NR2F2 is required in the embryonic testis for fetal Leydig cell development
- Université Côte d'Azur, CNRS, INSERM, iBV, France
eLife Assessment
This important study, which has been improved further upon revision, reveals a critical role of the transcription factor NR2F2 in mouse fetal Leydig cell (FLC) differentiation. With elegantly carried out experiments, the authors provide compelling evidence that NR2F2 helps to initiate the differentiation of certain interstitial cells into FLC until these cells mature into functional secretory cells that produce androgen and insulin-like peptide 3 (INSL3). The particular importance of the work comes from the fact that NR2F2 affects FLCs without altering paracrine signals known to be involved in FLC differentiation. The work will be of interest to colleagues studying reproductive development in mammals including humans or the biological functions of the nuclear receptor family.
https://doi.org/10.7554/eLife.103783.3.sa0Significance of the findings:
Important: Findings that have theoretical or practical implications beyond a single subfield
- Landmark
- Fundamental
- Important
- Valuable
- Useful
Strength of evidence:
Compelling: Evidence that features methods, data and analyses more rigorous than the current state-of-the-art
- Exceptional
- Compelling
- Convincing
- Solid
- Incomplete
- Inadequate
During the peer-review process the editor and reviewers write an eLife Assessment that summarises the significance of the findings reported in the article (on a scale ranging from landmark to useful) and the strength of the evidence (on a scale ranging from exceptional to inadequate). Learn more about eLife Assessments
Abstract
Male genital development in XY mammalian fetuses is triggered by the action of hormones, including testosterone, secreted by the developing testes. Defects in this process are a cause for differences in sex development (DSD), one of the most common congenital abnormalities in humans. Fetal Leydig cells (FLCs) play a central role in the synthesis of masculinizing hormones in the developing testes. Yet, the genetic cascade controlling their differentiation is poorly understood. Here, we investigate the role of the orphan nuclear receptor NR2F2 (COUP-TFII) in FLC development. We report that NR2F2 is expressed in interstitial progenitor cells of the mouse embryonic testes and is downregulated upon their differentiation into FLC. By using two mouse models for conditional mutation of Nr2f2 in the developing testes, we demonstrate that NR2F2 is required for testis morphogenesis and FLC development. NR2F2 acts in interstitial progenitors to regulate the initiation and progression of FLC differentiation. These results establish NR2F2 as an essential regulator of FLC development and steroid hormone synthesis in the mouse fetal testis and provide an entry point in understanding the etiology of 46,XY DSD associated with pathogenic NR2F2 variants.
Introduction
Sexual development in mammals is conditioned by the gonadal sex established during fetal life. In XY embryos, the initially undifferentiated gonads develop into testes that can produce testosterone which stimulates the differentiation of male internal and external genitalia (including epididymis, vas deferens, seminal vesicles, scrotum, and penis) leading to the masculinization of the fetus. In contrast, fetal ovaries do not produce testosterone, and female internal and external genitalia (oviducts, uterus, vagina, and vulva) develop in XX individuals. Abnormalities in the masculinization process are mainly associated with defects in androgen synthesis or signaling (Reyes et al., 2023). Although this molecular cascade is well defined, the differentiation of androgen-producing cells in the embryo is only partially understood.
Androgen production in the developing testis relies mainly on fetal Leydig cells (FLCs), which express all enzymes required for the biosynthesis of androstenedione from cholesterol (Miyabayashi et al., 2017; Inoue et al., 2016; Shima et al., 2013; Ademi et al., 2022). The final step of conversion of androstenedione into testosterone, catalyzed by HSD17B3, takes place in a different cell type, the fetal Sertoli cells (Shima et al., 2013; O’Shaughnessy et al., 2000). In addition to their role in androgen synthesis, FLCs also produce INSL3, a hormone required for testis descent (Nef and Parada, 1999; Zimmermann et al., 1999). Defects in testis descent, regulated also by androgens, result in cryptorchidism, a condition which impacts fertility and constitutes a risk factor for testicular cancer (Hutson et al., 2015). While a fraction of FLCs persist after birth, others de-differentiate or involute, and adult testosterone production is ensured by a distinct population of steroidogenic cells, the adult Leydig cells (ALCs), which differentiate at puberty (Shima and Morohashi, 2017; Shima, 2019).
FLCs differentiate from embryonic day 12.5 (E12.5) in mice and increase in number during fetal life through the recruitment and differentiation of Wnt5a positive proliferative progenitors located in the interstitial space of the testes (Ademi et al., 2022; Shima and Morohashi, 2017; Shima, 2019; Rotgers et al., 2018). Interstitial progenitors also give rise to the contractile peritubular myoid cells (PTM) that will surround the future seminiferous tubules (Ademi et al., 2022). Despite being the most abundant cell population in the fetal testis (Ademi et al., 2022; Mayère et al., 2022), little is known about the genetic control of the proliferation, specification, and differentiation of the interstitial steroidogenic progenitor cells.
Lineage tracing and single-cell transcriptomic analyses have revealed that interstitial progenitors have a dual origin. The majority are derived from the coelomic epithelium of the undifferentiated gonad, which harbors early bipotential progenitors able to differentiate along the supporting (the future Sertoli cells) or the steroidogenic lineage (Ademi et al., 2022; Karl and Capel, 1998; Stévant et al., 2018; Stévant et al., 2019). In addition, Nestin positive cells migrate from the adjacent mesonephros into the gonad and differentiate along the steroidogenic lineage from E13.5 to give rise to up to a third of the FLC population by the end of gestation (Ademi et al., 2022; DeFalco et al., 2011; Kumar and DeFalco, 2018).
Positive and negative paracrine signals drive FLC differentiation by upregulating the transcription factors NR5A1, GATA4, and GATA6, which in turn regulate the expression of genes related to cholesterol metabolism and steroidogenesis (Rotgers et al., 2018; Wen et al., 2016; Morohashi et al., 2020; Baba et al., 2018; Baba et al., 2014; Shima et al., 2018; Padua et al., 2015; Bielinska et al., 2007; Viger et al., 2022). Desert Hedgehog (DHH) is secreted by Sertoli cells and acts on the interstitial progenitors expressing the Hedgehog receptor Patched1 (PTCH1) and the Hedgehog effectors GLI1, -2, and -3 to trigger FLC and PTM differentiation (Yao et al., 2002; Barsoum and Yao, 2011; Barsoum et al., 2009; Park et al., 2000; Kothandapani et al., 2020; Pierucci-Alves et al., 2001; Clark et al., 2000). In addition, FLC development requires the activation of signaling pathways downstream of PDGFRA in the steroidogenic progenitors (Brennan et al., 2003; Inoue et al., 2022; Schmahl et al., 2008). On the other hand, ligands present in vascular and perivascular cells activate the NOTCH2 receptor and the expression of the effectors HES1 and HEYL in interstitial progenitors to maintain their undifferentiated state and inhibit FLC differentiation (Kumar and DeFalco, 2018; Tang et al., 2008; Liu et al., 2016; Defalco et al., 2013). In addition to the paracrine signals from adjacent cell populations, FLC differentiation is also regulated by the cell-autonomous action of the transcription factors ARX, TCF21, PBX1, MAF, and MAFB expressed in the interstitial steroidogenic progenitors, although their precise roles remain elusive (Ademi et al., 2022; Miyabayashi et al., 2013; Li et al., 2021; Cui et al., 2004; Schnabel et al., 2003).
Nuclear receptor subfamily 2 group F member 2 (NR2F2, also known as COUP-TFII) is abundantly expressed in interstitial progenitors of the fetal and adult testis in rodents and humans (Inoue et al., 2016; Ademi et al., 2022; Mendoza-Villarroel et al., 2014b; Kilcoyne et al., 2014; van den Driesche et al., 2012; Lottrup et al., 2014; Qin et al., 2008; Taelman et al., 2024). NR2F2 activates or represses transcription depending on the cellular context by directly binding to DNA responsive elements or by interacting with other transcription factors. NR2F2 regulates cell differentiation during organogenesis, adult tissue homeostasis, and tumorigenesis (Polvani et al., 2019). NR2F2 function is essential for cardiac and vascular development so that Nr2f2 mutation in mouse leads to embryonic lethality at mid-gestation (Pereira et al., 1999). The study of mouse conditional mutants has shown that NR2F2 is essential for ALC differentiation in the postnatal testis before puberty (Qin et al., 2008). However, the function of NR2F2 in the developing testis during fetal life has not been addressed, and its role in the interstitial progenitors that give rise to the FLC lineage remains unknown. It was initially proposed that NR2F2 could act as a negative regulator of steroidogenesis at fetal stages based on the inverse correlation between NR2F2 expression and steroidogenesis genes and testosterone levels in mouse and rat fetal testes treated with endocrine disruptors (van den Driesche et al., 2012). More recently, rare variants in NR2F2 have been associated with cryptorchidism, hypospadias, and defective penile growth in human patients (Zidoune et al., 2022; Ganapathi et al., 2023; Wankanit et al., 2024). These phenotypes can be attributed to defective testosterone and INSL3 production during gestation (Hutson et al., 2015; Amato et al., 2022), suggesting a positive role for NR2F2 in promoting FLC differentiation and/or function in the fetal testis.
In this study, we show that NR2F2 is expressed in interstitial progenitors of coelomic and mesonephric origin of the mouse fetal testes and is downregulated upon FLC differentiation. By using two Cre lines that drive Nr2f2 deletion in mouse embryonic gonads, we show that NR2F2 is required for fetal mouse testis morphogenesis and for FLC differentiation. Absence of Nr2f2 does not impair paracrine signals known to regulate FLC differentiation nor the proliferation or survival of the steroidogenic progenitor population. Our findings reveal that NR2F2 promotes the initiation of FLC differentiation as well as FLC maturation. Taken together, these results establish NR2F2 as an essential factor that positively regulates the development of steroidogenic cells in the mouse fetal testis.
Results
NR2F2 is expressed in steroidogenic progenitors of the developing testis
We analyzed the spatiotemporal distribution of NR2F2 in the developing testis by immunofluorescence. At E11.5 (18–21 tail somites [ts]), NR2F2 was detected in both the coelomic epithelium and the mesonephric mesenchyme adjacent to the gonads, two tissues that contribute to the population of interstitial steroidogenic progenitors (Ademi et al., 2022; Karl and Capel, 1998; Kumar and DeFalco, 2018; Figure 1A and C). The majority of mesenchymal cells in the gonad expressed the transcription factor RUNX1 (RUNX1+) (Nicol et al., 2019), indicating that most of the gonadal somatic cells at this stage belong to the supporting lineage (Figure 1A and D). Nevertheless, NR2F2+ RUNX1- cells were observed, revealing that interstitial progenitors were already present at this stage (Figure 1A). NR2F2+ cells were either GATA4+ or GATA4-, suggesting that interstitial progenitors of coelomic and mesonephric origins, respectively, were both present (Figure 1A–C).
Figure 1 with 1 supplement see all
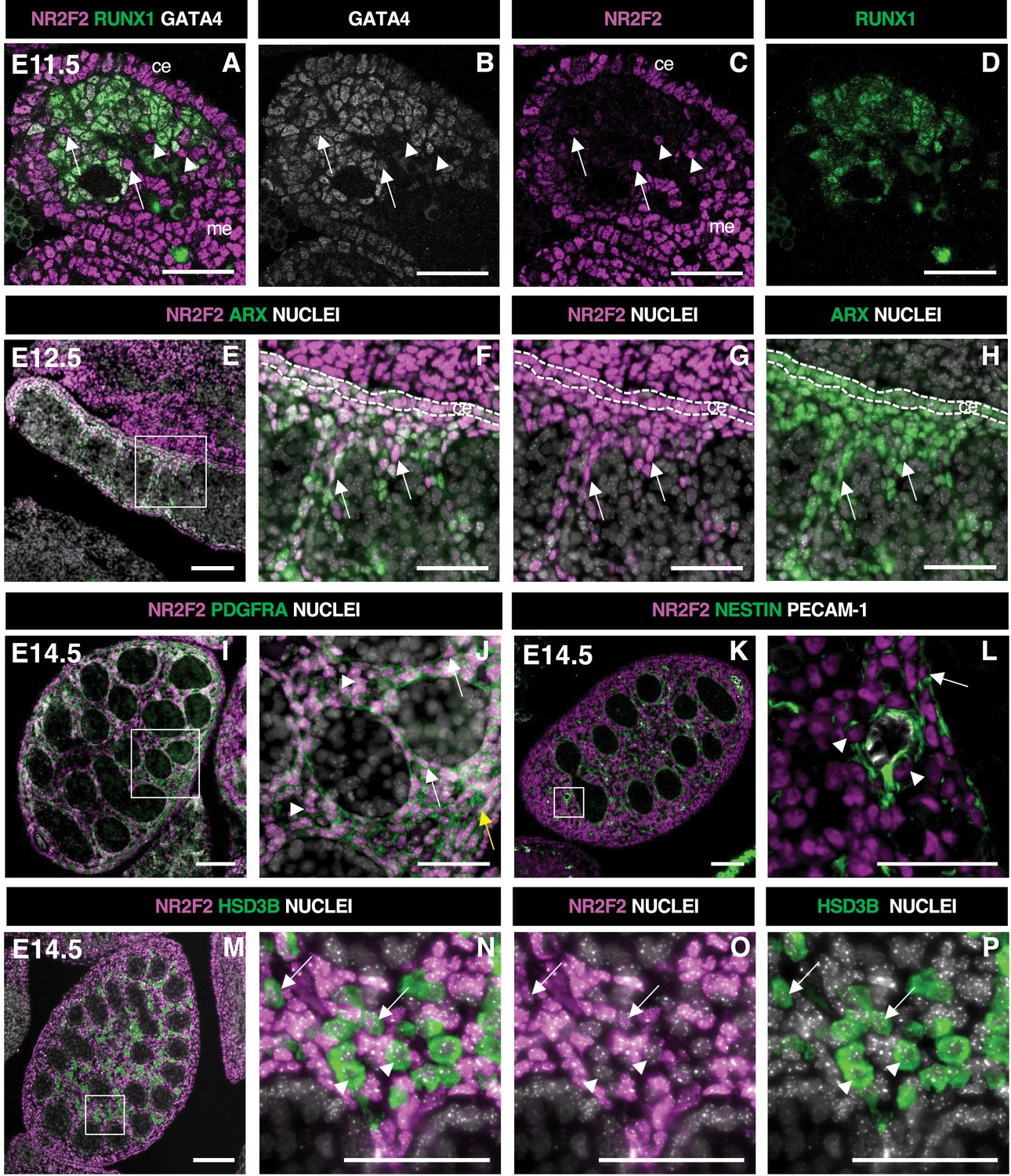
NR2F2 is expressed in steroidogenic progenitors of the fetal testis.
(A–D) Immunodetection of NR2F2, RUNX1, and GATA4 on embryonic day 11.5 (E11.5) (18 tail somites) XY gonad. NR2F2 is detected in coelomic epithelium (ce), mesonephros (me), and RUNX1 negative cells that are either GATA4 positive (arrows in A–C) or GATA4 negative (arrowheads in A–C). (E–H) Immunodetection of NR2F2 and ARX on E12.5 XY gonad. NR2F2 is co-expressed with ARX in the coelomic epithelium (ce, dotted lines in F–H) and in interstitial cells between the testis cords (arrows in F–H). (I,J) Immunodetection of NR2F2 and PDGFRA on E14.5 XY gonad. NR2F2 is detected in PDGFRA positive cells, including interstitial progenitors (arrowheads in J), peritubular myoid cell surrounding testis cords (arrows in J), and cells of the future tunica albuginea (yellow arrow in J). (K,L) Immunodetection of NR2F2, NESTIN, and PECAM-1 on E14.5 XY gonad. NR2F2 is detected in NESTIN positive interstitial progenitors, including perivascular cells (arrowheads in L) and peritubular myoid cells (arrow in L). (M–P) Immunodetection of NR2F2 and HSD3B on E14.5 XY gonad. NR2F2 is absent from the majority of HSD3B positive fetal Leydig cells (arrowheads in N–P) and is only detected at low levels in a few HSD3B positive elongated cells (arrows in N–P). Data are representative of triplicate biological replicates. Scale bar = 50 µm in A–D, F–H, J, L, and N–P. Scale bar = 100 µm in E, I, K, M.
In E12.5 testes, NR2F2+ cells were detected in the coelomic epithelium and in the interstitial space outside the developing testis cords (Figure 1E–H, Figure 1—figure supplement 1A–D). NR2F2+ cells co-expressed the steroidogenic progenitor marker ARX (Ademi et al., 2022; Miyabayashi et al., 2013) and were actively proliferating (Figure 1E–H, Figure 1—figure supplement 1E–H). At E14.5, NR2F2 expression was maintained in interstitial cells co-expressing PDGFRA (Brennan et al., 2003), as well as in the PTM lining the testis cords and in the cells beneath the surface of the testis that will contribute to the future tunica albuginea (Figure 1I and J, Figure 1—figure supplement 1M–P). NR2F2 expression was also found in NESTIN-expressing perivascular cells that correspond to the mesonephros-derived steroidogenic progenitors (Kumar and DeFalco, 2018; Figure 1K and L). In contrast, NR2F2 protein was either absent or detected at very low levels in FLC marked by the expression of the steroidogenic enzyme HSD3B (Figure 1M–P, Figure 1—figure supplement 1I–L).
Together, our results show that NR2F2 is expressed in the coelomic epithelium and the mesonephros, as well as in the interstitial progenitors derived from both sources, and is downregulated upon FLC differentiation in the developing testis.
NR2F2 is required for fetal testicular morphogenesis and FLC development
In order to investigate the function of NR2F2 in the developing mouse testis, we used an Nr2f2flox conditional allele (Hutson et al., 2015), and a knock-in Wt1CreERT2 line, in which tamoxifen-inducible CreERT2 is produced by Wt1-expressing cells (Shima and Morohashi, 2017; Figure 2A). Wt1 is expressed from E9.5 in the coelomic epithelium of the gonadal ridge and in the adjacent mesonephros (Armstrong et al., 1993), and Wt1CreERT2-mediated recombination can be induced in all somatic gonadal cells upon tamoxifen administration at E9.5 and E10.5 (Manuylov et al., 2008; Figure 2—figure supplement 1A).
Figure 2 with 1 supplement see all
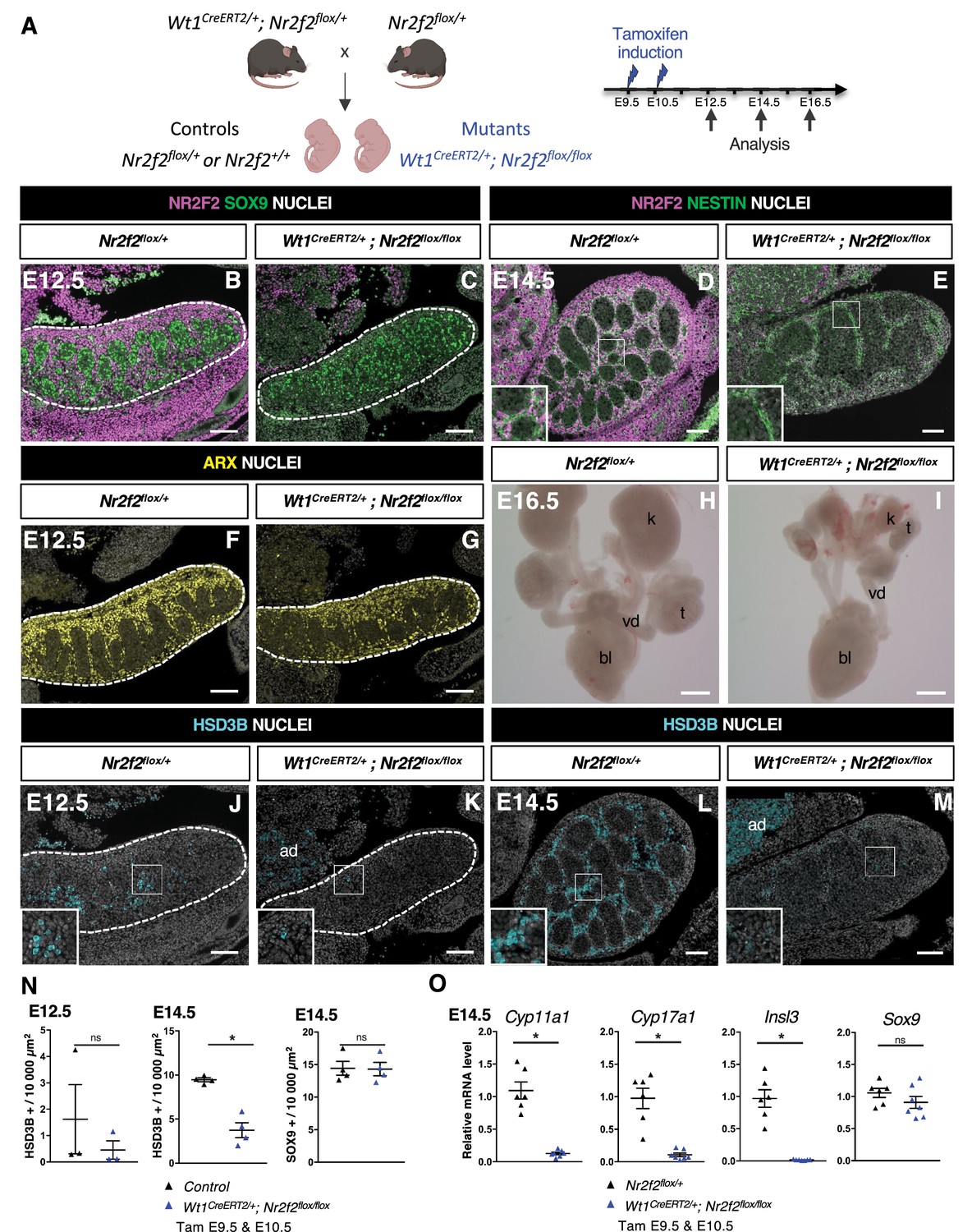
NR2F2 deletion by Wt1CreERT2 impairs Sertoli cell differentiation and fetal Leydig cell (FLC) development.
(A) Generation of Wt1CreERT2; Nr2f2flox/flox mutants. Tamoxifen was administered at embryonic day 9.5 (E9.5) and E10.5, and embryos were recovered at E12.5, E14.5, and E16.5. (B,C) Immunodetection of NR2F2 and SOX9 on E12.5 Nr2f2flox/+ and Wt1CreERT2; Nr2f2flox/flox testes (outlined by dotted lines) after tamoxifen treatment at E9.5 and E10.5. NR2F2 is efficiently deleted in the gonad and mesonephros. (D,E) Immunodetection of NR2F2 and NESTIN on E14.5 Nr2f2flox/+ and Wt1CreERT2; Nr2f2flox/flox testes. NR2F2 is efficiently deleted in NESTIN1 positive cells. (F,G) Immunodetection of ARX on E12.5 Nr2f2flox/+ and Wt1CreERT2; Nr2f2flox/flox testes (outlined by dotted lines). Interstitial cells are generated in Wt1CreERT2; Nr2f2flox/flox mutants. (H,I) Macroscopic view of the urogenital tract of XY E16.5 Nr2f2flox/+ and Wt1CreERT2; Nr2f2flox/flox testes dissected after tamoxifen treatment at E9.5 and E10.5. Testes (t) and kidneys (k) are hypoplastic in Wt1CreERT2; Nr2f2flox/flox mutants. bl: bladder. vd: vas deferens. (J,K) Immunodetection of HSD3B on E12.5 Nr2f2flox/+ and Wt1CreERT2; Nr2f2flox/flox testes (outlined by dotted lines). ad: adrenal. (L,M) Immunodetection of HSD3B on E14.5 Nr2f2flox/+ and Wt1CreERT2; Nr2f2flox/flox testes. ad: adrenal. (N) Quantification of the number of HSD3B positive cells per surface unit and of the number of SOX9 positive cells per surface unit in control (wild-type or Nr2f2flox/+) and Wt1CreERT2; Nr2f2flox/flox testes. Each triangle represents the mean number of HSD3B or SOX9 positive cells per surface unit of one individual measured on at least two sections per gonad. Data are shown as means ± SEM. Statistical significance was assessed by Mann-Whitney U two-tailed test. * indicates p-value≤0.05; ns indicates p-value>0.05. (O) Quantification of Cyp11a1, Cyp17a1, Insl3, and Sox9 transcripts in Nr2f2flox/+ and Wt1CreERT2; Nr2f2flox/flox testes treated with tamoxifen at E9.5 and E10.5 and dissected at E14.5 after normalization to Sdha and Tbp by RT-qPCR. Data are shown as means ± SEM. Statistical significance was assessed by Mann-Whitney U two-tailed test. * indicates p-value≤0.05; ns indicates p-value>0.05. Immunodetection data are representative of triplicate biological replicates. Scale bar = 100 µm in B–G, J–M. Scale bar = 500 µm in H, I.
-
Figure 2—source data 1
Source data for cell counts and RT-qPCR data in Figure 2.
- https://cdn.elifesciences.org/articles/103783/elife-103783-fig2-data1-v1.xlsx
NR2F2 is co-expressed with WT1 in the gonadal coelomic epithelium, in the mesonephros, and in interstitial cells (Yu et al., 2012; Figure 2—figure supplement 1C–F). Tamoxifen treatment at E9.5 and E10.5 triggered an efficient NR2F2 deletion in gonadal and mesonephric tissues of Wt1CreERT2; Nr2f2flox/flox embryos analyzed at E12.5 and E14.5 (Figure 2A–E, Figure 2—figure supplement 1B). NR2F2 expression was completely absent in gonadal interstitial cells, including NESTIN+ cells, demonstrating that all interstitial steroidogenic progenitors were targeted in Wt1CreERT2; Nr2f2flox/flox embryos (Figure 2D and E). Morphological examination of the urogenital system of E16.5 Wt1CreERT2; Nr2f2flox/flox embryos revealed hypoplastic undescended testes, as well as hypoplastic kidneys (Figure 2H and I), indicating that NR2F2 function is required for testicular and kidney development.
Sertoli cells expressing SOX9 (Figure 2B and C) and interstitial progenitors marked by ARX (Figure 2F and G) were present in Wt1CreERT2; Nr2f2flox/flox embryos at E12.5. However, differentiated steroidogenic FLC (marked by HSD3B expression) were almost completely absent in the mutant gonads (Figure 2J, K, and N). This phenotype was not due to a delay in the initiation of FLC differentiation, as the number of HSD3B positive cells remained strongly reduced in Wt1CreERT2; Nr2f2flox/flox mutant gonads at E14.5 (60% reduction, Figure 2L–N). In addition, the transcripts of Cyp11a1 and Cyp17a1, two genes encoding steroidogenic enzymes expressed in FLC, and of Insl3 were strongly reduced (Figure 2O), providing a possible explanation for the observed undescended testes phenotype (Figure 2—figure supplement 1G and H). The mutant gonads showed fewer testis cords that were enlarged and exhibited irregular shapes at E14.5 (Figure 2E, Figure 2—figure supplement 1J and M). Moreover, the expression of ACTA2 was strongly reduced both in the periphery of the gonad and in cells lining the testis cords, indicating that tunica cell and PTM development were impaired in Wt1CreERT2; Nr2f2flox/flox mutants (Figure 2—figure supplement 1G and H). Together, these results demonstrate that NR2F2 function is not essential for the initial specification of the interstitial and supporting cells of the testis but is required for testicular morphogenesis and for FLC development.
Sertoli cell development is impaired in Wt1CreERT2; Nr2f2flox/flox testes
FLC differentiation relies on signals produced by Sertoli cells such as DHH and PDGFA acting on interstitial cells (Rotgers et al., 2018; Wen et al., 2016). The number of Sertoli cells expressing SOX9 and the levels of Sox9 transcripts were not altered in Wt1CreERT2; Nr2f2flox/flox mutants at E14.5 (Figure 2N and O). However, the expression of Dhh, Pdgfa, and Amh, another marker of differentiated Sertoli cells, was reduced in Wt1CreERT2; Nr2f2flox/flox mutants compared to Nr2f2flox/+ control gonads (Figure 2—figure supplement 1K), indicating that Sertoli cell differentiation is abnormal in Wt1CreERT2; Nr2f2flox/flox gonads. WT1 is required for Sertoli cell development (Gao et al., 2006), and previous work suggested that Wt1 heterozygosity in Wt1CreERT2/+ gonads results in delayed testicular development (Carré et al., 2018). To discriminate between the effects of Nr2f2 loss of function and Wt1 heterozygosity in the phenotype of Wt1CreERT2; Nr2f2flox/flox mutants, we analyzed gene expression in Wt1CreERT2/+ testes compared to wild-type littermates. We found that the expression of the FLC markers Cyp11a1 and Cyp17a1 was not significantly different in Wt1CreERT2/+ testes compared to wild-type littermates (Figure 2—figure supplement 1L). In contrast, transcript levels of Dhh, Pdgfa, and Amh were significantly reduced (Figure 2—figure supplement 1L). While these results indicate that impaired FLC development in Wt1CreERT2; Nr2f2flox/flox mutants is associated with the loss of Nr2f2 function, the potential contribution of Sertoli cell defects caused by Wt1 heterozygosity to the FLC phenotype cannot be ruled out in this model.
NR2F2 is required in the steroidogenic lineage for FLC development
In order to elucidate the specific function of NR2F2 in the steroidogenic lineage for FLC development, we used the transgenic Nr5a1-Cre line which drives robust recombination in somatic gonadal cells from E11.5, after the supporting and steroidogenic lineages have been specified (Figure 3A, Figure 3—figure supplement 1A and B; Bingham et al., 2006; Manuylov et al., 2011).
Figure 3 with 1 supplement see all
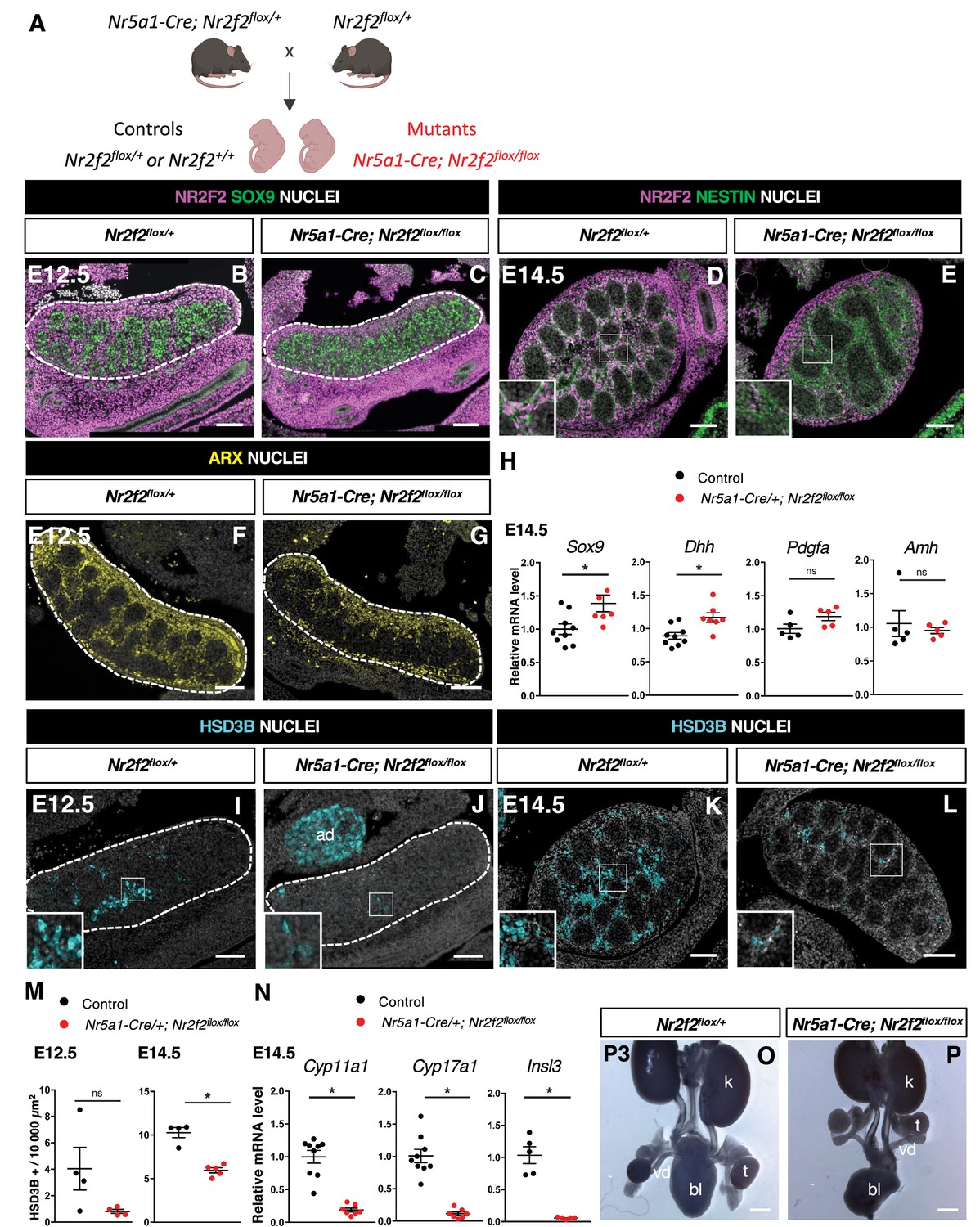
NR2F2 deletion by Nr5a1-Cre impairs fetal Leydig cell (FLC) development.
(A) Generation of Nr5a1-Cre; Nr2f2flox/flox mutants. (B,C) Immunodetection of NR2F2 and SOX9 on embryonic day 12.5 (E12.5) Nr2f2flox/+ and Nr5a1-Cre; Nr2f2flox/flox testes (outlined by dotted lines). NR2F2 is deleted in interstitial cells but is still present in the outermost layer of the testis. (D,E) Immunodetection of NR2F2 and NESTIN on E14.5 Nr2f2flox/+ and Nr5a1-Cre; Nr2f2flox/flox testes. NR2F2 is still detected in NESTIN1 positive cells. (F,G) Immunodetection of ARX on E12.5 Nr2f2flox/+ and Nr5a1-Cre; Nr2f2flox/flox testes (outlined by dotted lines). Interstitial cells are generated in Nr5a1-Cre; Nr2f2flox/flox mutants. (H) Quantification of Sox9, Dhh, Pdgfa, and Amh transcripts after normalization to Sdha and Tbp in control (wild-type or Nr2f2flox/+) and Nr5a1-Cre; Nr2f2flox/flox by RT-qPCR at E14.5. Data are shown as means ± SEM. Statistical significance was assessed by Mann-Whitney U two-tailed test. * indicates p-value≤0.05; ns indicates p-value>0.05. (I,J) Immunodetection of HSD3B on E12.5 Nr2f2flox/+ and Nr5a1-Cre; Nr2f2flox/flox testes (outlined by dotted lines). ad: adrenal. (K,L) Immunodetection of HSD3B on E14.5 Nr2f2flox/+ and Nr5a1-Cre; Nr2f2flox/flox testes. (M) Quantification of the number of HSD3B positive cells per surface unit in control (wild-type or Nr2f2flox/+) and Nr5a1-Cre; Nr2f2flox/flox testes at E12.5 and E14.5. Each circle represents the mean number of HSD3B positive cells per surface unit of one individual measured on at least two sections per gonad. Data are shown as means ± SEM. Statistical significance was assessed by Mann-Whitney U two-tailed test. * indicates p-value≤0.05; ns indicates p-value>0.05. (N) Quantification of Cyp11a1, Cyp17a1, and Insl3 after normalization to Sdha and Tbp in control (wild-type or Nr2f2flox/+) and Nr5a1-Cre; Nr2f2flox/flox by RT-qPCR at E14.5. Data are shown as means ± SEM. Statistical significance was assessed by Mann-Whitney U two-tailed test. * indicates p-value≤0.05; ns indicates p-value>0.05. (O,P) Macroscopic view of the urogenital tract of XY P3 Nr2f2flox/+ and Nr5a1-Cre; Nr2f2flox/flox mutants. The testes are in abdominal position in Nr5a1-Cre; Nr2f2flox/flox mutants. k: kidney, t: testis, bl: bladder. vd: vas deferens. Immunodetection data are representative of triplicate biological replicates. Scale bar = 100 µm in B–G, I–L. Scale bar = 500 µm in O, P.
-
Figure 3—source data 1
Source data for cell counts and RT-qPCR data in Figure 3.
- https://cdn.elifesciences.org/articles/103783/elife-103783-fig3-data1-v1.xlsx
At E11.5, NR5A1 is co-expressed with NR2F2 in the gonadal coelomic epithelium and in interstitial cells, but is absent from the mesonephros and mesonephros-derived cells (Stévant et al., 2018; Kumar and DeFalco, 2018; Figure 3—figure supplement 1D–G). Nr5a1-Cre; Nr2f2flox/flox mutants show efficient Nr2f2 deletion in gonadal interstitial cells at E12.5 and E14.5 (Figure 3B–E, Figure 3—figure supplement 1C). Consistent with previous reports on the activity of Nr5a1-Cre (Manuylov et al., 2011), NR2F2 was still detected in the coelomic epithelium layer and in interstitial cells just beneath it, particularly in the anterior part of the gonad of Nr5a1-Cre; Nr2f2flox/flox mutants (Figure 3B–E). NESTIN+ NR2F2+ cells were still present, confirming that steroidogenic progenitors of mesonephric origin were not targeted by Nr5a1-Cre (Figure 3D and E, Figure 3—figure supplement 1H and I; Kumar and DeFalco, 2018). Together, these results demonstrate that in Nr5a1-Cre; Nr2f2flox/flox mutants, NR2F2 is deleted after E11.5 in interstitial cells derived from the coelomic epithelium, except in the outermost layer of the testis.
Nr2f2 deletion by Nr5a1-Cre did not affect the initial formation of the supporting and steroidogenic lineages as evidenced by SOX9 (Figure 3B, C, and H, Figure 3—figure supplement 1N and O) and ARX expression (Figure 3F and G), nor the differentiation of Sertoli cells as shown by normal expression levels of Dhh, Pdgfa, and Amh (Figure 3H, Figure 3—figure supplement 1N). In addition, ACTA2 was detected in PTM and was only slightly reduced in the tunica albuginea of the posterior region in the mutant testes (Figure 3—figure supplement 1J and K). In contrast, the FLC population marked by HSD3B or CYP11A1 was decreased (40% reduction, Figure 3I–M, Figure 3—figure supplement 1L and M), and the expression of the FLC markers Cyp11a1, Cyp17a1, and Insl3 was strongly downregulated in Nr5a1-Cre; Nr2f2flox/flox mutant testes (Figure 3N). We conclude that Nr2f2 deletion after E11.5 by Nr5a1-Cre leads to FLC reduction without Sertoli cell defects, suggesting that NR2F2 is required cell-autonomously in the interstitial cells for FLC development.
In agreement with reduced Insl3 expression, the testes of Nr5a1-Cre; Nr2f2flox/flox embryos were undescended and exhibited an abnormal abdominal position at postnatal day (P) 3 (Figure 3O and P). In addition, the anogenital distance, a readout for androgen levels (Schwartz et al., 2019), was reduced in P0 and P1 Nr5a1-Cre; Nr2f2flox/flox males when compared to male controls (Figure 3—figure supplement 1P), consistent with the reduced expression of steroidogenic enzymes in the mutant testes. We conclude that defective FLC development in Nr2f2 mutants results in cryptorchidism and impaired masculinization of the external genitalia.
NR2F2 is required for the initiation of FLC differentiation
FLCs differentiate from proliferating interstitial progenitors that progressively lose their mitotic ability, downregulate the transcription factors ARX and NR2F2, upregulate the master regulator of steroidogenesis NR5A1, and activate the expression of steroidogenesis-related genes (Inoue et al., 2016; Ademi et al., 2022; Miyabayashi et al., 2013). We wanted to determine which of these steps of FLC differentiation are NR2F2 dependent.
The transcription factor ARX is required in the pool of proliferating interstitial progenitors for FLC development (Miyabayashi et al., 2013). We first examined whether NR2F2 regulates the survival, proliferation, or identity of the ARX+ cell population. We found that the expression of Arx mRNA was not modified in Nr5a1-Cre; Nr2f2flox/flox mutants (Figure 4—figure supplement 1A). The total number of gonadal cells, the percentage of ARX+ cells among the total number of gonadal cells, and the fraction of proliferating cells among the ARX+ population were similar to controls at E12.5 and E14.5 (Figure 4A, B, and E, Figure 4—figure supplement 1B–E). In addition, we did not find evidence of increased cell death in Nr5a1-Cre; Nr2f2flox/flox mutants (Figure 4—figure supplement 1F–H). Together, these results indicate that ARX+ steroidogenic progenitor cells are present and proliferate at normal rates in Nr5a1-Cre; Nr2f2flox/flox mutants.
Figure 4 with 1 supplement see all
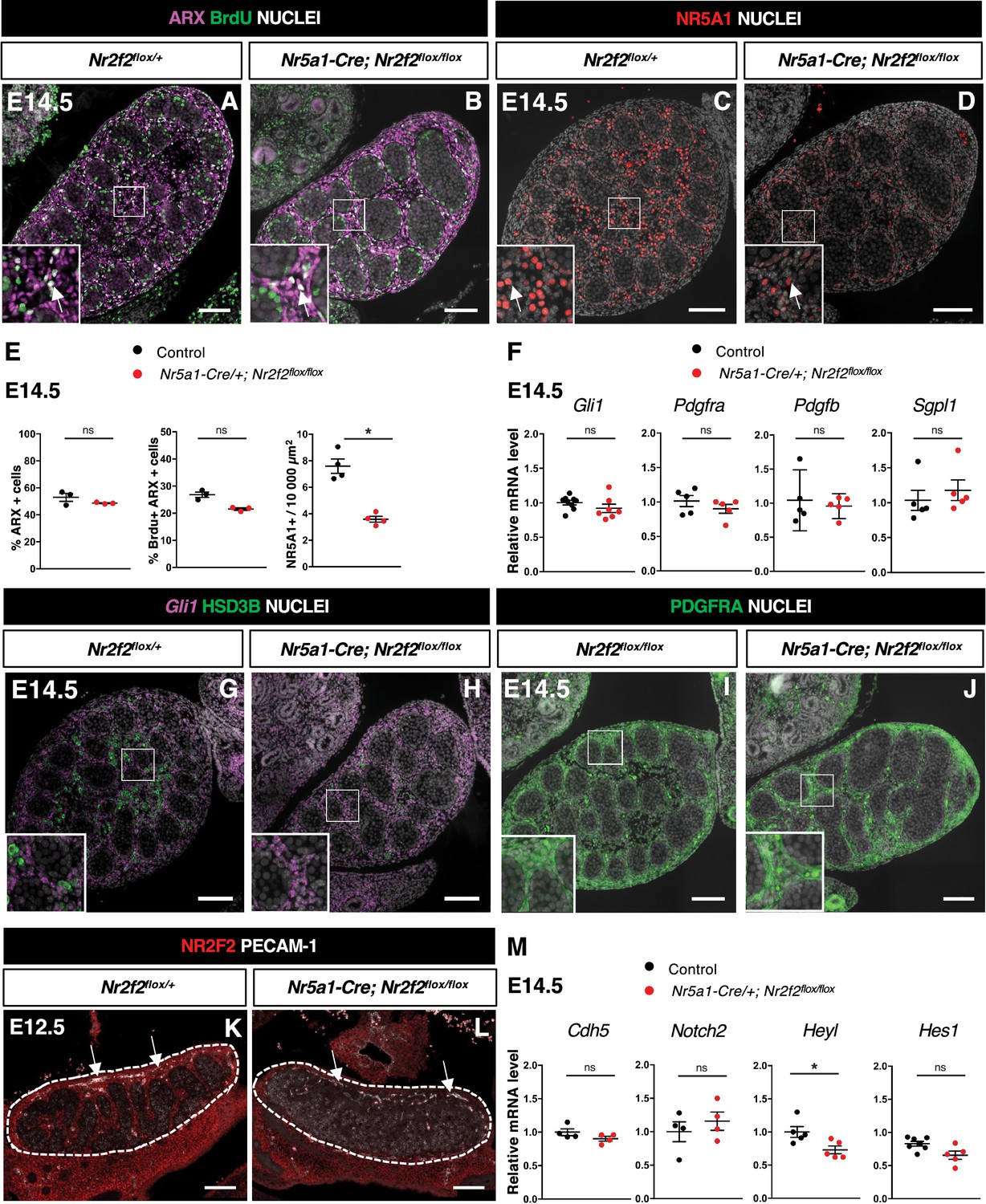
NR2F2 function is required for the initiation of fetal Leydig cell (FLC) differentiation.
(A,B) Immunodetection of ARX and BrdU (arrows) on embryonic day 14.5 (E14.5) XY Nr2f2flox/+ and Nr5a1-Cre; Nr2f2flox/flox testes. (C,D) Immunodetection of NR5A1 on E14.5 XY Nr2f2flox/+ and Nr5a1-Cre; Nr2f2flox/flox testes. Arrows indicate strong expression of NR5A1 positive FLC. (E) Quantification of the percentage of ARX positive cells (number of ARX positive nuclei relative to the total number of nuclei labeled by DAPI), of the percentage of ARX positive cells labeled by BrdU (number of nuclei positive for ARX and BrdU relative to the number of ARX positive nuclei) and of the number of NR5A1 positive cells per surface unit in E14.5 XY control (wild-type or Nr2f2flox/+) and Nr5a1-Cre; Nr2f2flox/flox testes. Each circle represents the mean percentage of ARX+ or ARX+/BrdU+ or NR5A1+ cells per surface unit of one individual measured on at least two sections per gonad. Data are shown as means ± SEM. Statistical significance was assessed by Mann-Whitney U two-tailed test. * indicates p-value≤0.05; ns indicates p-value>0.05. (F) Quantification of Gli1, Pdgfra, Pdgfb, and Sgpl1 transcripts after normalization to Sdha and Tbp in control (wild-type or Nr2f2flox/+) and Nr5a1-Cre; Nr2f2flox/flox by RT-qPCR at E14.5. Statistical significance was assessed by Mann-Whitney U two-tailed test. * indicates p-value≤0.05; ns indicates p-value>0.05. (G,H) In situ hybridization detection of Gli1 transcripts and immunodetection of HSD3B protein on E14.5 XY Nr2f2flox/+ and Nr5a1-Cre; Nr2f2flox/flox testes. (I,J) Immunodetection of PDGFRA on E14.5 XY Nr2f2flox/flox and Nr5a1-Cre; Nr2f2flox/flox testes. (K,L) Immunodetection of PECAM-1 and NR2F2 on E12.5 XY Nr2f2flox/+ and Nr5a1-Cre; Nr2f2flox/flox testes. PECAM-1 is expressed in germ cells and in endothelial cells (white arrows). (M) Quantification of Cdh5, Notch2, Heyl, and Hes1 transcripts after normalization to Sdha and Tbp in control (wild-type or Nr2f2flox/+) and Nr5a1-Cre; Nr2f2flox/flox by RT-qPCR at E14.5. Statistical significance was assessed by Mann-Whitney U two-tailed test. * indicates p-value≤0.05; ns indicates p-value>0.05. Immunodetection data are representative of triplicate biological replicates. Scale bar = 100 µm.
-
Figure 4—source data 1
Source data for cell counts and RT-qPCR data in Figure 4.
- https://cdn.elifesciences.org/articles/103783/elife-103783-fig4-data1-v1.xlsx
As the ARX+ steroidogenic progenitors adopt an FLC identity, NR2F2 is progressively lost, and the nuclear receptor NR5A1 is strongly upregulated (Inoue et al., 2016; Ademi et al., 2022; van den Driesche et al., 2012). NR5A1 directs FLC differentiation by regulating the expression of genes associated with cholesterol metabolism and steroidogenesis (Morohashi et al., 2020; Shima et al., 2018; Jeyasuria et al., 2004; Buaas et al., 2012). In Nr5a1-Cre; Nr2f2flox/flox mutants, the cells expressing high levels of NR5A1+ in the interstitial compartment were reduced compared to controls (Figure 4C–E, Figure 4—figure supplement 1I and J). This observation indicates that NR2F2 function is required for NR5A1 upregulation at the initial step of steroidogenic cell differentiation.
DHH, PDGFRA, and NOTCH pathways are not impaired in Nr5a1-Cre; Nr2f2flox/flox mutants
We next investigated the impact of Nr5a1-Cre; Nr2f2 flox/flox mutation on the activation of the signaling pathways involved in FLC differentiation. DHH secreted by Sertoli cells binds to its receptor PTCH1 expressed in interstitial progenitors and activates its target genes, including Gli1, to promote FLC differentiation (Yao et al., 2002; Barsoum and Yao, 2011). The expression levels of Dhh and of its target Gli1, a readout of Hedgehog pathway activation, were not modified in Nr5a1-Cre; Nr2f2flox/flox mutants (Figure 3H, Figure 4F, Figure 3—figure supplement 1N). Gli1 transcripts exhibited a similar distribution in interstitial cells of control and Nr5a1-Cre; Nr2f2flox/flox mutants at E14.5 (Figure 4G and H). PDGFA produced by Sertoli cells binds to the PDGFRA receptor expressed in interstitial progenitors to activate downstream signaling required for FLC development (Brennan et al., 2003; Schmahl et al., 2008). The expression levels of Pdgfa and Pdgfb, coding for two PDGFRA ligands expressed in developing testes (Brennan et al., 2003), were not altered in Nr5a1-Cre; Nr2f2flox/flox mutants (Figures 3H and 4F). Transcript levels of Pdgfra were unchanged in Nr5a1-Cre; Nr2f2flox/flox mutants (Figure 4F), and PDGFRA protein was detected at the plasma membrane of interstitial cells in both control and Nr5a1-Cre; Nr2f2flox/flox mutant testes at E14.5 (Figure 4I and J). In addition, the expression of Sgpl1, a PDGFRA signaling target involved in steroidogenic differentiation (Schmahl et al., 2008), was not reduced in Nr5a1-Cre; Nr2f2flox/flox mutants (Figure 4F). These observations indicate that the activities of Hedgehog and PDGFRA signaling, two pathways that positively regulate FLC differentiation, are not impaired in Nr5a1-Cre; Nr2f2flox/flox mutants.
In addition to the positive signals, FLC differentiation is also negatively modulated by NOTCH signaling triggered by ligands expressed in vascular and perivascular cells. An increase in testicular endothelial cells results in a reduction in FLC numbers (Kumar and DeFalco, 2018; Tang et al., 2008; Liu et al., 2016; Defalco et al., 2013). The distribution and abundance of endothelial cells marked by PECAM-1 was not altered in Nr5a1-Cre; Nr2f2flox/flox mutants (Figure 4K and L). In addition, Cdh5 transcript levels (a readout of the abundance of endothelial cells) were similar in controls and Nr5a1-Cre; Nr2f2flox/flox mutants (Figure 4M). These observations indicate that the reduction in FLCs in Nr5a1-Cre; Nr2f2flox/flox mutants was not associated with an increased population of endothelial cells. The receptor NOTCH2 expressed in interstitial cells is involved in restricting FLC differentiation (Kumar and DeFalco, 2018; Tang et al., 2008; Liu et al., 2016; Defalco et al., 2013). Notch2 mRNA levels were not altered in Nr5a1-Cre; Nr2f2flox/flox mutants (Figure 4M). NOTCH signaling activates the expression of its target genes (that also act as effectors of the pathway), including Heyl, specifically expressed in interstitial cells and strongly upregulated upon vascular depletion (Kumar and DeFalco, 2018) and Hes1, expressed in interstitial cells and involved in restricting FLC differentiation (Tang et al., 2008; Liu et al., 2016). Transcripts of the NOTCH pathway targets Hes1 and Heyl were detected at similar levels in controls and Nr5a1-Cre; Nr2f2flox/flox mutants (Figure 4M), indicating that the observed decrease in FLC numbers cannot be attributed to an increase in NOTCH signaling.
Together, these results indicate that NR2F2 deficiency does not impact the activity of the signaling pathways known to induce or repress FLC formation, suggesting that NR2F2 acts downstream or in parallel to these pathways to regulate FLC differentiation from interstitial progenitors.
NR2F2 is required for FLC maturation
A small fraction of HSD3B positive cells formed in the absence of NR2F2 function, even in the case of Wt1CreERT2; Nr2f2flox/flox mutant gonads that exhibit widespread NR2F2 deletion in steroidogenic progenitors of coelomic and mesonephric origin. One possibility is that these cells are ALCs that have prematurely differentiated in Nr2f2 mutant fetal testes. Bhmt (Betaine-homocysteine methyltransferase) is specifically expressed in ALC and absent in FLC (Sararols et al., 2021). Bhmt transcripts were not detected in control or Nr2f2 mutant testes by RT-qPCR at E14.5, while they were present in adult testes (Figure 5—figure supplement 1A), indicating that the steroidogenic cells of the mutants do not have adult characteristics.
To evaluate the steroidogenic capacities of the remaining FLC in Nr2f2 mutant gonads, we analyzed the transcript levels of Cyp11a1 and Cyp17a1 normalized to the FLC number as quantified by HSD3B immunofluorescence (Figure 2N, Figure 3M; Kothandapani et al., 2020). Normalized data showed reduced steroidogenic gene expression in Nr5a1-Cre; Nr2f2flox/flox and in Wt1CreERT2; Nr2f2flox/flox mutant testes (Figure 5A and B), in agreement with the reduced HSD3B expression levels detected by immunofluorescence in Nr2f2 mutant testes (Figures 2J–M–3I–L). These observations suggest that FLC formed in the mutant testes have reduced steroidogenic function.
Figure 5 with 1 supplement see all
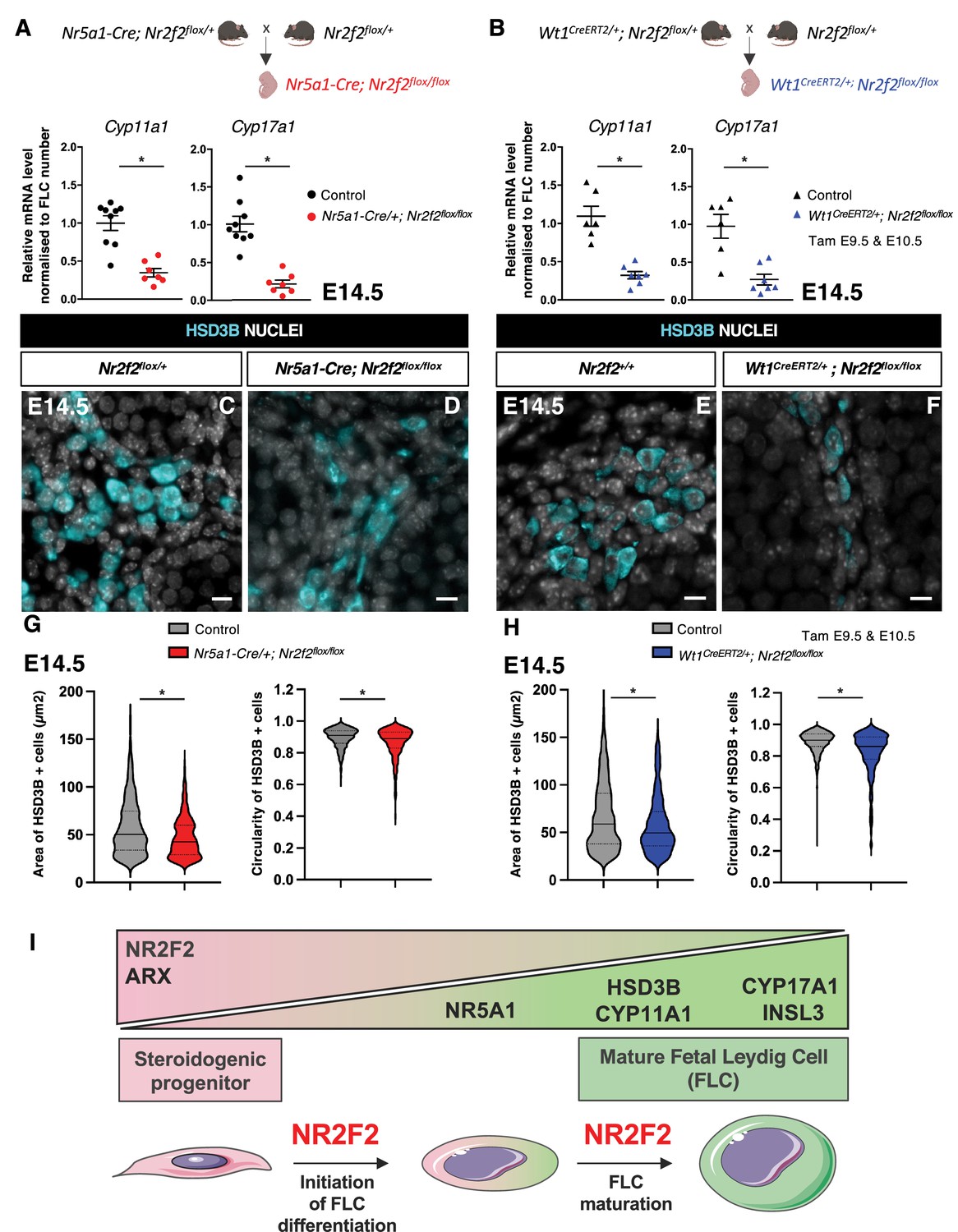
NR2F2 function is required for fetal Leydig cell (FLC) maturation.
(A) RT-qPCR quantification of Cyp11a1 and Cyp17a1 transcripts in control (wild-type or Nr2f2flox/+) and Nr5a1-Cre; Nr2f2flox/flox testes after normalization to Sdha and Tbp and to the number of FLC as quantified by HSD3B immunofluorescence at embryonic day 14.5 (E14.5). Data are shown as means ± SEM. Statistical significance was assessed by Mann-Whitney U two-tailed test. * indicates p-value≤0.05; ns indicates p-value>0.05. (B) RT-qPCR quantification of Cyp11a1 and Cyp17a1 transcripts in Nr2f2flox/+ and Wt1CreERT2; Nr2f2flox/flox gonads after normalization to Sdha and Tbp and to the number of FLC as quantified by HSD3B immunofluorescence at E14.5. Data are shown as means ± SEM. Statistical significance was assessed by Mann-Whitney U two-tailed test. * indicates p-value≤0.05; ns indicates p-value>0.05. (C,D) Immunodetection of HSD3B on E14.5 XY Nr2f2flox/+ and Nr5a1-Cre; Nr2f2flox/flox testes. (E,F) Immunodetection of HSD3B on E14.5 XY Nr2f2flox/+ and Wt1CreERT2; Nr2f2flox/flox testes after tamoxifen was administered at E9.5 and E10.5. (G) Quantification of the area and circularity of HSD3B positive cells in two E14.5 control (wild-type or Nr2f2flox/+, 737 cells, gray violin plot) and three Nr5a1-Cre; Nr2f2flox/flox (486 cells, red violin plot) testes. Statistical significance was assessed by Mann-Whitney U two-tailed test. * indicates p-value≤0.05; ns indicates p-value>0.05. (H) Quantification of the area and circularity of HSD3B positive cells in three E14.5 control (wild-type or Nr2f2flox/+, 1485 cells, gray violin plot) and three Wt1CreERT2; Nr2f2flox/flox (474 cells, blue violin plot) testes. Statistical significance was assessed by Mann-Whitney U two-tailed test. * indicates p-value≤0.05; ns indicates p-value>0.05. (I) Summary figure: NR2F2 is expressed in spindle-shaped interstitial steroidogenic progenitors together with ARX and is progressively downregulated upon FLC differentiation. NR2F2 is required for the initiation of FLC differentiation (marked by the upregulation of NR5A1) and for FLC maturation (characterized by the increase in cytoplasmic volume and the high expression of steroidogenic enzymes and Insl3). Immunodetection data are representative of triplicate biological replicates. Scale bar = 10 µm.
-
Figure 5—source data 1
Source data for cell measurements and RT-qPCR data in Figure 5.
- https://cdn.elifesciences.org/articles/103783/elife-103783-fig5-data1-v1.xlsx
FLC differentiation is accompanied by a change in cell shape from spindle-shaped progenitors to round-shaped FLC and an increase in cytoplasmic volume (van den Driesche et al., 2012; Haider, 2004). HSD3B positive FLC present in Nr5a1-Cre; Nr2f2flox/flox and in Wt1CreERT2; Nr2f2flox/flox mutants were smaller and more elongated than those in control littermates (Figure 5C–H, Figure 5—figure supplement 1). These cellular characteristics have been associated with immature FLC at the initial stages of FLC differentiation before the formation of large and round testosterone-producing FLC (van den Driesche et al., 2012; Haider, 2004).
Taken together, our results show that NR2F2 is required in interstitial steroidogenic progenitor cells for initiation of FLC differentiation but also for the subsequent step of FLC maturation leading to robust steroid production (Figure 5I).
Discussion
NR2F2 protein is expressed in interstitial cells of coelomic epithelium and mesonephric origin and is absent or detected at very low levels in FLC as soon as they are formed at E12.5. Our observations are consistent with NR2F2 positive cells being progenitors for FLC (Inoue et al., 2016; Ademi et al., 2022; Estermann et al., 2025) and indicate that NR2F2 is quickly downregulated upon differentiation of the fetal steroidogenic lineage, when the cells begin to synthesize steroid hormones.
We used two Cre lines to address the role of NR2F2 in the mouse fetal testis. Nr2f2 mutants exhibit reduced FLC numbers together with a strong reduction in Insl3 and steroidogenic gene expression, resulting in cryptorchidism and reduced anogenital distance. Importantly, the reduction of Insl3 expression is sufficient to impair testis descent even when a fraction of FLC is still present. Our findings identify NR2F2 as an essential regulator of FLC and male reproductive system development in the mouse. This conclusion is confirmed by an independent Nr2f2 conditional mutation resulting in a similar phenotype of FLC hypoplasia, cryptorchidism, and hypospadias (Estermann et al., 2025).
Nr2f2 deletion using Nr5a1-Cre does not impair the survival or proliferation of the interstitial steroidogenic progenitor population, yet the FLC population is reduced in Nr5a1-Cre; Nr2f2flox/flox mutant testes. We found that the expression of the master regulator of steroidogenic differentiation of NR5A1 is not upregulated in Nr2f2 mutants, which is sufficient to account for the decrease in FLC numbers (Morohashi et al., 2020; Baba et al., 2018; Baba et al., 2014; Shima et al., 2018). ChIP-seq analysis of whole E14.5 testes has identified NR2F2 binding peaks in the regulatory region of Nr5a1 (Estermann et al., 2025). In addition, NR2F2 regulates gene expression by directly interacting with NR5A1 in MA-10 cells, an in vitro model for immature ALCs (Mendoza-Villarroel et al., 2014b; Mendoza-Villarroel et al., 2014a; Di-Luoffo et al., 2022). It would be interesting to explore whether an interaction between NR2F2 and NR5A1 contributes to the strong upregulation of Nr5a1 itself in differentiating FLCs and/or whether NR2F2 cooperates with other factors for the regulation of Nr5a1.
The activity of the main paracrine pathways regulating FLC differentiation was not significantly altered in Nr5a1-Cre; Nr2f2flox/flox mutant testes, as shown by the expression of target genes of the DHH, PDGFRA, and NOTCH signaling. These observations indicate that NR2F2 is a permissive factor in steroidogenic progenitors acting downstream or in cooperation with the signaling pathways regulating FLC differentiation. NR2F2 expression has been found to be modulated by Hedgehog and/or NOTCH signaling in other cell types, and whether a similar regulation of Nr2f2 expression by DHH and NOTCH signaling occurs in the steroidogenic progenitors remains unknown (Lee et al., 2006; Swift et al., 2014).
A fraction of FLC differentiates in Nr2f2 mutants, including when progenitors of both coelomic epithelium and mesonephric origins are targeted. This observation indicates that additional factors cooperate with NR2F2 to regulate the transition from the steroidogenic progenitor state to the differentiating FLC. The homeodomain protein ARX is expressed in Nr2f2 mutant steroidogenic progenitors and is a good candidate to be such a factor. Arx mutants exhibit decreased FLC numbers without defects in paracrine signals driving their differentiation, similar to Nr2f2 mutants (Miyabayashi et al., 2013).
FLC and ALC are morphologically, transcriptionally, and functionally distinct (Shima, 2019; Sararols et al., 2021), yet NR2F2 function is required for the differentiation of both lineages. In contrast to the situation in the fetal testis, NR2F2 is maintained in cells that have started to express steroidogenesis genes in the postnatal testis (Inoue et al., 2016; Ademi et al., 2022; Mendoza-Villarroel et al., 2014b). NR2F2 regulates the transition from the adult progenitor Leydig cell (characterized by their elongated shape, their ability to proliferate, and a low level of testosterone synthesis) to the immature ALC (characterized by their round shape, low mitotic activity, and increased testosterone production) (Qin et al., 2008; Haider, 2004). In agreement with its in vivo role promoting maturation along the ALC lineage, NR2F2 cooperates with NR5A1 and GATA4 to activate the transcription of Insl3, Star (encoding the cholesterol transporter), and Amhr2 (Mendoza-Villarroel et al., 2014b; Mendoza-Villarroel et al., 2014a; Di-Luoffo et al., 2022; Mehanovic et al., 2021; Mehanovic et al., 2022) in MA-10 cells.
Here, we found that the steroidogenic cells that differentiate in Nr2f2 mutant fetal testes exhibit a small size, an elongated shape, and reduced steroidogenic gene expression, features of the earliest stages of FLC differentiation (Wen et al., 2016; van den Driesche et al., 2012; Haider, 2004). A similar phenotype is obtained when Nr5a1 is deleted after the onset of FLC differentiation (Buaas et al., 2012). This suggests that in addition to controlling the initial engagement of steroidogenic progenitors into the FLC lineage and similar to the situation in the postnatal testis, NR2F2 promotes FLC maturation possibly by directly regulating the initial expression of genes involved in steroidogenesis. In agreement with this hypothesis, NR2F2 binding peaks are found in the regulatory regions of genes involved in lipid metabolism and cholesterol biosynthesis in ChIP-seq analysis of whole E14.5 testes (Estermann et al., 2025).
NR2F2 is required for additional aspects of fetal testis morphogenesis and differentiation. Testis cords are enlarged and abnormally shaped in Nr2f2 mutants. Testis cord development involves the formation of Sertoli-germ cell masses after E11.5 and their subsequent partition by growing wedges of interstitial cells and associated vascular branches at E12.5 (Cool et al., 2012). How NR2F2-dependent regulation of interstitial cell adhesion or migration contributes to this process will be the aim of future research.
Pathogenic variants in NR2F2 have been associated with congenital malformations, including congenital heart disease, congenital diaphragmatic hernia, and syndromic 46,XX testicular or ovo-testicular difference/disorder in sex development (DSD) (Polvani et al., 2019; Bashamboo et al., 2018). More recently, defects in the external genitalia (micropenis, hypospadias) and cryptorchidism have been associated with rare heterozygous variants in NR2F2 in 46,XY patients (Zidoune et al., 2022; Ganapathi et al., 2023; Wankanit et al., 2024). These phenotypes can be attributed to defects in testosterone-dependent masculinization and INSL3-dependent testis descent during gestation and could be explained by a failure of FLC differentiation in the fetal testis. NR2F2 is abundantly expressed in interstitial cells of fetal human testes, a population that likely contains the progenitors for FLC (Kilcoyne et al., 2014; van den Driesche et al., 2012; Lottrup et al., 2014; Taelman et al., 2024). The present work demonstrating that NR2F2 is required in the steroidogenic progenitors of the murine fetal testis for the initiation and progression of FLC differentiation provides an entry point in understanding the etiology of 46,XY DSD associated with pathogenic NR2F2 variants.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene (Mus musculus) | Nr2f2 | MGI | MGI:1352452 | |
| Genetic reagent (Mus musculus) | Nr2f2tm1Vc | Dr. M. Vasseur-Cognet | MGI:3578106 | |
| Genetic reagent (Mus musculus) | Wt1tm2(cre/ERT2)Wtp | Dr. William T Pu | MGI:7528785 | |
| Genetic reagent (Mus musculus) | Tg(Nr5a1-cre)2Klp | Dr. Keith L Parker | MGI:5493455 | |
| Chemical compound | Tamoxifen | Sigma-Aldrich | T5648 | 200 mg/kg body weight |
| Chemical compound | 5-Bromo-2'-deoxy-uridine | Sigma-Aldrich | B5002 | 50 mg/kg body weight |
| Chemical compound | Paraformaldehyde | EMS | 15710-S | 4% in PBS |
| Antibody | Anti-ACTA2 (Mouse monoclonal) | Gift from Dr. Chaponnier | IF (1:500) | |
| Antibody | Anti- Activated Caspase 3 (Rabbit polyclonal) | R&D Systems | AF835 (RRID:AB_2243952) | IF (1:200) |
| Antibody | Anti- AMH (Mouse monoclonal) | Bio-Rad | MCA2246 (RRID:AB_2226471) | IF (1:50) |
| Antibody | Anti- ARX (Rabbit polyclonal) | Gift from Pr. Morohashi and Dr. Inoue | IF (1:200) | |
| Antibody | Anti-COL4A1 (Rabbit polyclonal) | Abcam | ab19808 (RRID:AB_445160) | IF (1:400) |
| Antibody | Anti-CYP11A1 (Rabbit polyclonal) | Gift from Dr. Wilhelm | IF (1:200) | |
| Antibody | Anti-GATA4 (Goat polyclonal) | Santa Cruz Biotechnology | Sc-1237 (RRID:AB_2108747) | IF (1:200) |
| Antibody | Anti-GFP (Chicken polyclonal) | Abcam | Ab13970 (RRID:AB_300798) | IF (1:200) |
| Antibody | Anti-HSD3B (Goat polyclonal) | Santa Cruz Biotechnology | Sc-30820 (RRID:AB_2279878) | IF (1:200) |
| Antibody | Anti-HSD3B (Rabbit polyclonal) | Invitrogen | PA5-76669 (RRID:AB_2720396) | IF (1:500) |
| Antibody | Anti-Ki67 (Rabbit monoclonal) | Spring Bioscience | M3062 (RRID:AB_11219741) | IF (1:200) |
| Antibody | Anti-LAMA1 (Rabbit polyclonal) | Sigma-Aldrich | L9393 (RRID:AB_477163) | IF (1:200) |
| Antibody | Anti-NESTIN (Rabbit) | BioLegend | 839801 (RRID:AB_2565443) | IF (1:1000) |
| Antibody | Anti-NR2F2 (Mouse monoclonal) | R&D Systems | PP-H7147-00 (RRID:AB_2155627) | IF (1:200) |
| Antibody | Anti-NR5A1 (Rabbit polyclonal) | Cosmo Bio | KO611(RRID:AB_2861370) | IF (1:200) |
| Antibody | Anti-PDGFRA (Rabbit polyclonal) | Santa Cruz Biotechnology | SC-338 (RRID:AB_631064) | IF (1:200) |
| Antibody | Anti-PECAM-1 (Goat polyclonal) | Santa Cruz Biotechnology | Sc-1506 (RRID:AB_2161037) | IF (1:200) |
| Antibody | Anti-RUNX1 (Rabbit monoclonal) | Abcam | ab92336 (RRID:AB_2049267) | IF (1:500) |
| Antibody | Anti-SOX9 (Rabbit polyclonal) | Sigma-Aldrich | HPA001758 (RRID:AB_1080067) | IF (1:250) |
| Antibody | Anti-WT1 (Goat polyclonal) | R&D Systems | AF5729 (RRID:AB_2216239) | IF (1:200) |
| Commercial assay or kit | BrdU detection kit | Roche | 11 296 736 001 | |
| Commercial assay or kit | RNAscope Multiplex Fluorescent Reagent Kit v2 | Bio-Techne | 323110 | |
| Sequence-based reagent | Gli1 probe | Bio-Techne | 311001 | |
| Commercial assay or kit | RNeasy Micro Kit | QIAGEN | 74004 | |
| Commercial assay or kit | SYBR Green I Master | Roche | 04887352001 | |
| Software, algorithm | RefFinder | https://www.ciidirsinaloa.com.mx/RefFinder-master/?type=reference | ||
| Software, algorithm | OMERO | https://www.openmicroscopy.org/omero/ | ||
| Software, algorithm | GraphPad Prism | Graphpad Software, Inc, La Jolla, CA, USA | GraphPad Prism 10.2.1 |
Mouse strains and genotyping
Request a detailed protocolThe experiments described herein were carried out in compliance with the guidelines of the French Regulations for Animal Care and with the approval of the local Ethical Committee (APAFIS APAFIS#12789-2017121515109323 v1 and APAFIS#44072-2023061915491990 v5). Mouse lines were kept on a mixed background B6CBAF1/JRj. The Nr2f2tm1Vc line where Nr2f2 exon 1 sequences (encoding the DNA binding domain) are deleted upon CRE-mediated recombination (referred to as Nr2f2flox), the knock-in Wt1tm2(cre/ERT2)Wtp line where tamoxifen-inducible CreERT2 is produced by WT1 expressing cells (referred to as Wt1CreERT2), and the transgenic Tg(Nr5a1-cre)2Klp line where Cre expression is driven by Nr5a1 regulatory sequences (referred to as Nr5a1-Cre) were genotyped as previously described (Bardoux et al., 2005; Zhou et al., 2008; Bingham et al., 2006). Wt1CreERT2; Nr2f2flox/+ or Nr5a1-Cretg/0; Nr2f2flox/+ males were crossed with Nr2f2flox/+ females to obtain mutant embryos at different stages. Embryos were named controls (Nr2f2+/+ or Nr2f2flox/+) or mutants (Wt1CreERT2; Nr2f2flox/flox or Nr5a1-Cretg/0; Nr2f2flox/flox). Genotypes of mice and embryos were determined using PCR assays on lysates from ear biopsies or tail tips. Genotyping primers are listed in Appendix 1. To activate the CreERT2 recombinase in embryos, tamoxifen (TAM, T5648, Sigma-Aldrich) was directly diluted in corn oil to a concentration of 40 mg/mL, and two TAM treatments (200 mg/kg body weight) were administered to pregnant females by oral gavage at E9.5 and E10.5. For proliferation assays, 5-bromo-2'-deoxy-uridine (BrdU) (B5002, Sigma-Aldrich) diluted to a concentration of 10 mg/mL in sterile H2O was administered to the pregnant females (50 mg/kg body weight) by intraperitoneal injection, and pregnant females and their embryos were humanely killed after 3 hr and 30 min. The day when a vaginal plug was found was designated as E0.5. E11.5–E12.5 embryos were staged by counting the number of ts with 18 ts corresponding to E11.5.
Immunofluorescence staining and in situ hybridization
Request a detailed protocolEmbryos were fixed in 4% (wt/vol) paraformaldehyde (PFA, 15,710-S, EMS) overnight, processed for paraffin embedding, and sectioned into 5-µm-thick sections. Immunofluorescence and DAPI staining were performed as described in Tang et al., 2020. Proliferation analysis was performed by using a BrdU detection kit (11 296 736 001, Roche). Gli1 mRNA was detected with the RNAscope technology (probe 311001) according to Advanced Cell’s instructions using the RNAscope Multiplex Fluorescent Reagent Kit v2 Assay. Images were obtained on a motorized Axio Imager Z1 microscope (Zeiss) coupled with an AxioCam MRm camera (Zeiss) and processed with Fiji (Bethesda, MD, USA). The DAPI staining marking the nuclei was adjusted to visualize the tissues and may vary between samples. However, for the immunofluorescence analysis, the exposure time of the acquisition of the fluorescent signal was identical in the same experiment to allow comparison between controls and mutants. Images were assembled using the open-source software platform OMERO (https://www.openmicroscopy.org/omero/). Antibodies are listed in the Key resources table. At least three embryos of each genotype were analyzed for each marker.
Cell quantifications
Request a detailed protocolThe gonadal area for each section was measured by creating a gonadal region of interest (ROI) drawn manually in Fiji. The number of HSD3B positive cells (2–493 cells per section depending on the stages and genotypes), SOX9 positive cells (231–882 cells per section depending on the stages and genotypes), NR5A1 positive cells (3–256 cells per section depending on the stages and genotypes), ARX positive cells (666–2487 cells per section depending on the stages and genotypes), ARX positive cells that had incorporated BrdU (232–656 cells per section depending on the stages and genotypes), activated caspase 3 positive cells (0–6 cells per section depending on the stages and genotypes), or the number of nuclei labeled by DAPI (1582–4279 cells per section depending on the stages and genotypes) were counted manually in the entire gonadal section using the Cell Counter Plugin from Fiji. For each genotype, gonads of three or four biological replicates were analyzed. Two to three coronal sections spaced by at least 30 µm in the medial regions of the gonads were analyzed for each individual. Statistical significance was assessed by Mann-Whitney U two-tailed test. * indicates p-value≤0.05; ns indicates p-value>0.05.
Quantification of area and circularity of HSD3B positive cells
Request a detailed protocolGonadal (ROI) was drawn manually, and HSD3B positive cells were segmented using the Stardist Deep Learning plugin of Fiji with a minimum area of 20 µm2 to remove small particles. The area and circularity of each segmented cell were measured with Fiji. Circularity = 4π*area/perimeter^2. A value of 1.0 indicates a perfect circle. As the value approaches 0.0, it indicates an increasingly elongated shape. For each genotype, two to three biological replicates were analyzed. The data are shown as violin plots (with median and quartiles) for control and mutant samples.
RNA extraction and quantitative PCR analysis
Request a detailed protocolIndividual gonads were dissected from the mesonephros in PBS, snap-frozen in liquid nitrogen, and kept at −80°C. RNA was extracted by RNeasy Micro Kit (74004, QIAGEN) and reverse-transcribed by M-MLV reverse transcriptase (M170A, Promega). The cDNA was used as a template for quantitative PCR analysis using the SYBR Green I Master (04887352001, Roche) and a LightCycler 480 System (Roche). Primer sequences are listed in Appendix 1.
All biological replicates of different genotypes (N=3–9) were run in the same plate and run as duplicate technical replicates. Relative gene expression of each gonad was normalized to the expression of the housekeeping genes Shda and Tbp (Yokoyama et al., 2018) by the 2-ΔΔCt calculation method. GeNorm, BestKeeper algorithms, and the comparative delta-Ct method provided through the online tool RefFinder (https://www.ciidirsinaloa.com.mx/RefFinder-master/?type=reference#) were used to confirm reference gene stability in the experimental datasets. Fold change in gene expression was obtained by dividing the normalized gene expression in gonads of a given genotype by the mean of the normalized gene expression in control gonads. Data are shown as means ± SEM. Statistical significance was assessed by Mann-Whitney U two-tailed test (GraphPad Prism 10.2.1). * indicates p-value≤0.05; ns indicates p-value>0.05.
Appendix 1
Primers
| Primer name | Application | Sequence (5’-3’) |
|---|---|---|
| CRE 1 | Genotyping Cre | CAGGATATACGTAATCTGGC |
| CRE 4 | Genotyping Cre | CACGGGCACTGTGTCCAGACCAG |
| CCR5mR | Genotyping internal control | ATGTGGATGGAGAGGAGTCG |
| CCR5mL | Genotyping internal control | CAACCGAGACCTTCCTGTTC |
| TF2.1 | Genotyping Nr2f2 | TGCCCACACTTTCCTACTCC |
| TF2.5 | Genotyping Nr2f2 | TTTCTGCAAGGAATGGGTTG |
| Amh-Fwd | RT-qPCR primers | GGGGAGACTGGAGAACAGC |
| Amh-Rev | RT-qPCR primers | AGAGCTCGGGCTCCCATA |
| Arx-Fwd | RT-qPCR primers | GCACCACGTTCACCAGTTAC |
| Arx-Rev | RT-qPCR primers | GCACCACGTTCACCAGTTAC |
| Bhmt-Fwd | RT-qPCR primers | CGTCAGCTTCATCGGGAGTT |
| Bhmt-Rev | RT-qPCR primers | CTTGCCGTGCAATGTCACAA |
| Cdh5-Fwd | RT-qPCR primers | TCCTCTGCATCCTCACCATCACA |
| Cdh5-Rev | RT-qPCR primers | GTAAGTGACCAACTGCTCGTGAAT |
| Cyp11a1-Fwd | RT-qPCR primers | TGGCCCCATTTACAGGGAGAA |
| Cyp11a1-Rev | RT-qPCR primers | GGCATCTGAACTCTTAAACAGGA |
| Cyp17a1-Rev | RT-qPCR primers | CAGAGAAGTGCTCGTGAAGAAG |
| Cyp17a1Fwd | RT-qPCR primers | CAGAGAAGTGCTCGTGAAGAAG |
| Dhh-Fwd | RT-qPCR primers | GGACCTCGTACCCAACTACAA |
| Dhh-Rev | RT-qPCR primers | CGATGGCTAGAGCGTTCACC |
| Gli1-Fwd | RT-qPCR primers | TGGTACCATGAGCCCTTCTT |
| Gli1-Rev | RT-qPCR primers | GTGGTACACAGGGCTGGACT |
| Hes1-Fwd | RT-qPCR primers | ATAGCTCCCGGCATTCCAAG |
| Hes1-Rev | RT-qPCR primers | ATAGCTCCCGGCATTCCAAG |
| HeyL-Fwd | RT-qPCR primers | CAGCCCTTCGCAGATGCAA |
| HeyL-Rev | RT-qPCR primers | CCAATCGTCGCAATTCAGAAAG |
| Insl3-Fwd | RT-qPCR primers | ATTGCTCCCCACCTCCTGGCTATG |
| Insl3-Rev | RT-qPCR primers | GGTCATGATGGGGCTTCTTGGGGA |
| Notch2-Fwd | RT-qPCR primers | ATATCGACGACTGCCCCAAC |
| Notch2-Rev | RT-qPCR primers | CCATAGCCTCCGTTTCGGTT |
| Nr2f2-Fwd | RT-qPCR primers | CGGAGGAACCTGAGCTACAC |
| Nr2f2-Rev | RT-qPCR primers | CGGAGGAACCTGAGCTACAC |
| Pdgfa-Fwd | RT-qPCR primers | CAGTGTCAAGGTGGCCAAAGT |
| Pdgfa-Rev | RT-qPCR primers | CAGTGTCAAGGTGGCCAAAGT |
| Pdgfb-Fwd | RT-qPCR primers | GATCTCTCGGAACCTCATCG |
| Pdgfb-Rev | RT-qPCR primers | GATCTCTCGGAACCTCATCG |
| Pdgfra-Fwd | RT-qPCR primers | TCCATGCTAGACTCAGAAGTCA |
| Pdgfra-Rev | RT-qPCR primers | TCCCGGTGGACACAATTTTT |
| Sdha-Fwd | RT-qPCR primers | TGTTCAGTTCCACCCCACA |
| Sdha-Rev | RT-qPCR primers | TCTCCACGACACCCTTCTG |
| Sgpl1-Fwd | RT-qPCR primers | CTGAAGGACTTCGAGCCTTATTT |
| Sgpl1-Rev | RT-qPCR primers | CTGAAGGACTTCGAGCCTTATTT |
| Sox9-Fwd | RT-qPCR primers | GCGGAGCTCAGCAAGACTCTG |
| Sox9-Rev | RT-qPCR primers | ATCGGGGTGGTCTTTCTTGTG |
| Tbp-Fwd | RT-qPCR primers | GCTCTGGAATTGTACCGCAG |
| Tbp-Rev | RT-qPCR primers | TGACTGCAGCAAATCGCTTG |
Data availability
All data generated or analysed during this study are included in the manuscript and supporting files.
References
-
The expression of the Wilms’ tumour gene, WT1, in the developing mammalian embryoMechanisms of Development 40:85–97.https://doi.org/10.1016/0925-4773(93)90090-k
-
Glycolytic genes are targets of the nuclear receptor Ad4BP/SF-1Nature Communications 5:3634.https://doi.org/10.1038/ncomms4634
-
Loss of function of the nuclear receptor NR2F2, encoding COUP-TF2, causes testis development and cardiac defects in 46,XX childrenAmerican Journal of Human Genetics 102:487–493.https://doi.org/10.1016/j.ajhg.2018.01.021
-
GATA-4 is required for sex steroidogenic cell development in the fetal mouseDevelopmental Dynamics 236:203–213.https://doi.org/10.1002/dvdy.21004
-
Testis formation in the fetal mouse: dynamic and complex de novo tubulogenesisWiley Interdisciplinary Reviews. Developmental Biology 1:847–859.https://doi.org/10.1002/wdev.62
-
Two distinct origins for Leydig cell progenitors in the fetal testisDevelopmental Biology 352:14–26.https://doi.org/10.1016/j.ydbio.2011.01.011
-
Heterozygous rare variants in NR2F2 cause a recognizable multiple congenital anomaly syndrome with developmental delaysEuropean Journal of Human Genetics 31:1117–1124.https://doi.org/10.1038/s41431-023-01434-5
-
Cell biology of Leydig cells in the testisInternational Review of Cytology 233:181–241.https://doi.org/10.1016/S0074-7696(04)33005-6
-
Regulation of testicular descentPediatric Surgery International 31:317–325.https://doi.org/10.1007/s00383-015-3673-4
-
Cell-specific knockout of steroidogenic factor 1 reveals its essential roles in gonadal functionMolecular Endocrinology 18:1610–1619.https://doi.org/10.1210/me.2003-0404
-
Sertoli cells of the mouse testis originate from the coelomic epitheliumDevelopmental Biology 203:323–333.https://doi.org/10.1006/dbio.1998.9068
-
A perivascular niche for multipotent progenitors in the fetal testisNature Communications 9:4519.https://doi.org/10.1038/s41467-018-06996-3
-
Indian hedgehog is a major mediator of progesterone signaling in the mouse uterusNature Genetics 38:1204–1209.https://doi.org/10.1038/ng1874
-
The INSL3 gene is a direct target for the orphan nuclear receptor, COUP-TFII, in Leydig cellsJournal of Molecular Endocrinology 53:43–55.https://doi.org/10.1530/JME-13-0290
-
Coordination of multiple cellular processes by NR5A1/NR5A1Endocrinology and Metabolism 35:756–764.https://doi.org/10.3803/EnM.2020.402
-
The orphan nuclear receptor COUP-TFII is required for angiogenesis and heart developmentGenes & Development 13:1037–1049.https://doi.org/10.1101/gad.13.8.1037
-
A developmental study of the Desert hedgehog-null mouse testisBiology of Reproduction 65:1392–1402.https://doi.org/10.1095/biolreprod65.5.1392
-
Genetic control of typical and atypical sex developmentNature Reviews. Urology 20:434–451.https://doi.org/10.1038/s41585-023-00754-x
-
Specific transcriptomic signatures and dual regulation of steroidogenesis between fetal and adult mouse Leydig cellsFrontiers in Cell and Developmental Biology 9:695546.https://doi.org/10.3389/fcell.2021.695546
-
The PDGF signaling pathway controls multiple steroid-producing lineagesGenes & Development 22:3255–3267.https://doi.org/10.1101/gad.1723908
-
Contribution of Leydig and Sertoli cells to testosterone production in mouse fetal testesMolecular Endocrinology 27:63–73.https://doi.org/10.1210/me.2012-1256
-
Leydig progenitor cells in fetal testisMolecular and Cellular Endocrinology 445:55–64.https://doi.org/10.1016/j.mce.2016.12.006
-
Development of fetal and adult Leydig cellsReproductive Medicine and Biology 18:323–330.https://doi.org/10.1002/rmb2.12287
-
Development, function and fate of fetal Leydig cellsSeminars in Cell & Developmental Biology 59:89–98.https://doi.org/10.1016/j.semcdb.2016.03.003
-
Desert Hedgehog/Patched 1 signaling specifies fetal Leydig cell fate in testis organogenesisGenes & Development 16:1433–1440.https://doi.org/10.1101/gad.981202
-
Identification of reference genes for quantitative PCR analyses in developing mouse gonadsThe Journal of Veterinary Medical Science 80:1534–1539.https://doi.org/10.1292/jvms.18-0417
-
Targeted disruption of the Insl3 gene causes bilateral cryptorchidismMolecular Endocrinology 13:681–691.https://doi.org/10.1210/mend.13.5.0272
Article and author information
Author details
Aitana Perea-Gomez

Natividad Bellido Carreras
Marie-Christine Chaboissier
Funding
Agence Nationale de la Recherche (ANR-23-CE14-0012)
- Marie-Christine Chaboissier
China Scholarship Council
- Furong Tang
Agence Nationale de la Recherche (Heterosex)
- Marie-Christine Chaboissier
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We acknowledge the help from members of the Experimental Histopathology Platform, the PRISM Imaging Platform, and the Animal House at iBV (Institut de Biologie Valrose, Université Côte d'Azur, CNRS, Inserm, iBV, France). We are grateful to members of the A Schedl, MC Chaboissier, and S Nef groups for helpful discussions and to D Wilhelm for critical reading of the manuscript. We are indebted to Dr. Chaponnier, Dr. Inoue, Pr Morohashi, Dr. M Vasseur-Cognet, and Dr. Wilhelm for sharing mouse lines and reagents. This research was funded by Agence Nationale de la Recherche ANR-23-CE14-0012, Heterosex, and by a scholarship from the China Scholarship Council (to FT).
Ethics
The experiments described herein were carried out in compliance with the relevant institutional and French animal welfare laws, guidelines, and policies (APAFIS APAFIS#12789-2017121515109323 v1 and APAFIS#44072-2023061915491990 v5).
Version history
- Preprint posted:
- Sent for peer review:
- Reviewed Preprint version 1:
- Reviewed Preprint version 2:
- Version of Record published:
Cite all versions
You can cite all versions using the DOI https://doi.org/10.7554/eLife.103783. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2025, Perea-Gomez et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,355
- views
-
- 93
- downloads
-
- 8
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 5
- citations for umbrella DOI https://doi.org/10.7554/eLife.103783
-
- 1
- citation for Reviewed Preprint v1 https://doi.org/10.7554/eLife.103783.1
-
- 1
- citation for Reviewed Preprint v2 https://doi.org/10.7554/eLife.103783.2
-
- 1
- citation for Version of Record https://doi.org/10.7554/eLife.103783.3
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
NR2F2 is required in the embryonic testis for fetal Leydig cell development
eLife 14:RP103783.
https://doi.org/10.7554/eLife.103783.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}