Ionic mechanisms underlying history-dependence of conduction delay in an unmyelinated axon
- New Jersey Institute of Technology, United States
- NJIT and Rutgers University, United States
Figures
Figure 1
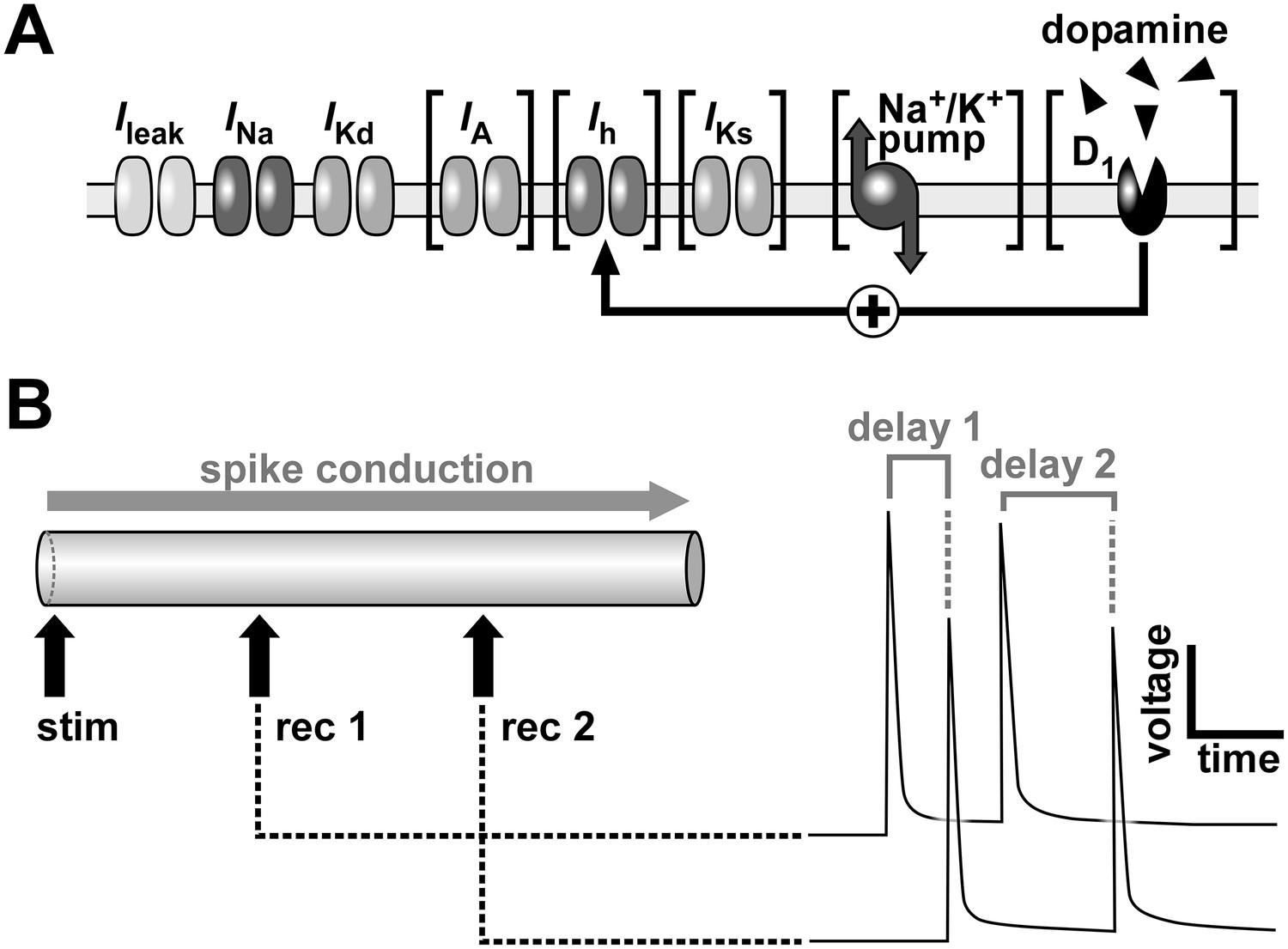
Schematic of the model axon and conduction delay.
(A) Schematic of ion channels, the Na+/K+ pump and dopamine receptors in the model axon. Brackets indicate components that were omitted in some simulations. (B) The model axon was stimulated at one end and the conduction delay was measured between two positions at 0.3 and 0.7 times the length of the axon. Right panel shows conduction delays between the recording sites for two consecutive action potentials.
Figure 2
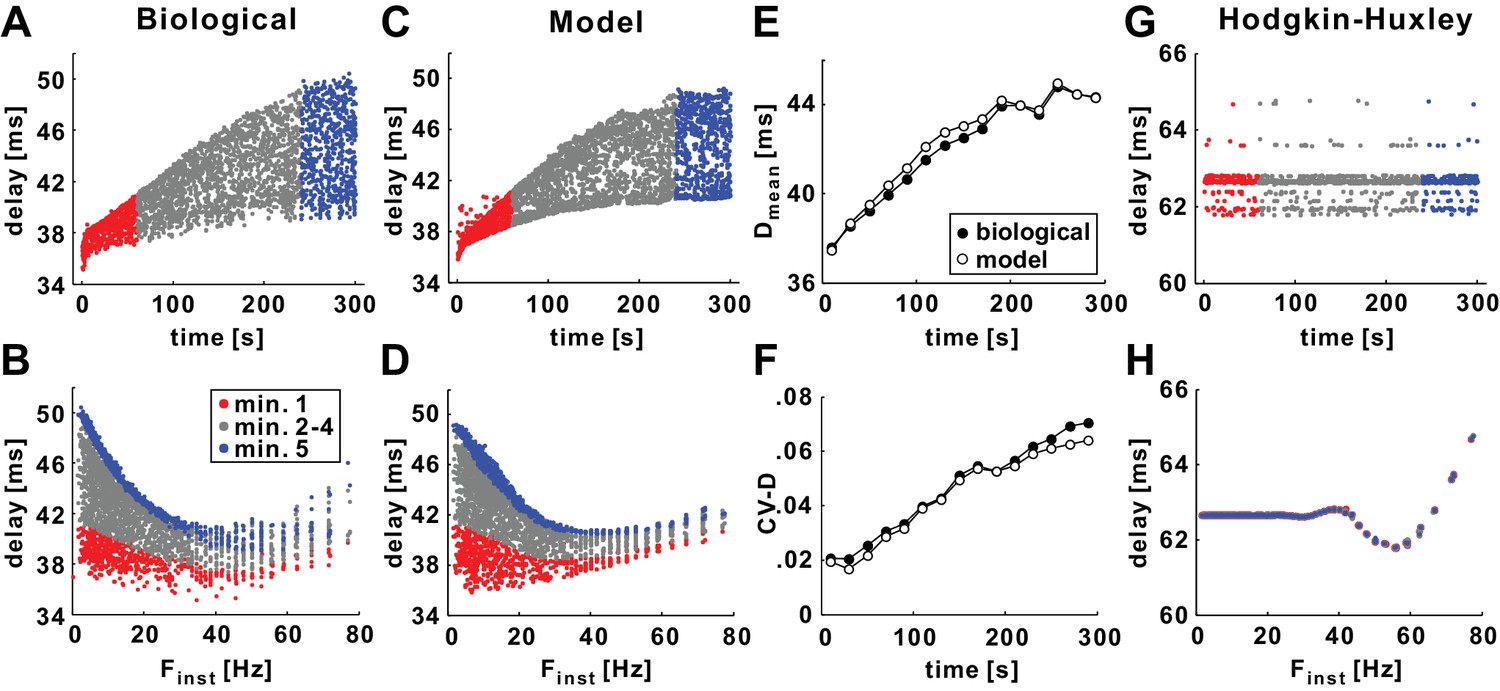
History-dependence of conduction delay in the model axon (with gh = 0).
(A) Delay shown as a function of stimulus time for a 10 Hz (mean rate), 5 min Poisson stimulation of the axon of the biological PD neuron. Red and blue, respectively, mark delay values for the first and fifth minute of stimulation. (B) The data in panel A, plotted as a function of the instantaneous stimulus frequency (Finst). Colors as in panel A. (C–D) The same Poisson stimulation as in panel A, applied to the model axon produces results that are quantitatively similar to the experimental data. (E–F) The mean (Dmean) and the coefficient variation (CV-D) of the data in panels A and C, measured in 20 s bins. (G–H) The same Poisson stimulation of panels C-D, applied to the standard Hodgkin-Huxley model axon.
Figure 3
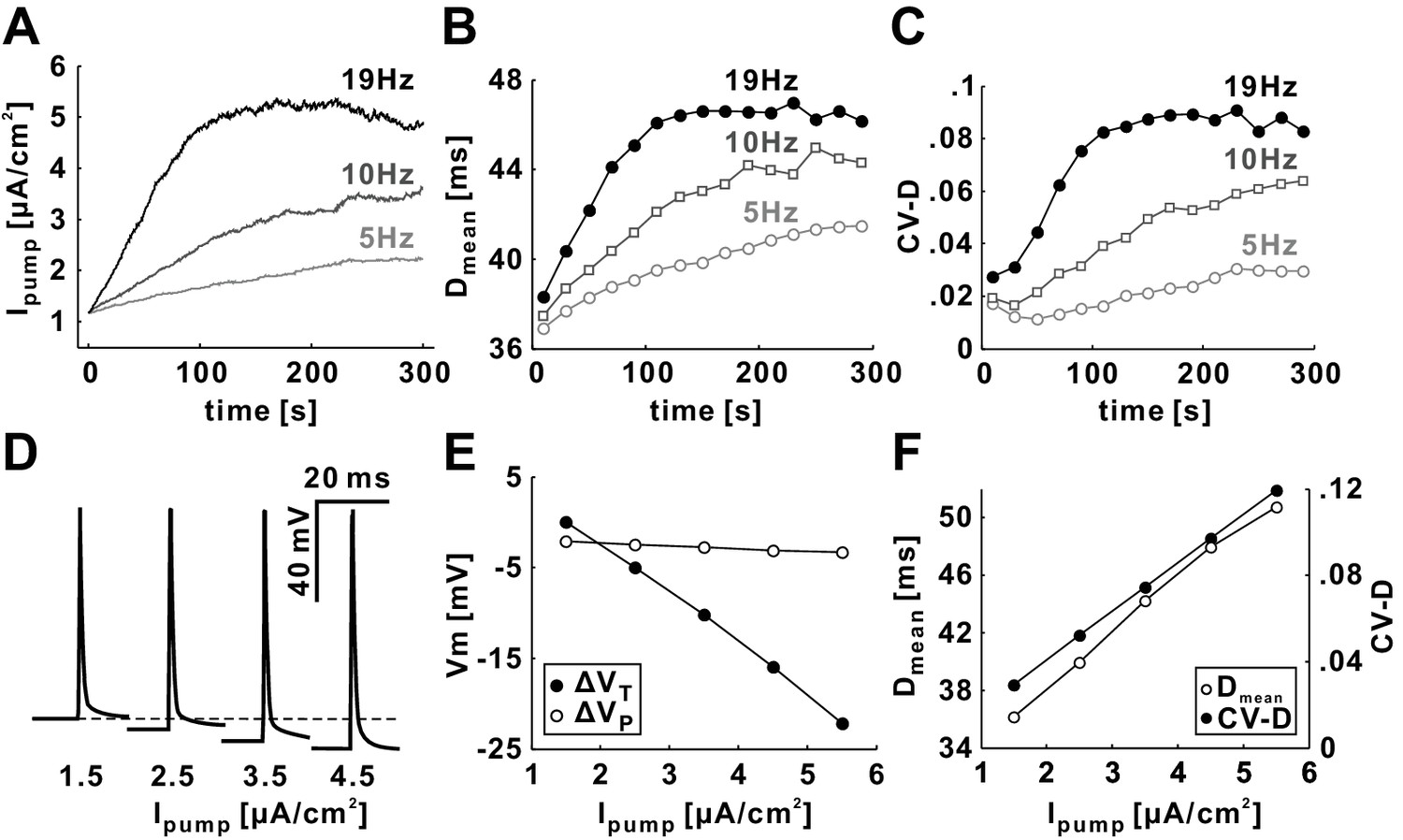
Dmean and CV-D strongly depend on the activity level of Ipump at all Poisson stimulation frequencies.
(A) The value of Ipump for Poisson stimulations with different mean rates (5, 10 and 19 Hz). (B–C) Dmean and CV-D increase with time as well as with the mean rate of Poisson stimulation. (D) The baseline membrane potential is hyperpolarized as the level of Ipump is increased. (E) The trough voltage (VT) linearly decreases with Ipump, but the peak voltage (VP) does not change much when Ipump is increased. (F) Dmean and CV-D linearly increase with the level of Ipump (set to a constant value in each simulation run).
Figure 4
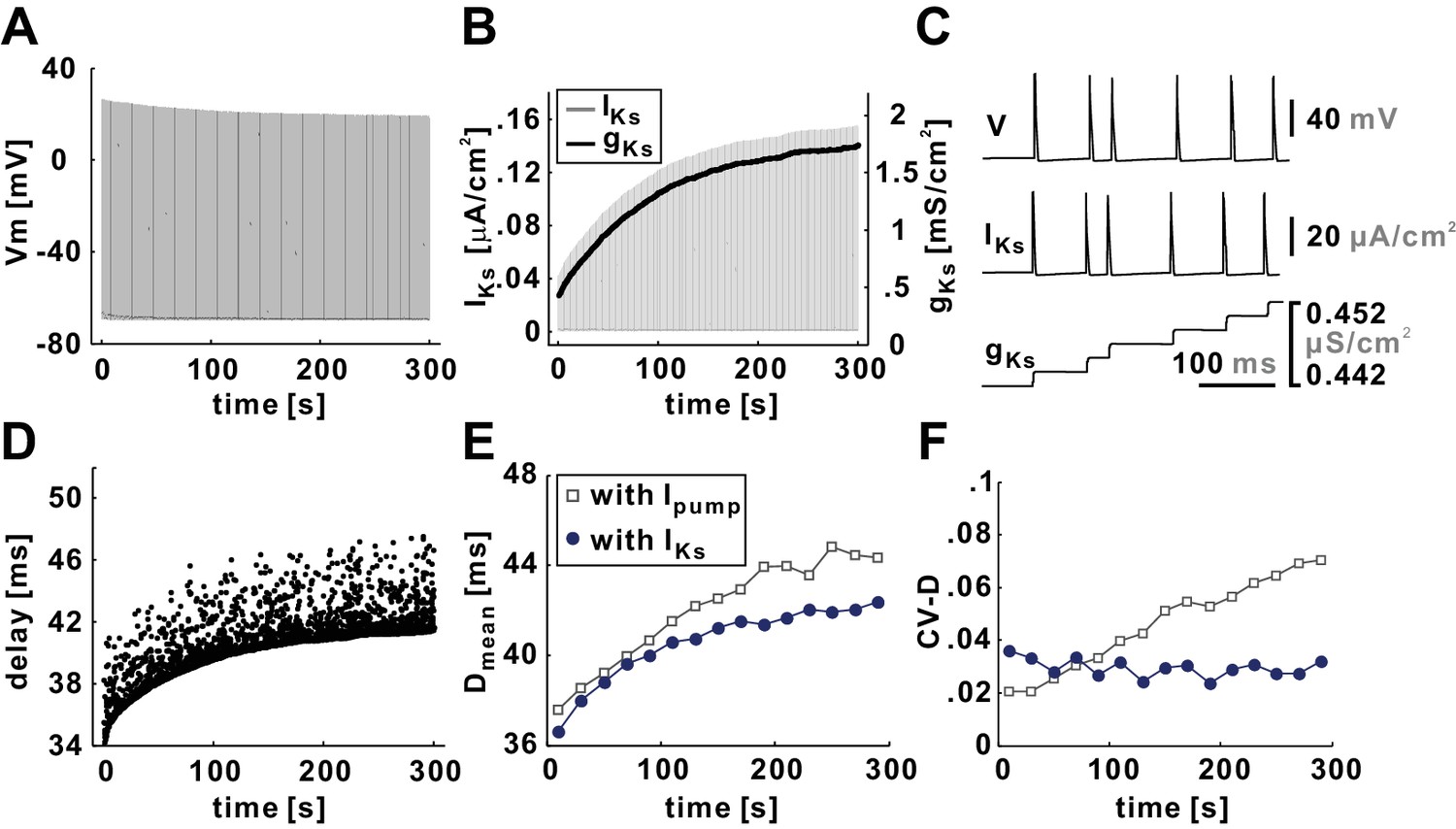
A slow potassium current (IKs) fails to capture the slow timescale effects on conduction delay.
(A) Voltage trace of the model axon, with IKs substituted for Ipump, for a 5 min, 10 Hz Poisson stimulation. (B) Both gKs and the peak values of IKs increase with time during the stimulation process. (C) A small time-window shows that, although gKs accumulates with time, IKs vanishes after each action potential because of the drop in the driving force. (D) Conduction delay of the model axon in response to a 5 min, 10 Hz Poisson stimulation. (E) Dmean increases with time but does not quite match the results of the model axon with Ipump. (F) CV-D does not increase with time as it does in the model axon with Ipump.
Figure 5

Changing the level of Ih in the model mimics the experimental effects of CsCl and DA.
(A1–B1) At the slow timescale, DA results in an increase of gh, which in turn causes Dmean and CV-D stay to remain relatively constant in response to a 5 min 10 Hz Poisson stimulation. In contrast, blocking Ih with CsCl results in an increase in Dmean and CV-D (Ballo et al., 2012). (A2–B2) This effect is mimicked by changing gh levels and the activation curve of Ih in the model. (C1) Changes in gh levels by DA or CsCl result in changes in the minimum delay (Dmin) and curvature (κmin) values in the delay vs. Finst plots (for the 5th minute of the Poisson stimulation). (C2) These effects are mimicked by the models with different activity level of Ih (see Materials and methods and Tables 1 and 2 for details).
Figure 6
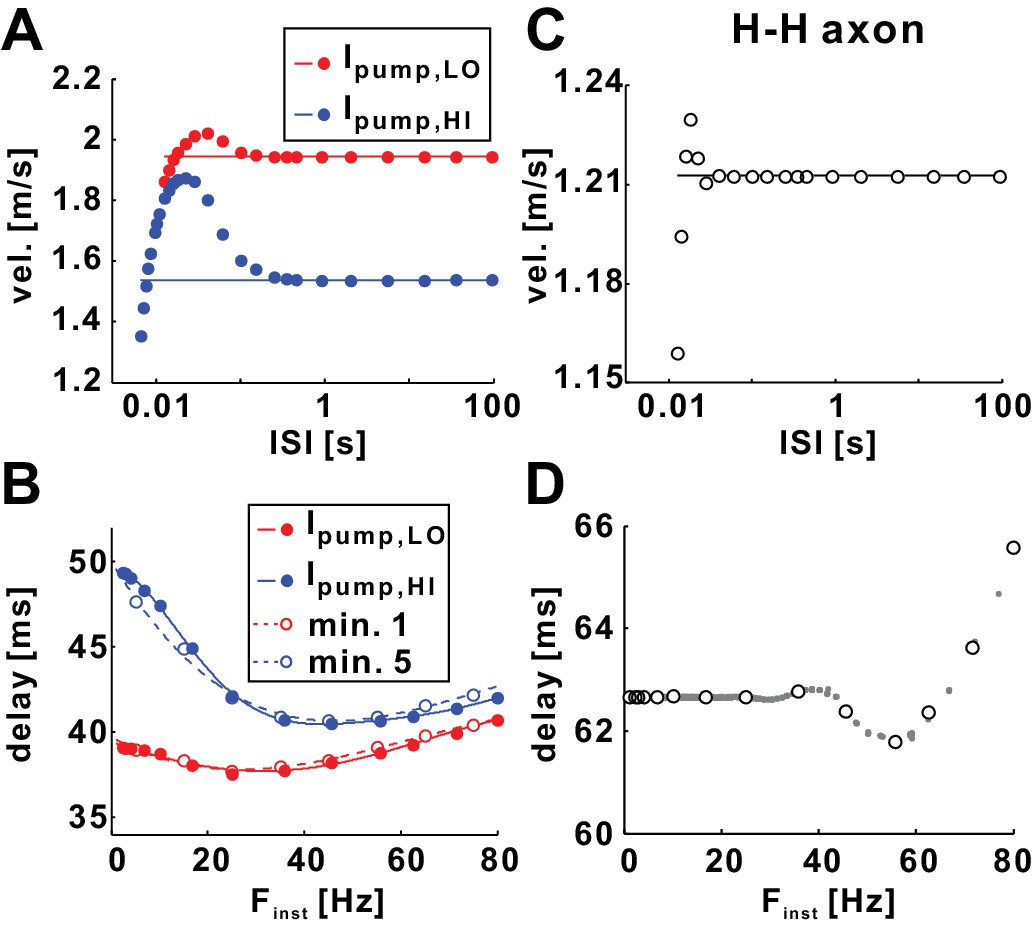
The fast timescale effect can be predicted by paired-pulse stimulations.
(A) Conduction velocity of conditioning (horizontal line) and test (filled circles) pulses plotted as a function of interspike interval (ISI). The graph shows the refractory and supernormal phases, where the velocity of the test pulse is, respectively, less or more than that of the conditioning pulse. Simulations were done with two constant pump currents: Ipump,LO and Ipump,HI, respectively equal to the mean value of Ipump during minute 1 and minute 5 of the 10 Hz Poisson stimulation (Figure 2). Horizontal lines show the velocity of the conditioning spikes. (B) Data of panel A plotted as conduction delay vs. Finst (=1/ISI), together with the Poisson stimulation results of minutes 1 and 5 (from Figure 2D). Solid curves are polynomial fits for the Ipump,LO and Ipump,HI datasets. (C) Paired-pulse stimulations of the Hodgkin-Huxley model axon show similar refractory and supernormal phases. (D) The data from the panel C compared with the Poisson stimulation data (from Figure 2H) of the Hodgkin-Huxley model axon show a perfect match.
Figure 7
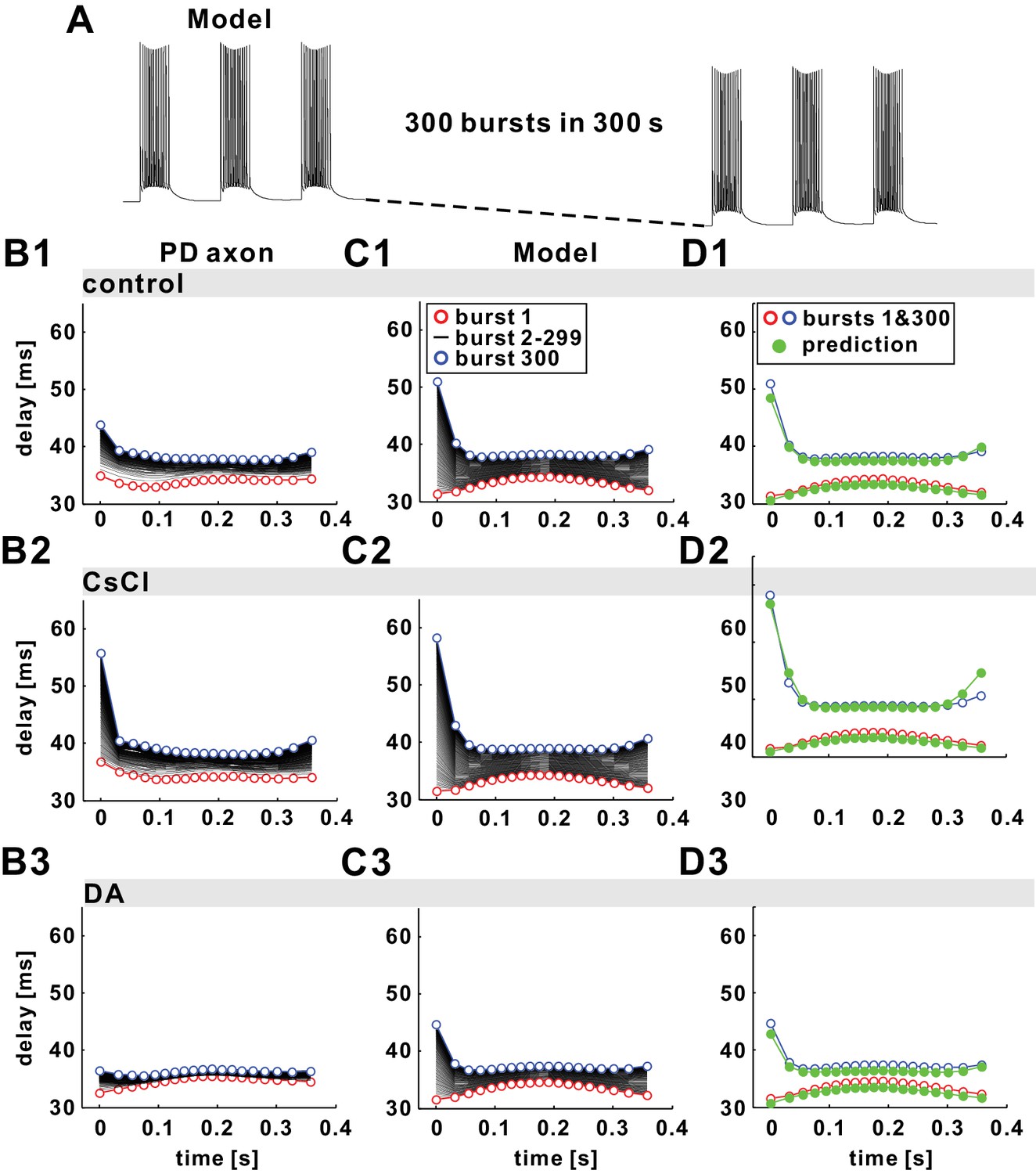
The fast timescale effect observed in the burst stimulations can be predicted by the paired-pulse stimulation results.
(A) The model responses to 300 parabolic burst stimuli (350 ms duration; 1 s period) over 300 s show a drop in baseline membrane potential. (B1–B3) Experimental results of conduction delay of spikes in each burst shown, superimposed as a function of time from the burst onset. Data of the first (red) and the 300th (blue) burst are colored. (C1–C3) Model replication of the experimental results, with different Ih levels (see Tables 1 and 2 for model parameters). (D1–D3) Simulation results of the first and the 300th burst in panel C1-C3 are predicted by estimating history-dependence using paired-pulse simulations (see Figure 6). Paired-pulse simulations were done with a constant low or high Ipump level, equal, respectively, to the Ipump mean value during the first and the 300th burst, fit with polynomial functions and used to obtain the plotted predictions (green).
Figure 8
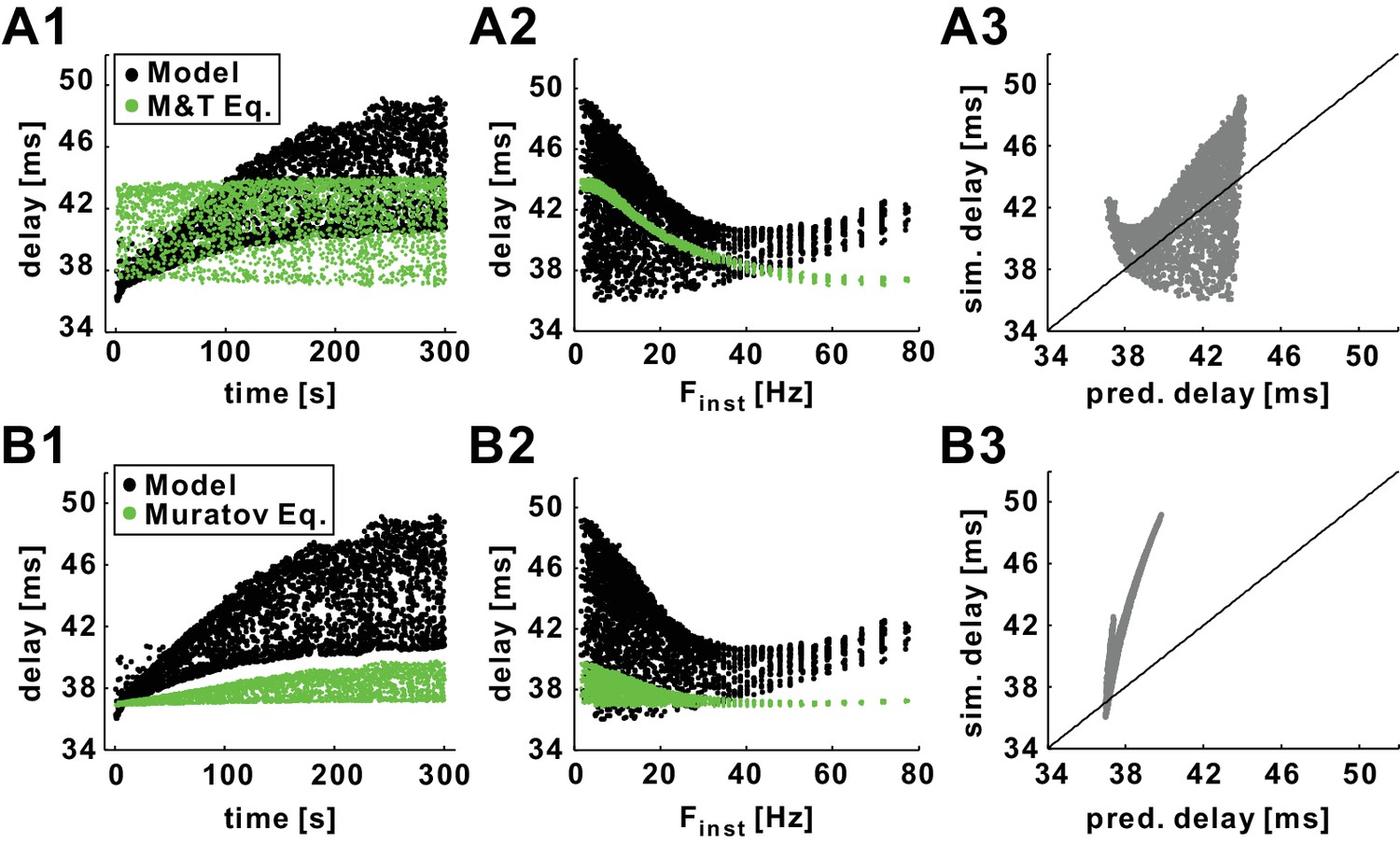
Predictions of history-dependence of conduction delay by previously published equations of action potential velocity.
(A1) Conduction delays of the biophysical model in response to 10 Hz Poisson stimulation and delays predicted by the Matsumoto and Tasaki equation (Tasaki and Matsumoto, 2002). (A2) Data in panel A1 plotted versus Finst. (A3) Simulation delay versus delay predicted by Matsumoto and Tasaki equation. (B1–B3) Comparison between simulation delays in the biophysical model (as in A1-C1) and the delays predicted by the Muratov equation (Muratov, 2000). Diagonal lines are y=x.
Figure 9
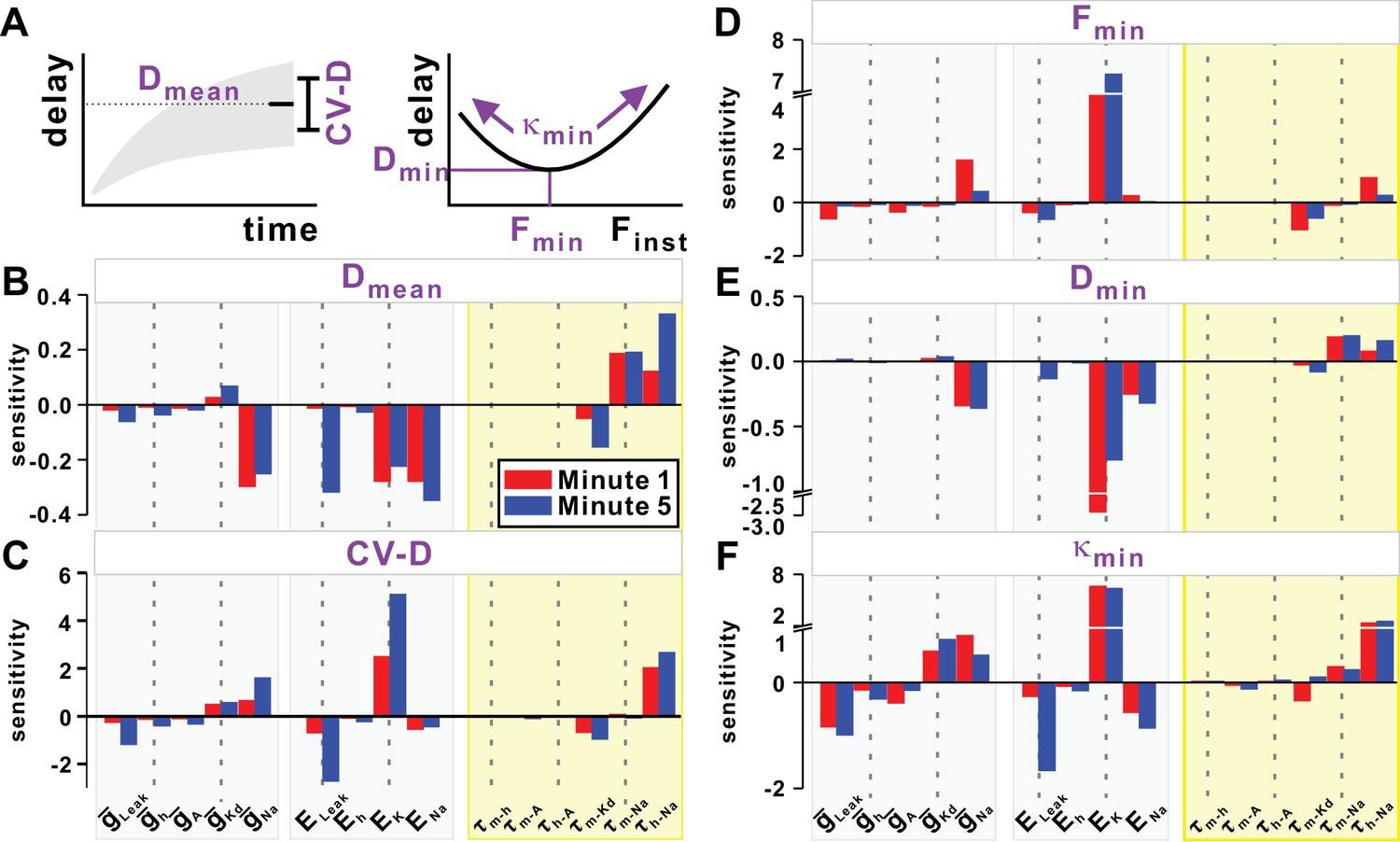
Sensitivity of the slow (STS) and fast timescale (FTS) effects to the model parameters.
(A) Top left inset schematically shows the mean (Dmean) and variability of conduction delay (CV-D = coefficient of variation), measured in the last 20 s of simulation. Top right inset schematically shows the minimum delay point (Fmin, Dmin) and the curvature κmin of the quadratic fit at this point. (B–C) STS effect. Sensitivity of Dmean and CV-D to model parameters in the first and five minute of a 10 Hz Poisson stimulation. (D–F) FTS effect. Sensitivity of Fmin, Dmin and κmin to model parameters in the first and fifth minute of a 10 Hz Poisson stimulation.
Figure 10
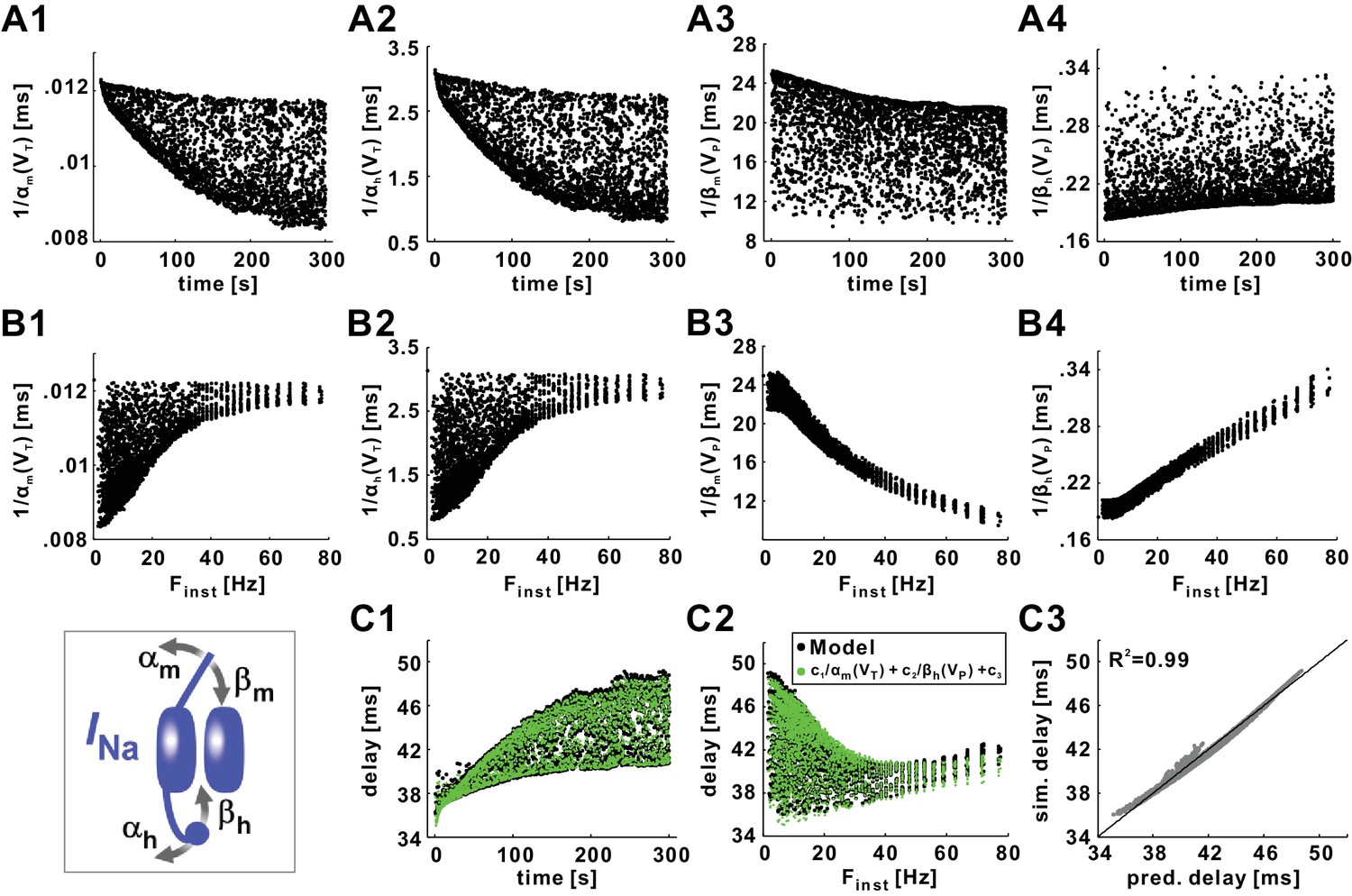
Conduction delay is determined by the INa activation and inactivation variables, evaluated at the action potential trough (VT) or peak (VP) voltage.
(A1–A2) Reciprocal of the INa activation rate, evaluated at VT. (A3–A4) Reciprocal of the INa inactivation rate, evaluated at VP. (B1–B4) Data in panel A plotted as a function of Finst. (C1) Conduction delay of the model compared with the predicted delay of c1/αm(VT)+c2/βh(VP)+c3 (c1 = −3361.48, c2 = 32.80, c3 = 70.14). (C2) Data in panel C1 plotted as a function of Finst. (C3) Simulation delays compared with predicted delays from the empirical equation (R2=0.99). The line is y=x. The inset is the schematic of the fast sodium channel.
Figure 11
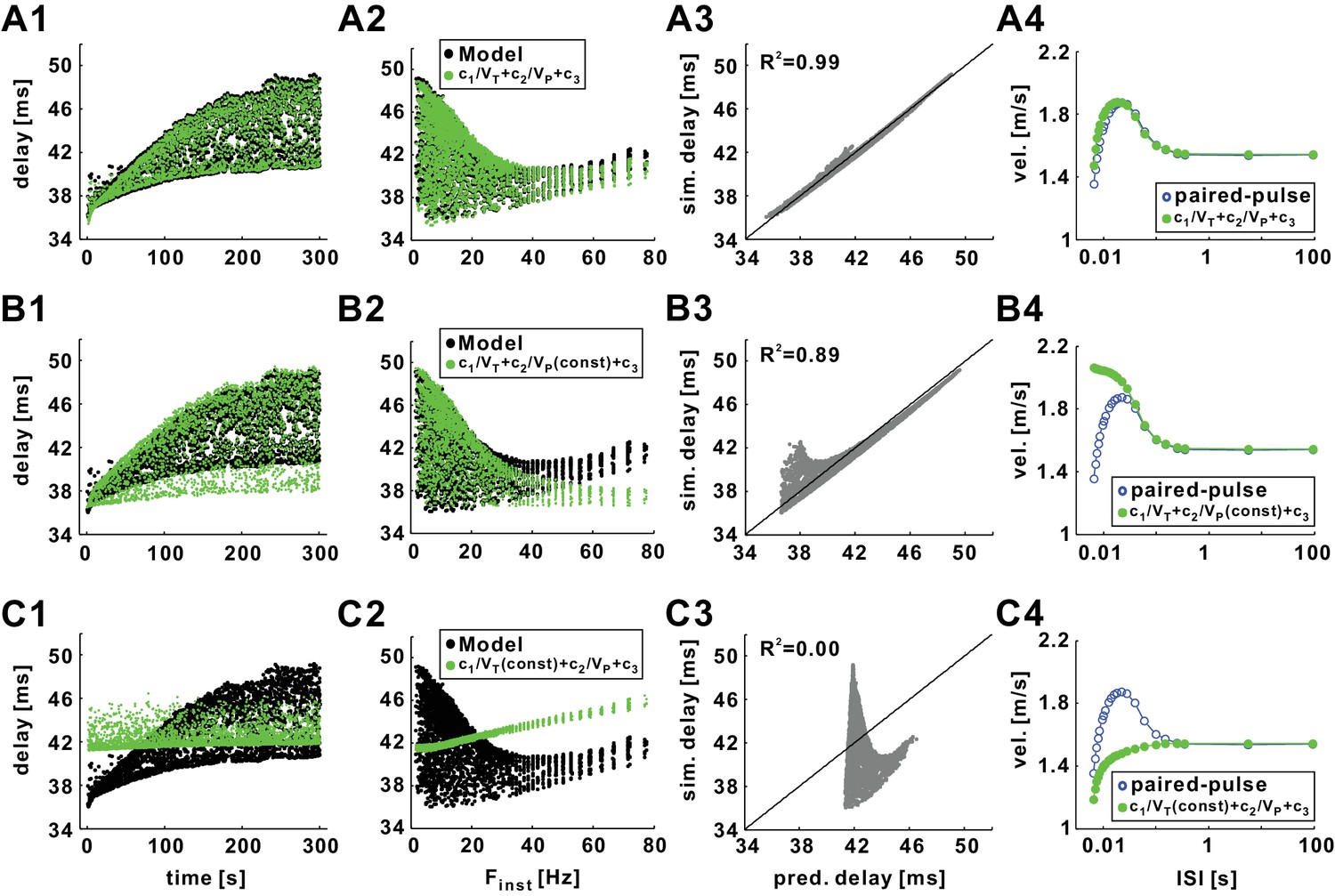
Conduction delay can be almost perfectly predicted by the trough and peak voltages of the action potentials.
(A1) Superimposed graph of conduction delays of the biophysical model (black) in response to 10 Hz Poisson stimulation and the predicted delays from the nonlinear regression equation: d = c1/VT +c2/VP +c3 (c1 = 4072.88, c2 = 302.36, c3 = 82.94; the same values of c1-c3 are used in all panels). (A2) Data in panel A1 plotted as a function of Finst. (A3) Simulation delays (training data) compared with predicted delays (R2 = 0.99). (A4) Conduction delays of the biological model (blue) in response to paired-pulse stimulation (same as in Figure 6A) and the predicted delays from the regression equation in panel A1. (B1–B3) Simulation results (same as in A1-A3) compared with predicted delays from the single variable regression: d = c1/VT +c2/VP (mean) +c3, where VP (mean) is the mean value of Vp. (B4) Simulation results (same as in A4) compared with predicted delays from the single variable regression: d = c1/VT +c2/VP (cond) +c3, where VP (cond) is the value of Vp of the conditioning pulse. (C1–C3) Simulation results (same as in A1-A3) compared with predicted delays from the single variable regression: d = c1/VT(mean)+c2/VP +c3, where VT (mean) is the mean value of VT. The lines in A3, B3, and C3 are y=x. (C4) Simulation results (same as in A4) compared with predicted delays from the single variable regression: d = c1/VT(cond)+c2/VP +c3, where VT (cond) is the value of VT of the conditioning pulse.
Figure 12
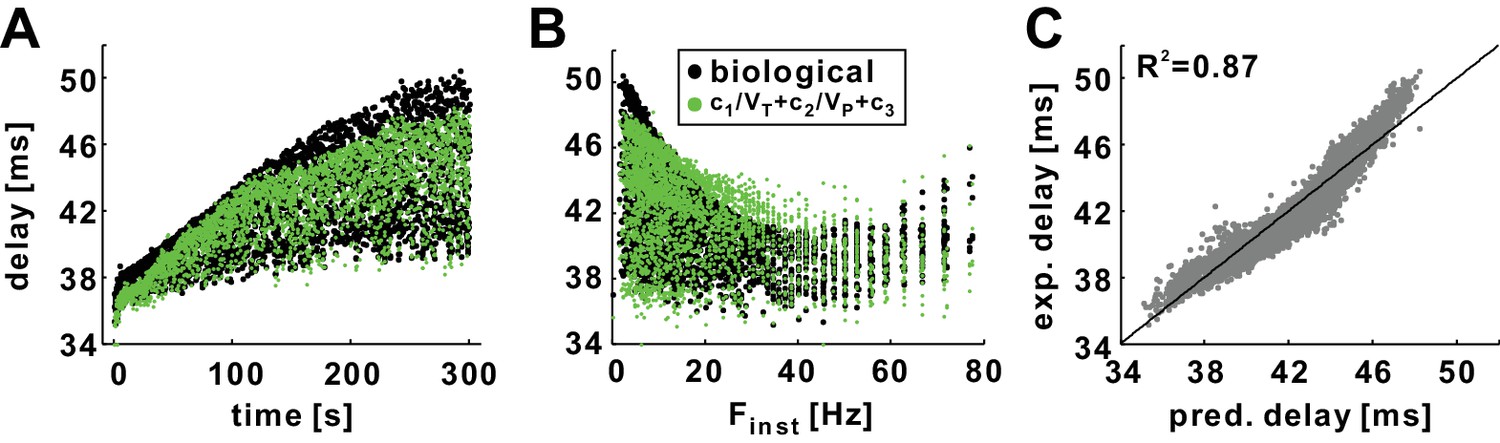
The history dependence of conduction delay in the biological PD axon can be predicted by the empirical equation without need for computational modeling.
(A) Conduction delays in the biological PD axon in response to a 5 min, 10 Hz, Poisson stimulation (black; same as in Figure 2A) compared with delays predicted by empirical equation (c1=4817.75, c2 = 500.5, c3 = 105) using VT and VP values from intracellular recordings (not shown). (B) Data in panel A plotted as a function of Finst. (C) Experimental delays plotted versus the predicted delays (R2 = 0.87). The line is y=x.
Tables
Table 1
Model parameters.
| Ionic Curr | [mS/cm2] | [mV] |
|---|---|---|
| INa | 14 | Dynamic |
| IKd | 3 | −70 |
| ILeak | 0.125 | −65 |
| IA | 5 | −70 |
| Ih (ctrl) | 0.05 | −32 |
| Ih (DA) | 0.1 | −25 |
| IKs | 3 | −70 |
| Other | Value | |
| Imax,pump | 2 mA/cm2 | |
| [Na+]1/2 | 78 mM | |
| [Na+]S | 2 mM | |
| α | 7.4 | |
| Vol | 7850 µm3 | |
| F | 96485 C/mol | |
Table 2
Parameters of ionic current steady-state activation and inactivation and their time constants.
| Ionic Curr | m, h | [ms] | |
|---|---|---|---|
| INa | m3 | ||
| h | |||
| IKd | m4 | ||
| IA | m3 | ||
| h | 50 | ||
| Ih (ctrl) | m | 3700 | |
| Ih (DA) | m | 3800 | |
| IKs | m |
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Ionic mechanisms underlying history-dependence of conduction delay in an unmyelinated axon
eLife 6:e25382.
https://doi.org/10.7554/eLife.25382




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}