Hawkmoths evaluate scenting flowers with the tip of their proboscis
- Max-Planck Institute for Chemical Ecology, Germany
Figures
Figure 1 with 1 supplement
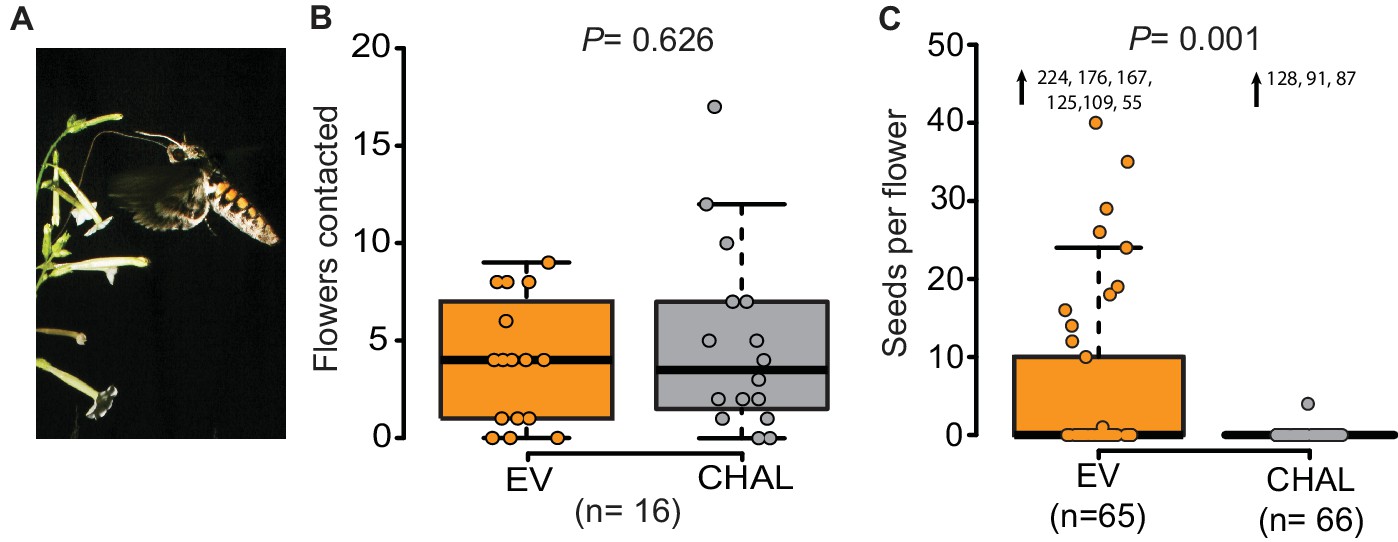
Even though M. sexta visited the same number of benzyl acetone (BA)-emitting (EV) and non-emitting flowers (CHAL), BA-emitting flowers received superior pollination services, increasing seed production.
(A) M. sexta feeding from N. attenuata flowers. (B) Number of EV and CHAL flowers visited per moth on each foraging flight when 10 randomly placed plants per line were presented in a two-choice, free-flight tent assay (Wilcoxon signed rank test). (C) Seeds matured per antherectomized flower after visitations by a moth experimentally loaded with pollen (Wilcoxon rank sum test). Extreme values are shown as numbers.
Figure 1—figure supplement 1
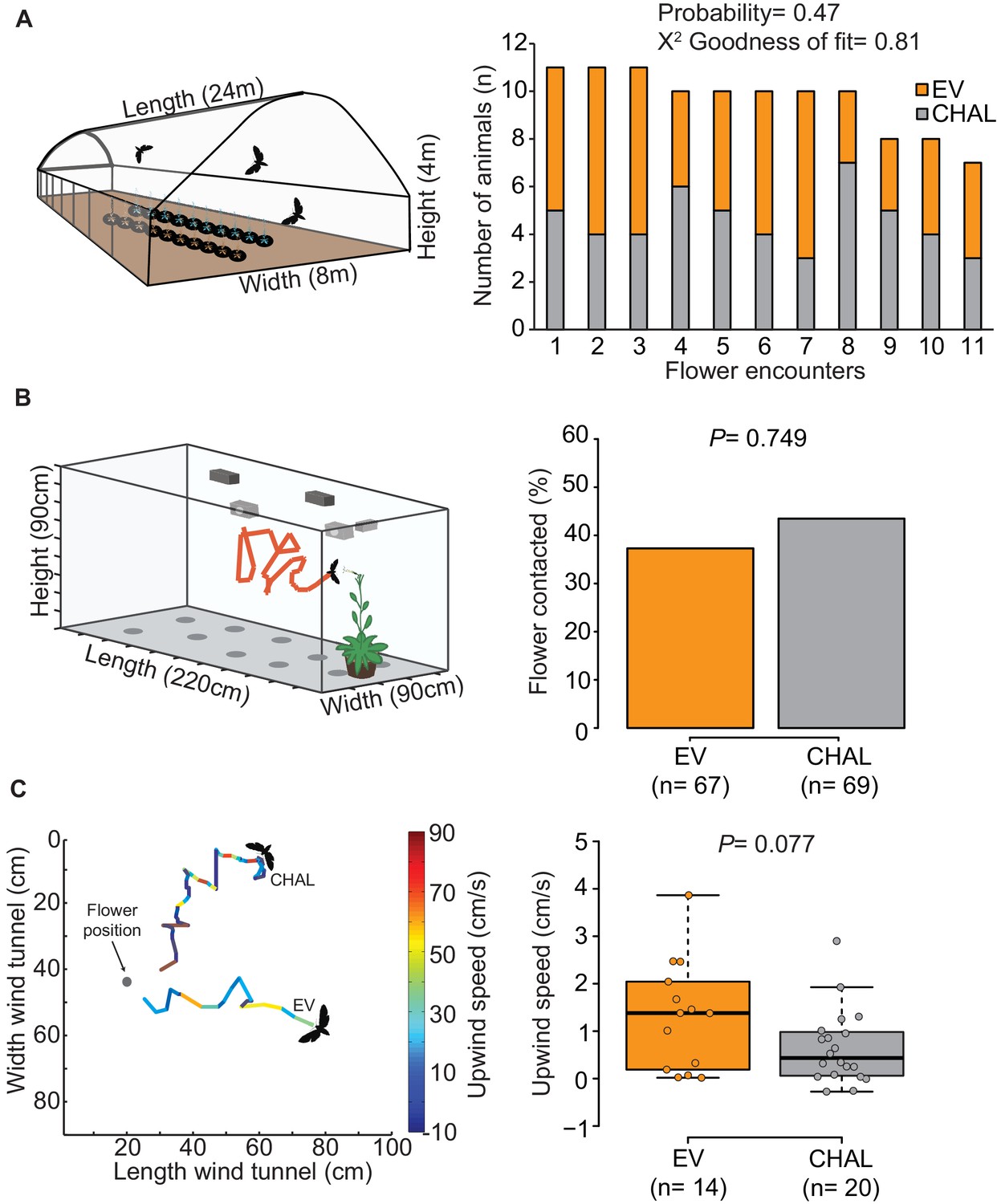
Free flight response of M. sexta towards flowers emitting and not emitting BA.
(A) In a dual-choice assay in a free flight tent, moths visited emitting and non-emitting flowers in a random sequence (probability of two subsequent visits at the same plant type= 0.47). EV and CHAL plants were arranged into two lanes of ten plants each; with positions altered in between different moths. (B) Moths visited a similar percentage of emitting and non-emitting flowers in a no-choice wind tunnel test. (Fisher’s exact test, P = 0.7493)). (C) Top view, 2D representation of a 3D video tracking from two example flights approaching either an EV or a CHAL flower. Flight tracks are color-coded regarding the upwind speed of the moth. The analyses revealed a tendency for a higher upwind speed towards EV flowers emitting BA (Wilcoxon rank sum test, P = 0.077).
Figure 2 with 1 supplement
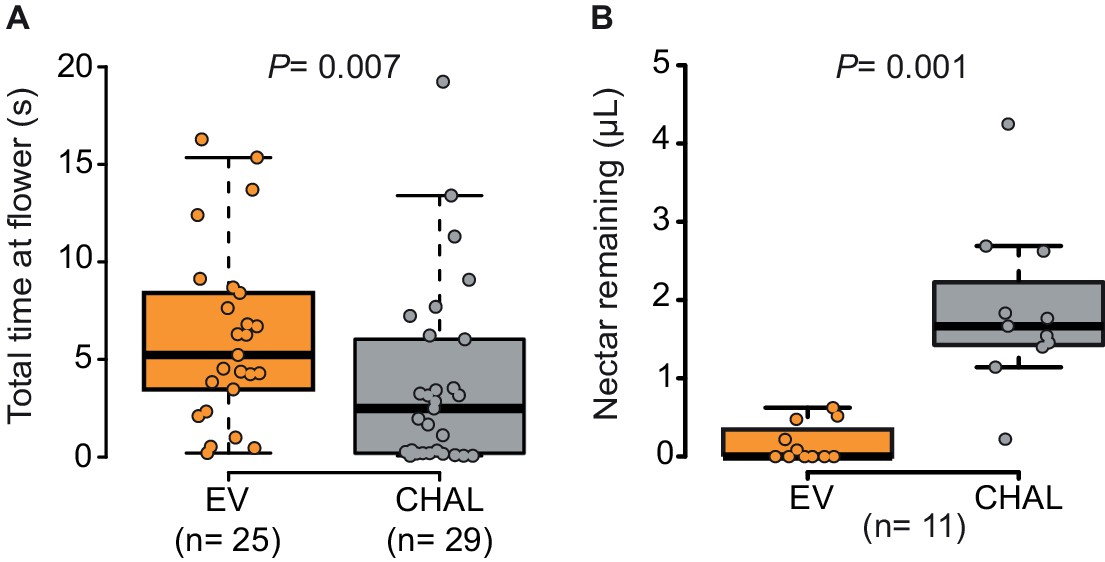
Moths spent more time and removed more nectar from BA-emitting (EV) flowers than from scentless (CHAL) flowers.
(A) Time spent by moths at single flowers (Wilcoxon rank sum test) in a wind tunnel assay. (B) Nectar remaining in flowers after moths attempted to feed in a two choice tent assay (Wilcoxon signed rank test).
Figure 2—figure supplement 1
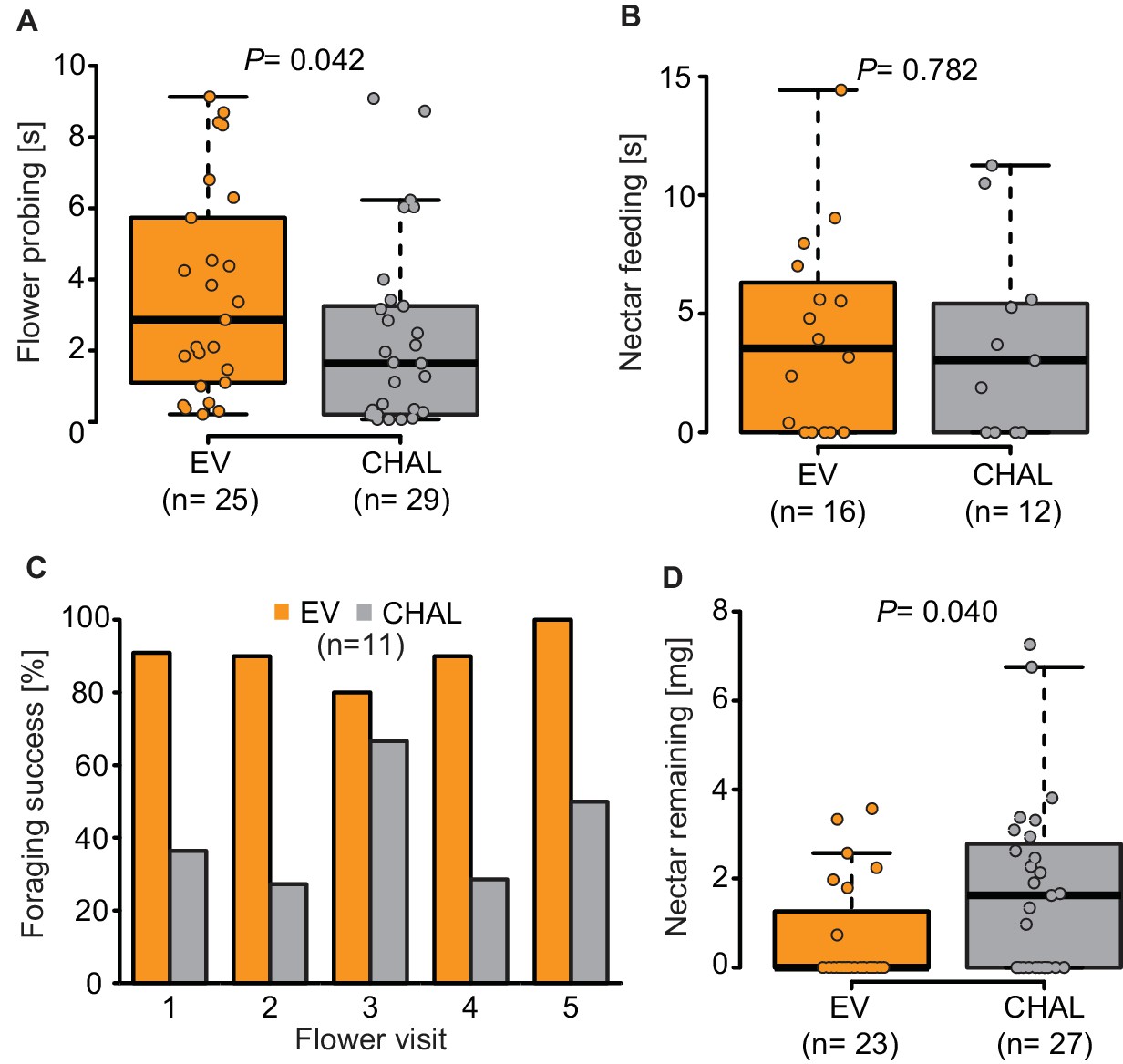
Behavior of M. sexta while foraging on EV and CHAL.
(A) Moth spent significantly more time probing EV flowers in comparison to CHAL flowers, (B) but did not differ in their time feeding from the different flowers. (C) Moth maintained their initial percentage of success full flower visits also over several flower visits. (D) Moth removed significantly less nectar from CHAL plants in a no-choice wind tunnel assay. Wilcoxon signed rank test was used in all cases.
Figure 3 with 1 supplement
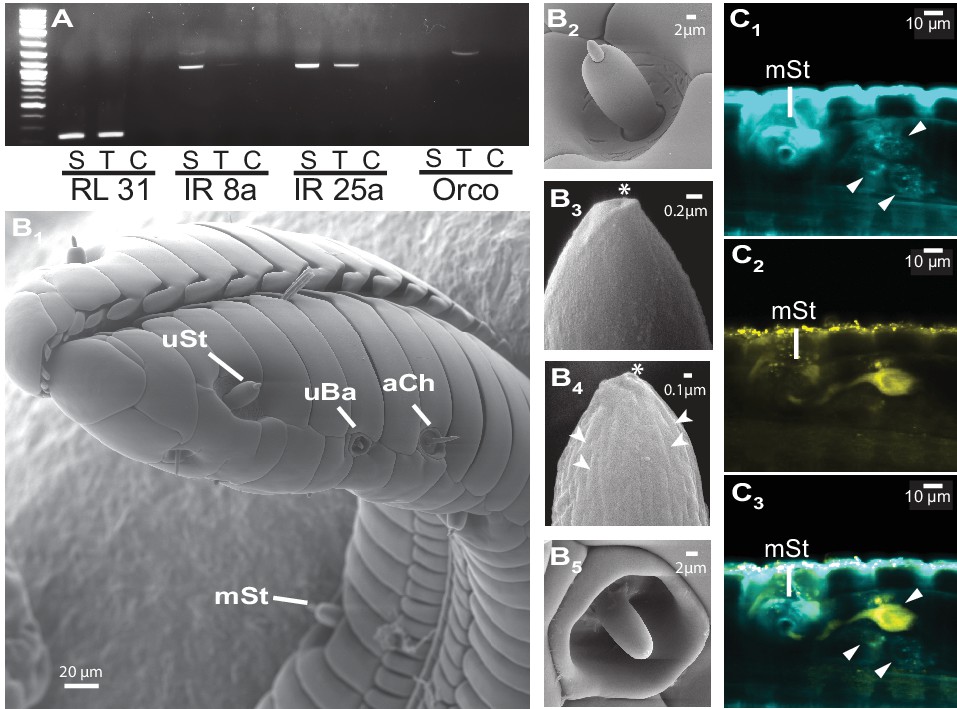
The M. sexta proboscis harbors sensilla, which house sensory neurons expressing olfactory genes.
(A) Reverse transcription PCR using either the proboscis shaft (S), the first centimeter of the proboscis tip (T) or a water control (C) with primers for the three major olfactory co-receptors (IR8a, IR25a, Orco). The transcripts of the ribosomal gene RL131 and water were included as positive and negative control. (B1) Scanning electron microscopy images of the M. sexta proboscis tip show three types of potential chemosensory sensilla: sensilla styloconica (B2) with a uniporous (uSt (B3)) and multiporous (mSt (B4)) cone and uniporous sensilla basiconica (uBa (B5, Figure 3—figure supplement 1B)) as well as aporouse sensilla chaetica (aCh, Figure 3—figure supplement 1A). Asterisks mark tip pore, arrowheads indicate side pores. Neuronal labeling using anti-bodies against horseradish peroxidase (C1) and against Orco(C2) indicate three neurons close to the first mSt sensillum (arrows), of which one expresses the olfactory co-receptor Orco (C3, Figure 3—figure supplement 1).
Figure 3—figure supplement 1
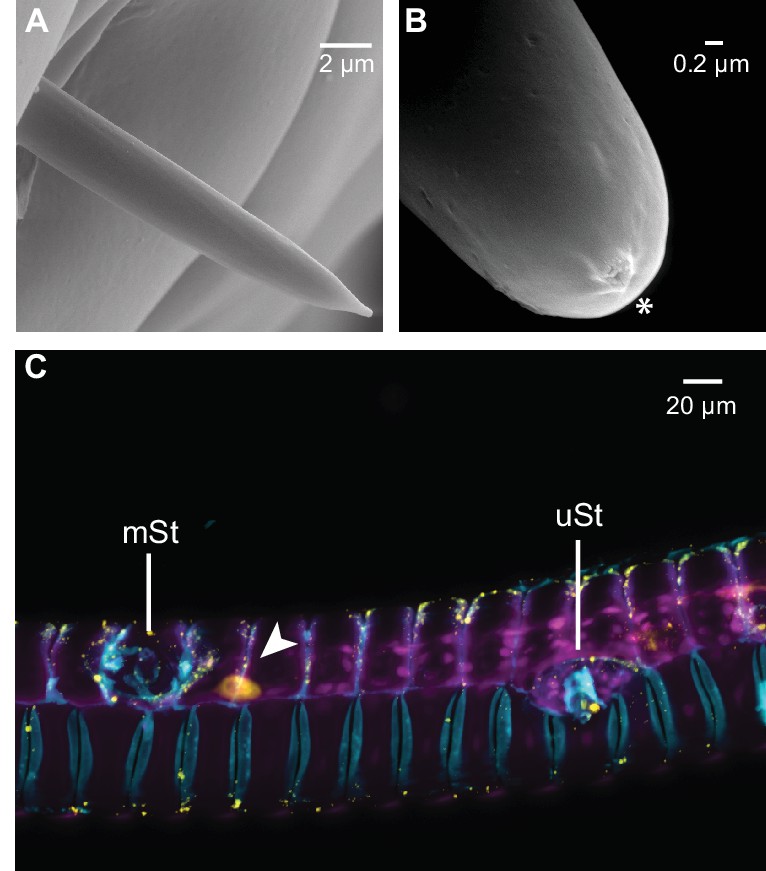
Different sensillum types were found in tip region of the M. sexta proboscis, but only multiporous sensilla styloconica (mSt) were determined to be Orco positive.
(A) Aporouse sensillum chaeticum (B) tip of a uniporous sensillum basiconicum (uBa, Figure 3b5) asterisk indicates tip pore. (C) Proboscis section showing the first multiporous sensillum stylochonicum (mSt) and a single Orco-positive cell body in close proximity (arrowhead) as well as a uniporous senillum stylochinicum (uSt) without Orco-positive staining.
Figure 4 with 1 supplement
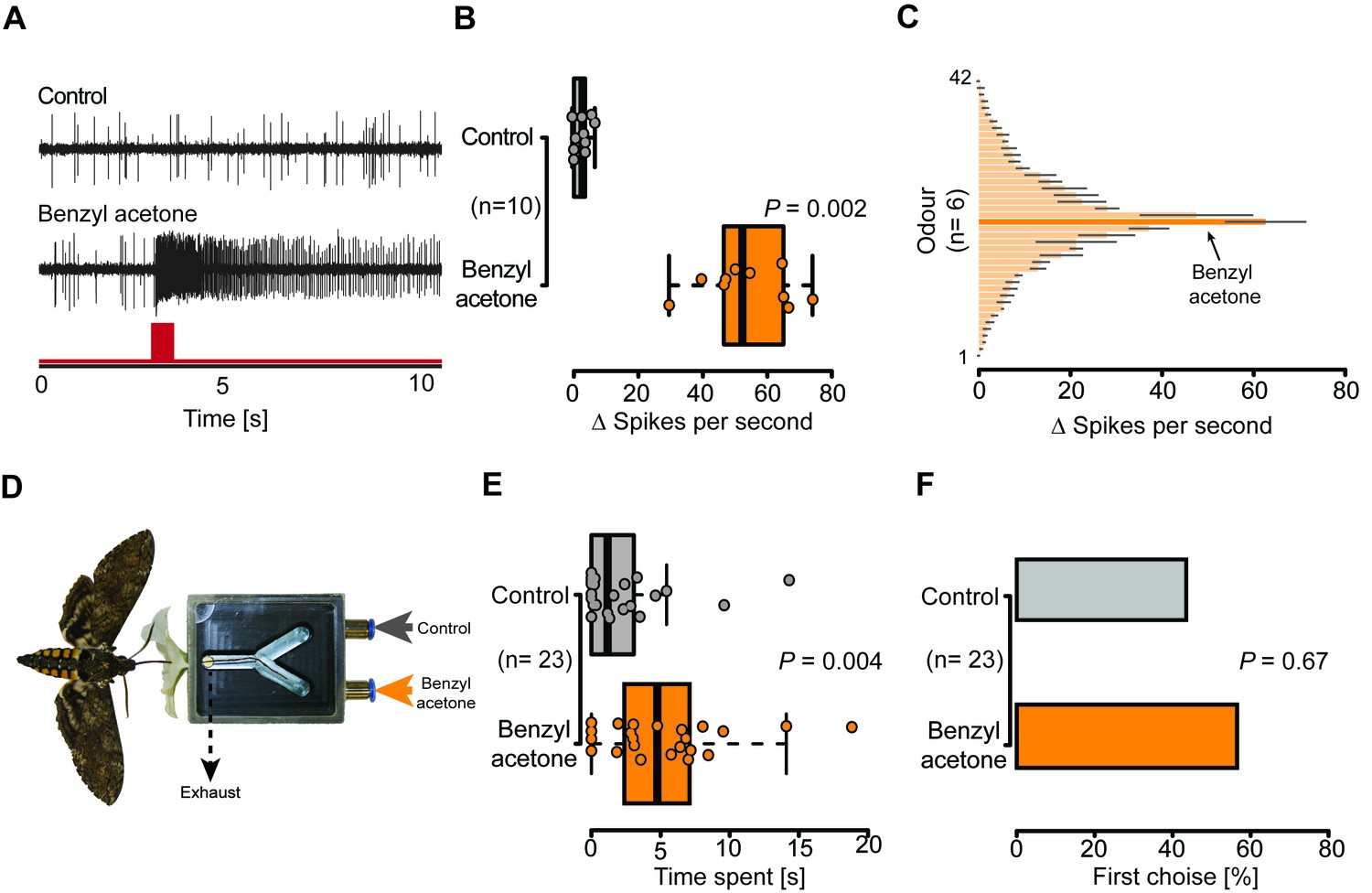
Olfactory sensory neurons housed in proboscis sensilla respond to BA and are sufficient for flower evaluation.
(A) Single sensillum recordings from the first multiporous sensilla styloconica. Upper trace depicts a characteristic response to the water control; lower trace shows a response to BA from the neuron in the same sensillum. Red bar indicates time of stimulus. (B) Boxplot shows ∆ spikes per second recorded from the first mSt when stimulating with water control or BA (0.1 mM) for 0.5 s. Neurons responded with a significantly higher spike rate to BA than to the water control (Wilcoxon signed-rank test). (C) Response profile of the first mSt to 42 different odorants. Black bars indicate S.E.M. Names and spike rate of each odorant can be found in Figure 4C—source data 1. (D) Behavioral assay to test the response to either humidified air with BA (0.1 mM) or humidified air only. Exhaust excludes antennal olfactory input. (E) Moth inserted their proboscis significantly longer into the arm in which BA was present (Wilcoxon signed-rank test). (F) Moths chose equally often between Y-tube arms containing BA-scented air or solvent control at the first approach (Exact binominal test).
-
Figure 4—source data 1
Response spectrum of the first multpourus sensillum to ecological relevant odors.
- https://doi.org/10.7554/eLife.15039.011
Figure 4—figure supplement 1
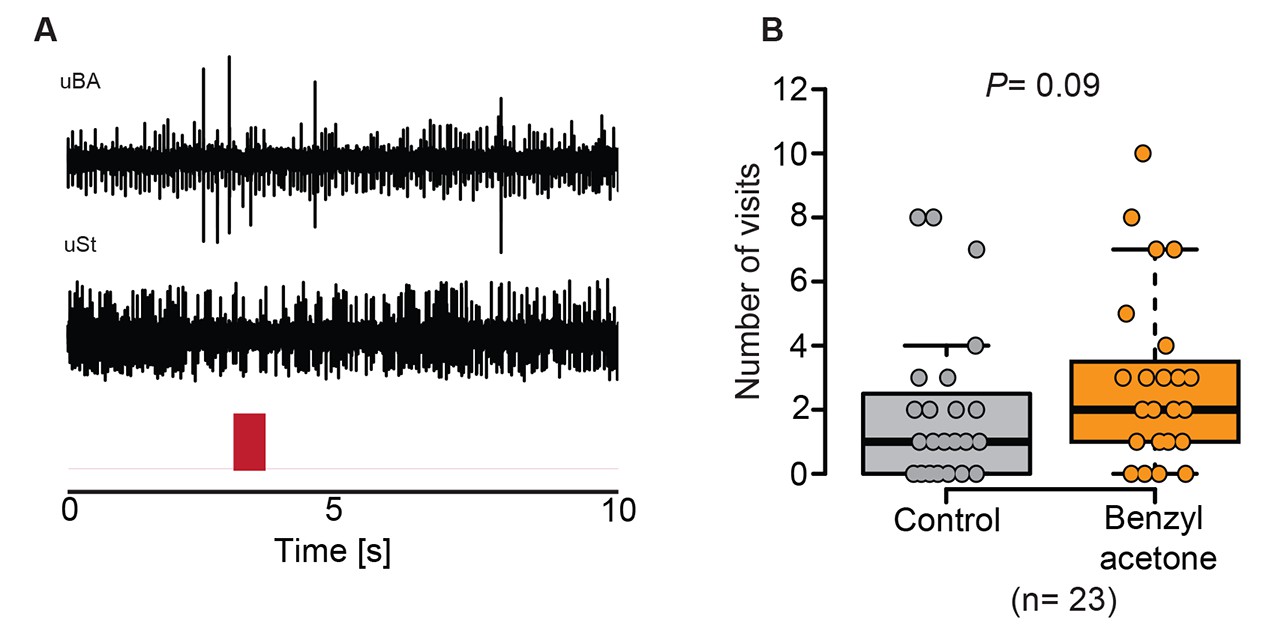
BA does not influence all types of sensilla on the proboscis of M. sexta and does also not enable the moth to navigate actively towards BA.
(A) Example traces of a uniporous sensillum basiconicum (uBa) and a uniporous sensillum stylochonicum (uSt) stimulated with the headspace from a 0.1 mM BA suspension, showing no response towards the stimulus. (B) Moths entered the arm containing the solvent control as often as they entered the arm containing BA-scented air (Wilcoxon signed rank test).
Videos
Video 1
M. sexta foraging on EV flowers emitting BA in a free-flight tent.
https://doi.org/10.7554/eLife.15039.004
Video 2
M. sexta attempting to forage on Chal flowers not emitting BA in a free-flight tent.
https://doi.org/10.7554/eLife.15039.007
Video 3
Y-maze of the proboscis choice assay, BA-scented air is provided at the right arm and the solvent control in the left arm.
Odors were removed by applying a vacuum at the entrance of the Y-maze.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Hawkmoths evaluate scenting flowers with the tip of their proboscis
eLife 5:e15039.
https://doi.org/10.7554/eLife.15039




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}