Selections that isolate recombinant mitochondrial genomes in animals
- University of California, San Francisco, United States
Figures
Figure 1
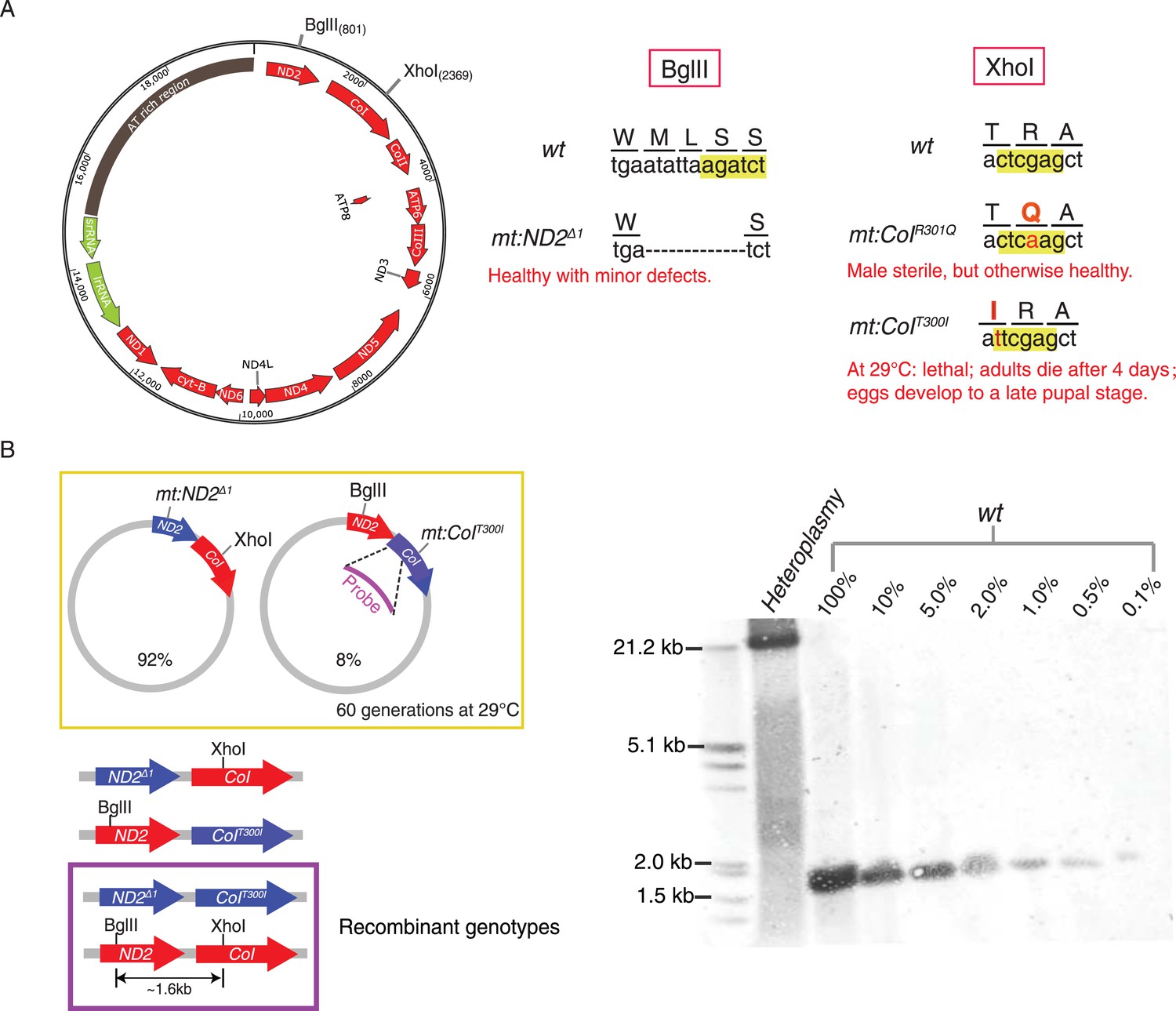
Without selection, no recombination was detected in a stable heteroplasmic line after 60 generations.
(A) Mutants at the BglII and XhoI sites of mtDNA used in this study (Xu et al., 2008; Ma et al., 2014). The mitochondrial genome of Drosophila resembles that of mammals. It has little intergenic spacing, and encodes 13 polypeptides, all of which are involved in oxidative phosphorylation as well as 22 tRNAs and two rRNAs required for mtDNA translation. A single non-coding region (∼5 kb) called the ‘AT-rich region’ (dark brown) as it contains >90% A and T residues includes origins of replication and some repeated sequences of unknown function (Lewis et al., 1994). The genome contains one BglII and one XhoI site in the coding regions of mt:ND2 and mt:CoI, respectively. (B) No wild-type genome was detected by Southern blotting analysis in heteroplasmic lines where mt:ND2del1 and mt:CoIT300I were maintained in the same population for more than 60 generations at 29°C. In the heteroplasmic lane, 40 adults were sacrificed and their mtDNA were cut with both BglII and XhoI, and probed by a DIG-labeled sequence that hybridizes to mt1579–mt2369. The sensitivity of the Southern analysis was measured by loading a series of dilutions of wild-type mtDNA cut with both enzymes from 40 adult flies.
Figure 2 with 1 supplement
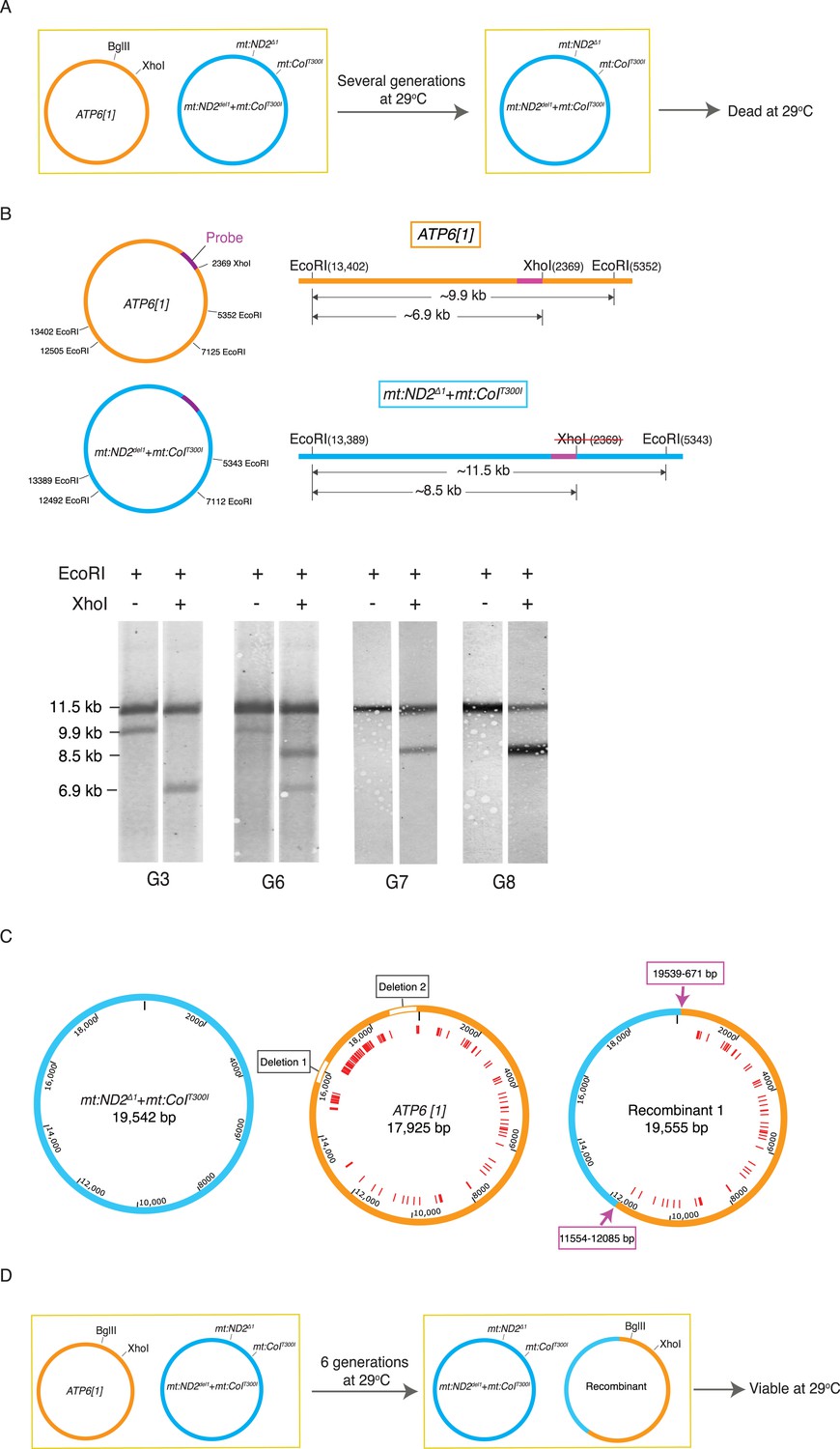
Selection revealed homologous recombination in a heteroplasmic line containing the ATP6[1] genome and the temperature-sensitive double-mutant: mt:ND2del1 + mt:CoIT300I.
(A) The abundance the ATP6[1] genome declined when co-resident with mt:ND2del1 + mt:CoIT300I at 29°C. After several generations, the flies at 29°C started to die. (B) A combination of a restriction fragment length polymorphism and a restriction site difference reveals the emergence of a recombinant genome. mtDNA isolated from 40 adults from each generation was cut with EcoRI in the presence or absence of XhoI. The schematics show the distribution of the EcoRI and XhoI sites on the whole parental genomes (left) and a detail of the largest EcoRI fragment with the position (purple bar) of a hybridization probe (center). Southern analysis shows single and double cut samples taken at different generations during the selection. Only the two parental bands were detected early, from G0 to G5 (shown for G3). From G6 onward, a third EcoRI fragment appeared that had a length characteristic of the mt:ND2del1 + mt:CoIT300I genome but with a XhoI site. By G7, the ATP6[1] specific fragment was not detected while a new apparently recombinant genome dominated the population. (C) Detailed maps of three genomes sequenced by PacBio SMRT. Red lines indicate mismatches between ATP6[1] and mt:ND2del1 + mt:CoIT300I sequences. The ATP6[1] genome also lacks ∼1.6 kb of the AT-rich region (two type I repeats and two type II repeats, see Figure 2—figure supplement 1A for details). Pink arrows indicate approximate points of exchange with the given range defined by the nearest neighboring polymorphisms (see deposited full sequences in GenBank as KT174472, KT174473 and KT174474). (D) Proposed progression leading to the recombinant. The original ATP6[1] genome is still displaced, but the newly emerged recombinant competes effectively and persists. By generation 6, the abundance of the recombinant genome is sufficient to complement the temperature-sensitive genome so that some viable flies sustain the line. Over subsequent generations at 29°C, the recombinant genome increases in relative abundance because of selection against the temperature-sensitive genome.
Figure 2—figure supplement 1
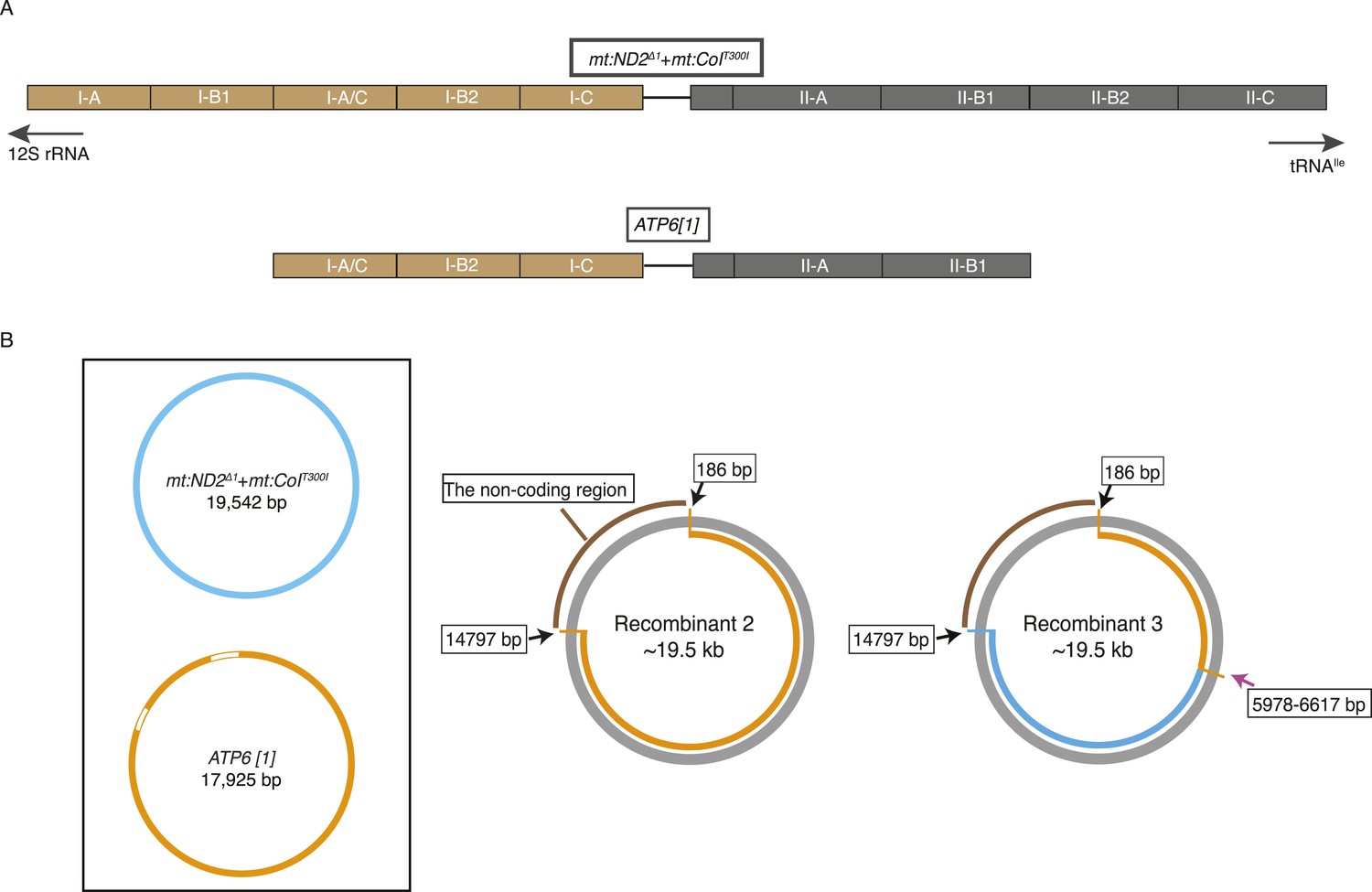
mtDNA maps for the two parental genotypes (ATP6[1], mt:ND2del1 + mt:CoIT300I) and two other recombinants.
(A) Schematic illustration of repeat structure of the AT-rich regions of ATP6[1] and mt:ND2del1 + mt:CoIT300I genomes. (B) mtDNA map of two other recombinants. The size of the recombinant genomes was determined by Southern analysis as described in Figure 2B. The coding region was amplified by PCR followed by standard sequencing. We do not have accurate sequence for the non-coding region. The pink arrow indicates approximate points of exchange with the given range defined by the nearest neighboring polymorphisms. Black arrows indicate the two ends of the coding region that has been sequenced.
Figure 3

Introducing DSBs into both parental genomes vastly induces homologous recombination in two heteroplasmic lines.
(A) Expression of both mito-BglII and mito-XhoI enzymes in the germline effectively sterilizes most females carrying either mt:ND2del1 or mt:CoIR301Q genome, resistant to only one of the enzymes, but resistant progeny appear at a much higher frequency if the female carries both types of sensitive genomes. The resistant progeny from the heteroplasmic parent are homoplasmic for a newly generated recombinant genotype carrying both parental alleles: mt:ND2del1 + mt:CoIR301Q. (B) The same level of rescue was observed when a different heteroplasmic line was used, which had a different starting ratio of two parental genomes. Results are means ±SD (n = 4 × 100 for each heteroplasmic line).
Figure 4
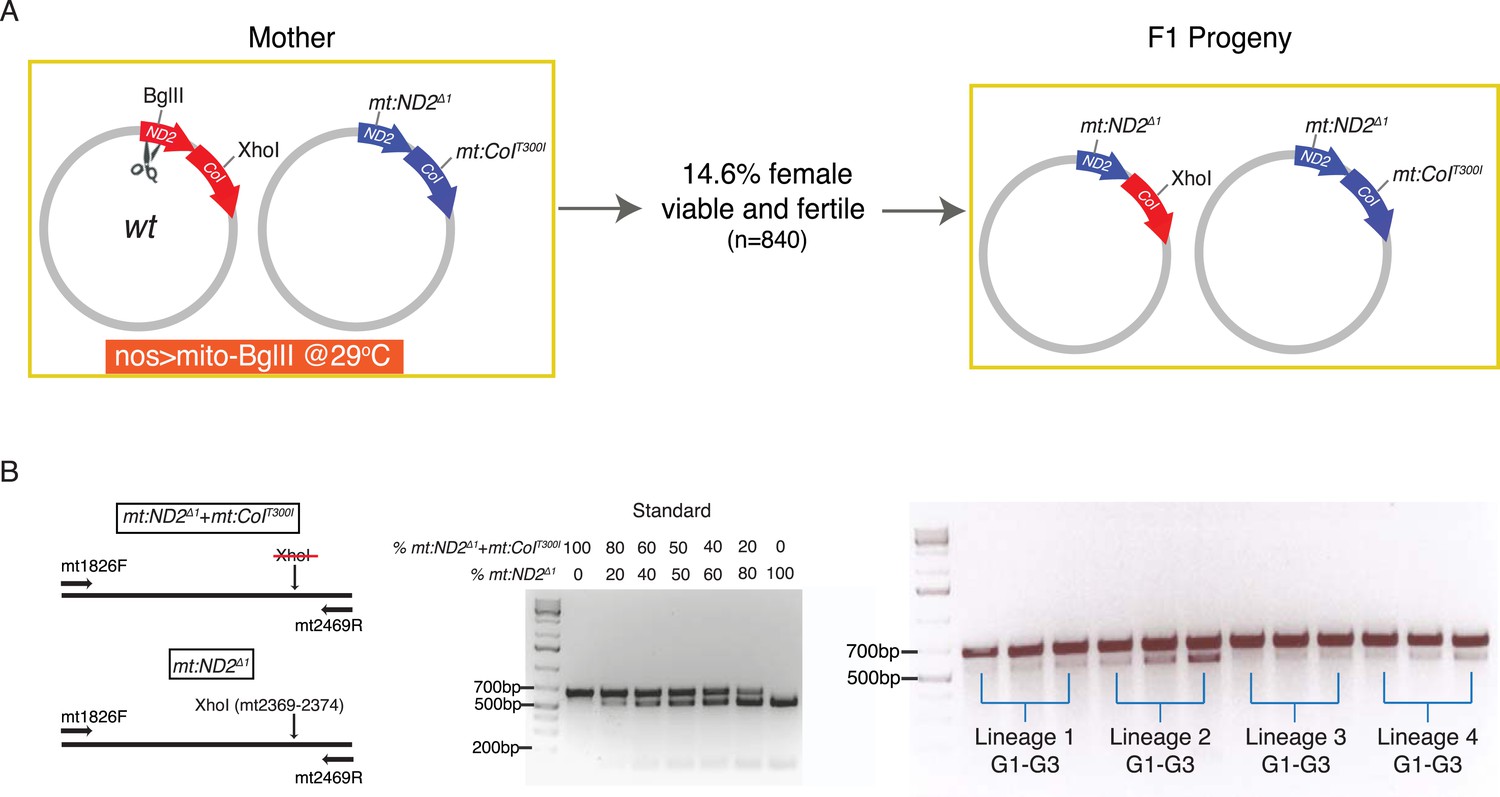
Homology-dependent conversion of the wild-type BglII site into the sequence of the mt:ND2del1 allele following cutting of the BglII site.
(A) Expression of mito-BglII in the germline of flies heteroplasmic for wild-type and mt:ND2del1 + mt:CoIT300I genomes at 29°C led to isolation of progeny with repaired wild-type genome at the BglII site and converted it to mt:ND2del1. (B) The starting abundance for the newly generated mt:ND2del1 was low, but it increased over generations as the genome possessed a selective advantage at 29°C. The abundance of mt:ND2del1 was measured by PCR amplifying a mtDNA region (mt1826–mt2496) using mtDNA from 40 adults as template followed by restriction digestion using XhoI in four heteroplasmic lineages from generation 1 to 3.
Figure 5 with 1 supplement
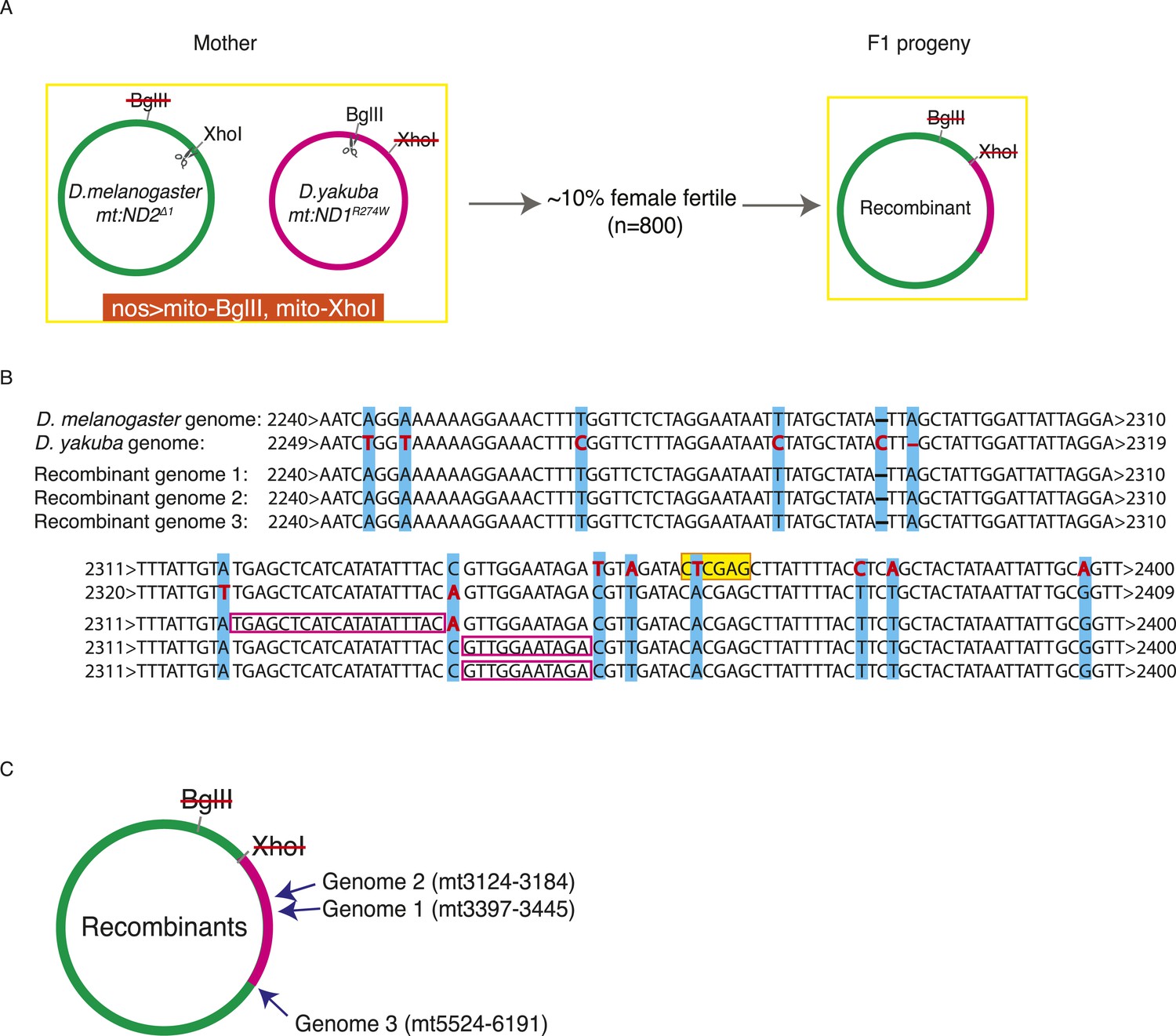
Exchange between two mitochondrial genomes of different species that share 93% sequence homology.
(A) A heteroplasmic line containing both a Drosophila yakuba (mt:ND1R274W) genome and a Drosophila melanogaster (mt:ND2del1) genome was made. 90% of heteroplasmic females induced to express both mito-BglII and mito-XhoI enzymes in the germline were sterile, but the escaper progeny from the fertile females contained newly generated recombinant genotypes. (B) Sequences of the parental and three recombinant genomes reveal that one recombination junction is close to the XhoI site of the D. melanogaster genome. The XhoI site (yellow highlight) present in the D. melanogaster parent sequence (top line) is absent in all the other sequences. Pink rectangles indicate the interval in which recombination occurred. (C) A schematic illustration of where the second crossover was for the recombinant genomes. Arrows indicate the approximate position and the numbers, which are based on the D. melanogaster sequence, given the positions of bounding polymorphisms.
Figure 5—figure supplement 1
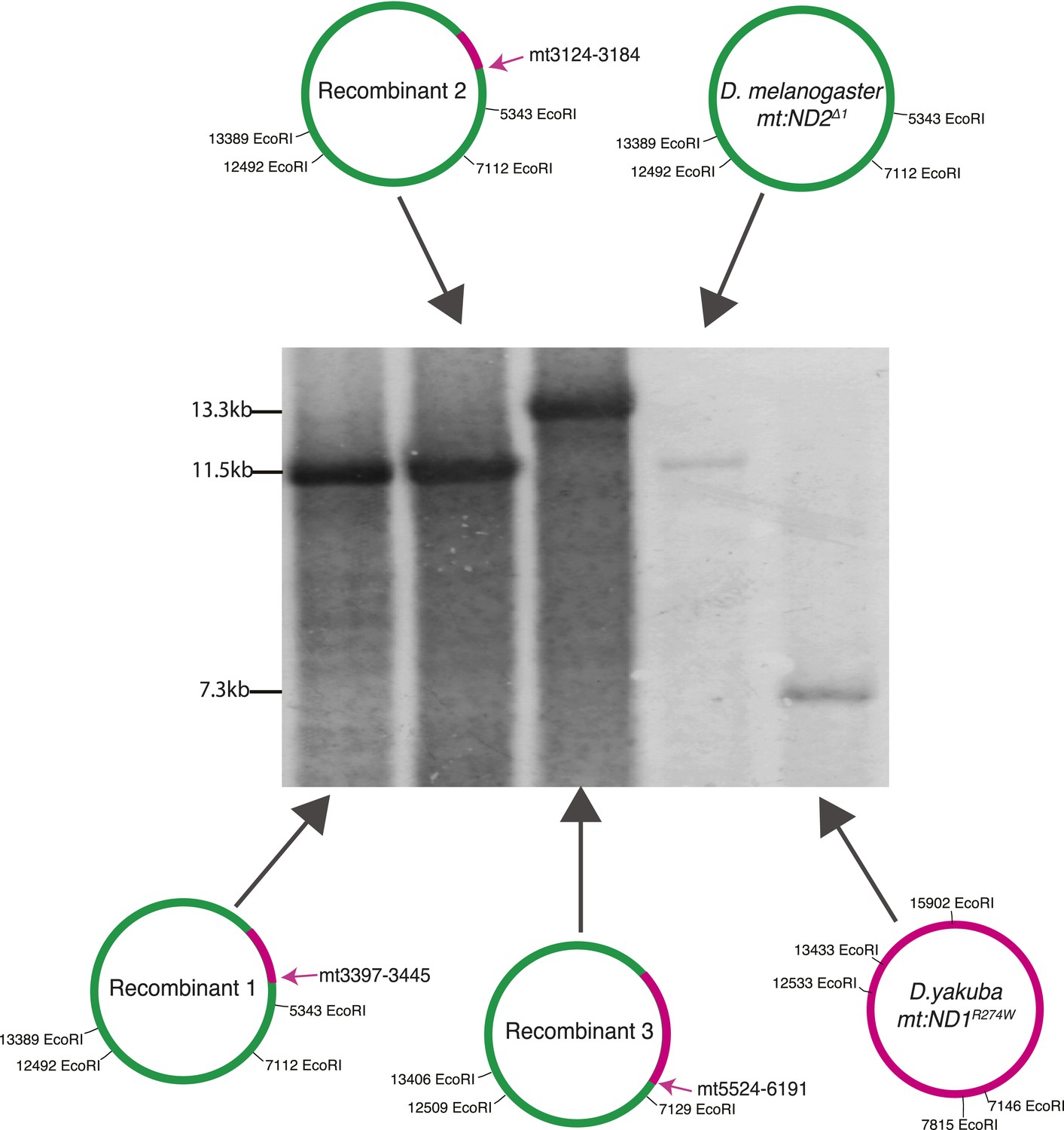
Southern Blot analysis of the long EcoRI segment covering the AT-rich region of two parental genomes (D. yakuba mt:ND1R274W and D. melanogaster mt:ND2del1) and three recombinant genomes.
None of the recombinant genomes (three left lanes) carry the short (7.3 kb) EcoRI fragment produced from the AT-rich region of the D. yakuba mtDNA (rightmost lane). We conclude that all the recombinants carry the D. melanogaster AT-rich region and attribute the shift in the size of the fragment (from 11.5 to 13.3 kb) in recombinant 3 to loss of the EcoRI site at position 5343 as demonstrated by sequencing. The probe used recognizes mt1579–mt2369.
Additional files
-
Supplementary file 1
HR-based repair was also able to rescue in somatic tissues like eye. High-level eye-antennal disc expression of both mito-BglII and mito-XhoI using ey-Gal4 at 29°C resulted in pupal lethality (headless pupa) in wild-type flies or in flies carrying mt genomes resistant to only one of the two enzymes. A few (about 1%) escapers eclosed and the majority of eclosed flies had no eye or small eye phenotype (rows 1 and 2). Having both genomes increased the eclosion rate to 10% (row 3) and most of the eclosed flies had normal-sized eyes (not shown).
- https://doi.org/10.7554/eLife.07247.010
-
Supplementary file 2
A list of primers used in this study.
- https://doi.org/10.7554/eLife.07247.011
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Selections that isolate recombinant mitochondrial genomes in animals
eLife 4:e07247.
https://doi.org/10.7554/eLife.07247




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}