Assembling the Tat protein translocase
- University of Oxford, United Kingdom
- University of Dundee, United Kingdom
Figures
Figure 1 with 1 supplement
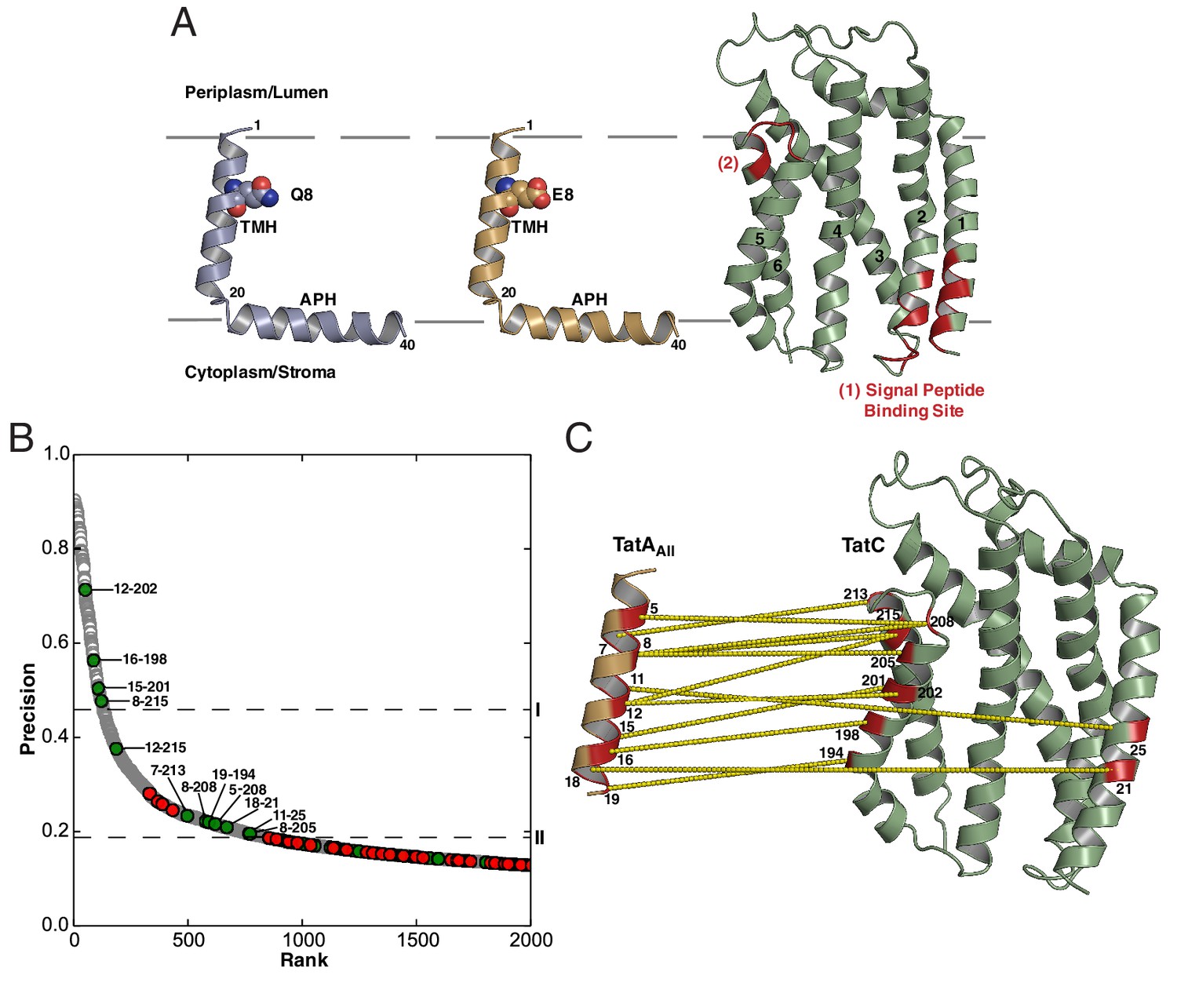
Sequence-coevolution analysis of interactions between TatA family proteins and TatC.
(A) Structures of the E. coli Tat components. The transmembrane (TMH) and amphipathic (APH) helices of TatA and TatB are indicated. Areas of highest surface sequence conservation on TatC are indicated in red and include the binding site for the signal peptide twin-arginine motif. The natively unstructured tails of TatA and TatB are not depicted. (B) Predicted co-evolutionary residue contacts for the TatAAll–TatC dataset using the program PSICOV. Filled circles are predicted inter-subunit co-evolutionary contacts that are either (green) less than 15 Å apart along the membrane normal as expected of authentic direct contacts or (red) at greater than this value and therefore unlikely to correspond to direct interaction pairs. Unfilled gray circles are predicted intra-subunit contacts. Dashed line I marks the evolutionary coupling precision score (0.46) at 7SD above the mean for the whole dataset. Dashed line II marks the evolutionary coupling precision score (0.19) that is 6SD above the mean for the inter-subunit contact dataset. (C) A structural representation of the predicted TatAAll-TatC contacts above threshold level II. See also Table 1.
Figure 1—figure supplement 1
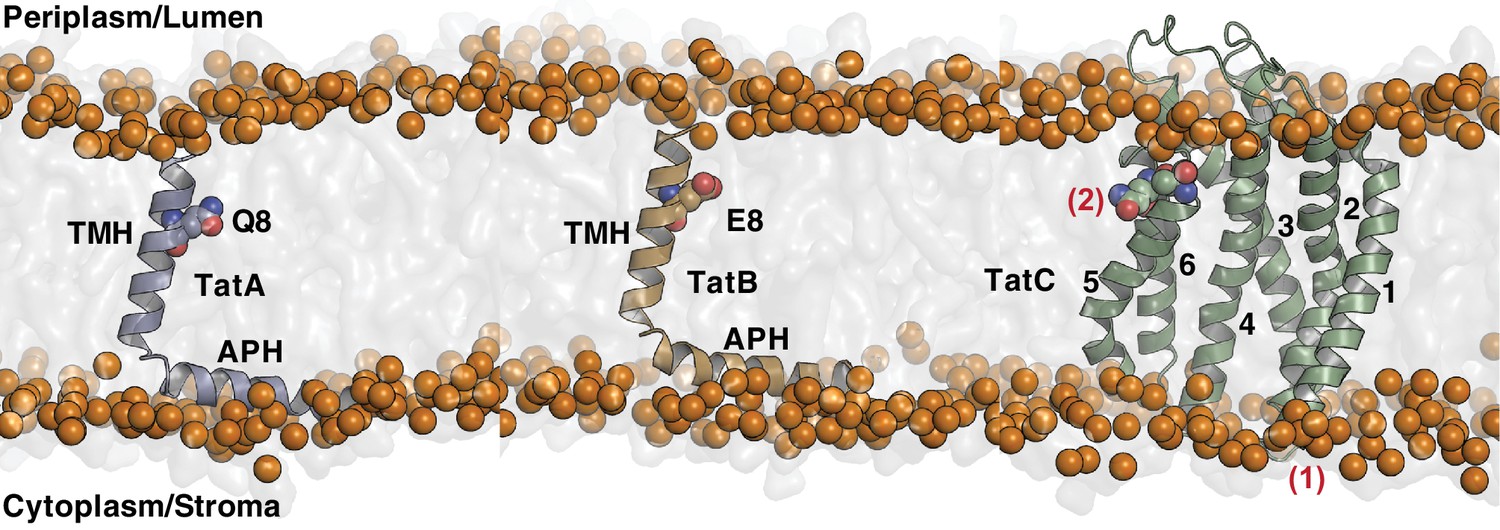
Components of the Tat translocase within a lipid bilayer.
End snapshots of coarse grain-molecular dynamics simulations of the individual Escherichia coli Tat components in a 1:4 phosphatidylglycerol (POPG): 1-palmitoly, 2-oleoyl, phosphatidylethanolamine (POPE) membrane. Orange spheres represent the phosphate group of the membrane. The time-averaged positions of the proteins in the membrane bilayer were used to filter out topologically unrealistic co-evolutionary contacts as described in Materials and methods. Residues predicted to form an interacting intramembrane polar cluster are shown in space-filling mode. All other features are labeled as in Figure 1A.
Figure 2
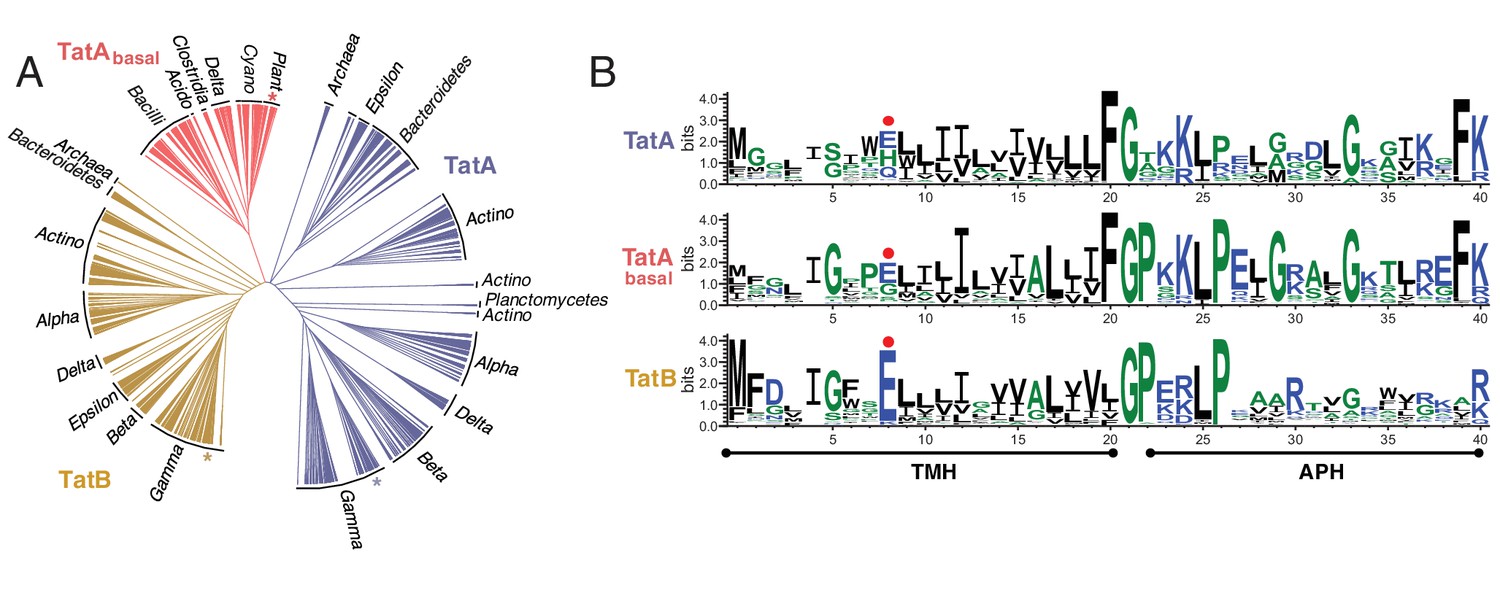
Phylogeny and sequence conservation of the TatA family.
(A) Phylogenetic tree of TatA family members based on a sequence comparison to the end of the APH. The phylogenetic origins of the sequences are given around the edge of the tree: `Alpha’, `Beta’, `Gamma’, `Delta’, and `Epsilon’ refer to the five classes of the phylum Proteobacteria; `Bacilli’ and `Clostridia’ are classes within the phylum Firmicutes; `Actino’ refers to the phylum Actinobacteria; `Acido’ refers to the phylum Acidobacteria; `Cyano’ refers to the phylum Cyanobacteria; `Plants’ refers to plant chloroplast proteins. Asterisks mark the sequences of E. coli TatA/TatE (blue), E. coli TatB (orange), and the pea thylakoid TatA and TatB proteins Tha4 and Hcf106 (red). (B) Sequence logos for the three Tat subsets identified in (A). The logos correspond to the sequence region analyzed in (A) and use the sequence numbering of E. coli TatA and TatB. The secondary structure elements of the Tat proteins are shown under the logos and the position of the TMH polar residue is indicated with a red dot. The figure was generated using WebLogo 3.4 (Crooks et al., 2004).
Figure 3 with 1 supplement

Modeling the interaction of TatA and TatB with the C-terminal end of TatC.
The interaction between the TatA/B TMH and TatC TM5/TM6 modeled for E. coli TatBC and TatAC pairs. The models are based on A. aeolicus TatC crystal packing contacts. Evolutionary couplings for the TatAAll dataset are shown (dotted lines). Couplings retained in just the TatB or TatA datasets with precisions greater than 0.5 (Figure 3—figure supplement 1A) are colored orange or blue, respectively. See also (Table 1).
Figure 3—figure supplement 1
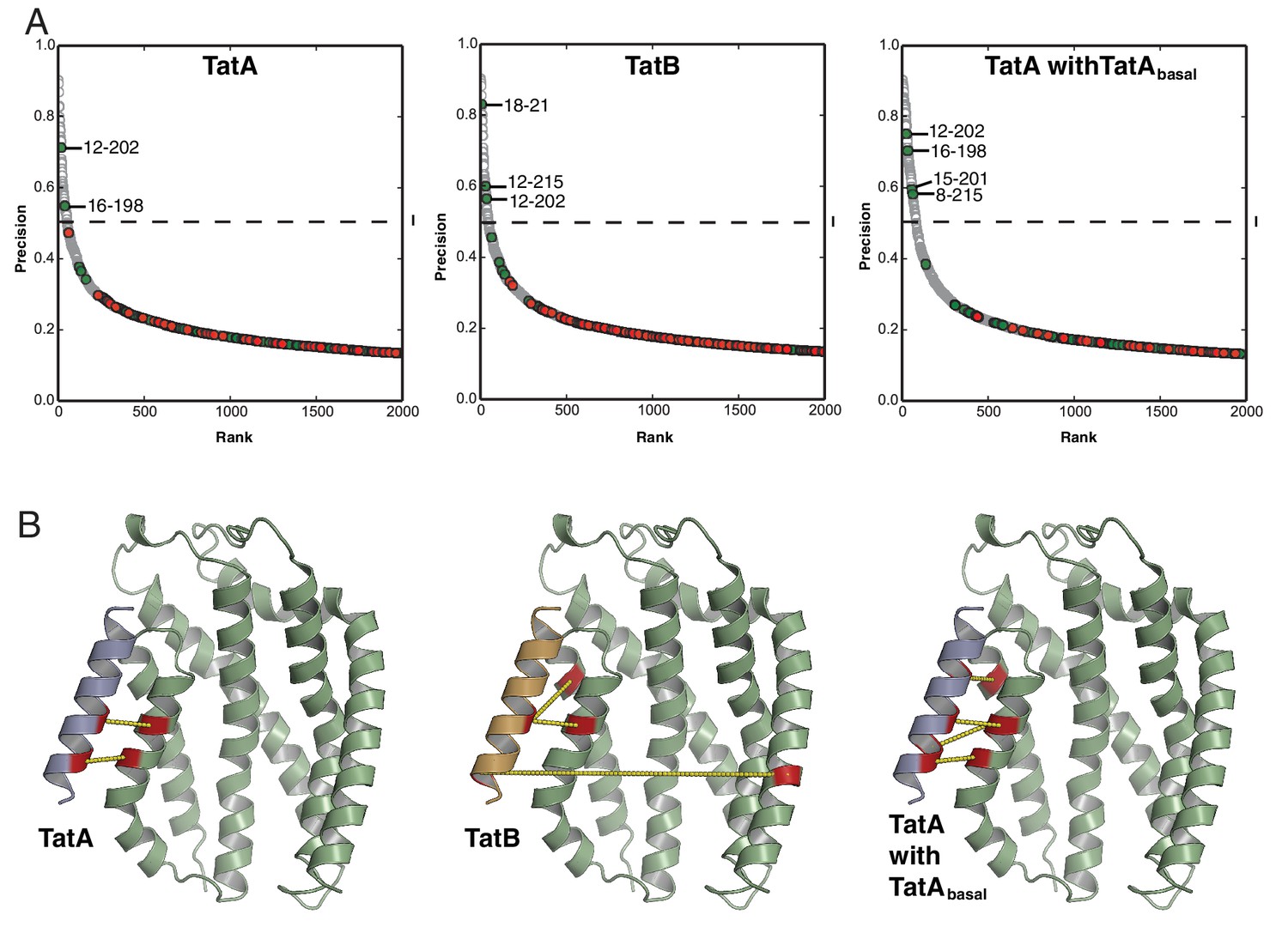
Co-evolution analysis of inter-subunit contacts in the Tat system.
(A) Co-evolution data for TatA subsets with TatC, predicted using PSICOV (Jones et al., 2012). The TatA and TatB subsets were defined phylogenetically as shown in Figure 2A. The comparison using the TatAbasal dataset alone did not identify significant co-variances due to the limited number of sequences in the dataset. However, combining the TatAbasal dataset with the TatA dataset recovered more of the TMH-TatC TM5/TM6 covariances seen with the TatAall dataset than using the TatA dataset alone. This suggests that TatAbasal also binds at this site. Combining the TatAbasal dataset with the TatB dataset did not change the set of high-scoring co-varying pairs recovered with the TatB dataset alone. A cut-off of 0.5 precision (dashed line) is used for increased confidence in the predicted residue pairs. (B) Inter-subunit contacts above 0.5 precision are depicted (yellow dashed lines) on the TatA/B TMH-TatC complex models derived in Figure 3.
Figure 4 with 1 supplement

Molecular simulations of the interactions of TatA and TatB with the C-terminal end of TatC.
(A) Root-mean square deviation (RMSD) of the distances between predicted contact pairs during atomistic MD simulations of the indicated TatBC (left) and TatAC (right) models in a membrane environment taken from three simulations. Both raw data (light gray) and data averaged over a rolling window of 0.35 ns (bold) are shown. Except where indicated, TatB E8 was deprotonated in the simulations. (B) Alanine substitution of the TMH polar residue disrupts the interaction between TatC and the TMHs of TatB (orange) or TatA (blue). The output structures from 100 ns MD simulations are shown with the helix displacements seen in the variants (right hand panel in each pair) relative to the wild-type proteins (left hand panel in each pair) denoted by arrows. (C) Snapshots of the MD simulations of the TatBC and TatAC models showing hydrogen bonding interactions between residues in the inter-subunit polar cluster. Simulations were run with TatB E8 either deprotonated (left panel) or protonated (center panel). See also Videos 1–2.
Figure 4—figure supplement 1
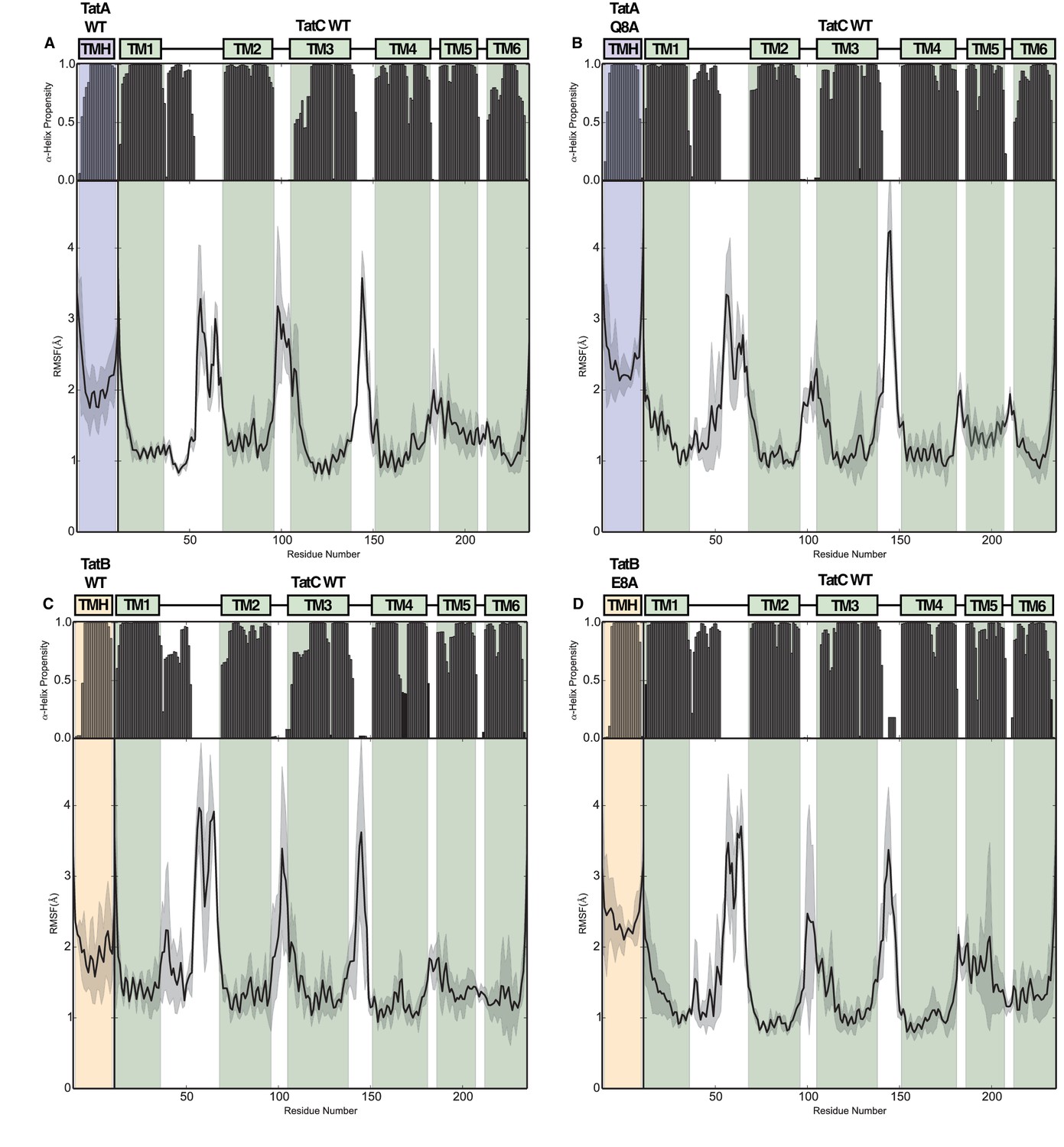
Structural stability plots for the modeled Tat protein complexes from molecular simulations.
Each plot details the secondary structure propensity and Cα-root-mean-square-fluctuations (RMSF) for either the TatA or TatB (deprotonated E8) TMH bound to TatC. The complexes analyzed are (A) TatA with TatC, (B) TatA Q8A with TatC, (C) TatB with TatC, (D) TatB E8A with TatC.
Figure 5 with 1 supplement
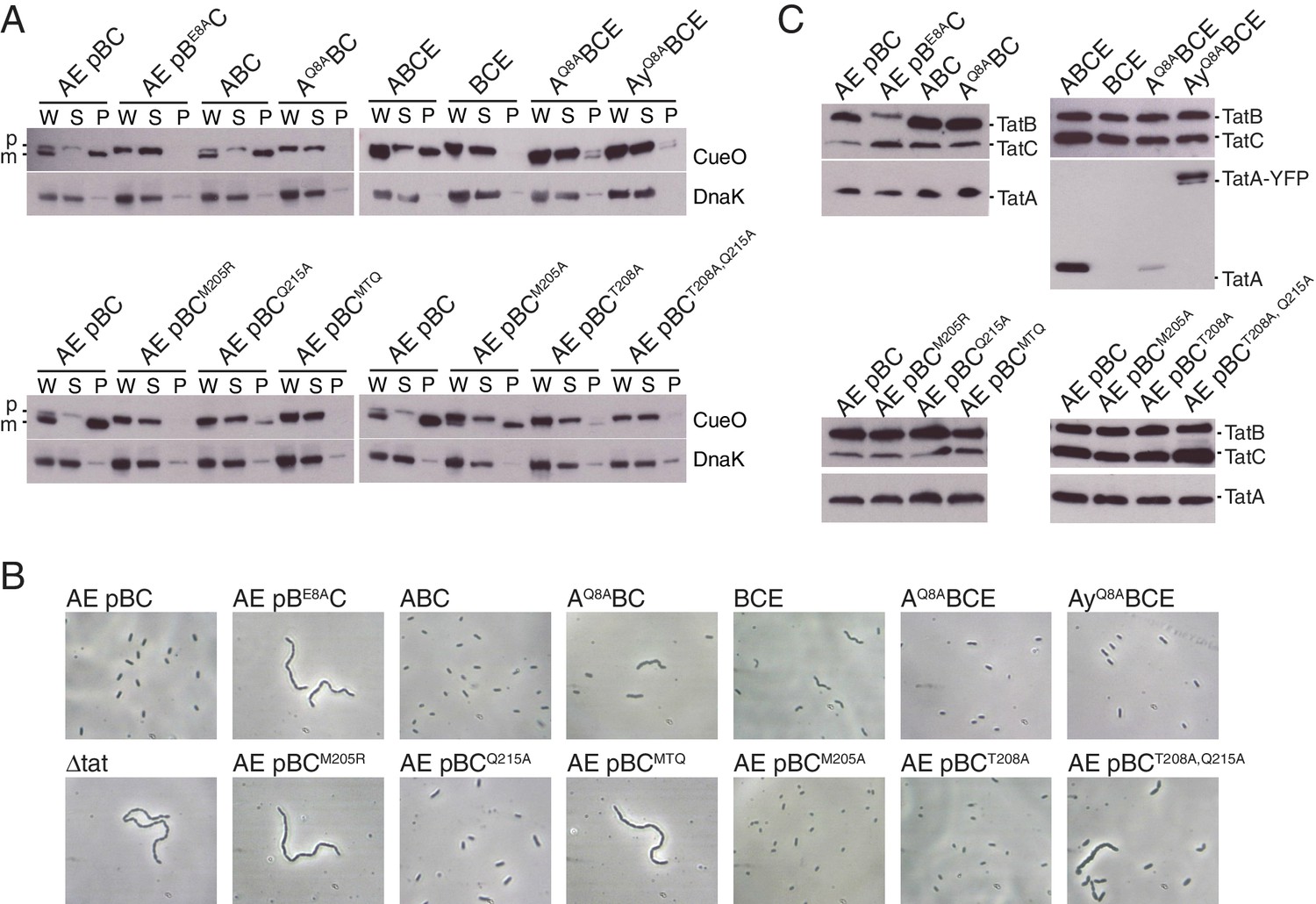
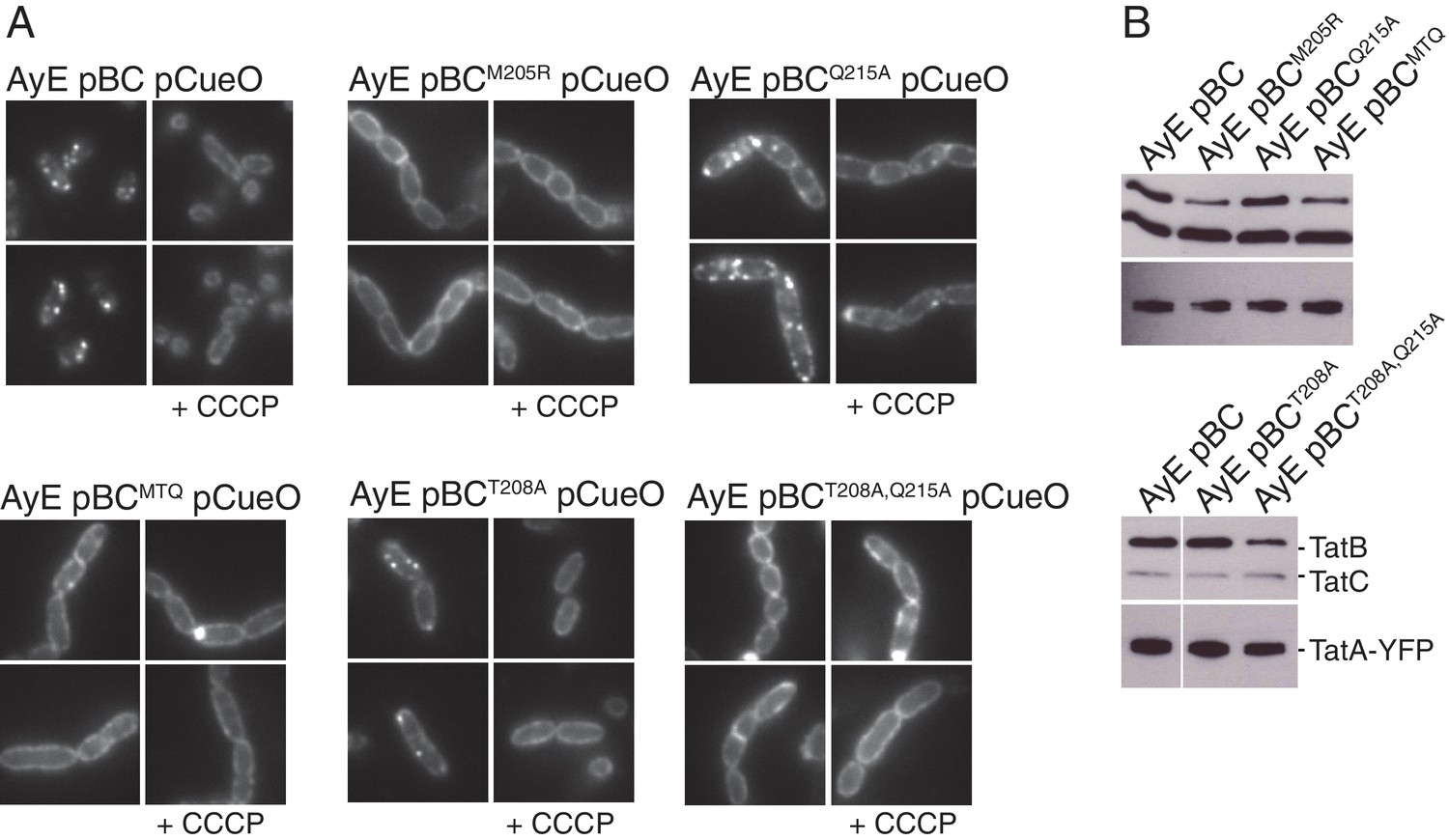
Polar cluster substitutions impair Tat transport.
Strains contained the indicated amino acid substitutions in chromosomally encoded TatA or plasmid-encoded TatB or TatC. TatCMTQ is a combination of the three substitutions M205A, T208A, and Q215A. TatAy is TatA with a C-terminal fusion to yellow fluorescent protein. (A) Transport activity of strains overproducing the Tat substrate CueO. Whole cell (W), spheroplast (S) and periplasm (P) fractions were subject to immunoblotting with antibodies against CueO or the cytoplasmic marker protein DnaK. m is the transported form of CueO from which the signal peptide has been removed and p the precursor protein. (B) Phase contrast images of the strains. `Δtat’ is the complete tat deletion strain DADE-A. (C) Membranes from the same strains were isolated and immunoblotted with a combination of TatB and TatC antibodies or with TatA antibodies.
Figure 5—figure supplement 1
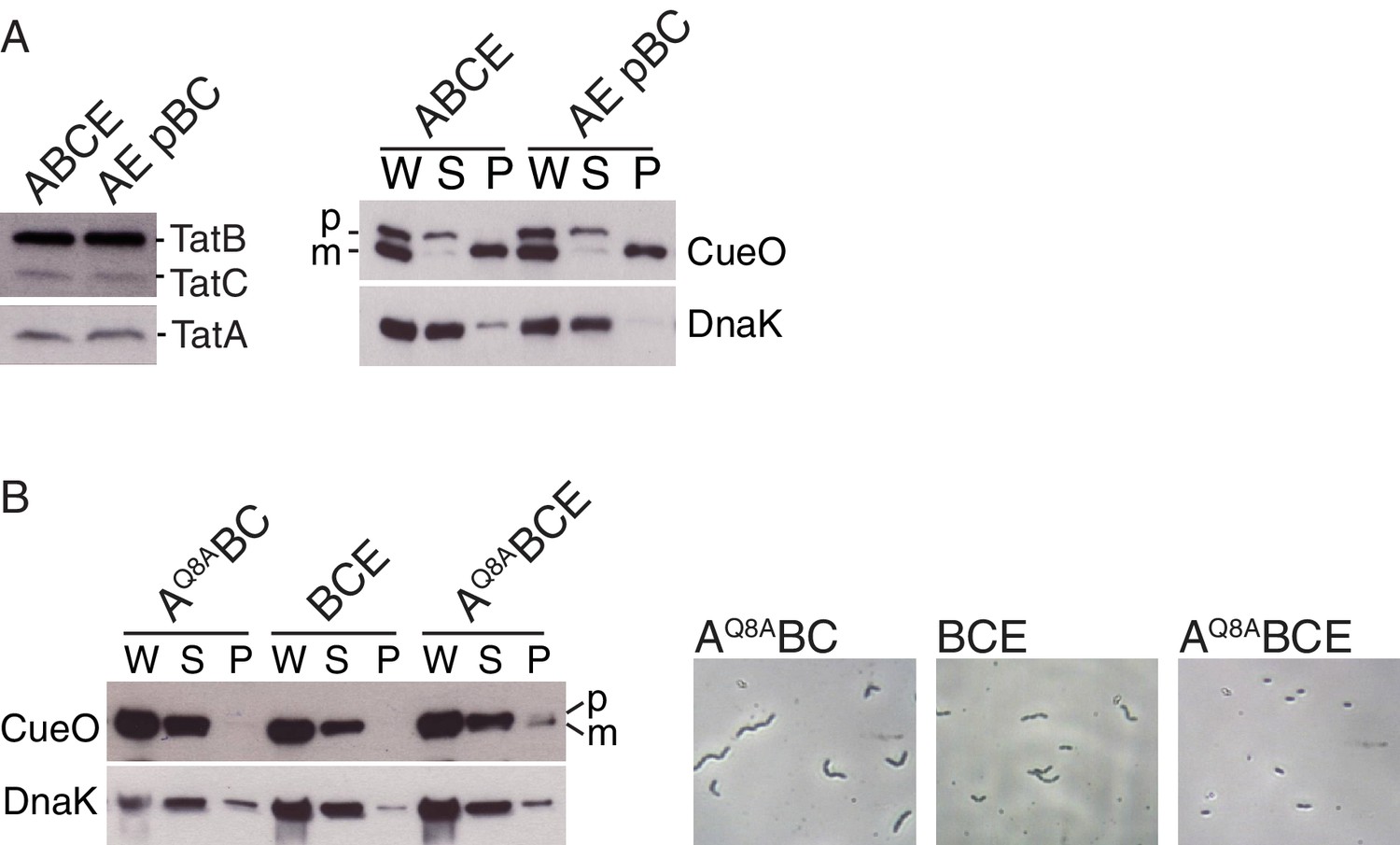
Analysis of Tat transport activity.
(A) Plasmid p101C*TatBC produces wild-type levels of TatB and TatC. (Left) Immunoblot of membranes isolated from the indicated strains probed with a combination of TatB and TatC antibodies, or with TatA antibodies. (Right) Transport activity of strains overproducing the Tat substrate CueO. Whole cell (W), periplasm (P), or spheroplast (S) fractions were subject to immunoblotting with antibodies against CueO (top panel) or the cytoplasmic marker protein DnaK (bottom panel). m is the transported form of CueO from which the signal peptide has been removed and p the precursor protein. (B) The TatAQ8A variant is able to increase the Tat pathway activity of a TatE-containing strain. (Left) Transport activity of strains overproducing the Tat substrate CueO, labeled as in (A). (Right) Phase contrast images of the indicated strains.
Figure 6
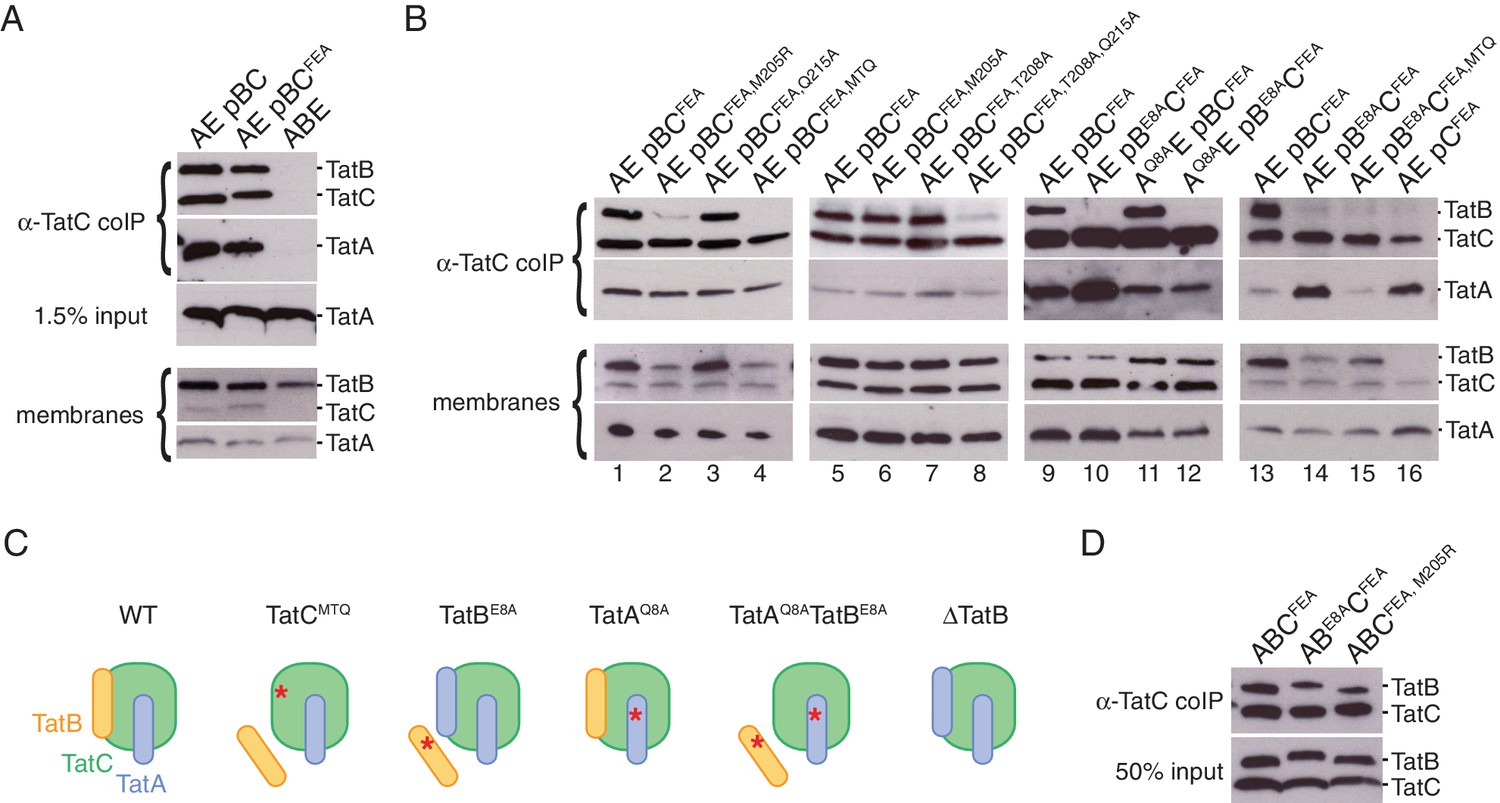
Polar cluster substitutions impair TatBC interactions.
Digitonin-solubilized cell lysates of the indicated strains were immunoprecipitated with antibodies against TatC and then immunoblotted with either a combination of TatB and TatC antibodies or with TatA antibodies (`α-TatC coIP’ panels). Where indicated a portion of the total cell lysate was also immunoblotted (‘input’ panels). (A,B) Co-immunoprecipitation analysis of strains expressing native levels of Tat proteins. Corresponding immunoblots of membranes isolated from the same strains are also shown (`membranes’). The TatCFEA variant is blocked in substrate interactions. The TatCMTQ variant carries the three polar cluster substitutions M205A, T208A, and Q215A. (C) Schematic representation of the results of polar cluster substitutions on Tat receptor complex composition. The red asterisks indicate the location of the polar cluster substitutions. (D) The indicated Tat proteins were overproduced approximately 500-fold in the ΔtatABCDΔtatE strain DADE-A from pUT2DH series plasmids and immunoprecipitated as in (A).
Figure 7 with 1 supplement
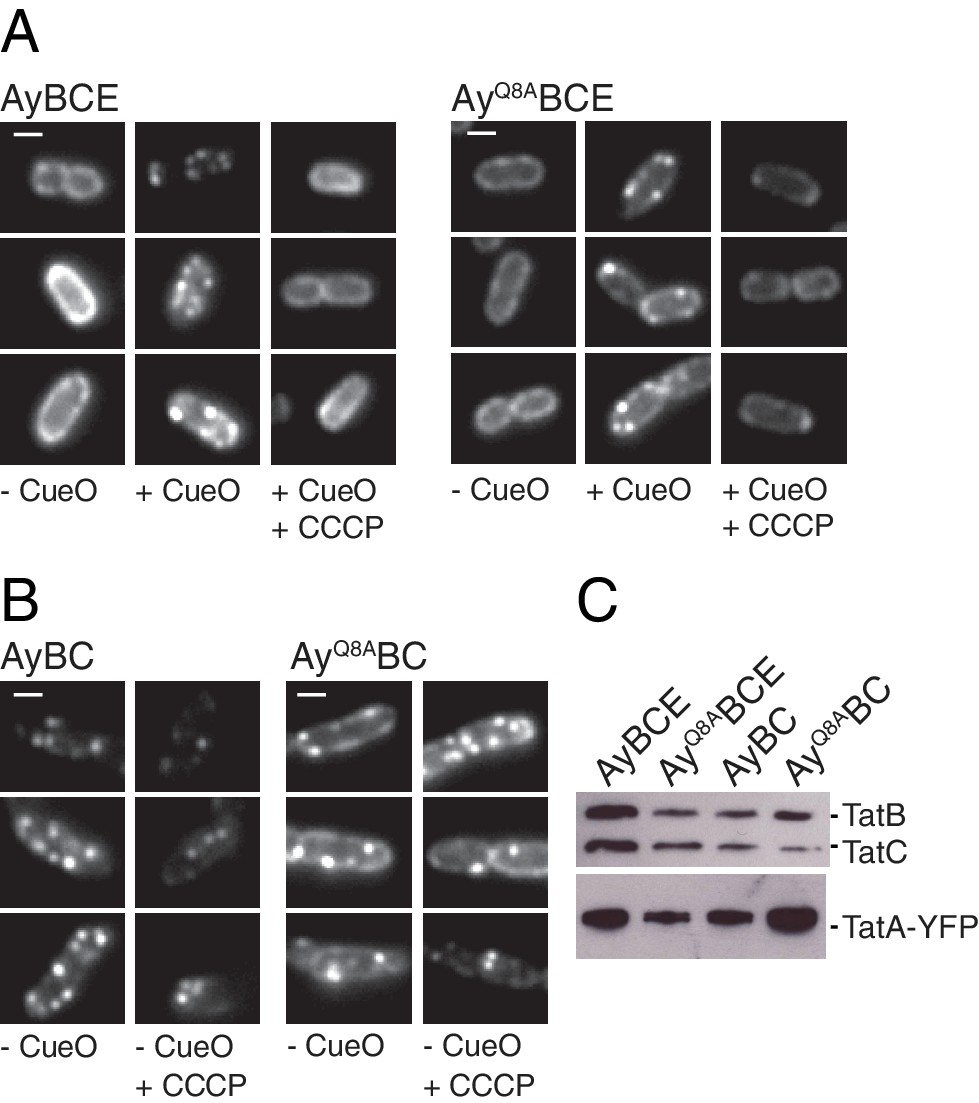
The TatA polar cluster residue is not required for TatA oligomerization.
(A,B) Fluorescence images of TatA-YFP in living cells. The indicated strains were either left untreated (-CueO columns) or the Tat substrate protein CueO was overproduced from plasmid pQE80-CueO by adding 1 mM IPTG to early exponential phase cultures for 30 min prior to imaging (+CueO columns). 50 μM CCCP was subsequently added as indicated (+CCCP columns). Scale bar = 1 μM (C) Membranes isolated from the same strains were immunoblotted using a combination of TatB and TatC antibodies or with TatA antibodies to assess protein expression levels.
Figure 7—figure supplement 1
The effect of TatC polar cluster substitutions on the substrate-induced assembly of TatA.
(A) Fluorescence images of TatA-YFP in live cells overproducing the substrate protein CueO. Overproduction of CueO was induced in early exponential phase cultures of the indicated strains with 1 mM IPTG 30 min prior to immobilizing the cells for microscopy. Additional images were collected after addition of 50 μM CCCP (+CCCP). (B) Membranes from the same strains were isolated and immunoblotted with a combination of TatB and TatC antibodies, or with TatA antibodies. MTQ indicates the three substitutions M205A, T208A, and Q215A.
Figure 8 with 4 supplements
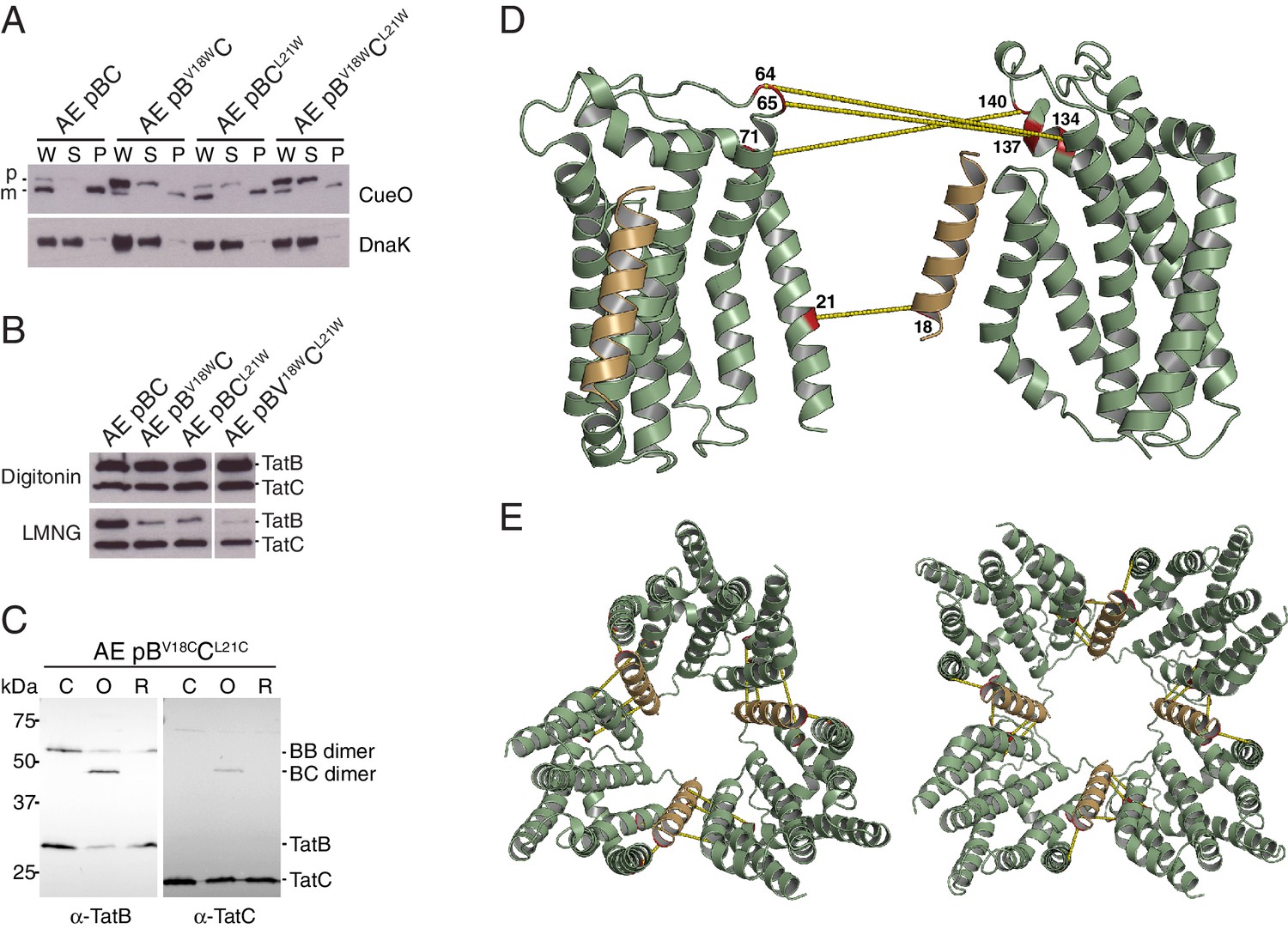
Identification of a second TatBTMH-TatC contact site.
(A) Tat transport activity of strains with tryptophan substitutions targeting the predicted interface between the TatB TMH and TatC TM1. Methodology and labels are as for Figure 5A. (B) Effects of the tryptophan substitutions on TatBC interactions. Cell lysates were solubilized in either digitonin (top panel) or LMNG (bottom panel), immunoprecipitated with antibodies against TatC, and then immunoblotted with a combination of TatB and TatC antibodies. (C) Disulfide crosslinks can be detected at the predicted interface between the TatB TMH and TatC TM1. Cells carrying the indicated cysteine substituted Tat variants were subjected to a mock incubation (‘C’, no oxidant or reductant), oxidizing (‘O’, copper phenanthroline) or reducing (‘R’, DTT) conditions. Membranes were then isolated and subjected to immunoblotting with TatB (left panels) or TatC (right panels) antibodies. (D) Structural representation of the highest-scoring co-evolution-predicted contacts between TatBTMHC heterodimers (precision >0.6). (E) Model for the TatBC complex based on docking either three (Left) of four (Right) TatBTMH-TatC heterodimers to optimize agreement with the co-evolution data in (D). The complexes are viewed from the cytoplasmic side of the membrane. See also Figure 8—figure supplement 3, Video 3 and Supplementary files 1 and 2.
Figure 8—figure supplement 1
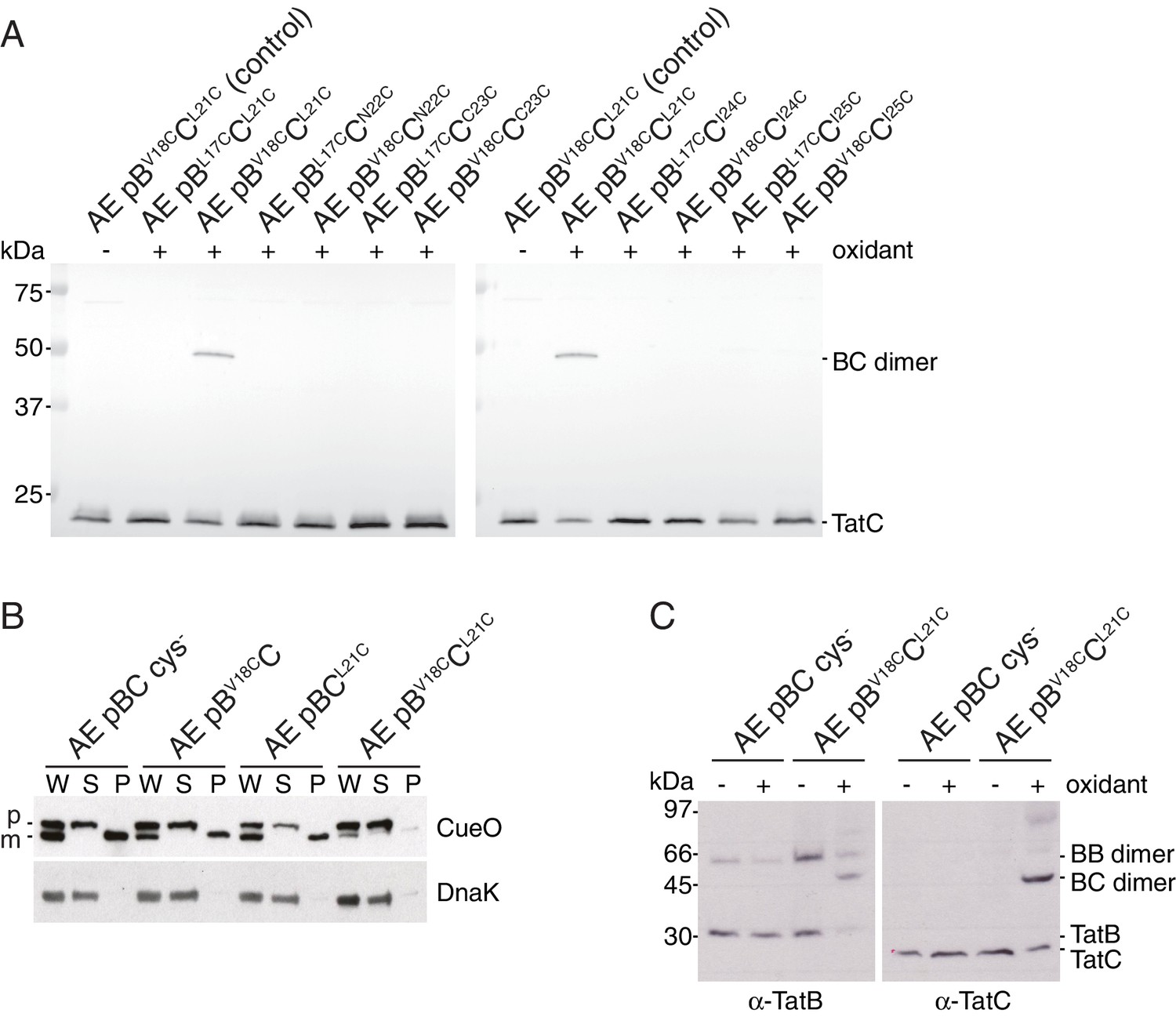
TatB V18C and TatC L21C substitutions permit crosslinking of TatB to TatC and impair transport activity.
(A) The indicated strains were subjected to control (no oxidant) or oxidizing (1.8 mM copper phenantrholine) conditions. Membranes were then isolated and subjected to immunoblotting with TatC antibodies. (B) Transport activity of strains overproducing the Tat substrate CueO. Whole cell (W), periplasm (P), and spheroplast (S) fractions were subject to immunoblotting with antibodies against CueO (top panel) or the cytoplasmic marker protein DnaK (bottom panel). m is the transported, signal peptide-cleaved form of CueO, and p the precursor protein. (C) The cultures used in (B) were subjected to control (no oxidant) or oxidizing (0.9 mM copper phenanthroline) conditions. Membranes were isolated and subjected to immunoblotting with TatB (left panel) or TatC (right panel) antibodies.
Figure 8—figure supplement 2
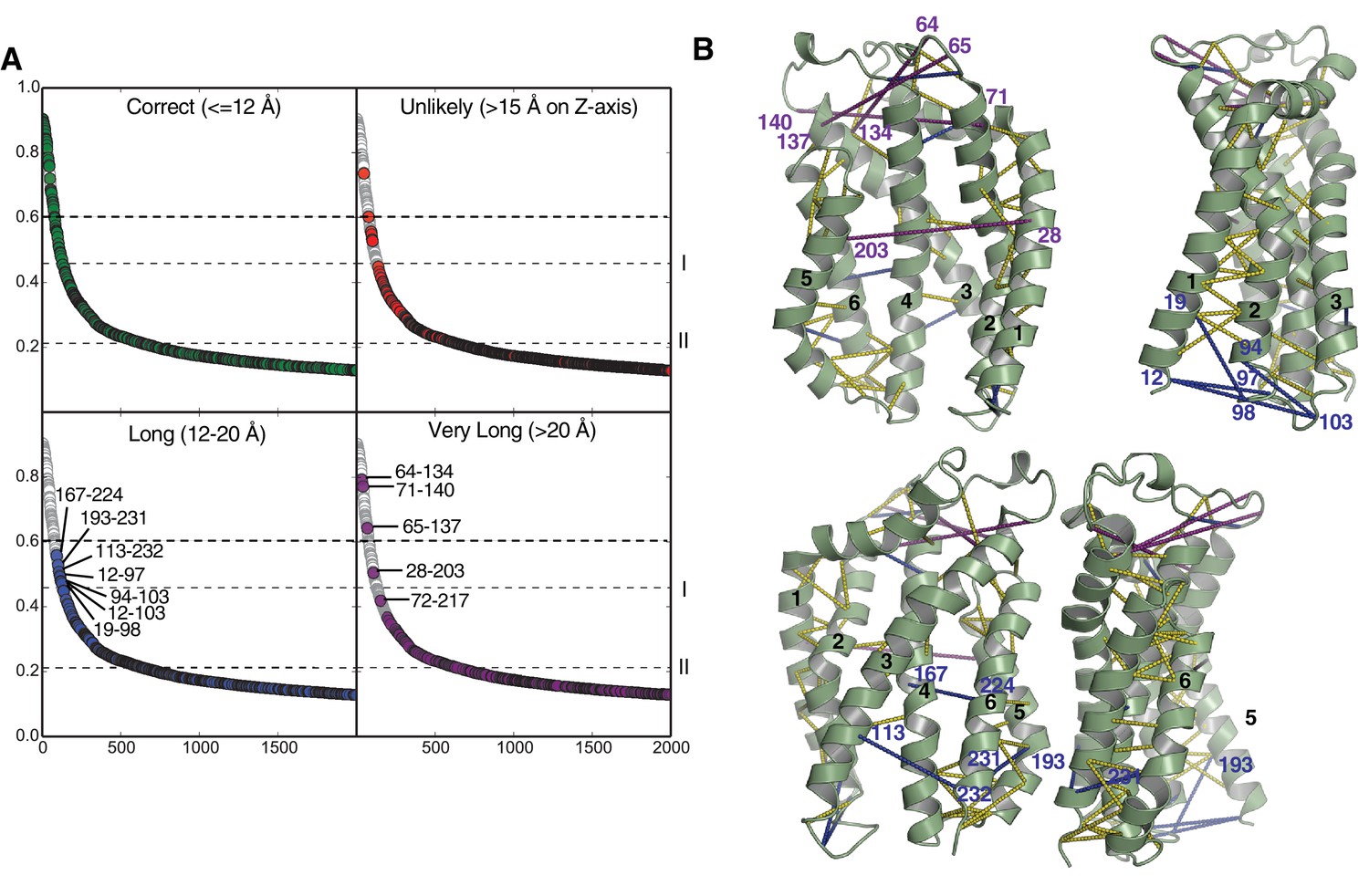
Evolutionary contacts predicted by PSICOV for TatC.
(A) Predicted TatC-TatC contacts from the analysis in Figure 1B were mapped on to the modeled structure of E. coli TatC and then sorted by the distance between the Cα-atoms of the interacting residues. Contacts within 12 Å were considered to be consistent with the crystal structure of TatC (green). Contacts with a separation of greater than 15 Å across the membrane (direction of the z-axis) were deemed to be spurious under any model (red). Contacts with separations between 12 and 20 Å may indicate contacts in alternative conformational states (blue). Contacts with separations over 20 Å are candidates for inter-subunit contacts within the TatBC multimer (purple). Dashed line I marks the co-variance score that is 7SD above the mean for the full contact dataset shown in Figure 1B. Dashed line II marks the co-variance score that is 6SD above the mean for the inter-subunit contacts shown in Figure 1B. See also Table 1. (B) Four orthogonal views of the TatC protein showing predicted TatC-TatC contacts above the 7SD significance level (dotted lines). Plotted are all contacts (yellow), contacts with separations between 12 and 20 Å (blue), and contacts with separations over 20 Å (purple). Contacts with separations between 12 and 20 Å cluster at the cytoplasmic face of the membrane and include residues known to be involved in signal peptide binding (H12, F94, and E103, E. coli TatC numbering). These putative contacts may, therefore, characterize a TatC conformer that is reached after substrate binding.
Figure 8—figure supplement 3
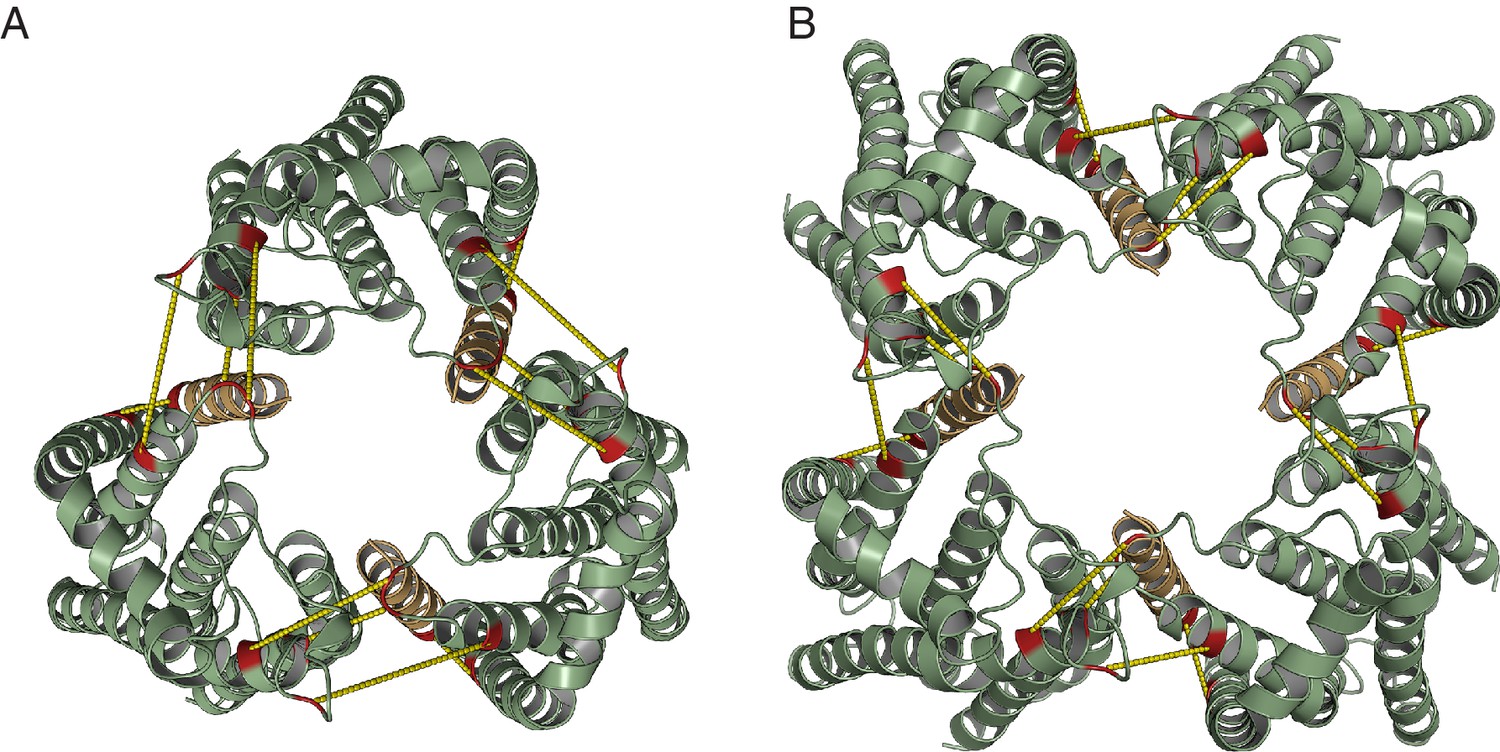
Model for the TatBC complex based on docking either (A) three or (B) four TatBTMH-TatC heterodimers to optimize agreement with the co-evolution data in Figure 8D.
The complex is viewed from the periplasmic side of the membrane.
Figure 8—figure supplement 4
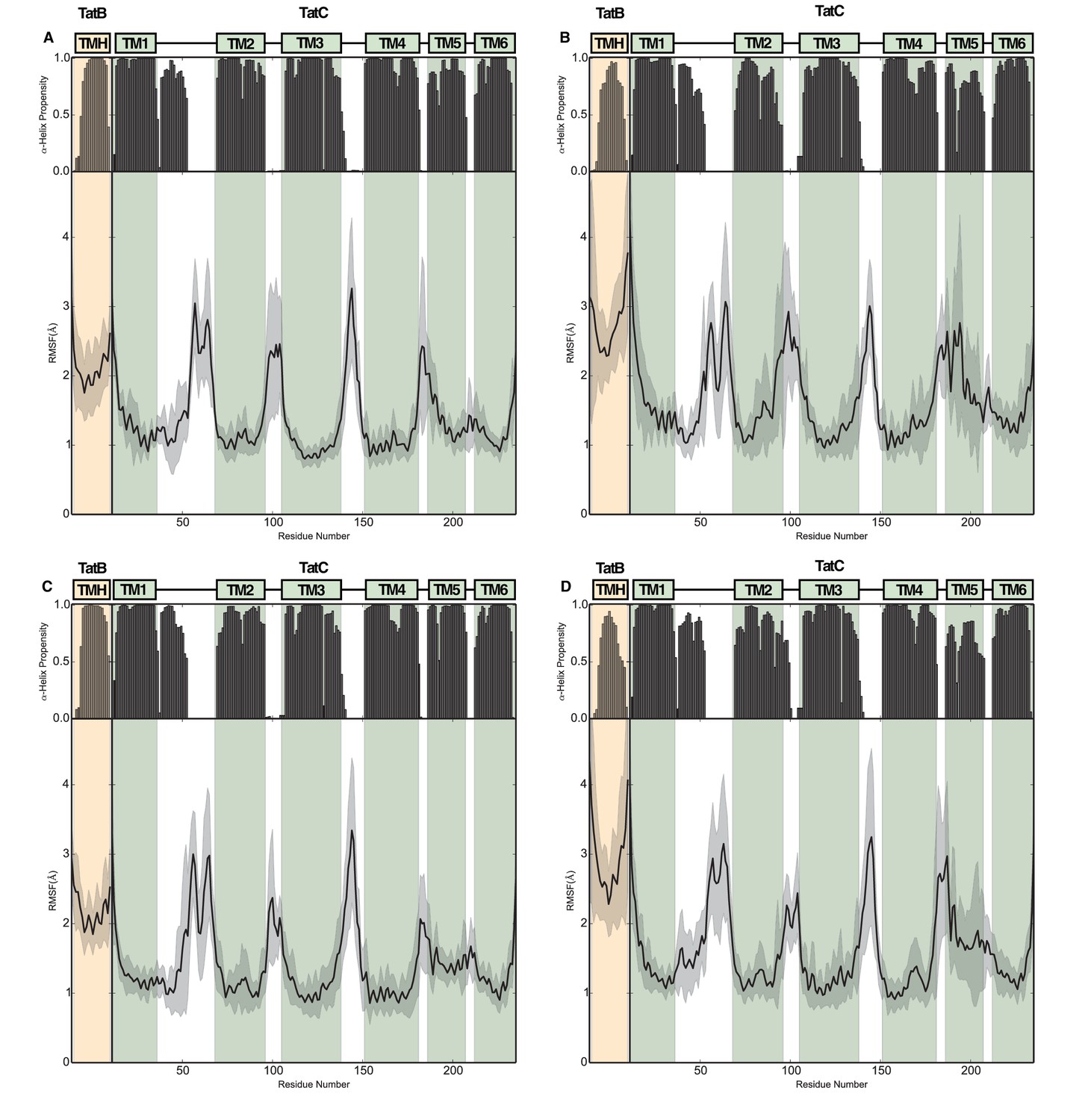
Structural stability plots for the modeled trimeric and tetrameric Tat protein complexes from molecular simulations.
(A) The (TatBC)3 complex with lipids in the central pore, (B) the (TatBC)3 complex with water in the central pore, (C) the (TatBC)4 complex with lipids in the central pore, and (D) the (TatBC) 4 complex with water in the central pore.
Figure 9
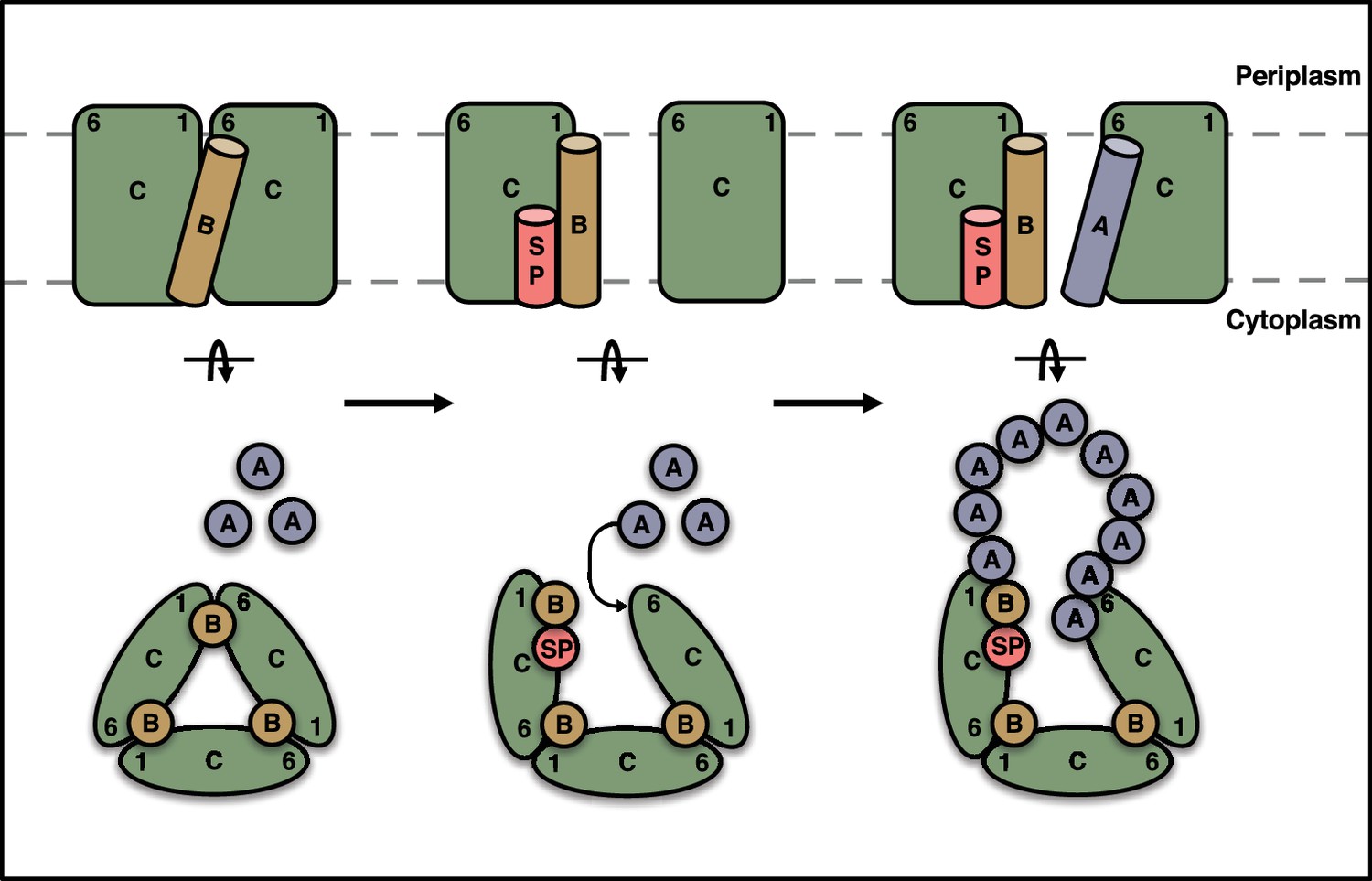
Schematic model for substrate activation of the Tat receptor complex.
The TMHs of TatB molecules (orange) in the receptor complex are sandwiched between TM1 of one TatC molecule and TM6 of the adjacent TatC molecule (green). Signal peptide (SP, red) binding to a TatC subunit transmits a conformational change through TM1 that reduces the affinity of TatB for the TM6 site on the adjacent TatC molecule and favors uptake of TatA (blue) into this site. The incoming TatA molecule is envisaged to nucleate the formation of the TatA oligomer. Current data are insufficient to determine the final location of the displaced TatB molecule and so the position shown should be regarded as speculative. A (TatBC)3 oligomer is shown for simplicity, but this mechanism is generally applicable to a (TatBC)n complex.
Videos
Video 1
Representative atomistic molecular dynamics simulations of E. coli TatA-TatC and TatB-TatC heterodimers in a phospholipid bilayer.
The interactions of the wild-type TatA TMH (blue; left panel panel) or TatB TMH (deprotonated E8) (orange; right hand panel) with TatC (green) were assessed in a 100 ns molecular simulation. Yellow dashed lines connect the predicted co-evolving residue pairs shown in Figure 3.
Video 2
Polar cluster interactions between TatC residues T208 and Q215 (green) and either deprotonated TatB residue E8 (orange), or protonated TatB residue E8 (orange), or TatA residue Q8 (blue) during 100 ns of molecular simulation.
Black dashed lines indicate hydrogen bonds.
Video 3
Animations of the (TatBC)3 and (TatBC)4 complex models.
https://doi.org/10.7554/eLife.20718.022
Video 4
Exemplar atomistic 100 ns molecular simulations of the (TatBC)3 (Left) and (TatBC)4 (Right) complexes in a membrane bilayer.
The complexes are viewed from the cytoplasmic side of the membrane. The simulations were run with phospholipids filling the internal pore of the complex.
Tables
Table 1
Comparison of PSICOV predictions for Tat component contacts with those of other co-evolutionary methods. The programs PSICOV (Jones et al., 2012), CCMPRED (Seemayer et al., 2014), and FreeContact (mfDCA) (Hopf et al., 2012; Kaján et al., 2014) use methodologically distinct algorithms to calculate sequence co-evolution. Contacts predicted by all three of the co-evolutionary methods are colored green. Meta-PSICOV (Jones et al., 2015) aggregates information from the other three prediction methods as well as other sequence information in Stage 1, then weights by vicinity to other contacts in Stage 2. The table is divided to include the co-evolutionary contacts for TatAAll, TatA, TatB, TatA with TatAbasal, TatA with TatBbasal, and TatC-TatC contacts with separations greater than 20 Å. For the TatC-TatC contacts the three high-probabilty contacts identified by the three distinct methods suggest plausible contacts between the TM2-3 linker and residues at the C-terminal end of TM3 (green).
PSICOV | FreeContact (mfDCA) | CCMPRED | Meta-PSICOV Stage 1 | Meta-PSICOV Stage 2 | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
TMH | TatC | Score | Rank | TMH | TatC | Score | Rank | TMH | TatC | Score | Rank | TMH | TatC | Score | Rank | TMH | TatC | Score | Rank | |
TatAAll | 12 | 202 | 0.71 | 51 | 12 | 202 | 1.46 | 108 | 12 | 202 | 0.30 | 28 | 12 | 202 | 0.75 | 22 | 12 | 202 | 0.46 | 112 |
16 | 198 | 0.56 | 88 | 15 | 201 | 1.00 | 305 | 16 | 198 | 0.27 | 59 | 16 | 198 | 0.71 | 30 | 15 | 201 | 0.40 | 145 | |
15 | 201 | 0.5 | 109 | 16 | 198 | 0.95 | 335 | 15 | 201 | 0.23 | 117 | 15 | 201 | 0.64 | 49 | 16 | 198 | 0.34 | 191 | |
8 | 215 | 0.48 | 123 | 8 | 215 | 0.84 | 422 | 12 | 215 | 0.21 | 175 | 12 | 215 | 0.19 | 414 | 12 | 201 | 0.15 | 444 | |
12 | 215 | 0.38 | 189 | 12 | 215 | 0.75 | 549 | 18 | 21 | 0.19 | 313 | 15 | 198 | 0.14 | 609 | 12 | 198 | 0.08 | 771 | |
7 | 213 | 0.23 | 497 | 18 | 21 | 0.56 | 1010 | 11 | 212 | 0.18 | 323 | 18 | 21 | 0.14 | 611 | 8 | 215 | 0.05 | 1171 | |
8 | 208 | 0.22 | 578 | 8 | 208 | 0.53 | 1128 | 8 | 215 | 0.18 | 388 | 8 | 215 | 0.11 | 734 | 4 | 205 | 0.04 | 1332 | |
19 | 194 | 0.22 | 591 | 14 | 172 | 0.45 | 1577 | 15 | 198 | 0.18 | 400 | 8 | 208 | 0.09 | 860 | 8 | 205 | 0.04 | 1593 | |
5 | 208 | 0.22 | 619 | 8 | 205 | 0.44 | 1630 | 5 | 208 | 0.18 | 404 | 12 | 201 | 0.06 | 1072 | 5 | 208 | 0.03 | 1726 | |
18 | 21 | 0.21 | 670 | 5 | 126 | 0.43 | 1717 | 11 | 174 | 0.16 | 674 | 12 | 198 | 0.06 | 1083 | 15 | 198 | 0.03 | 1897 | |
11 | 25 | 0.2 | 768 | 15 | 198 | 0.42 | 1761 | 8 | 208 | 0.15 | 753 | 14 | 203 | 0.05 | 1175 | 8 | 208 | 0.03 | 1963 | |
8 | 205 | 0.19 | 772 | 17 | 85 | 0.42 | 1851 | 4 | 205 | 0.15 | 793 | 4 | 205 | 0.05 | 1176 | 11 | 202 | 0.03 | 2044 | |
8 | 166 | 0.19 | 855 | 5 | 124 | 0.39 | 2105 | 17 | 227 | 0.15 | 802 | 5 | 208 | 0.04 | 1367 | 8 | 198 | 0.02 | 2299 | |
12 | 198 | 0.18 | 911 | 5 | 132 | 0.38 | 2131 | 19 | 194 | 0.15 | 960 | 11 | 25 | 0.04 | 1424 | 15 | 202 | 0.02 | 2569 | |
17 | 227 | 0.17 | 1003 | 12 | 194 | 0.33 | 2870 | 8 | 214 | 0.15 | 979 | 11 | 174 | 0.04 | 1544 | 4 | 208 | 0.02 | 2846 | |
15 | 198 | 0.17 | 1150 | 12 | 75 | 0.32 | 2995 | 12 | 198 | 0.15 | 1036 | 15 | 219 | 0.03 | 1574 | 8 | 202 | 0.02 | 3287 | |
12 | 201 | 0.16 | 1183 | 12 | 198 | 0.32 | 3008 | 12 | 201 | 0.14 | 1158 | 10 | 25 | 0.03 | 1607 | 4 | 206 | 0.01 | 3437 | |
TatA | 12 | 202 | 0.71 | 18 | 12 | 202 | 1.81 | 16 | 12 | 202 | 0.21 | 10 | 12 | 202 | 0.72 | 12 | 12 | 202 | 0.46 | 91 |
16 | 198 | 0.55 | 39 | 16 | 198 | 1.12 | 124 | 16 | 198 | 0.19 | 15 | 16 | 198 | 0.72 | 13 | 9 | 206 | 0.27 | 229 | |
14 | 216 | 0.38 | 122 | 12 | 215 | 1.06 | 155 | 9 | 206 | 0.14 | 49 | 9 | 206 | 0.40 | 82 | 16 | 198 | 0.27 | 230 | |
16 | 82 | 0.37 | 133 | 5 | 136 | 0.91 | 246 | 15 | 163 | 0.14 | 67 | 15 | 163 | 0.35 | 98 | 4 | 206 | 0.18 | 378 | |
9 | 206 | 0.34 | 162 | 5 | 210 | 0.86 | 287 | 3 | 205 | 0.14 | 72 | 15 | 202 | 0.29 | 137 | 9 | 202 | 0.11 | 630 | |
5 | 162 | 0.29 | 262 | 15 | 163 | 0.86 | 297 | 12 | 215 | 0.14 | 74 | 16 | 82 | 0.28 | 146 | 8 | 206 | 0.09 | 830 | |
5 | 136 | 0.29 | 270 | 8 | 212 | 0.77 | 408 | 13 | 197 | 0.14 | 79 | 15 | 201 | 0.24 | 184 | 5 | 210 | 0.09 | 835 | |
9 | 39 | 0.28 | 290 | 19 | 170 | 0.74 | 470 | 16 | 82 | 0.14 | 86 | 19 | 20 | 0.20 | 271 | 15 | 202 | 0.09 | 866 | |
12 | 215 | 0.26 | 339 | 15 | 202 | 0.72 | 530 | 2 | 219 | 0.13 | 121 | 12 | 215 | 0.18 | 317 | 18 | 232 | 0.09 | 880 | |
TatB | 18 | 21 | 0.83 | 7 | 18 | 21 | 2.19 | 6 | 18 | 21 | 0.23 | 9 | 12 | 198 | 0.72 | 12 | 12 | 198 | 0.61 | 41 |
12 | 215 | 0.60 | 30 | 12 | 215 | 1.45 | 22 | 12 | 198 | 0.18 | 14 | 12 | 202 | 0.63 | 23 | 12 | 202 | 0.47 | 97 | |
12 | 202 | 0.56 | 36 | 12 | 198 | 1.39 | 24 | 18 | 24 | 0.17 | 16 | 18 | 21 | 0.61 | 29 | 18 | 21 | 0.41 | 125 | |
20 | 185 | 0.46 | 65 | 12 | 202 | 1.13 | 63 | 12 | 202 | 0.16 | 17 | 18 | 24 | 0.53 | 47 | 18 | 24 | 0.32 | 194 | |
12 | 198 | 0.39 | 110 | 19 | 13 | 1.04 | 89 | 12 | 215 | 0.16 | 21 | 12 | 215 | 0.49 | 55 | 5 | 212 | 0.20 | 350 | |
5 | 213 | 0.36 | 127 | 5 | 212 | 0.97 | 117 | 17 | 227 | 0.14 | 71 | 19 | 13 | 0.39 | 79 | 12 | 215 | 0.20 | 355 | |
7 | 204 | 0.35 | 141 | 5 | 126 | 0.87 | 164 | 14 | 167 | 0.14 | 72 | 14 | 24 | 0.25 | 175 | 4 | 206 | 0.16 | 462 | |
5 | 212 | 0.28 | 281 | 18 | 24 | 0.81 | 221 | 5 | 213 | 0.13 | 88 | 19 | 198 | 0.22 | 225 | 5 | 208 | 0.14 | 529 | |
TatA with TatAbasal | 12 | 202 | 0.75 | 28 | 12 | 202 | 1.85 | 26 | 16 | 198 | 0.31 | 10 | 16 | 198 | 0.80 | 8 | 12 | 202 | 0.47 | 94 |
16 | 198 | 0.70 | 35 | 16 | 198 | 1.26 | 62 | 12 | 202 | 0.30 | 11 | 12 | 202 | 0.77 | 14 | 16 | 198 | 0.43 | 111 | |
15 | 201 | 0.60 | 58 | 8 | 215 | 1.01 | 130 | 15 | 201 | 0.22 | 40 | 15 | 201 | 0.68 | 36 | 15 | 201 | 0.39 | 136 | |
8 | 215 | 0.58 | 64 | 15 | 201 | 1.01 | 131 | 11 | 212 | 0.17 | 126 | 8 | 215 | 0.18 | 309 | 12 | 201 | 0.08 | 644 | |
19 | 194 | 0.38 | 141 | 12 | 215 | 0.69 | 378 | 8 | 215 | 0.16 | 143 | 14 | 203 | 0.14 | 419 | 8 | 215 | 0.06 | 832 | |
8 | 205 | 0.27 | 309 | 5 | 136 | 0.67 | 426 | 12 | 215 | 0.16 | 159 | 8 | 208 | 0.09 | 605 | 4 | 205 | 0.06 | 852 | |
TatB with TatAbasal | 18 | 21 | 0.48 | 72 | 18 | 21 | 1.42 | 44 | 18 | 21 | 0.23 | 19 | 12 | 198 | 0.65 | 36 | 12 | 198 | 0.48 | 94 |
12 | 198 | 0.46 | 79 | 12 | 198 | 0.99 | 173 | 12 | 198 | 0.19 | 37 | 12 | 202 | 0.56 | 55 | 4 | 206 | 0.43 | 119 | |
12 | 202 | 0.38 | 123 | 12 | 202 | 0.84 | 281 | 12 | 202 | 0.19 | 39 | 18 | 21 | 0.56 | 56 | 18 | 21 | 0.28 | 214 | |
20 | 185 | 0.37 | 134 | 12 | 215 | 0.76 | 383 | 18 | 24 | 0.18 | 44 | 15 | 201 | 0.34 | 122 | 12 | 202 | 0.28 | 218 | |
12 | 215 | 0.33 | 180 | 15 | 201 | 0.73 | 427 | 14 | 167 | 0.17 | 69 | 4 | 206 | 0.28 | 168 | 4 | 207 | 0.28 | 225 | |
5 | 213 | 0.30 | 252 | 5 | 212 | 0.68 | 514 | 5 | 213 | 0.17 | 88 | 18 | 24 | 0.27 | 175 | 4 | 205 | 0.16 | 415 | |
TatC-TatC | 64 | 134 | 0.79 | 33 | 27 | 199 | 2.04 | 39 | 64 | 134 | 0.28 | 51 | 64 | 134 | 0.6 | 62 | 64 | 134 | 0.64 | 60 |
71 | 140 | 0.77 | 41 | 65 | 137 | 1.72 | 62 | 71 | 140 | 0.26 | 75 | 71 | 140 | 0.54 | 84 | 27 | 199 | 0.47 | 108 | |
65 | 137 | 0.64 | 69 | 71 | 140 | 1.63 | 72 | 65 | 137 | 0.21 | 191 | 27 | 199 | 0.48 | 103 | 65 | 137 | 0.3 | 227 | |
28 | 203 | 0.51 | 10864 | 64 | 134 | 1.51 | 93 | 113 | 228 | 0.19 | 319 | 65 | 137 | 0.39 | 150 | 64 | 137 | 0.24 | 283 | |
72 | 217 | 0.42 | 155 | 64 | 157 | 1.05 | 284 | 67 | 140 | 0.18 | 345 | 28 | 203 | 0.37 | 162 | 33 | 167 | 0.23 | 289 | |
Table 2
Strains used in this study.
Strain name* | Abbreviation | Genotype | Reference |
|---|---|---|---|
MC4100 | ABCE | F-, ΔlacU169, araD139, rpsL150, relA1, ptsF, rbsR, flbB5301 | |
MC4100-A | ABCE | arabinose-resistant derivative of MC4100 | |
J1M1 | ABC | MC4100 ΔtatE | |
| ELV16 | BCE | MC4100 ΔtatA | |
B1LK0 | ABE | MC4100 ΔtatC | |
MΔBC | AE | MC4100 ΔtatBC | |
DADE-A | Δtat | MC4100 ΔtatABC ΔtatE | |
MΔABC-A λAry | AyE | MC4100-A ΔtatABC::apra, attB::PtatAtatA-EAK-eyfpA206K(kanr) | |
JARV16 λA(Q8A) | AQ8ABC | MC4100 ΔtatA ΔtatE, attB::PtatAtatAQ8A (kanr) | This work |
MΔABC-A λA(Q8A) | AQ8AE | MC4100-A ΔtatABC::apra, attB::PtatAtatAQ8A(kanr) | This work |
ELV16 λA(Q8A) | AQ8ABCE | MC4100 ΔtatA, attB::PtatAtatAQ8A (kanr) | This work |
ELV16 λAry(Q8A) | AyQ8ABCE | MC4100 ΔtatA, attB::PtatAtatAQ8A-EAK-eyfpA206K (kanr) | This work |
ELV16-A λAry | AyBCE | MC4100-A ΔtatA, attB::PtatAtatA-EAK-eyfpA206K (kanr) | |
JARV16 λAry | AyBC | MC4100 ΔtatA ΔtatE, attB::PtatAtatA-EAK-eyfpA206K(kanr) | |
JARV16 λAry(Q8A) | AyQ8ABC | MC4100 ΔtatA ΔtatE, attB::PtatAtatAQ8A-EAK-eyfpA206K(kanr) | This work |
-
*All strains designated `–A’ are arabinose-resistant derivatives (Ize et al., 2002)
Table 3
Plasmids used in this study.
Plasmid name | Abbreviation | Description | Reference |
|---|---|---|---|
pTH19cr | Low copy number pSC101-derived replicon. Chlr. | ||
p101C*TatBC | pBC | pTH19cr derivative. Expression of tatBC from the tatA promoter with a modified RBS. | |
p101C*BC M205R | pBCM205R | p101C*TatBC tatC-met205arg | This work |
p101C*BC Q215A | pBCQ215A | p101C*TatBC tatC-gln215ala | This work |
p101C*BC MTQ | pBCMTQ | p101C*TatBC tatC- met205ala- thr208ala- gln215ala | This work |
p101C*BC M205A | pBCM205A | p101C*TatBC tatC- met205ala | This work |
p101C*BC T208A | pBCT208A | p101C*TatBC tatC- thr208ala | This work |
p101C*BC TQA | pBCT208A,Q215A | p101C*TatBC tatC- thr208ala- gln215ala | This work |
p101C*BC FEA | pBCFEA | p101C*TatBC tatC- Phe 94ala- glu103ala | |
p101C*BC FEAMR | pBCFEA, M205R | p101C*BC FEA tatC-met205arg | This work |
p101C*BC FEAQA | pBCFEA, Q215A | p101C*BC FEA tatC-gln215ala | This work |
p101C*BC FEAMTQ | pBCFEA, MTQ | p101C*BC FEA tatC- met205ala- thr208ala- gln215ala | This work |
p101C*BC FEAMA | pBCFEA,M205A | p101C*BC FEA tatC- met205ala | This work |
p101C*BC FEATA | pBCFEA,T208A | p101C*BC FEA tatC- thr208ala | This work |
p101C*BC FEATQA | pBCFEA,T208A, Q215A | p101C*BC FEA tatC- thr208ala- gln215ala | This work |
p101C*BC E8A | pBE8AC | p101C*TatBC tatB-glu8ala | This work |
p101C*BC EAFEA | pBE8ACFEA | p101C*BC FEA tatB-glu8ala | This work |
p101C*BC EFM | pBE8ACFEA, MTQ | p101C*BC FEAMTQ tatB-glu8ala | This work |
p101CTatC | pTH19cr derivative expressing tatC from the tatA promoter | This work | |
p101C*TatC | pC | p101CTatC with a modified RBS and GTG start codon. | This work |
p101C*TatC FEA | pCFEA | p101C*TatC tatC- Phe 94ala- glu103ala | This work |
p101C*BC V18W | pBV18WC | p101C*TatBC tatB-val18trp | This work |
p101C*BC L21W | pBCL21W | p101C*TatBC tatC-leu21trp | This work |
p101C*BC VLW | pBV18WCL21W | p101C*TatBC V18W tatC-leu21trp | This work |
pTat101 cys less | Very low copy number vector expressing tatABCC23A,C33A,C179A,C224A from the tat promoter. Kanr. | ||
p101C*BC cys less | pBC cys- | p101C*TatBC tatC-cys23ala-cys33ala-cys179ala-cys224ala | This work |
p101C*BC V18C | pBV18CC | p101C*BC cys less tatB-val18cys | This work |
p101C*BC L21C | pBCL21C | p101C*BC cys less tatC-leu21cys | This work |
p101C*BC 17C 21C | pBL17CCL21C | p101C*BC L21C tatB-leu17cys | This work |
p101C*BC 18C 21C | pBV18CCL21C | p101C*BC L21C tatB-val18cys | This work |
p101C*BC 17C 22C | pBL17CCN22C | p101C*BC cys less tatB-leu17cys tatC-asn22cys | This work |
p101C*BC 18C 22C | pBV18CCN22C | p101C*BC V18C tatC-asn22C | This work |
p101C*BC 17C 23C | pBL17CCC23C | p101C*BC cys less tatB-leu17cys tatC-ala23cys | This work |
p101C*BC 18C 23C | pBV18CCC23C | p101C*BC V18C tatC-ala23cys | This work |
p101C*BC 17C 24C | pBL17CCI24C | p101C*BC cys less tatB-leu17cys tatC-ile24cys | This work |
p101C*BC 18C 24C | pBV18CCI24C | p101C*BC V18C tatC-ile24cys | This work |
p101C*BC 17C 25C | pBL17CCI25C | p101C*BC cys less tatB-leu17cys tatC-ile25cys | This work |
p101C*BC 18C 25C | pBV18CCI25C | p101C*BC V18C tatC-ile25cys | This work |
pUnitat2 | Expression of tatABChis under the control of a T5 promoter | ||
pUT2DH | Expression of tatABC under the control of a T5 promoter. | This work | |
pUT2DH FEA | pUT2DH tatC- Phe 94ala- glu103ala | This work | |
pUT2DH FEAMR | pUT2DH FEA tatC-met205arg | This work | |
pUT2DH EAFEA | pUT2DH FEA tatB-glu8ala | This work | |
pQE80-CueO | Synthesis of E. coli CueO with a C-terminal his6 tag. | ||
pKSUniA | tatA under control of the tat promoter in pBluescript KS+ | ||
pKSUniA Q8A | pKSUniA tatA-gln8ala | This work | |
pRS552 | lambda attachment site shuttle vector | ||
p552TatA Q8A | pRS552 carrying PtatAtatAQ8A | This work |
Additional files
-
Supplementary file 1
Molecular model for the (TatBC)3 complex in PDB format.
- https://doi.org/10.7554/eLife.20718.027
-
Supplementary file 2
Molecular model for the (TatBC)4 complex in PDB format.
- https://doi.org/10.7554/eLife.20718.028
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Assembling the Tat protein translocase
eLife 5:e20718.
https://doi.org/10.7554/eLife.20718




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}