The dynamic assembly of distinct RNA polymerase I complexes modulates rDNA transcription
- Centro de Investigaciones Biológicas, Spain
- Institute for Research in Biomedicine, Spain
- The Barcelona Institute of Science and Technology, Spain
- CSIC-Universidad de Salamanca, Spain
- Cooperative Center for Research in Biosciences CIC bioGUNE, Spain
Figures
Figure 1 with 5 supplements
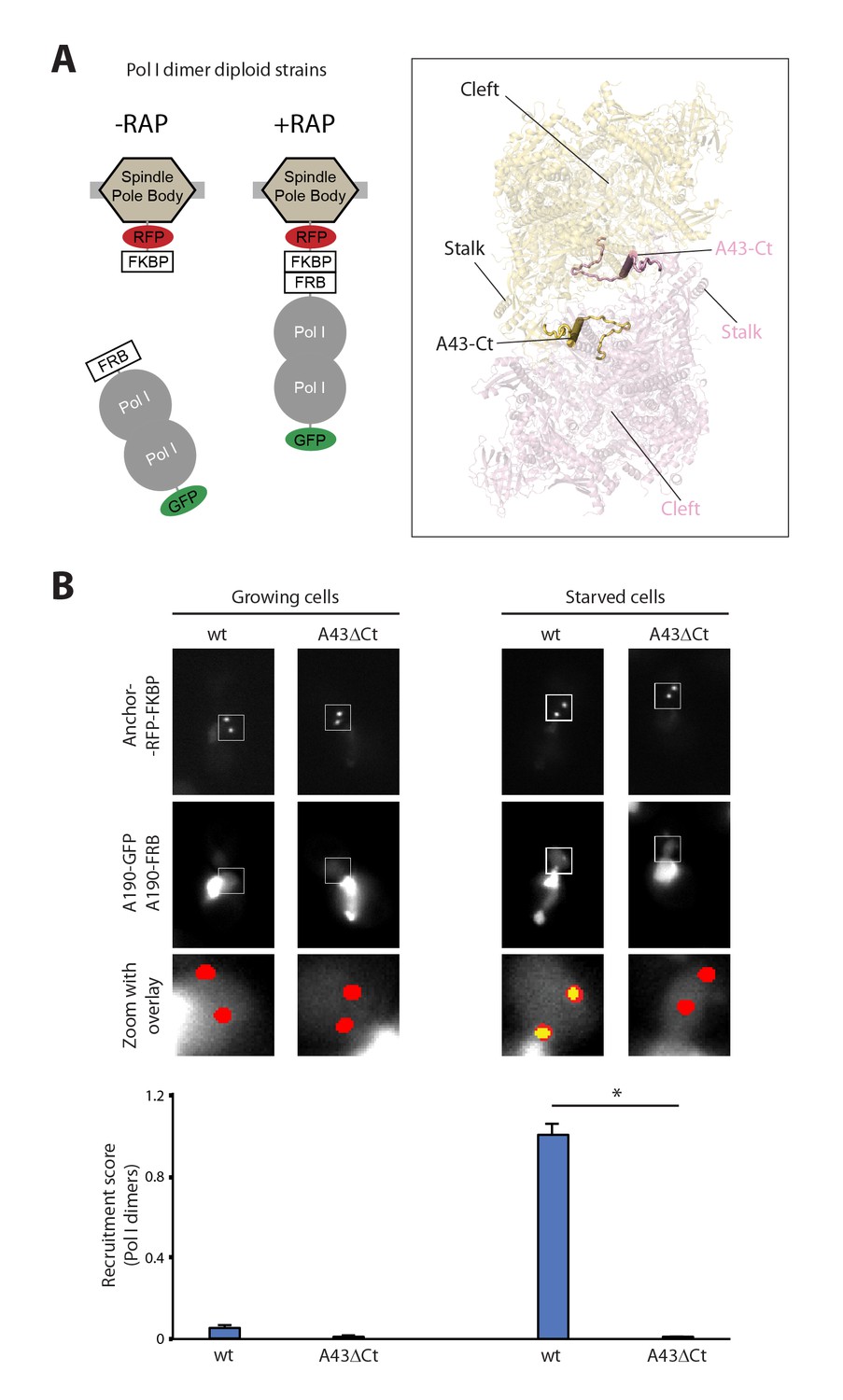
Live-cell imaging of Pol I homodimerization.
(A) Scheme of the diploid strains designed to study Pol I homodimers in vivo. The crystal structure of inactive Pol I homodimers, critically maintained by the A43 C-terminal tail, is shown on the right with monomers in yellow and pink. (B) Representative PICT images of the RFP-tagged anchor (upper row), GFP-tagged A190 (middle row) and a zoom of a 2.6 × 2.6 µm square around the anchoring platforms (bottom row). Below, quantification of the A190-GFP recruitment score, normalized to the measurement of the wild-type strain in starving medium (Mean ± SD, p-value * < 0.01 t-test).
Figure 1—figure supplement 1
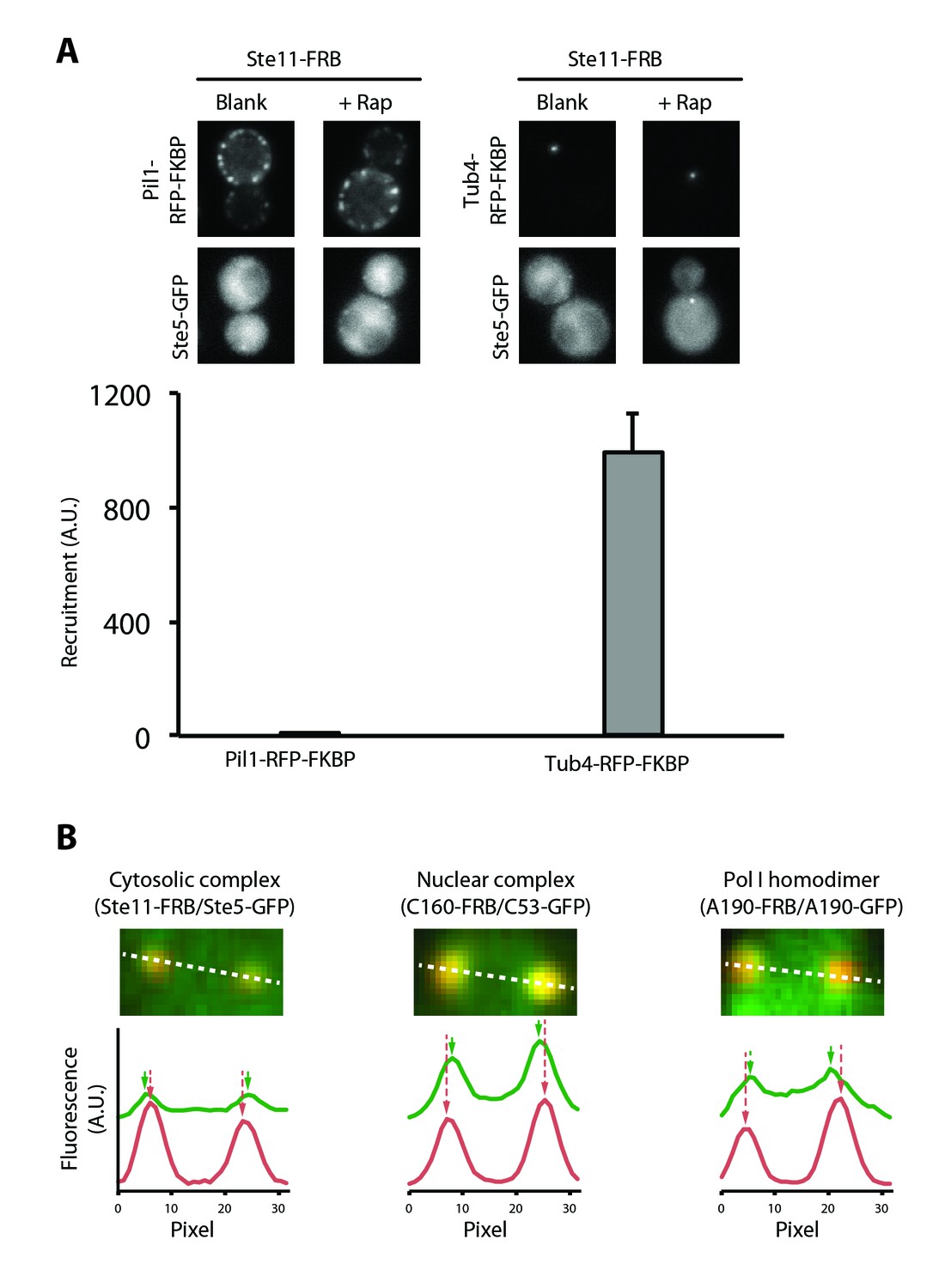
A more sensitive PICT assay to detect cytosolic and nuclear complexes.
(A) Comparison of Pil1-RFP-FKBP (left) and Tub4-RFP-FKBP (right) anchoring platforms to detect the Ste5-Ste11 complex. Ste11-FRB was used as bait. Representative images of the anchor-RFP-FKBP (upper row) and Ste5-GFP (bottom row). Below, quantification of Ste5-GFP recruitment to the corresponding anchoring platform in rapamycin-treated cells (Mean ± SD). (B) PICT assay to detect nuclear and cytosolic complexes. Above, close-up view around two anchoring platforms in a representative cell. Below, intensity profile through the white dashed line for the prey-GFP (green) and Tub4-RFP-FKBP (red) channels, with arrows indicating the brightest pixel for each peak of intensity.
Figure 1—figure supplement 2

Methodology for PICT quantification.
Segmentation workflow for RFP-anchors (red channel, left column) and prey-GFP spots (green channel, right column). For both channels the same operations are performed: (1) raw images, (2) noise and background attenuation by Gaussian blurring followed by top hat opening (ImageJ ‘subtract background’), (3) local mean threshold with radius adjusted to expected spot size, and (4) small and large particle removal by median filtering and area opening (area adjusted to expected spot size). The bottom row image shows an overlay of the raw GFP image, segmented RFP-anchors (red) and segmented prey-GFP spots (green), yellow pixels correspond to overlap between prey-GFP spots and RFP-anchors. The recruitment score is computed as the ratio between GFP mean intensity measured inside yellow pixel regions multiplied by the summed yellow area divided by the summed red area. In this image no false GFP positive spot counts toward the recruitment score since none overlaps with a segmented anchor. For both channels, RFP-anchors are indicated by red arrows.
Figure 1—figure supplement 3
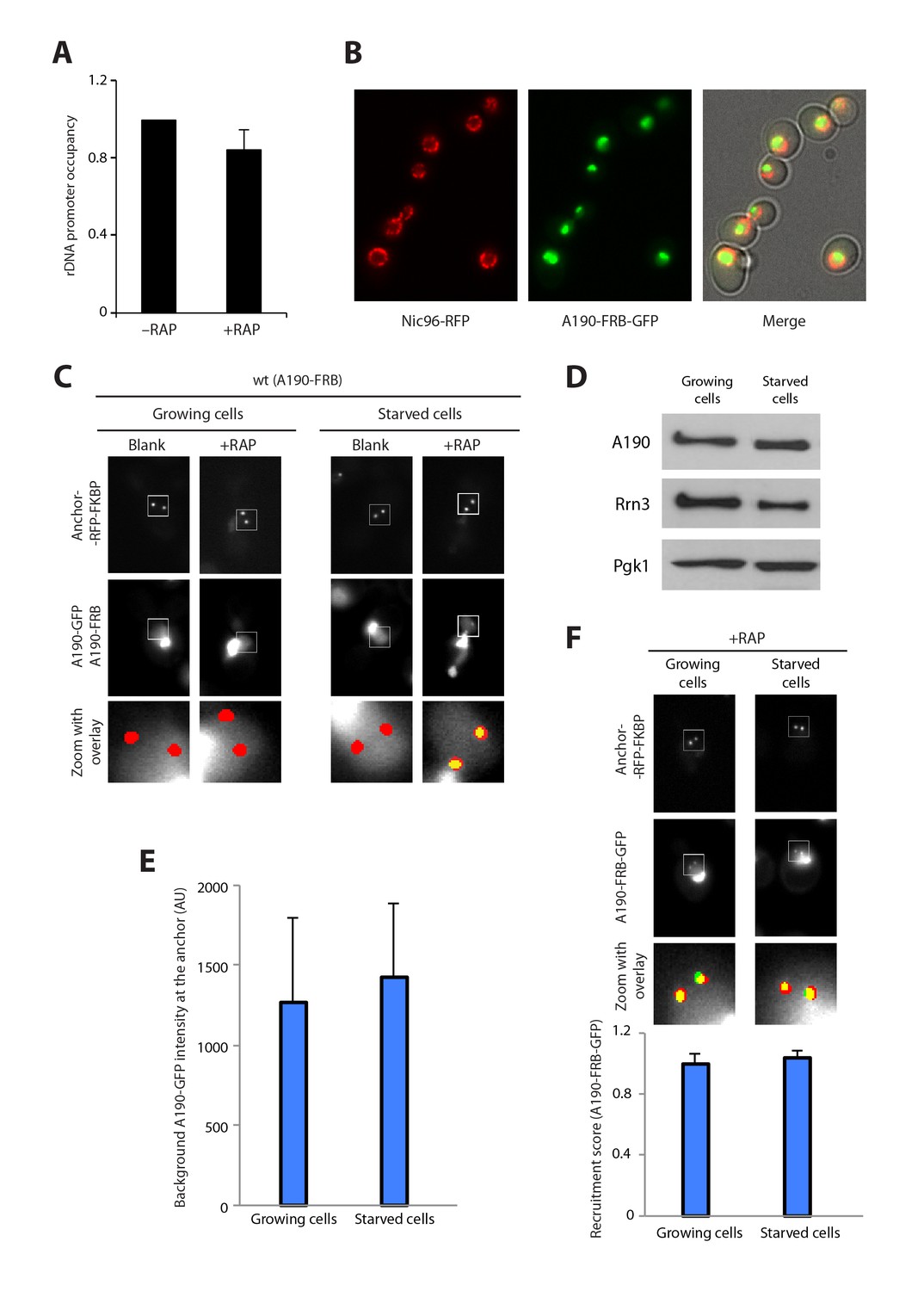
Additional control experiments.
(A) ChIP of A190-FRB + A190 GFP strain in the absence and presence of rapamycin (Mean ± SD). T-test shows that the difference is not statistically significant. (B) Visualization of the nuclear envelope by labelling of Nuc96. The A190-FRB-GFP signal is confined within the nuclear space. (C) Control experiments for live cell imaging of Pol I homodimerization using PICT. (D) Western-blot analysis of Pol I and Rrn3 levels in cells growing in rich and starving medium, using rabbit anti-A190 (gift of Michel Riva) and mouse anti-GFP (Takara, JL-8) antibodies, respectively. (E) Quantification of the A190-GFP background signal using PICT (Mean ± SD). (F) Quantification of A190-FRB-GFP recruitment to anchoring platforms in growing and starving cells (Mean ± SD).
Figure 1—figure supplement 4
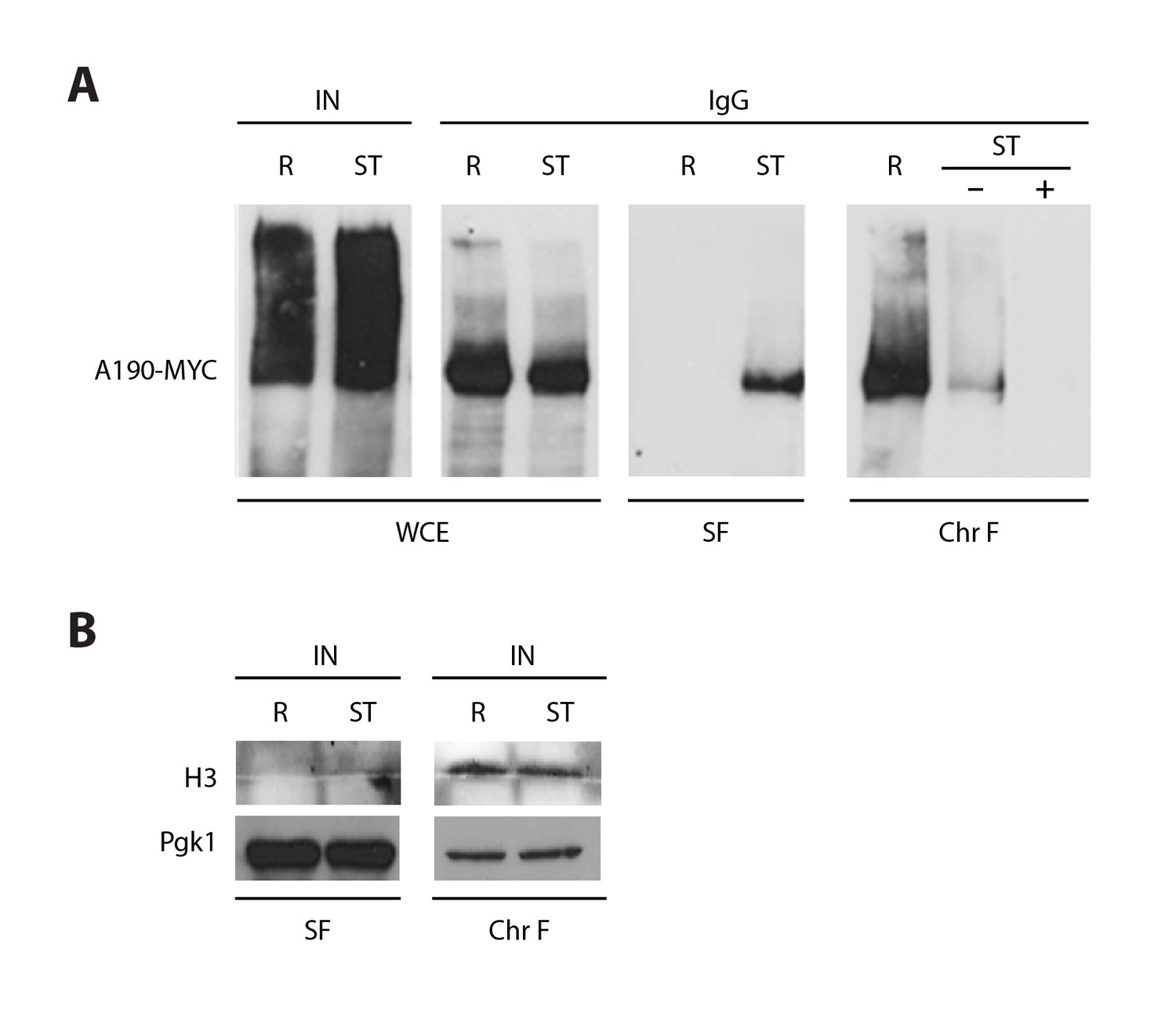
Co-immunoprecipitation on Pol I dimerization.
(A) Whole cell extracts (WCE) were obtained from crosslinked cells grown in rich (R) or starving media (ST). Centrifugation of the cell lysate allowed the separation of the protein soluble fraction (SF) and the pellet with the chromatin insoluble fraction and associated proteins, which was thereafter solubilized (Chr F). To analyze dimer formation, A190-TAP was precipitated from both fractions (SF and Chr F) and the amount of associated A190-MYC analyzed by western blot. The ST sample of Chr F was incubated in the presence and absence of DNase I (‘+’ and ‘−’) after immunoprecipitation. A control experiment with WCE was used to show that A190-TAP is efficiently immunoprecipitated in both conditions, R and ST, where IN is the input before immunoprecipitation and IgG the precipitated samples after being decrosslinked. (B) The quality of both fractions was verified by western blot of the chromatin associated protein histone H3, present in Chr F only, and the cytosolic protein Pgk1, present in SF mostly.
Figure 1—figure supplement 5
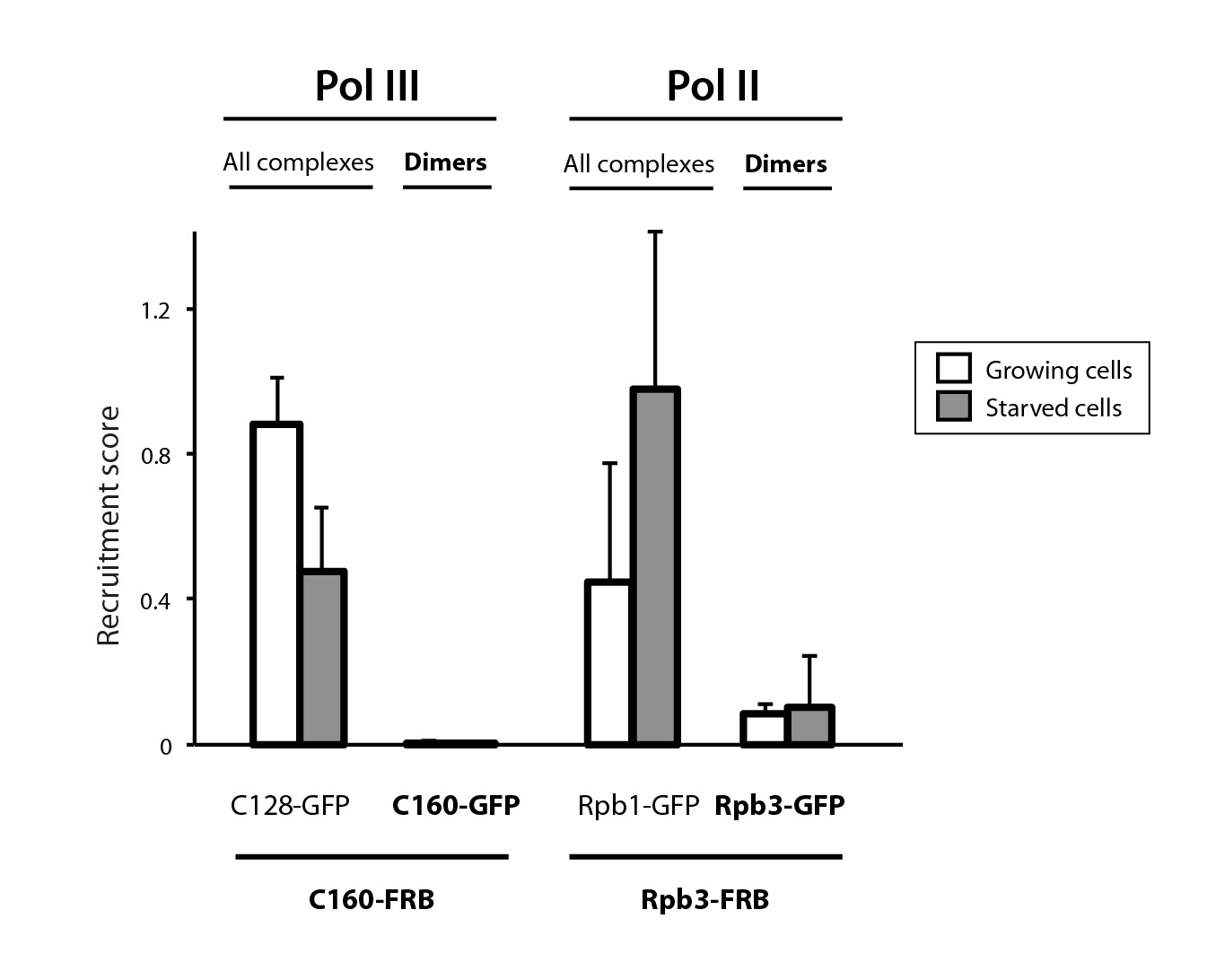
Analysis of Pol II and Pol III complexes.
Quantification of detected Pol III and Pol II complexes in diploid yeast strains (Mean ± SD).
Figure 2
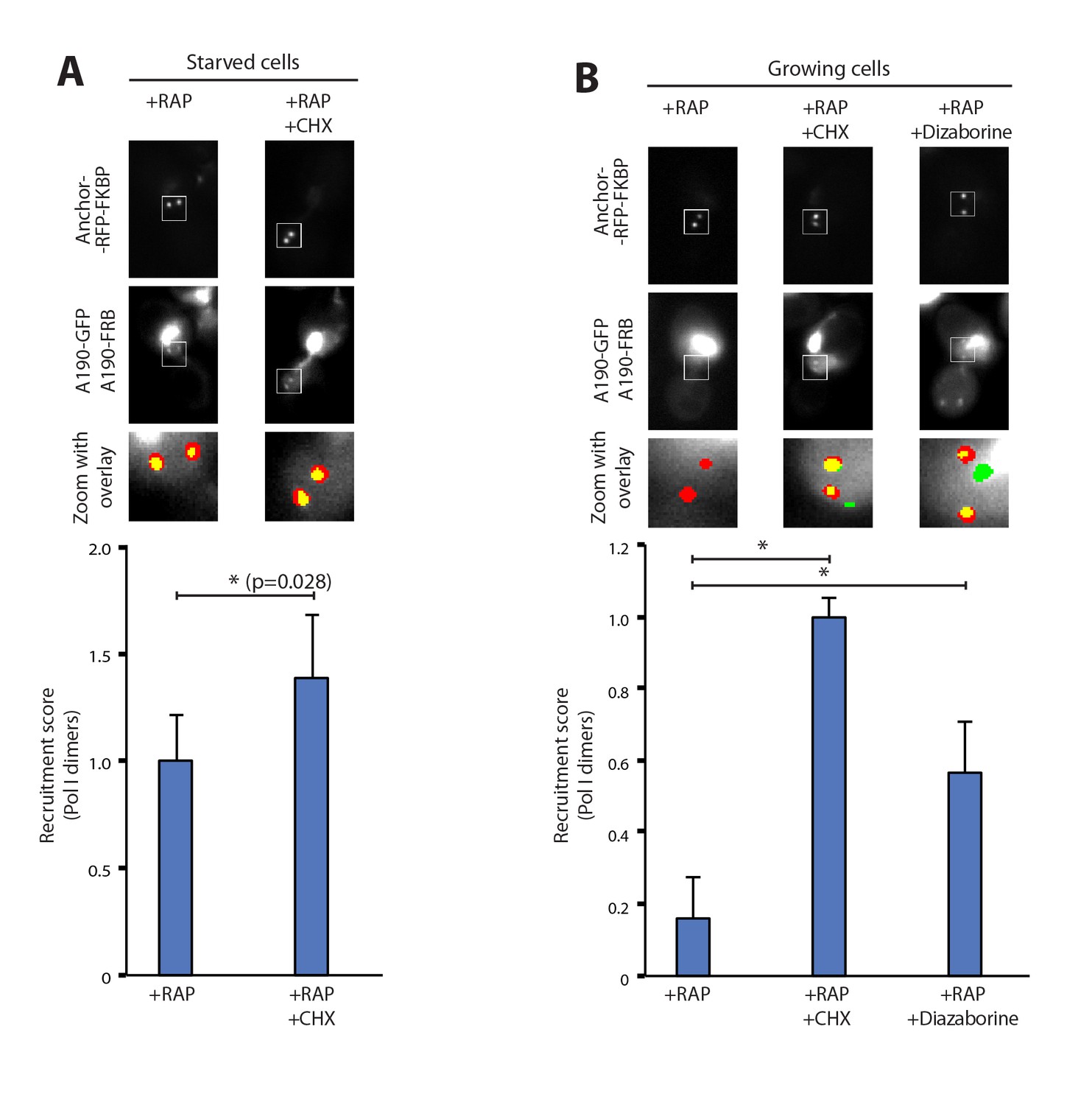
Pol I homodimerization upon ribosome perturbation.
Representative PICT images of the RFP-tagged anchor (upper row), GFP-tagged A190 (middle row) and a zoom of a 2.3 × 2.3 µm square around anchoring platforms (bottom row). Below, quantification of the A190-GFP recruitment score, normalized to the measurement of untreated cells (Mean ± SD, p-value * < 0.01 t-test). (A) Effect of cycloheximide (CHX; 0.2 µg/ml) in starved cells. (B) Effect of cycloheximide (0.2 µg/ml) and diazaborine (10 µg/ml) in growing cells.
Figure 3 with 2 supplements

Live-cell imaging of Pol I–Rrn3 complexes.
(A) Scheme of the haploid strains designed to study Pol I–Rrn3 in vivo. (B) Representative PICT images of the RFP-tagged anchor (upper row), GFP-tagged Rrn3 (middle row) and a zoom of a 2.3 × 2.3 µm square around anchoring platforms (bottom row). Below, quantification of the Rrn3-GFP recruitment score, normalized to the measurement in rich medium (Mean ± SD, p-value * < 0.01 t-test). (C) Schematic representation of the 35S rDNA. Below, approximate location of the primer pairs used for ChIP experiments in the following panel. (D) ChIP experiments showing the relative occupancy of A190 and Rrn3 on the rDNA gene in different culture media (Mean ± SD).
Figure 3—figure supplement 1
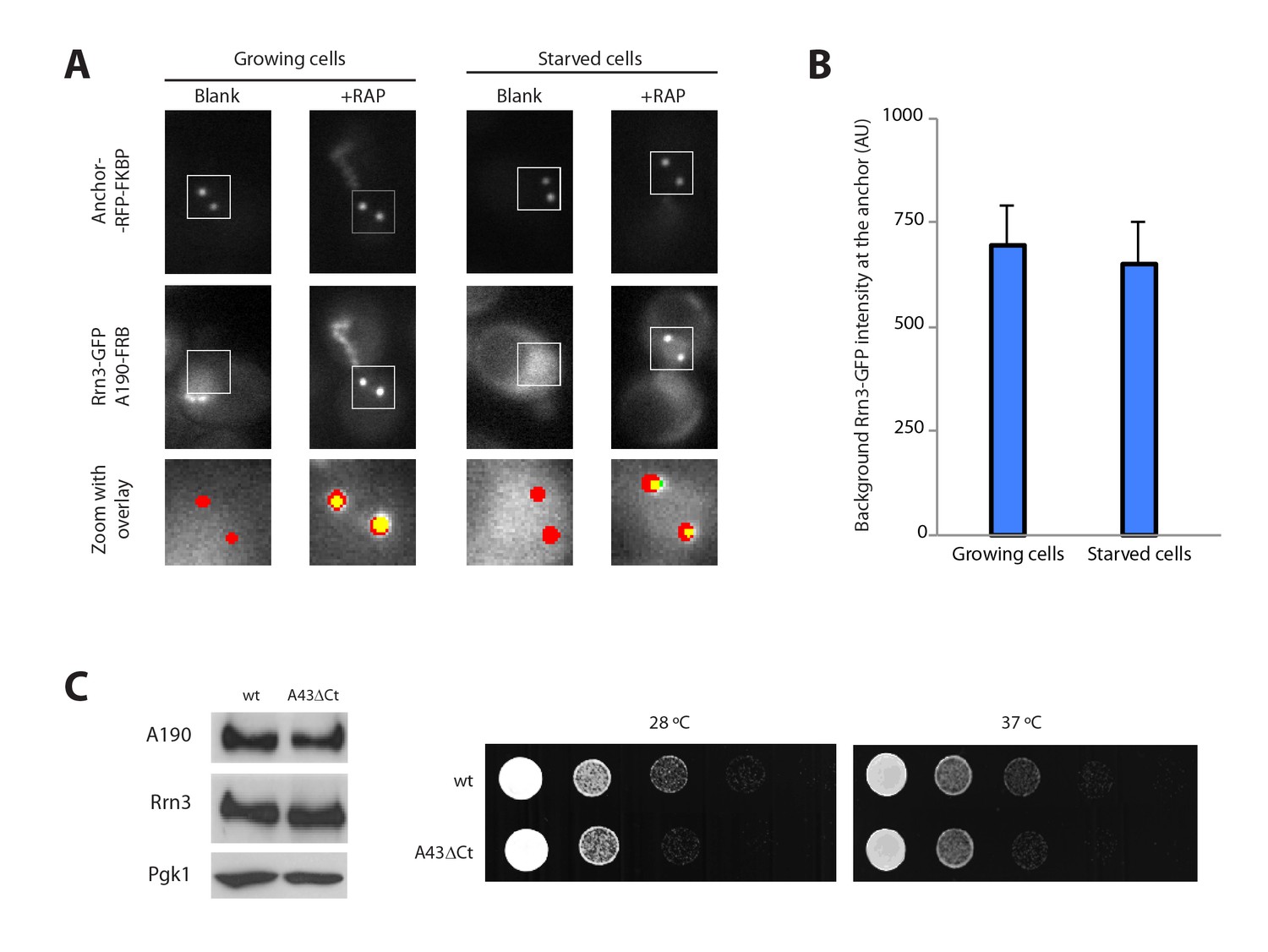
Additional control experiments.
(A) Control experiments for live cell imaging of Pol I–Rrn3 complexes using PICT. (B) Quantification of the Rrn3-GFP background signal using PICT (Mean ± SD). (C) Western-blot analysis of A190 and Rrn3 in whole cell extracts, using rabbit anti-A190 and mouse anti-GFP, respectively (left panel). Comparative growth phenotype at 28°C and 37°C, obtained by serial dilutions (1:10) of wild-type and A43ΔCt mutant strains spotted on either YPD or selective SC medium, and grown for 2–3 days (right panel).
Figure 3—figure supplement 2

Analysis of the CARA strain.
(A) ChIP experiments showing the relative association of A190 to the rDNA promoter in the CARA and wild-type strains, in different culture media (Mean ± SD). (B) Comparative growth phenotype at 28°C and 37°C for the CARA and wild-type strains in rich medium after growing the cells either in rich or starving conditions.
Figure 4 with 1 supplement
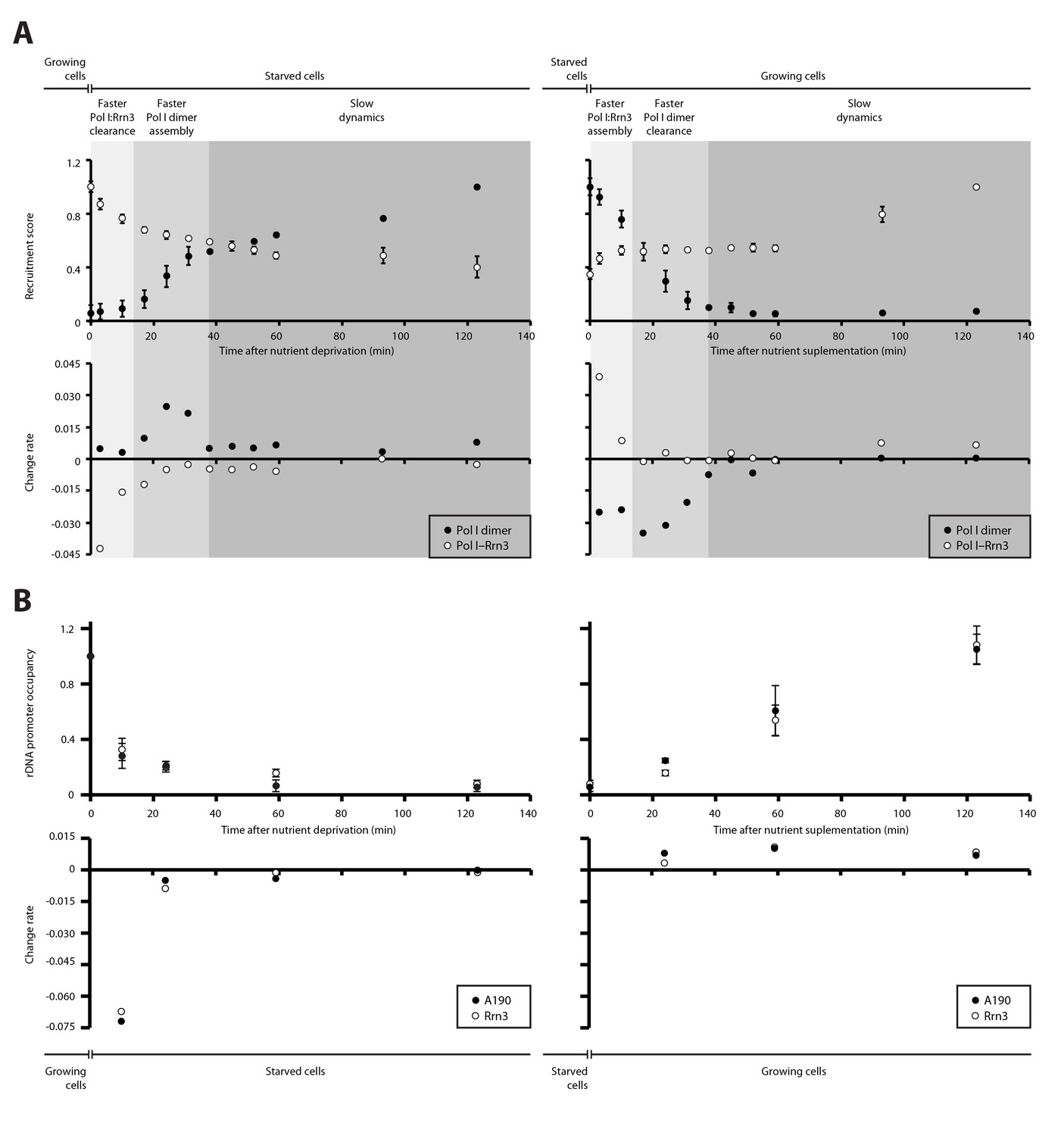
Dynamics in the cellular levels of Pol I complexes in response to nutrient availability.
(A) Upper plots show the relative levels of Pol I–Rrn3 complexes and Pol I homodimers as detected by PICT. Measurements were done at the indicated time points after growing cells were switched to starving medium (left) or cells starved for 2 hr were switched to nutrient-rich medium (right). Values were normalized to the highest measurement of the corresponding complex. Bottom plots show the rate of assembly (positive values) and disassembly (negative values) of each complex between two consecutive measurements. Grey shadows indicate the different stages observed in response to nutrient availability: faster adjustment of Pol I–Rrn3 levels (lighter grey), faster adjustment of Pol I homodimer levels (middle grey) and slow consolidation of the levels of each Pol I complex (dark grey). (B) Upper plots show the relative A190 and Rrn3 occupancy at the rDNA promoter for the indicated conditions and time points, as measured by ChIP. Values were normalized to the value obtained for cells growing in rich medium (Mean ± SD). Bottom plots show the rate of assembly and disassembly, calculated as in panel A.
Figure 4—figure supplement 1
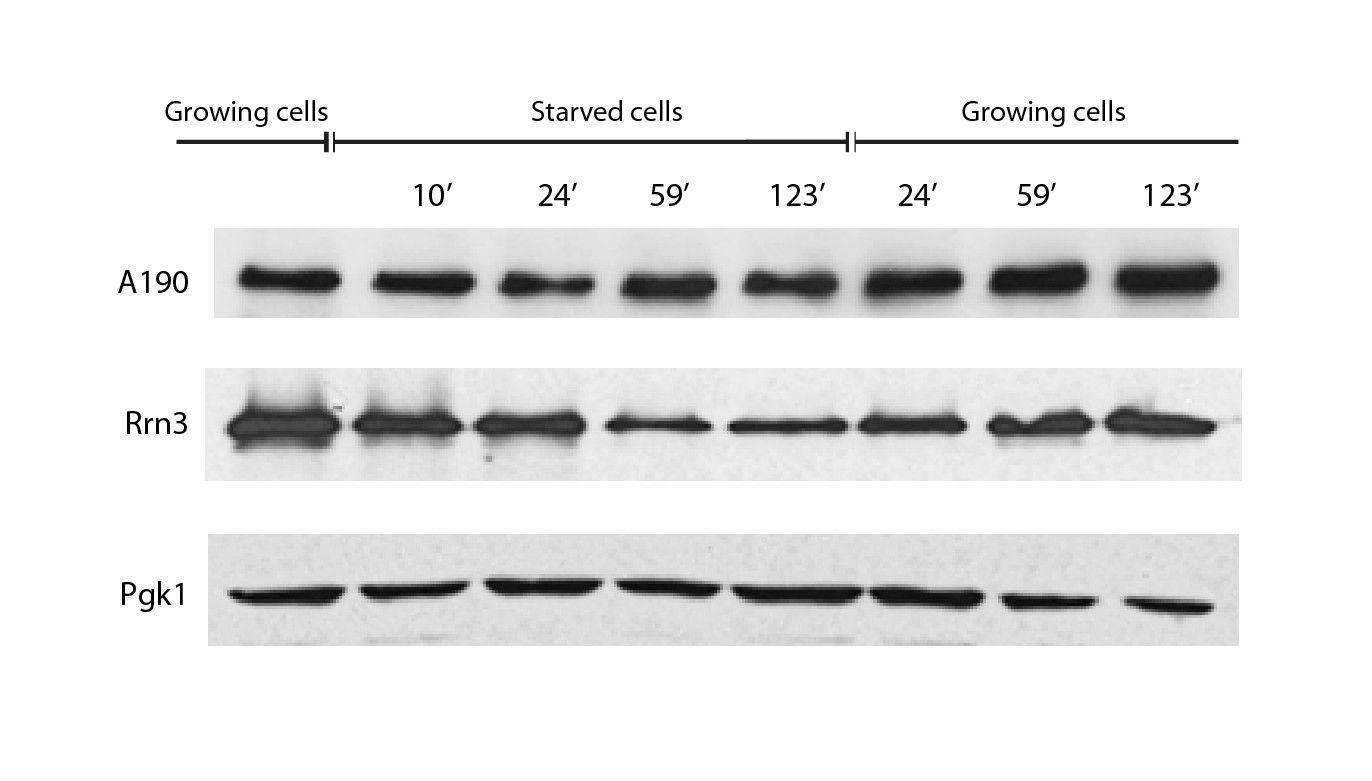
A190 and Rrn3 levels in response to nutrient availability.
Western-blot analysis of the A190 and Rrn3 levels in whole cell extract from wild-type cells grown in the indicated conditions and time points, using rabbit anti-A190 and mouse anti-GFP (Living colors, JL-8 Clontech) antibodies, respectively. Mouse anti-Pgk1 was used as a loading control.
Figure 5 with 3 supplements
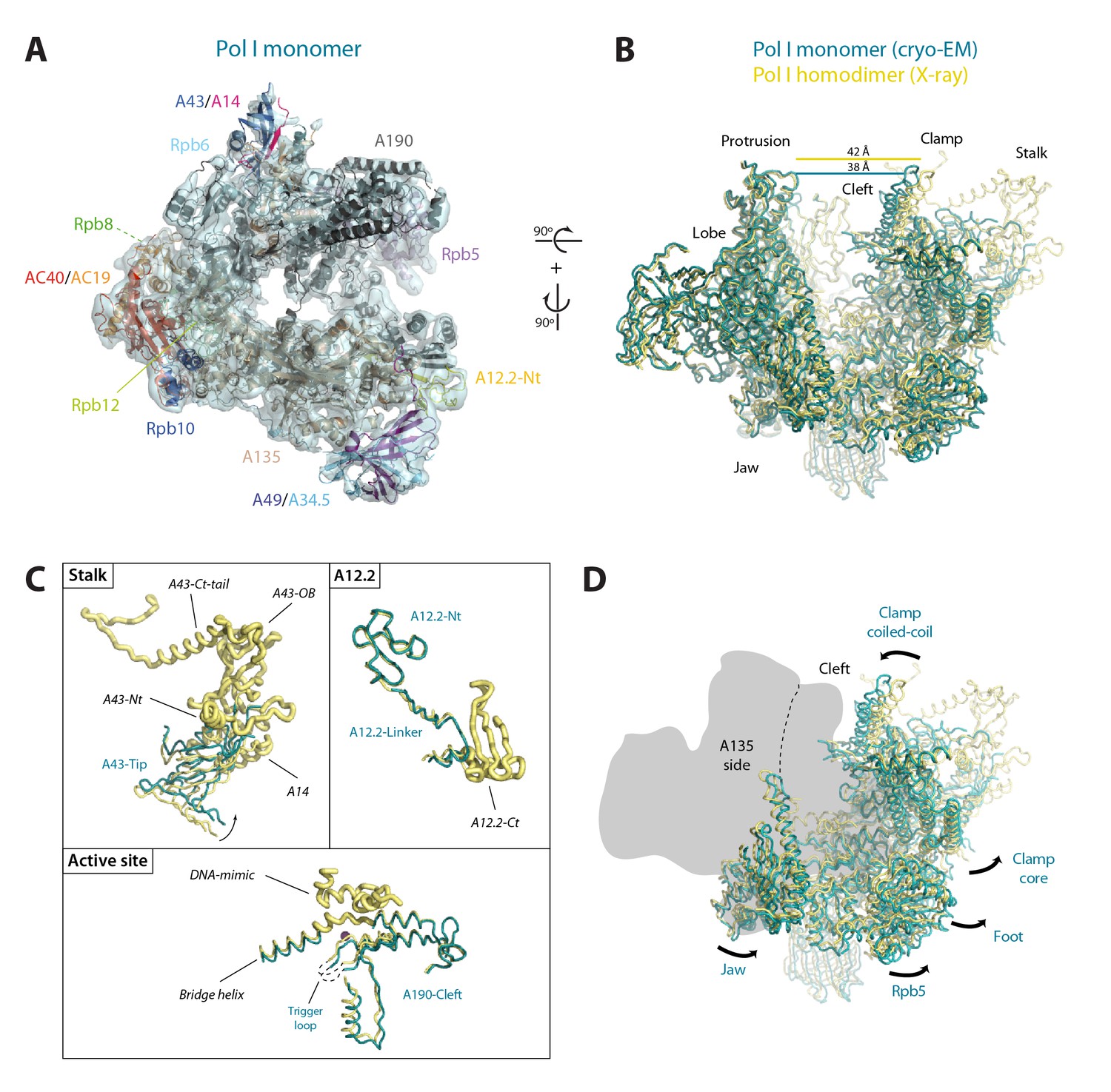
Structure of monomeric Pol I in solution.
(A) Cryo-EM reconstruction of Pol I at 4.9 Å resolution superposed with the derived pseudo-atomic model. (B) Comparison between the structural models of the Pol I dimer (PDB-4C3H) and the Pol I monomer (this report) in yellow and cyan, respectively, with labelled structural domains. (C) Close-up views of regions becoming flexible in the transition from dimeric to monomeric Pol I, in the same colors and orientation as in panel B. Flexible regions are depicted with thicker ribbon trace and labelled in italics. (D) Representation of the conformational changes associated with the transition from dimeric to monomeric Pol I, in the same colors and orientation as in panel B.
Figure 5—figure supplement 1
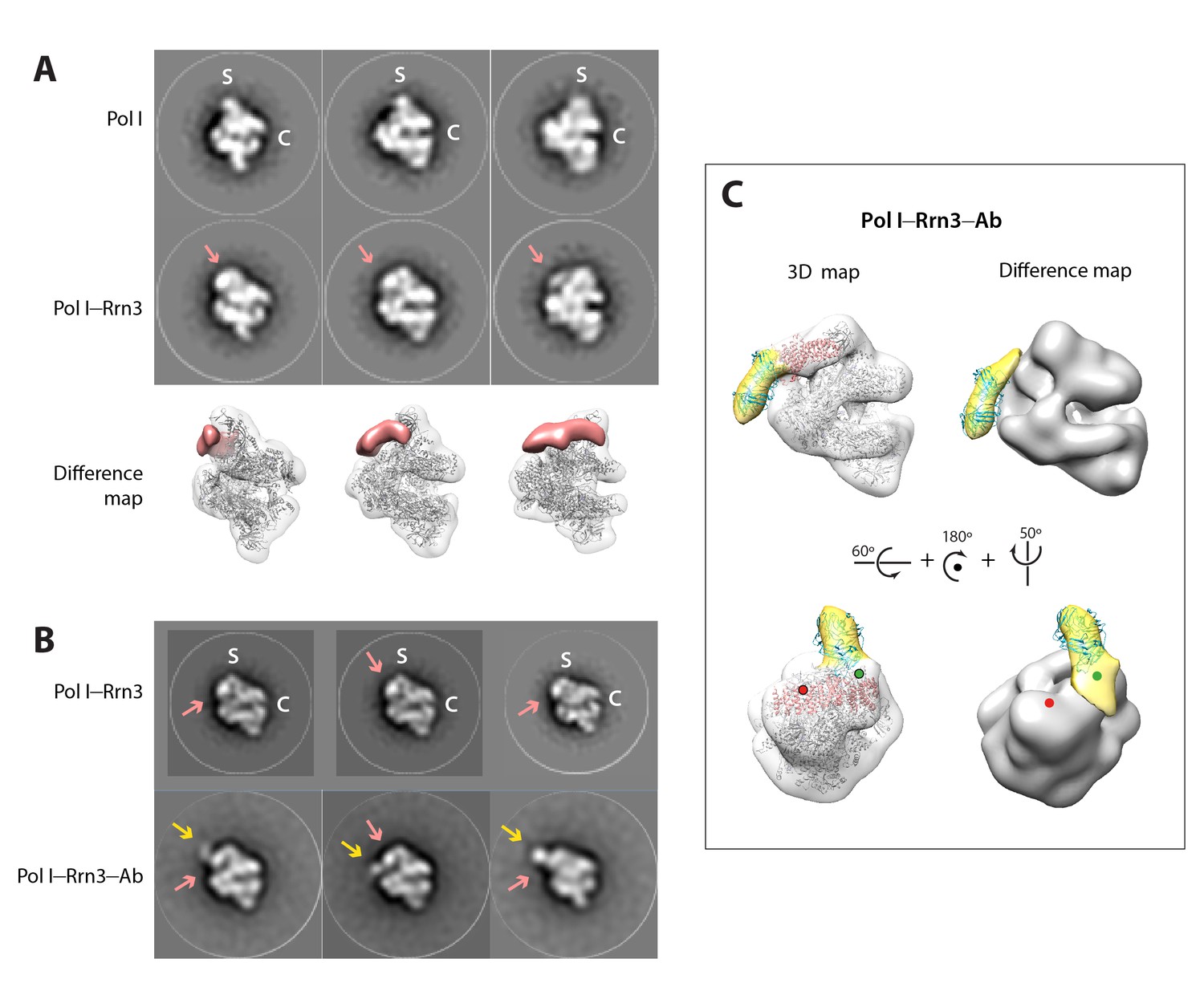
Negative-staining EM of the yeast Pol I–Rrn3 complex.
(A) Reference-free 2D averages of free Pol I (upper row) and Pol I in complex with Rrn3 (middle row). Averages derive from 2D classes containing between 100 and 200 images each. The cleft and stalk are labelled C and S. A salmon arrow indicates the position of the additional mass in the Pol I–Rrn3 complex. In the bottom row, difference map between the two previous reconstructions, showing the presence of Rrn3 in salmon. (B) Reference-free 2D averages of a Pol I–Rrn3 complex labelled with an anti-Strep-tag antibody bound to the C-terminus of Rrn3 (bottom row), compared with similar averages of the Pol I–Rrn3 complex (top row). Averages derive from 2D classes containing between 200 and 500 images each. Salmon and yellow arrows indicate densities assigned to Rrn3 and the Fab part of the antibody, respectively. (C) Reconstructed 3D volume of the Pol I–Rrn3–anti-Strep with fitted crystal structures of Pol I in grey, Rrn3 in salmon, and an Fab in cyan. In yellow is the density assigned to the Fab part of the antibody, as shown by difference mapping with the Pol I–Rrn3 map. Green and red spheres indicate the position of the Rrn3 C-terminus after fitting inside the Pol I–Rrn3 map in two opposite orientations, with only the green being compatible with antibody labelling.
Figure 5—figure supplement 2

Cryo-EM structure of the yeast Pol I–Rrn3 complex.
(A) Typical field of our cryo-EM grids. The scale bar represents 50 nm. (B) Initial reference-free 2D averages showing a significant level of detail. An arrow indicates the position of Rrn3. (C) Data processing strategy, showing the initial volume, the result of 3D classification, and the three refined maps with their corresponding local resolution and FSC curves.
Figure 5—figure supplement 3
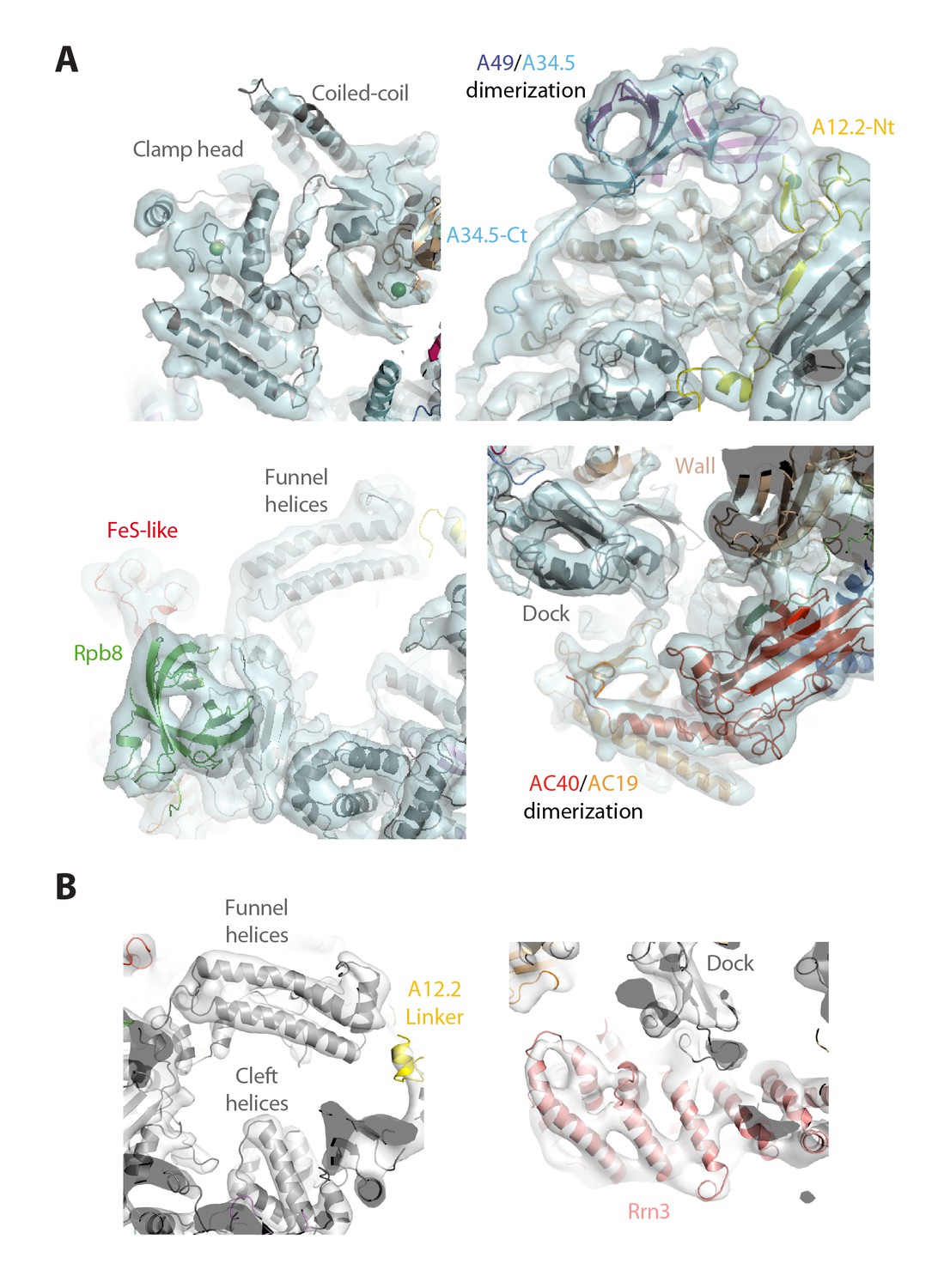
Structural details of the monomeric Pol I and Pol I–Rrn3 cryo-EM structures.
(A) Close-up views of significant regions in the structure of monomeric Pol I at 4.9 Å resolution: clamp (upper-left), lobe (upper-right), funnel (bottom-left), and groove (bottom-right). (B) Close-up views of significant regions in the structure of monomeric Pol I at 7.7 Å resolution: funnel (left) and Rrn3 (right).
Figure 6 with 1 supplement

Structure of the Pol I–Rrn3 complex.
(A) Cryo-EM reconstruction of Pol I in complex with Rrn3 (left) superposed with the derived pseudo-atomic model (right). The different Pol I structural domains and subunits are labelled. (B) Stalk fixation in the transition from free to Rrn3-bound Pol I, in a lateral view as indicated. Subunit A14 and the different domains in subunit A43 are indicated.
Figure 6—figure supplement 1
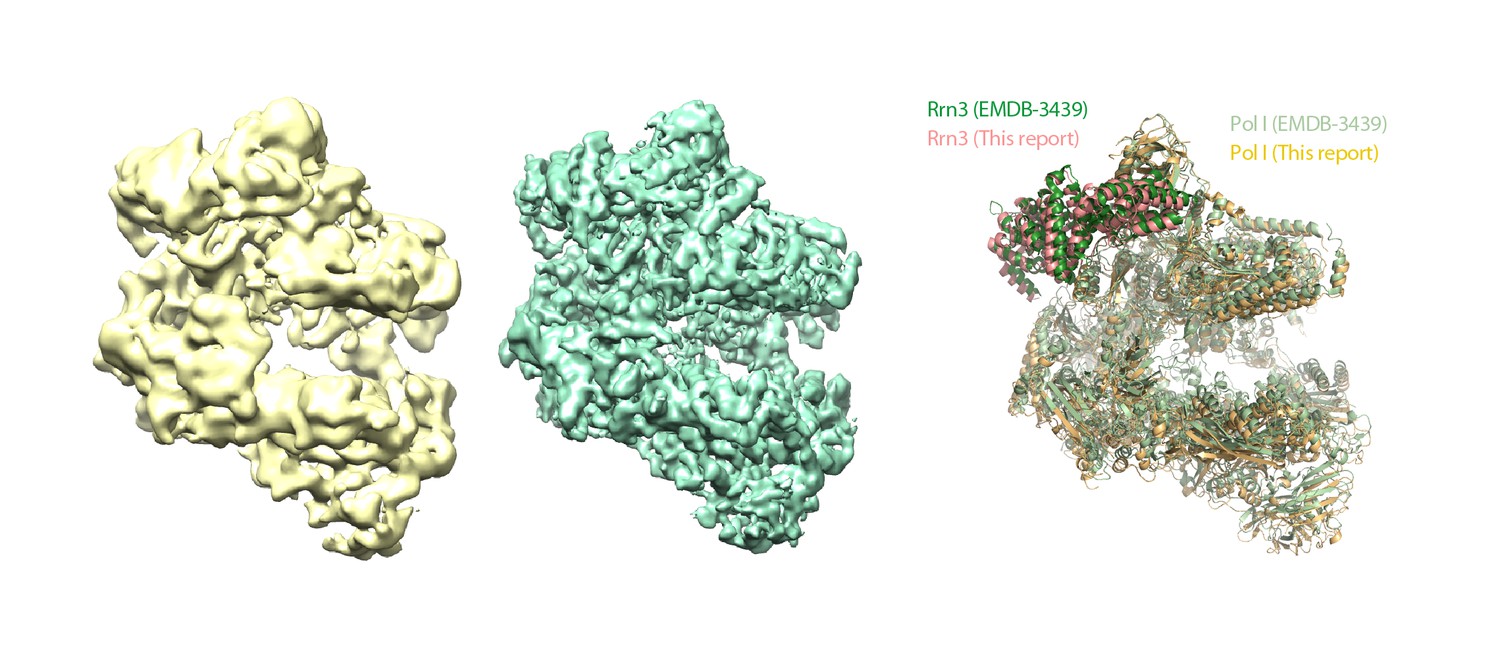
Structural comparison of Pol I–Rrn3 cryo-EM structures.
Comparison between the Pol I–Rrn3 structure reported here (yellow) and EMDB-3439 (green), with derived atomic models on the right side.
Figure 7 with 2 supplements
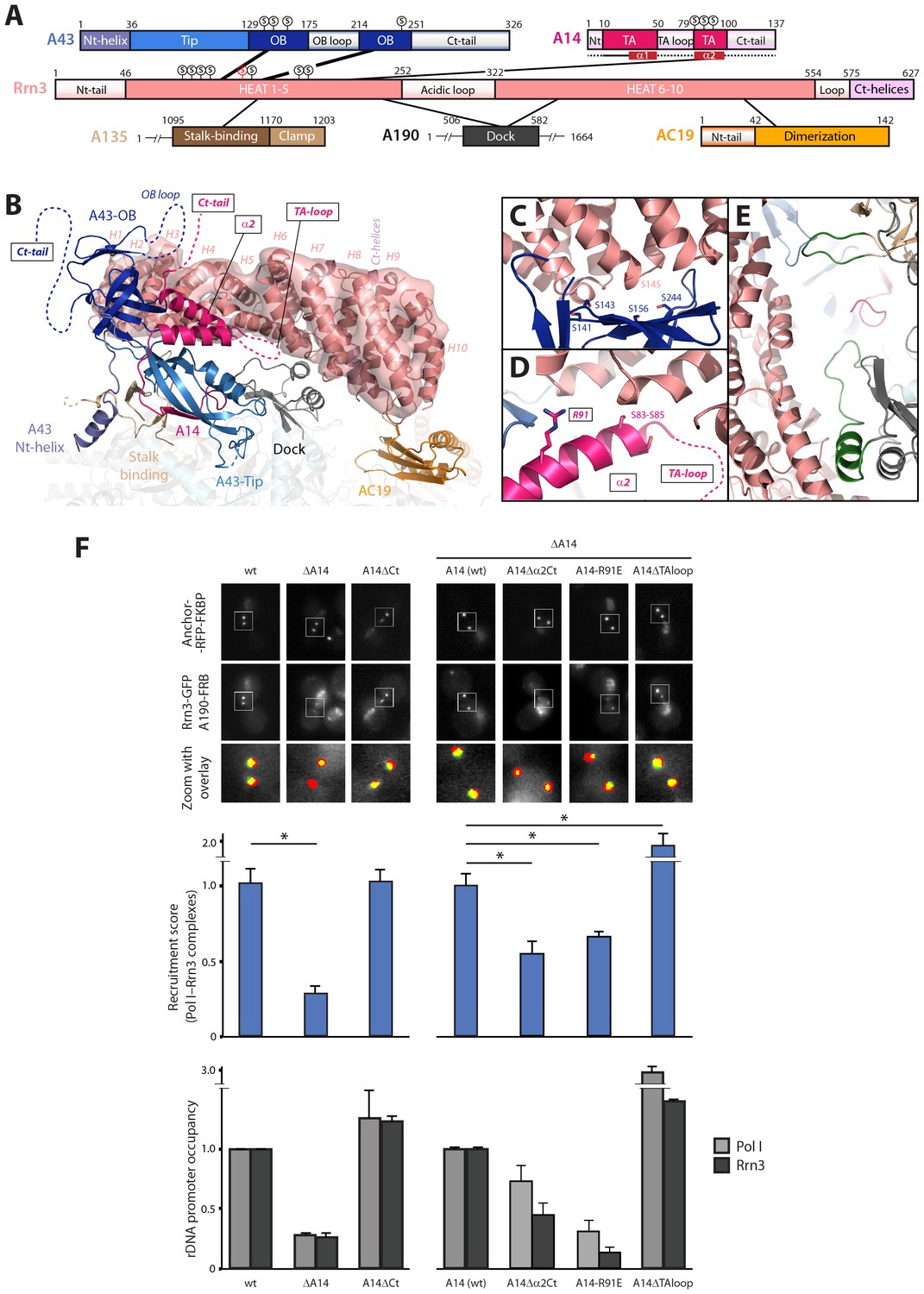
Mutational analysis of Pol I stalk subunits contacting Rrn3.
(A) Bar diagrams of the Pol I regions involved in Rrn3 binding, with connecting lines as derived from the cryo-EM structure. α-helices 1 and 2 in the A14 TA domain are marked in dark red. Serine residues in the A43 cradle, A14 stretch and Rrn3 patch are shown above the corresponding bars, with Rrn3-S145 in red. (B) Close-up view of the Pol I–Rrn3 interaction in a similar orientation to that in Figure 6B. HEAT repeats in Rrn3 labelled H1 to H10. Dotted lines represent disordered regions in the Pol I and Rrn3 crystal structures. Boxed-text marks truncated regions in the yeast mutants of panel F. (C) Close-up view of the serine cradle in A43 that accommodates serine 145 in Rrn3. (D) Close-up view of A14 helix α2, which lies in the vicinity of Rrn3. (E) Pol I specific insertions in subunits A190 and A135 are shown in green. (F) Representative PICT images of the RFP-tagged anchor (upper row), GFP-tagged Rrn3 (middle row) and a zoom of a 2.6 × 2.6 µm square around anchoring platforms (bottom row) of different mutant strains. Below, quantification of the Rrn3-GFP recruitment score, normalized to the measurement of the wild-type strain (Mean ± SD, p-value * < 0.01 t-test). At the bottom, ChIP experiments showing the relative association of A190 (light) and Rrn3 (dark) to the rDNA promoter region. All ChIP experiments were normalized to the value of the wild-type strain in rich medium (Mean ± SD).
Figure 7—figure supplement 1
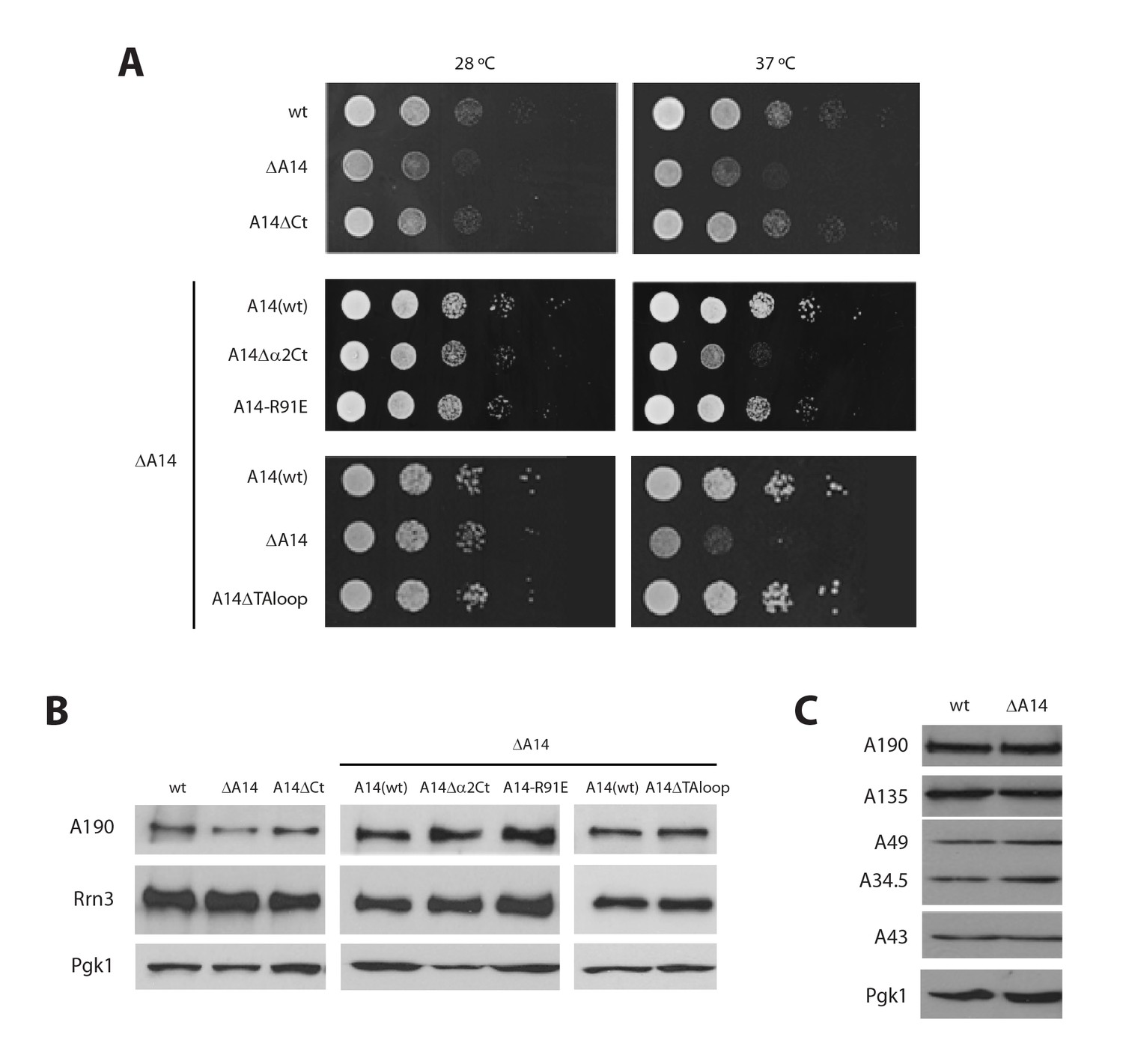
Additional characterization of Pol I stalk mutants.
(A) Comparative growth phenotype at 28°C and 37°C of the different A14 mutants. Serial dilutions (1:10) of wild-type and mutant strains were spotted on either YPD or selective SC medium and grown for 2–3 days at the indicated temperatures. (B) Western-blot analysis of A190 and Rrn3 of the whole cell extract from the different A14 mutants, using rabbit anti-A190 and mouse anti-GFP, respectively. Mouse anti-Pgk1 was used as a loading control. (C) Western-blot analysis of different Pol I subunits in the ΔA14 strain. All antibodies against Pol I subunits are from rabbit.
Figure 7—figure supplement 2
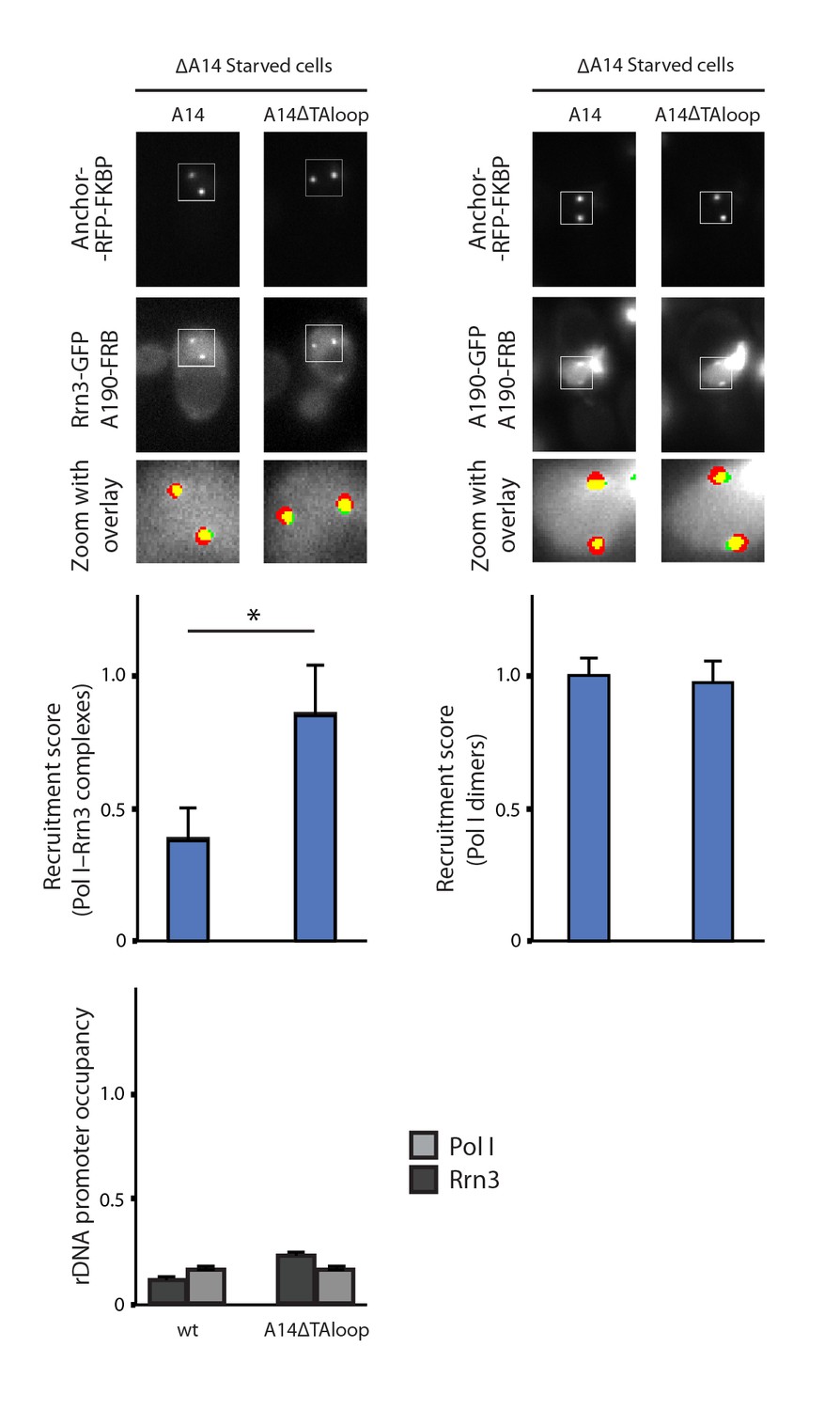
Characterization of A14ΔTAloop in starving conditions.
Representative PICT images of the RFP-tagged anchor (upper row), GFP-tagged Rrn3 (middle-left) or A190 (middle-right), and a zoom of a 2.6 × 2.6 µm square around the anchoring platforms (bottom row). Below, quantification of the Rrn3- or A190-GFP recruitment score, normalized to the measurement of the wild-type strain in rich (Rrn3-GFP) or starving (A190-GFP) medium (Mean ± SD, p-value * < 0.01 t-test). At the bottom, ChIP experiments showing the relative association of A190 (light) and Rrn3 (dark) to the rDNA promoter region. All ChIP experiments were normalized to the value of the wild-type strain in rich medium.
Figure 8 with 1 supplement
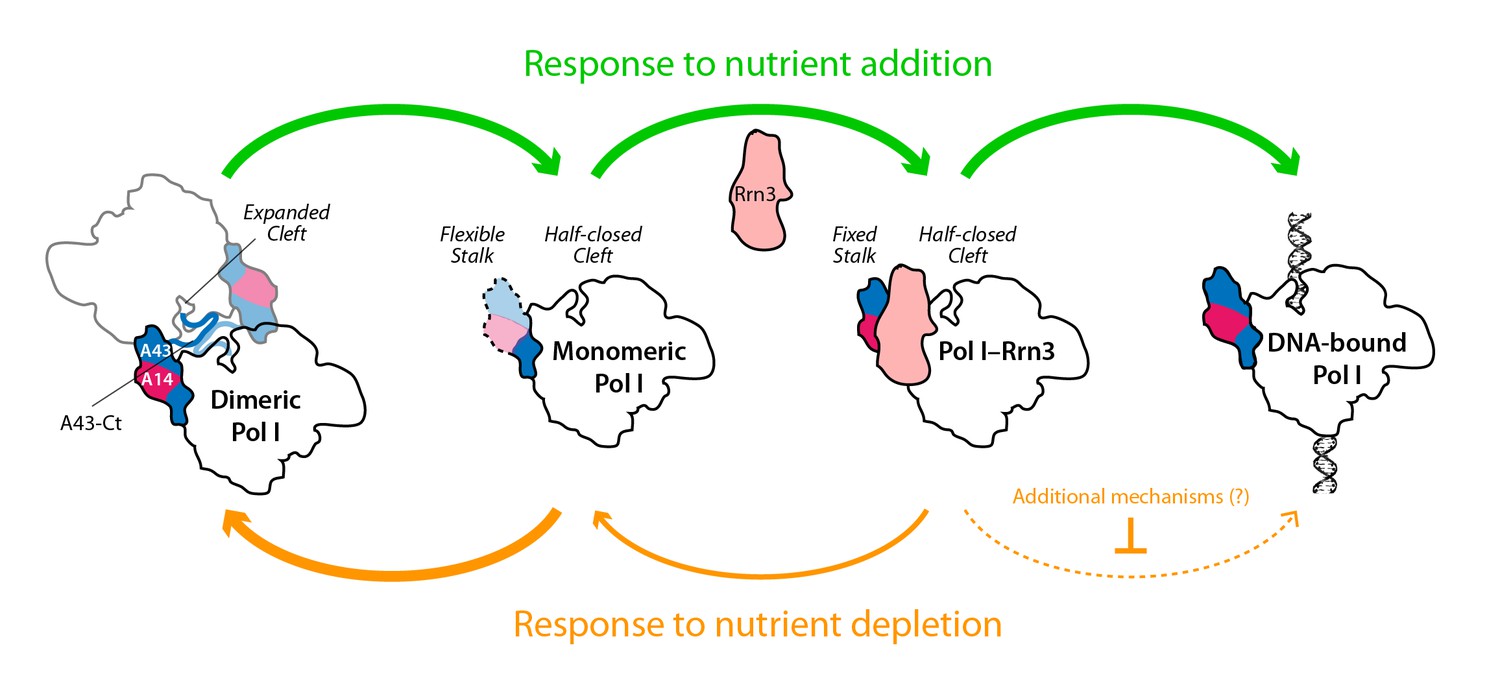
Model for the influence of nutrient availability on the assembly of Pol I complexes.
Nutrient addition to starved cells induces formation of Pol I–Rrn3 complexes to activate transcription, while Pol I homodimers disrupt to generate fresh monomeric Pol I (green arrows). Nutrient depletion causes partial disruption of Pol I–Rrn3 complexes and formation of Pol I homodimers (orange arrows). Both events downregulate rDNA transcription, while additional regulatory mechanisms may also participate.
Figure 8—figure supplement 1
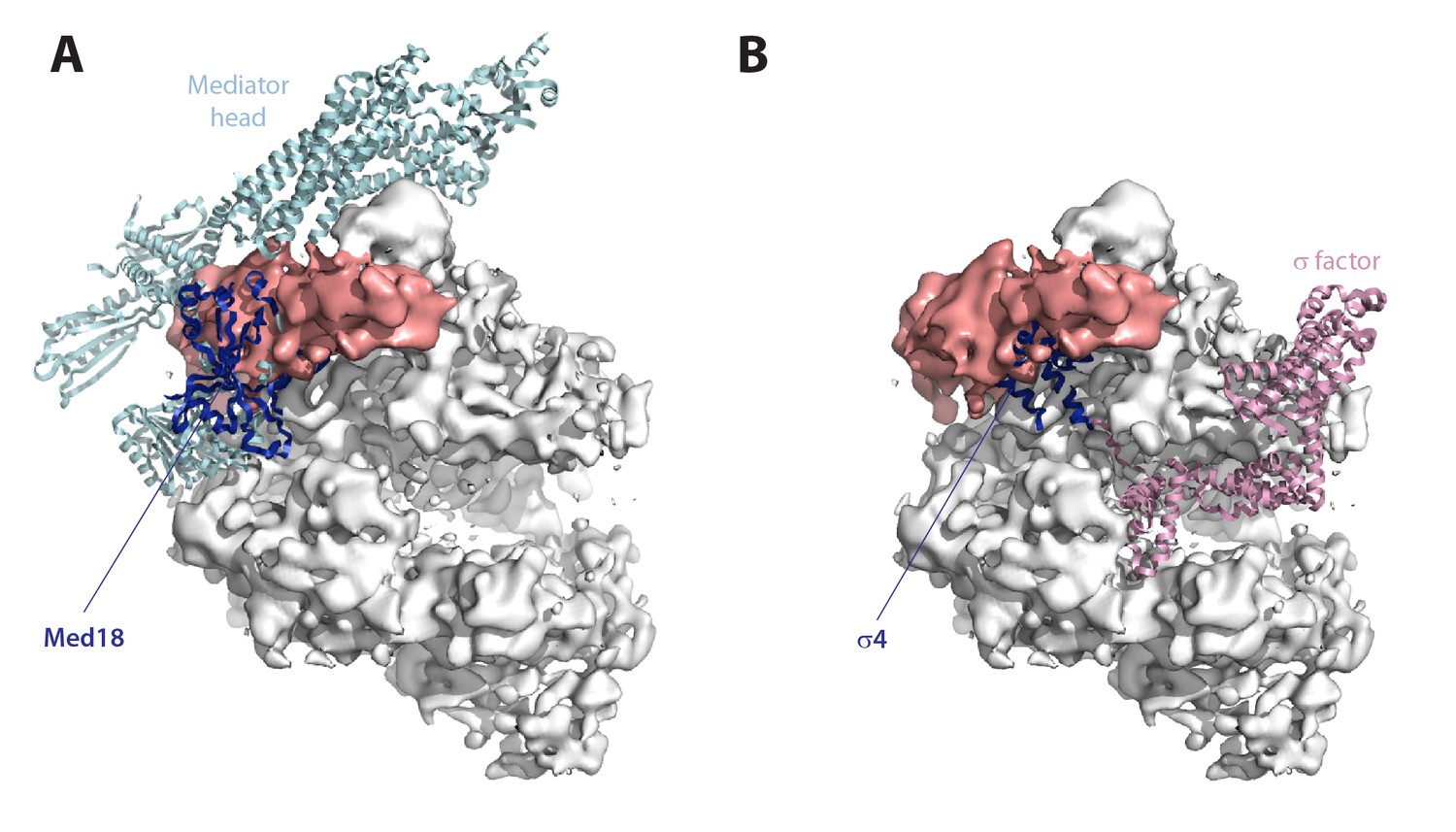
Comparison with other transcription systems.
(A) Cryo-EM reconstruction of the Pol I–Rrn3 complex with fitted Pol II–Mediator head complex (PDB 4V1O). The mediator head is in cyan, while its Med18 subunit is in blue. (B) Cryo-EM reconstruction of the Pol I–Rrn3 complex with fitted bacterial RNA polymerase holoenzyme (PDB 1IW7). σ70 is in pink, while its σ4 domain is in blue.
Videos
Video 1
Engineered anchoring platform associated with the spindle pole body.
Yeast cell expressing GFP and Tub4-RFP-FKBP imaged in z-stacks of 250 nm incremental steps. Imaris software was used to obtain the 3D reconstruction. A maximum of two anchoring platforms could be observed in each cell.
Video 2
Structural transition from Pol I homodimers to monomers.
The Pol I enzyme is presented in the same view as Figure 5B, with the clamp on the right and the protrusion on the left.
Additional files
-
Supplementary file 1
Table of yeast strains used in this study.
- https://doi.org/10.7554/eLife.20832.027
-
Supplementary file 2
Table of statistics for the cryo-EM structures described in this study.
- https://doi.org/10.7554/eLife.20832.028
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The dynamic assembly of distinct RNA polymerase I complexes modulates rDNA transcription
eLife 6:e20832.
https://doi.org/10.7554/eLife.20832




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}