CatSperζ regulates the structural continuity of sperm Ca2+ signaling domains and is required for normal fertility
- Howard Hughes Medical Institute, Boston Children's Hospital, United States
- Harvard Medical School, United States
- Yale School of Medicine, United States
- Harvard University, United States
- James J. Perters VA Bronx, Icahn School of Medicine at Mount Sinai, United States
Figures
Figure 1 with 2 supplements
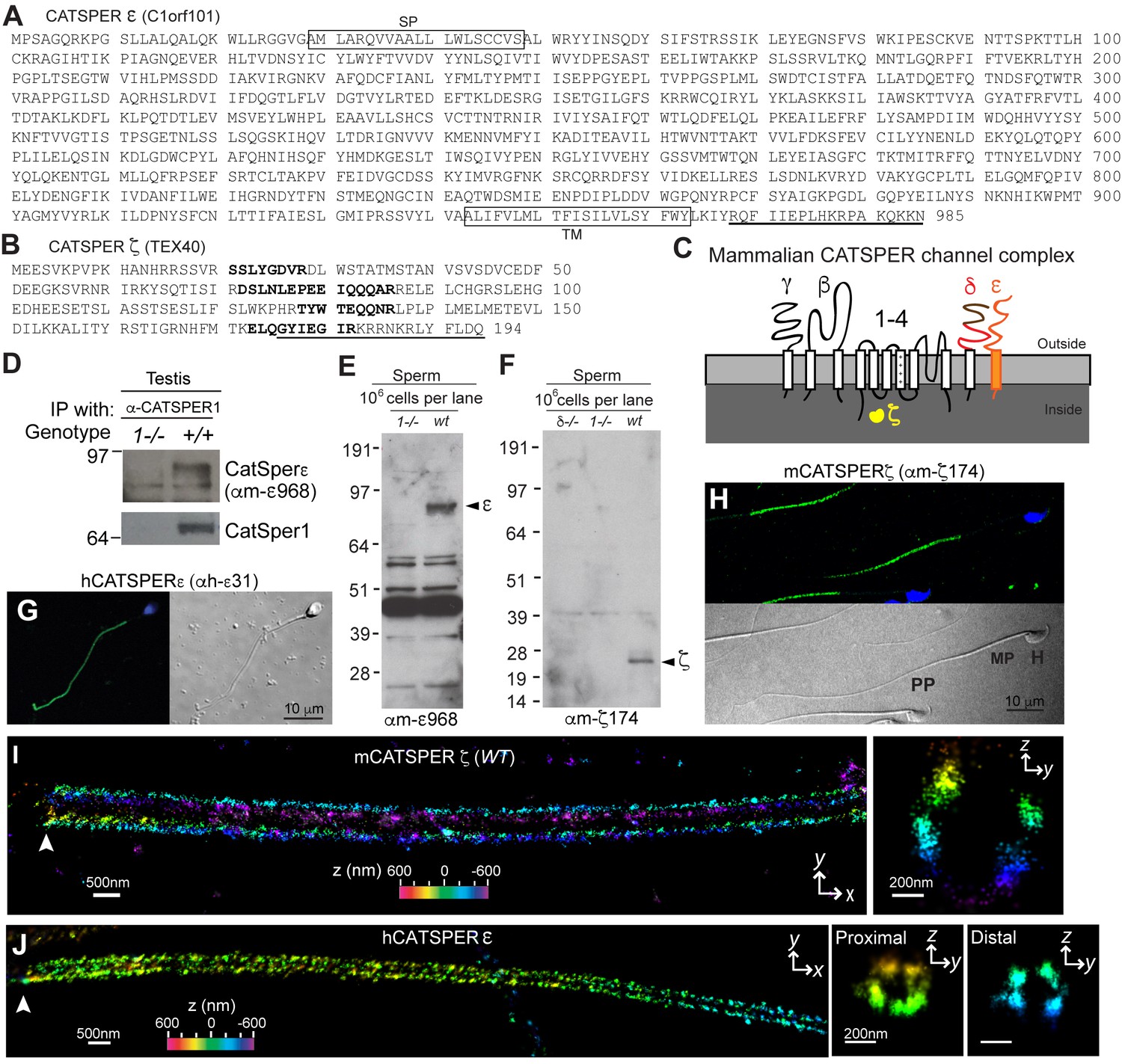
CatSper ε and ζ, two new accessory proteins of CatSper channel complex.
(A and B) Mouse protein sequences of CatSper ε (A) and ζ (B). (C) Cartoon of the predicted topology of 9 CatSper subunits. (D) Association of CatSperε with CatSper1 in testis. (E and F) Dependence of CatSper ε (E) and ζ (F) proteins on CatSper1 in mouse sperm cells. (G and H) Confocal fluorescence and the corresponding phase-contrast images of immunostained human CatSperε (G) and mouse CatSperζ (H). (I) 3D STORM images of mouse CatSperζ in capacitated wt sperm. x-y projection (left) and a y-z cross-section (right) at 0.5 um from the annulus. The color encodes the relative distance from the focal plane along the z axis (color scale bar in x-y projection). (J) 3D STORM images of human CatSperε in x-y projection (left), in y-z cross-sections (right). Colors indicate the z positions (see color scale bar). See also Figure 1—figure supplements 1–2.
Figure 1—figure supplement 1
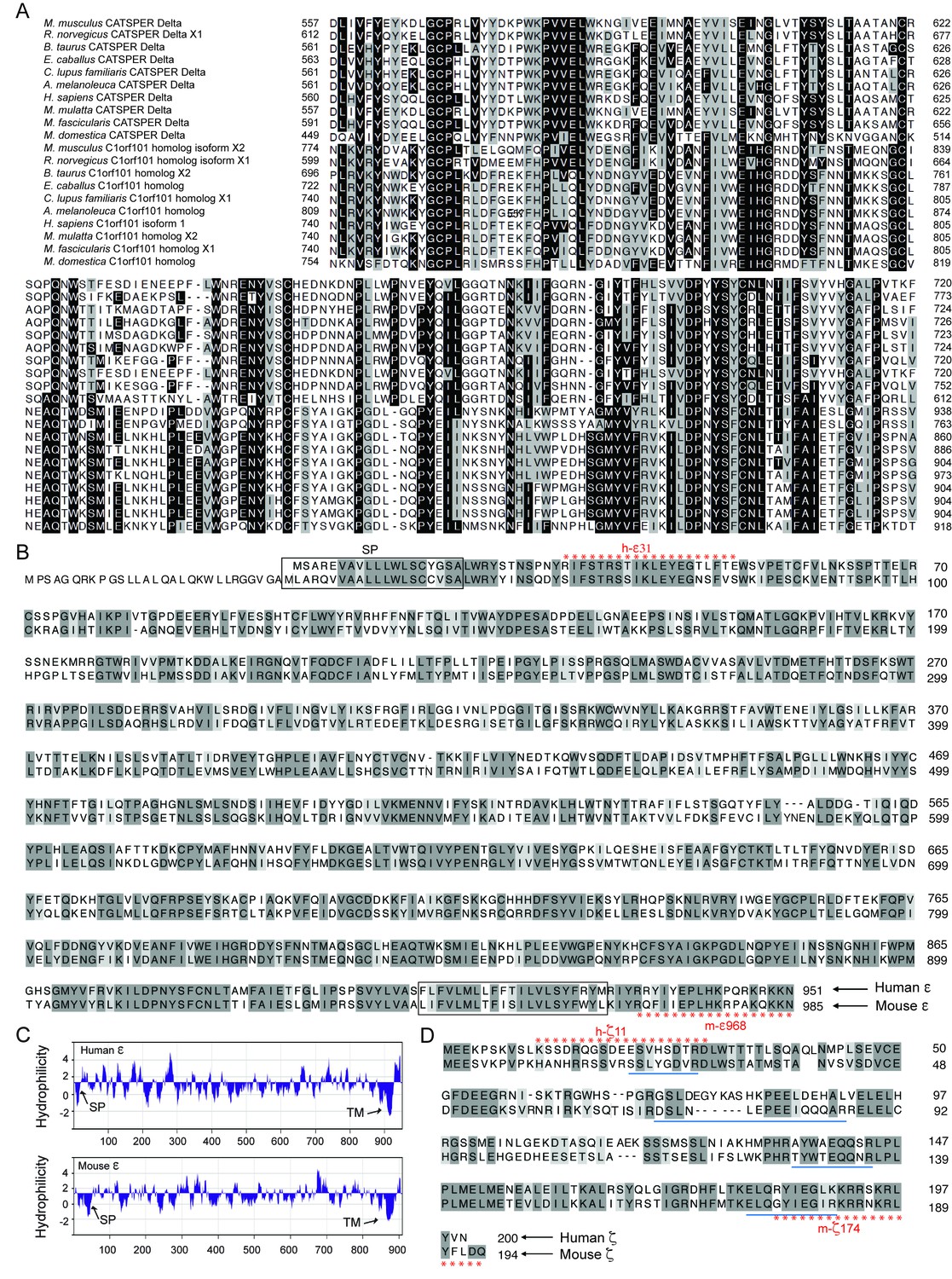
Identification of CatSper ε and ζ, two novel accessory proteins of the CatSper channel complex, related to Figure 1.
(A) Multiple sequence alignment of CatSperδ identifies C1orf101 as δ homologs from various species. Alignments originate ~165 amino acids before the predicted single transmembrane (TM) domain, showing the highly conserved region in the proteins’ C-terminal half. Identical (black) and similar (gray) residues highlighted. (B) CatSperε is a protein containing a putative single transmembrane (TM) domain localized to the sperm tail. Pairwise alignment of the predicted human (upper, C1orf101 isoform 1) and mouse (lower, C1orf101-like isoform X2) CatSper ε protein sequences. The predicted signal peptide (SP) (Frank and Sippl, 2008) and TM domain are boxed. (C) von Heijne hydrophilicity plot (window size = 11) of human and mouse CatSperε proteins. (D) Sequence alignment between the human (upper) and mouse (lower) CatSperζ proteins encoded by Tex40 genes. The four peptides from mouse CatSperζ (identified by mass spectrophotometry from CatSper1 affinity purification but not annotated in the previous study) (Chung et al., 2011), are underlined.
Figure 1—figure supplement 2
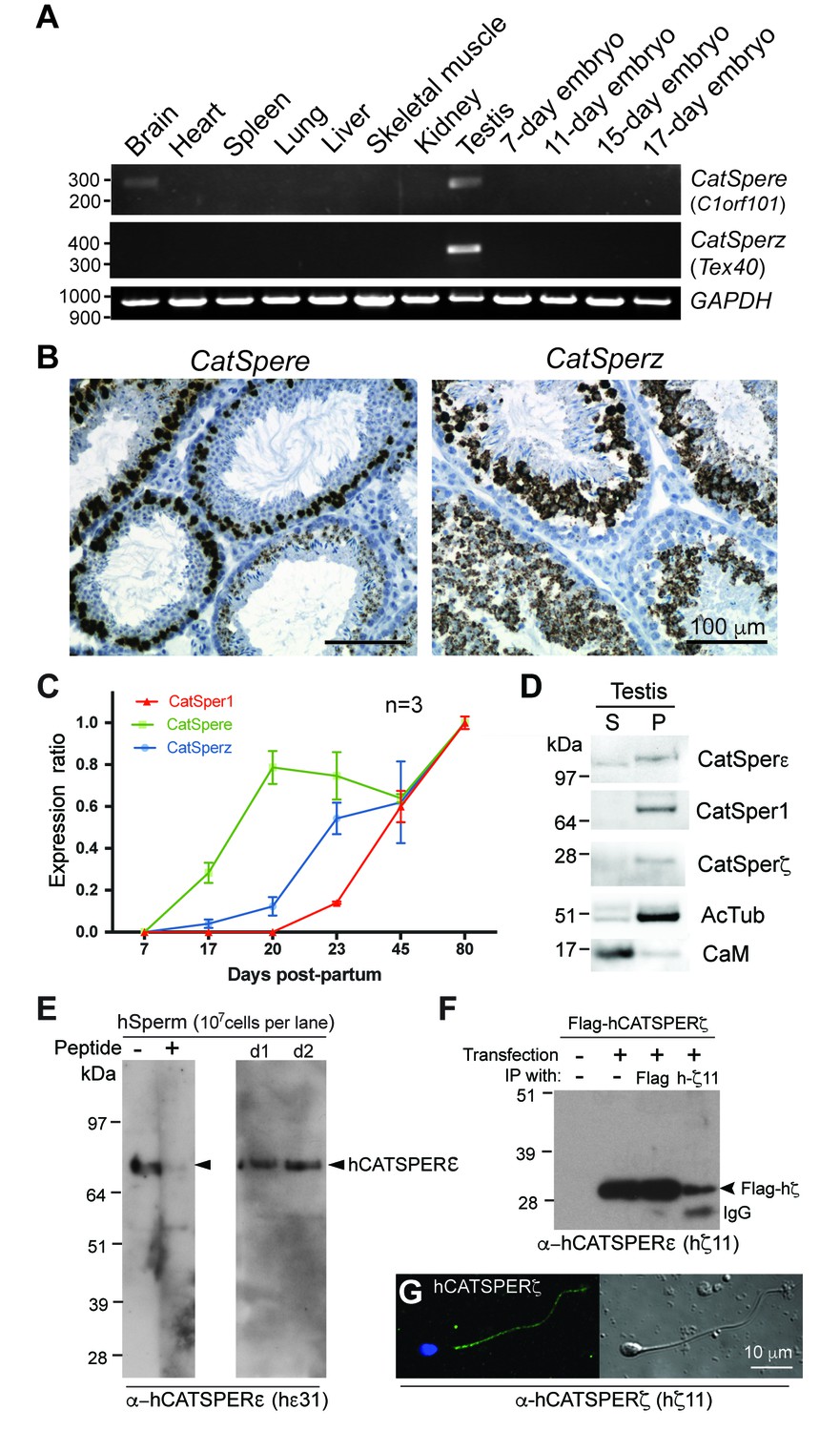
Expression of CatSper e and z mRNAs and proteins, related to Figure 1.
(A) Tissue expression profile of CatSper e and z. Reverse transcription PCR of CatSper e (upper), z (middle), and G3pdh (control; lower) from 12 mouse cDNAs. CatSper e and z are enriched in testis. (B) Spatial localization of CatSpere and z mRNA in the testis. Representative fields of in situ hybridization by gene-specific oligonucleotides against CatSper e (left) and z (right) in mouse testis (RNAscope). (C) Temporal expression of CatSper1, CatSpere, and CatSperz mRNAs during postnatal testis development. The mRNA levels of CatSper1 (orange), CatSpere (green), and CatSperz (blue) are measured by real-time RT PCR, normalized to HPRT and expressed as ratios relative to 80-day old adult mouse testis. The data are presented as mean ± SEM. N = 3. (D) Partitioning of CatSperε and CatSperζ in fractionated extracts of testis from wt mice. Both CatSperε and CatSperζ are enriched in the microsomal pellet (P), not in supernatant (S). (E–G) Specific recognition of CatSperε and CatSperζ in human spermatozoa. Immunoblotting of (E) total human sperm extracts and (F) recombinant human CatSperζ by rabbit polyclonal CatSperε (hε31) and CatSperζ (hζ11) antibodies, respectively. d1 and d2 indicate sperm from donors 1 and 2. (G) Confocal image and the corresponding phase-contrast image of CatSperζ in human sperm, immunostained with hζ11.
-
Figure 1—figure supplement 2—source data 1
Temporal expression of CatSper1, CatSpere, and CatSperz mRNAs during postnatal testis development.
- https://doi.org/10.7554/eLife.23082.006
Figure 2 with 2 supplements
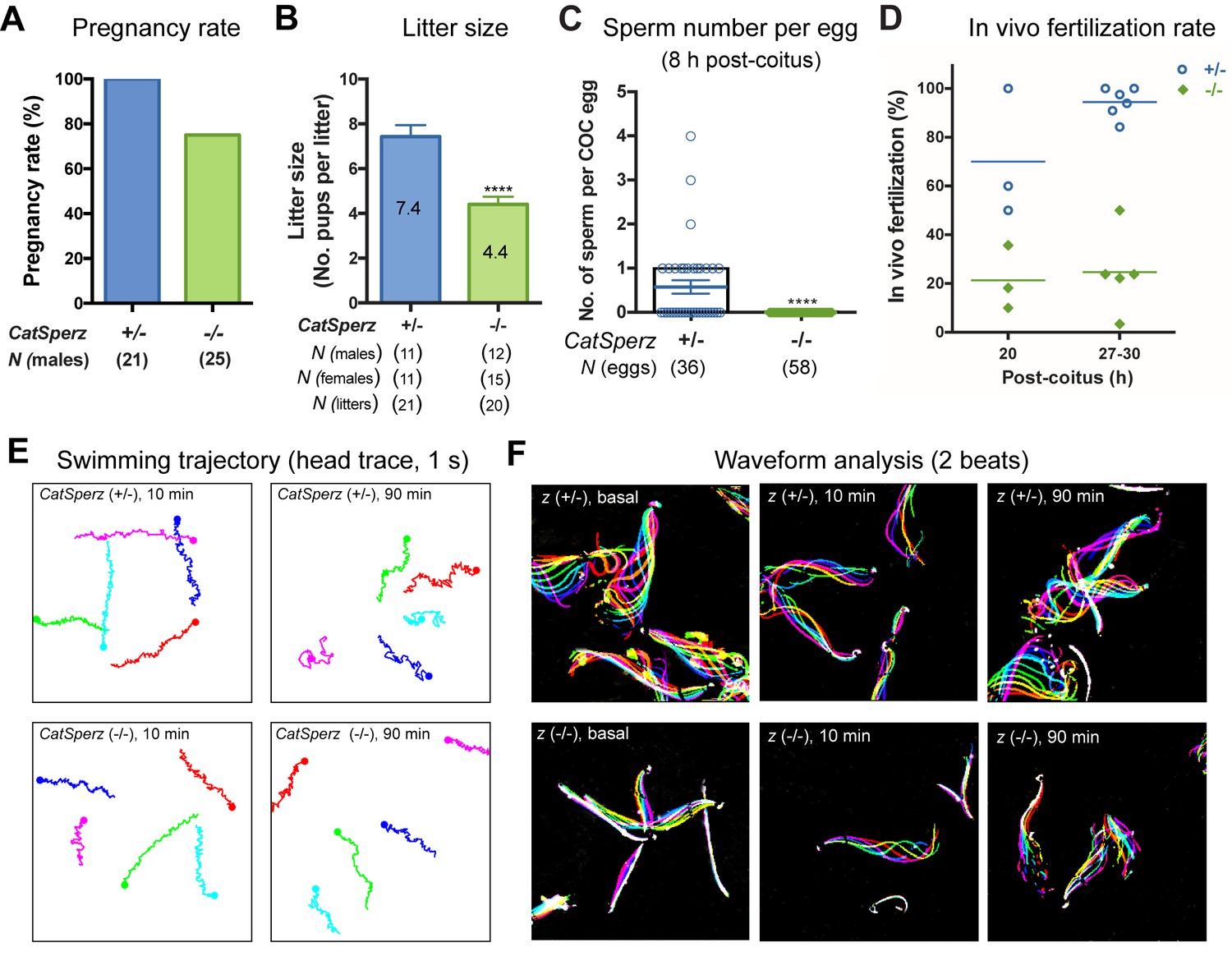
Deletion of the mouse CatSperζ subunit severely impairs male fertility.
(A) Percent pregnancy rate over three months. (B) Average litter size resulting from CatSperz+/- (7.4 ± 0.5) and CatSperz-/- (4.4 ± 0.3) males. (C) Sperm number per egg at the fertilization site 8 hr after 1 hr window-timed coitus with CatSperz+/- (0.58 ± 0.15) and CatSperz-/- (0, none) males, quantified from eggs collected from ampullae. (B) and (C) Data are mean ± SEM. ****p<0.0001. (D) In vivo fertilization rate: Scatter plot with mean % of 2 cell fertilized eggs from CatSperz+/- (70% and 94.4%) and CatSperz-/- (21.3% and 24.6%) mated females at 20 and 27–30 hr after coitus, respectively. (E) Head trace of free swimming CatSperz+/- (top) and CatSperz-/- (bottom) sperm cells at 10 min (left) and 90 min (right) after capacitation. Traces are from 1 s movies taken at 37°C. (F) Flagellar waveform traces. Movies recorded at 200 fps: CatSperz+/- (top) and CatSperz-/- (bottom) sperm cells attached on glass coverslips before capacitation (left), and 10 min (middle), and 90 min (right) after capacitation. Overlays of flagellar traces from two beat cycles are generated by hyperstacking binary images; time coded in color. See also Figure 2—figure supplements 1–2 and Figure 6—figure supplement 1.
-
Figure 2—source data 1
Impaired male fertility in CatSperz-/- mice: pregnancy rate, litter size, sperm number per egg, and in vivo fertilization rate.
- https://doi.org/10.7554/eLife.23082.008
Figure 2—figure supplement 1
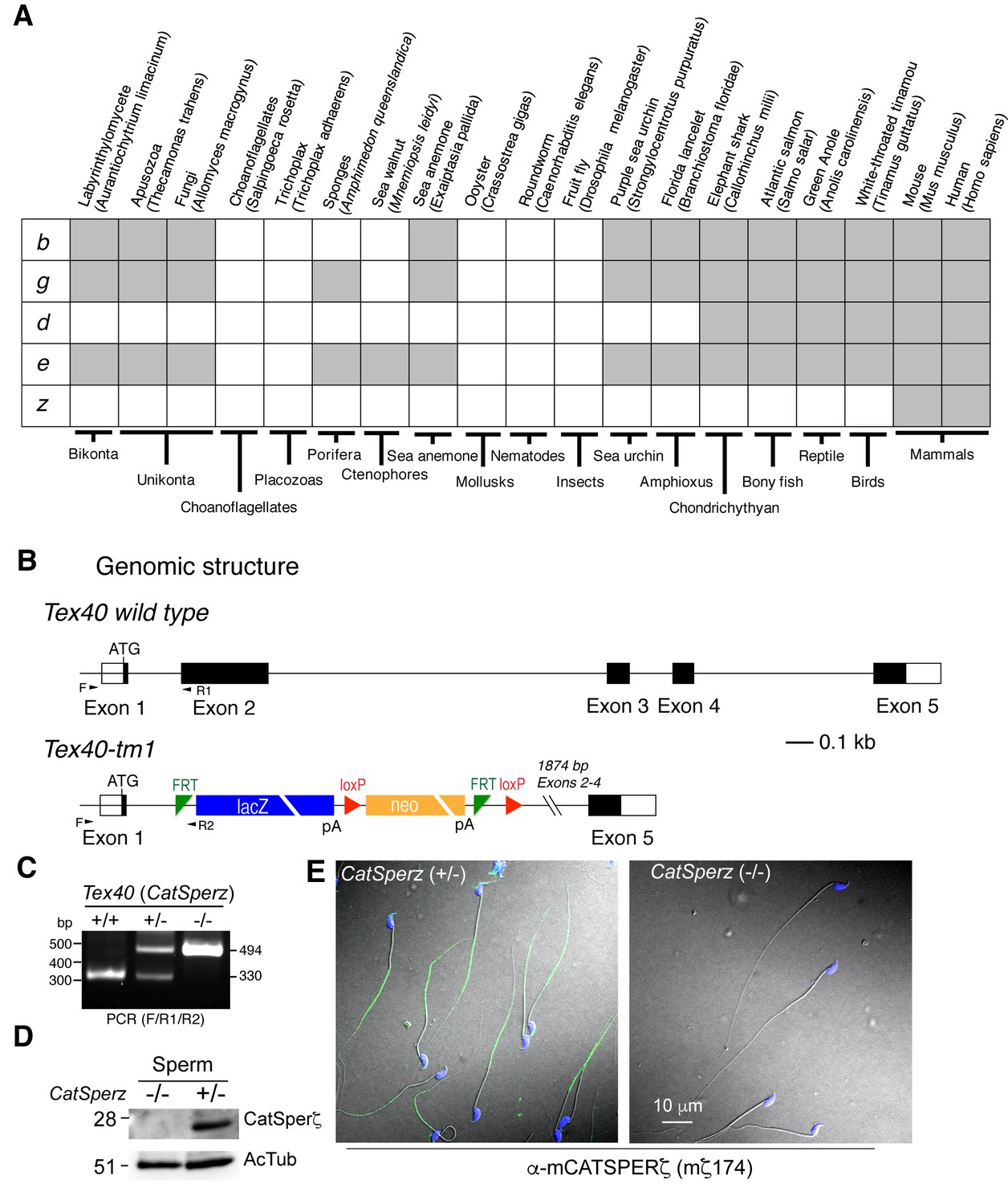
Generation of CatSperz-/- mice, related to Figure 2.
(A) Distribution of CatSper subunits in eukaryotes. (B) ES cells (Project ID: CSD33943) from the KOMP Repository were used to produce KO mice. Exons 2–4 deleted by gene trap. (C and D) Genotyping (primers F/R1/R2) (C) and immunoblotting (D) analysis of CatSperz-/-. (E) Normal sperm morphology despite the absence of ζ protein in CatSperz-/- spermatozoa. Overlay of confocal images and the corresponding phase-contrast images of mouse sperm cells from CatSperz+/- and CatSperz-/- mice immunostained with mζ174 (also used in (D) and Figure 1F and H. The principal piece labeling is not observed in CatSperz-null sperm, validating the specific subcellular distribution of the signal to the sperm tail. Hoechst dye stains the sperm head DNA (blue).
Figure 2—figure supplement 2
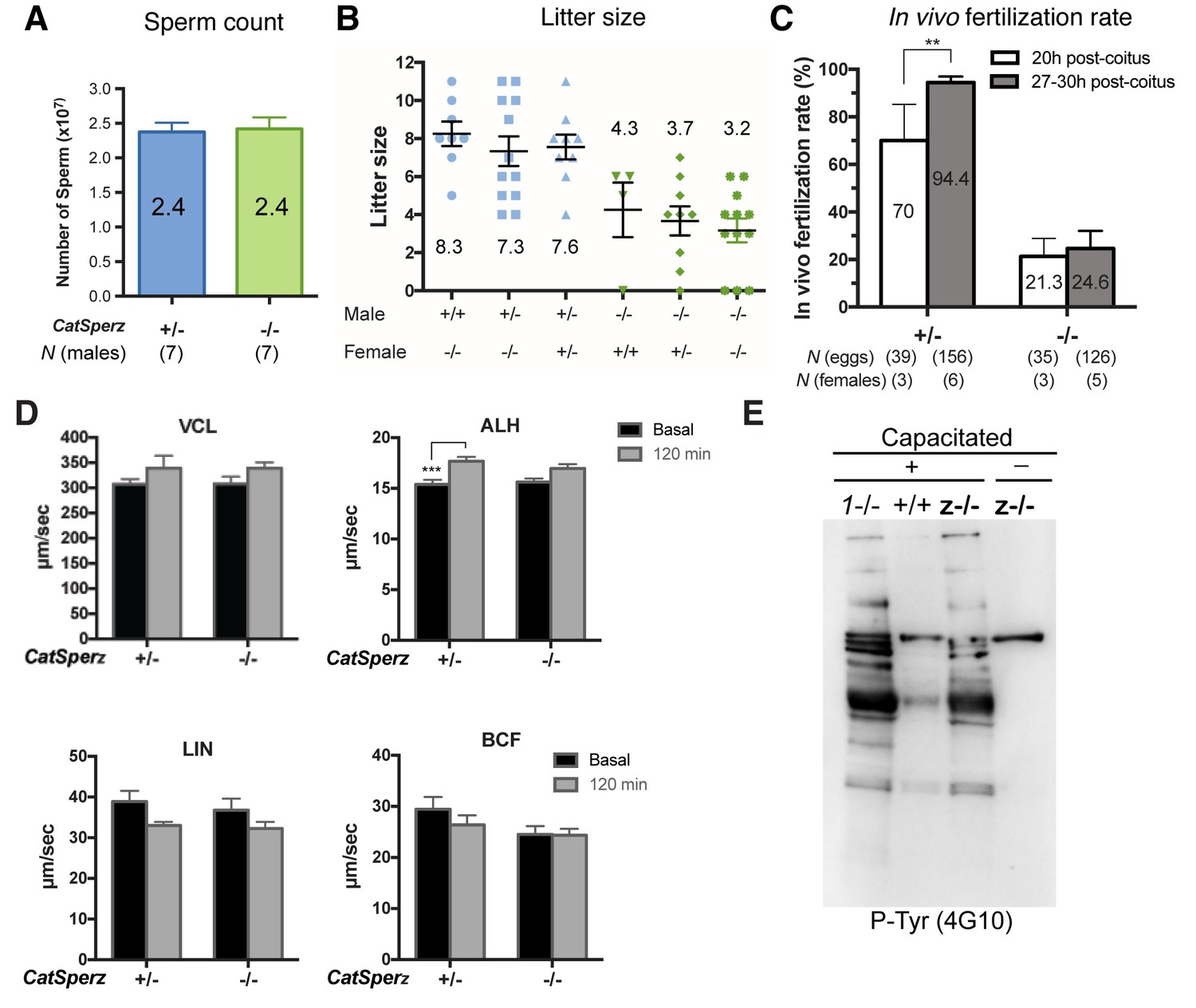
Sperm count and fertility of CatSperz-/- mice; sperm motility analysis and development of P-Tyr, related to Figures 2 and 3.
(A) Epididymal sperm count (mean ± SEM) from littermates at ages 2–3 months. CatSperz het (+/-, blue; 2.4 ± 0.1) versus null (-/-, green; 2.4 ± 0.2) cells (107). (B) Average litter size from all males in the mating test, grouped by male and female genotype. (C) IVF rate calculated by counting fertilized eggs (2 cell stage) 20 and 27–30 hr after coitus. Data are expressed as a scatter plot of mean percentage from CatSperz+/- (20 hr, 70 ± 15; 27–30 hr, 94.4 ± 2.5) and CatSperz-/- (20 hr, 21.3 ± 7.6; 27–30 hr, 24.6 ± 7.4). **p=0.0097 (One-way ANOVA and F test). See also Figure 2D. (D) Sperm motility parameters measured by computer assisted sperm analysis (CASA) from CatSperz het (+/-) versus null (-/-) male mice. 5 min (basal, gray) and 90 min (capacitated, black) after incubation in HTF. ALH of CatSperz+/- (basal, 15.5 ± 0.4; 120 min, 17.3 ± 0.4, p=0.0002). Data are mean ± SEM. N = 4. (E) Capacitation-associated protein tyrosine phosphorylation of CatSperz -/- spermatozoa.
-
Figure 2—figure supplement 2—source data 1
Impaired male fertility in CatSperz-/- mice: sperm count, litter size per genotype, in vivo fertilization rate, and CASA parameters.
- https://doi.org/10.7554/eLife.23082.011
Figure 3

ICatSper, but not ATP-activated P2X2 current, is reduced in CatSperz-null spermatozoa.
(A) CatSperz-/- and (B) CatSperz+/- ICatSper. Left panels show the current-voltage relations of monovalent ICatSper in response to voltage ramps at the time points indicated. Right traces are representative time courses of ICatSper measured in the standard bath solution (1, HS), ATP-activated P2X2 current (2, ATP), and nominally divalent-free solution (3, DVF) at −100 mV (gray circles) and +100 mV (black circles). ICatSper in CatSperz-null sperm cells is ~60% of that recorded from wt. Inward IATP current induced by 100 µM ATP is similar in both phenotypes and indistinguishable from previously published wt IATP (Navarro et al., 2011). (C) Average ICatSper measured from CatSperz+/- (−683 ± 77 pA) and CatSperz-/- (−426 ± 50 pA) sperm cells at −100 mV. Data are mean ± SEM. p=0.0297. Cartoon shows the standard pipette solution (mM); internal Cs used to block K+ currents.
-
Figure 3—source data 1
Inward CatSper current at −100 mV.
- https://doi.org/10.7554/eLife.23082.013
Figure 4
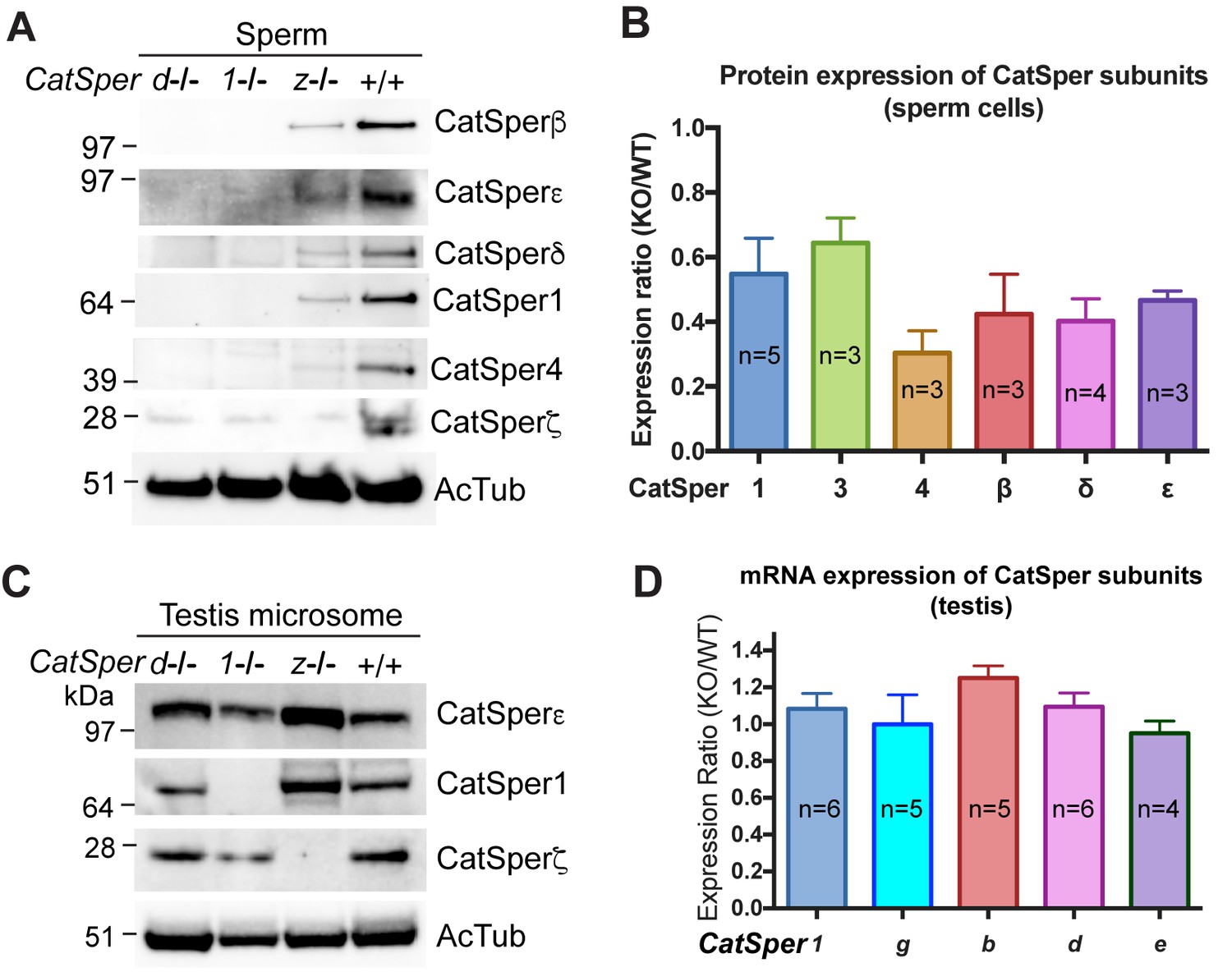
CatSper proteins are reduced in sperm from CatSperz-null mice despite protein expression during spermatogenesis.
(A and B) Reduced expression of CatSper subunits in sperm cells of CatSperz homozygous null mice compared with their complete absence in CatSper1 and d-null mice. Immunoblotting of (A) total mouse sperm extracts and (B) protein expression ratio (z-KO/wt) of CatSper 1 (0.5 ± 0.1), 3 (0.6 ± 0.08), 4 (0.3 ± 0.07), β (0.4 ± 0.1), δ (0.4 ± 0.07), and ε (0.5 ± 0.03). Data are mean ± SEM. (C) Increased expression of CatSper1 and ε in mouse testis in CatSperz-null mutants. (D) Quantitative gene expression analysis (qRT-PCR) from adult CatSperz-het and null testes: expression ratio (2-ddCT) and mean ddCt (null-het); TATA binding protein (TBP) is the internal control. The expression ratio of CatSper1 (1.1 ± 0.1) and all accessory g (1.0 ± 0.2), b (1.3 ± 0.07), d (1.1 ± 0.08), and e (0.95 ± 0.07) subunits are mean ± SEM.
-
Figure 4—source data 1
Protein and mRNA expression of CatSper subunits.
- https://doi.org/10.7554/eLife.23082.018
Figure 5 with 1 supplement
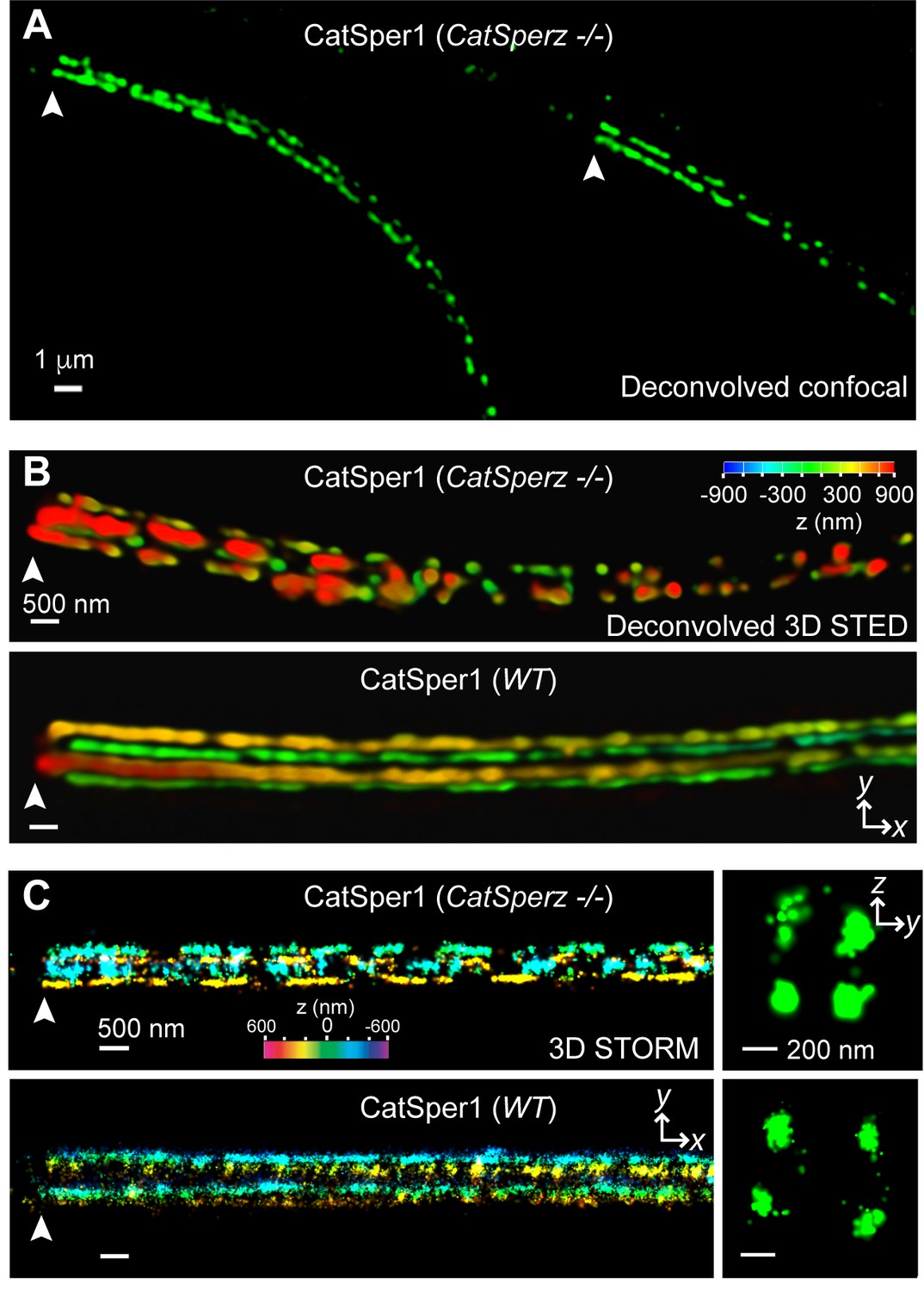
CatSperz deletion disrupts the continuity of the CatSper linear domains.
Application of different modes of fluorescence microscopy to observe CatSper localization. (A) Deconvolved confocal image of α-CatSper1 immunostained CatSperz -null spermatozoa. Scale bar, 1 μm. (B and C) 3D super-resolution images of CatSper1. 3D STED (B) and 3D STORM (C) images of CatSperz -null (top) and wt (bottom) sperm flagella, respectively. x-y projection colors encode the relative distance from the focal plane along the z axis. Scale bar, 500 nm. Arrowheads indicate the junction between the mid-piece and the principal piece (annulus) of the tail. 3D STORM, y-z cross-section images are shown on the right. Scale bar, 200 nm. See also Figure 5—figure supplement 1.
Figure 5—figure supplement 1
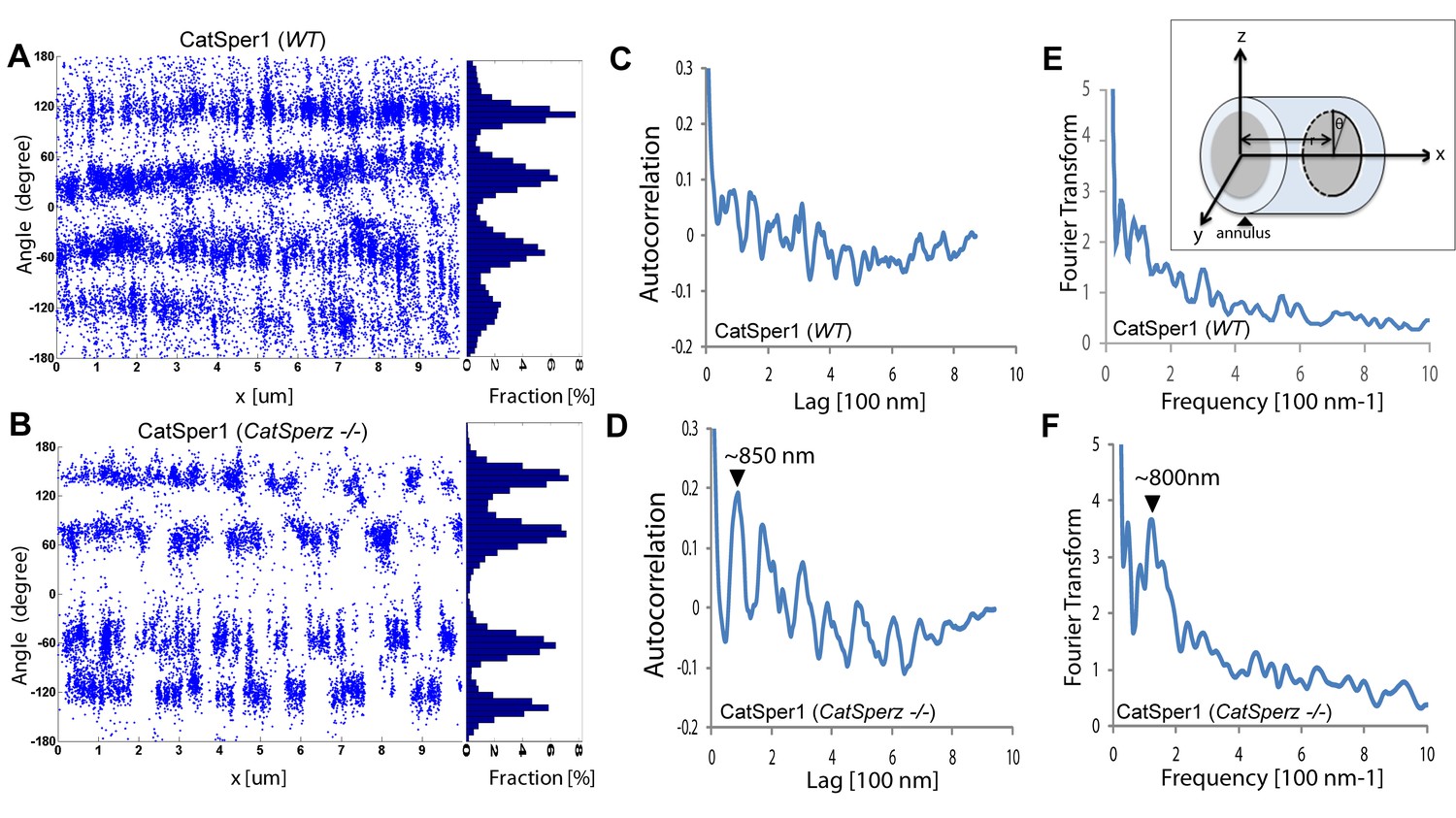
Subcellular distribution of immunolocalized CatSper proteins; related to Figure 5.
(A and B) Angular distributions (left) and profiles (right) of the surface-localized molecules of CatSper1 in wt (A) and CatSperz -/- spermatozoa (B) of Figure 5. (C and D) Averaged autocorrelation functions along the longitudinal axis (x-axis shown in E, inset) calculated from multiple CatSper domains in wt (C) and CatSperz -/- (D) spermatozoa. (n = 8). The longitudinal axis (x) is placed at the flagellar center and the origin at the annulus. (E and F) Fourier transformation of the 1D localization distribution shown in (A) and (B), showing a fundamental frequency of (800 nm)−1 in CatSperz-/- spermatozoa. (E, inset) Cartoon of cylindrical coordinate system for defining the radius and angles of molecular coordinates in STORM images.
Figure 6 with 1 supplement

CatSperz-null sperm rheotax poorly due to low torque.
(A) In-capillary sperm rheotaxis. Rheotactic ability is reduced in sperm lacking CatSperz at all flow rates tested (12–65 µm/s). (B) Rheotactic sperm cells are expressed as the % of total motile spermatozoa counted from 9 s-movies (CatSperz+/-, n = 32; CatSperz-/-, n = 24). Data are expressed in scatter plots; mean ± SEM (colored bars) of CatSperz+/- (88 ± 2) and CatSperz-/- (24 ± 4) as well as median with interquartile ranges (black boxes) of CatSperz+/- (96, IQR 78–100) and CatSperz-/- (22.5, IQR 0–40). ****p<0.0001. (C) Trajectory of free-swimming sperm in 0.3% methyl cellulose. Movies were taken at 50 fps to compare CatSperz+/- (left) and CatSperz-/- (right) sperm cells; bottom of glass dish, 37°C, 5 min after incubation in capacitation medium (HTF). Overlays of flagellar traces (20 frames, 2 s movie) are generated by hyperstacking binary images with gray intensity scale; end frame in black. Arrows indicate sperm heads in each trace. (D) Sperm rotation rate from CatSperz+/- (5 min, 2.4 ± 0.2; 90 min, 3.1 ± 0.2, p=0.0064) and CatSperz-/- (5 min, 3.8 ± 0.2; 90 min, 3.4 ± 0.3) males after incubation in HTF. The sperm rotation rate is calculated as previously reported (Miki and Clapham, 2013). Data are mean ± SEM. ****p<0.0001. (E and F) IVF with epididymal and/or ejaculated CatSperz+/- and CatSperz-/- spermatozoa. 2 cell stage eggs were counted 24 hr after insemination. (E) IVF rate with cumulus-intact oocytes from CatSperz+/- (epididymal, 71 ± 6) and CatSperz-/- (epididymal, 0.7 ± 0.7; ejaculate, 20 ± 6, p=0.0051). (F) IVF rate of cumulus-free/ZP-intact eggs with (CatSperz+/-, 80 ± 8; CatSperz-/-, 7 ± 7, p=0.0005; wt, 78 ± 6) or without (CatSperz+/-, 88 ± 6; CatSperz-/-, 0.8 ± 0.8, p=0.0002) glutathione-containing (GSH; 2 mM) media. Data are mean ± SEM. See also Figure 6—figure supplement 1.
-
Figure 6—source data 1
In-capillary sperm rheotaxis and in vitro fertilization.
- https://doi.org/10.7554/eLife.23082.023
Figure 6—figure supplement 1
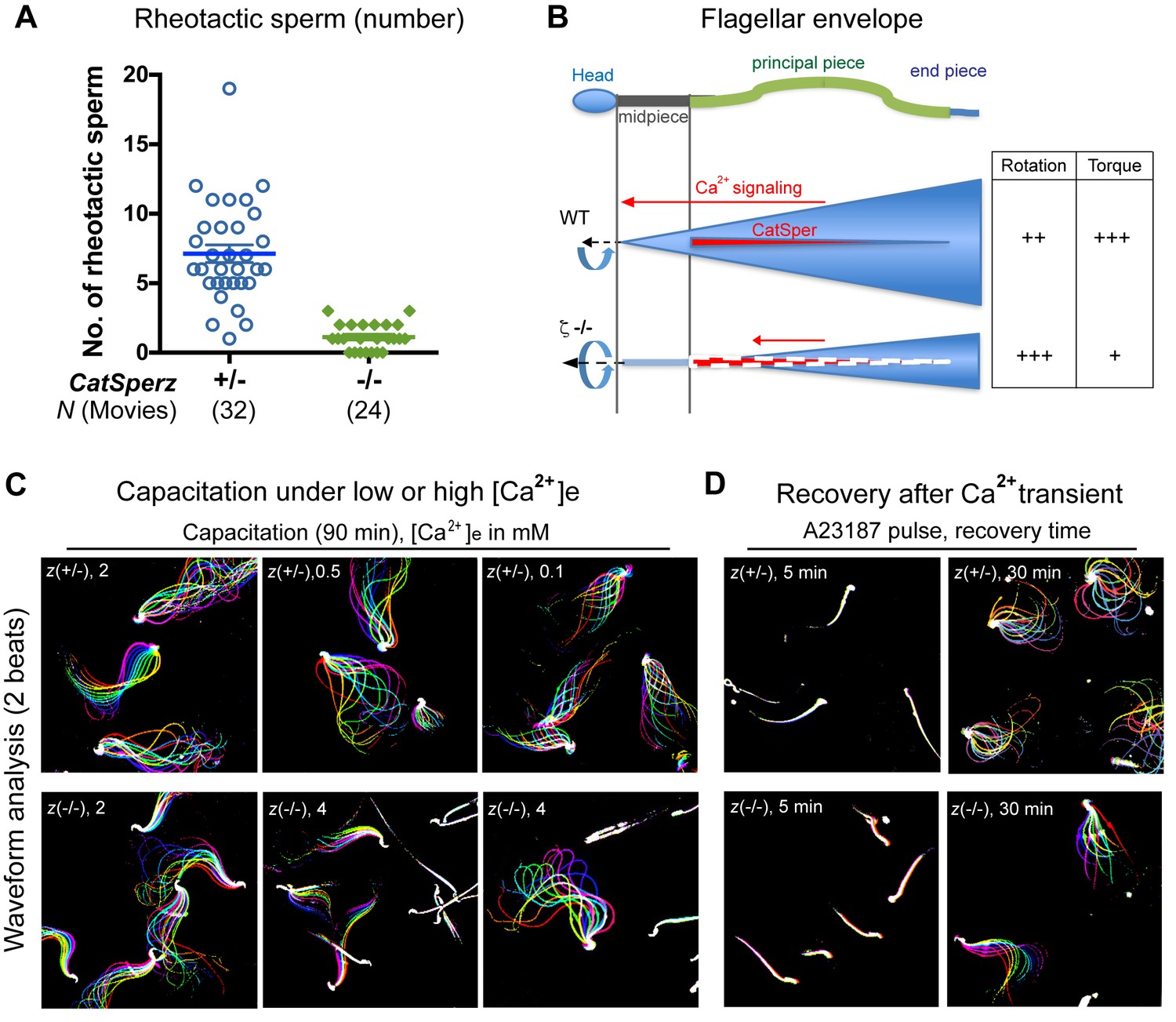
CatSper-mediated Ca2+signaling and development of the flagellar envelope; related to Figure 6.
(A) In-capillary sperm rheotaxis. Number of rheotactic sperm cells from each 9 s movie at all flow rates in the range of 12–65 µm/s; from Figure 6A. Data are expressed as a scatter plot with mean ± SEM (colored bars) from CatSperz+/- (7.1 ± 0.6) and CatSperz-/- (1.1 ± 0.2). (B) Working model illustrating the integrity of CatSper Ca2+ signaling domains and their relation to flagellar envelopes during sperm rotation. In wt spermatozoa, the CatSper channel forms four linear continuous Ca2+ signaling domains confined to the principal piece of the flagella. Ca2+ entry through the CatSper channels potentiates sperm rotation during capacitation (Miki and Clapham, 2013). With the resulting increased asymmetry and change in wave amplitude, the flagellar envelope is mapped out as a 3-dimensional cone in space, orienting sperm into the flow. Deficits in ICatSper and loss of the continuity of the linear domains in CatSperz-/- null spermatozoa compromise Ca2+ signaling and result in rigidity in the proximal region. The inflexibility of CatSperz-null spermatozoa from midpiece to halfway through the principal piece constrains the flagellar envelope to a narrower, rod like spatial map. The still active distal tail rotation then drives the more static rod-like structure. This causes the sperm to rotate faster but with less torque, thereby inefficiently orienting them into the flow and yielding less force in orthogonal directions needed to push aside the cumulus cells. (C and D) Flagellar waveform traces of spermatozoa. Movies recorded at 200 fps: CatSperz+/- (top) and CatSperz-/- (bottom) sperm cells tethered on glass coverslips. Overlays of flagellar traces from two beat cycles are generated by hyperstacking binary images; time coded in color. (C) Spermatozoa capacitated under low (0.5, 0.1 mM) or high (4 mM) extracellular calcium. (D) Sperm motility after Ca2+ transients by A23187 treatment followed by incubation under capacitation conditions.
Videos
Video 1
Movement of free swimming CatSperz +/- and -/- spermatozoa; related to Figure 2.
Uncapacitated (left) and 90 min capacitated (right) spermatozoa were allowed to disperse for 10 min pre-incubation in a 37°C chamber containing HEPES-HTF; free swimming sperm cells recorded within the next 5 min; video rate 20 fps (1/5 speed), 1 s movies; head trace to track swimming trajectory. (A) CatSperz+/- and (B) CatSperz-/- spermatozoa.
Video 2
Motility of tethered CatSperz +/- and -/- spermatozoa; uncapacitated, related to Figure 2.
Uncapacitated epididymal spermatozoa in non-capacitating M2 media were tethered to the fibronectin-coated glass bottom dish; sperm motility was recorded at 37°C; video rate 200 fps, 2 s movies. (A) CatSperz+/- and (B) CatSperz-/- spermatozoa.
Video 3
Motility of tethered CatSperz +/- and -/- spermatozoa; 90 min capacitated, related to Figure 2.
After 90 min incubation in HTF, capacitated epididymal spermatozoa were tethered to a fibronectin-coated glass bottom dish; sperm motility was recorded at 37°C; video rate 100 fps (1/2 speed), 1 s movies. (A) CatSperz+/- and (B) CatSperz-/- spermatozoa.
Video 4
In-capillary rheotaxis of CatSperz +/- and -/- spermatozoa; capacitated, related to Figure 6.
Capacitated epididymal spermatozoa in HTF for 90 min were loaded into the capillary and transferred to a 37°C chamber; sperm cells swimming against the flow and down were recorded; video rate 33 fps, 9 s movies. (A) CatSperz+/- and (B) CatSperz-/- spermatozoa.
Video 5
Movement of CatSperz +/- and -/- sperm in viscous medium; uncapacitated, related to Figure 6.
Uncapacitated spermatozoa were allowed to disperse for 10 min pre-incubation in a 37°C chamber containing HEPES-HTF supplemented with 0.3% methylcellulose; swimming sperm cells were recorded within the next 5 min; video rate 50 fps, 2 s movies. (A) CatSperz+/- and (B) CatSperz-/- spermatozoa.
Video 6
Movement of CatSperz +/- and -/- sperm in viscous medium; 90 min capacitated, related to Figure 6.
Spermatozoa capacitated in HTF were allowed to disperse for 10 min pre-incubation in a 37°C chamber containing HEPES-HTF supplemented with 0.3% (left), 0.4% (middle), or 0.5% (right) methylcellulose; swimming sperm cells were recorded within the next 5 min; video rate 50 fps, 2 s movies. (A) CatSperz +/- and (B) CatSperz-/- spermatozoa.
Video 7
Motility of tethered CatSperz-/- sperm in high external calcium; 90 min capacitated, related to Figure 6—figure supplement 1.
After 90 min incubation in Ca2+-HTF under capacitating conditions, CatSperz-/- spermatozoa were tethered to a fibronectin-coated glass bottom dish; sperm motility was recorded within the next 5 min at 37°C; video rate 100 fps (1/2 speed), 1 s movies. (A) 2 Ca2+-HTF and (B) 4 Ca2+-HTF (in mM).
Video 8
Motility of tethered CatSperz+/- sperm in low external calcium; 90 min capacitated, related to Figure 6—figure supplement 1.
After 90 min incubation in Ca2+-HTF under capacitating conditions, CatSperz+/- spermatozoa were tethered to a fibronectin-coated glass bottom dish; sperm motility was recorded within the next 5 min at 37°C; video rate 100 fps (1/2 speed), 1 s movies. (A) 2 Ca2+-HTF, (B) 0.5 Ca2+-HTF and (C) 0.1 Ca2+-HTF (in mM).
Video 9
Motility of tethered CatSperz +/- and -/- sperm after A23187 treatment; 5 min after wash, related to Figure 6—figure supplement 1.
Spermatozoa treated with 20 µM A23187 in H-HTF for 10 min were washed and incubated in HTF under capacitating conditions for 5 min; sperm were tethered to a fibronectin-coated glass bottom dish and the motility was recorded in H-HTF within the next 5 min at 37°C; video rate 100 fps (1/2 speed), 1 s movies. (A) CatSperz+/- and (B) CatSperz-/- spermatozoa.
Video 10
Motility of tethered CatSperz +/- and -/- sperm after A23187 treatment; 30 min after wash, related to Figure 6—figure supplement 1.
Spermatozoa treated with 20 µM A23187 in H-HTF for 10 min were washed and incubated in HTF under capacitating conditions for 30 min; sperm were tethered to a fibronectin-coated glass bottom dish and the motility was recorded in H-HTF within the next 5 min at 37°C; video rate 100 fps (1/2 speed), 1 s movies. (A) CatSperz+/- and (B) CatSperz-/- spermatozoa.
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
CatSperζ regulates the structural continuity of sperm Ca2+ signaling domains and is required for normal fertility
eLife 6:e23082.
https://doi.org/10.7554/eLife.23082




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}