Salmonella exploits host- and bacterial-derived β-alanine for replication inside host macrophages
- National Key Laboratory of Intelligent Tracking and Forecasting for Infectious Diseases, TEDA Institute of Biological Sciences and Biotechnology, Nankai University, China
eLife Assessment
The authors use a multidisciplinary approach to provide a link between Beta-alanine and S. Typhimurium (STM) infection and virulence. This valuable work shows how Beta-alanine synthesis mediates zinc homeostasis regulation, possibly contributing to virulence. The work is convincing as it adds to the existing knowledge of metabolic flexibility displayed by STM during infection.
https://doi.org/10.7554/eLife.103714.4.sa0Significance of the findings:
Valuable: Findings that have theoretical or practical implications for a subfield
- Landmark
- Fundamental
- Important
- Valuable
- Useful
Strength of evidence:
Convincing: Appropriate and validated methodology in line with current state-of-the-art
- Exceptional
- Compelling
- Convincing
- Solid
- Incomplete
- Inadequate
During the peer-review process the editor and reviewers write an eLife Assessment that summarises the significance of the findings reported in the article (on a scale ranging from landmark to useful) and the strength of the evidence (on a scale ranging from exceptional to inadequate). Learn more about eLife Assessments
Abstract
Salmonella is a major foodborne pathogen that can effectively replicate inside host macrophages to establish life-threatening systemic infections. Salmonella must utilize diverse nutrients for growth in nutrient-poor macrophages, but which nutrients are required for intracellular Salmonella growth is largely unknown. Here, we found that either acquisition from the host or de novo synthesis of a nonprotein amino acid, β-alanine, is critical for Salmonella replication inside macrophages. The concentration of β-alanine is decreased in Salmonella-infected macrophages, while the addition of exogenous β-alanine enhances Salmonella replication in macrophages, suggesting that Salmonella can uptake host-derived β-alanine for intracellular growth. Moreover, the expression of panD, the rate-limiting gene required for β-alanine synthesis in Salmonella, is upregulated when Salmonella enters macrophages. Mutation of panD impaired Salmonella replication in macrophages and colonization in the mouse liver and spleen, indicating that de novo synthesis of β-alanine is essential for intracellular Salmonella growth and systemic infection. Additionally, we revealed that β-alanine influences Salmonella intracellular replication and in vivo virulence partially by increasing expression of the zinc transporter genes znuABC, which in turn facilitates the uptake of the essential micronutrient zinc by Salmonella. Taken together, these findings highlight the important role of β-alanine in the intracellular replication and virulence of Salmonella, and panD is a promising target for controlling systemic Salmonella infection.
eLife digest
Salmonella, a type of bacterium, is one of the most common foodborne pathogens and is responsible for illnesses ranging from gastroenteritis to typhoid fever. Each year, it infects over 100 million people globally, leading to 350,000 deaths.
Immune cells called macrophages are important for defending against bacterial infections as they can engulf and destroy harmful bacteria. However, Salmonella is able to survive and multiply within these very immune cells that are meant to eliminate it. To do so, the bacteria require nutrients such as amino acids, which are the building blocks of proteins. These are either produced by the bacteria or obtained from the infected host. However, the specific nutrients that Salmonella require to survive and multiply, as well as their source, remained unknown.
To investigate, Ma, Yang et al. measured amino acid levels in macrophages that had been infected with Salmonella and compared them to those in uninfected macrophages. This revealed that the levels of an amino acid called β-alanine – which differs from many amino acids because it is not used to make proteins – are lower in infected macrophages. Furthermore, providing infected macrophages with more β-alanine increased bacterial replication. This suggests that the bacteria acquire this amino acid from the macrophages in order to survive and replicate.
To determine whether Salmonella can also make β-alanine themselves, Ma et al. prevented them from producing it, which slowed bacterial growth and led to milder infections in mice. This ability to produce β-alanine required a gene known as PanD. Further experiments also showed that β-alanine assists Salmonella in acquiring the essential micronutrient zinc from macrophages.
Taken together, the findings of Ma et al. reveal the critical role of β-alanine in Salmonella growth within macrophages and its ability to cause disease. The panD gene that enables Salmonella to synthesize β-alanine could serve as a potential target for new treatments or vaccines. By targeting this specific aspect of Salmonella's survival strategy, researchers may be able to develop more effective methods to prevent and treat these dangerous infections.
Introduction
Salmonella is a major foodborne pathogen worldwide that can cause self-limiting gastroenteritis or life-threatening systemic disease in a wide range of animals (Fàbrega and Vila, 2013; Ohl and Miller, 2001). Salmonella infection remains a significant global public health concern. An estimated 93.8 million cases of gastroenteritis and 27 million cases of systemic diseases caused by Salmonella species occur annually worldwide, with 355,000 deaths (Kim et al., 2019; Majowicz et al., 2010). The ability to survive and replicate in host macrophages is a key determinant for Salmonella to induce systemic infection (Fields et al., 1986; LaRock et al., 2015; Leung and Finlay, 1991). After internalization by macrophages, Salmonella delivers a set of more than 30 effector proteins to the macrophage cytoplasm, mainly through a type III secretion system (T3SS) encoded by Salmonella pathogenicity island-2 (SPI-2) (Pillay et al., 2023). SPI-2 effectors manipulate diverse cellular processes to promote the formation of a membrane-bound compartment, termed the Salmonella-containing vacuole (SCV), a niche where Salmonella resides and grows (Castanheira and García-del Portillo, 2017; Rosenberg et al., 2022; Steeb et al., 2013). SCV protects Salmonella from contact with antimicrobial agents in macrophages (Figueira and Holden, 2012; Li et al., 2023). Moreover, SPI-2 effectors induce the formation of specific tubular membrane compartments that extend from the SCV, known as Salmonella-induced filaments (SIFs). These filaments allow Salmonella to access various types of endocytosed nutrients, thereby facilitating efficient replication within macrophages (Brumell et al., 2001; Liss et al., 2017; Rajashekar et al., 2008).
As the SCV of macrophages is a nutrient-poor environment (Fields et al., 1986; Kehl et al., 2020), to effectively replicate in the SCV, Salmonella needs to acquire a wide range of host nutrients or host-derived metabolites and synthesize metabolites de novo that cannot be sufficiently accessed from the host (Dandekar et al., 2014; Röder et al., 2021; Tuli and Sharma, 2019). Nutrients/metabolites are used by intracellular Salmonella either as carbon sources to generate energy or for the synthesis of fatty acids and proteins (Dandekar et al., 2014; Steeb et al., 2013). Moreover, several metabolites were found to be employed by Salmonella as environmental cues to induce the expression of virulence genes (Jiang et al., 2021; Wang et al., 2023). In recent years, an increasing number of studies have focused on the intracellular nutrition of Salmonella (Bumann and Schothorst, 2017; Liss et al., 2017; Röder et al., 2021); however, the nutrients that are required for Salmonella replication in macrophages remain largely unknown.
β-Alanine, also known as 3-aminopropionic acid (3-AP), is the only naturally occurring β-type amino acid and is found in all living organisms (Song et al., 2023). β-Alanine can be synthesized de novo by bacteria, fungi, and plants, whereas animals need to obtain it from food or generate it via the catabolism of cytosine and uracil (Gojković et al., 2001; Wang et al., 2021b). In bacteria, β-alanine is synthesized via the decarboxylation of L-aspartate, a reaction catalyzed by L-aspartate decarboxylase (PanD) (Begley et al., 2001; Schmitzberger et al., 2003; West et al., 1985). The panD gene is conserved among most bacteria (Nozaki et al., 2012). Although β-alanine is a nonprotein amino acid that is not incorporated into proteins, it has important physiological functions in the metabolism of organisms (Wang et al., 2021b; Yuan et al., 2022). First, β-alanine forms a part of pantothenate (vitamin B5), which is the key precursor for the biosynthesis of coenzyme A (CoA) (Webb et al., 2004; White et al., 2001). CoA is an essential cofactor involved in many metabolic pathways, including the synthesis and degradation of fatty acids, pyruvate oxidation through the tricarboxylic acid (TCA) cycle, and the production of secondary metabolites (Davaapil et al., 2014; Gout, 2019; Sibon and Strauss, 2016; Theodoulou et al., 2014). Second, β-alanine is a limiting precursor of carnosine, a nonenzymatic free radical scavenger and a natural antioxidant, with anti-inflammatory and neuroprotective effects in animals (Boldyrev et al., 2013; Hoffman et al., 2018). In the past 15 y, β-alanine has become one of the most commonly used sports supplements worldwide (Bellinger, 2014; Hoffman et al., 2018; Huerta Ojeda, Tapia Huerta Ojeda et al., 2020). Although both Salmonella and host cells are capable of producing β-alanine, whether β-alanine contributes to the pathogenicity and intracellular growth of Salmonella remains unknown.
In this work, using targeted metabolic profiling, in vitro and in vivo infection assays, and many other molecular techniques, we demonstrated that the utilization of β-alanine is essential for Salmonella replication in host macrophages and virulence in mice. Salmonella acquires β-alanine both via the uptake of β-alanine from host macrophages and the de novo synthesis of β-alanine. Further investigation revealed the molecular mechanism underlying the contribution of β-alanine to Salmonella intracellular replication and pathogenicity, wherein β-alanine promotes the expression of zinc transporter genes to facilitate the uptake of the essential micronutrient zinc by intracellular Salmonella, therefore promoting Salmonella replication in macrophages and subsequent systemic infection. Taken together, these findings demonstrate a correlation between Salmonella β-alanine utilization and zinc uptake during intracellular infection and provide new insights into the intracellular nutrition of Salmonella. The rate-limiting gene (panD) in the β-alanine synthesis pathway of Salmonella might be a future target for the prevention and treatment of this pathogen.
Results
Host-derived β-alanine promotes Salmonella replication inside macrophages
To explore changes in the levels of different amino acids inside macrophages upon Salmonella infection, we performed targeted metabolomics analysis of mouse RAW264.7 macrophages that were mock-infected or infected with wild-type Salmonella (Salmonella enterica serovar Typhimurium ATCC 14,028 s, STM) for 8 hr using liquid chromatography-tandem mass spectrometry (LC‒MS/MS) (Figure 1A). Principal component analysis (PCA) demonstrated a clear separation between the mock- and Salmonella-infected groups (Figure 1B). A total of 26 free amino acids were analyzed, and eight showed significant differences in abundance between the two groups (VIP (Variable Importance in the Projection) >1 and p<0.05; FC (fold change) > 1.5 or < 0.667) (Figure 1C and D). Compared with those in the mock-infected group, the concentrations of three amino acids (L-hydroxyproline, L-citrulline and L-cysteine) were upregulated (Figure 1C), and five amino acids (L-asparagine, L-serine, L-aspartate, β-alanine and γ-aminobutyric acid) were downregulated (Figure 1D) in the Salmonella-infected group. Consistent with previous findings, intracellular serine concentrations were downregulated due to the reprogramming of macrophage glucose metabolism during Salmonella infection (Jiang et al., 2021). Salmonella can use host-derived aspartate and asparagine for growth in macrophages (Popp et al., 2015); therefore, the decrease in intracellular aspartate and asparagine upon Salmonella infection is likely due to their utilization by bacteria. Interestingly, β-alanine concentrations were also downregulated in the Salmonella-infected group (Figure 1D), suggesting that intracellular Salmonella may use host-derived β-alanine for growth.
Figure 1 with 8 supplements see all
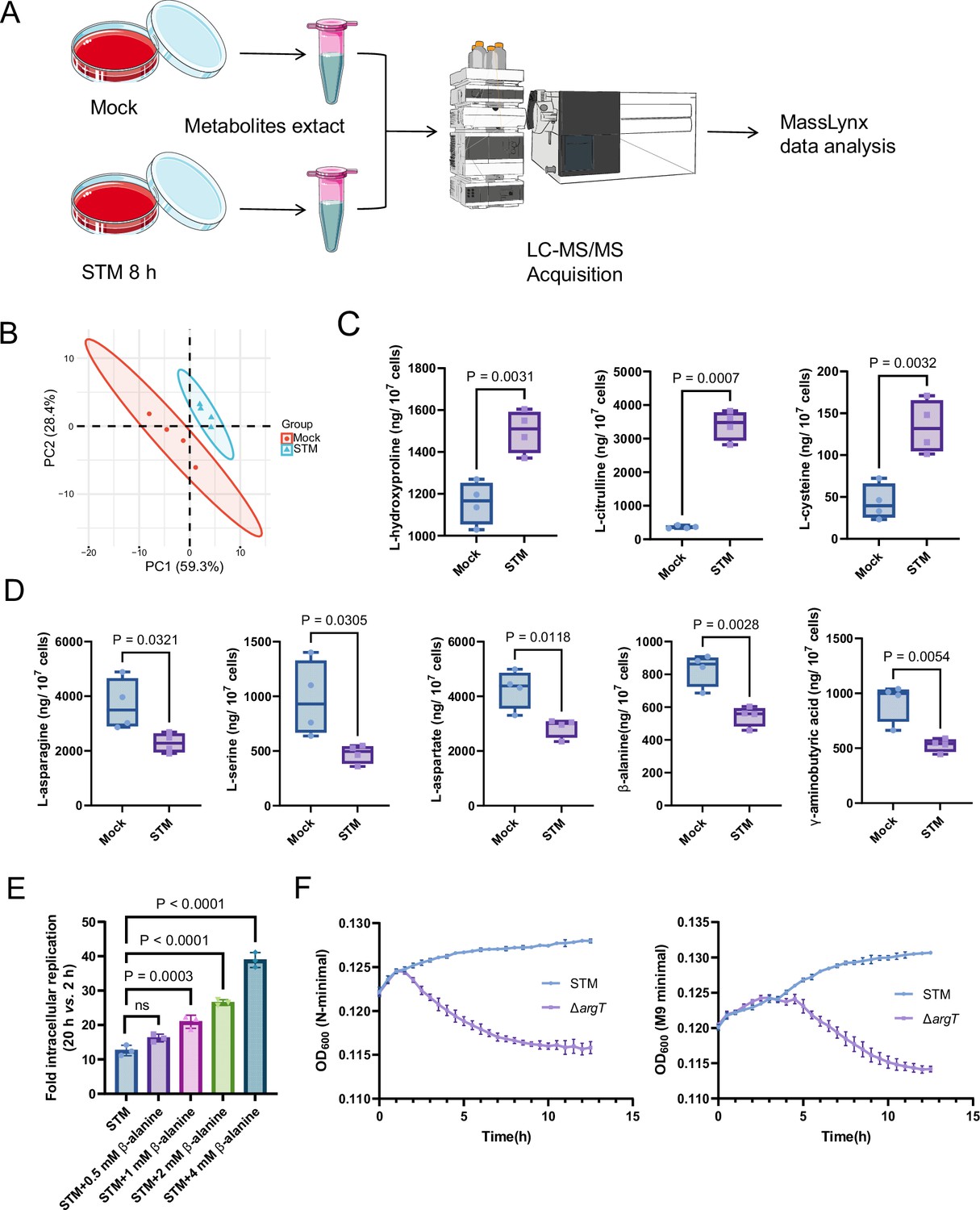
Host-derived β-alanine promotes Salmonella replication inside macrophages.
(A) Schematic workflow for targeted metabolomics investigation of mock- and Salmonella-infected (STM) mouse RAW264.7 macrophages. Picture materials were used from bioicons (https://bioicons.com/). (B) Principal component analysis (PCA) score plots of metabolic profiles in the mock- and Salmonella-infected (STM) groups (n=4 biologically independent samples). (C) The concentrations of upregulated amino acids in the mock- and Salmonella-infected groups (n=4 biologically independent samples). (D) The concentrations of downregulated amino acids in the mock- and Salmonella-infected groups (n=4 biologically independent samples). (E) Fold intracellular replication (20 hr vs. 2 hr) of Salmonella WT in RAW264.7 cells in the presence of 0.5, 1, 2, 4 mM β-alanine. Data are presented as the mean ± SD, n=3 independent experiments. (F) Growth curves of Salmonella WT and the argT mutant (ΔargT) in N-minimal (left) and M9 minimal (right) medium supplemented with β-alanine (1 mM) as the sole carbon source. Data are presented as mean ± SD, n=4 independent experiments. Statistical significance was assessed using two-sided Student’s t-test (C, D) and one-way ANOVA (E).
-
Figure 1—source data 1
The numerical source data corresponds to Figure 1.
- https://cdn.elifesciences.org/articles/103714/elife-103714-fig1-data1-v1.xlsx
To investigate whether host-derived β-alanine can promote intracellular Salmonella replication, we added an additional 0.5, 1, 2, 4 mM β-alanine (Schneider et al., 2004) to the culture medium (RPMI) of RAW264.7 cells and then infected them with Salmonella to test the influence of β-alanine addition on the ability of Salmonella to replicate in macrophages. The results showed that the replication of Salmonella in RAW264.7 cells significantly (p<0.001) increased with the addition of 1, 2, or 4 mM β-alanine (Figure 1E). Furthermore, β-alanine enhanced Salmonella intracellular replication in a dose-dependent manner (Figure 1E). The results suggest that host-derived β-alanine facilitates Salmonella replication inside macrophages. We then investigated whether β-alanine-mediated Salmonella growth promotion is due to the changes in antimicrobial activity of the macrophages. We observed that the addition of 1 mM β-alanine did not influence the ROS (reactive oxygen species) and RNS (reactive nitrogen species) levels in Salmonella-infected RAW264.7 cells (Figure 1—figure supplement 1). Flow cytometry analysis indicated that the addition of 1 mM β-alanine did not affect the percentage of pro-inflammatory M1 macrophages (CD86+) and anti-inflammatory M2 macrophages (CD163+) during Salmonella infection (Figure 1—figure supplement 2), implying that the addition of β-alanine to macrophages does not change their immune response. Combining these results, we can further infer that Salmonella use host-derived β-alanine for intracellular growth.
Direct validation of Salmonella using host-derived β-alanine for intracellular growth requires a mutant that has a defect in β-alanine uptake. Escherichia coli uptakes β-alanine via the transporter protein CycA (Schneider et al., 2004). However, the Salmonella ΔcycA mutant was able to use β-alanine as the sole carbon source for growth in minimal medium (Figure 1—figure supplements 3 and 4), indicating that CycA is not a transporter for β-alanine in Salmonella. Consistent with these results, mutation of cycA did not influence the replication of Salmonella in RAW264.7 cells (Figure 1—figure supplement 5) or colonization in mouse systemic tissues (liver and spleen; Figure 1—figure supplement 6). In E. coli, GabP transports γ-aminobutyric acid (GABA), a structural analog of β-alanine, and may also transport β-alanine (Pavić et al., 2021). Nevertheless, the Salmonella ∆gabP mutant displayed no growth defect in minimal medium with β-alanine as the sole carbon source (Figure 1—figure supplements 7 and 8), indicating that GabP is not involved in β-alanine uptake in Salmonella. Strikingly, the ΔargT mutant—defective in arginine uptake—showed markedly decreased growth in the minimal medium with β-alanine as the sole carbon source (Figure 1F), suggesting that ArgT also transports β-alanine in Salmonella. It has been reported that ArgT is essential for Salmonella replication within macrophages and full virulence in vivo (Das et al., 2010). Given that ArgT is involved in both arginine and β-alanine uptake (as verified in this study), whether the attenuated virulence of the ∆argT mutant is due to a deficiency in β-alanine or arginine requires further investigation.
De novo β-alanine synthesis is critical for Salmonella replication inside macrophages
Salmonella can de novo synthesize β-alanine via the decarboxylation of L-aspartate, which is catalyzed by L-aspartate decarboxylase (PanD) (Figure 2A) and is reportedly the rate-limiting step of β-alanine generation (Begley et al., 2001; Schmitzberger et al., 2003; West et al., 1985). To further assess the role of β-alanine in Salmonella intracellular replication, we analyzed the expression level of the Salmonella panD gene in macrophages and the impact of panD mutation on the ability of Salmonella to replicate in macrophages. Quantitative real-time PCR (qRT‒PCR) assays revealed that the expression level of panD was significantly (p<0.01) greater in RAW264.7 cells than in RPMI-1640 medium (Figure 2B). Increased expression of panD was also observed in N-minimal medium, a widely used medium that mimics the conditions inside macrophages, as revealed by qRT‒PCR and bioluminescent reporter assays (Figure 2C and D). These results demonstrate that panD expression is enhanced during Salmonella growth inside macrophages, suggesting a relationship between panD expression and intracellular Salmonella growth.
Figure 2 with 3 supplements see all
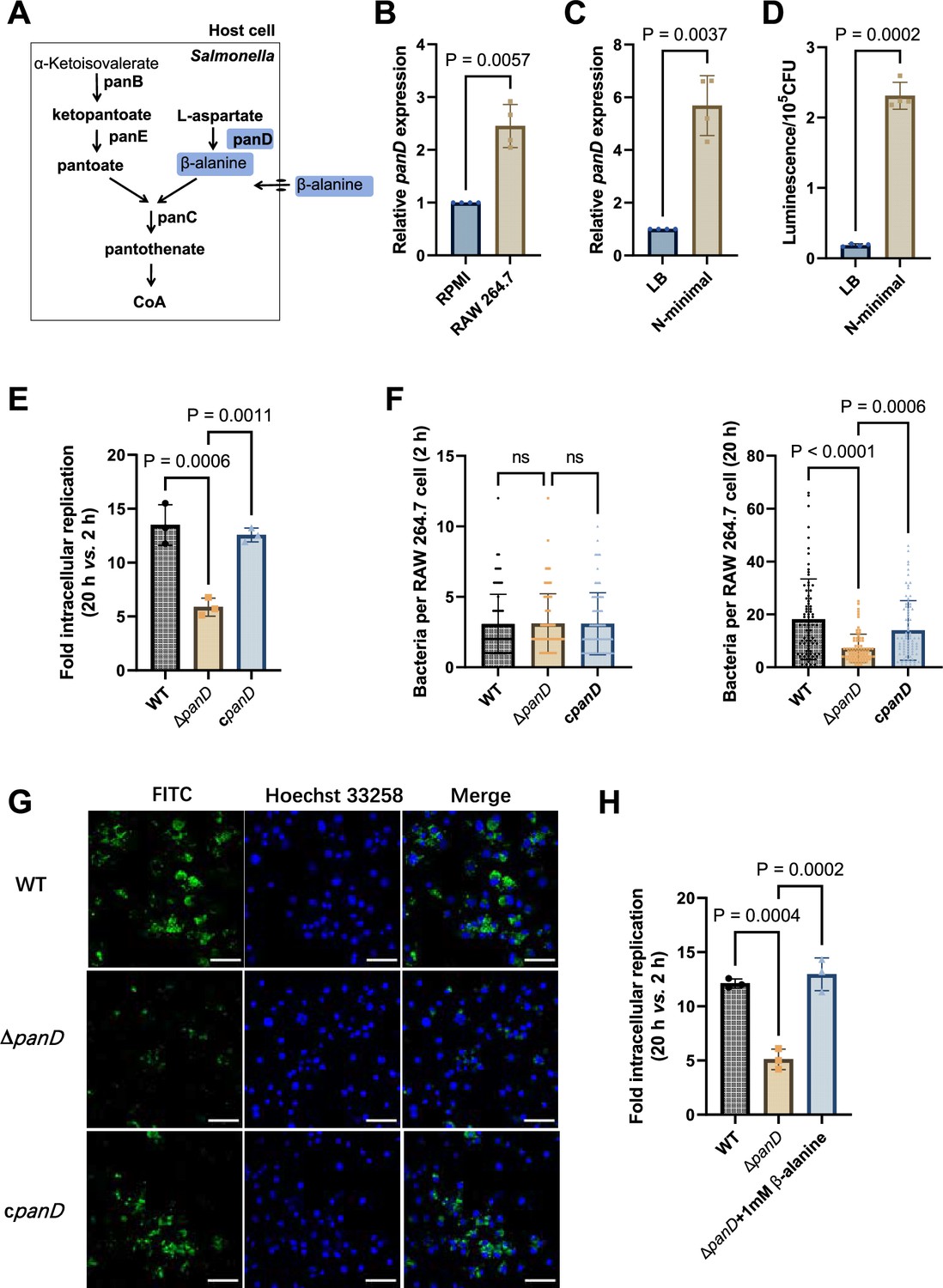
De novo β-alanine synthesis is critical for Salmonella replication inside macrophages.
(A) Scheme of β-alanine and the downstream CoA biosynthesis pathway in Salmonella. (B) Quantitative real-time PCR (qRT‒PCR) analysis of the expression of the Salmonella panD gene in RAW264.7 cells (8 hr post-infection) and RPMI-1640 medium. (C) qRT‒PCR analysis of the expression of the Salmonella panD gene in N-minimal medium and LB medium. (D) Expression of the panD-lux transcriptional fusion in N-minimal medium and LB medium. Luminescence values were normalized to 105 bacterial CFUs. (E) Fold intracellular replication (20 hr vs. 2 hr) of Salmonella Typhimurium 14,028 s wild-type (WT), the panD mutant (ΔpanD), and the complemented strain (cpanD) in RAW264.7 cells. (F) Number of intracellular Salmonella WT, ΔpanD, and cpanD strains per RAW264.7 cell at 2 and 20 hr post-infection. The number of intracellular bacteria per infected cell was estimated in random fields, n=80 cells per group from three independent experiments. (G) Representative immunofluorescence images of Salmonella WT, ΔpanD, and cpanD in RAW264.7 cells at 20 hr post-infection (green, Salmonella; blue, nuclei; scale bars, 50 µm). Images are representative of three independent experiments. (H) Replication of Salmonella WT and ΔpanD in RAW264.7 cells in the presence or absence of 1 mM β-alanine. The data are presented as the mean ± SD, n=3 (B–E, H) independent experiments. Statistical significance was assessed using a two-sided Student’s t-test (B–D) or one-way ANOVA (E, F, H). ns, not Significant.
-
Figure 2—source data 1
The numerical source data corresponds to Figure 2.
- https://cdn.elifesciences.org/articles/103714/elife-103714-fig2-data1-v1.xlsx
We then constructed the panD mutant strain ΔpanD and compared the replication ability of the ΔpanD strain and the Salmonella Typhimurium 14,028 s wild-type (WT) strain in RAW264.7 cells. Gentamicin protection assays showed that the replication of ΔpanD in RAW264.7 cells decreased 2.4-fold at 20 hr post-infection compared with that of the WT strain (p<0.001), while complementation of ΔpanD with the panD gene restored the replication ability of the mutant strain in RAW264.7 cells (Figure 2E). Immunofluorescence analysis revealed that the number of ΔpanD in each infected RAW264.7 cell was comparable to that of the WT strain at the initial infection stage (2 hr), but at 20 hr post-infection, the number of ΔpanD in each infected RAW264.7 cell was significantly (p<0.0001) lower than that of the WT strain (Figure 2F and G). These results indicate that panD contributes to Salmonella replication in macrophages. The growth rates of ΔpanD in LB medium and RPMI medium resembled those of the WT (Figure 2—figure supplements 1 and 2), indicating that the impaired intracellular replication ability of the mutant was not due to a growth defect. Moreover, the replication defect of ΔpanD in RAW264.7 cells was relieved by the addition of 1 mM β-alanine to the RPMI medium (Figure 2H). Furthermore, we examined the role of β-alanine synthesis in the intracellular replication of Salmonella within another typical serovar, Salmonella enterica serovar Typhi (S. Typhi), a serovar specific to humans and the causative agent of typhoid fever (de Jong et al., 2012). We found that the replication of S. Typhi ΔpanD in human THP-1 macrophages was reduced by 2.6-fold compared to the S. Typhi Ty2 WT strain (p<0.01) (Figure 2—figure supplement 3), suggesting that panD also facilitates S. Typhi replication within human macrophages.
These data collectively suggest that β-alanine synthesis is critical for Salmonella replication inside macrophages.
De novo β-alanine synthesis is critical for systemic Salmonella infection in mice
As replication in macrophages is a key determinant of systemic Salmonella infection, we reasoned that β-alanine synthesis also influences Salmonella systemic infection in vivo. To determine whether β-alanine influences systemic Salmonella infection, we conducted mouse infection assays using intraperitoneal (i.p.) injection. This method allows Salmonella to disseminate directly to systemic sites via the lymphatic and bloodstream systems, bypassing the need for intestinal invasion (Pandeya et al., 2023; Silva et al., 2016). BALB/c mice were infected by i.p. injection of 5000 CFU of WT, ΔpanD, or the complemented strain cpanD. The survival rate, body weight, bacterial burden in the liver and spleen, and liver histopathological alterations of the infected mice were measured (Figure 3A). The WT-infected mice exhibited high lethality and marked loss of body weight within 5 d, and all mice died within 9 d of infection (Figure 3B and C). In contrast, the ΔpanD-infected mice displayed significantly improved survival rates and body weights, and no mice died within the 10 d surveillance period (Figure 3B and C). Consistent with these results, the bacterial burden in the liver and spleen of the ΔpanD-infected mice was significantly decreased, and the body weight was significantly increased compared with that of the WT-infected mice on day 3 post-infection (Figure 3D). Complementation of ΔpanD with panD significantly decreased the survival rate and body weight of infected mice but significantly increased the bacterial burden in the liver and spleen of the infected mice (Figure 3B–D). Through immunofluorescence staining, we examined the bacterial count in liver macrophages of mice infected with WT, ΔpanD, and complemented strain. The results showed that the bacterial count in each macrophage from ΔpanD-infected mice was significantly (P<0.0001) lower than that in WT-infected mice, on day 5 post-infection. Complementation of ΔpanD with panD restored the bacterial count in each macrophage to WT level (Figure 3E). Furthermore, H&E staining revealed increased aggregation of inflammatory cells and pyknosis in the livers of the WT-infected mice on day 5 post-infection, while these histopathological alterations were obviously reduced in the livers of ΔpanD-infected mice (Figure 3F). Taken together, these results reveal that β-alanine synthesis is critical for systemic Salmonella infection in mice.
Figure 3
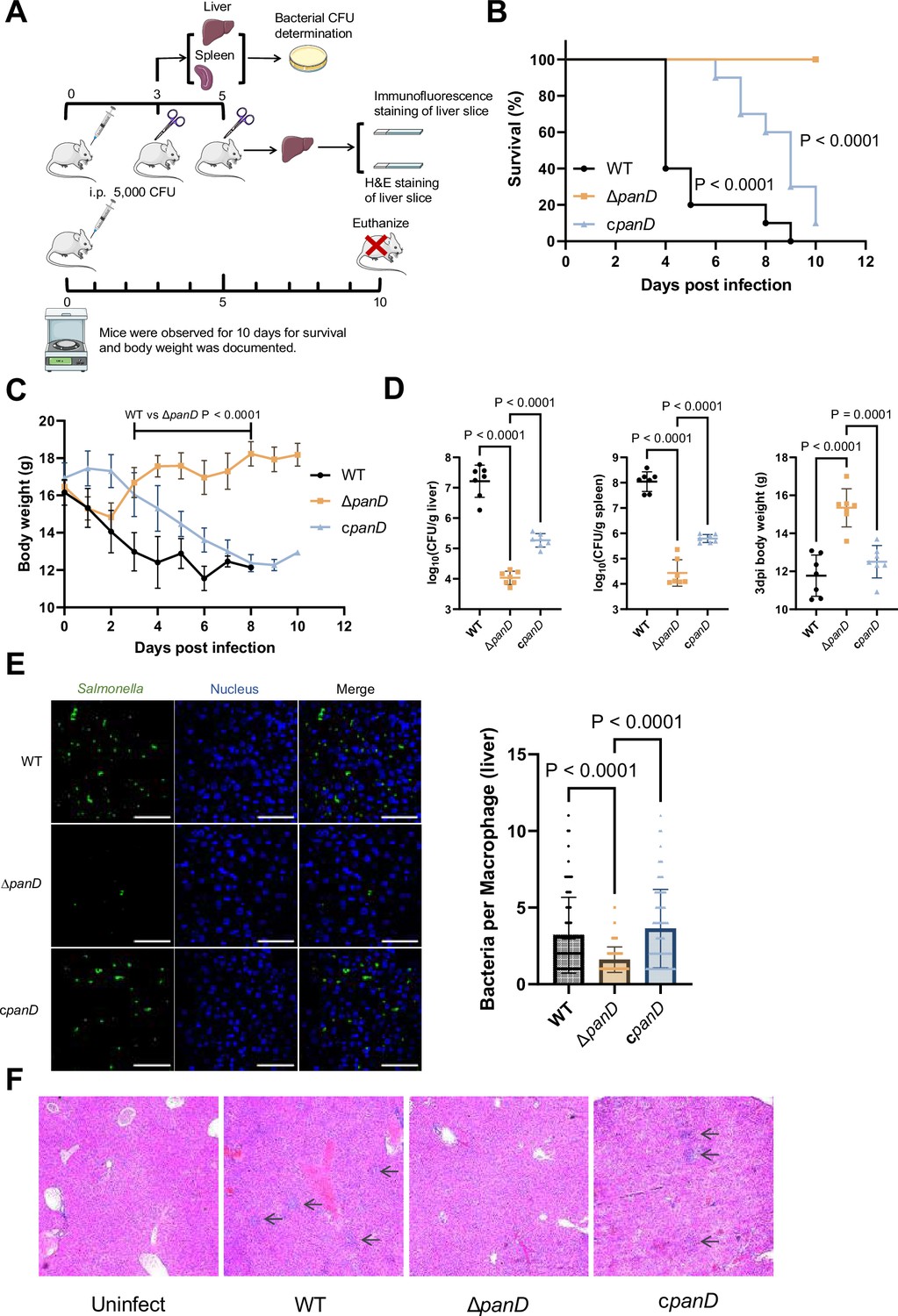
De novo β-alanine synthesis is critical for systemic Salmonella infection in mice.
(A) Schematic illustration of the mouse infection assays. Picture materials were used from bioicons (https://bioicons.com/). (B, C) Survival curves (B) and body weight dynamics (C) of mice infected i.p. with Salmonella wild-type (WT), ΔpanD, or cpanD. n=10 randomly assigned mice per group. (D) Liver and spleen bacterial burdens and body weights of mice infected with Salmonella WT, ΔpanD, or cpanD on day 3 post-infection. n=7 randomly assigned mice per group. (E) Representative immunofluorescence images and intracellular bacterial counts of Salmonella WT, ΔpanD, and cpanD in mouse liver at 5 d post-infection (green, Salmonella; blue, nuclei; scale bars, 50 µm). Images are representative of three independent experiments. The number of intracellular bacteria per infected cell was estimated in random fields, with n=80 cells per group from three independent experiments. (F) Representative H&E-stained liver sections from mice that were left uninfected or infected with Salmonella WT, ΔpanD, or cpanD on day 5 post-infection. Arrows indicate severe inflammatory cell infiltration in the mouse liver. Images are representative of three independent experiments. The data are presented as the mean ± SD (B–E). Statistical significance was assessed using the log-rank Mantel–Cox test (B), two-sided Student’s t-test (C), or one-way ANOVA (D, E).
-
Figure 3—source data 1
The numerical source data corresponds to Figure 3.
- https://cdn.elifesciences.org/articles/103714/elife-103714-fig3-data1-v1.xlsx
β-Alanine is involved in the regulation of several metabolic pathways in Salmonella
To explore the mechanism(s) associated with β-alanine-mediated promotion of Salmonella replication in macrophages and in vivo virulence, we performed RNA sequencing (RNA-seq) to reveal the differences in gene transcripts between Salmonella WT and ΔpanD strains grown in N-minimal medium. PCA plot of the global transcriptomic profiles clearly demonstrated separation between the WT and ΔpanD strains (Figure 4A). Remarkable transcriptional changes were observed due to the mutation of panD. Compared with those in the WT strain, 1379 genes were differentially expressed in the ΔpanD strain, with 561 upregulated genes and 618 downregulated genes (fold change ≥2 and p-value <0.05; Figure 4B). Gene Ontology (GO) enrichment analysis revealed that the differentially expressed genes (DEGs) were mainly involved in the metabolism and biosynthesis of several amino acids (including arginine, leucine, histidine, and branched amino acids), carboxylic acid metabolism, small molecule biosynthesis, and aerobic respiration (Figure 4C). Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis also revealed a high frequency of terms related to metabolism, including amino acid metabolism, lipid metabolism, carbohydrate metabolism, energy metabolism, and nucleotide metabolism (Figure 4D). These data collectively indicate that β-alanine is involved in the regulation of a series of metabolic pathways in Salmonella.
Figure 4 with 3 supplements see all
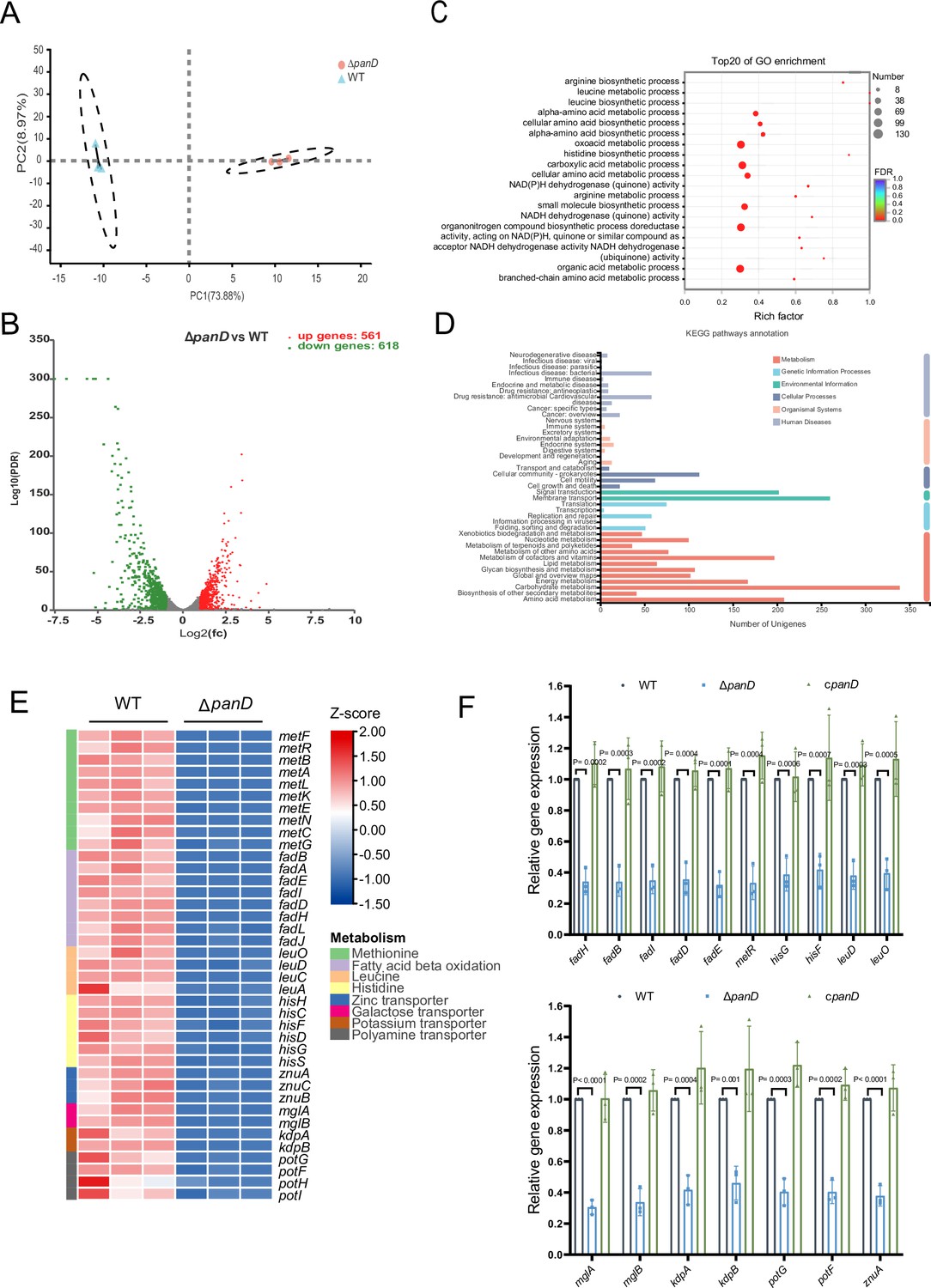
β-Alanine is involved in the regulation of several metabolic pathways in Salmonella.
(A) Principal component analysis (PCA) score plots of transcriptomic profiles of Salmonella wild-type (WT) and ΔpanD (n=3 biologically independent samples). (B) Volcano plot of the differentially expressed genes (DEGs) in Salmonella WT versus ΔpanD. The upper right section (red dots) indicates the upregulated DEGs, and the upper left section (green dots) indicates the downregulated DEGs. (C) Gene Ontology (GO) enrichment analysis of DEGs. Bubble chart showing the top 20 enriched Gene Ontology (GO) terms. (D) Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway enrichment analysis of DEGs. (E) Expression of the downregulated pathways (activated by PanD) is shown in the Z score-transformed heatmap, with red representing higher abundance and blue representing lower abundance. (F) Quantitative real-time PCR (qRT‒PCR) analysis of the mRNA levels of 16 selected downregulated DEGs in Salmonella WT, ΔpanD, and cpanD. The data are presented as the mean ± SD, n=3 independent experiments. Statistical significance was assessed using two-way ANOVA.
-
Figure 4—source data 1
The numerical source data corresponds to Figure 4.
- https://cdn.elifesciences.org/articles/103714/elife-103714-fig4-data1-v1.xlsx
Further analysis of the downregulated DEGs (activated by PanD) revealed that mutation of panD decreased the expression of genes involved in even pathways that are associated with the virulence of Salmonella or other bacterial pathogens, including methionine metabolism, fatty acid β-oxidation, histidine biosynthesis, and the transport of zinc, galactose, potassium, and polyamine (Figure 4E). Zinc and potassium uptake are associated with the virulence of Salmonella (zinc acquisition promotes Salmonella Typhimurium virulence in mice, and potassium acquisition promotes Salmonella Enteritidis virulence in chickens) (Ammendola et al., 2007; Battistoni et al., 2017; Ilari et al., 2016; Liu et al., 2013), while the remaining 5 pathways are involved in the pathogenicity of other pathogens (Basavanna et al., 2013; Paiva et al., 2016; Feldman et al., 2016; Lauriano et al., 2004; Martínez-Guitián et al., 2019). In addition, the expression of the LysR-type transcriptional regulator LeuO, which activates the expression of the leuABCD leucine synthesis operon and numerous virulence genes in Salmonella Typhimurium (Dillon et al., 2012; Guadarrama et al., 2014; Hernández-Lucas et al., 2008), was also downregulated in the ΔpanD strain (Figure 4E). In line with the decreased expression of leuO, the expression of leuABCD was downregulated in the ΔpanD strain (Figure 4E).
We selected 16 downregulated DEGs (including the regulatory gene leuO and genes from the above 7 pathways) for qRT‒PCR analysis. The results showed that the expression of all 16 genes significantly (p<0.05) decreased in the ΔpanD mutant compared with the WT strain (Figure 4F), and complementation of ΔpanD with panD restored the gene expression to the WT level (Figure 4F), thus confirming the positive regulation of these pathways and LeuO by β-alanine.
Salmonella pathogenesis largely depends on virulence genes encoded by Salmonella pathogenicity islands (SPIs), with SPI-1 to SPI-5 being well-characterized for their involvement in Salmonella virulence (Han et al., 2024). SPI-2 gene expression is essential for Salmonella replication in macrophages and systemic infection (Deng et al., 2017), yet the expression of SPI-2 genes was unaffected by panD mutation (Figure 4—figure supplement 1). Moreover, the gene expression of four other virulence-associated SPIs, SPI-1, SPI-3, SPI-4, and SPI-5, is also unaffected by panD mutation (Figure 4—figure supplements 2 and 3).
Taken together, these data suggest that β-alanine might promote Salmonella intracellular replication and virulence by activating virulence-associated pathway(s) or activating the virulence-associated regulator LeuO, rather than by activating the expression of virulence genes encoded within pathogenicity islands.
β-alanine promotes Salmonella virulence in vivo partially by increasing the expression of zinc transporter genes
Next, we inactivated the seven downregulated pathways mentioned above, as well as the regulatory gene leuO in Salmonella, to uncover the mechanism(s) by which β-alanine promotes Salmonella virulence in vivo. Mouse infection assays revealed that mutations in fadAB, metR, hisABCDFGHL, kdpABC, mglABC, and potFGHI, which are associated with fatty acid β-oxidation, methionine metabolism, histidine biosynthesis, potassium uptake, galactose uptake, and polyamine uptake, respectively, did not influence Salmonella colonization in the mouse liver or spleen (Figure 5A and B) or the body weight of infected mice (Figure 5C). Interestingly, although LeuO has been reported to be associated with the regulation of a diverse set of virulence factors (Dillon et al., 2012; Guadarrama et al., 2014), mutation of the regulatory gene leuO did not influence Salmonella colonization in the mouse liver or spleen (Figure 5A and B) or the body weight of infected mice (Figure 5C). In contrast, mutation of the zinc transporter gene znuA significantly decreased Salmonella colonization in the mouse liver and spleen (Figure 5D, left and middle panels); this result is consistent with previous studies (Ammendola et al., 2007; Battistoni et al., 2017; Ilari et al., 2016). Accordingly, the body weight of ΔznuA-infected mice was significantly (p<0.001) greater than that of the WT-infected mice (Figure 5D, right panel). These results indicate that β-alanine might promote Salmonella virulence in vivo by promoting zinc uptake.
Figure 5
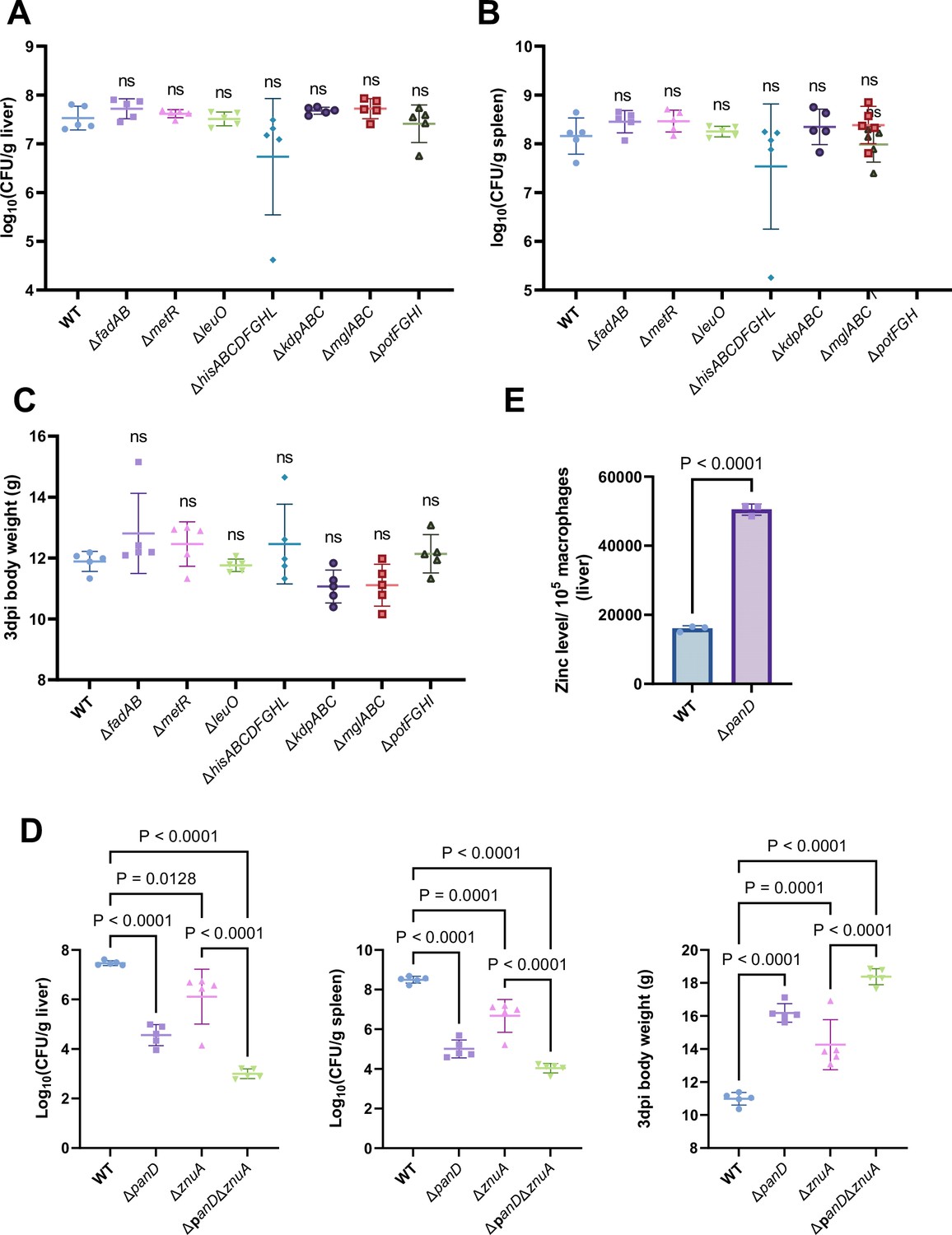
β-alanine promotes Salmonella virulence in vivo partially by increasing the expression of zinc transporter genes.
(A, B, C) Liver (A) and spleen (B) bacterial burdens and body weight (C) of mice infected with Salmonella wild-type (WT), ΔfadAB, ΔmetR, ΔhisABCDFGHL, ΔkdpABC, ΔmglABC, ΔpotFGHI, or ΔleuO on day 3 post-infection. n=5 mice per group. (D) Liver and spleen bacterial burdens and body weights of mice infected with Salmonella WT, ΔpanD, ΔznuA or ΔpanDΔznuA on day 3 post-infection. n=5 mice per group. (E) The zinc levels in the livers of mice infected with either Salmonella WT or ΔpanD for 3 d, n=5 mice per group. The data are presented as the mean ± SD (A–E). Statistical significance was assessed using one-way ANOVA (A-D), two-sided Student’s t-test (E). ns, not Significant.
-
Figure 5—source data 1
The numerical source data corresponds to Figure 5.
- https://cdn.elifesciences.org/articles/103714/elife-103714-fig5-data1-v1.xlsx
To test this hypothesis, we initially evaluated the zinc content in the livers of WT- and ΔpanD-infected mice, on day 3 post-infection. We observed that the zinc concentration in the macrophages of ΔpanD-infected mouse livers was 3.2-fold higher than in those of WT-infected mice (p<0.0001; Figure 5E), suggesting that the panD gene and β-alanine are crucial for Salmonella to obtain zinc from host cells.
Next, we constructed a double mutant, ΔpanDΔznuA, and compared colonization of the mouse liver and spleen of the double mutant to that of the single mutant, ΔznuA. The results showed that colonization of the liver and spleen of infected mice by ΔpanDΔznuA was significantly lower than that of infected mice colonized by ΔznuA (Figure 5D, left and middle panels). In agreement with these results, the body weight of ΔpanDΔznuA-infected mice was greater than that of ΔznuA-infected mice (Figure 5D, right panel), suggesting that the contribution of panD to the virulence of Salmonella is partially dependent on znuA.
Collectively, these data indicate that β-alanine promotes in vivo virulence of Salmonella partially by increasing the expression of zinc transporter genes.
β-alanine promotes Salmonella replication within macrophages partially by increasing the expression of zinc transporter genes
To determine whether β-alanine influences Salmonella intracellular replication by acting on zinc transporters, the zinc content in RAW 264.7 macrophages infected with WT and ΔpanD was also examined. We observed that the zinc concentration in ΔpanD-infected RAW 264.7 cells increased by 1.8-fold compared to WT-infected cells (p<0.0001; Figure 6A), further indicating the panD gene and β-alanine are crucial for Salmonella to absorb zinc from macrophages.
Figure 6
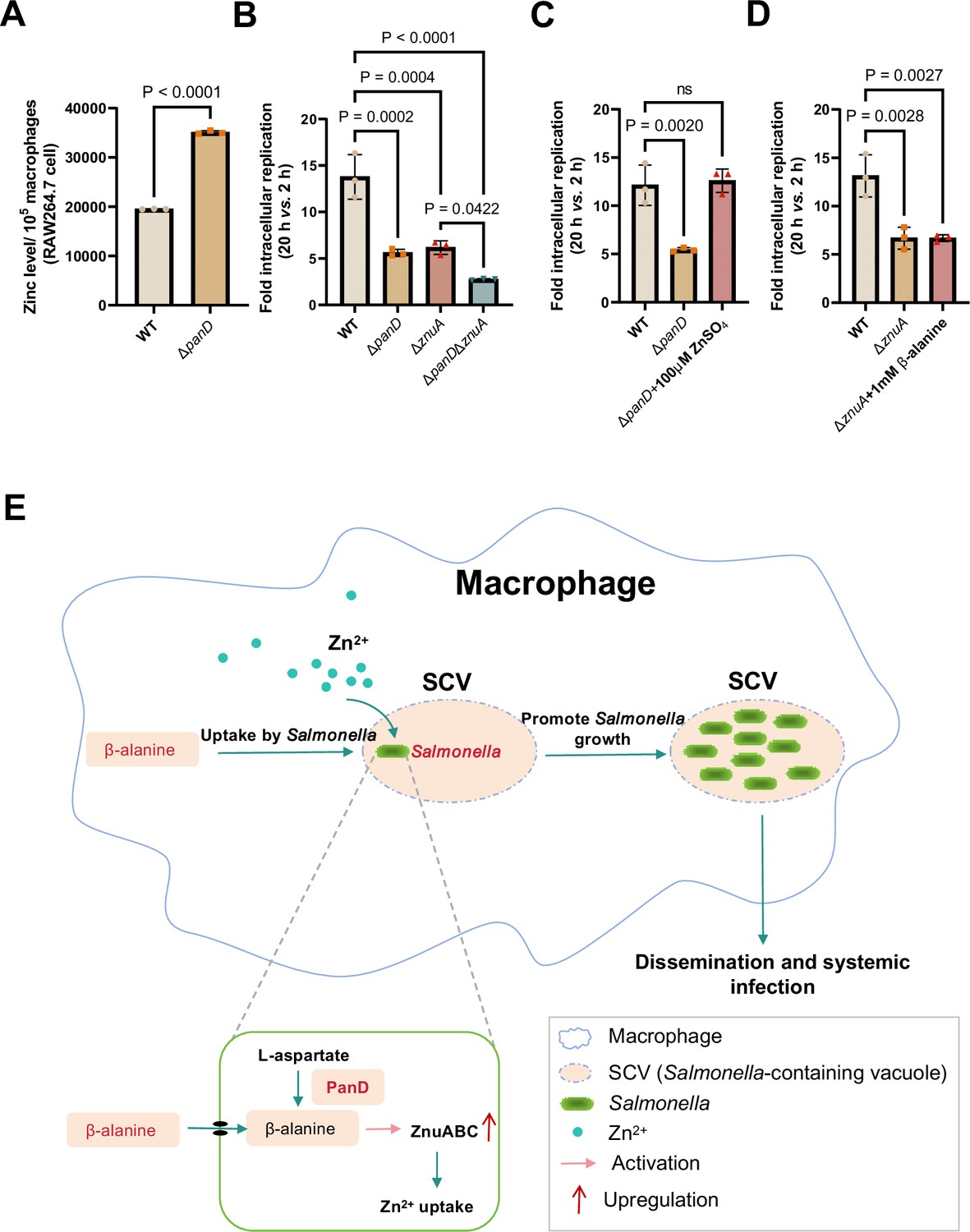
β-alanine promotes Salmonella replication within macrophages partially by increasing the expression of zinc transporter genes.
(A) The zinc levels in RAW264.7 cells after infection with Salmonella wild-type (WT) or ΔpanD for 8 hr. (B) Replication of Salmonella WT, ΔpanD, ΔznuA, and ΔpanDΔznuA in RAW264.7 cells. (C) Replication of Salmonella WT and ΔpanD in RAW264.7 cells in the presence or absence of 100 μM ZnSO4. (D) Replication of Salmonella WT and ΔznuA in RAW264.7 cells in the presence or absence of 1 mM β-alanine. The data are presented as the mean ± SD, n=3 independent experiments (A–D). Statistical significance was assessed using a two-sided Student’s t-test (A), one-way ANOVA (B-D). ns, not Significant. (E) Schematic model of β-alanine-mediated Salmonella replication inside macrophages. In macrophages, Salmonella acquires β-alanine both via the uptake of β-alanine from host macrophages and the de novo synthesis of β-alanine. β-alanine promotes the expression of zinc transporter genes ZnuABC, which facilitate the uptake of zinc by intracellular Salmonella, therefore, promote Salmonella replication in macrophages and subsequent systemic infection.
-
Figure 6—source data 1
The numerical source data corresponds to Figure 6.
- https://cdn.elifesciences.org/articles/103714/elife-103714-fig6-data1-v1.xlsx
We then analyzed the ability of ΔznuA and ΔpanDΔznuA to replicate in RAW264.7 macrophages via gentamicin protection assays. The results showed that the replication of ΔpanDΔznuA in RAW264.7 cells was significantly reduced compared with that of the single mutant ΔznuA (Figure 6B), implying that the contribution of panD to the intracellular replication of Salmonella is partially dependent on znuA. The addition of 100 μM zinc to RPMI medium increased the replication of ΔpanD in RAW264.7 cells (Figure 6C), while the addition of 1 mM β-alanine to RPMI medium did not increase the replication of ΔznuA (Figure 6D), suggesting that the impaired replication due to the decrease in β-alanine can be relieved by zinc supplementation.
Taken together, these data indicate that β-alanine promotes Salmonella replication within macrophages by increasing the expression of zinc transporter genes.
Discussion
Replication within host macrophages is a crucial step for Salmonella to cause life-threatening systemic infection in the host (Bomjan et al., 2019; Lathrop et al., 2015), while the crosstalk between Salmonella and macrophages at the metabolic interface is critical for intracellular Salmonella replication (Dandekar et al., 2014; Herrero-Fresno and Olsen, 2018; Lynch and Lesser, 2021; Rosenberg et al., 2021; Thompson et al., 2018). Emerging evidence suggests that several metabolites affect the replication of Salmonella in macrophages. The promotion of intracellular replication by metabolites is possibly achieved in three ways: (i) metabolites are utilized by Salmonella as nutrients for intracellular growth (Bowden et al., 2009; Eisenreich et al., 2010; Wang et al., 2021a); (ii) Salmonella senses metabolites as environmental cues to activate the expression of virulence genes (Jiang et al., 2021; Wang et al., 2023); and (iii) metabolites can regulate the immune responses of macrophages (Michelucci et al., 2013; Peace and O’Neill, 2022; Yang and Cong, 2021). In this study, we demonstrated that Salmonella promotes its replication inside macrophages by utilizing both host- and bacterial-derived β-alanine (Figure 6E). We showed that β-alanine promotes Salmonella intracellular replication and systemic infection partially by increasing the expression of zinc transporter genes and therefore, the uptake of zinc by intracellular Salmonella (Figure 6E). Therefore, this work identified another metabolite that can influence the replication of Salmonella in macrophages and illustrated the mechanism by which β-alanine promotes intracellular Salmonella replication.
We observed that Salmonella-infected macrophages contained lower β-alanine levels than mock-infected macrophages, while β-alanine supplementation in the cell medium increased the replication of Salmonella in macrophages, revealing that Salmonella uptakes host-derived β-alanine to promote intracellular replication. In addition, a deficiency in the biosynthesis of β-alanine (via mutation of the rate-limiting gene panD) reduced Salmonella replication in macrophages and systemic infection in mice, suggesting that Salmonella also utilizes bacterial-derived β-alanine to promote intracellular replication and pathogenicity. It is known that bacteria are quite stringent with their energy resources (Bergkessel, 2020; Bosdriesz et al., 2015), while the results of this work indicate that either acquisition from the host or de novo synthesis of β-alanine is critical for Salmonella replication inside macrophages. We speculate that Salmonella relies on a large amount of β-alanine to efficiently replicate in macrophages, thereby highlighting the importance of β-alanine for Salmonella intracellular growth. Nevertheless, unlike the closely related species E. coli, which takes up β-alanine via the transporter protein CycA (Schneider et al., 2004), Salmonella does not use CycA to uptake β-alanine. GabP, the GABA transporter, is potentially involved in the uptake of β-alanine in E. coli (Pavić et al., 2021). However, Salmonella does not utilize GabP to uptake β-alanine either. Ultimately, we revealed that ArgT, the transporter of arginine, is involved in the transport of β-alanine in Salmonella. ArgT has been reported to be essential for Salmonella replication within macrophages and for full virulence in vivo (Das et al., 2010). However, the attenuated virulence of the ∆argT mutant due to a deficiency in β-alanine or arginine requires further investigation.
Several amino acids, including lysine, proline, arginine, aspartate, and asparagine, have previously been reported to be associated with the pathogenicity of Salmonella (Popp et al., 2015; Steeb et al., 2013). These amino acids are involved in the synthesis of proteins in Salmonella. In contrast, β-alanine is not incorporated into proteins but can participate in the regulation of bacterial activity through the synthesis of pantothenate and CoA (Webb et al., 2004; White et al., 2001; Yuan et al., 2022). Accordingly, our transcriptome data showed that a deficiency in β-alanine biosynthesis affected the expression of Salmonella genes involved in a series of important metabolic pathways. Importantly, although β-alanine does not influence the gene expression of SPIs, it activates methionine metabolism; fatty acid β-oxidation; histidine biosynthesis; and the transport of zinc, galactose, potassium, and polyamine, which have been previously known to be associated with the virulence of Salmonella and other bacterial pathogens. Further analysis revealed that β-alanine promotes Salmonella intracellular replication and systemic infection partially by promoting the uptake of zinc. As for the other virulence-related metabolic pathways activated by β-alanine, methionine metabolism, fatty acid β-oxidation, and histidine biosynthesis contribute to the virulence of Streptococcus pneumoniae (Basavanna et al., 2013), Yersinia pestis (Feldman et al., 2016), and Acinetobacter baumannii (Martínez-Guitián et al., 2019), respectively; the uptake of galactose and polyamine influences the pathogenicity of Francisella tularensis (Lauriano et al., 2004) and the avian pathogenic Escherichia coli (Paiva et al., 2016), respectively; and the uptake of potassium is associated with the virulence of Salmonella Enteritidis in chickens (Liu et al., 2013). However, blocking these pathways did not influence the systemic Salmonella Typhimurium infection in mice, implying that different bacterial pathogens adopt different virulence strategies to establish infection. Determining other mechanism(s) by which β-alanine promotes the intracellular replication and systemic infection of Salmonella requires further investigation.
We observed that β-alanine also activates the expression of the LysR-type transcriptional regulator LeuO, which is known to regulate the expression of a wide variety of Salmonella genes that impact the stress response and virulence (Dillon et al., 2012; Guadarrama et al., 2014; Hernández-Lucas et al., 2008). Typically, LeuO activates the synthesis of the quiescent porins OmpS1 and OmpS2, which are required for Salmonella virulence in mice (De la Cruz et al., 2007; Fernández-Mora et al., 2004; Rodríguez-Morales et al., 2006). Consistent with the positive regulation of OmpS1 and OmpS2 by LeuO, lack of leuO in Salmonella also attenuated virulence in a mouse model (Rodríguez-Morales et al., 2006). However, the attenuated phenotypes of the leuO mutant in mice were not evident after i.p. injection relative to oral infection, as a previous report showed that the competitive index for the leuO mutant indicated approximately 1000-fold reduced colonization in mouse systemic tissues after oral infection but much less reduced colonization after i.p. injection (less than 10-fold) (Rodríguez-Morales et al., 2006). These results imply that LeuO might be predominantly associated with the invasion and intestinal infection of Salmonella but is weakly implicated in Salmonella intracellular replication and systemic infection. Therefore, it is not surprising that lack of leuO did not significantly affect Salmonella colonization in the systemic tissues of mice after i.p. injection, as revealed by our results.
Zinc is an essential micronutrient for all living organisms and is used as a cofactor for various enzymes and proteins (Bock et al., 2016; Ilari et al., 2016; Tan et al., 2004). In bacteria, zinc-binding proteins account for approximately 5% of the bacterial proteome and play crucial roles in bacterial metabolism and virulence (Andreini et al., 2006). Knockout of the zinc transporter ZnuABC reduces the virulence of Salmonella, Campylobacter jejuni, Haemophilus ducreyi, Moraxella, and urinary tract pathogenic Escherichia coli (UPEC) in the host (Ilari et al., 2016). Moreover, zinc is also utilized by Salmonella to subvert the antimicrobial host defense of macrophages by inhibiting NF-кB activation and impairing NF-кB-dependent bacterial clearance (Jennings et al., 2018; Wu et al., 2017). Therefore, the efficient acquisition of zinc may be crucial for Salmonella’s survival and replication within macrophages, where zinc availability is limited (Ammendola et al., 2007; Ilari et al., 2016). It has been reported that Salmonella utilizes the high-affinity ZnuABC zinc transporter to optimize zinc availability in host cells (Ammendola et al., 2007). In this study, we found that β-alanine can increase the expression of the zinc transporter genes znuABC, which could represent an additional mechanism for the efficient zinc uptake of Salmonella in macrophages. The results demonstrate a correlation between Salmonella β-alanine utilization and zinc uptake during intracellular infection and provide evidence that β-alanine can influence the macrophage immune response by acting on zinc uptake.
Overall, our findings suggest a model in which Salmonella exploits host- and bacterial-derived β-alanine to efficiently replicate in host macrophages and cause systemic disease. We propose that Salmonella requires a large amount of β-alanine during intracellular infection. The utilization of β-alanine promotes Salmonella uptake of the essential micronutrient zinc, which was previously shown to be required for the metabolic needs of intracellular Salmonella and to subvert the antimicrobial defense of macrophages by Salmonella. These observations provide new insight into Salmonella pathogenesis and the crosstalk between Salmonella and macrophages during intracellular infection. Considering that the panD gene is present in the genomes of all Salmonella strains and that mutation of panD markedly reduced Salmonella replication ability in macrophages, as well as virulence in the mouse model, this gene may be used as a potential target to control systemic Salmonella infection.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Biological sample (M. musculus) | BALB/c mice | Beijing Vital River Laboratory Animal Technology | Cat# 213 | 6-wk-old female |
| Cell line (M. musculus) | RAW264.7 | ATCC | Cat# TIB-71; RRID:CVCL_0493 | mouse macrophage-like cell line |
| Antibody | FITC-conjugated anti-Salmonella antibody (Rabbit polyclonal) | Abcam | Cat# ab20320; RRID:AB_445509 | IF (1:100) |
| Antibody | anti-CD86 antibody (Rat monoclonal) | Abcam | Cat# ab119857; RRID:AB_10902800 | Flow Cyt (1:100) |
| Antibody | anti-CD163 antibody (Rabbit monoclonal) | Abcam | Cat# ab182422; RRID:AB_2753196 | Flow Cyt (1:100) |
| Antibody | goat anti-rat IgG H&L (Alexa Fluor 488) antibody | Abcam | Cat# ab150165; RRID:AB_2650997 | Flow Cyt (1:100) |
| Antibody | donkey anti-rabbit IgG H&L (Alexa Fluor 647) antibody | Abcam | Cat# ab150075; RRID:AB_2752244 | Flow Cyt (1:100) |
| Chemical compound, drug | β-alanine | Solarbio | Cat# A9770 | N/A |
| Commercial assay or kit | EASYspinPlus bacterial RNA rapid extraction kit | Aidlab | Cat# RN0802 | N/A |
| Commercial assay or kit | 2×RealStar Power SYBR qPCR Mix | Genstar | Cat# A304 | N/A |
| Commercial assay or kit | StarScript III RT Kit | Genstar | Cat# A232 | N/A |
| Commercial assay or kit | ROS fluorescence probe (BBoxiProbe O06) | Bestbio | Cat# BB-46051 | N/A |
| Commercial assay or kit | RNS fluorescence probe (BBoxiProbe O52) | Bestbio | Cat# BB-470567 | N/A |
| Commercial assay or kit | gentleMACS/Mouse Liver Dissociation Kit | Miltenyi Biotec | Cat# 130-105-807 | N/A |
| Commercial assay or kit | Zinc fluorescence probe (Zinquin ethyl ester) | MKBio | Cat# MX4516 | N/A |
| Commercial assay or kit | Hematoxylin-Eosin(HE) staining kit | Sangon Biotech | Cat# E607318 | N/A |
| Software, algorithm | GraphPad Prism 9.5.1 | GraphPad | RRID:SCR_002798 | http://www.graphpad.com/ |
| Software, algorithm | Bowtie 2 | Bowtie | RRID:SCR_016368 | https://bowtie-bio.sourceforge.net/bowtie2/index.shtml |
| Software, algorithm | Xcalibur 4.0 | Thermo Fisher | RRID:SCR_014593 | https://www.thermofisher.cn/order/catalog/product/OPTON-30965?SID=srch-srp-OPTON-30965 |
| Software, algorithm | ZEN 2.3 ((blue edition)) | Carl Zeiss | RRID:SCR_013672 | https://www.zeiss.com/microscopy/en/products/software/zeiss-zen.html |
| Software, algorithm | Image J | National Institutes of Health | RRID:SCR_003070 | https://imagej.nih.gov/ij/ |
| Other | DAPI stain | Invitrogen | Cat# 21490 | N/A |
Ethics statement
Request a detailed protocolSix-wk-old female BALB/c mice were obtained from Beijing Vital River Laboratory Animal Technology (Beijing, China). Mice were housed in barrier facilities under specific pathogen-free conditions with a 12 hr light/dark cycle at a temperature of 24 ± 2°C and a relative humidity of 50 ± 5%. Mice were fed a standard mouse chow diet, and they consumed food and water ad libitum throughout the experiment. All animal experiments were conducted in accordance with the policies of the Institutional Animal Care Committee of Nankai University (Tianjin, China) and performed under protocol no. 2021-SYDWLL-000029.
Cell culture
Request a detailed protocolThe RAW264.7 mouse macrophage-like cell line (ATCC TIB-71) was obtained from the Shanghai Institute of Biochemistry and Cell Biology of the Chinese Academy of Sciences (Shanghai, China). This cell line originates from ATCC and is not included in the list of commonly misidentified cell lines. Additionally, this cell line is free of mycoplasma contamination. Cells were cultured in RPMI-1640 medium (Gibco #11879020) supplemented with 10% (v/v) fetal bovine serum (FBS, Gibco #10100147) at 37℃ with 5% CO2. Cells were seeded in 24-well tissue culture plates at 1×105 cells per well 24 hr before infection.
Bacterial strains, plasmids, and growth conditions
Request a detailed protocolThe bacterial strains and plasmids used in this study are listed in Supplementary file 1. The Salmonella enterica serovar Typhimurium (STM) strain ATCC 14,028 s was used as the WT strain throughout this study and for the construction of the mutants. Mutant strains were generated using the λ Red recombination system with the plasmid pSIM17 (Jiang et al., 2017). To construct the complemented strain of ΔpanD, the amplified DNA fragments of the panD ORF and its upstream promoter were digested and inserted into the BamHI–EcoRI site of the low-copy-number plasmid pBR322. To generate the panD-lux reporter fusion, the amplification products of the panD promoter region were digested and cloned into the XhoI–BamHI site of the plasmid pMS402, which carries a promoter-less luxCDABE reporter gene cluster (Liang et al., 2008). The sequences of primers used for the construction of the strains are listed in Supplementary file 2. All the strains were verified by PCR amplification and sequencing.
Bacterial strains were conventionally grown overnight in Luria–Bertani (LB) medium (10 g/L tryptone, 5 g/L yeast extract, and 10 g/L NaCl) or in N-minimal medium (10 μM MgCl2, 110 μM KH2PO4, 7.5 mM (NH4)2SO4, 0.5 mM K2SO4, 5 mM KCl, 38 mM glycerol, and 0.1% [w/v] casamino acids) supplemented with appropriate antibiotics at 37 °C with shaking at 180 rpm or on LB agar plates. All antibiotics were used at their standard concentrations (chloramphenicol, 25 μg/mL; kanamycin, 50 μg/mL; ampicillin, 100 μg/mL; gentamicin, 10 or 100 μg/mL) unless otherwise mentioned.
Growth curve
Request a detailed protocolBacterial strains were conventionally grown overnight in LB medium. The next day, they were subcultured (1:100) in new LB medium and RPMI-1640 medium or subcultured in N-minimal medium supplemented with glycerol or β-alanine as the sole carbon source at 37 °C with shaking at 180 rpm. To measure the growth of bacteria, 200 μL of the bacterial cultures were transferred to the microplate wells. The absorbance (OD600) of the bacteria was measured every half hour for 12 hr with a Spark multimode microplate reader (Tecan).
Bioluminescent reporter assays
Request a detailed protocolSTM WT carrying the panD-luxCDABE fusion plasmid was conventionally grown overnight in LB medium, and the next day, the cells were subcultured (1:100) in new LB medium or N-minimal medium for 8 hr. The luminescence of the cultured bacteria (200 μl) was measured with a Spark multimode microplate reader (Tecan). Moreover, the cultured bacteria (100 μl) were serially diluted and plated on LB agar plates to estimate bacterial CFUs. Bacterial CFUs were used to normalize luminescence values.
Salmonella infection of macrophages
Request a detailed protocolBacterial strains were conventionally grown overnight in LB medium to the late stationary phase, and the next day, the bacteria were diluted to 2×106 CFUs/mL and opsonized in RPMI-1640 medium supplemented with 10% FBS for 15 min. The macrophage monolayers were infected with the opsonized bacteria culture (0.5 mL/well, multiplicity of infection (MOI)=10) and centrifuged at 800×g for 5 min to synchronize infection. The infected cells were incubated for 30 min at 37 °C in 5% CO2 and then washed three times with 1×PBS. Fresh RPMI-1640 medium containing 100 μg/mL gentamicin was added to the infected cells to kill extracellular bacteria. After 1 hr, fresh RPMI-1640 medium containing 10 μg/mL gentamicin was added to the infected cells for the remainder of the experiment. To assess the intracellular growth of Salmonella, the infected cells were lysed with 1% Triton X-100 at 2 hpi and 20 hpi, and the abundance of the intracellular bacteria CFUs was estimated on LB agar plates. The relative fold replication of intracellular bacterial strains was denoted as the CFUs recovered at 20 hpi relative to those at 2 hpi. The relative fold change in replication was normalized to the number of RAW264.7 cells. When indicated, 1 mM β-alanine or 100 μM ZnSO4 were added after 1 hr of gentamicin treatment.
Targeted metabolomics analysis of amino acids in macrophages
Request a detailed protocolTargeted metabolomics was conducted as previously described (Jiang et al., 2021), with the combined metabolites of infected cells and intracellular bacteria extracted for analysis. The metabolites have been confirmed to be dominated by the host metabolites (Jiang et al., 2021). RAW264.7 cells were mock-infected or infected with STM WT for 8 hr. The cells were harvested and washed with precooled PBS solution to remove the medium. Cellular metabolites were extracted using ice-cold extraction solvent (40:40:20 vol/vol/vol acetonitrile:methanol:water, 0.1 M formic acid), incubated at –20℃ for 20 min, and then centrifuged for 10 min at 12,000×g and 4℃ to obtain the supernatant. Subsequently, the supernatant was transferred to an LC-MS vial and analyzed using ultrahigh-performance liquid chromatography (Acquity; Waters, Milford, MA, USA) coupled with mass spectrometry (Q Exactive Hybrid Quadrupole-Orbitrap; Thermo Fisher Scientific, Waltham, MA, USA). Metabolites were separated with a Luna NH2 column (2 mm × 100 mm, 3 µm particle size; Phenomenex). Mobile phase A was 20 mM ammonium acetate (pH 9.0), and mobile phase B was acetonitrile containing 0.1% formic acid. The flow rate was 0.4 ml/min. Xcalibur 4.0 software (Thermo Fisher) was used for data acquisition and processing. Metabolite identification was achieved by high-resolution mass and retention time matching to authentic standards. The absolute quantification of amino acids was performed using the standard curve method, and the values were normalized to the cell number. Four biological replicates of each sample were analyzed.
Immunofluorescence staining
Request a detailed protocolRAW264.7 cells were infected with STM WT or ΔpanD and the complemented strain cpanD as described above. After 2 and 20 hr of cultivation, the infected cells were fixed for 15 min in 4% paraformaldehyde, washed in PBS, and permeabilized with 0.1% Triton X-100 in PBS for 15 min. The fixed samples were blocked in 5% bovine serum albumin for 30 min, followed by staining with a FITC-conjugated anti-Salmonella antibody (1:100 dilution, Abcam #ab20320) for 1 hr at room temperature in the dark. The nuclei were then stained with DAPI (Invitrogen #D21490) for 2 min. A confocal laser scanning microscope (Zeiss LSM800) and ZEN 2.3 software (blue edition) were used to acquire and analyze the cell images (Objective lens: 40x; The number of intracellular bacteria per infected cell was estimated in random fields by Fiji-ImageJ).
RNA isolation
Request a detailed protocolRNA was extracted from Salmonella strains cultured in N-minimal medium or LB medium. To investigate the expression of the Salmonella panD gene inside macrophages, we obtained RNA from intracellular bacteria in RAW264.7 cells at 8 hr post-infection and from bacteria in RPMI medium. RNA was extracted using an EASYspinPlus bacterial RNA rapid extraction kit (Aidlab #RN0802) according to the manufacturer’s protocol. RNA quantity and purity were determined using a NanoDrop 2000 spectrophotometer (NanoDrop Technologies). RNA samples were stored at −80 °C before use.
Quantitative real-time PCR (qRT‒PCR)
Request a detailed protocolAccording to the manufacturer’s protocols, qRT‒PCR was performed using 2x RealStar Power SYBR qPCR Mix (Genstar #A304) in a QuantStudio 5 real-time PCR system (Applied Biosystems). cDNA was synthesized using a StarScript III RT Kit (Genstar #A232). Each sample was subjected to qRT‒PCR in triplicate. The expression level of the 16 S rRNA gene was used to normalize that of the target genes. We estimated the expression of each target gene using the 2−ΔΔCt method.
RNA sequencing and analyses
Request a detailed protocolThe STM WT and ΔpanD strains were conventionally grown overnight in LB medium, subcultured (1:100) in N-minimal medium for 8 hr, and then collected by centrifugation for RNA extraction. Sequencing libraries were generated using the NEBNext Ultra RNA Library Prep Kit for Illumina (New England Biolabs) according to the manufacturer’s instructions, and sequencing was conducted using the Illumina HiSeq 2000 platform at Shanghai Majorbio Bio-Pharm Technology Co., Ltd. (Shanghai, China). The sequencing data were deposited in the NCBI Sequence Read Archive under accession number (SRA, PRJNA1124637). The clean reads were mapped to the STM ATCC 14028 reference genome (CP001363 and CP001362) by using the short-sequence alignment software Bowtie 2. Gene expression was evaluated using the fragments per kilobase of transcript per million mapped reads (FPKM) method. DEGs in the panD mutant relative to the WT were determined using the R statistical package software EdgeR. The thresholds for statistically significant differences were set to a fold change ≥ 2 or ≤ 0.5 and a false discovery rate (FDR) ≤ 0.05. p-values were adjusted using the Benjamini–Hochberg procedure for controlling the FDR. Enrichment analysis of DEGs was conducted using GO and KEGG enrichment analyses.
Mouse infection
Request a detailed protocolSalmonella strains were conventionally cultured overnight in LB medium, and the next day, they were subcultured (1:100) in new LB medium and grown at 37 °C with shaking at 200 rpm to stationary phase (OD600~2). The bacteria were diluted to 5×104 CFUs/mL in 0.9% NaCl. Groups of BALB/c mice were infected i.p. with 0.1 mL of the NaCl suspension. For survival assays, we recorded and monitored the mortality and body weight of the infected mice daily for 10 d. To analyze the bacterial burden of the mouse liver and spleen, we weighed the infected mice first on day 3 post-infection and then harvested the liver and spleen. The liver and spleen of infected mice were homogenized in ice-cold PBS, serially diluted, and plated on LB plates containing the appropriate antibiotics to determine bacterial CFUs. H&E staining of the mouse liver was performed to investigate the histopathological alterations in the liver of infected mice. To evaluate the histopathological alterations in the mouse liver, we harvested the liver of the infected mice on day 5 post-infection. The mouse liver was washed with 0.9% NaCl, fixed with 10% neutral formalin for 48 hr, and subsequently processed for routine paraffin embedding. Paraffin-embedded tissues were sectioned at a thickness of 5 μm and then stained with hematoxylin (Sangon Biotech #E607318) and eosin (Sangon Biotech #E607318) for histopathological examination. The stained sections were then examined by light microscopy (Leica DM2500 LED).
Flow cytometry
Request a detailed protocolRAW264.7 cells were infected with Salmonella WT for 8 hr, in the absence or presence of 1 mM β-alanine, which was added to the infected cells at 1 hr post-infection. The infected cells were fixed for 15 min in 4% paraformaldehyde, washed in PBS, and permeabilized with 0.1% Triton X-100 in PBS for 15 min. The fixed samples were blocked in 5% bovine serum albumin for 30 min, followed by staining with an anti-CD86 antibody (Abcam, #ab119857), an anti-CD163 antibody (Abcam, #ab182422) for 30 min, and a goat anti-rat IgG H&L (Alexa Fluor 488) (Abcam, #ab150165), a donkey anti-rabbit IgG H&L (Alexa Fluor 647) (Abcam, #ab150075) for 30 min in the dark. The infected cells were analyzed using a BD FACSAria Flow Cytometer (BD Biosciences).
Assessment of intracellular ROS/ RNS levels
Request a detailed protocolRAW264.7 cells were infected with Salmonella WT for 8 hr, either in the absence or presence of 1 mM β-alanine. The β-alanine was added to the infected cells at 1 hr post-infection. Following the infection, the cells were washed three times with PBS, and the levels of ROS and RNS were measured using a ROS fluorescence probe (BBoxiProbe O06, Bestbio, #BB-46051) and an RNS fluorescence probe (BBoxiProbe O52, Bestbio, #BB-470567), respectively, in accordance with the manufacturer’s protocol. The ROS/RNS levels were normalized to the cell count.
Assessment of zinc levels in mouse liver and RAW 264.7 macrophages
Request a detailed protocolTo assess the zinc level in mouse liver, mice were i.p. infected with approximately 5000 CFUs of Salmonella WT or ΔpanD for 3 d, with three mice in each injection group. Following infection, mouse livers were collected and then dissociated into single cells using the gentleMACS/Mouse Liver Dissociation Kit (Miltenyi Biotec, #130-105-807) in accordance with the manufacturer’s protocol. The zinc concentration in mouse liver was determined using a zinc fluorescence probe (Zinquin ethyl ester, MKBio, #MX4516) following the manufacturer’s instructions and normalized by the cell count.
To assess the zinc level in RAW 264.7 macrophages, RAW 264.7 cells were infected with Salmonella WT or ΔpanD for 8 hr. The infected cells were washed three times with PBS, and zinc concentration was measured using a zinc fluorescence probe (Zinquin ethyl ester, MKBio, MX4516) following the manufacturer’s protocol. The zinc concentration was normalized by the cell count.
Statistical analysis
Request a detailed protocolThe data are presented as the mean ± SD. All in vitro experiments were conducted in duplicate and repeated at least three times (n≥3). Mouse assays were performed twice, with at least two mice (n≥2) in each injection group, and the combined data from the two experiments was used for statistical analysis. Statistical analyses were performed using GraphPad InStat software (version GraphPad Prism 9.5.1, San Diego, CA, USA) with two-sided Student’s t-tests, one-way ANOVA, two-way ANOVA, log-rank Mantel–Cox tests, or Mann‒Whitney U tests according to the test requirements (as stated in the figure legends). A p-value <0.05 indicated a statistically significant difference. ns represents no statistical significance.
Data availability
The RNA-seq data generated in this study have been deposited in the NCBI Sequence Read Archive (SRA) database under the accession number PRJNA1124637. All other data associated with this study are available in the main text and supplementary materials. Source data are provided in supplementary data.
-
NCBI BioProjectID PRJNA1124637. A Study on the Pathogenic Mechanism of Salmonella.
References
-
Zinc through the three domains of lifeJournal of Proteome Research 5:3173–3178.https://doi.org/10.1021/pr0603699
-
A novel antimicrobial approach based on the inhibition of zinc uptake in Salmonella entericaFuture Medicinal Chemistry 9:899–910.https://doi.org/10.4155/fmc-2017-0042
-
The biosynthesis of coenzyme A in bacteriaVitamins and Hormones 61:157–171.https://doi.org/10.1016/s0083-6729(01)61005-7
-
β-Alanine supplementation for athletic performance: an updateJournal of Strength and Conditioning Research 28:1751–1770.https://doi.org/10.1519/JSC.0000000000000327
-
Regulation of protein biosynthetic activity during growth arrestCurrent Opinion in Microbiology 57:62–69.https://doi.org/10.1016/j.mib.2020.07.010
-
Crystal structure of AibC, a reductase involved in alternative de novo isovaleryl coenzyme A biosynthesis in Myxococcus xanthusActa Crystallographica. Section F, Structural Biology Communications 72:652–658.https://doi.org/10.1107/S2053230X16011146
-
Physiology and pathophysiology of carnosinePhysiological Reviews 93:1803–1845.https://doi.org/10.1152/physrev.00039.2012
-
YshB promotes intracellular replication745 and is required for Salmonella virulenceJournal of Bacteriology 201:e00314-19.https://doi.org/10.1128/JB.00314-19
-
Salmonella populations inside host cellsFrontiers in Cellular and Infection Microbiology 7:432.https://doi.org/10.3389/fcimb.2017.00432
-
Salmonella-how a metabolic generalist adopts an intracellular lifestyle during infectionFrontiers in Cellular and Infection Microbiology 4:191.https://doi.org/10.3389/fcimb.2014.00191
-
Signalling functions of coenzyme A and its derivatives in mammalian cellsBiochemical Society Transactions 42:1056–1062.https://doi.org/10.1042/BST20140146
-
Host-pathogen interaction in invasive salmonellosisPLOS Pathogens 8:e1002933.https://doi.org/10.1371/journal.ppat.1002933
-
LeuO antagonizes H-NS and StpA-dependent repression in Salmonella enterica ompS1Molecular Microbiology 66:727–743.https://doi.org/10.1111/j.1365-2958.2007.05958.x
-
Assembly, structure, function and regulation of type III secretion systemsNature Reviews. Microbiology 15:323–337.https://doi.org/10.1038/nrmicro.2017.20
-
LeuO is a global regulator of gene expression in Salmonella enterica serovar typhimuriumMolecular Microbiology 85:1072–1089.https://doi.org/10.1111/j.1365-2958.2012.08162.x
-
Carbon metabolism of intracellular bacterial pathogens and possible links to virulenceNature Reviews. Microbiology 8:401–412.https://doi.org/10.1038/nrmicro2351
-
Salmonella enterica serovar typhimurium skills to succeed in the host: virulence and regulationClinical Microbiology Reviews 26:308–341.https://doi.org/10.1128/cmr.00066-12
-
A high-coverage Yersinia pestis genome from a sixth-century justinianic plague victimMolecular Biology and Evolution 33:2911–2923.https://doi.org/10.1093/molbev/msw170
-
OmpR and LeuO positively regulate the Salmonella enterica serovar Typhi ompS2 porin geneJournal of Bacteriology 186:2909–2920.https://doi.org/10.1128/jb.186.10.2909-2920.2004
-
Coenzyme A: a protective thiol in bacterial antioxidant defenceBiochemical Society Transactions 47:469–476.https://doi.org/10.1042/BST20180415
-
Infection biology of Salmonella entericaEcoSal Plus 12:eesp00012023.https://doi.org/10.1128/ecosalplus.esp-0001-2023
-
Effects of β-alanine supplementation on carnosine elevation and physiological performanceAdvances in Food and Nutrition Research 84:183–206.https://doi.org/10.1016/bs.afnr.2017.12.003
-
Structure-function analyses of the bacterial zinc metalloprotease effector protein GtgA uncover key residues required for deactivating NF-κBThe Journal of Biological Chemistry 293:15316–15329.https://doi.org/10.1074/jbc.RA118.004255
-
Eat in or take out? Metabolism of intracellular Salmonella entericaTrends in Microbiology 28:644–654.https://doi.org/10.1016/j.tim.2020.03.005
-
Spatial and temporal patterns of typhoid and paratyphoid fever outbreaks: a worldwide review, 1990-2018Clinical Infectious Diseases 69:S499–S509.https://doi.org/10.1093/cid/ciz705
-
Salmonellae interactions with host processesNature Reviews. Microbiology 13:191–205.https://doi.org/10.1038/nrmicro3420
-
Replication of Salmonella enterica serovar typhimurium in human monocyte-derived macrophagesInfection and Immunity 83:2661–2671.https://doi.org/10.1128/IAI.00033-15
-
Strategies adopted by Salmonella to survive in host: a reviewArchives of Microbiology 205:362.https://doi.org/10.1007/s00203-023-03702-w
-
The global burden of nontyphoidal Salmonella gastroenteritisClinical Infectious Diseases 50:882–889.https://doi.org/10.1086/650733
-
Involvement of HisF in the persistence of Acinetobacter baumannii during a pneumonia infectionFrontiers in Cellular and Infection Microbiology 9:310.https://doi.org/10.3389/fcimb.2019.00310
-
Salmonella: a model for bacterial pathogenesisAnnual Review of Medicine 52:259–274.https://doi.org/10.1146/annurev.med.52.1.259
-
In vivo influence of in vitro up-regulated genes in the virulence of an APEC strain associated with swollen head syndromeAvian Pathology: Journal of the W.V.P.A 45:94–105.https://doi.org/10.1080/03079457.2015.1125995
-
The role of itaconate in host defense and inflammationThe Journal of Clinical Investigation 132:e148548.https://doi.org/10.1172/JCI148548
-
Role of host cell-derived amino acids in nutrition of intracellular Salmonella entericaInfection and Immunity 83:4466–4475.https://doi.org/10.1128/IAI.00624-15
-
Comprehensive single cell analyses of the nutritional environment of intracellular Salmonella entericaFrontiers in Cellular and Infection Microbiology 11:624650.https://doi.org/10.3389/fcimb.2021.624650
-
Immunometabolic crosstalk during bacterial infectionNature Microbiology 7:497–507.https://doi.org/10.1038/s41564-022-01080-5
-
Identification and characterization of the main beta-alanine uptake system in Escherichia coliApplied Microbiology and Biotechnology 65:576–582.https://doi.org/10.1007/s00253-004-1636-0
-
Coenzyme A: to make it or uptake it?Nature Reviews. Molecular Cell Biology 17:605–606.https://doi.org/10.1038/nrm.2016.110
-
Advances in the synthesis of β-alanineFrontiers in Bioengineering and Biotechnology 11:1283129.https://doi.org/10.3389/fbioe.2023.1283129
-
Effect of Zn on acetyl coenzyme a synthase: evidence for a conformational change in the alpha subunit during catalysisJournal of the American Chemical Society 126:5954–5955.https://doi.org/10.1021/ja039600z
-
Coenzyme A and its derivatives: renaissance of a textbook classicBiochemical Society Transactions 42:1025–1032.https://doi.org/10.1042/BST20140176
-
The metabolic pathways utilized by Salmonella typhimurium during infection of host cellsEnvironmental Microbiology Reports 10:140–154.https://doi.org/10.1111/1758-2229.12628
-
How to do business with lysosomes: Salmonella leads the wayCurrent Opinion in Microbiology 47:1–7.https://doi.org/10.1016/j.mib.2018.10.003
-
Salmonella enterica serovar typhi induces host metabolic reprogramming to increase glucose availability for intracellular replicationInternational Journal of Molecular Sciences 22:10003.https://doi.org/10.3390/ijms221810003
-
Advances in biotechnological production of β-alanineWorld Journal of Microbiology & Biotechnology 37:79.https://doi.org/10.1007/s11274-021-03042-1
-
A Salmonella typhimurium strain defective in uracil catabolism and beta-alanine synthesisJournal of General Microbiology 131:1083–1090.https://doi.org/10.1099/00221287-131-5-1083
-
Saccharomyces cerevisiae is capable of de novo pantothenic acid biosynthesis involving a novel pathway of beta-alanine production from spermineThe Journal of Biological Chemistry 276:10794–10800.https://doi.org/10.1074/jbc.M009804200
-
Gut microbiota-derived metabolites in the regulation of host immune responses and immune-related inflammatory diseasesCellular & Molecular Immunology 18:866–877.https://doi.org/10.1038/s41423-021-00661-4
Article and author information
Author details
Funding
National Natural Science Foundation of China (32170110)
- Lingyan Jiang
Natural Science Foundation of Tianjin Municipality (grant no. 22JCYBJC01060)
- Lingyan Jiang
Fundamental Research Funds for the Central Universities (63241588)
- Lingyan Jiang
National Natural Science Foundation of China (32270191)
- Lingyan Jiang
Fundamental Research Funds for the Central Universities (63243161)
- Lingyan Jiang
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
This work was supported by the National Natural Science Foundation of China (NSFC) Program (grant no. 32170110, 32270191), the Natural Science Foundation of Tianjin (grant no. 22JCYBJC01060), and the Fundamental Research Funds for the Central Universities, Nankai University (grant no. 63241588, 63243161).
Ethics
All animal experiments were conducted in accordance with the policies of the Institutional Animal Care Committee of Nankai University (Tianjin, China) and performed under protocol no. 2021-SYDWLL-000029.
Version history
- Preprint posted:
- Sent for peer review:
- Reviewed Preprint version 1:
- Reviewed Preprint version 2:
- Reviewed Preprint version 3:
- Version of Record published:
Cite all versions
You can cite all versions using the DOI https://doi.org/10.7554/eLife.103714. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2024, Ma, Yang et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,839
- views
-
- 116
- downloads
-
- 5
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 2
- citations for umbrella DOI https://doi.org/10.7554/eLife.103714
-
- 3
- citations for Version of Record https://doi.org/10.7554/eLife.103714.4
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Salmonella exploits host- and bacterial-derived β-alanine for replication inside host macrophages
eLife 13:RP103714.
https://doi.org/10.7554/eLife.103714.4




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}