A mitochondrial DNA hypomorph of cytochrome oxidase specifically impairs male fertility in Drosophila melanogaster
- Fred Hutchinson Cancer Research Center, United States
- Howard Hughes Medical Institute, United States
- Vanderbilt University, United States
- University of Michigan, United States
- University of Washington, United States
- University of Washington Medical Center, United States
Figures
Figure 1 with 1 supplement
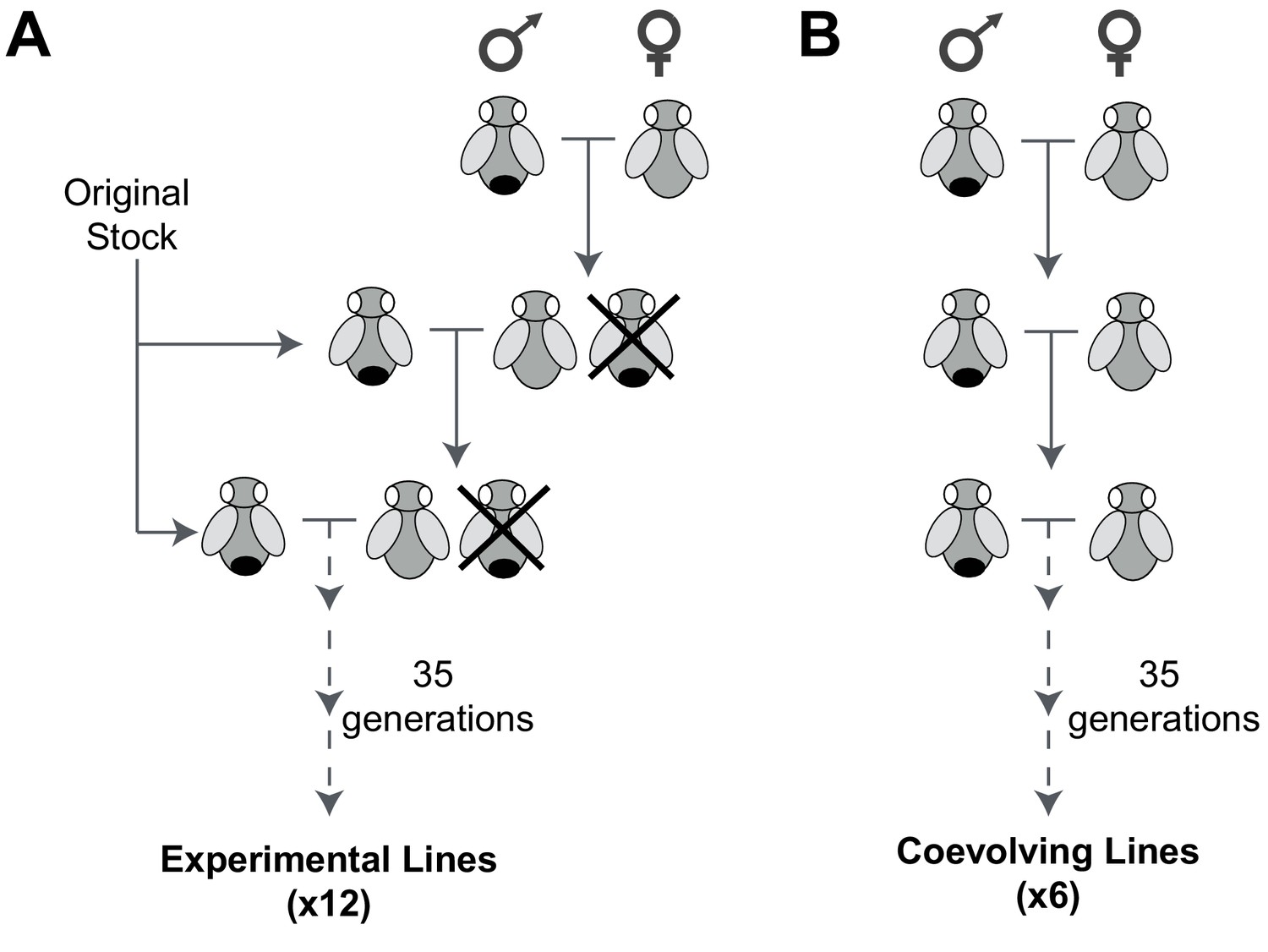
An experimental evolution strategy to recover male-harming mtDNA mutations.
(A) In 12 D. melanogaster lines undergoing experimental evolution, 300 virgin female progeny in every generation were prevented from mating with sibling males (shown as being ‘crossed out’) and instead were mated with 100 males from the original stock. (B) In six lines undergoing coevolution, we allowed 300 virgin female and 100 male siblings to mate in every generation. Crosses for the 12 experimental and six coevolving lines were carried out for 35 generations.
Figure 1—figure supplement 1

Status of Wolbachia infection in w1118 derived stocks.
Using PCR for a Wolbachia-specific WSP gene, we find no evidence of Wolbachia infection in the re-isolated stocks of wildtype and COIIG177S mtDNA. As a positive control for Wolbchia infection, we used the cinnabar brown (cnbw) strain of D. melanogaster.
Figure 2
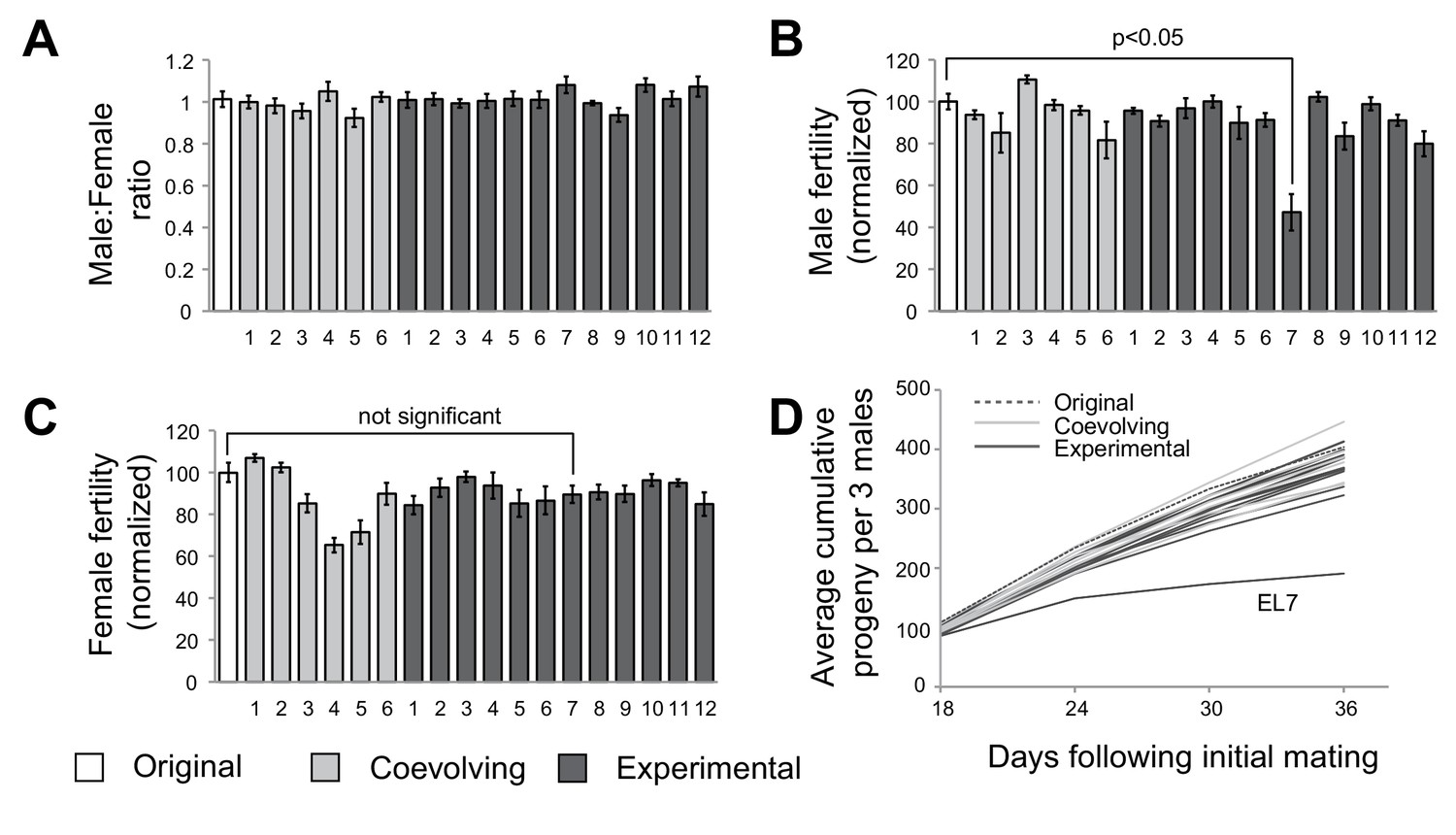
A single experimental line with male-specific fertility defects.
(A) We measured average sex ratio of progeny by mating five females from each of the original, experimental, or coevolving lines, with three males from the original stock. Average sex ratio was found to be close to 50:50 in each of the lines, suggesting that there were no gross viability differences between male and female progeny in any of the lines. All error bars represent standard error of the mean. All experiments were done in replicates of 10 per group. (B) We measured male fertility by mating three males from each of the original stock, experimental, or coevolving lines with five females from the original stock, and calculating average number of resulting progeny. Male fertility is represented as a normalized percentage of progeny relative to the original stock. Only one line, experimental line 7 (EL7) showed significant reduction relative to the others. All error bars represent standard error of the mean. All experiments were done in replicates of 10 per group. (C) We measured female fertility by mating five females from each of the original, experimental, or coevolving lines with three males from the original stock, and calculating average number of resulting progeny. Like in (B), female fertility is represented as a normalized percentage of progeny relative to the original stock. EL7 female fertility is not significantly different from the original stock. All error bars represent standard error of the mean. All experiments were done in replicates of 10 per group. (D) To gain further insight into altered male fertility in EL7 (B), we plotted the cumulative number of progeny sired by (three) males as a function of time since initial mating (10 replicates per line). We find that the cumulative number of progeny sired by EL7 males is normal until day 18 but is subsequently significantly lower than for all other lines. This finding suggests an age-dependent decline in male fertility in the EL7 line.
Figure 3 with 3 supplements

A single missense mutation (G177S) in subunit II of cytochrome c oxidase underlies lower male fertility in EL7.
(A) The five complexes of the electron transport chain are schematized. These complexes are comprised of subunits encoded by both nuclear and mitochondrial genomes; the latter are indicated in pink. Subunit II of cytochrome c oxidase (COII) with the glycine to serine mutation at position 177 (G177S) in Experimental Line 7 is highlighted in bright pink. (B) Crystal structure of Bos taurus COII (in gold) in complex with COI (in blue) (PDB number: 2OCC). Glycine with its side chains at position 177 (G177) in COII is indicated. (C) Partial amino acid sequence alignment of COII from representative animal species highlights the conserved glycine residue at position 177 (boxed in yellow).
Figure 3—figure supplement 1
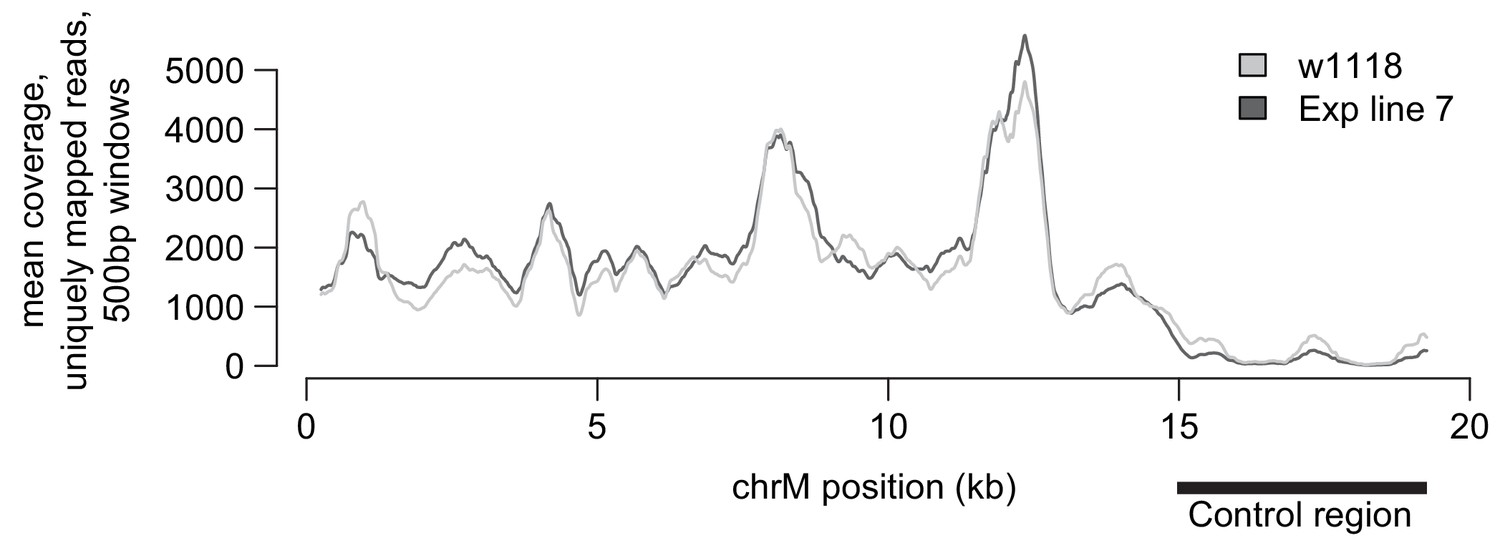
Whole genome mtDNA sequencing coverage.
Coverage of the mtDNA genome sequencing from the original w1118 stock compared to the EL7 strain. With the exception of the AT-rich ‘control region’ (which has relatively modest coverage), the rest of the mtDNA has good coverage in both the original w1118 and the EL7 strains, allowing unambiguous detection of mutations.
Figure 3—figure supplement 2

COIIG177S is present in many experimental and coevolving lines.
(A) 15 females from each of the coevolving, and (B) experimental lines were pooled. From these, DNA was extracted and the COII gene was PCR amplified and sequenced. Sequence traces of the pooled PCR products highlight the nucleotide position with G to A mutation responsible for the G177S mutation in COII (indicated by a vertical line). These reveal a qualitative measure of the degree of heteroplasmy at G177S.
Figure 3—figure supplement 3
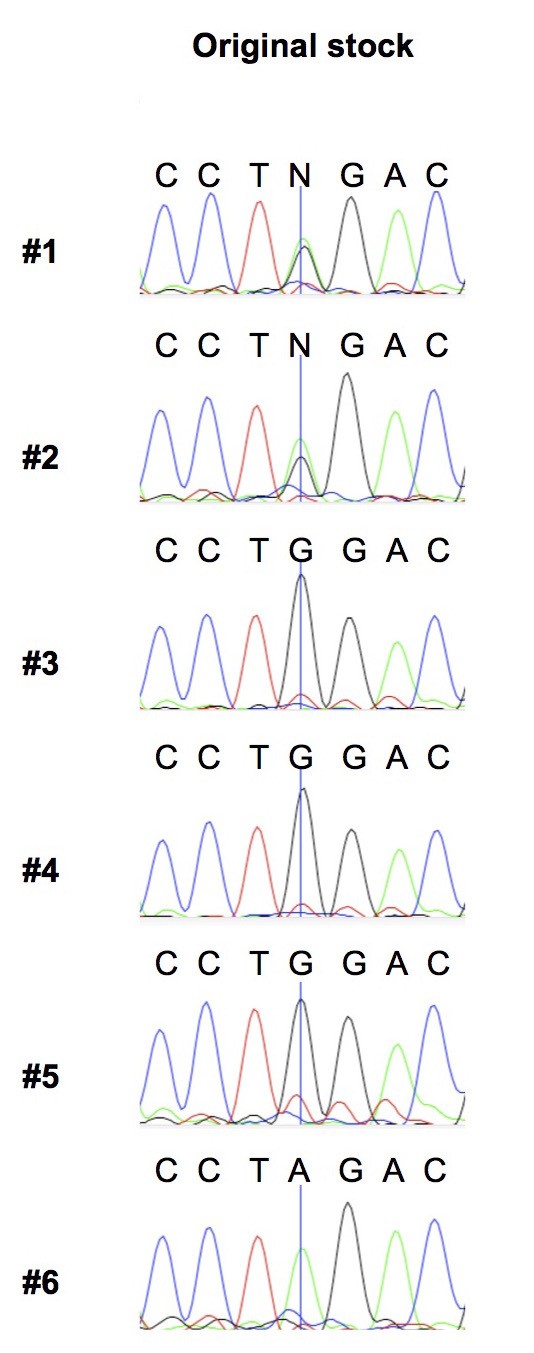
COIIG177S is present at variable levels in flies of the original w1118 stock.
We extracted DNA from 6 individual females from the original w1118 stock and carried out PCR and sequencing of the COII mtDNA gene. We present sequence traces highlighting the nucleotide position where the G to A mutation in COII occurs (indicated by a vertical line). Note that first two individuals are clearly heteroplasmic.
Figure 4
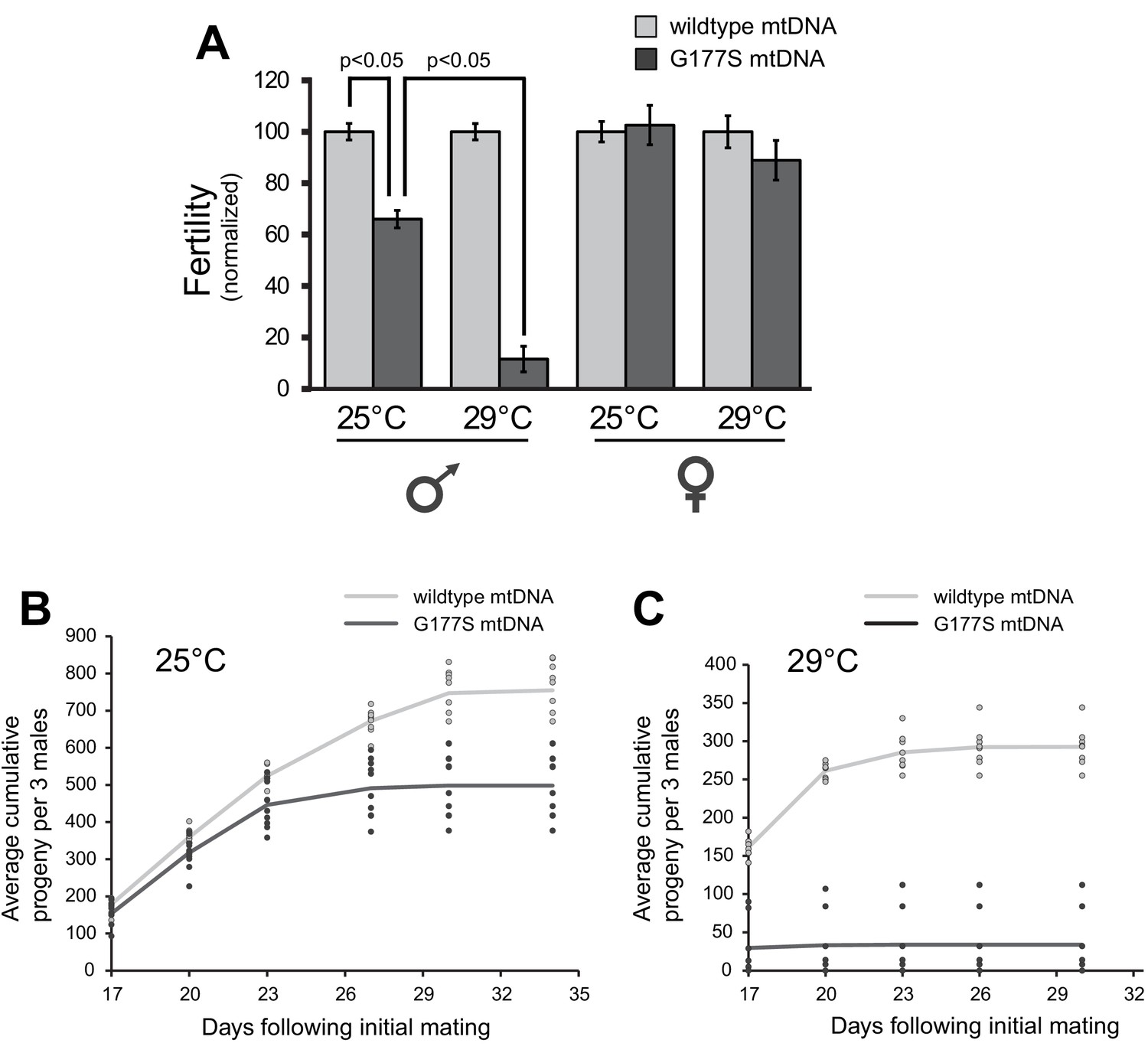
Males with COIIG177S mutation have decreased fertility at elevated temperatures.
(A) To measure male fertility, we compared the fertility of males homoplasmic for either wildtype or COIIG177S mutant mtDNA reestablished from the original stock. For each experiment, three males from homoplasmic stocks were mated with five females homoplasmic for wildtype mtDNA. Male fertility is presented as a normalized percentage of progeny produced, relative to the number produced by wildtype mtDNA males. COIIG177S mutant mtDNA males produce fewer progeny at 25°C, but this is further reduced at 29°C. To measure female fertility, five females from homoplasmic wildtype or COIIG177S mutant mtDNA stocks were mated with three males with wildtype mtDNA. Female fertility is presented as a normalized percentage of progeny produced, relative to wildtype mtDNA females. All error bars represent standard error of the mean. All experiments were done in replicates of 10 per group. (B) Number of progeny sired by wildtype or COIIG177S mutant males at 25°C is cumulatively plotted as a function of time since initial mating (10 replicates per line). Actual number of cumulative progeny from each replicate is indicated by dots at given time points. (C) Number of progeny sired by wildtype or COIIG177S mutant males at 29°C is cumulatively plotted as a function of time since initial mating (10 replicates per line). The reduction in male fertility is more significant at the higher temperature.
-
Figure 4—source data 1
Near homoplasmy of wildtype and COIIG177S mtDNA in re-isolated lines.
Summary of data from duplex sequencing of COIIG177S mutant and wildtype mtDNA strains re-isolated from the ancestral, heteroplasmic w1118 stock. Individual fly heads from the mutant COIIG177S mtDNA stock (Mut Fly 1, 2, 6–10) were subjected to duplex sequencing. Pooled samples represent sequencing data generated by sequencing combined DNA from 10 individuals from either the mutant or wildtype mtDNA strains.
- https://doi.org/10.7554/eLife.16923.011
Figure 5
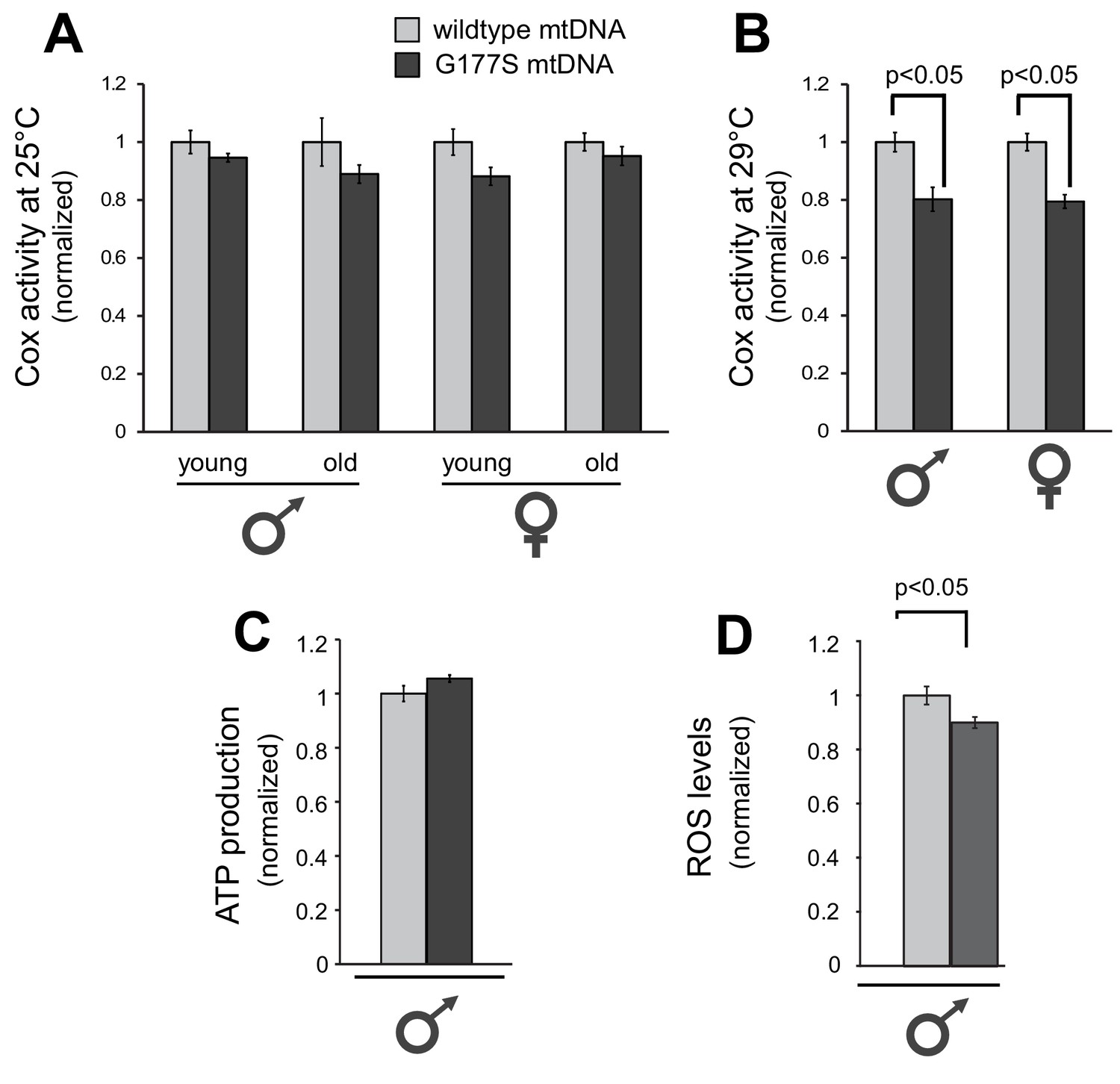
Biochemical consequences of COIIG177S mtDNA mutation.
We measured COX activity from young (3–4 days) or old (21 days) flies raised at 25°C (A), or young flies raised at 29°C (B). (C) We measured ATP levels from 3–4 day old males raised at 29°C. (D) We measured ROS levels from 3–4 day old males raised at 29°C. 4–5 replicates per group for all experiments. Data is normalized to wildtype flies for each group in all experiments. Error bars represent standard error of the mean.
Figure 6 with 2 supplements
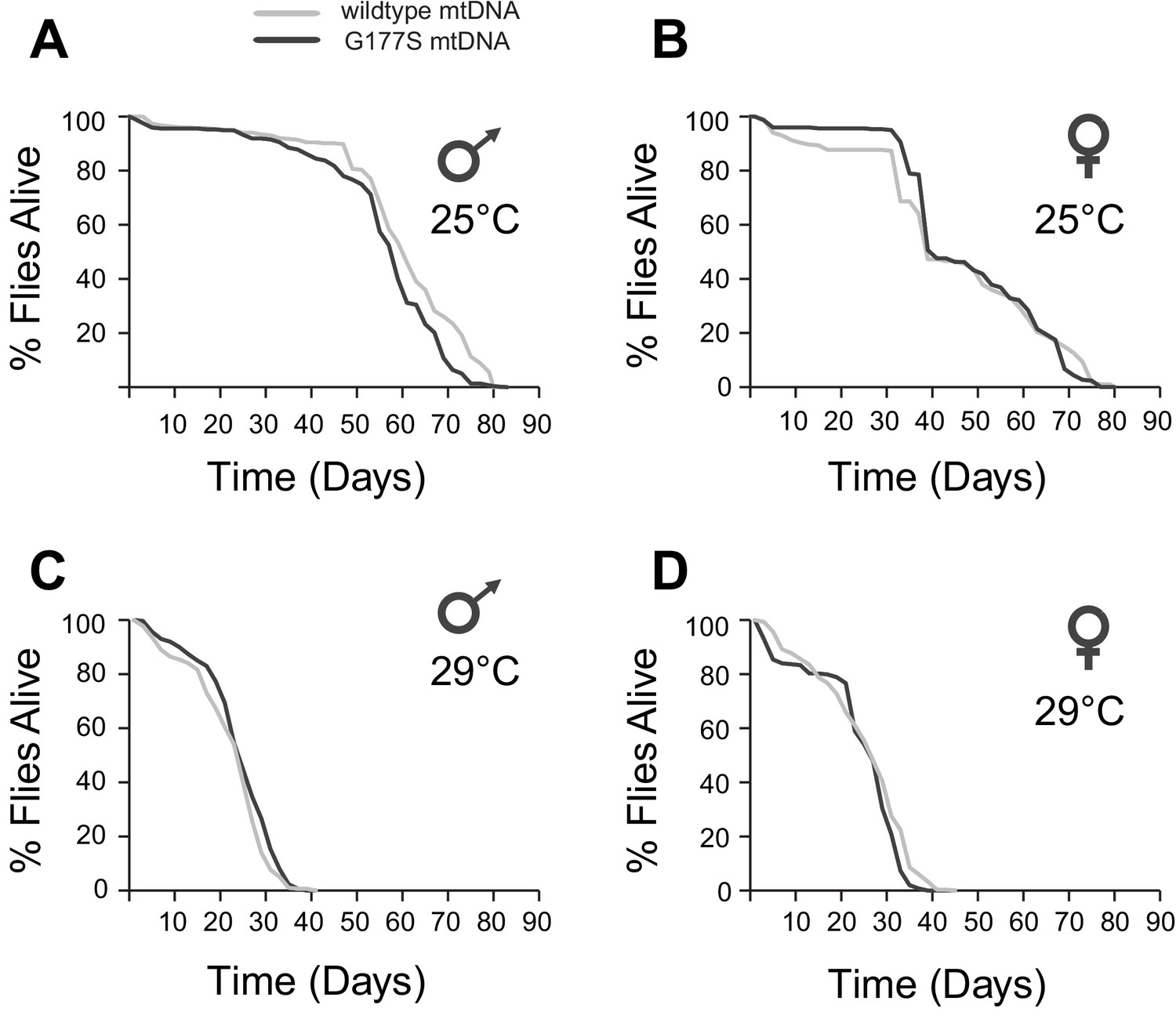
COIIG177S mtDNA mutation does not significantly affect male or female lifespan.
We measured average lifespan of males and females at 25°C (A,B) or 29°C (C,D). N = 100 flies per group.
Figure 6—figure supplement 1

COIIG177S mutation does not affect heat tolerance of flies raised at 29°C.
Time it takes males (A) or females (C) to become paralyzed within allotted time of 6 min upon 39°C heat shock. Time it takes males (C) or females (D) to recover after 39°C heat shock for 6 min. All flies raised at 29°C. Error bars represent standard error of the mean. N = 16 flies per group.
Figure 6—figure supplement 2
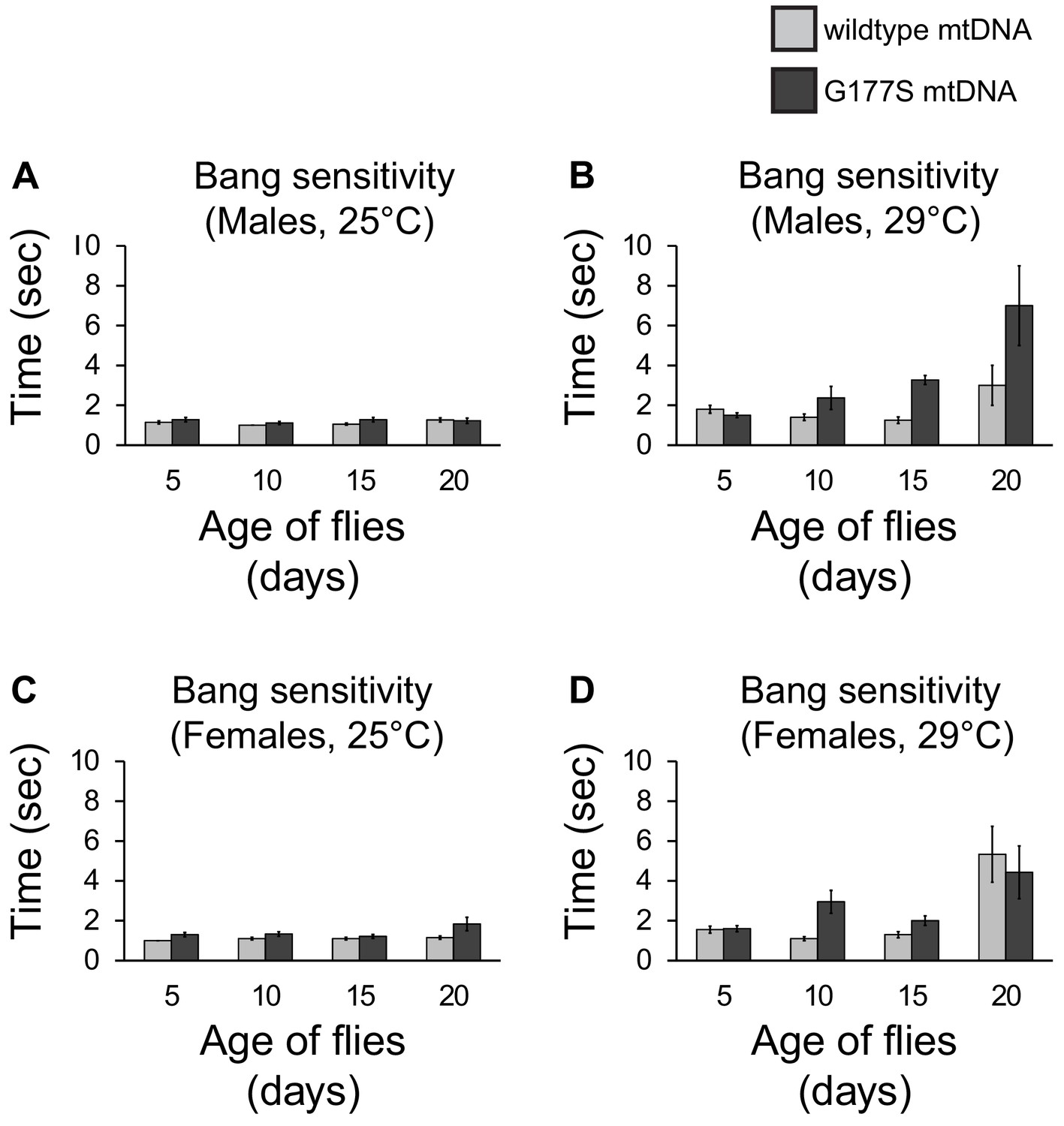
COIIG177S mutation does not affect bang sensitivity.
Amount of time it takes males raised at 25°C (A) or (B) 29°C to right themselves after being vortexed at maximum speed for 10 s. Amount of time it takes females raised at 25°C (C) or (D) 29°C to right themselves after being vortexed at maximum speed for 10 s. Error bars represent standard error of the mean. N = 18–20 flies per group. Note that there is no difference in females carrying wildtype or COIIG177S mutant mtDNA. There is a trend to increased bang sensitivity in males carrying COIIG177S mutant mtDNA (B) but this is not statistically significant.
Figure 7 with 1 supplement
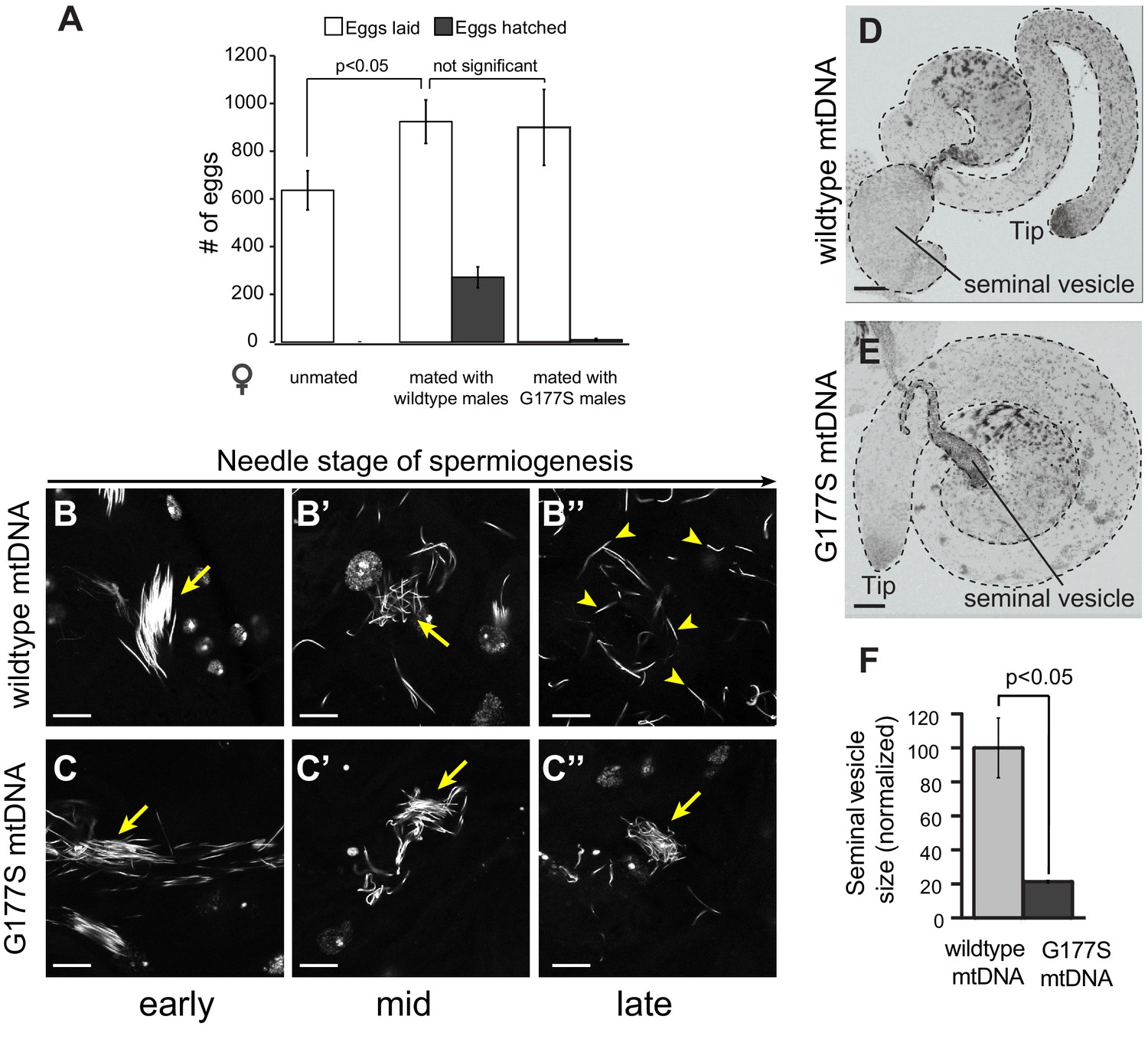
Normal mating but defective sperm development in COIIG177S mutant mtDNA males.
(A) We measured whether the COIIG177S mtDNA mutation affected the mating success of males. We measured eggs laid by virgin wildtype females that were either unmated, or mated with 2–5 day old wildtype, or COIIG177S mutant males at 29°C. We determined the fraction of eggs hatched by counting unhatched eggs and larva 24 hrs after eggs were laid. All experiments were done in replicates of 6 per group. Error bars represent standard error of the mean. Our results show that the number of eggs laid after mating to wildtype mtDNA males is not significantly different from those mated to COIIG177S mutant mtDNA males; in the latter case, most of the eggs are unfertilized and do not hatch. (B,C) We present maximum projection representative images of DAPI stained testis from 2–5 day old wildtype mtDNA male flies grown at 29°C at early (B), middle (B’), and late (B’’) needle stage of sperm development. Note that the sperm are organized during early needle stage (arrow) and then break up into individual sperm by late needle stage (arrowheads). We also present maximum projection images of DAPI stained testis from COIIG177S mutant mtDNA male flies grown at 29°C at early (C), middle (C’), and late (C’’) needle stage of sperm development. Note also that the sperm in COIIG177S mutant are ‘clumped’ and disorganized early in the needle stage and remain so through remainder of spermiogenesis (arrow). Scale bar, 20 μm. (D,E) Representative DAPI stained images of whole testis (outlined in dotted line) from 2–5 day old virgin wildtype mtDNA (D) and COIIG177S mutant mtDNA males (E) raised at 29°C. For orientation, in both images, we identify the tip of the testis (where germ stem cells reside) as well as the seminal vesicle (the storage organ for mature sperm). Note the much smaller seminal vesicle size in the mutant males. Scale bar, 50 μm. (F) Quantification of the seminal vesicle size, as measured by cross-sectional area, normalized to wildtype. Average calculated from 5–7 testes. Error bars represent standard error of the mean.
Figure 7—figure supplement 1
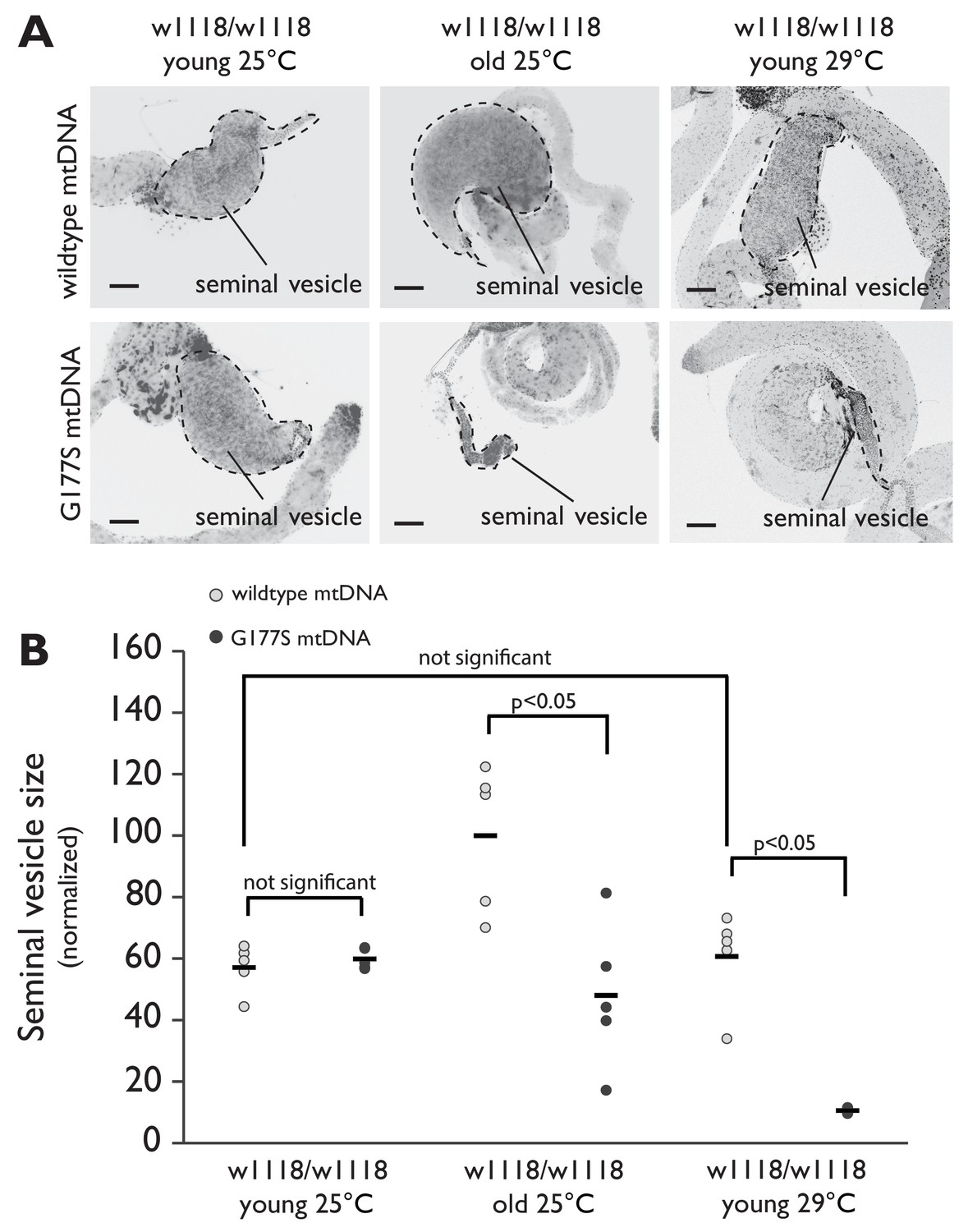
Aged males recapitulate the male sterility defects at 25°C.
(A) Representative DAPI stained images of seminal vesicles (outlined in dotted line) from w1118 males containing either wildtype mtDNA or COIIG177S mutant mtDNA. We analyzed both young virgin males (3–4 day adults) raised at 25°C or 29°C, as well as aged virgin males (21 day old adults) raised at 25°C. Note the much smaller seminal vesicle sizes in COIIG177S mutant mtDNA containing aged males raised at 25°C as well as young males raised at 29°C. Scale bar, 100 μm. (B) Quantification of the seminal vesicle size, as measured by cross-sectional area, normalized to aged wildtype mtDNA males raised at 25°C. Average calculated from 5–7 testes. Error bars represent standard error of the mean.
Figure 8
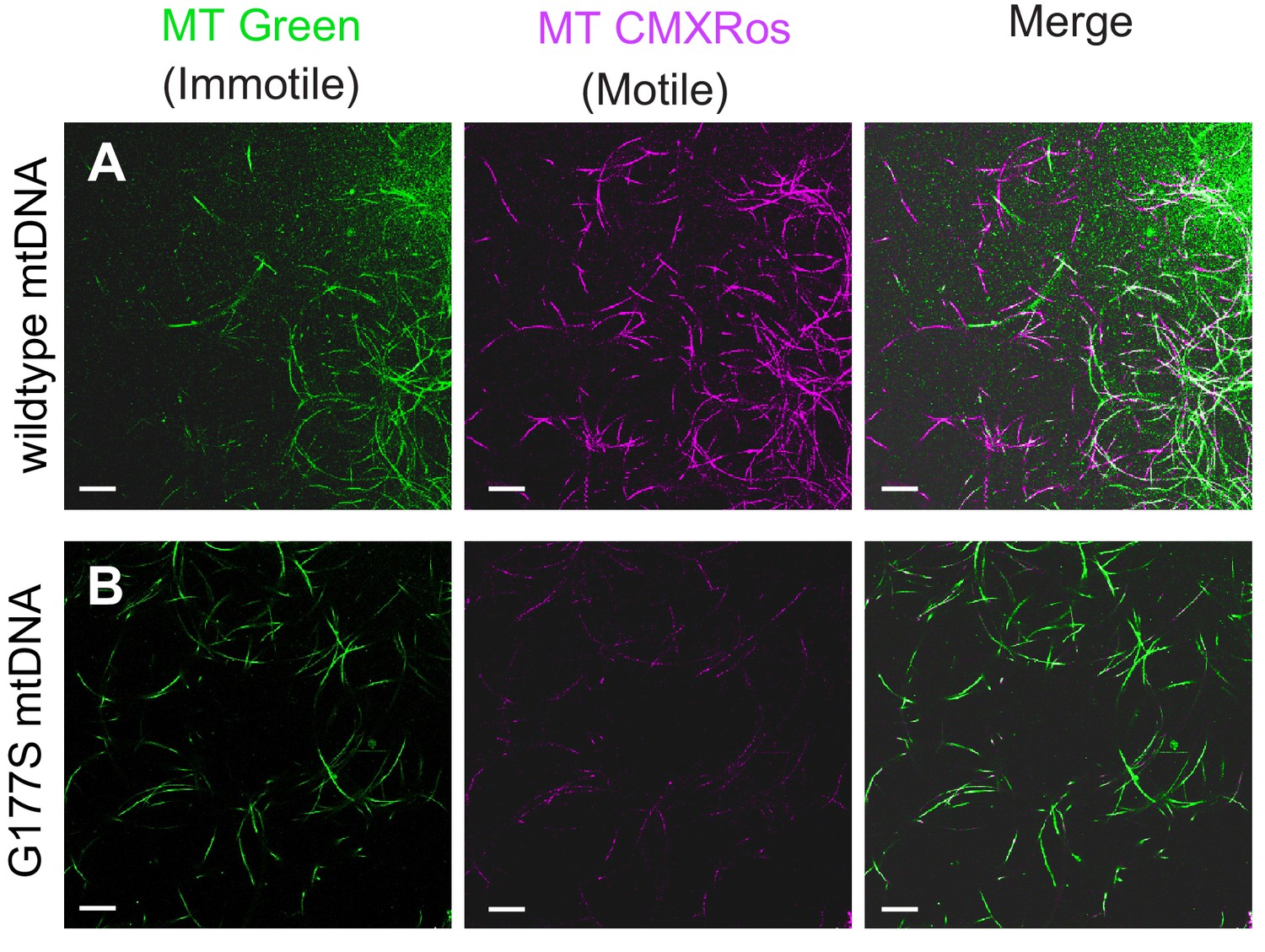
Lower motility of sperm from COIIG177S mutant mtDNA males.
Sperm from wildtype mtDNA (A) and COIIG177S mutant mtDNA (B) males raised at 29°C stained with mitotracker Green (green), which stains immotile sperm and mitotracker CMS Rox (magenta), which stains motile sperm preferentially. Scale bar, 20 μm.
Figure 9 with 1 supplement
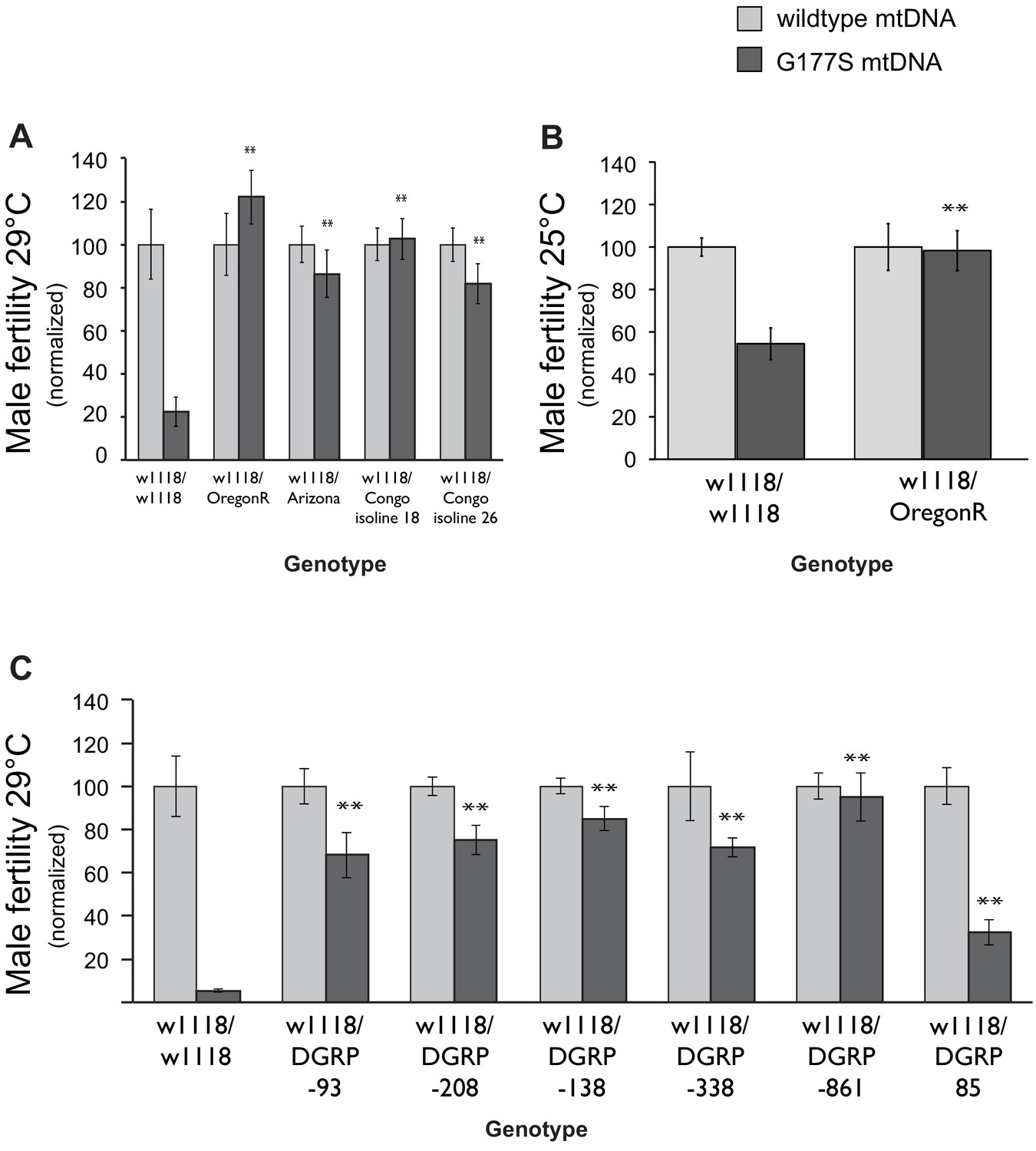
Dominant suppression of COIIG177S associated male fertility defects by diverse nuclear genome backgrounds.
(A) We measured male fertility of heterozygous males derived from crosses of w1118 females (carrying wildtype or COIIG177S mutant mtDNA) to males from a variety of different nuclear backgrounds (i.e., Oregon R, Arizona, Congo isoline 18, Congo isoline 26). In each case, three heterozygous males were mated with 5 w1118 females at 29°C (10 replicates per group). Male fertility was measured as average number of resulting progeny, normalized as a percentage of progeny of corresponding males with wildtype mtDNA. Raw progeny numbers are given in Figure 9—figure supplement 1. Most nuclear backgrounds show mild to complete suppression of the male fertility defects observed in w1118/w118 males carrying COIIG177S mutant mtDNA (**p<0.05). Error bars represent standard error of the mean. (B) We examined lifetime male fertility in the w1118/Oregon R background at 25°C (note that male fertility is generally higher at lower temperatures) to quantify the suppression of male sterility. (C) We measured male fertility of heterozygous males derived from crosses of w1118 females (carrying wildtype or COIIG177S mutant mtDNA) to males from six DGRP strains, at 29°C (10 replicates per group) as in (A). At least five of the tested DGRP strains show partial to complete suppression of male sterility.
Figure 9—figure supplement 1
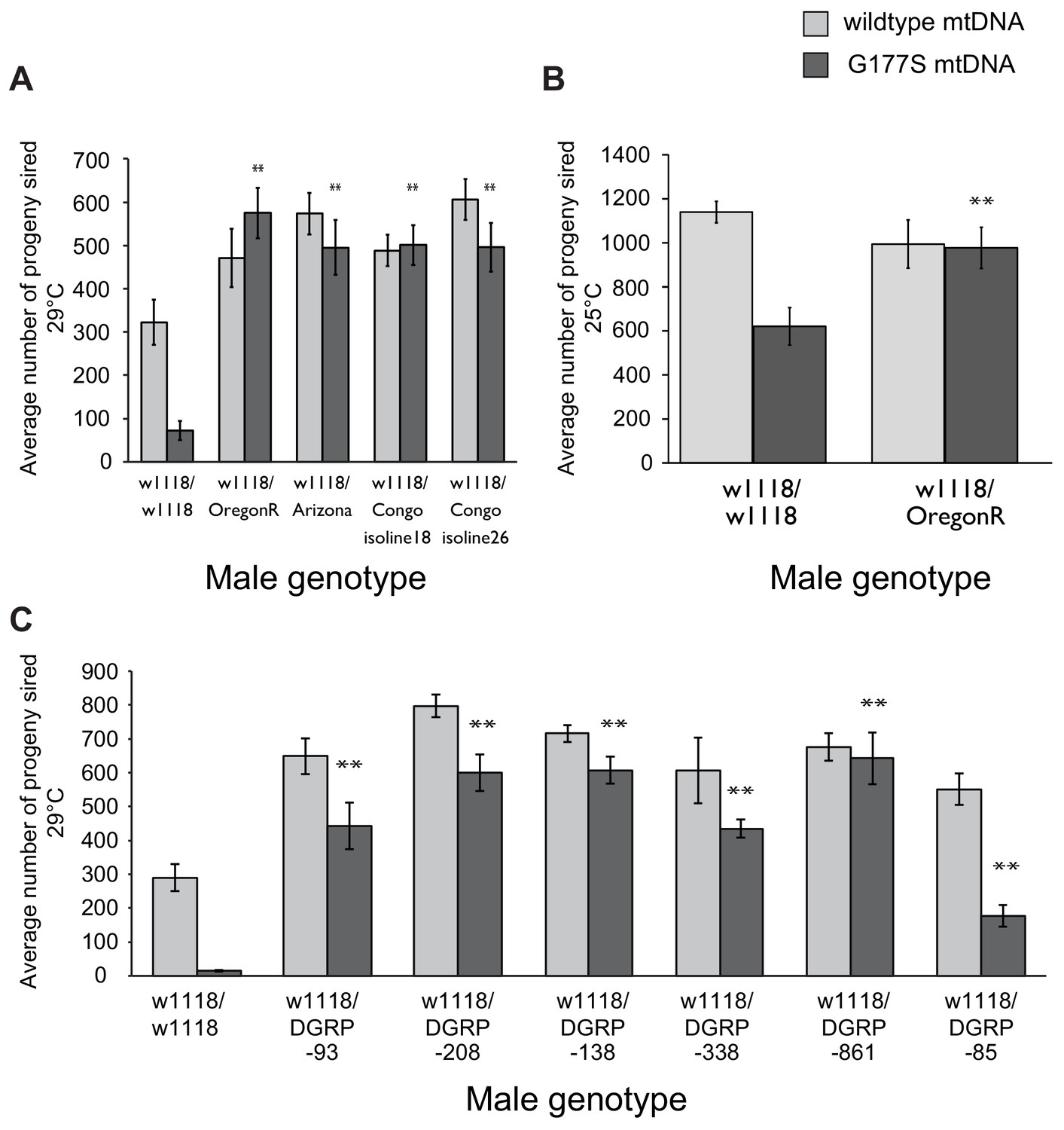
Dominant suppression of COIIG177S-associated male fertility by diverse nuclear genome backgrounds.
(A–C) We measured male fertility of heterozygous males derived from crosses of w1118 females (carrying wildtype or COIIG177S mutant mtDNA) to males from a variety of different nuclear backgrounds. In each case, 3 heterozygous males were mated with 5 w1118 females at 29°C (10 replicates per group) except in panel (B) when fertility was measured at 25°C. Male fertility was measured as average number of resulting progeny; this data is presented as normalized to the wildtype mtDNA background in Figure 9. Most nuclear backgrounds show mild to complete suppression of the male fertility defects observed in w1118/w118 males carrying COIIG177S mutant mtDNA (**p<0.05). Error bars represent standard error of the mean.
Figure 10 with 2 supplements
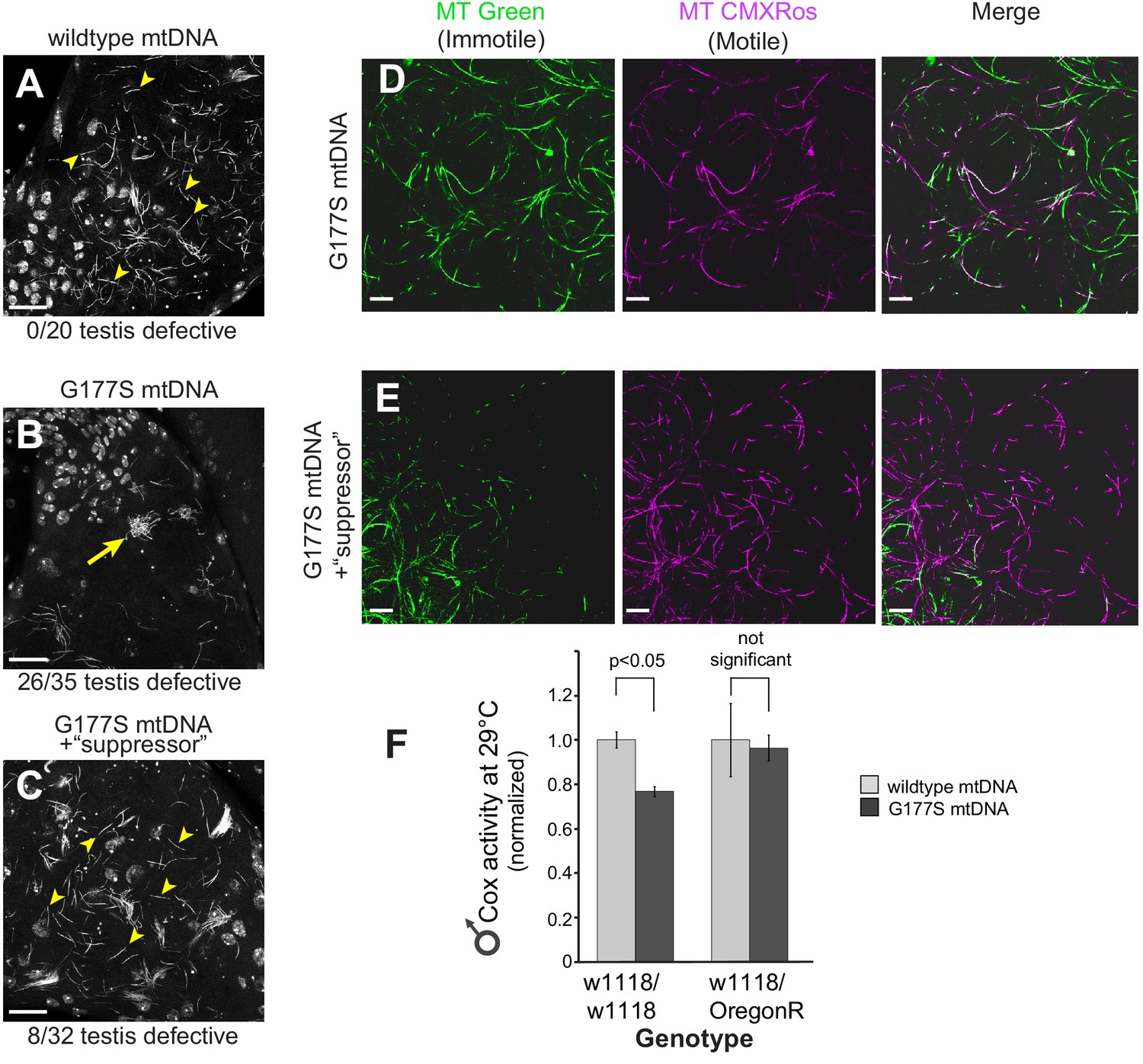
w1118/OreR heterozygous males carrying COIIG177S mutant mtDNA show rescued sperm development and COX activity.
Representative DAPI stained images of the late needle stage of sperm development from w1118/w1118 males carrying either wildtype mtDNA (A), or COIIG177S mtDNA (B) compared to heterozygous w1118/OreR males carrying COIIG177S mtDNA (C). Numbers of examined males with defects in sperm development are indicated. Arrow points to ‘clumped’ sperm. Note the presence of many individualized sperm (arrowheads) in A and C but few in B. Scale bar, 20 μm. We stained sperm from homozygous w1118/w1118 males (D) and w1118/OreR heterozygous males (E), both carrying COIIG177S mutant mtDNA with mitotracker Green (green) and mitotracker CMS Rox (magenta), which stain immotile and motile sperm respectively. All flies were raised at 29°C. Scale bar, 20 μm. (F) COX activity measured from young (3–4 day old) w1118/w1118 males and w1118/OreR heterozygous males both carrying COIIG177S mutant mtDNA. All flies were raised at 29°C. Data is normalized to flies with wildtype mtDNA in the corresponding nuclear background. Error bars represent standard error of the mean.
Figure 10—figure supplement 1
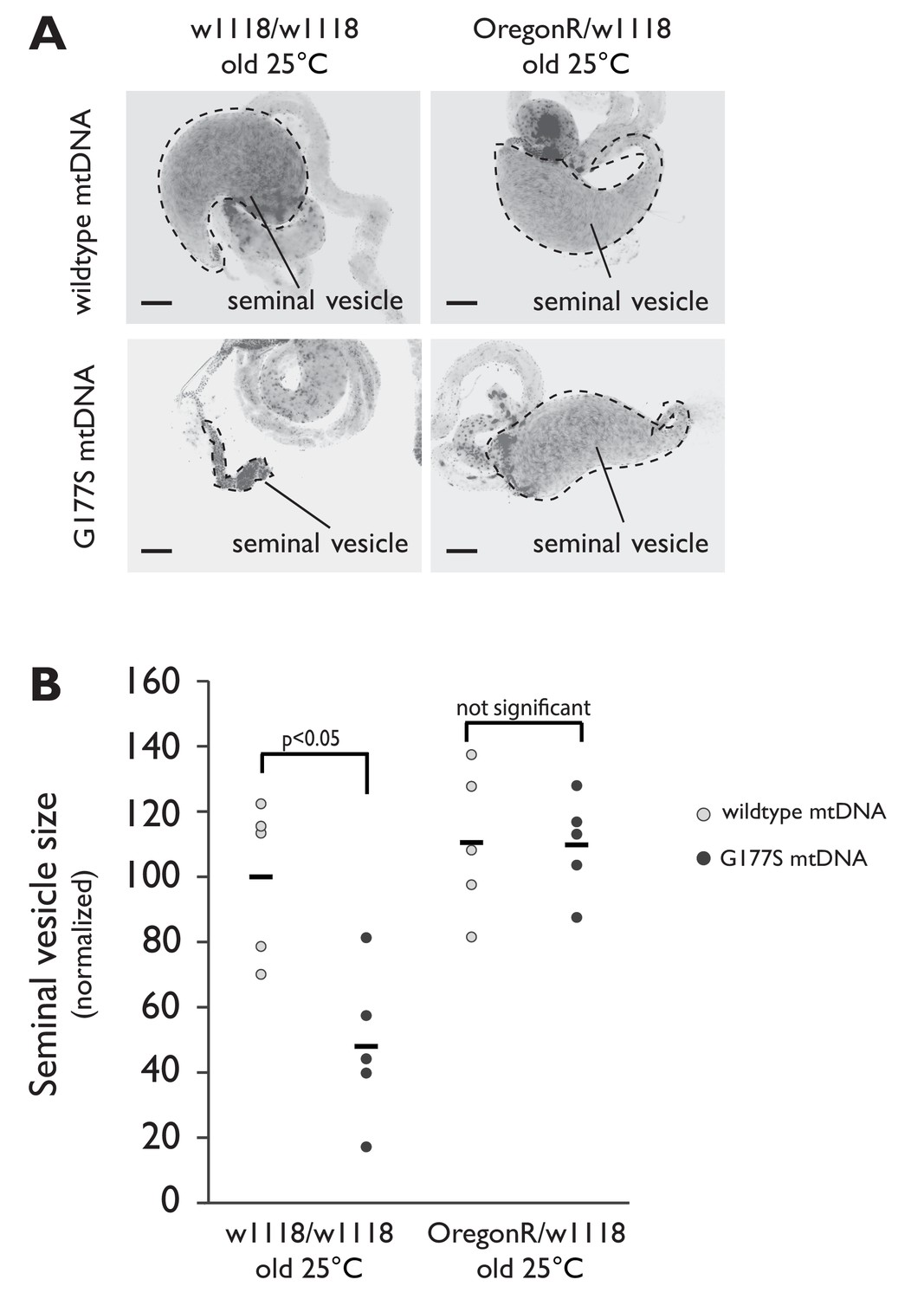
Seminal vesicle size is restored in aged 'suppressor' backgrounds in males containing COIIG177S mutant mtDNA raised at 25°C.
(A) Representative DAPI stained images of seminal vesicles (outlined in dotted line) from either w1118/w1118 or w1118/OregonR males containing either wildtype mtDNA or COIIG177S mutant mtDNA. All males were raised at 25°C and allowed to age before dissection. Note the much smaller seminal vesicle sizes in COIIG177S mutant mtDNA containing aged w1118/w1118 males are completely restored in the w1118/OregonR males. Scale bar, 100 μm. (B) Quantification of the seminal vesicle size, as measured by cross-sectional area, normalized to aged wildtype mtDNA males raised at 25°C (data from Figure 7—figure supplement 1 is reproduced here for easy comparison). Average calculated from 5–7 testes. Error bars represent standard error of the mean.
Figure 10—figure supplement 2
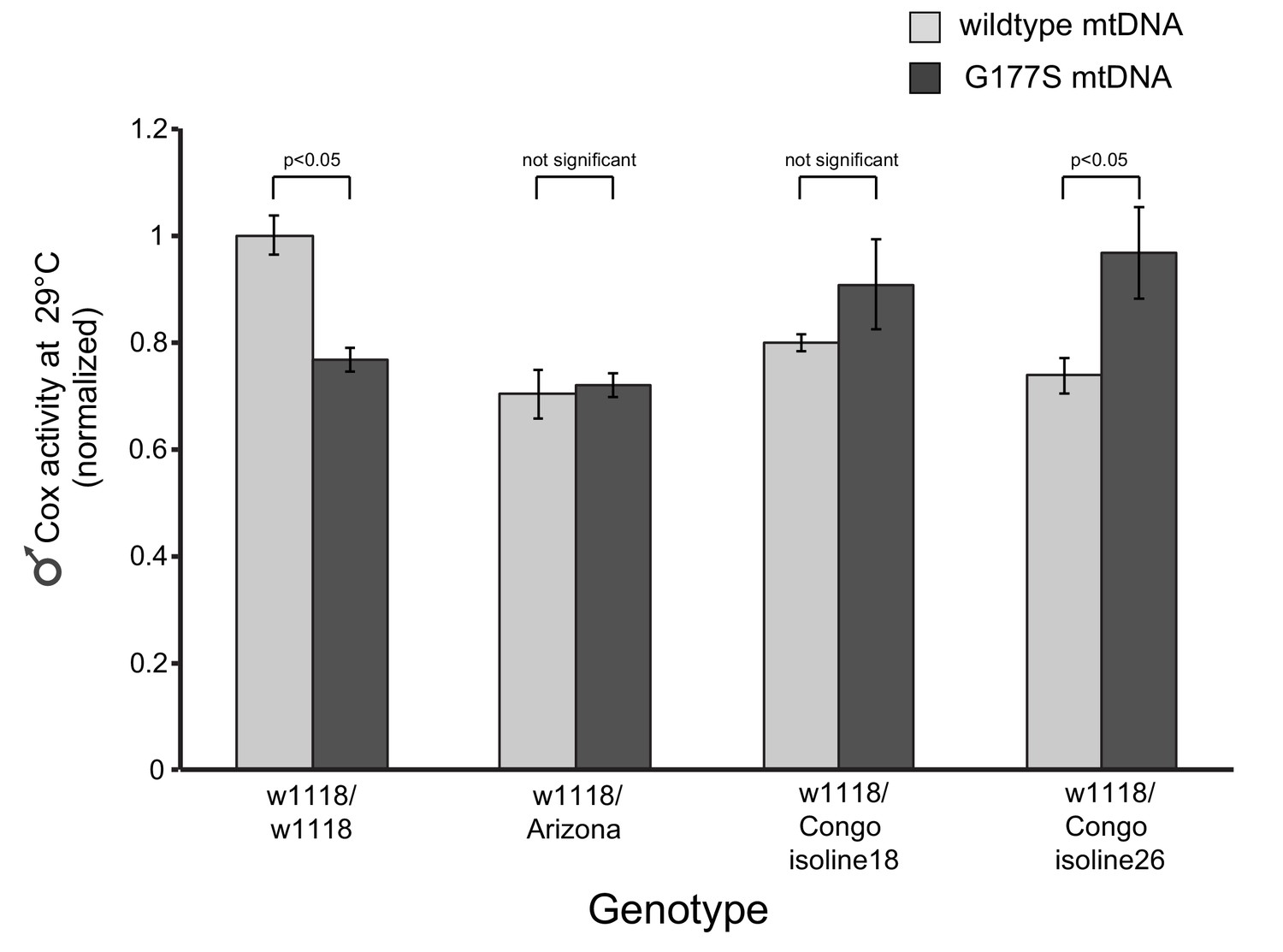
COX activity is specifically restored in ‘suppressor’ nuclear backgrounds.
We measured COX activity from young (3–4 day old) w1118/w1118 males carrying COIIG177S mutant mtDNA; the latter showed a 20% reduction in COX activity. We also measured COX activity in three heterozygous nuclear backgrounds (similar to Figure 10F) and found there was no reduction in COX activity in flies carrying COIIG177S mutant mtDNA. Instead, in one instance, in w1118/Congo26 males, we observed a significant enhancement of COX activity associated with the COIIG177S mutant mtDNA. All flies were raised at 29°C. Data is normalized to flies with wildtype mtDNA in the w1118/w1118 nuclear background.
Videos
Video 1
Sperm motility assays in w1118 males carrying wildtype mtDNA.
From a representative male grown at 29°C. Sperm stained with mitotracker Green (green), which stains immotile sperm and mitotracker CMS Rox (red), which stains motile sperm preferentially (related to Figure 8).
Video 2
Sperm motility assays in w1118 males carrying COIIG177S mutant mtDNA.
From a representative male grown at 29°C. Sperm stained with mitotracker Green (green), which stains immotile sperm and mitotracker CMS Rox (red), which stains motile sperm preferentially (related to Figure 8).
Video 3
Sperm motility assays in w1118/Oregon R males ('nuclear suppressor' background) carrying COIIG177S mutant mtDNA.
From a representative male grown at 29°C. Sperm stained with mitotracker Green (green), which stains immotile sperm and mitotracker CMS Rox (red), which stains motile sperm preferentially (related to Figure 10).
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
A mitochondrial DNA hypomorph of cytochrome oxidase specifically impairs male fertility in Drosophila melanogaster
eLife 5:e16923.
https://doi.org/10.7554/eLife.16923




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}