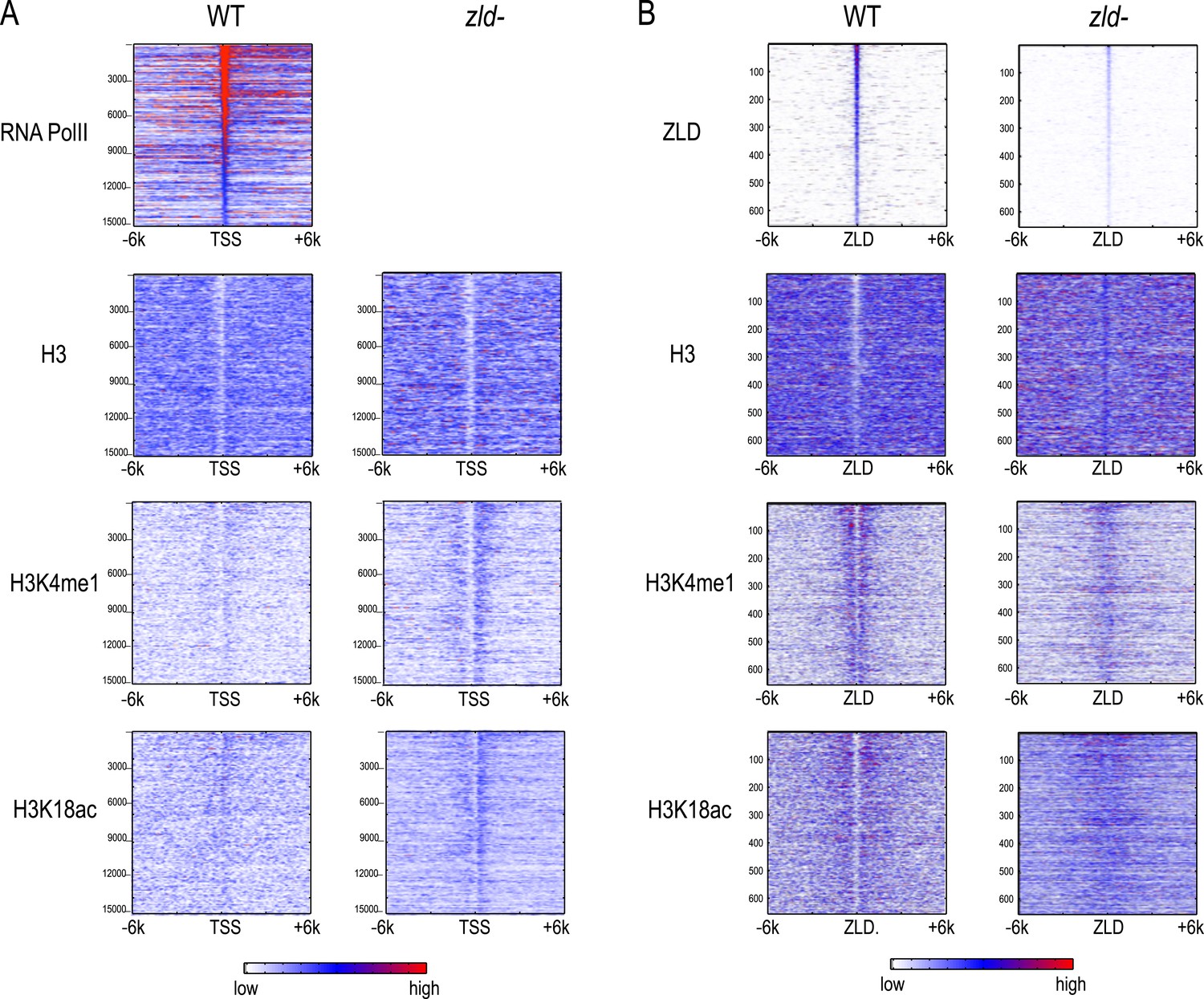
Effect of zld mutation on histone occupancy and modifications.
Heatmaps show ChIP-seq data from WT embryos (left) and embryos lacking maternal zld (right). (A) Heatmaps centered at transcription start sites (TSS) and ordered by cycle 14 RNA polymerase II binding. (B) Heatmaps centered around intergenic ZLD peaks (Harrison et al., 2011). Shown are all the intergenic and intronic peaks, among top 1000 ZLD bound regions (total of 656), ordered by ZLD ChIP signal and aligned by peak position. TSS, transcription start site.
Figure 9
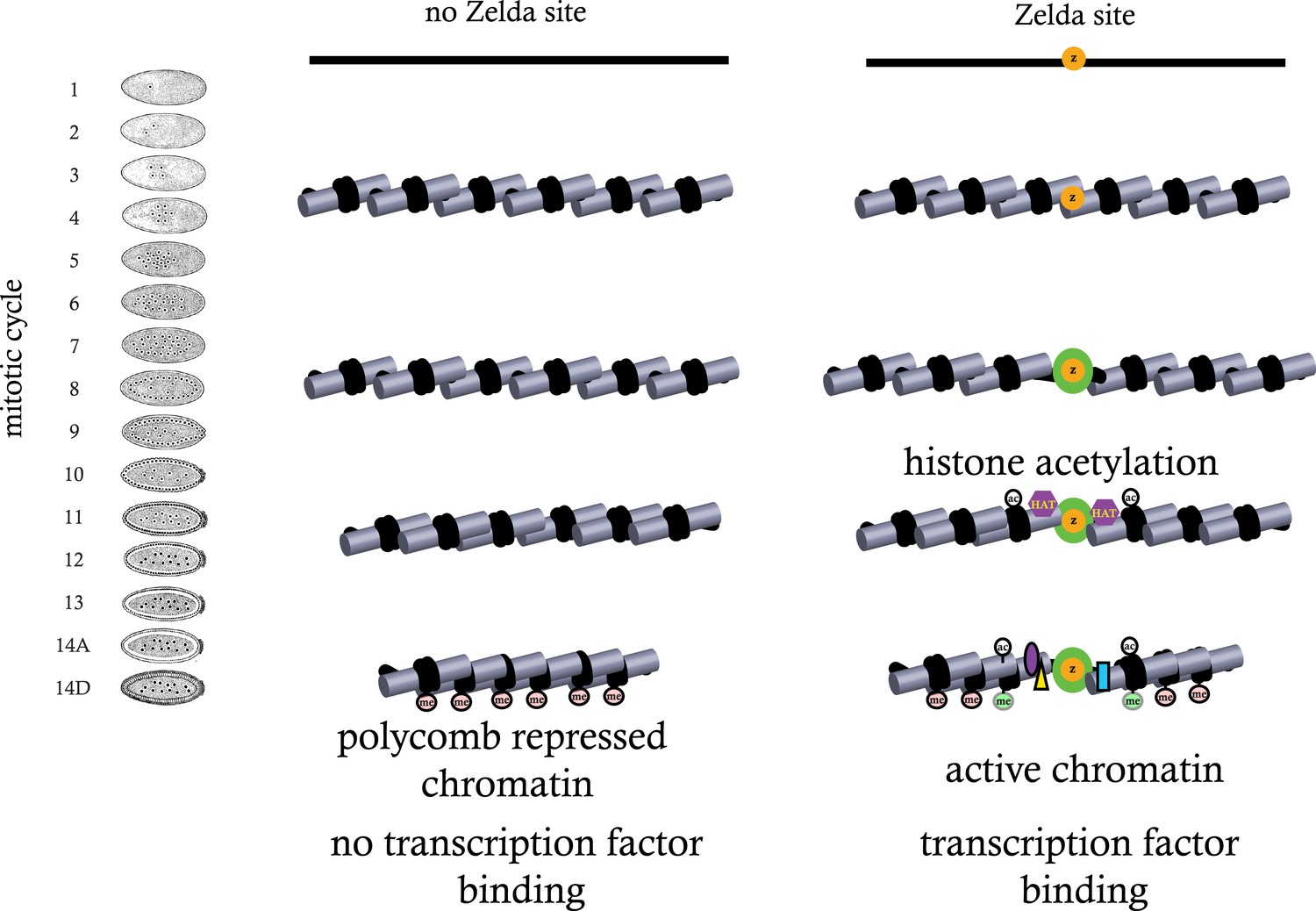
Model for ZLD function during zygotic genome activation.
ZLD binds to enhancers in pre-MBT embryos at as early as cycle 8. This leads to histone acetylation and nucleosome remodeling around ZLD binding sites, which facilitates binding by other transcription factors, and in many other cases leads to additional deposition of histone marks including H3K4me1 while at the same time prevents local deposition of repressive histone mark H3K27me3 and presumably formation of repressive higher order chromatin structure.
Tables
Table 1
Antibodies used in this study
| Mark | AB source | AB catalog # |
|---|---|---|
| H3 | Abcam | ab1791 |
| H4K5ac | Millipore | 07-327 |
| H4K8ac | Abcam | ab15823 |
| H3K18ac | Abcam | ab1191 |
| H3K27ac | Abcam | ab4729 |
| H3K4me1 | Abcam | ab8895 |
| H3K4me3 | Abcam | ab8580 |
| H3K9ac | ActiveMotif | 39,138 |
| H3K27me3 | Millipore | 07-449 |
| H3K36me3 | Abcam | ab9050 |
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Establishment of regions of genomic activity during the Drosophila maternal to zygotic transition
eLife 3:e03737.
https://doi.org/10.7554/eLife.03737




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}