PRDM16 functions as a co-repressor in the BMP pathway to suppress neural stem cell proliferation
- Department of Molecular Bioscience, the Wenner-Gren Institute, Stockholm University, Sweden
- Division of Genome Sciences and Cancer, The John Curtin School of Medical Research, The Australian National University and Australian Research Council Centre of Excellence for the Mathematical Analysis of Cellular Systems, Australia
eLife Assessment
This valuable work presents how PRDM16 plays a critical role during colloid plexus development, through regulating BMP signaling. Solid evidence supports the context-dependent gene regulatory mechanisms both in vivo and in vitro. The work will be of broad interest to researchers working on growth factor signaling mechanisms and vertebrate development.
https://doi.org/10.7554/eLife.104076.3.sa0Significance of the findings:
Valuable: Findings that have theoretical or practical implications for a subfield
- Landmark
- Fundamental
- Important
- Valuable
- Useful
Strength of evidence:
Solid: Methods, data and analyses broadly support the claims with only minor weaknesses
- Exceptional
- Compelling
- Convincing
- Solid
- Incomplete
- Inadequate
During the peer-review process the editor and reviewers write an eLife Assessment that summarises the significance of the findings reported in the article (on a scale ranging from landmark to useful) and the strength of the evidence (on a scale ranging from exceptional to inadequate). Learn more about eLife Assessments
Abstract
BMP signaling acts as an instructive cue in various developmental processes such as tissue patterning, stem cell proliferation, and differentiation. However, it is not fully understood how this signaling pathway generates different cell-specific outputs. Here, we have identified PRDM16 as a key co-factor for BMP signaling in the mouse brain. PRDM16 contributes to a repressive role of BMP signaling on neural stem cell (NSC) proliferation. We demonstrate that PRDM16 regulates the genomic distribution of BMP pathway transcription factors, the SMAD4/pSMAD complex, preventing the activation of cell proliferation genes. When Prdm16 is lost, the SMAD complex relocates to nearby genomic regions, leading to abnormal upregulation of BMP target genes. This function of PRDM16 is also required for the specification of choroid plexus (ChP) epithelial cells. Through a single-cell resolution fluorescent in situ approach, we have observed that genes co-repressed by SMAD and PRDM16, such as Wnt7b and several cell cycle regulators, become overexpressed in Prdm16 mutant ChP. Our findings elucidate a mechanism through which SMAD4 and pSMAD1/5/8 repress gene expression. Moreover, our study suggests a regulatory circuit composed of BMP and Wnt signaling, along with PRDM16, in controlling stem cell behaviors.
Introduction
Uncontrollable cell proliferation can lead to tumor growth, while premature differentiation can result in tissue degeneration. The balance between stem cell proliferation and differentiation is a crucial aspect in developmental and stem cell biology. BMP (Bone morphogenetic proteins) signaling is a key cell-signaling pathway in regulating stem cell proliferation and maintaining adult stem cell quiescence (Mira et al., 2010; Colak et al., 2008; Zhang et al., 2016). Moreover, BMP signaling is essential in various cell specification processes (Zhang and Li, 2005; Hogan, 1996; Massagué, 1998).
The ability of a single pathway to play a diverse range of roles relies on context-specific transcriptional outputs. BMPs signal through two types of SMAD proteins: the receptor SMADs (R-SMADs) - Smad1, 5, and 8, and the co-SMAD protein SMAD4. Upon ligand binding, heterodimeric receptors like BMPRI and BMPRII phosphorylate R-SMADs, leading to the assembly and nuclear translocation of the SMAD complex with SMAD4. This complex then regulates gene expression by binding to enhancers of BMP target genes. Apart from transducing BMP signaling, SMAD4 is an essential effector in TGF-β/Activin signaling. In response to ligands such as TGF-β and Activin, SMAD4 associates with two other R-SMADs - phosphorylated SMAD 2 and 3, regulating downstream genes of the TGF-β/Activin signaling pathway. These SMADs recognize and directly bind to two main types of short DNA motifs in target enhancers via their N-terminal MH1 domain (Martin-Malpartida et al., 2017). However, since the binding is generally weak, SMAD complexes rely on various co-factors such as transcription factors, co-activators and co-repressors to activate or repress target gene expression (Hill, 2016).
In the mammalian brain, BMP and Wnt signaling pattern the brain midline where an essential structure, the ChP, emerges. Neural epithelial cells at the presumptive ChP site lose neural potential and exit the cell cycle. Only a small number of cells at the border of the ChP primordium and cortical hem (CH) persist as slowly dividing ChP progenitors, leading to the expansion of the monolayered ChP epithelial cells (Liddelow et al., 2010). BMP signaling is essential for ChP epithelium specification. Conditional depletion of the BMP receptor BMPr1a diminishes ChP development (Hébert et al., 2002), whereas ectopic BMP transforms neural progenitors into ChP cells (Watanabe et al., 2012). Wnt signaling, peaking at CH, is also necessary for ChP epithelial cell specification (Parichha et al., 2022). Conditional depletion of ß-Catenin results in defective ChP, and overexpression of ß-Catenin expands CH at the expense of the ChP epithelium. These findings emphasize the importance of tightly controlling Wnt and BMP signaling levels for proper cell specification.
The PR domain-containing (PRDM) family protein PRDM16 determines cell fate specification in various cell types (Aguilo et al., 2011; Chuikov et al., 2010; Kajimura et al., 2008; Shimada et al., 2017; Chui et al., 2020; Baizabal et al., 2018; Inoue et al., 2017). Previous studies, including our own, demonstrated that Prdm16 knockout (KO) mouse brains show severely reduced ChP structures (He et al., 2021; Bjork et al., 2010; Strassman et al., 2017). Interestingly, it was shown that PRDM16 can interact with TGF-ß pathway SMAD proteins in vitro and impact TGF-ß signaling output in craniofacial tissues (Bjork et al., 2010; Warner et al., 2007). PRDM16 is also a downstream effector of BMP signaling during brown adipocyte specification (Seale et al., 2007; Tseng et al., 2008). A recent study reported that PRDM16 and its ortholog PRDM3 (also known as Evi1) regulate Wnt signaling during craniofacial development in zebrafish (Shull et al., 2022). These findings suggest that PRDM16 may be more broadly involved in regulating BMP/TGF-ß and Wnt signaling. However, the underlying molecular mechanisms remain unclear.
Consistent with its essential roles in normal development, mutations and dysregulation of Prdm16 are linked with several human diseases, including those identified in patients with 1p36 chromosomal aberrations and cardiomyopathy (Arndt et al., 2013). PRDM16 exhibits versatile functions at the molecular level, regulating chromatin accessibility and epigenetic states of its bound enhancers (Baizabal et al., 2018; He et al., 2021). Depending on associated cofactors, PRDM16 can either repress or activate gene expression. This dual role poses a challenge when considering PRDM16 as a therapeutic target, as an undesired outcome may occur. Thus, a comprehensive understanding of the regulatory roles of this protein in each specific process is crucial for effective disease treatment strategies.
In this study, we have investigated the mechanisms that regulate the transition between stem cell proliferation and differentiation during ChP development. We show that Prdm16 mutant ChP cells fail to exit the cell cycle, a similar phenotype to when BMP signaling is abolished (Hébert et al., 2002). Using primary NSC culture, we dissected the molecular interaction of SMAD4/pSMAD1/5/8 proteins with PRDM16, and found that PRDM16 functions as a co-repressor that holds the SMAD complex at distal enhancers, repressing genes involved in cell proliferation. Finally, we validated that some of the co-regulated genes by BMP signaling and PRDM16 become de-repressed in the Prdm16 mutant ChP. These findings uncover an essential function of PRDM16 in stem cell regulation and BMP and Wnt signalling.
Results
PRDM16 promotes cell cycle exiting of neural epithelial cells at the ChP primordium
To understand how Prdm16 regulates ChP epithelial specification, we first investigated the expression of this gene in the developing mouse brain. At embryonic day 10.5 (E10.5), BMP signaling specifies the presumptive ChP (Hébert et al., 2002). Prdm16 mRNAs and nuclear localization of the PRDM16 protein become detectable in the presumptive ChP at this stage (Figure 1A, Figure 1—figure supplement 1A). As previously shown, Prdm16cGT, a Prdm16 knockout allele (Prdm16 KO), displayed severely reduced ChP structure at E13.5 (Figure 1B; He et al., 2021; Bjork et al., 2010; Strassman et al., 2017). Expression of Ttr, a classic ChP marker gene, is reduced in both the lateral ventricle (tChPs) (Figure 1C) and the fourth ventricle (hChP) (Figure 1—figure supplement 1B) in the Prdm16 KO mutant, suggesting that the function of PRDM16 is not area-restricted but generally required for ChP development.
Figure 1 with 1 supplement see all
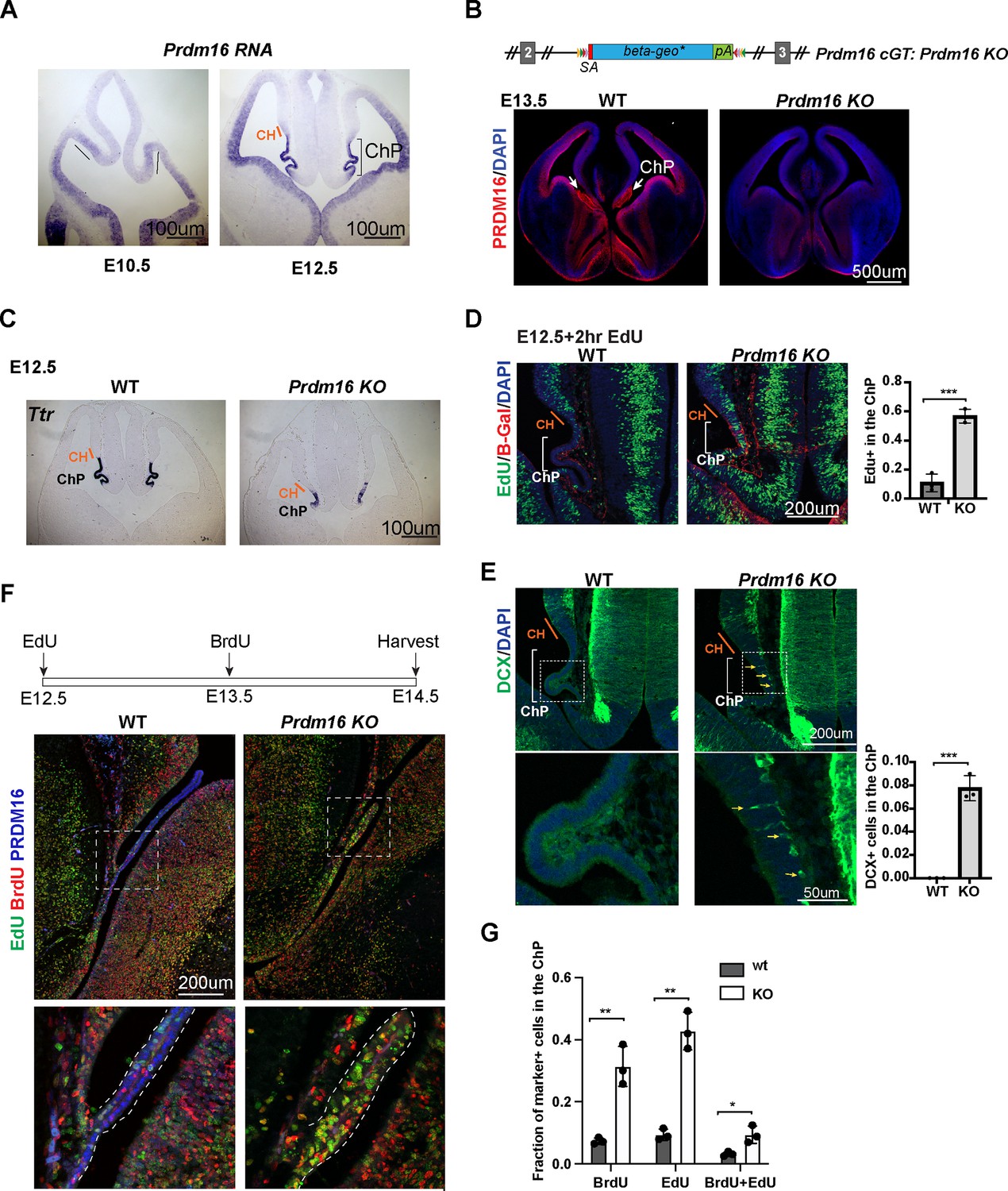
Prdm16 mutant choroid plexus (ChP) cells fail to exit the cell cycle and retain neural progenitor identity.
(A) RNA in situ hybridization using a Prdm16 probe shows Prdm16 expression in E10.5 and E12.5 brains. The line and bracket mark the developing CH and ChP. (B) Top: Schematic of the Prdm16 GT allele, as reported in Strassman et al., 2017. Bottom: immunostaining of PRDM16 in E13.5 control and null mutant brains. Red: PRDM16; blue: DAPI. White arrows indicate the ChP in the control brain. (C) RNA in situ hybridization for Ttr on E12.5 control and Prdm16 null mutant forebrains. Cortical hem (CH) regions are outlined based on morphology. (D) Two-hour EdU labeling in E12.5 control and Prdm16 homozygous brains. Sections are co-stained with anti-β-Gal antibody to identify mutant ChP cells in the KO brain. Note: the β-Geo trap is absent in wild-type animals. Brackets mark ChP regions. Red: β-Gal; blue: DAPI; green: EdU. (E) Immunostaining for Doublecortin (DCX) on E12.5 control and Prdm16 homozygous brain slices. Bottom panels show magnified views of the ChP (dashed rectangles). Yellow arrows indicate some of the DCX-positive cells. Right panels show quantification of EdU+ (D) and DCX+ cells (E). Biological replicates: N=3. Error bars represent standard deviation (SD). Statistical significance is calculated using unpaired t-test. ***p<0.001. (F) Schematic of the double S-phase labeling experiment (top). Brain sections from E14.5 control and prdm16 KO embryos were immunostained with PRDM16 and BrdU antibodies and processed for EdU detection. Bottom panels show magnified views of the ChP region. (G) Quantification of EdU+ and BrdU+ cells from the double labeling experiment. Biological replicates: N=3. Error bars represent standard deviation (SD). Statistical significance is calculated using unpaired t-test. **p<0.01. *p<0.05.
-
Figure 1—source data 1
Counting of EdU+ and DAPI stained cells, related to Figure 1D.
- https://cdn.elifesciences.org/articles/104076/elife-104076-fig1-data1-v1.xlsx
-
Figure 1—source data 2
Counting of Dcx+ and DAPI stained cells, related to Figure 1E.
- https://cdn.elifesciences.org/articles/104076/elife-104076-fig1-data2-v1.xlsx
-
Figure 1—source data 3
Counting of EdU+, BrdU+, double positive and DAPI stained cells, related to Figure 1G.
- https://cdn.elifesciences.org/articles/104076/elife-104076-fig1-data3-v1.xlsx
To assess the patterning of the CH and ChP regions, we analyzed the expression of Wnt2b and BMP4 using conventional RNA in situ hybridization. Wnt2b expression, which marks the CH, appeared comparable between Prdm16 KO and control brains at E11.5 and E12.5 (Figure 1—figure supplement 1C), indicating that CH development is not affected by the loss of Prdm16. BMP4, which labels the ChP and CH, also showed normal expression levels and spatial distribution in the mutant brain. These findings suggest that the BMP-dependent patterning of the ChP and CH domains remains intact in the absence of Prdm16.
However, the ChP epithelial layer in Prdm16 mutants remained abnormally thick, resembling the adjacent neural epithelium (Figure 1C, Figure 1—figure supplement 1C). This observation led us to investigate the underlying cellular changes responsible for the ChP defects in the mutant. To assess cell proliferation, we performed a 2 hr EdU labeling at E12.5. In control embryos, ChP cells were largely EdU negative and formed a monolayer, indicating that most had exited the cell cycle. In contrast, Prdm16 mutant ChP cells marked by β-Gal remained highly proliferative (Figure 1D). These results suggest several possibilities: ChP epithelial cells are not properly specified in the mutant, mutant ChP cells are specified but fail to exit the cell cycle to differentiate, or both processes are impaired.
To explore these possibilities, we first examined the cell type composition of Prdm16 mutant ChP cells at E12.5. We stained brain sections from control and mutant embryos with the neuronal marker Doublecortin (Dcx). In controls, Dcx-positive cells were absent from the ChP, indicating that neural epithelial cells had successfully transitioned into ChP epithelial cells and lost their neural potential (Figure 1E). In contrast, the Prdm16 mutant ChP exhibited numerous Dcx-positive cells along the basal side of the epithelium, suggesting ongoing neurogenesis.
We next examined the expression of the NSC marker SOX2. In control brains, SOX2 is highly expressed in NSCs adjacent to the ChP epithelium but is significantly downregulated within the presumptive ChP region (Figure 1—figure supplement 1D). This downregulation was absent in the Prdm16 mutant. Together, these results indicate that at E12.5, Prdm16 depletion disrupts the normal transition of neural progenitors into ChP epithelial cells, maintaining cells in a proliferative, undifferentiated state.
We then asked whether PRDM16 not only regulates ChP epithelial specification but also directly restricts cell proliferation. To test this, we examined Prdm16 mutant ChP cells at a later stage. We performed double S-phase labeling by injecting EdU at E12.5 and BrdU at E13.5, followed by sample collection at E14.5. By this stage, the wild-type ChP had developed into an elongated, monolayered epithelium in which most cells were negative for both EdU and BrdU, indicating cell cycle exit. In contrast, the Prdm16 mutant ChP remained a disorganized cluster of cells, many of which were EdU-positive, BrdU-positive, or both (Figure 1F–G). This finding suggests that although the mutant ChP can separate from the neuroepithelium at later stages, it fails to exit the cell cycle and differentiate, supporting a direct role of PRDM16 in restricting proliferation.
Given that BMP signaling is a key regulator of ChP formation and that the BMP pathway mutants, such as Foxg1cre::Bmpr1afl/fl also exhibit ectopic cell proliferation in the developing ChP (Hébert et al., 2002), we next investigated whether PRDM16 interacts with the BMP pathway to control cell proliferation.
PRDM16 and BMP signaling collaborate to induce NSC quiescence in vitro
To investigate the molecular interplay between PRDM16 and BMP signaling in regulating cell proliferation, we turned to primary NSCs as an in vitro model. BMP signaling is known to maintain quiescence in adult NSC in vivo and can induce proliferative NSCs into quiescence in vitro (Mira et al., 2010). Furthermore, previous studies have shown that embryonic cortical NSCs are responsive to BMP4 treatment (Hu et al., 2008), making them a suitable system for probing downstream BMP signaling events. Based on this, we established a cell culture assay to evaluate the effects of BMP4 and PRDM16 on NSC proliferation and quiescence (Figure 2A).
Figure 2 with 1 supplement see all
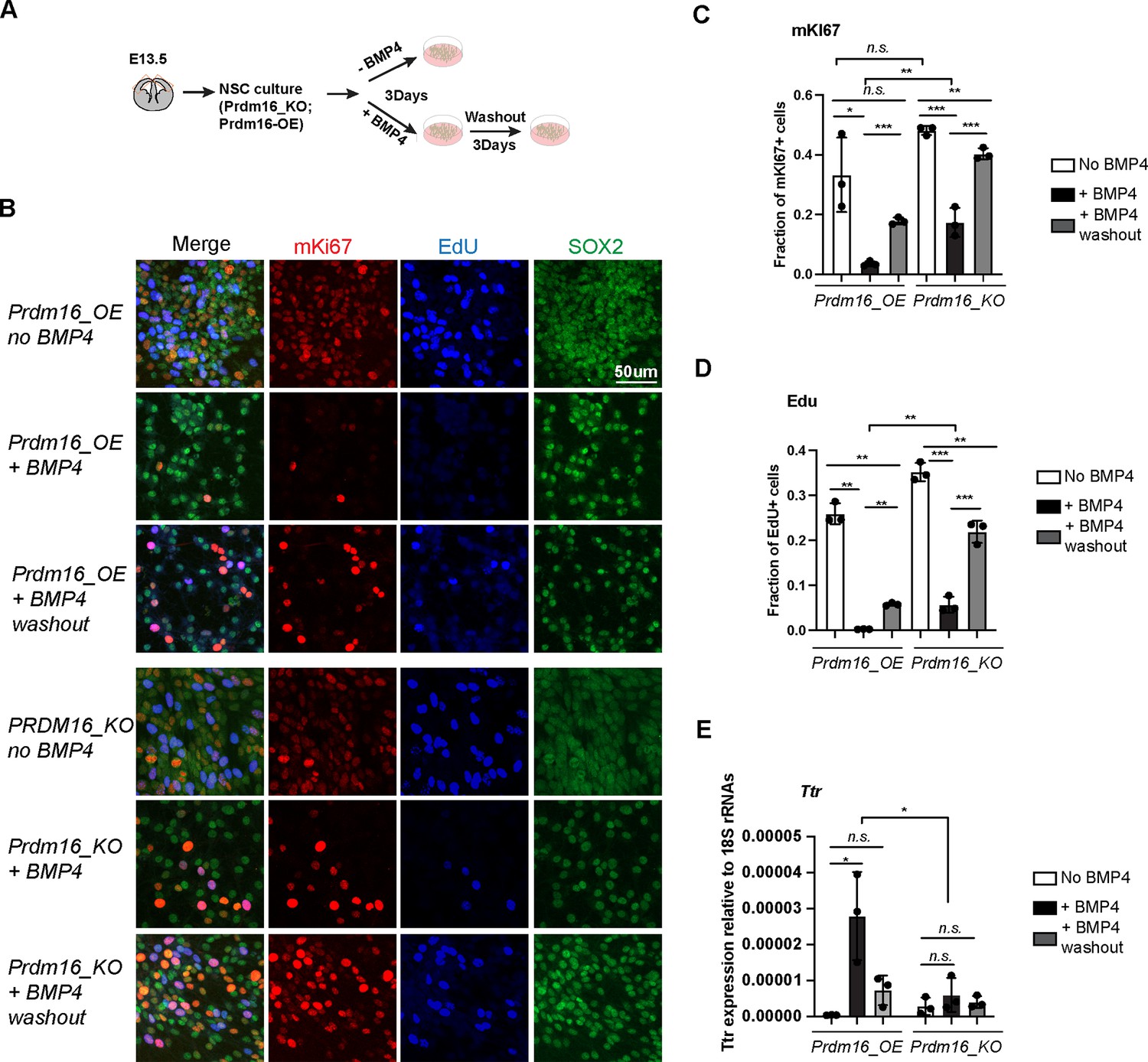
Prdm16 is required for BMP4-induced neural stem cell (NSC) quiescence.
(A) Schematic of the NSC culture assay. (B) Immunostaining of mKI67 in red and SOX2 in green and EdU labeling in blue on indicated NSC genotypes and treatment. (C–D) Quantification of the fraction of mKI67+ (C) and EdU+ (D) cells among the total cell number marked by SOX2. (E) RT-qPCR measurement of Ttr levels relative to 18 S RNAs. Biological replicates: N=3. Error bars represent standard deviation (SD). Statistical significance is calculated using unpaired t-test. ***p<0.001; **p<001; *p<0.05; n.s., non-significant.
-
Figure 2—source data 1
Counting of mKI67 + and DAPI-stained cells, related to Figure 2C.
- https://cdn.elifesciences.org/articles/104076/elife-104076-fig2-data1-v1.xlsx
-
Figure 2—source data 2
Counting of EdU+ and DAPI-stained cells, related to Figure 2D.
- https://cdn.elifesciences.org/articles/104076/elife-104076-fig2-data2-v1.xlsx
-
Figure 2—source data 3
RTqPCR measurement of Ttr and 18srRNA expression, related to Figure 2E.
- https://cdn.elifesciences.org/articles/104076/elife-104076-fig2-data3-v1.xlsx
Unexpectedly, we found that PRDM16 protein levels were undetectable in cultured NSCs despite high levels of Prdm16 mRNAs (Figure 2—figure supplement 1A–B). Given that the PRDM16 protein is normally restricted to the nucleus of NSCs in embryonic brain tissues (Figure 1B, Figure 1—figure supplement 1A; He et al., 2021), this observation suggests that in vivo mechanisms may regulate PRDM16 protein stability and nuclear localization. Post-translational modification of PRDM16 has been identified as a key regulatory mechanism in brown adipocytes (Wang et al., 2022), and we speculate that a similar mechanism may operate in NSCs.
To bypass this limitation and examine PRDM16’s molecular function in regulating NSC proliferation and gene regulation, we generated a lentiviral construct expressing 3xNSL_Flag_Prdm16 under a constitutive promoter. Infection of wild-type primary NSCs with this construct yielded a cell line with robust Prdm16 mRNA expression and detectable nuclear PRDM16 protein (Figure 2—figure supplement 1A–B), which we referred to as Prdm16_overexpressing (Prdm16_OE). For comparison, we attempted to establish three additional lines: wild-type NSCs infected with the empty vector (wt_CDH), Prdm16_KO NSCs infected with the empty vector (KO_CDH), and Prdm16_KO NSCs infected with 3xNSL_Flag_Prdm16. However, we were unable to generate the last line despite repeated attempts. This failure was likely due to low viral production (the Prdm16 coding sequence exceeds 3 kb) and the increased sensitivity of KO NSCs to viral infection. Nevertheless, we proceeded with comparative analyses of Prdm16_OE cells against wt_CDH, KO_CDH, and uninfected KO NSCs.
To assess cell proliferation rates, we labeled NSCs with EdU and mKi67. BMP4 treatment of Prdm16_OE NSCs led to a marked reduction in the number of mKi67- and EdU-positive populations (Figure 2B–D). Following BMP4 washout, Prdm16_OE NSCs restored EdU and mKi67 labeling, indicating re-entry into the cell cycle. This reversible reduction confirms that BMP4 induces a quiescent, non-proliferative state in NSCs. In contrast, a larger proportion of Prdm16_KO cells failed to exit the cell cycle in response to BMP4, as shown by a less pronounced decrease in mKi67- and EdU-positive populations (Figure 2B–D). Notably, Prdm16_KO cells exhibited similar properties regardless of whether they were infected with the control viral vector (Figure 2—figure supplement 1D–F), and we, therefore, used both lines interchangeably in our analyses.
Previous studies have shown that BMP4 can program ChP cell fate and activate ChP-specific genes in neural progenitors cultured in vitro (Watanabe et al., 2012). To test whether PRDM16 cooperates with BMP signaling to induce ChP gene expression, we measured Ttr mRNA levels using reverse transcription followed by quantitative PCR (RT-qPCR). In Prdm16_OE NSCs, Ttr expression was robustly induced following BMP4 treatment. In contrast, Prdm16_KO NSCs and NSCs lacking the Prdm16_OE construct failed to upregulate Ttr in response to BMP4 (Figure 2E, Figure 2—figure supplement 1G).
Together, these results indicate that both BMP signaling and PRDM16 are required not only to restrict NSC proliferation but also to induce ChP gene expression. We next investigated the molecular mechanisms underlying this cooperation.
BMP signaling and PRDM16 cooperatively repress proliferation genes
To understand how BMP signaling and PRDM16 suppress cell proliferation, we aimed to determine the transcriptional targets of SMAD4 and pSMAD1/5/8 in cells with and without BMP4. We first applied Cleavage Under Targets and Tagmentation (CUT&TAG) experiments using a PRDM16 antibody and the available SMAD antibodies to profile their genomic binding sites. The PRDM16 antibody worked efficiently, but none of the SMAD antibodies produced libraries with sufficient sequencing reads. Subsequently, we employed chromatin immunoprecipitation followed by deep sequencing (ChIP-seq) for the SMAD proteins. Given that SMAD4 forms a complex with SMAD2/3 only in response to TGF-β/activin-type ligands, we included an antibody targeting SMAD3 as a control for non-BMP4-induced SMAD4 binding. Thus, our experiment settings enabled us to profile genomic binding sites for PRDM16, SMAD4, and two types of R_SMADs under conditions with endogenous BMPs and TGF-β/Activin, as well as ectopic BMP4.
In Prdm16_OE cells without BMP4, we identified several hundred to a few thousand ChIP-seq peaks for all three classes of SMAD proteins after normalizing to the input reads (FDR <10%), indicating endogenous levels of BMP and TGF-β signaling in these cells (Figure 3A, Figure 3—figure supplement 1A–C). Following BMP4 addition, pSMAD1/5/8 exhibited approximately a 750-fold increase in peak number, while the number of SMAD3 peaks increased by less than threefold (Figure 3A, Figure 3—figure supplement 1A). Furthermore, in cells treated with BMP4, the pSMAD1/5/8 peaks largely overlap with the SMAD4 peaks, but much less with the SMAD3 peaks (Figure 3—figure supplement 1B). For example, pSMAD1/5/8 binds to two enhancers in the intronic region of Prdm16, with the binding intensity dramatically increased in response to BMP4 (Figure 3—figure supplement 1D). This suggests that Prdm16 itself is a transcriptional target of BMP signaling. Indeed, Prdm16 mRNA levels were elevated by BMP4 in wild-type but not Prdm16_KO cells (Figure 2—figure supplement 1B). By contrast, SMAD3 showed little ChIP-seq signal in the Prdm16 gene locus even in the presence of BMP4. These results confirm that the response of pSMAD1/5/8 is specific to BMP4, while SMAD3 is generally unresponsive to BMP4. The gained Smad3 sites likely result from an indirect effect of BMP signaling.
Figure 3 with 1 supplement see all
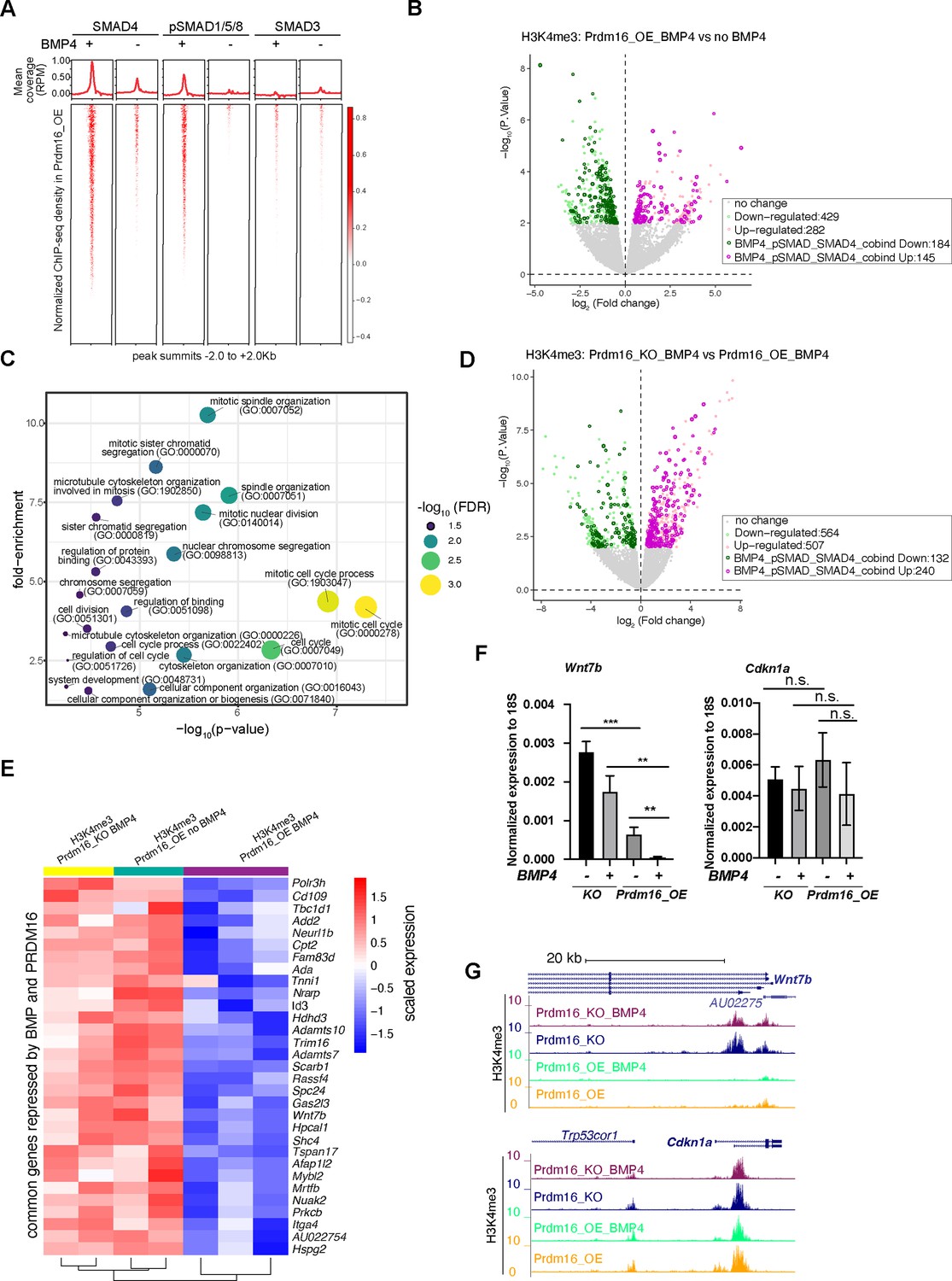
BMP signaling represses cell proliferation genes to induce cell quiescence.
(A) Heatmaps of ChIP-seq coverage (normalized by library depths and input) centered at peak summits for SMAD4, pSMAD1/5/8 and SMAD3 in Prdm16_OE cells with or without BMP4. (B) Volcano plot displaying differential H3K4me3 signal at TSS in Prdm16_OE cells with BMP4 versus untreated controls. Genes with significant changes (p<0.01) are colored. (C) Bubble plot of GO terms significantly enriched among SMAD-repressed genes (highlighted in dark green in panel B). (D) Volcano plot of differential H3K4me3 signal at TSS comparing Prdm16_KO and Prdm16_OE cells, both treated with BMP4. Significantly changed genes (p<0.01) are colored. (E) Heatmap of normalized H3K4me3 read counts at the transcription start sites (TSS) of 31 genes co-repressed by PRDM16 and SMADs (overlapping genes from dark green in B and dark pink in D). (F) RT-qPCR quantification of Wnt7b and Cdkn1a mRNA levels across indicated genotypes and treatment conditions. Data represent three biological replicates, each with two technical replicates. Statistical significance was calculated using unpaired t-test (***p<0.001; **p<0.01; n.s., non-significant). (G) Genome browser tracks of H3K4me3 CUT&TAG profiles at the Wnt7b and Cdkn1a loci.
-
Figure 3—source data 1
RTqPCR measurement of Wnt7b, Cdkn1a, and 18srRNA expression, related to Figure 3F.
- https://cdn.elifesciences.org/articles/104076/elife-104076-fig3-data1-v1.xlsx
To evaluate the regulatory activity of SMAD4/pSMAD1/5/8 in NSCs, we determined the state of transcription activation using H3K4me3 CUT&TAG signals at annotated gene transcription start sites (TSS) in BMP4-treated and untreated Prdm16_OE cells. As cell identity genes are known to be marked by extended breadth of H3K4me3 (Benayoun et al., 2014), we reasoned that compared to RNA-seq, which measures all gene products, changes in the H3K4me3 signal at the promoter might bias toward cell identity genes between proliferation and quiescence states. Thus, we determined all BMP4-repressed and activated genes by measuring changes in the H3K4me3 CUT&TAG read coverage at annotated TSS in BMP4-treated Prdm16_OE cells compared to untreated, identifying 282 up-regulated and 429 down-regulated genes (Figure 3B) (p<0.01). To pinpoint genes directly regulated by SMAD4 and pSMAD1/5/8, we intersected the dysregulated genes with those whose regulatory regions contain overlapping SMAD4 and pSMAD1/5/8 ChIP-seq peaks. This analysis led to the identification of 145 up-regulated and 184 down-regulated SMAD4 and pSMAD1/5/8 target genes.
To elucidate the function of BMP-regulated targets, we performed gene ontology (GO) analyses for the genes that changed expression in response to BMP4 in Prdm16_OE cells and were also bound by SMAD4 and pSMAD1/5/8. Remarkably, the downregulated genes (BMP4-repressed genes), but not the up-regulated genes (BMP4-activated genes), showed significantly enriched functional categories, with nearly all GO terms related to cell proliferation/cell cycle (Figure 3C). A recent study reported that BMP2-induced genes are enriched for neuronal and astrocyte differentiation (Katada et al., 2021), while our analysis did not identify these categories. One possibility for the discrepancy is that we overexpress Prdm16 in cultured NSCs, which may reinforce BMP signaling activities in cell proliferation. Other possibilities could be the use of different BMP ligands (BMP4 versus BMP2), differences in the origin of NSC culture (ours was from E13 instead of E11 and E14), or different profiling methods (H3K4me3 versus RNA-seq).
To identify PRDM16-repressed and activated genes under active BMP signaling conditions, we compared changes in H3K4me3 coverage at TSSs between BMP4-treated Prdm16_KO cells and BMP4-treated Prdm16_OE cells. We found approximately twice as many up-regulated genes (240) as down-regulated genes (132) (Figure 3D), suggesting that the cooperative activity of PRDM16 and BMP signaling mainly represses gene expression. We further overlapped the 184 genes repressed by BMP4 and the 240 genes repressed by PRDM16, identifying 31 common ones. These 31 genes displayed low H3K4me3 coverage in Prdm16_OE cells with BMP4 but higher H3K4me3 signal in both Prdm16_KO cells treated with BMP4 and untreated Prdm16_OE cells (Figure 3E).
Next, we attempted to validate whether changes in TSS H3K4me3 intensity correspond to changes in mRNA levels, by conducting RT-qPCR for selected genes whose function is associated with cell proliferation from the gene ontology analysis: Wnt7b, Mybl2, Id3, Spc24 and the three Spc24-related genes (Spc25, Ndc80 and Nuf2) (Figure 3F–G, Figure 3—figure supplement 1E–K). SPC24 forms the NDC80 kinetochore complex along with three other proteins: SPC25, NDC80, and NUF2 (Tooley and Stukenberg, 2011) (Illustrated in Figure 3—figure supplement 1H), and their function is essential for chromosome segregation and spindle checkpoint activity. Notably, Spc25, Ndc80, and Nuf2 appeared repressed by PRDM16 and BMP4 based on changes in H3K4me3 at the TSS (Figure 3—figure supplement 1E), despite not being identified as the top 31 candidates. mRNA levels for most of the tested genes followed a similar pattern to the H3K4me3 intensity: BMP4-treated Prdm16_OE cells showed the lowest expression, while either the loss of Prdm16 or absence of BMP4 led to upregulation. Spc24 gene products showed minimal amplification in qPCR, likely due to poor primer sequence quality or unstable mRNAs. Another exception is Id3 whose expression increased upon BMP4 treatment or Prdm16 depletion, indicating PRDM16 repressing Id3 but BMP4 inducing Id3. However, the activation of Id3 by BMP4 is significantly stronger in Prdm16_KO cells than that in Prdm16_OE cells, suggesting that BMP signaling normally activates Id3, but PRDM16 suppresses such activation. In contrast, Cdkn1a, a target gene of TGF-β pathway encoding the cell cycle inhibitor P21, exhibited consistent H3K4me3 coverage and mRNA levels across BMP4-treated and non-treated cells, as well as between Prdm16_OE and KO cells (Figure 3F–G), confirming that neither PRDM16 nor BMP signaling influences Cdkn1a expression in NSCs.
PRDM16 assists genomic binding of SMAD4 and pSMAD1/5/8
To understand how PRDM16 interacts with BMP signaling, we integrated PRDM16 CUT&TAG data with SMAD ChIP-seq data, focusing on assessing PRDM16’s influence on the genomic binding of SMAD proteins.
The SEACR peak caller software (Meers et al., 2019) identified 3337 and 7639 PRDM16 CUT&TAG common peaks from four replicates of BMP4-treated and untreated Prdm16_OE samples (FDR <10%, see Methods), respectively (Figure 4A). Upon BMP4 treatment, there were 5936 lost and 1634 gained PRDM16 CUT&TAG peaks, suggesting that PRDM16 regulates a distinct subset of genes in proliferating versus quiescent NSCs. Samples from Prdm16_KO cells lacked these sites, confirming the specificity of the PRDM16 CUT&TAG signal (Figure 4B).
Figure 4 with 1 supplement see all
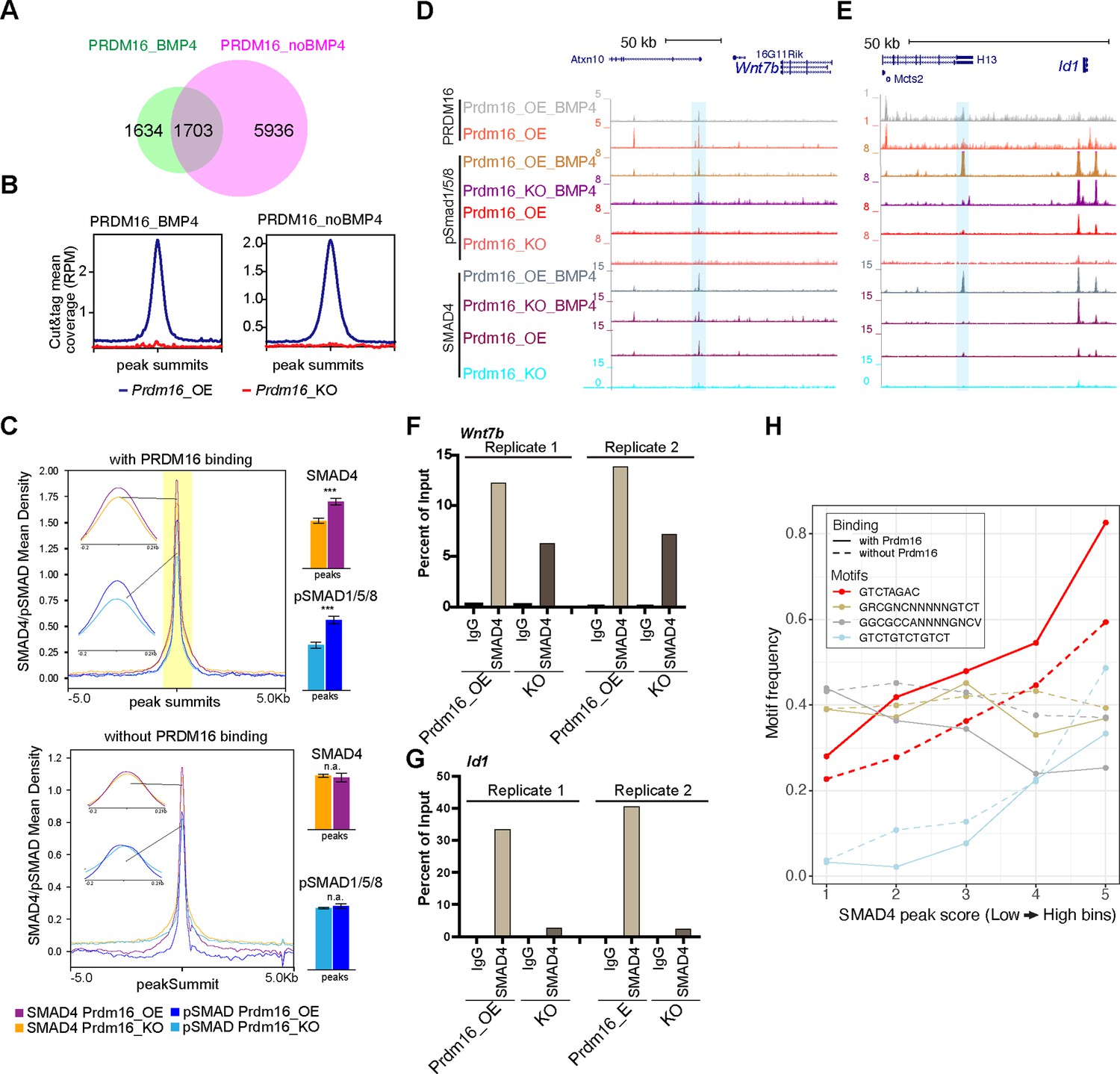
PRDM16 anchors SMAD proteins at specific genomic regions to mediate gene repression.
(A) Venn diagram showing overlap of PRDM16 CUT&TAG peak in BMP4-treated and untreated Prdm16_OE cells. (B) Average PRDM16 CUT&Tag signal (normalized to library size) at peak summits in BMP4-treated, untreated Prdm16_OE, and Prdm16_KO cells (4 replicates combined). (C) Metaplots of SMAD4 and pSMAD1/5/8 ChIP-seq coverage centered on peak summits with (top) and without (bottom) PRDM16 co-binding in BMP4-treated Prdm16_OE and Prdm16_KO cells. Bar plots (right) show average coverage; significant reduction in the KO cells is seen only at PRDM16 co-bound sites (***p<0.001; **p<0.01; *p<0.05; n.a. p>0.05). (D–E) Genome browser view of Wnt7b and Id1 loci showing PRDM16 CUT&TAG and SMAD ChIP-seq tracks. Co-bound regions are highlighted. (F–G) ChIP-qPCR validation of SMAD4 binding at highlighted regions in D-E from Prdm16_OE and Prdm16_KO cells (2 replicates). (H) Frequency of four SMAD motif types across SMAD ChIP-seq peaks binned by binding strength. The palindromic motif GTCTAGAC is most enriched at PRDM16 co-bound sites (solid red) and notably reduced in non-co-bound peaks (dashed red).
-
Figure 4—source data 1
ChIP-qPCR measurement of SMAD4 and PRDM16 co-bound site in Wnt7b, related to Figure 4F.
- https://cdn.elifesciences.org/articles/104076/elife-104076-fig4-data1-v1.xlsx
-
Figure 4—source data 2
ChIP-qPCR measurement of SMAD4 and PRDM16 co-bound site in Id1, related to Figure 4G.
- https://cdn.elifesciences.org/articles/104076/elife-104076-fig4-data2-v1.xlsx
From our overlapping analyses (see Methods), we found that over 50% of SMAD4 and pSMAD1/5/8 binding peaks were consistent in Prdm16_OE and Prdm16_KO cells (Figure 4—figure supplement 1A–B), indicating that deletion of Prdm16 does not affect general genomic binding ability of these proteins. Using Homer’s mergePeaks function with PRDM16 CUT&TAG and SMAD ChIP-seq data, we identified co-bound sites by PRDM16 and SMAD proteins in BMP4-treated Prdm16_OE cells. PRDM16 CUT&TAG peaks mainly overlap with SMAD4 and pSMAD1/5/8 peaks, but not much with SMAD3 peaks (Figure 4—figure supplement 1A–C). This result suggests that PRDM16 mainly collaborates with the SMAD4/pSMAD1/5/8 complex but not the SMAD3/SMAD4 complex in cells with high levels of BMP4.
Further examination of SMAD4 and pSMAD1/5/8 binding revealed significantly lower enrichment of SMAD4 and pSMAD1/5/8 at PRDM16 co-bound sites in Prdm16_KO cells compared to Prdm16_OE cells (Figure 4C) (p=2.5E-6, and p=4.7E-3, respectively, two-tailed t-test). As a control, the SMAD4 and pSMAD1/5/8 sites without PRDM16 co-binding did not show such a change (p>0.05). This result suggests that SMAD binding to the PRDM16 co-binding sites depends on PRDM16.
To validate the co-binding of SMAD4 and PRDM16, we selected the candidate loci, Wnt7b and Id1 (Figure 4D–E) and applied sequential ChIP-qPCR. This experiment confirmed simultaneous binding of SMAD4 and PRDM16 to the same DNA molecules at these loci (Figure 4—figure supplement 1D), as SMAD4 was more enriched in the chromatin pulled by the PRDM16 antibody compared to the IgG control. Additionally, ChIP-qPCR confirmed reduced SMAD binding at the SMAD/PRDM16 co-bound site in Prdm16_KO cells compared to Prdm16_OE cells (Figure 4F–G). Thus, PRDM16 enhances genomic binding of the SMAD proteins to specific genome regions.
PRDM16 facilitates SMAD4 binding to regions enriched for SMAD palindromic motifs
We then wondered whether there is a sequence feature that distinguishes the regions co-bound by SMADs and PRDM16 from those only bound by SMADs. To this end, we checked SMAD-bound regions with PRDM16 binding and those without for two types of SMAD4 binding motifs (Hill, 2016). Together with pSMAD1/5/8 or pSMAD3, SMAD4 may bind to a palindromic motif, GTCTAGAC or direct repeats of GTCT like GTCTGTCTGTCT (Zawel et al., 1998; Dennler et al., 1998; Wong et al., 1999); together with pSMAD1/5/8, SMAD4 associates with GC-rich SBEs (SMAD-binding elements) including GGCGCC-AN4-GNCV and GRCGNCNNNNNGTCT (Pyrowolakis et al., 2004; Gao et al., 2005; Weiss et al., 2010). We calculated the occurrence frequency of each of these motifs in binned SMAD4 ChIP-seq peaks from Prdm16_OE cells treated with BMP4 (bin 1–5 with low to high peak scores, Figure 4H). Interestingly, the palindromic motif is the most enriched one. Its occurrence frequency increases with peak scores but becomes lower in the SMAD4 peaks absent for PRDM16 co-binding. The frequency of the GTCT-triplet motif also increases with higher SMAD4 peak scores, while there is no reduction in regions without PRDM16 binding. The two GC-rich motifs show a distinct trend: they are not as highly enriched, their frequencies do not increase with higher SMAD4 peak scores, and the occurrence frequency in PRDM16 co-bound regions is either comparable to or even lower than the regions without PRDM16 binding. Together, this result suggests that PRDM16 may separate the SMAD4/pSMAD1/5/8 proteins into two types of genomic regions, one enriched for the palindromic motif where PRDM16 is present and the other with the GC-rich motifs.
SMAD4 and pSMAD1/5/8 switch genomic locations in the absence of PRDM16
Our careful inspection of the Wnt7b and Id1 loci surprisingly revealed that there are multiple ectopic SMAD4 and pSMAD1/5/8 peaks in the Prdm16 mutant sample (arrow-indicated peaks in Figure 5A–B). We then globally assessed the extent of ectopic SMAD binding surrounding the SMAD/PRDM16 co-bound sites by quantifying differential SMAD binding intensity between Prdm16_KO and Prdm16_OE cells (Figure 5C, FDR <10%). In agreement with the metaplots (Figure 4C), all of the SMAD/PRDM16 co-bound sites (blue dots) showed reduced ChIP-seq read coverage in Prdm16_KO cells. By contrast, the flanking regions within the 200 kb range of a co-bound site (red dots) displayed a trend of increase in SMAD binding in Prdm16_KO cells. This result agrees with the aforementioned motif analysis that PRDM16 helps to localize the SMAD complex at specific genomic regions, and it also suggests that without Prdm16, the SMAD factors are partly redirected to neighboring genomic regions.
Figure 5
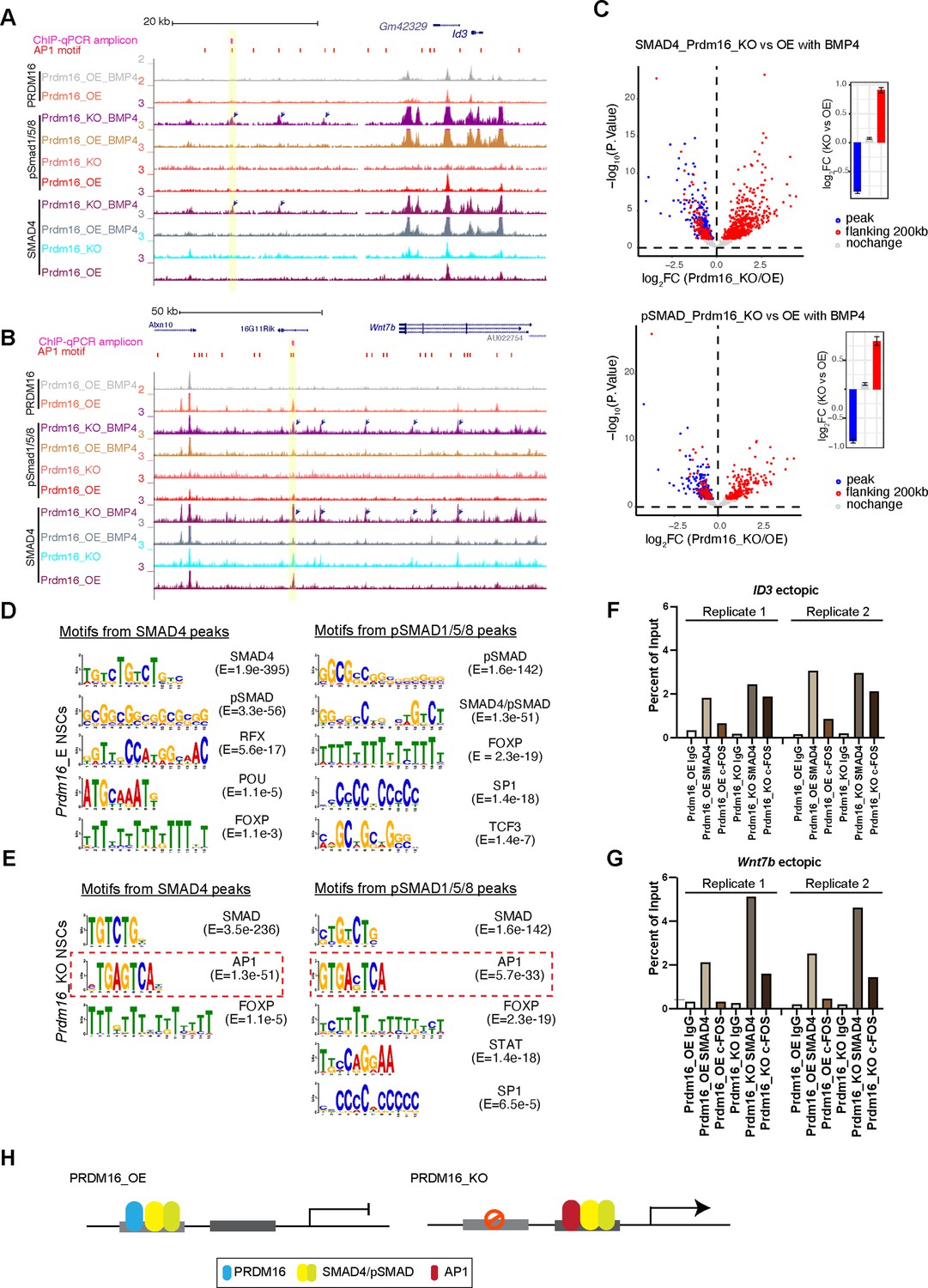
The SMAD complex shifts genome binding in the absence of Prdm16.
(A–B) Genome browser views of the Id3 (top) and Wnt7b (bottom) loci showing ectopic SMAD peaks (arrows) in Prdm16 KO cells. (C) Volcano plots showing differential ChIP-seq signal for SMAD4 and pSMAD1/5/8 in BMP4-treated Prdm16_KO cells versus BMP4-treated Prdm16_OE cells. Blue dots represent sites co-bound by PRDM16; red dots indicate sites not bound by PRDM16 but located within 200 kb of a PRDM16-cobound region. Mean log2 fold-change values are plotted to the right. (D–E) De novo motif discovery from SMAD4 and pSMAD1/5/8 ChIP-seq peaks in Prdm16_OE cells (D) and Prdm16_KO cells (E). (F–G) ChIP-qPCR validation of increased SMAD4 and c-FOS occupancy at the indicated ectopic binding sites in Prdm16_KO versus Prdm16_OE cells. Locations of PCR amplicons are shown in A and B. (H) Model illustrating enhancer switching: in the absence of PRDM16, SMAD complexes alter their co-factors and function as transcriptional activators.
-
Figure 5—source data 1
ChIP-qPCR measurement of ectopic SMAD4 and AP1 site in Id3, related to Figure 5F.
- https://cdn.elifesciences.org/articles/104076/elife-104076-fig5-data1-v1.xlsx
-
Figure 5—source data 2
ChIP-qPCR measurement of ectopic SMAD4 and AP1 site in Wnt7b, related to Figure 5G.
- https://cdn.elifesciences.org/articles/104076/elife-104076-fig5-data2-v1.xlsx
AP1 is the potential co-factor that interacts with relocated SMAD proteins
We intended to find out how SMAD4 and pSMAD1/5/8 relocate to ectopic sites in the absence of Prdm16. By running de novo motif discovery (see Methods), we identified a number of significantly enriched DNA motifs from pSMAD1/5/8 and SMAD4 ChIP-seq peaks in Prdm16_OE and Prdm16_KO cells (Figure 5D–E). Interestingly, in addition to the known SMAD motifs, the only other motif identified in both pSMAD1/5/8 and SMAD4 peaks from Prdm16_KO cells but not Prdm16_OE cells is the AP1 motif. Furthermore, the AP1 motif is more frequently found in the pSMAD1/5/8 and SMAD4 regions lacking PRDM16 binding than those with PRDM16 binding (Figure 4—figure supplement 1E). The interaction between the AP-1 complex and the SMAD proteins was reported previously (Zhang et al., 1998). Our result implies that AP-1 is a potential cofactor for the SMAD proteins in the absence of Prdm16. To validate this finding, we performed ChIP-qPCR with an antibody against c-FOS, one of the subunits of AP1, on Prdm16_OE and KO cells. Supporting the global analyses, c-FOS is enriched in the Smad4-bound regions at the Wnt7b and Id3 loci in the Prdm16_KO condition (Figure 5F–G). There is little c-FOS binding to these regions in cells expressing Prdm16, suggesting that there is cooperativity of SMAD and AP-1 which facilitates each other’s binding when PRDM16 is absent. Alternatively, PRDM16 may suppress genomic accessibility of these regions for SMAD and AP-1 proteins, a function we reported for PRDM16 in cortical NSCs (He et al., 2021).
Together, our genomic data and validation suggest an enhancer switch model (Figure 5H): PRDM16 assists SMAD protein binding to repressive cis-regulatory elements that are enriched for GTCTAGAC palindromic motifs; loss of PRDM16 results in genomic relocation of SMAD proteins, presumably via the association with coactivators such as AP1; the consequence of SMAD relocation is de-repression of cell proliferation regulators.
SMADs and PRDM16 co-repressed genes in NSCs are suppressed in the developing ChP
Next, we investigated whether genes co-regulated by PRDM16 and BMP signaling in NSCs (Figure 3C) are also involved in ChP development, where both factors limit NSC proliferation (Hébert et al., 2002 and Figure 1). Using published E12.5 mouse brain single cell RNA-seq (scRNA-seq) data (La Manno et al., 2021), we analyzed the expression of the 31 co-repressed genes across ChP epithelial (Ttr+), CH (BMP4+/ Ttr-) and a few forebrain-specific radial glia (RG) (Sox2+/Hes5+) clusters (Figure 6A, Figure 6—figure supplement 1A). Except for one gene Mrtfb that was not found in the scRNA-seq data, thirteen showed no or weak expression (Figure 6—figure supplement 1A), and seventeen had moderate to high expression in at least one cluster (Figure 6A). Notably, these genes were generally expressed at lower levels in ChP clusters than in CH or RG clusters, suggesting repression in the ChP.
Figure 6 with 1 supplement see all
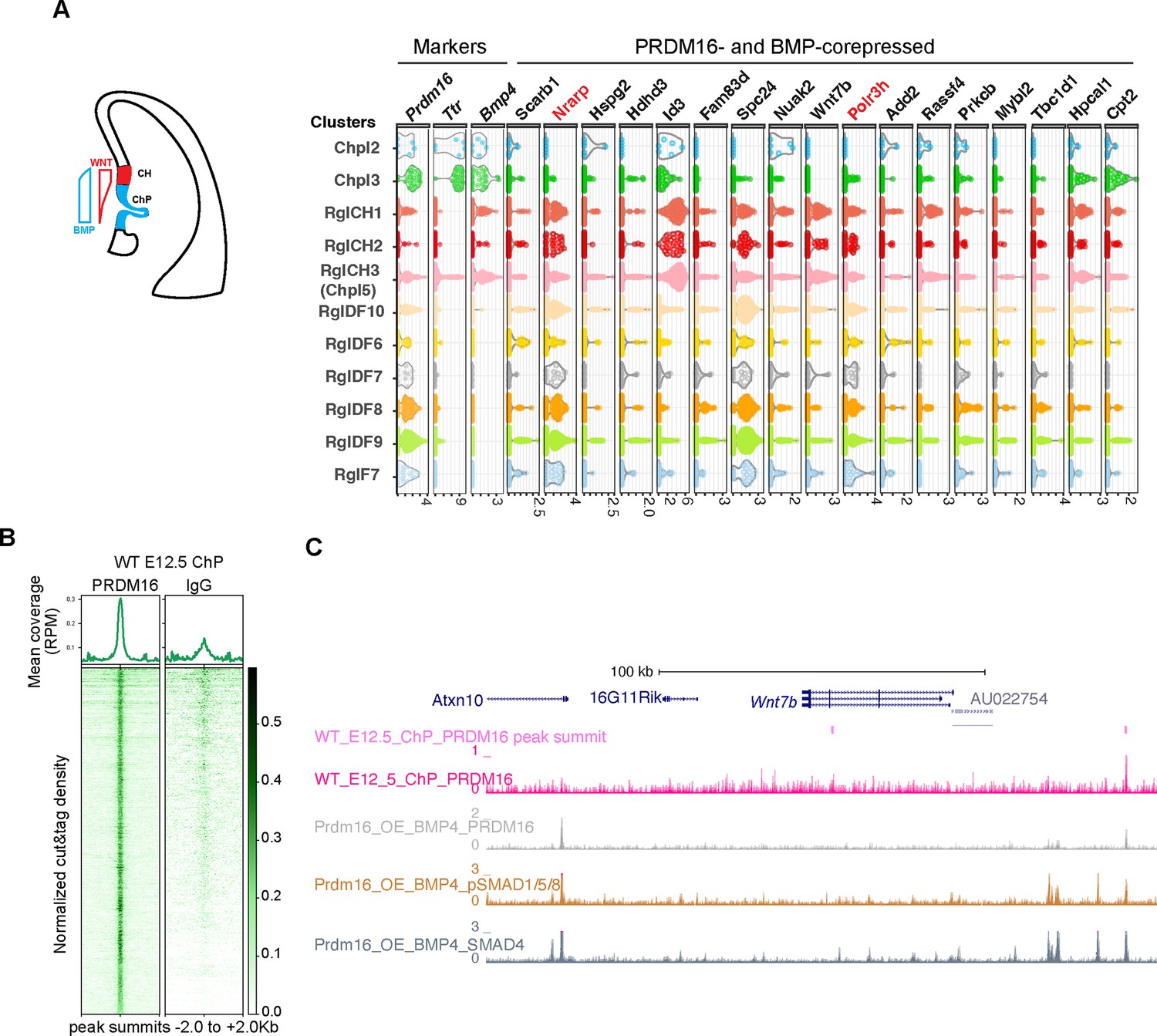
PRDM16 and SMADs co-regulated genes exhibit differential expression in choroid plexus (ChP) and neural cells.
(A) Schematic illustrating the cortical hem (CH) and ChP regions, which exhibit opposing Wnt and BMP signaling gradients. Shown alongside is a violin plot of scRNA-seq expression at E12.5 for the indicated genes across cell type clusters (Pyrowolakis et al., 2004). Cell clusters were defined based on marker gene expression from E9.5-E18.5 brain tissues, including choroid plexus clusters 1–5 (Chpl), with Chpl5 newly identified in this study based on Ttr expression as well as radial glia clusters: RglCH 1–3, RglDF 6–10, and RglF7. CH: cortical hem; DF: dorsal forebrain; F: forebrain. Gene expression per cell is represented as log2-normalized counts. (B) Metaplots and heatmaps showing CUT&TAG signal for PRDM16 and IgG, centered on peak summits and normalized for library size, from E12.5 dorsal midline tissues. (C) Genome browser view of the Wnt7b locus, displaying CUT&TAG tracks for PRDM16 and ChIP-seq tracks for SMAD proteins.
To test for direct PRDM16 regulation, we performed CUT&TAG assays using the PRDM16 antibody on dissected dorsal midline tissues that mainly contain CH and the ChP (Figure 6B and Figure 6—figure supplement 1B) and identified 3238 common peaks across three replicates. Manual inspection of the 31 loci revealed PRDM16 binding near the TSS of 25 genes using the browser Embryonic Mouse Brain Epigenome Atlas (Rhodes et al., 2022) (Text in black in Figure 6A, Figure 6—figure supplement 1A). Using an unbiased peak-to-promoter mapping, 24/31 co-repressed and 84/153 BMP-only-repressed genes had PRDM16 binding in E12.5 ChP. The enrichment in the co-repressed group (Figure 6—figure supplement 1C Fisher’s Exact Test, P=0.015) indicates a stronger regulatory role of PRDM16 on these genes in the developing ChP.
As an example, Wnt7b has two PRDM16 peaks (Figure 6C), one unique to the dorsal midline tissue. Given Wnt signaling’s role in ChP specification and NSC proliferation (Parichha et al., 2022; Kalani et al., 2008). Wnt7b likely represents a direct PRDM16 target in the developing ChP.
PRDM16 represses Wnt7b and Wnt activity in the developing ChP
We sought to determine which of the identified genes from NSC culture are regulated by PRDM16 in the developing ChP. In addition to Wnt7b, several other Wnt ligands are expressed in the CH and ChP region (Harrison-Uy and Pleasure, 2012). For example, Wnt3a is expressed in the dorsal-midline region but not in the forebrain, which explains why it was not identified from the forebrain-derived NSC datasets. Although there is no PRDM16 CUT&TAG peak within the Wnt3a locus in cultured NSCs, a PRDM16 CUT&TAG peak is present in the intronic region of Wnt3a in the dorsal midline tissue (Figure 6—figure supplement 1D). We thus examined the scRNA-seq data (La Manno et al., 2021) and confirmed that multiple components in the Wnt and BMP pathways are present in the CH and ChP clusters (Figure 6—figure supplement 1E). To systematically measure expression changes of PRDM16 target genes in the Prdm16 mutant brain, we applied a multiplexed fluorescent in situ approach, Single-Cell Resolution in Situ Hybridization On Tissues (SCRINSHOT) (Sountoulidis et al., 2020). In this method, each mRNA molecule is hybridized with three gene-specific padlock probes and visualized by fluorescent dye-conjugated detection probes. A dot of fluorescent signal corresponds to one mRNA molecule, allowing quantitative measurement of gene expression in single-cell resolution.
We designed probes for ten Wnt and five BMP pathway components, other genes that are co-repressed by BMP and PRDM16 in NSC culture, as well as cell-type-specific markers (the ChP marker gene Ttr and Foxj1, the neural progenitor markers Sox2, Hes5, and Zbtb20, and the neuronal marker Ngn2), and conducted SCRINSHOT in three pairs of control and Prdm16 mutant brains.
In the control brain, the Ttr and Foxj1 probes detect the ChP epithelium but not the adjacent tissues at E11.5 and E12.5, confirming the signal specificity of SCRINSHOT. Several Wnt (Wnt2b, Wnt7b, Wnt5a, Wnt3a, Fzd10, and Axin2) and BMP components (Bmp4 and Bmp7) also display tissue specificity: the Wnt genes are more enriched at CH, while the BMP genes are more at the ChP. The representative images from E11.5 are presented in Figure 7A and B and Figure 7—figure supplement 1A. Except for Wnt2b, the other tested Wnt genes exhibit low expression in the ChP cells, consistent with the notion that Wnt signaling is present and required at the ChP (Parichha et al., 2022).
Figure 7 with 4 supplements see all
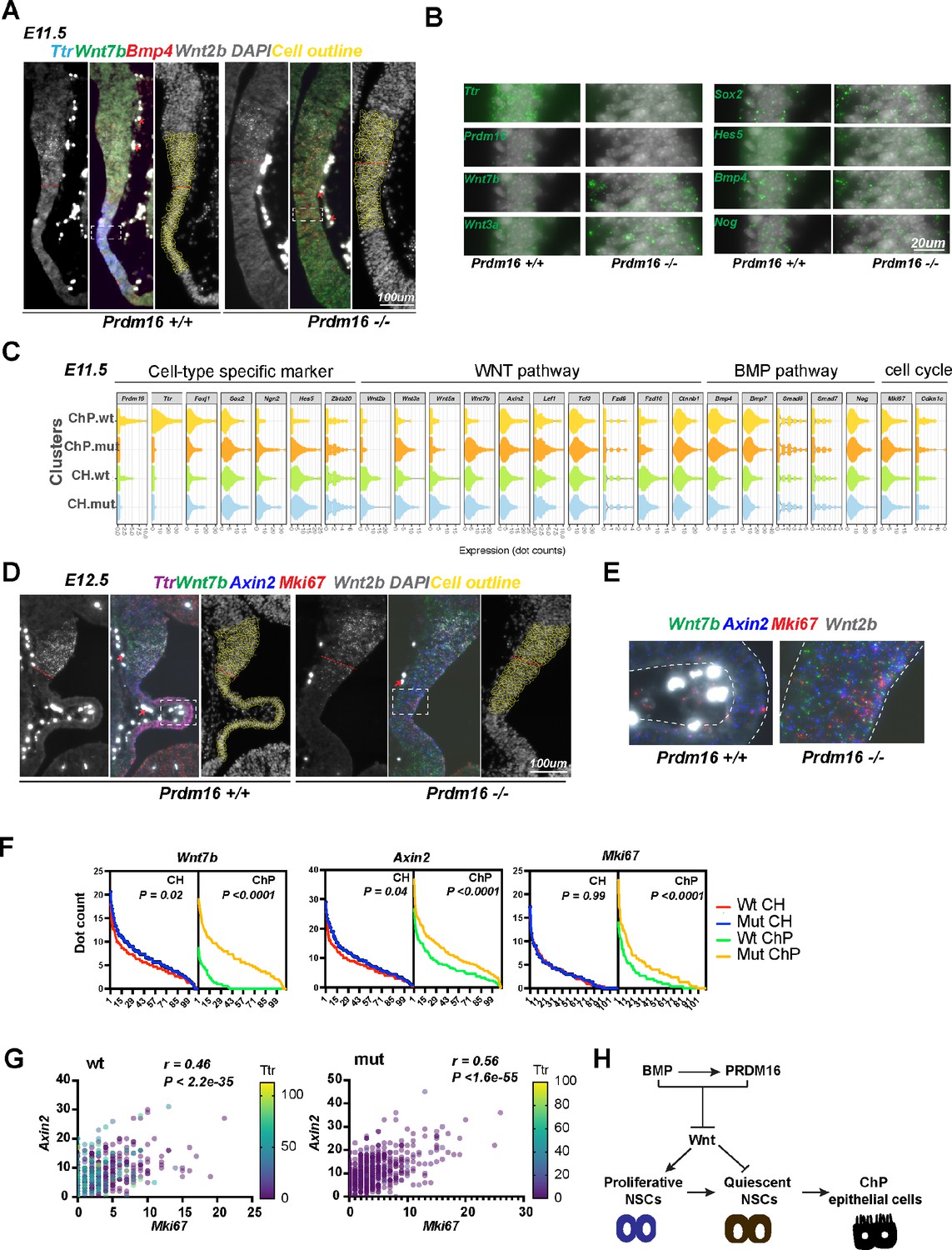
PRDM16 suppresses Wnt signaling and cell proliferation in choroid plexus (ChP) epithelial cells.
(A) SCRINSHOT images of E11.5 wild-type and Prdm16 mutant brains probed for Bmp4, Ttr, Wnt7b and Wnt2b. Red arrows indicate autofluorescent blood cells (non-specific signal in all channels). Cells quantified are circled in yellow, and the red dashed line marks the cortical hem (CH) and ChP boundary. (B) Enlarged views of the boxed region in (A), showing increased expression of Wnt and neural genes and decreased Ttr and Prdm16 in mutants. (C) Violin plots displaying dot counts for the 24 genes in 330 CH and 330 ChP cells from three wild-type and three Prdm16 mutant samples. (D) SCRINSHOT images of E12.5 wild-type and Prdm16 mutant brains with probes for Axin2, Ttr, Wnt7b, Wnt2b, and Mki67. (E) Enlarged views of the boxed areas in (D), with ChP regions outlined by dashed lines. (F) Distribution of dot counts for indicated genes in wild-type (wt) and Prdm16 mutant (mut) CH and ChP cells. X-axis: 110 cells per replicate; Y-axis: mean dot count across three biological replicates. P values calculated using area under the ROC curve (Prism Graphpad). (G) Multi-variant linear regression of Mki67, Axin2 and Ttr expression across 660 CH and ChP cells per genotype. Pearson correlation and significance were calculated using Prism Graphpad. (H) Model illustrating the regulatory circuit through which PRDM16 promotes the transition from proliferative to quiescent NSCs, a prerequisite step for ChP epithelial cell specification.
-
Figure 7—source data 1
Dot counts in three pairs of wt and Prdm16 mutant CH (1-330) and ChP (331-660) cells from E11.5 brains, related to Figure 7C.
- https://cdn.elifesciences.org/articles/104076/elife-104076-fig7-data1-v1.xlsx
Next, we quantified expression changes of these genes in Prdm16 mutant CH and ChP cells. As expression of Wnt2b was relatively unchanged in the mutant (Figure 1—figure supplement 1C, Figure 7A), we used Wnt2b-expressing cells to define the CH cells and those expressing Ttr to define ChP cells in the wild-type brain slice. There were about 110 cells in each region of one brain slice. Because the mutant brain slice showed little Ttr signal, we only used the Wnt2b-expressing cells to define the anterior border between CH and neocortex and included 110 cells under the border for the CH cells, and the further 110 cells for the mutant ChP cells, as shown in the images with DAPI and cell outlines (Figure 7A, Figure 7—figure supplement 1B and Figure 7D). We then measured the dot count of each gene in these 220 cells in each sample, and summarized the changes from three pairs of animals in the violin plots (Figure 7C and Figure 7—figure supplement 2A). Expression of Ttr and Foxj1 is severely reduced, while Sox2, Hes5, and Ngn2 all become upregulated in the mutant ChP, indicating a fate transformation of ChP epithelial to neural cells. This agrees with our immunostaining and conventional in situ results (Figure 1 and Figure 1—figure supplement 1). Similar to Bmp4, other BMP components we tested including Bmp7, Smad6, Smad7 and Nog are unchanged, suggesting that PRDM16 is not an upstream regulator of the BMP pathway.
Moreover, Wnt3a and Wnt7b are significantly upregulated in the Prdm16 mutant ChP (Figure 7B-E, Figure 7—figure supplement 2A-C). In line with this, the Wnt target gene Axin2 is also upregulated, indicating aberrantly elevated Wnt signaling in the Prdm16 mutant ChP. Thus, the normal function of Prdm16 represses WNT signaling in the developing ChP.
Levels of Wnt activity correlate with cell proliferation in the developing ChP and CH
We further assessed the relationship between Wnt signaling and cell proliferation in the ChP and CH at E12.5, by correlating expression levels of Mki67 with Wnt3a, Wnt7b, and Axin2 levels in wild-type and mutant samples. A significant increase in Mki67 expression in Prdm16 mutant ChP cells at E12.5 is accompanied by significantly increased Wnt gene expression (Figure 7F and Figure 7—figure supplement 2B–C). In contrast, little or no increase in Mki67 signal is found in the mutant CH, suggesting that the effect of PRDM16 is mainly cell-autonomous. We then performed linear regression and Pearson correlation analyses to assess correlations between ChP markers, BMP, and Wnt genes. Interestingly, the level of Axin2 best correlates with the level of Mki67, even better than that with other Wnt genes in both wild-type (r=0.46; p<2.2 E-35) and mutant (r=0.56; p<1.6 E-55) CH and ChP cells (Figure 7G and Figure 7—figure supplement 2D). This result suggests that Wnt activity may be responsible for cell proliferation in the CH and ChP regions. Supporting this finding, it was shown that ectopic Wnt signaling converts ChP cells into proliferative CH neural cells (Parichha et al., 2022). Similarly, in the fourth ventricle, two Wnt genes, Wnt1 and Wnt3a, and Mki67 all became ectopically expressed in Prdm16 mutant ChP cells (Figure 7—figure supplement 3). Thus, PRDM16 also suppresses Wnt signaling in the hindbrain ChP.
Additionally, six cell-proliferation-related genes (Spc24, Spc25, Nuf2, Ndc80, Id3, and Mybl2) exhibited upregulation in Prdm16 mutant ChP (Figure 7—figure supplement 4). Interestingly, all four NDC80 complex genes became upregulated, pointing to its potential role in promoting NSC proliferation. Taken together, our results suggest that specification of the ChP epithelium requires a process transitioning proliferating NSCs into quiescence, and that this process is mediated by a suppressive role of PRDM16 on Wnt signaling and cell cycle regulators.
Discussion
In this study, we reveal a novel molecular mechanism by which BMP signaling regulates cell proliferation and gene expression. We find that PRDM16 acts as a tethering factor to localize the SMAD4/pSMAD1/5/8 complex at specific genomic sites and facilitate the repressive role of the SMAD proteins. The genes co-repressed by PRDM16 and BMP signaling include those encoding Wnt pathway ligands and other cell proliferation regulators.
Combinatory activities of morphogens are exploited throughout animal development and tissue homeostasis. The crosstalk between BMP and WNT signaling is complex as it can be synergistic or antagonistic (Itasaki and Hoppler, 2010). Surprisingly, both types of effects exist in the specification of ChP epithelium. Here, BMP signaling induces NSC quiescence, as evidenced by the presence of ectopic proliferating cells at the ChP in Bmpr1a mutant mice (Hébert et al., 2002), while Wnt signaling promotes proliferation since the gain-of-function condition of beta-catenin phenocopies Bmpr1a mutant animals (Parichha et al., 2022). On the other hand, adding a low dose of Wnt activator to cell culture medium enhances programing efficiency of ChP epithelial cells induced by BMP4 (Pellegrini et al., 2020), and loss-of-function of beta-catenin resulted in under-developed ChP structure (Parichha et al., 2022), suggesting that BMP and WNT signaling collaborate to promote ChP epithelial cell differentiation. Our work demonstrated that PRDM16 ensures the right balance of BMP and Wnt activity by maintaining a low level of Wnt gene expression in the developing ChP.
PRDM16 was shown to physically interact with SMAD3 (Warner et al., 2007) and antagonize TGF-β-induced cell cycle arrest (Takahata et al., 2009). However, we did not detect stable PRDM16/SMAD4/pSMAD1/5/8 or PRDM16/SMAD4/SMAD3 complexes in co-immunoprecipitation assays (data not shown). Instead, sequential ChIP revealed that PRDM16 and SMAD4 bind to the same DNA molecule, supporting their cooperative function at co-occupied regulatory elements. Notably, SMADs when associated with PRDM16 compared to those without PRDM16, with SMADs preferentially bind palindromic motifs in the presence of PRDM16 and GC-rich SBE motifs in its absence, suggesting that PRDM16 modulates SMAD DNA-binding specificity and shifts SMAD regulatory output.
A recent study similarly reported that PRDM16 acts as a SMAD4 co-repressor during pancreatic ductal adenocarcinoma progression (Hurwitz et al., 2023). There, SMAD4 stably binds the PRDM16 promoter and switches its co-factor from the co-repressor PRDM16 (under low TGF-β) to the co-activator SMAD3 (under high TGF-β), depending on TGF-β levels. This mechanism differs from our model. Our data indicate that high BMP4 levels promote SMAD4 and pSMAD1/5/8 binding to target loci, including Prdm16, resulting in both gene repression and activation. Notably, PRDM16 contributes to the repression function of the SMAD complex even under elevated BMP conditions.
Although PRDM16 has been shown to promote proliferation by antagonizing the anti-proliferation activity of TGF-β and SMAD3 (Takahata et al., 2009), our data demonstrate that PRDM16 collaborates with SMAD4 and pSMAD1/5/8 to repress proliferation genes, which is consistent with BMP-induced quiescence. Similar roles have been observed in other contexts, such as osteoblast differentiation and maturation (Wu et al., 2016). Supporting this, genomic profiling revealed that under high BMP4 and PRDM16 conditions, pSMAD1/5/8 and SMAD4 occupy numerous loci without SMAD3, suggesting PRDM16 primarily modulates BMP target genes in these settings.
Moreover, while it was reported that PRDM16 enhances Wnt signaling by stabilizing nuclear beta-Catenin in the craniofacial tissue (Shull et al., 2022), we detected the opposite outcome in the ChP and cultured NSCs, ectopic WNT gene expression in the absence of Prdm16, suggesting that PRDM16 represses Wnt genes in these contexts. Notably, PRDM16, Wnt, and BMP co-exist in various developmental settings, such as craniofacial development (Bjork et al., 2010; Shull et al., 2022; Nie et al., 2006), heart formation (Arndt et al., 2013; van Wijk et al., 2007; Gessert and Kühl, 2010), limb patterning (Bjork et al., 2010; Geetha-Loganathan et al., 2008; Robert, 2007), and adult intestinal stem cells (Stine et al., 2019; Korinek et al., 1998; He et al., 2004). We speculate a similar regulatory circuit is used in some, if not all, of these settings.
ChP epithelial cells derive from proliferating NSCs, without intermediate fate-commitment steps. This property endows ChP cells with higher plasticity and makes them more vulnerable to abnormal genetic and cellular change, to which high occurrence frequency of ChP tumors in fetuses and young children may be attributed (Wolff et al., 2002). As an essential brain structure, the ChP releases cerebrospinal fluid (CSF) and acts as a brain-blood barrier (Johansson, 2014; Lehtinen et al., 2013; Lun et al., 2015). Its dysfunction has been linked with several types of human diseases including hydrocephalus, Alzheimer’s disease, and multiple sclerosis (Lun et al., 2015). Deepening our knowledge on the development of ChP will provide new insights into potential therapeutics to prevent or treat ChP tumors and other ChP-related diseases.
Footnote: a study on the antagonism between PRDM16 and SMAD4 was published on Feb 24, 2023 (Hurwitz, 2023) after we posted our manuscript at BioRxiv and during our submission process. Materials and methods.
Materials and methods
Animals
All animal procedures were approved by the Swedish Agriculture Board (Jordbruksverket) with document numbers Dnr 11553–2017 and 11766–2022. The Prdm16cGT mice (Strassman et al., 2017) were maintained by outcrossing with the FVB/NJ line.
In situ hybridization
Request a detailed protocolTo make probes for conventional RNA in situ hybridization, genomic regions covering one exon or full-length cDNA were PCR amplified to generate fragments with restriction enzyme overhangs. The sequences of all oligos were included in the Key Resources Table. The fragments were inserted into the pBluescript SK(II) vector. In vitro transcription was performed as previously described (He et al., 2021). The mouse brains at defined ages were dissected and fixed for 12 hr in 4% PFA, dehydrated in 25% sucrose, cryoprotected, and embedded in O.C.T. The brain samples were then sectioned at 18 μm thickness on Leica cryostats CM3050s. RNA in situ hybridization was performed using digoxigenin-labeled riboprobes as described previously. Detailed protocols are available upon request. Images were taken using a Leica DMLB microscope.
Immunostaining
Request a detailed protocolImmunostaining was performed according to standard protocols as previously used (Dai et al., 2013a). For EdU and BrdU labeling, EdU (5-ethynyl-2′-deoxyuridine) and BrdU (5-bromo-2'-deoxyuridine) (5–20 μg/g of body weight) were injected into the peritoneal cavity of pregnant mice at desired ages. EdU incorporation was detected with the Click-iT assay (Invitrogen) according to the manufacturer’s instructions. BrdU incorporation was measured by immunostaining using an antibody against rat-BrdU (Abcam). Imaging was taken on a Zeiss confocal microscope. ZEN (ZeissLSM800), ImageJ (NIH), and Photoshop (Adobe) were used for analysis and quantification.
NSC culture
Request a detailed protocolControl and mutant embryonic cortices from E13.5 animals were dissected and dissociated into single-cell suspension and digested with Accutase (Sigma). Cells were maintained in proliferation media (STEMCELL Technologies). Lentivirus expressing Flag-Prdm16 and a puromycin-resistant gene in the pCDH vector was produced in 293t cells (ATCC) and used to infect a wild-type NSC line derived from E13.5 forebrain, and the selected NSCs were maintained in puromycin-containing medium for three passages before puromycin withdrawal. In the quality control specifications, mycoplasma was not detected.
RT-qPCR
Request a detailed protocolPrdm16_OE, wild-type, and mutant NSCs were cultured and treated with or without BMP4 (Sigma). After 48 hr treatment, total RNAs were extracted using TRIzol reagent (Invitrogen). Total RNA was further cleaned with Turbo DNase (Ambion) and used in reverse transcription with RT master mix (Thermo Fisher). To ensure the absence of genomic DNA, control qPCR was performed on the mock-reverse-transcribed RNA samples. The list of qPCR primers is included in the Key resources table.
SCRINSHOT
Request a detailed protocolBrain sections were prepared in the same way as for regular in situ hybridization (described above). The SCRINSHOT experiments were carried out according to the published method (Sountoulidis et al., 2020). In brief, 3 padlock probes and 3 corresponding detection probes were designed for each gene of interest. The slides with cryo-sectioned brain slices were pretreated at 45 °C to reduce moisture and fixed in 4% PFA in 1 X PBS, followed by washing in PBS Tween-20 0.05% twice. Permeabilization of tissues was done by washing slides in 0.1 M HCl for 2 mins 15 s, followed by washing in PBS Tween-20 0.05% twice. Then a stepwise dehydration was performed for the slides in 70%, 85%, 100% Ethanol and air. The SecureSeal hybridization chamber (GRACE BIO-LABS, 621501) was then mounted to cover each pair of control and Prdm16 mutant brain slices. Samples were then blocked in a probe-free hybridization reaction mixture of 1X Amplifase buffer (Lucigen, A1905B), 0.05 M KCl, 20% Formamide deionized (Millipore S4117), 0.1 uM Oligo-dT, 0.1 ug/ul BSA (New England Biolabs, B9000S), 1 U/ul RiboLock (Thermo, EO0384), and 0.2 ug/ul tRNAs (Ambion, AM7119). Then hybridization of padlock probes was done by incubating samples with padlock probes (with the concentration of each one 0.01 uM) mixed in blocking reagents used before (no oligo dT used in this step). The slide was then put into a PCR machine to denature at 55 °C for 15 min and hybridize at 45 °C for 120 min. Padlock probes were then ligated by using SplintR ligase (NEB M0375) at 25 °C for 16 hr followed by RCA (rolling cycle amplification) at 30 °C for 16 hr by using phi29 polymerase (Lucigen, 30221–2) and RCA primer1. Then a fixation step was applied to stabilize RCA product in 4% PFA for 15 min followed by washing in PBS Tween-20 0.05%. Then the hybridization of the first 3 genes was done by mixing all 3 3’ fluorophore-conjugated detection probes of each gene in reaction reagent 2XSSC, 20% Formamide deionized, 0.1 ug/ul BSA, and 0.5 ng/μl DAPI (Biolegend, 422801) followed by hybridization at 30 °C for 1 hr. The slides were washed in 20% formamide in 2 X SSC and then in 6 X SSC, followed by dehydration in 70%, 85%, and 100% ethanol until the chamber was removed. Then the samples were preserved in SlowFade Gold Antifade mountant (Thermo, S36936) and kept in the dark before imaging. After image acquisition, the first 3 detection probes were removed by using Uracil-DNA Glycosylase (Thermo, EN0362), and the slides were ready for the next round of detection probe hybridization. The procedure was repeated until all genes were hybridized and imaged.
Images were acquired with the Zeiss Axio Observer 7 fluorescent microscope with an automated stage setting to fix the imaging region for different hybridization rounds. Image analysis was done according to the published method (Sountoulidis et al., 2020). In brief, the DAPI channel from each round was extracted to measure the shift of imaging. The images were then aligned for all gene channels in Zen by creating an image subset with the shifting value. After alignment, images were exported into TIFF files and the threshold analysis was carried out for individual channels one by one in CellProfiler with scripts provided by the published method (Sountoulidis et al., 2020). Then, nuclear segmentation was done manually in the Fiji ROI manager to obtain nuclear ROIs, followed by an expansion of 2 μm in CellProfiler to obtain cell ROIs. Then signal dots were counted in these cell ROIs for each gene in CellProfiler and Fiji. Summary of the dot counts for each gene was exported to Excel files.
CUT&TAG
Request a detailed protocolCUT&TAG was performed according to the published method (Kaya-Okur et al., 2019). In brief, Prdm16_OE and Prdm16 mutant NSCs were cultured in the NeuroCult Proliferation Kit (Stem Cell Technologies, 05702) for 3 days with or without BMP4 at concentration of 25 ng/ml medium (Sigma, H4916). Cells on day 3 were then shortly rinsed with 1 X PBS and resuspended in 1 ml cold NE1 buffer on ice. Nuclei were lightly fixed with 0.1% formaldehyde to 1 ml PBS at RT for 2 min and neutralized with 75 mM Glycine. Nuclei were then washed three times, resuspended in 1 mL wash buffer and mixed with 90 μL Concanavalin A-coated magnetic beads (Polyscience 86057). The nuclei/beads mix was then blocked with 800 μL cold Antibody buffer for 5 min and resuspended in 1.2 mL Antibody buffer and aliquoted into 8 tubes with 150 μL each. 2 μL Anti-PRDM16 (Generous gift from Bryan Bjork lab) or IgG (SIGMA-ALDRICH, I5006) was added into each tube for an overnight incubation at 4 °C. The beads/nuclei/antibody mix was then washed with Dig-wash buffer and incubated with secondary antibody (1:100 dilution) for 1 hr. After further washes with the Dig-wash buffer. a pA-Tn5 adaptor complex in Dig-300 buffer was added to the beads for 1 hr reaction. The beads were then washed with Dig-300 buffer before the incubation with 300 μl Tagmentation buffer at 37 °C for 1 hr. Then 10 μl 0.5 M EDTA, 3 μL 10% SDS and 2.5 μL 20 mg/mL proteinase K (Invitrogen) were used to stop the reaction. Then the resultant DNA was purified with the DNA Clean & Concentrator kit (ZYMO Research D4013) and eluted in 25 uL Elution buffer. To generate libraries, 21 μL fragment DNA was mixed with 2 μL 10 uM Universal i5 primer, 2 μL 10 μM uniquely barcoded i7 primer and 25 μL PCR master mix (NEB Phusion High-Fidelity PCR Master Mix with HF Buffer, M0531). The PCR condition was as follows: 72 °C 5 min, 98 °C 30 s, repeat 12 times (98 °C 10 s, 63 °C 10 s), 72 °C 1 min and hold 4 °C. The libraries were cleaned with standard Ampure XP beads as previously described. Libraries from four biological replicates were produced for each condition.
ChIP
Request a detailed protocolChIP was performed as previously described (Dai et al., 2013b). For each ChIP reaction, 10 million Prdm16_OE, Prdm16_KO NSCs with or without 3 day BMP4 treatment were fixed, lysed, sonicated and made into chromatin extract. After precleared with gamma-bind-G beads, the chromatin extract was incubated with 2 μg PRDM16, 2 μg SMAD4 (Proteintech, 10231–1-AP), 2 μg pSMAD1/5/8 (Millipore, AB3848-1), or 2 μg SMAD3 (abcam, ab227223) in each ChIP reaction. In sequential ChIP assays, the same chromatin lysate was precleared and used, but with more antibodies in each reaction: 5 μg of IgG or PRDM16 antibody in the first round of immunoprecipitation. The elutes were then precleared again using gamma-bind G beads, divided into two equal halves and immunoprecipitated with 2 μg of IgG or the SMAD4 antibody. In ChIP-qPCR experiments, 2 μg SMAD4 and 2 μg c-FOS antibodies (Invitrogen, MA5-15055) were used in each reaction. The precipitated DNA was reverse cross-linked, and then purified using the Qiagen PCR purification kit.
Computation analyses
ChIP-seq libraries and analyses
Request a detailed protocol0.2% input and ChIPed DNA were made into libraries using the NEBNext Ultra II DNA Library Prep Kit and sequenced on the Illumina Nextseq500 platform. Three replicates of ChIP-seq samples, after the adaptor trimming by Trimmomatic, were mapped to the UCSC Mus musculus (mm10) genome assembly using Bowtie2 with the default parameters. The uniquely mapped reads (with mapping quality ≥ 20) were used for further analyses. The peaks were called by HOMER (v4.10) (Heinz et al., 2010). The reproducibility between replicates was estimated by Irreproducibility Discovery Rate (IDR), using the HOMER IDR pipeline (https://github.com/karmel/homer-idr; Allison, 2015). As suggested by the Encode IDR guideline, we used a relatively relaxed parameter ‘-F 2 -fdr 0.3 P.1 L 3 -LP.1’ for the true/pseudo/pooled replicates by the HOMER peak calling. The final confident peaks were determined by an IDR <5%. The peaks that were overlapped with the mm10 blacklist were also removed.
CUT&TAG analyses
Request a detailed protocolThe CUT&TAG samples of Prdm16_OE and Prdm16 mutant NSCs in four replicates, and of Prdm16 from CH and ChP area at stage 12.5 embryos in three replicates, were mapped to mm10 genome assembly using Bowtie2 (bowtie2 --end-to-end --very-sensitive --no-mixed --no-discordant --phred33 -I 10 X 700). The CUT&TAG coverage for these samples was generated by bedtools genomeCoverageBed and normalized to the library depths to give a read per million per base (RPM). By using the peak caller SEACR (Meers et al., 2019), which was designed for calling peaks from sparse chromatin profiling data such as CUT&TAG [22], we performed peak calling with FDR <10% for each replicated sample (SEACR_1.3.sh normalized_coverage 0.1 non-stringent output_peaks). To identify confident peaks, we selected the common peaks, which were called from all replicates for each condition. Peak overlapping analysis was performed by the Homer mergePeaks function with default parameters. Peak overlap analysis utilized HOMER’s mergePeaks function, specifically employing the default '-d given' parameter. This approach ensures that only peaks directly overlapping with each other are merged, maintaining the precise distances between peaks as provided in the input peak calls. This method is particularly beneficial for retaining the integrity of the original peak boundaries, avoiding any alterations to their relative positions. De novo motif discovery from SMAD4 and pSMAD1/5/8 peaks in Prdm16_OE NSCs and Prdm16 mutant NSCs was performed by MEME-ChIP software (Machanick and Bailey, 2011) (parameters: -ccut 100 -meme-p 5 -dna -meme-mod anr -minw 5 -maxw 15 -filter-thresh 0.05).
The CUT&TAG samples of H3K4me3 BMP4-treated and non-treated Prdm16_OE and Prdm16_KO cells in two or three replicates were mapped to the mm10 genome assembly using Bowtie2 (bowtie2 --end-to-end --very-sensitive --no-mixed --no-discordant --phred33 -I 10 X 700). Differential analysis of TSS up-stream and down-stream 500 bp between Prdm16_OE BMP-treated vs non-BMP-treated and Prdm16_KO vs Prdm16_OE BMP-treated was performed by the Limma R package (Ritchie et al., 2015). Gene Ontology (GO) enrichment analysis of up-regulated and down-regulated regions was performed by PANTHER (Thomas et al., 2022).
scRNA-seq analyses
Request a detailed protocolThe scRNA-seq data of the developing mouse brain were obtained from La Manno et al., 2021. The counts per cell were normalized by the logNormCounts function in the Bioconductor package ‘scater’ and the normalized expression data per cell were used to generate gene expression violin plots. To test whether Prdm16 target gene sets of Prdm16 with and without BMP4 and ChP E12.5 are significantly enriched among the scRNA-seq gene mean expression in each identified cell type cluster, the gene set enrichment analysis (GESA) ‘fgesa’ Bioconductor package was used (Korotkevich et al., 2021).
Appendix 1
Appendix 1—key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Genetic reagent (Mus musculus) | FVB/NJ | Strassman, A., et al., 2017 | A gift from Bryan C. Bjork’s laboratory | |
| Genetic reagent (Mus musculus) | Prdm16 GT | Strassman, A., et al., 2017 | A gift from Bryan C. Bjork’s laboratory | |
| Antibody | Rabbit polyclonal anti-PRDM16 | Gift from Bryan Bjork’s lab | IF (1:500–1000), WB (1:2000) Cut&Tag (1:150) | |
| Antibody | Goat polyclonal anti-SOX2 | R&D systems | AF2018 | IF (1:1000) |
| Antibody | Rabbit polyclonal anti-Ki67 | Abcam | ab15580 | IF (1:1000) |
| Antibody | Goat polyclonal anti-Doublecortin | SANTA CRUZ Biotechnology | sc-8066 | IF (1:500) |
| Antibody | beta Galactosidase Polyclonal Antibody | Thermo Fisher | A-11132 | IF (1:500) |
| Antibody | Rabbit polyclonal anti-SMAD4 | Proteintech | 10231–1-AP | IF (1:500) WB (1:2000) ChIP (1:150) |
| Antibody | Rabbit polyclonal anti-pSMAD1/5/8 | Millipore | AB3848-1 | IF (1:500) WB (1:2000) ChIP (1:150) |
| Antibody | Secondary antibodies conjugated to Cy3 or Cy5 or Alexa Fluor-488 | Jackson ImmunoResearch | IF (1:1000) | |
| Antibody | mouse anti-Rabbit IgG light chain specific | Jackson ImmunoResearch | Cat# 211-032-171 | WB (1:40000) |
| Antibody | Rabbit polyclonal anti-SMAD3 | abcam | ab227223 | IF (1:500) WB (1:1000) ChIP (1:150) |
| Antibody | Rabbit monoclonal anti-C-Fos | Invitrogen | MA5-15055 | ChIP (1:150) |
| Antibody | Rabbit polyclonal to Histone H3 (tri methyl K4) | abcam | ab8580 | CUT&TAG (1:150) |
| Antibody | IgG from rabbit serum | SIGMA-ALDRICH | I5006 | ChIP (1:150) CUT&TAG (1:150) |
| Antibody | Guinea Pig anti-Rabbit IgG (Heavy & Light Chain) Antibody - Preadsorbed | antibodies-online | ABIN101961 | ChIP (1:150) CUT&TAG (1:150) |
| Cell line | 293t | ATCC | CRL-3216 | |
| Sequence-based reagent | Ttr ISH F | This paper | PCR primers | GCTCTAGAGGATGGCTTCCCTTCGAC |
| Sequence-based reagent | Ttr ISH R | This paper | PCR primers | CCCAAGCTTAAAAATGCTTCCCGGCAT |
| Sequence-based reagent | Wnt2b ISH F | This paper | PCR primers | GCTCTAGAGAAGCTGCAGGGTGAGGAT |
| Sequence-based reagent | Wnt2b ISH R | This paper | PCR primers | GGAATTCTCATATCGCCTCCTCAGGTAGT |
| Sequence-based reagent | Bmp4 ISH F | This paper | PCR primers | CGGGATCCCCTGCAGCGATCCAGTCT |
| Sequence-based reagent | Bmp4 ISH R | This paper | PCR primers | GGAATTCGCCCAATCTCCACTCCCT |
| Sequence-based reagent | 18srRNA qRTF | This paper | RTqPCR primers | GTAACCCGTTGAACCCCATT |
| Sequence-based reagent | 18srRNA qRTR | This paper | RTqPCR primers | CCATCCAATCGGTAGTAGCG |
| Sequence-based reagent | Mki67 qRTF1F | This paper | RTqPCR primers | TGCCTCAGATGGCTCAAAGA |
| Sequence-based reagent | Mki67 qRTF1R | This paper | RTqPCR primers | TCTGCCAGTGTGCTGTTCTA |
| Sequence-based reagent | Wnt7b qRTF | This paper | RTqPCR primers | TGCCTTCACCTATGCCATCA |
| Sequence-based reagent | Wnt7b qRTR | This paper | RTqPCR primers | GTCACAGCCACAATTGCTCA |
| Sequence-based reagent | Prdm16 qRTF | This paper | RTqPCR primers | CACCGGGTCAGAGGAGAAAT |
| Sequence-based reagent | Prdm16 qRTR | This paper | RTqPCR primers | CGACATGTCAGGGCTCCTAT |
| Sequence-based reagent | Cdkn1a qRTF | This paper | RTqPCR primers | CAAAGTGTGCCGTTGTCTCT |
| Sequence-based reagent | Cdkn1a qRTR | This paper | RTqPCR primers | CGAAGTCAAAGTTCCACCGT |
| Sequence-based reagent | Mybl2 qRTF | This paper | RTqPCR primers | CCTGGAGCAGAGAGACAACA |
| Sequence-based reagent | Mybl2 qRTR | This paper | RTqPCR primers | GTACTGGCATTGCTGGTCTG |
| Sequence-based reagent | Ndc80 qRTF | This paper | RTqPCR primers | CGCCTCTCTATGCAGGAGTT |
| Sequence-based reagent | Ndc80 qRTR | This paper | RTqPCR primers | TTCCGATGTCGGTTTGTGTG |
| Sequence-based reagent | Nuf2 qRTF | This paper | RTqPCR primers | GAACAAACCGGACAAGTCGT |
| Sequence-based reagent | Nuf2 qRTR | This paper | RTqPCR primers | CTTCCTGGTGCACATTGCTT |
| Sequence-based reagent | Spc24 qRTF | This paper | RTqPCR primers | TGCTTTGAGACAGCATGGAG |
| Sequence-based reagent | Spc24 qRTR | This paper | RTqPCR primers | AGCCCTCTCTAACCTCTCCT |
| Sequence-based reagent | Spc25 qRTF | This paper | RTqPCR primers | AGAGTCGGAAGAGCTGACTG |
| Sequence-based reagent | Spc25 qRTR | This paper | RTqPCR primers | CGCTGATTTCTGCAGTCCTT |
| Sequence-based reagent | Wnt7b Co-bound enhancer ChIP qPCRF | This paper | ChIPqPCR primers | GTCATGTGCTTTCAGGGCAT |
| Sequence-based reagent | Wnt7b Co-bound enhancer ChIP qPCRR | This paper | ChIPqPCR primers | TCCTAGGAGTCACCATGGAA |
| Sequence-based reagent | Id1 Co-bound enhancer ChIP qPCRF | This paper | ChIPqPCR primers | GAGGCCACTCCAGTTCCAAT |
| Sequence-based reagent | Id1 Co-bound enhancer ChIP qPCRR | This paper | ChIPqPCR primers | GCGGTTCACATCTGGAGTCT |
| Sequence-based reagent | Wnt7b ectopic enhancer ChIP qPCRF | This paper | ChIPqPCR primers | CAGCGGTCAAGTTTAAGCGA |
| Sequence-based reagent | Wnt7b ectopic enhancer ChIP qPCRR | This paper | ChIPqPCR primers | CCACAGATGTCCCAGAGTGA |
| Sequence-based reagent | Id3 ectopic enhancer ChIP qPCRF | This paper | ChIPqPCR primers | CTTGGGCAATGTGAAAGGCT |
| Sequence-based reagent | Id3 ectopic enhancer ChIP qPCRR | This paper | ChIPqPCR primers | CAGTTGGTCCCTGTCAGTGT |
| Sequence-based reagent | Prdm16_PL_4 | This paper | SCRINSHOT padlock probe | /5Phos/AAGTCTTCAGAGATCTGCTTTTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCAGAACTTCTCGCTACCC |
| Sequence-based reagent | Ttr_Pl-1 | This paper | SCRINSHOT padlock probe | /5Phos/CTTCTACAAACTTCTCATCTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCTACTATCTTCTTTCAGTTCTACTCTGTACACTC |
| Sequence-based reagent | Foxj1_PL_1 | This paper | SCRINSHOT padlock probe | CGTGATCCACTTGTAGATGGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCGGAAGTAGCAGAAGTTGTC |
| Sequence-based reagent | Foxj1_PL_2 | This paper | SCRINSHOT padlock probe | /5Phos/GCCAAGAAGGTCTCATCAAAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGATGCTGTAGGAAGGATGTG |
| Sequence-based reagent | Foxj1_PL_3 | This paper | SCRINSHOT padlock probe | /5Phos/CACTTTGATGAAGCACTTGTTCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTCATCCTTCTCCCGAGG |
| Sequence-based reagent | Sox2_Pl-2 | This paper | SCRINSHOT padlock probe | /5Phos/TCGGCAGCCTGATTCCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCACGTCTATCTTCTTTTAGTTCAAATATATACATGGATTC |
| Sequence-based reagent | Sox2_Pl-3 | This paper | SCRINSHOT padlock probe | /5Phos/CTCTAAATATTACTGGTAAAGACTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCACGTCTATCTTCTTTCATCGCCCGGAGTCTAG |
| Sequence-based reagent | Ngn2_Pl-1 | This paper | SCRINSHOT padlock probe | /5Phos/TTGGTTTGACAGGTGAAATTCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCATCGCTATCTTCTTTCACATCAGAGAGGGAAAGT |
| Sequence-based reagent | Ngn2_Pl-2 | This paper | SCRINSHOT padlock probe | /5Phos/ACGCTTGCATTCAACCCTTTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCATCGCTATCTTCTTTGAGAATTCAGCCTAAATTTCC |
| Sequence-based reagent | Ngn2_Pl-3 | This paper | SCRINSHOT padlock probe | /5Phos/ATAAACCAGAGCTGGTCTCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCATCGCTATCTTCTTTCCATAAATCCTCACATCTTCA |
| Sequence-based reagent | Hes5_Pl-1 | This paper | SCRINSHOT padlock probe | /5Phos/CAAGAGTTCAAATTCTAAGTACTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCAGCTCTATCTTCTTTTGTGTTCTCCCACATGAC |
| Sequence-based reagent | Hes5_Pl-2 | This paper | SCRINSHOT padlock probe | /5Phos/CACACATTCTCTAAGAATGACTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCAGCTCTATCTTCTTTCCCAAATGACAACTCTGCA |
| Sequence-based reagent | Hes5_Pl-3 | This paper | SCRINSHOT padlock probe | /5Phos/TTAAGGATCATCGTGGAGACTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCAGCTCTATCTTCTTTCACCCATACAAAGGAATCCT |
| Sequence-based reagent | Zbtb20_PL-1 | This paper | SCRINSHOT padlock probe | /5Phos/CCATTTCCTGCTTGATGTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCATAGTAGTCATAGTCATCTT |
| Sequence-based reagent | Zbtb20_PL-2 | This paper | SCRINSHOT padlock probe | /5Phos/TACTTATCCGTCAGACACATTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGTAAGAAAGAGAGAAAGGTAC |
| Sequence-based reagent | Zbtb20_PL-3 | This paper | SCRINSHOT padlock probe | /5Phos/GAATGTGTCGAGTGGATGAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGAGCGTGAGAGTTTGTCAGT |
| Sequence-based reagent | Wnt2b_Pl-1 | This paper | SCRINSHOT padlock probe | /5Phos/TGAAGACAGACCTCTCCTATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCCGTACTATCTTCTTTTTAGTGCAACTCAAATCACTC |
| Sequence-based reagent | Wnt2b_Pl-2 | This paper | SCRINSHOT padlock probe | /5Phos/GTAATGGATGTTGTCACTACATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCCGTACTATCTTCTTTCTTGGCAAAGCGAACACC |
| Sequence-based reagent | Wnt2b_Pl-3 | This paper | SCRINSHOT padlock probe | /5Phos/CGTGCAGTAGTTAACAGTTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCCGTACTATCTTCTTTCTGTTTAAGTCGATCGTGGAA |
| Sequence-based reagent | Wnt3a_Pl-1 | This paper | SCRINSHOT padlock probe | /5Phos/CCTCTCTACTCAGGATAGAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCCATAGAGATTCCTAGAGAAC |
| Sequence-based reagent | Wnt3a_Pl-2 | This paper | SCRINSHOT padlock probe | /5Phos/ATGAGACCCTCACAGGTAGGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCAGAAACCGGGTCCTTAGGT |
| Sequence-based reagent | Wnt3a_Pl-3 | This paper | SCRINSHOT padlock probe | /5Phos/CATGTATCTAGATAGAGCAGTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCACCCTGAAGCACCCTCT |
| Sequence-based reagent | Wnt5a_PL-1 | This paper | SCRINSHOT padlock probe | /5Phos/TTAAGAGCCACAGGACTGATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTAATGAATATTGTTGGGCAATAAG |
| Sequence-based reagent | Wnt5a_PL-2 | This paper | SCRINSHOT padlock probe | /5Phos/TAGAGACCACCAAGAATTAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTTGAACAGGGTTATTCATACC |
| Sequence-based reagent | Wnt5a_PL-3 | This paper | SCRINSHOT padlock probe | /5Phos/CTTCATTGTTGTGTAAGTTCATGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTATACTGTCCTACGGCCTG |
| Sequence-based reagent | Wnt7b_PL-1 | This paper | SCRINSHOT padlock probe | /5Phos/AGGGATTTGTCTCCATTGGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCACTTTCCCAAAGAGAAGTAA |
| Sequence-based reagent | Wnt7b_PL-2 | This paper | SCRINSHOT padlock probe | /5Phos/CATTGTTGTGAAGGTTCATGAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGAACCTTTCTGCCCGCCT |
| Sequence-based reagent | Wnt7b_PL-3 | This paper | SCRINSHOT padlock probe | /5Phos/CCGGTCACAGCCACAATTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGTTGTAGTAGCCTTGCTTCTC |
| Sequence-based reagent | Axin2_PL-1 | This paper | SCRINSHOT padlock probe | /5Phos/AAGTCAAGAACACCTGGTAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCATATTCCAGGTAAATGTCAG |
| Sequence-based reagent | Axin2_PL-2 | This paper | SCRINSHOT padlock probe | /5Phos/TACGTGATAAGGATTGACTGGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCAAAGACATAGCCGGAACC |
| Sequence-based reagent | Axin2_PL-3 | This paper | SCRINSHOT padlock probe | /5Phos/TCCTGTATGGAATTTCTTCTCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTTTGAGCCTTCAGCATCC |
| Sequence-based reagent | Lef1_Pl-1 | This paper | SCRINSHOT padlock probe | /5Phos/CATATTGGGCATCATTATGTAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCACTGCTATCTTCTTTCATGTACGGGTCGCTGTT |
| Sequence-based reagent | Lef1_Pl-2 | This paper | SCRINSHOT padlock probe | /5Phos/CGTGCTAGTTCATAGTATTTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCACTGCTATCTTCTTTGTGTAGCTGTCTCTCTTTC |
| Sequence-based reagent | Lef1_Pl-3 | This paper | SCRINSHOT padlock probe | /5Phos/CTTCATCAGACACTTCACAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCACTGCTATCTTCTTTGGGTTTGGCTATCTCTTTAG |
| Sequence-based reagent | Tcf3_PL_1 | This paper | SCRINSHOT padlock probe | /5Phos/AGGTCACTCAGTTCCTTGTCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTACATCATGCTGAAGTCCAGG |
| Sequence-based reagent | Tcf3_PL_2 | This paper | SCRINSHOT padlock probe | /5Phos/GAGACCTGTCTCATCCAGAATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTTCTCTGCTCATGCTTCGATG |
| Sequence-based reagent | Tcf3_PL_3 | This paper | SCRINSHOT padlock probe | /5Phos/TCTTTCTCCTCCCGCTTGATTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTGATGCGATTTCCTCATCC |
| Sequence-based reagent | Fzd9-PL-1 | This paper | SCRINSHOT padlock probe | /5Phos/CACATAGAAAGGAAGATAATCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCAAGGAGTAGACATTGTAG |
| Sequence-based reagent | Fzd9-PL-2 | This paper | SCRINSHOT padlock probe | /5Phos/TTCATAAACGTAGCAGACAATGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGAAGTCCATGTTGAGGCG |
| Sequence-based reagent | Fzd9-PL-3 | This paper | SCRINSHOT padlock probe | /5Phos/CACTGGACCCTGCCCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTTAAATTACATCATTAAATAACTCTG |
| Sequence-based reagent | Fzd10-PL-1 | This paper | SCRINSHOT padlock probe | /5Phos/CTCAAGGTAGACTTCATGTATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTTACATTCAGCTGTTCTGAAC |
| Sequence-based reagent | Fzd10-PL-2 | This paper | SCRINSHOT padlock probe | /5Phos/TGGAGAGGAAGATGATAGGATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCGAATAAACGCAGTAGCACA |
| Sequence-based reagent | Fzd10-PL-3 | This paper | SCRINSHOT padlock probe | /5Phos/GATTGTTCATCTTACACTTGTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCAGTCAGGTGTCTTGGTCT |
| Sequence-based reagent | Ctnnb1_Pl-1 | This paper | SCRINSHOT padlock probe | /5Phos/GTGTCAACATCTTCTTCCTCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCATGCCTATCTTCTTTCATTCATAAAGGACTTGGGAG |
| Sequence-based reagent | Ctnnb1_Pl-2 | This paper | SCRINSHOT padlock probe | /5Phos/GTCGCTGCATCTGAAAGGTTTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCATGCCTATCTTCTTTCTTCCATCCCTTCCTGCTTA |
| Sequence-based reagent | Ctnnb1_Pl-3 | This paper | SCRINSHOT padlock probe | /5Phos/CAGGCCAGCTGATTGCTATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCATGCCTATCTTCTTTGATTTACAGGTCAGTATCAAAC |
| Sequence-based reagent | Bmp4_Pl-1 | This paper | SCRINSHOT padlock probe | /5Phos/AGAATTAACAGAGTTGACTAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCATCTATCTTCTTTAACAGGCCTTAGGGATACT |
| Sequence-based reagent | Bmp4_Pl-2 | This paper | SCRINSHOT padlock probe | /5Phos/GTGATGGAAACTCCTCACAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCATCTATCTTCTTTGATGTTCTCCAGATGTTCTTC |
| Sequence-based reagent | Bmp4_Pl-3 | This paper | SCRINSHOT padlock probe | /5Phos/GTGGTTGAATGGGAACGTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCATCTATCTTCTTTCAGTTTGTGTGGTATGTGTAG |
| Sequence-based reagent | Bmp7_Pl-1 | This paper | SCRINSHOT padlock probe | /5Phos/TTCTTCAGGATGACATTAGAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCTACGCTATCTTCTTTGACCACCATGTTTCTGTAC |
| Sequence-based reagent | Bmp7_Pl-2 | This paper | SCRINSHOT padlock probe | /5Phos/AGTCCTTATAGATCCTGAATTCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCTACGCTATCTTCTTTCAAATCGCTCCCGGATGT |
| Sequence-based reagent | Bmp7_Pl-3 | This paper | SCRINSHOT padlock probe | /5Phos/TGTCGCAGAACCTTCTGTTATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCTACGCTATCTTCTTTCTCAGATCCAGAAACCAATC |
| Sequence-based reagent | Smad6_PL-1 | This paper | SCRINSHOT padlock probe | /5Phos/ACTAAGGCTTGTGGATAACTCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTTTTCCCTGCAGGATCCGAA |
| Sequence-based reagent | Smad6_PL-2 | This paper | SCRINSHOT padlock probe | /5Phos/CAGTGTAAGACAATGTAGAATCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTAGTTGGTGGCCTCGGTTT |
| Sequence-based reagent | Smad6_PL-3 | This paper | SCRINSHOT padlock probe | /5Phos/AGTAGGATCTCCAGCCAACTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCATTGCTATCTGTGGTTGTTG |
| Sequence-based reagent | Smad7_PL-1 | This paper | SCRINSHOT padlock probe | /5Phos/GCCTGCAGTTGGTTTGAGAATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGAAGGTACAGCATCTGGACA |
| Sequence-based reagent | Smad7_PL-2 | This paper | SCRINSHOT padlock probe | /5Phos/GTACCAGCTGACTCTTGTTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGATCTTGCTCCGCACTTTCT |
| Sequence-based reagent | Smad7_PL-3 | This paper | SCRINSHOT padlock probe | /5Phos/CCTCTTGACTTCCGAGGAATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGTAAGATTCACAGCAACACAG |
| Sequence-based reagent | Nog_PL-1 | This paper | SCRINSHOT padlock probe | /5Phos/AGGGTCTGGATGTTCGATGATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTTCTCCTTAGGGTCAAAGAT |
| Sequence-based reagent | Nog_PL-2 | This paper | SCRINSHOT padlock probe | /5Phos/GATATAAATAGCAGGTTAACATTATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCATGCGAAGGGTACTGG |
| Sequence-based reagent | Nog_PL-3 | This paper | SCRINSHOT padlock probe | /5Phos/TGCATTACAGGAACCAGAAATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCATTCCTACACAGTTAAACAG |
| Sequence-based reagent | mKi67_PL_1 | This paper | SCRINSHOT padlock probe | /5Phos/GGACTTTCCTGGAAATTCTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTTGATGGTTCCTTTCCAAG |
| Sequence-based reagent | mKi67_PL_2 | This paper | SCRINSHOT padlock probe | /5Phos/ACTTTCCTTGCATCTTTGAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGAACCTTCACTTAATTCACC |
| Sequence-based reagent | mKi67_PL_3 | This paper | SCRINSHOT padlock probe | /5Phos/CTGAGGCTTTATCAAGATTTCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCAGAGACTCCTTTCTCTTC |
| Sequence-based reagent | Cdkn1c_PL_1 | This paper | SCRINSHOT padlock probe | /5Phos/CAAGTTTAAGAGCAATCTAATGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTTTTCGACTGTCTGGTCAC |
| Sequence-based reagent | Cdkn1c_PL_2 | This paper | SCRINSHOT padlock probe | /5Phos/CGCATGGATAAATACAAATATTAATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGTGGCAGAGGGATCCA |
| Sequence-based reagent | Cdkn1c_PL_3 | This paper | SCRINSHOT padlock probe | /5Phos/GCCTCTAAACTAACTCATCTCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCAAGTTCTCTCTGGCCGTTA |
| Sequence-based reagent | Id3-1 | This paper | SCRINSHOT padlock probe | /5Phos/AGAGTAGAGATAGAGAGGGAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTCAGCGCCTTCATGTTGG |
| Sequence-based reagent | Id3-2 | This paper | SCRINSHOT padlock probe | /5Phos/GGGCTGGGTTAAGATCGAATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTTGAGTTCAGGGTAAGTGAAGA |
| Sequence-based reagent | Id3-3 | This paper | SCRINSHOT padlock probe | /5Phos/TGCTGCCTTGGCAGAGTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGAAACCAGAAGAACAGCTCTTA |
| Sequence-based reagent | Spc24-1 | This paper | SCRINSHOT padlock probe | /5Phos/GCTGCTCTTCTATAGAATACTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTCAACAATTAAGAGCACTG |
| Sequence-based reagent | Spc24-2 | This paper | SCRINSHOT padlock probe | /5Phos/AAATCAATGAACGCACTCACTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTGTCTCAAAGCAAGTAGAT |
| Sequence-based reagent | Spc24-3 | This paper | SCRINSHOT padlock probe | /5Phos/CACAGCTAACAGACACACATTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTCTAACCTCTCCTGGAATA |
| Sequence-based reagent | Nrarp-1 | This paper | SCRINSHOT padlock probe | /5Phos/GGTACCATGCTCTCAGAATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTTATAATAGAAGTCACTAGGAAG |
| Sequence-based reagent | Nrarp-2 | This paper | SCRINSHOT padlock probe | /5Phos/ACTTCATACAGATAGAAGAGACTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTTAGGAGTGTGCCACAG |
| Sequence-based reagent | Nrarp-3 | This paper | SCRINSHOT padlock probe | /5Phos/TAGTATGAGAGATTCACAGTCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCCTTCCACTCATTCAGCAA |
| Sequence-based reagent | Rassf4-1 | This paper | SCRINSHOT padlock probe | /5Phos/ACTGAGGTCTTGTGGTTATAAATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCCATAGGCAGGAGTGAAC |
| Sequence-based reagent | Rassf4-2 | This paper | SCRINSHOT padlock probe | /5Phos/CATCAAGAAGATCTTGACAATTTTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTCGCTCAAGTCAGCTTC |
| Sequence-based reagent | Rassf4-3 | This paper | SCRINSHOT padlock probe | CTCGGTGTCTTAGCTGAAAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGATCAGAGTCCCTTCTTCCT |
| Sequence-based reagent | Nuak2-1 | This paper | SCRINSHOT padlock probe | /5Phos/TTGCTGCTATTCTCAAACACTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTACTCCATGACAATCACAATC |
| Sequence-based reagent | Nuak2-2 | This paper | SCRINSHOT padlock probe | /5Phos/GGACTTCTTAAGAGAATGTTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCCATGTCATTCTCCTTTCG |
| Sequence-based reagent | Nuak2-3 | This paper | SCRINSHOT padlock probe | /5Phos/GTCGAGACTTCTTAAGGATGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGTAGTAACCAGATTCACGCT |
| Sequence-based reagent | Fam83d-1 | This paper | SCRINSHOT padlock probe | /5Phos/AATTGTCCGAACTGTCATCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTTGCATAGTAGATATTTCCTGT |
| Sequence-based reagent | Fam83d-2 | This paper | SCRINSHOT padlock probe | /5Phos/GTAGAAGCCTCAGAGTCAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGAAATAGTCTTCATCGCTGATG |
| Sequence-based reagent | Fam83d-3 | This paper | SCRINSHOT padlock probe | /5Phos/GTTGGTAGAGGCTCTGGTAATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTATGTCCCTCACAGAAACCAG |
| Sequence-based reagent | Mybl2-1 | This paper | SCRINSHOT padlock probe | /5Phos/AAGTAGCAGCTATGGCAATCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTATGTCCGAGTTCTTTAGCAG |
| Sequence-based reagent | Mybl2-2 | This paper | SCRINSHOT padlock probe | /5Phos/TTCTGATGATGGATACTTCTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTACTTCTGATCTGGGAGTAC |
| Sequence-based reagent | Mybl2-3 | This paper | SCRINSHOT padlock probe | /5Phos/TTGACCAACTCAATGACCTTTTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTGTTTGGTGCCATACTTC |
| Sequence-based reagent | Gas2-1 | This paper | SCRINSHOT padlock probe | /5Phos/TTCCTGAAGACTTTGATGAATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTTAAGTTTCCAGTACTCTTCT |
| Sequence-based reagent | Gas2-2 | This paper | SCRINSHOT padlock probe | /5Phos/CTGTTATCTCTTTCCCTAATATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTTTCTCCATGAAGGTTTCTG |
| Sequence-based reagent | Gas2-3 | This paper | SCRINSHOT padlock probe | /5Phos/TCTTTGAATTTCTCCTGCACATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTTGTTGGCATCCATACTC |
| Sequence-based reagent | Ada-1 | This paper | SCRINSHOT padlock probe | /5Phos/GAGAGCTTCATCCTCGATTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTTTTCTTTCAGTAGTCTGTTGTA |
| Sequence-based reagent | Ada-2 | This paper | SCRINSHOT padlock probe | /5Phos/AAACTTGGCCAGGAAGCCTTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTAATCACAGGCATGTAGTAGTC |
| Sequence-based reagent | Ada-3 | This paper | SCRINSHOT padlock probe | /5Phos/CAGCTCCAACACCTCAAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTTCTTCTGATTGTACTTCTTACA |
| Sequence-based reagent | Hspg2-1 | This paper | SCRINSHOT padlock probe | /5Phos/CTTCCTGTAGGCATCTTTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGAAGGTGATCTTGATCTCAAA |
| Sequence-based reagent | Hspg2-2 | This paper | SCRINSHOT padlock probe | /5Phos/TGGATATCAGGGTCCAAGATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTATGATGTTATTGCCCGTAATC |
| Sequence-based reagent | Hspg2-3 | This paper | SCRINSHOT padlock probe | /5Phos/CTTGAGCCATGTACCTGATGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTTCACCTGGAGAAGTCTCAGT |
| Sequence-based reagent | shc4-1 | This paper | SCRINSHOT padlock probe | /5Phos/TCATGGTCTCTGTCTTCAGGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCCGGGATGGAGTTGTAATAG |
| Sequence-based reagent | shc4-2 | This paper | SCRINSHOT padlock probe | /5Phos/AGTCCTGTTTAGATTGGATTCTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGGTGCAGGTGCAGGTCC |
| Sequence-based reagent | shc4-3 | This paper | SCRINSHOT padlock probe | /5Phos/GGTTGTCAAGGTTCATCAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTGGTGATTTGCAATAATCTGTT |
| Sequence-based reagent | Nuf2-1 | This paper | SCRINSHOT padlock probe | /5Phos/AATCACTTTCAATCTGGTCCTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCAGTTTCTTTAGTTCCGATG |
| Sequence-based reagent | Nuf2-2 | This paper | SCRINSHOT padlock probe | /5Phos/GGCTTTCATGTAAATCATGTTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTACTCCATACACTAACTGTAA |
| Sequence-based reagent | Nuf2-3 | This paper | SCRINSHOT padlock probe | /5Phos/GTGGTTACTTGCTCACAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTTGTGGATTTCTTGATTAACA |
| Sequence-based reagent | Ndc80-1 | This paper | SCRINSHOT padlock probe | /5Phos/ATACTCTCTGTCGGCTTTATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTATCTTCAGACATGAATTCTTC |
| Sequence-based reagent | Ndc80-2 | This paper | SCRINSHOT padlock probe | /5Phos/GGATCCATGTCCGCTAGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTAATATACCAAGTTGACTATTCCT |
| Sequence-based reagent | Ndc80-3 | This paper | SCRINSHOT padlock probe | /5Phos/CTGTTCATTTAGTGCTTTGTTTTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTCCTCCAGTCTTGCAAT |
| Sequence-based reagent | Spc25-1 | This paper | SCRINSHOT padlock probe | /5Phos/CCCATGTTTAGGTAGACTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTAGATACCTCTATCATCTAGGT |
| Sequence-based reagent | Spc25-2 | This paper | SCRINSHOT padlock probe | /5Phos/ACAGATCCGCTGATTTCTGTCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTCTAGTCCAAGGTAATCTCTAT |
| Sequence-based reagent | Spc25-3 | This paper | SCRINSHOT padlock probe | /5Phos/AAATGGTTTCCCTCTTCCTATCCTCTATGATTACTGACTGCGTCTATTTAGTGGAGCCGCCCCTATCTTCTTTATTAGCTTTGTTAGCAGTGG |
| Sequence-based reagent | Prdm16_Det_4 | This paper | SCRINSHOT detection probe | TCGCUACCCAAGUCTTCAGA/3Cy3Sp/ |
| Sequence-based reagent | Ttr_Det-1 | This paper | SCRINSHOT detection probe | TACTCUGTACACUCCTTCUACAAACTTC-FITC |
| Sequence-based reagent | Foxj1_Det_1 | This paper | SCRINSHOT detection probe | AGAAGTUGTCCGTGAUCCAC/3Cy5Sp/ |
| Sequence-based reagent | Foxj1_Det_2 | This paper | SCRINSHOT detection probe | AGGAAGGAUGUGGCCAAG/3Cy5Sp/ |
| Sequence-based reagent | Foxj1_Det_3 | This paper | SCRINSHOT detection probe | CGAGGCACUTTGAUGAAGCA/3Cy5Sp/ |
| Sequence-based reagent | Sox2_Det-2 | This paper | SCRINSHOT detection probe | ATATAUACATGGATUCTCGGCAGC-cy3 |
| Sequence-based reagent | Sox2_Det-3 | This paper | SCRINSHOT detection probe | CGGAGTCUAGCTCTAAAUATTACTGG-cy3 |
| Sequence-based reagent | Ngn2_Det-1 | This paper | SCRINSHOT detection probe | GGGAAAGUTTGGTTUGACAGGT-Cy3 |
| Sequence-based reagent | Ngn2_Det-2 | This paper | SCRINSHOT detection probe | GCCTAAAUTTCCACGCUTGC-Cy3 |
| Sequence-based reagent | Ngn2_Det-3 | This paper | SCRINSHOT detection probe | ATCCTCACAUCTTCAAUAAACCAGAG-Cy3 |
| Sequence-based reagent | Hes5_Det-1 | This paper | SCRINSHOT detection probe | CCCACAUGACCAAGAGUTCAAAT-Cy3 |
| Sequence-based reagent | Hes5_Det-2 | This paper | SCRINSHOT detection probe | CAACTCUGCACACACAUTCTCT-Cy3 |
| Sequence-based reagent | Hes5_Det-3 | This paper | SCRINSHOT detection probe | CAAAGGAAUCCTTUAAGGAUCATCGT-Cy3 |
| Sequence-based reagent | Zbtb20_Det-1 | This paper | SCRINSHOT detection probe | GTCATAGUCATCTTCCAUTTCCTGC/3Cy5Sp/ |
| Sequence-based reagent | Zbtb20_Det-2 | This paper | SCRINSHOT detection probe | GAGAAAGGUACTACTTAUCCGTCAG/3Cy5Sp/ |
| Sequence-based reagent | Zbtb20_Det-3 | This paper | SCRINSHOT detection probe | AGTTTGUCAGTGAAUGTGTCGA/3Cy5Sp/ |
| Sequence-based reagent | Wnt2b_Det-1 | This paper | SCRINSHOT detection probe | CAACUCAAAUCACTCUGAAGACAGA-Cy5 |
| Sequence-based reagent | Wnt2b_Det-2 | This paper | SCRINSHOT detection probe | CGAACACCGUAATGGAUGTTGT-Cy5 |
| Sequence-based reagent | Wnt2b_Det-3 | This paper | SCRINSHOT detection probe | ATCGUGGAACGUGCAGUAGT-Cy5 |
| Sequence-based reagent | Wnt3a_Det-1 | This paper | SCRINSHOT detection probe | TTCCUAGAGAACCCUCTCTACTC-Cy5 |
| Sequence-based reagent | Wnt3a_Det-2 | This paper | SCRINSHOT detection probe | GTCCTTAGGUATGAGACCCUCA-Cy5 |
| Sequence-based reagent | Wnt3a_Det-3 | This paper | SCRINSHOT detection probe | AAGCACCCUCTCATGTAUCTAGATA-Cy5 |
| Sequence-based reagent | Foxj1_Det_2 | This paper | SCRINSHOT detection probe | TTGGGCAAUAAGTUAAGAGCCA/3Cy3Sp/ |
| Sequence-based reagent | Foxj1_Det_3 | This paper | SCRINSHOT detection probe | CAGGGTTAUTCATACCUAGAGACCA/3Cy3Sp/ |
| Sequence-based reagent | Sox2_Det-2 | This paper | SCRINSHOT detection probe | CGGCCUGCTTCATUGTTG/3Cy3Sp/ |
| Sequence-based reagent | Sox2_Det-3 | This paper | SCRINSHOT detection probe | AGAGAAGUAAAGGGATUTGTCTCC/36-FAM/ |
| Sequence-based reagent | Ngn2_Det-1 | This paper | SCRINSHOT detection probe | CCCGCCUCATTGUTGTGA/36-FAM/ |
| Sequence-based reagent | Ngn2_Det-2 | This paper | SCRINSHOT detection probe | CTTGCTUCTCCCGGUCAC/36-FAM/ |
| Sequence-based reagent | Ngn2_Det-3 | This paper | SCRINSHOT detection probe | AGGTAAATGUCAGAAGUCAAGAACA/36-FAM/ |
| Sequence-based reagent | Hes5_Det-1 | This paper | SCRINSHOT detection probe | CGGAACCUACGTGAUAAGGATT/36-FAM/ |
| Sequence-based reagent | Hes5_Det-2 | This paper | SCRINSHOT detection probe | TCAGCAUCCTCCTGUATGGA/36-FAM/ |
| Sequence-based reagent | Hes5_Det-3 | This paper | SCRINSHOT detection probe | TCGCUACCCAAGUCTTCAGA/3Cy3Sp/ |
| Sequence-based reagent | Zbtb20_Det-1 | This paper | SCRINSHOT detection probe | TACTCUGTACACUCCTTCUACAAACTTC-FITC |
| Sequence-based reagent | Zbtb20_Det-2 | This paper | SCRINSHOT detection probe | AGAAGTUGTCCGTGAUCCAC/3Cy5Sp/ |
| Sequence-based reagent | Zbtb20_Det-3 | This paper | SCRINSHOT detection probe | AGGAAGGAUGUGGCCAAG/3Cy5Sp/ |
| Sequence-based reagent | Wnt2b_Det-1 | This paper | SCRINSHOT detection probe | CGAGGCACUTTGAUGAAGCA/3Cy5Sp/ |
| Sequence-based reagent | Wnt2b_Det-2 | This paper | SCRINSHOT detection probe | ATATAUACATGGATUCTCGGCAGC-cy3 |
| Sequence-based reagent | Wnt2b_Det-3 | This paper | SCRINSHOT detection probe | CGGAGTCUAGCTCTAAAUATTACTGG-cy3 |
| Sequence-based reagent | Wnt3a_Det-1 | This paper | SCRINSHOT detection probe | GGGAAAGUTTGGTTUGACAGGT-Cy3 |
| Sequence-based reagent | Wnt3a_Det-2 | This paper | SCRINSHOT detection probe | GCCTAAAUTTCCACGCUTGC-Cy3 |
| Sequence-based reagent | Wnt3a_Det-3 | This paper | SCRINSHOT detection probe | ATCCTCACAUCTTCAAUAAACCAGAG-Cy3 |
| Sequence-based reagent | Wnt5a_Det-1 | This paper | SCRINSHOT detection probe | CCCACAUGACCAAGAGUTCAAAT-Cy3 |
| Sequence-based reagent | Wnt5a_Det-2 | This paper | SCRINSHOT detection probe | CAACTCUGCACACACAUTCTCT-Cy3 |
| Sequence-based reagent | Wnt5a_Det-3 | This paper | SCRINSHOT detection probe | CAAAGGAAUCCTTUAAGGAUCATCGT-Cy3 |
| Sequence-based reagent | Wnt7b_Det-1 | This paper | SCRINSHOT detection probe | GTCATAGUCATCTTCCAUTTCCTGC/3Cy5Sp/ |
| Sequence-based reagent | Wnt7b_Det-2 | This paper | SCRINSHOT detection probe | GAGAAAGGUACTACTTAUCCGTCAG/3Cy5Sp/ |
| Sequence-based reagent | Wnt7b_Det-3 | This paper | SCRINSHOT detection probe | AGTTTGUCAGTGAAUGTGTCGA/3Cy5Sp/ |
| Sequence-based reagent | Axin2_Det-1 | This paper | SCRINSHOT detection probe | CAACUCAAAUCACTCUGAAGACAGA-Cy5 |
| Sequence-based reagent | Axin2_Det-2 | This paper | SCRINSHOT detection probe | CGAACACCGUAATGGAUGTTGT-Cy5 |
| Sequence-based reagent | Axin2_Det-3 | This paper | SCRINSHOT detection probe | ATCGUGGAACGUGCAGUAGT-Cy5 |
| Sequence-based reagent | Lef1_Det-1 | This paper | SCRINSHOT detection probe | TCGCTGUTCATATUGGGCAT-FITC |
| Sequence-based reagent | Lef1_Det-2 | This paper | SCRINSHOT detection probe | TCTCTCTTUCCGTGCUAGTTCA-FITC |
| Sequence-based reagent | Lef1_Det-3 | This paper | SCRINSHOT detection probe | GCTATCTCTUTAGCTTCAUCAGACAC-FITC |
| Sequence-based reagent | Tcf3_Det_1 | This paper | SCRINSHOT detection probe | GAAGUCCAGGAGGUCACTC/36-FAM/ |
| Sequence-based reagent | Tcf3_Det_2 | This paper | SCRINSHOT detection probe | GCTTCGAUGGAGACCUGTC/36-FAM/ |
| Sequence-based reagent | Tcf3_Det_3 | This paper | SCRINSHOT detection probe | GATTTCCUCATCCTCTUTCTCCTC/36-FAM/ |
| Sequence-based reagent | Fzd9-Det-1 | This paper | SCRINSHOT detection probe | GTAGACATUGTAGCACAUAGAAAGGA/36-FAM/ |
| Sequence-based reagent | Fzd9-Det-2 | This paper | SCRINSHOT detection probe | TGAGGCGTUCATAAACGUAGC/36-FAM/ |
| Sequence-based reagent | Fzd9-Det-3 | This paper | SCRINSHOT detection probe | TAAATAACUCTGCACUGGACCC/36-FAM/ |
| Sequence-based reagent | Fzd10-Det-1 | This paper | SCRINSHOT detection probe | GCTGTUCTGAACCUCAAGGTA/3Cy5Sp/ |
| Sequence-based reagent | Fzd10-Det-2 | CGCAGUAGCACAUGGAGAG/3Cy5Sp/ | ||
| Sequence-based reagent | Fzd10-Det-3 | This paper | SCRINSHOT detection probe | TGTCTTGGUCTGATTGTUCATCTT/3Cy5Sp/ |
| Sequence-based reagent | Ctnnb1_Det-1 | This paper | SCRINSHOT detection probe | GACUTGGGAGGUGTCAACAT-Cy5 |
| Sequence-based reagent | Ctnnb1_Det-2 | This paper | SCRINSHOT detection probe | CCTGCUTAGTCGCUGCA-Cy5 |
| Sequence-based reagent | Ctnnb1_Det-3 | This paper | SCRINSHOT detection probe | TTACAGGUCAGTAUCAAACCAGG-Cy5 |
| Sequence-based reagent | Bmp4_Det-1 | This paper | SCRINSHOT detection probe | GCCTTUGGGATACUAGAATUAACAGAG-FITC |
| Sequence-based reagent | Bmp4_Det-2 | This paper | SCRINSHOT detection probe | CAGAUGTTCTUCGTGAUGGAAACT-FITC |
| Sequence-based reagent | Bmp4_Det-3 | This paper | SCRINSHOT detection probe | GTGGUATGTGUAGGTGGUTGAATG-FITC |
| Sequence-based reagent | Bmp7_Det-1 | This paper | SCRINSHOT detection probe | CATGTTTCUGTACTTCTUCAGGATGA-Cy5 |
| Sequence-based reagent | Bmp7_Det-2 | This paper | SCRINSHOT detection probe | TCCCGGAUGTAGTCCUTATAGATC-Cy5 |
| Sequence-based reagent | Bmp7_Det-3 | This paper | SCRINSHOT detection probe | GAAACCAAUCTGUCGCAGAAC-Cy5 |
| Sequence-based reagent | Smad6_Det-1 | This paper | SCRINSHOT detection probe | CAGGAUCCGAAACUAAGGCTT/36-FAM/ |
| Sequence-based reagent | Smad6_Det-2 | This paper | SCRINSHOT detection probe | CCTCGGUTTCAGTGUAAGACAA/36-FAM/ |
| Sequence-based reagent | Smad6_Det-3 | This paper | SCRINSHOT detection probe | CTGTGGUTGTTGAGUAGGATCT/36-FAM/ |
| Sequence-based reagent | Smad7_Det-1 | This paper | SCRINSHOT detection probe | CUGGACAGCCUGCAGT/36-FAM/ |
| Sequence-based reagent | Smad7_Det-2 | This paper | SCRINSHOT detection probe | CGCACUTTCTGUACCAGCT/36-FAM/ |
| Sequence-based reagent | Smad7_Det-3 | This paper | SCRINSHOT detection probe | ACACAGCCUCTTGACUTCC/36-FAM/ |
| Sequence-based reagent | Nog_Det-1 | This paper | SCRINSHOT detection probe | GTCAAAGAUAGGGTCUGGATGTTC/3Cy5Sp/ |
| Sequence-based reagent | Nog_Det-2 | This paper | SCRINSHOT detection probe | GCGAAGGGUACTGGGATAUAAATAG/3Cy5Sp/ |
| Sequence-based reagent | Nog_Det-3 | This paper | SCRINSHOT detection probe | CTACACAGUTAAACAGUGCATTACAG/3Cy5Sp/ |
| Sequence-based reagent | mKi67_Det_1 | This paper | SCRINSHOT detection probe | CCTTUCCAAGGGACUTTCCT/3Cy3Sp/ |
| Sequence-based reagent | mKi67_Det_2 | This paper | SCRINSHOT detection probe | ACTTAATUCACCACTTUCCTTGCA/3Cy3Sp/ |
| Sequence-based reagent | mKi67_Det_3 | This paper | SCRINSHOT detection probe | CTTTCTCTUCCTGAGGCUTTATCA/3Cy3Sp/ |
| Sequence-based reagent | Cdkn1c_Det-1 | This paper | SCRINSHOT detection probe | TCTGGUCACCAAGTUTAAGAGC/36-FAM/ |
| Sequence-based reagent | Cdkn1c_Det-2 | This paper | SCRINSHOT detection probe | GGGAUCCACGCAUGGATAA/36-FAM/ |
| Sequence-based reagent | Cdkn1c_Det-3 | This paper | SCRINSHOT detection probe | TGGCCGUTAGCCTCUAAAC/36-FAM/ |
| Sequence-based reagent | Id3-Det-1 | This paper | SCRINSHOT detection probe | CCTTCATGUTGGAGAGUAGAGATAG/3Cy3Sp/ |
| Sequence-based reagent | Id3-Det-2 | This paper | SCRINSHOT detection probe | TAAGUGAAGAGGGCUGGGT/3Cy3Sp/ |
| Sequence-based reagent | Id3-Det-3 | This paper | SCRINSHOT detection probe | GAACAGCUCTTATGCUGCCT/3Cy3Sp/ |
| Sequence-based reagent | Spc24-Det-1 | This paper | SCRINSHOT detection probe | GAGCACUGGCTGCUCTT/36-FAM/ |
| Sequence-based reagent | Spc24-Det-2 | This paper | SCRINSHOT detection probe | AAAGCAAGUAGATAAAUCAATGAACGC/36-FAM/ |
| Sequence-based reagent | Spc24-Det-3 | This paper | SCRINSHOT detection probe | CCTGGAAUACACAGCUAACAGAC/36-FAM/ |
| Sequence-based reagent | Nrarp-Det-1 | This paper | SCRINSHOT detection probe | AGTCACUAGGAAGGGUACCATG/3Cy5Sp/ |
| Sequence-based reagent | Nrarp-Det-2 | This paper | SCRINSHOT detection probe | GUGCCACAGACUTCAUACAGATAG/3Cy5Sp/ |
| Sequence-based reagent | Nrarp-Det-3 | This paper | SCRINSHOT detection probe | ACTCATUCAGCAATAGUATGAGAGAUTC/3Cy5Sp/ |
| Sequence-based reagent | Rassf4-Det-1 | This paper | SCRINSHOT detection probe | AGGAGUGAACACUGAGGTCT/3Cy3Sp/ |
| Sequence-based reagent | Rassf4-Det-2 | This paper | SCRINSHOT detection probe | TCAGCUTCCAUCAAGAAGAUCTTG/3Cy3Sp/ |
| Sequence-based reagent | Rassf4-Det-3 | This paper | SCRINSHOT detection probe | CCTTCUTCCTCUCGGTGTC/3Cy3Sp/ |
| Sequence-based reagent | Nuak2-Det-1 | This paper | SCRINSHOT detection probe | GACAAUCACAAUCTTGCTGCUATTC/36-FAM/ |
| Sequence-based reagent | Nuak2-Det-2 | This paper | SCRINSHOT detection probe | TCTCCTTUCGGGACTUCTTAAGA/36-FAM/ |
| Sequence-based reagent | Nuak2-Det-3 | This paper | SCRINSHOT detection probe | TCACGCTGUCGAGACUTCT/36-FAM/ |
| Sequence-based reagent | Fam83d-Det-1 | This paper | SCRINSHOT detection probe | GTAGATATTUCCTGTAATUGTCCGAACT/36-FAM/ |
| Sequence-based reagent | Fam83d-Det-2 | This paper | SCRINSHOT detection probe | TTCATCGCUGATGGUAGAAGC/36-FAM/ |
| Sequence-based reagent | Fam83d-Det-3 | This paper | SCRINSHOT detection probe | ACCAGGUTGGUAGAGGCT/36-FAM/ |
| Sequence-based reagent | Mybl2-Det-1 | This paper | SCRINSHOT detection probe | GTTCTTUAGCAGAAGUAGCAGCT/3Cy5Sp/ |
| Sequence-based reagent | Mybl2-Det-2 | This paper | SCRINSHOT detection probe | CTGGGAGUACTTCTGAUGATGG/3Cy5Sp/ |
| Sequence-based reagent | Mybl2-Det-3 | This paper | SCRINSHOT detection probe | GCCAUACTTCTUGACCAACUCAAT/3Cy5Sp/ |
| Sequence-based reagent | Gas2-Det-1 | This paper | SCRINSHOT detection probe | CCAGTACUCTTCTTTCCUGAAGAC/36-FAM/ |
| Sequence-based reagent | Gas2-Det-2 | This paper | SCRINSHOT detection probe | TGAAGGTTUCTGCTGTTAUCTCTTTC/36-FAM/ |
| Sequence-based reagent | Gas2-Det-3 | This paper | SCRINSHOT detection probe | CATCCATACUCTCTTTGAAUTTCTCCT/36-FAM/ |
| Sequence-based reagent | Ada-Det-1 | This paper | SCRINSHOT detection probe | AGTAGUCTGTTGUAGAGAGCUTCAT/36-FAM/ |
| Sequence-based reagent | Ada-Det-2 | This paper | SCRINSHOT detection probe | CATGTAGUAGTCAAACUTGGCCA/36-FAM/ |
| Sequence-based reagent | Ada-Det-3 | This paper | SCRINSHOT detection probe | GTACTTCUTACACAGCUCCAACA/36-FAM/ |
| Sequence-based reagent | Hspg2-Det-1 | This paper | SCRINSHOT detection probe | CTTGATCUCAAACTTCCUGTAGGC/3Cy3Sp/ |
| Sequence-based reagent | Hspg2-Det-2 | This paper | SCRINSHOT detection probe | GCCCGTAAUCTGGAUATCAGG/3Cy3Sp/ |
| Sequence-based reagent | Hspg2-Det-3 | This paper | SCRINSHOT detection probe | AGAAGTCUCAGTCTUGAGCCA/3Cy3Sp/ |
| Sequence-based reagent | Shc4-Det-1 | This paper | SCRINSHOT detection probe | GGAGTTGUAATAGTCAUGGTCTCTG/3Cy3Sp/ |
| Sequence-based reagent | Shc4-Det-2 | This paper | SCRINSHOT detection probe | GCAGGUCCAGTCCUGTTTAG/3Cy3Sp/ |
| Sequence-based reagent | Shc4-Det-3 | This paper | SCRINSHOT detection probe | GCAATAAUCTGTTGGTUGTCAAGG/3Cy3Sp/ |
| Sequence-based reagent | Nuf2-Det-1 | This paper | SCRINSHOT detection probe | AGTTCCGAUGAATCACTUTCAATCT/3Cy5Sp/ |
| Sequence-based reagent | Nuf2-Det-2 | This paper | SCRINSHOT detection probe | TACACTAACUGTAAGGCTUTCATGTAA/3Cy5Sp/ |
| Sequence-based reagent | Nuf2-Det-3 | This paper | SCRINSHOT detection probe | ATTTCTTGAUTAACAGTGGUTACTTGC/3Cy5Sp/ |
| Sequence-based reagent | Ndc80-1 | This paper | SCRINSHOT detection probe | GACATGAATUCTTCATACUCTCTGTCG/3Cy3Sp/ |
| Sequence-based reagent | Ndc80-2 | This paper | SCRINSHOT detection probe | CAAGTUGACTATTCCUGGATCCA/3Cy3Sp/ |
| Sequence-based reagent | Ndc80-3 | This paper | SCRINSHOT detection probe | GTCTTGCAAUCTGTTCATUTAGTGC/3Cy3Sp/ |
| Sequence-based reagent | Spc25-Det-1 | This paper | SCRINSHOT detection probe | TCTATCATCUAGGTCCCAUGTTTAGG/3Cy5Sp/ |
| Sequence-based reagent | Spc25-Det-2 | This paper | SCRINSHOT detection probe | AGGTAATCUCTATACAGAUCCGCTG/3Cy5Sp/ |
| Sequence-based reagent | Spc25-Det-3 | This paper | SCRINSHOT detection probe | GTTAGCAGUGGAAATGGUTTCC/3Cy5Sp/ |
Data availability
All sequencing data produced in this study has been submitted to Gene Expression Omnibus (GEO accession number: GSE275758).
-
NCBI Gene Expression OmnibusID GSE275758. PRDM16 functions as a co-repressor in the BMP pathway to suppress neural stem cell proliferation.
-
NCBI Sequence Read ArchiveID PRJNA637987. Single cell sequencing of the development mouse brain.
References
-
Fine mapping of the 1p36 deletion syndrome identifies mutation of PRDM16 as a cause of cardiomyopathyAmerican Journal of Human Genetics 93:67–77.https://doi.org/10.1016/j.ajhg.2013.05.015
-
Prdm16 is required for normal palatogenesis in miceHuman Molecular Genetics 19:774–789.https://doi.org/10.1093/hmg/ddp543
-
Adult neurogenesis requires Smad4-mediated bone morphogenic protein signaling in stem cellsThe Journal of Neuroscience 28:434–446.https://doi.org/10.1523/JNEUROSCI.4374-07.2008
-
Dpp-responsive silencers are bound by a trimeric Mad-Medea complexThe Journal of Biological Chemistry 280:36158–36164.https://doi.org/10.1074/jbc.M506882200
-
Wnt signaling and forebrain developmentCold Spring Harbor Perspectives in Biology 4:a008094.https://doi.org/10.1101/cshperspect.a008094
-
Transcriptional control by the SMADsCold Spring Harbor Perspectives in Biology 8:a022079.https://doi.org/10.1101/cshperspect.a022079
-
Bone morphogenetic proteins: multifunctional regulators of vertebrate developmentGenes & Development 10:1580–1594.https://doi.org/10.1101/gad.10.13.1580
-
Antagonism between Prdm16 and Smad4 specifies the trajectory and progression of pancreatic cancerThe Journal of Cell Biology 222:e202203036.https://doi.org/10.1083/jcb.202203036
-
Crosstalk between Wnt and bone morphogenic protein signaling: a turbulent relationshipDevelopmental Dynamics 239:16–33.https://doi.org/10.1002/dvdy.22009
-
The choroid plexuses and their impact on developmental neurogenesisFrontiers in Neuroscience 8:340.https://doi.org/10.3389/fnins.2014.00340
-
The choroid plexus and cerebrospinal fluid: emerging roles in development, disease, and therapyThe Journal of Neuroscience 33:17553–17559.https://doi.org/10.1523/JNEUROSCI.3258-13.2013
-
Development of the lateral ventricular choroid plexus in a marsupial, Monodelphis domesticaCerebrospinal Fluid Research 7:16.https://doi.org/10.1186/1743-8454-7-16
-
Development and functions of the choroid plexus-cerebrospinal fluid systemNature Reviews. Neuroscience 16:445–457.https://doi.org/10.1038/nrn3921
-
MEME-ChIP: motif analysis of large DNA datasetsBioinformatics 27:1696–1697.https://doi.org/10.1093/bioinformatics/btr189
-
TGF-beta signal transductionAnnual Review of Biochemistry 67:753–791.https://doi.org/10.1146/annurev.biochem.67.1.753
-
Peak calling by Sparse Enrichment Analysis for CUT&RUN chromatin profilingEpigenetics & Chromatin 12:42.https://doi.org/10.1186/s13072-019-0287-4
-
BMP signalling in craniofacial developmentThe International Journal of Developmental Biology 50:511–521.https://doi.org/10.1387/ijdb.052101xn
-
Bone morphogenetic protein signaling in limb outgrowth and patterningDevelopment, Growth & Differentiation 49:455–468.https://doi.org/10.1111/j.1440-169X.2007.00946.x
-
Generation of a multipurpose Prdm16 mouse allele by targeted gene trappingDisease Models & Mechanisms 10:909–922.https://doi.org/10.1242/dmm.029561
-
SKI and MEL1 cooperate to inhibit transforming growth factor-beta signal in gastric cancer cellsThe Journal of Biological Chemistry 284:3334–3344.https://doi.org/10.1074/jbc.M808989200
-
PANTHER: Making genome-scale phylogenetics accessible to allProtein Science 31:8–22.https://doi.org/10.1002/pro.4218
-
The Ndc80 complex: integrating the kinetochore’s many movementsChromosome Research 19:377–391.https://doi.org/10.1007/s10577-010-9180-5
-
Role of bone morphogenetic proteins in cardiac differentiationCardiovascular Research 74:244–255.https://doi.org/10.1016/j.cardiores.2006.11.022
-
PRDM16/MEL1: a novel Smad binding protein expressed in murine embryonic orofacial tissueBiochimica et Biophysica Acta 1773:814–820.https://doi.org/10.1016/j.bbamcr.2007.03.016
-
BMP4 sufficiency to induce choroid plexus epithelial fate from embryonic stem cell-derived neuroepithelial progenitorsThe Journal of Neuroscience 32:15934–15945.https://doi.org/10.1523/JNEUROSCI.3227-12.2012
-
A conserved activation element in BMP signaling during Drosophila developmentNature Structural & Molecular Biology 17:69–76.https://doi.org/10.1038/nsmb.1715
-
Choroid plexus tumoursBritish Journal of Cancer 87:1086–1091.https://doi.org/10.1038/sj.bjc.6600609
-
Smad3-Smad4 and AP-1 complexes synergize in transcriptional activation of the c-Jun promoter by transforming growth factor betaMolecular and Cellular Biology 19:1821–1830.https://doi.org/10.1128/MCB.19.3.1821
-
BMP signaling and stem cell regulationDevelopmental Biology 284:1–11.https://doi.org/10.1016/j.ydbio.2005.05.009
Article and author information
Author details
Funding
Vetenskapsrådet (2020-03543)
- Qi Dai
Cancerfonden (CAN 2017/529)
- Qi Dai
Cancerfonden (20 1046 PjF 01 H)
- Qi Dai
Australian Research Council Centre of Excellence for the Mathematical Analysis of Cellular Systems (CE230100001)
- Jiayu Wen
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank the animal experimental core facility and the imaging facility of Stockholm University and Bioinformatics and Expression Analysis Core Facility at Karolinska Institute, Sweden, for their service and support. We thank Christos Samakovlis and his lab members for technical help on SCRINSHOT experiments. We also appreciate technical help from Adrian Martinez Martin. JW is funded by the Australian Research Council Centre of Excellence for the Mathematical Analysis of Cellular Systems (CE230100001) and ANU Future Scheme. The project was supported by the research project grant from Swedish Research Council (Vetenskapsrådet, 2020–03543) and the research grants from Swedish Cancerfonden (CAN 2017/529 and 20 1046 PjF 01 H) to QD.
Ethics
All animal work was performed in strict accordance with the ethic permit approved by Swedish Board of Agriculture (Dnr 11553-2017 and 11766-2022).
Version history
- Preprint posted:
- Sent for peer review:
- Reviewed Preprint version 1:
- Reviewed Preprint version 2:
- Version of Record published:
Cite all versions
You can cite all versions using the DOI https://doi.org/10.7554/eLife.104076. This DOI represents all versions, and will always resolve to the latest one.
Copyright
© 2025, He et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,179
- views
-
- 79
- downloads
-
- 2
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 1
- citation for umbrella DOI https://doi.org/10.7554/eLife.104076
-
- 1
- citation for Version of Record https://doi.org/10.7554/eLife.104076.3
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
PRDM16 functions as a co-repressor in the BMP pathway to suppress neural stem cell proliferation
eLife 14:RP104076.
https://doi.org/10.7554/eLife.104076.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}