The mucosal adjuvant cyclic di-GMP enhances antigen uptake and selectively activates pinocytosis-efficient cells in vivo
- Albany Medical College, United States
Peer review process
This article was accepted for publication as part of eLife's original publishing model.
History
- Version of Record published
- Accepted Manuscript published
- Accepted
- Received
Decision letter
-
Fiona M PowrieReviewing Editor; Oxford University, United Kingdom
eLife posts the editorial decision letter and author response on a selection of the published articles (subject to the approval of the authors). An edited version of the letter sent to the authors after peer review is shown, indicating the substantive concerns or comments; minor concerns are not usually shown. Reviewers have the opportunity to discuss the decision before the letter is sent (see review process). Similarly, the author response typically shows only responses to the major concerns raised by the reviewers.
Thank you for sending your work entitled “The Mucosal Adjuvant Cyclic di-GMP Enhances Antigen Uptake and Selectively Activates Pinocytosis-efficient Cells in vivo” for consideration at eLife. Your article has been favorably evaluated by Tadatsugu Taniguchi (Senior editor), a Reviewing editor, and two reviewers.
The Reviewing editor and the reviewers discussed their comments before we reached this decision, and the Reviewing editor has assembled the following comments to help you prepare a revised submission.
This is a very comprehensive and thorough study of mucosal responses to cyclic-di-GMP, a potent vaccine adjuvant. The ability of cyclic-di-GMP and related cyclic-di nucleotides to bind and activate STING, an ER membrane protein involved in type I IFN responses is well appreciated. Most of the work to date however in this area has focused on the mechanism of action of cyclic-di GMP and STING signaling in cells in culture. This study explores the in vivo mucosal adjuvant activity of cyclic-di GMP and the underlying mechanisms involved. In an elegant series of experiments the authors compare cyclic-di-GMP with cyclic GMP-AMP (cGAMP) an endogenous second messenger also known to bind STING. The authors provide evidence for the superior adjuvant activity of the bacterial ci-di-nucleotide over cGAMP. Moreoever, they provide evidence that cyclic di-GMP promotes its adjuvant activity in vivo by enhancing Ag uptake and via its ability to mobilize mucosal DCs in vivo, to promote its adjuvant effects. The authors generated CD11c-specific deletion of STING and uncover DC intrinsic roles for STING in mediating the adjuvant activities observed. Several really interesting aspects come out of these studies. Firstly, the authors convincingly demonstrate that the effects of cyclic-di-GMP are independent of type I IFN, but rather involve type II and type III IFNs. Secondly, their studies indicate that some in vivo effects of cyclic-di-GMP proceed independently of STING. These latter observations indicate that there are likely additional cyclic-di-GMP receptors important in vivo.
Specific comments:
1) STING-dependency is shown only for immune correlates (for example; cytokines, antigen uptake, etc). Whether actual immunity to a pathogen (after immunization with CDN adjuvants) requires STING has not been tested in this study. It would therefore be interesting to challenge STING-deficient mice (both germline knockout and conditional CD11c-Cre) with S.pneumoniae following PspA+c-di-GMP vaccination (as done in Figure 1E with WT mice). The STING is indeed crucial to successful vaccination with CDNs, pathogen burden in STING knockouts should be similar to that seen in WT mice vaccinated with non-adjuvanted PspA. This experiment is important, as it would implicate STING in the actual outcome/effectiveness of CDN vaccination rather than in immune correlates of the immunization.
2) The observation that the responsiveness of STING for CDNs varies between species raises some concern. C-di-GMP (the CDN used in this study) is far more potent in mouse cells in vitro compared to human cells (for example, Cavlar et al., EMBO J 2013, PMID 23604073, Figure 4). Therefore, whether c-di-GMP is superior to cGAMP as an adjuvant in humans is questionable. It is important that the authors discuss this point and amend the Abstract and other parts of the text to make it clear that c-di-GMP elicits stronger responses than cGAMP in mice. It is also noteworthy that different STING alleles found in humans encode proteins responding differentially to cGAMP isoforms (for example, Gao et al., Cell 2013, PMID 23910378).
Minor comments:
1) Figure 4A: To help those readers who are unfamiliar with the DQ-OVA+OVA647 assay, could the authors include a cartoon of how these compounds work here?
2) Figure 8F-J: Have the authors measured epithelial cell derived cytokines (as in Figure 3)? These should not be affected by knockout of STING in CD11c+ cells.
https://doi.org/10.7554/eLife.06670.016Author response
1) STING-dependency is shown only for immune correlates (for example; cytokines, antigen uptake, etc). Whether actual immunity to a pathogen (after immunization with CDN adjuvants) requires STING has not been tested in this study. It would therefore be interesting to challenge STING-deficient mice (both germline knockout and conditional CD11c-Cre) with S.pneumoniae following PspA+c-di-GMP vaccination (as done in Figure 1E with WT mice). The STING is indeed crucial to successful vaccination with CDNs, pathogen burden in STING knockouts should be similar to that seen in WT mice vaccinated with non-adjuvanted PspA. This experiment is important, as it would implicate STING in the actual outcome/effectiveness of CDN vaccination rather than in immune correlates of the immunization.
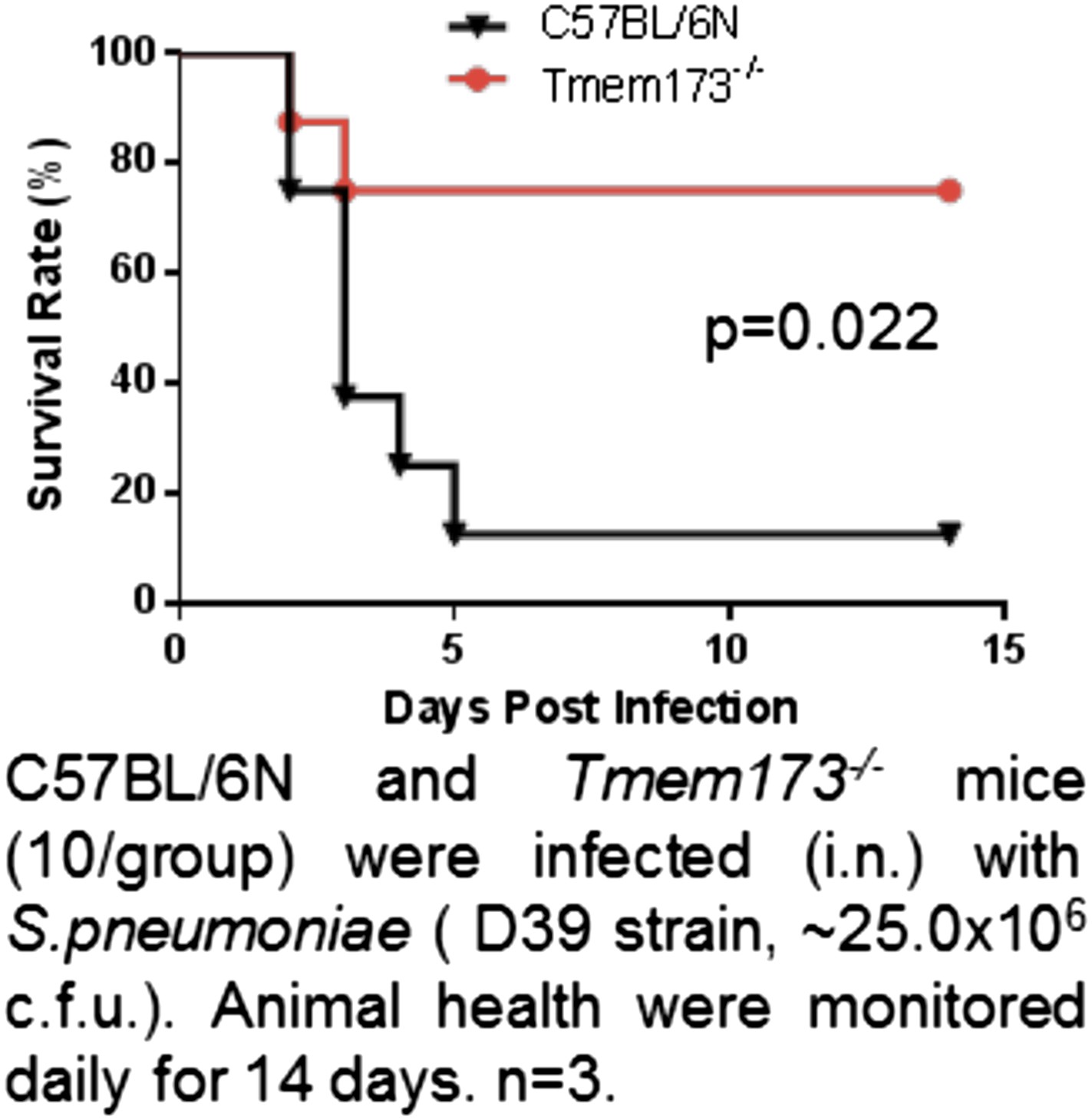
As requested, we now included the pneumococcal challenge studies done on CDG/PspA or PspA immunized WT vs Tmem173-/- mice (Figure 7G) and WT vs ItgaxCreTmem173Flox/Flox mice (Figure 9E). As expected, there is no significantly CFU difference between CDG/PspA and PspA immunized Tmem173-/- mice. However, we found that PspA immunized Tmem173-/- mice have significantly lower CFU than PspA immunize WT mice (Figure 7G). We further found that Tmem173-/- mice, without PspA immunization, are more resistant to S. pneumoniae infection than the WT mice (see Author response image 1). This is a novel unexpected finding and points to a detrimental role of MPYS in host defense against S. pneumoniae infection. Currently, we are actively pursuing this novel and exciting discovery.
Author response image 1
Similarly, CDG/PspA immunization did not lower bacterial burden in the ItgaxCreTmem173Flox/Flox mice (Figure 9E). However, differently from the Tmem173-/- mice, PspA immunized Tmem173Flox/Flox mice had similar bacterial burden as the PspA immunized ItgaxCreTmem173Flox/Flox mice (Figure 9E). We further found that ItgaxCreTmem173Flox/Flox mice, without PspA immunization, were not resistant to S. pneumoniae infection (see Author response image 2). This suggested that MPYS expression in DCs and alveolar macrophages are not responsible for its detrimental role in host response to S. pneumoniae infection. Currently, we are actively pursuing this novel and exciting discovery.
Author response image 2

2) The observation that the responsiveness of STING for CDNs varies between species raises some concern. C-di-GMP (the CDN used in this study) is far more potent in mouse cells in vitro compared to human cells (for example, Cavlar et al., EMBO J 2013, PMID 23604073, Figure 4). Therefore, whether c-di-GMP is superior to cGAMP as an adjuvant in humans is questionable. It is important that the authors discuss this point and amend the Abstract and other parts of the text to make it clear that c-di-GMP elicits stronger responses than cGAMP in mice. It is also noteworthy that different STING alleles found in humans encode proteins responding differentially to cGAMP isoforms (for example, Gao et al., Cell 2013, PMID 23910378).
We have amended the manuscript to emphasize that our observation was made in mice. We further added two paragraphs in the Discussion to address the issues raised here.
Minor comments:
1) Figure 4A: To help those readers who are unfamiliar with the DQ-OVA+OVA647 assay, could the authors include a cartoon of how these compounds work here?
Yes. We added Figure 4A and 4B to illustrate the mechanism of action of DQ-OVA and OVA-647. Thanks for the suggestion.
2) Figure 8F-J: Have the authors measured epithelial cell derived cytokines (as in Figure 3)? These should not be affected by knockout of STING in CD11C+ cells.
We added Figure 8K showing the IL-33 and TSLP production in lung from cyclic di-GMP treated ItgaxCreTmem173Flox/Flox mice. The lung TSLP production was slightly lower in cyclic di-GMP treated CD11CCreMPYSflox/flox mice than in the Tmem173Flox/Flox, but it was not statistically significant (Figure 8K). However, the lung IL-33 production was significantly lower in cyclic di-GMP treated ItgaxCreTmem173Flox/Flox mice than in the Tmem173Flox/Flox (Figure 8K).
MPYS expression in CD11C+ cells is responsible for the proinflammatory cytokines TNFα and IL-1β production, which can act on lung epithelial cells (Figure 8J). Furthermore IFN-λ receptors are uniquely expressed on lung epithelial cells and ItgaxCreTmem173Flox/Flox mice lack cyclic di-GMP induced IFN-λ production (Figure 8I). We favored the idea that there is a crosstalk/communication between lung epithelial cells and CD11C+ cells during cyclic di-GMP induced immune response.
https://doi.org/10.7554/eLife.06670.017Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The mucosal adjuvant cyclic di-GMP enhances antigen uptake and selectively activates pinocytosis-efficient cells in vivo
eLife 4:e06670.
https://doi.org/10.7554/eLife.06670




{kind=link}
{kind=link}