Initial elevations in glutamate and dopamine neurotransmission decline with age, as does exploratory behavior, in LRRK2 G2019S knock-in mice
- University of British Columbia, Canada
- McGill University, Canada
- Mayo Clinic, United States
Figures
Figure 1 with 1 supplement

Increased exploratory behavior in young GKI mice declines with age.
(A) Animal body weight over time. There was no main effect of genotype upon mouse weight, but there was a significant interaction between weight and age (see text and Figure 1—figure supplement 1). Weights were not significantly different at any age up to 500 days (i; animal n shown in ii); however, in the oldest age group there was a statistically significant increase in GKI mean weight (post-test *p=0.018). (B) Spontaneous exploratory behavior in young and old mice. Analysis of open field exploration showed no genotype (p=0.9) or interaction (p=0.9) effects on total distance travelled (i). In the cylinder test for vertical exploration (ii) there was a significant interaction between genotype and age upon rearing events (p=0.008). At < 6 months, GKI mice exhibited a significant 24.8% increase in vertical exploration (post-test **p=0.009), which declined significantly with age (post-test ****p=0.0001). No significant age-related decline was observed in WT littermates (post-test p=0.3).
Figure 1—figure supplement 1
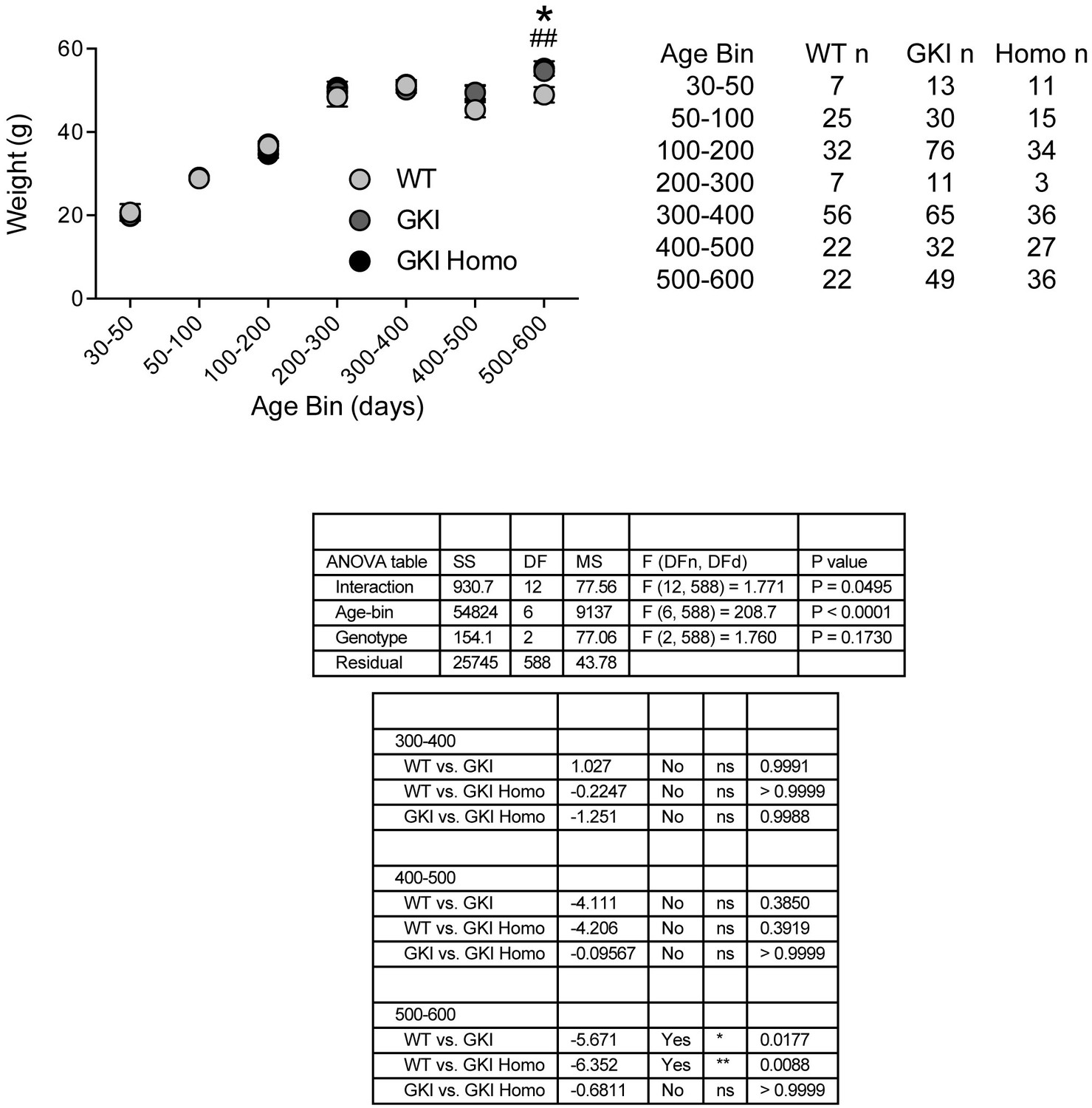
Weights of WT, GKI and GKI homozygous animals are similar until advanced ages.
Animal body weight over time. There was no main effect of genotype upon mouse weight, but there was a significant interaction between weight and age (2-way ANOVA, genotype p=0.17, interaction p=0.049). Weights were not significantly different at any age up to 500 days; however, in the oldest age group there was a statistically significant increase in GKI (Holm-Sidak post-test *p=0.018) and GKI Homo (post-test ##p=0.009) mean weight, relative to WT animals.
Figure 2 with 7 supplements
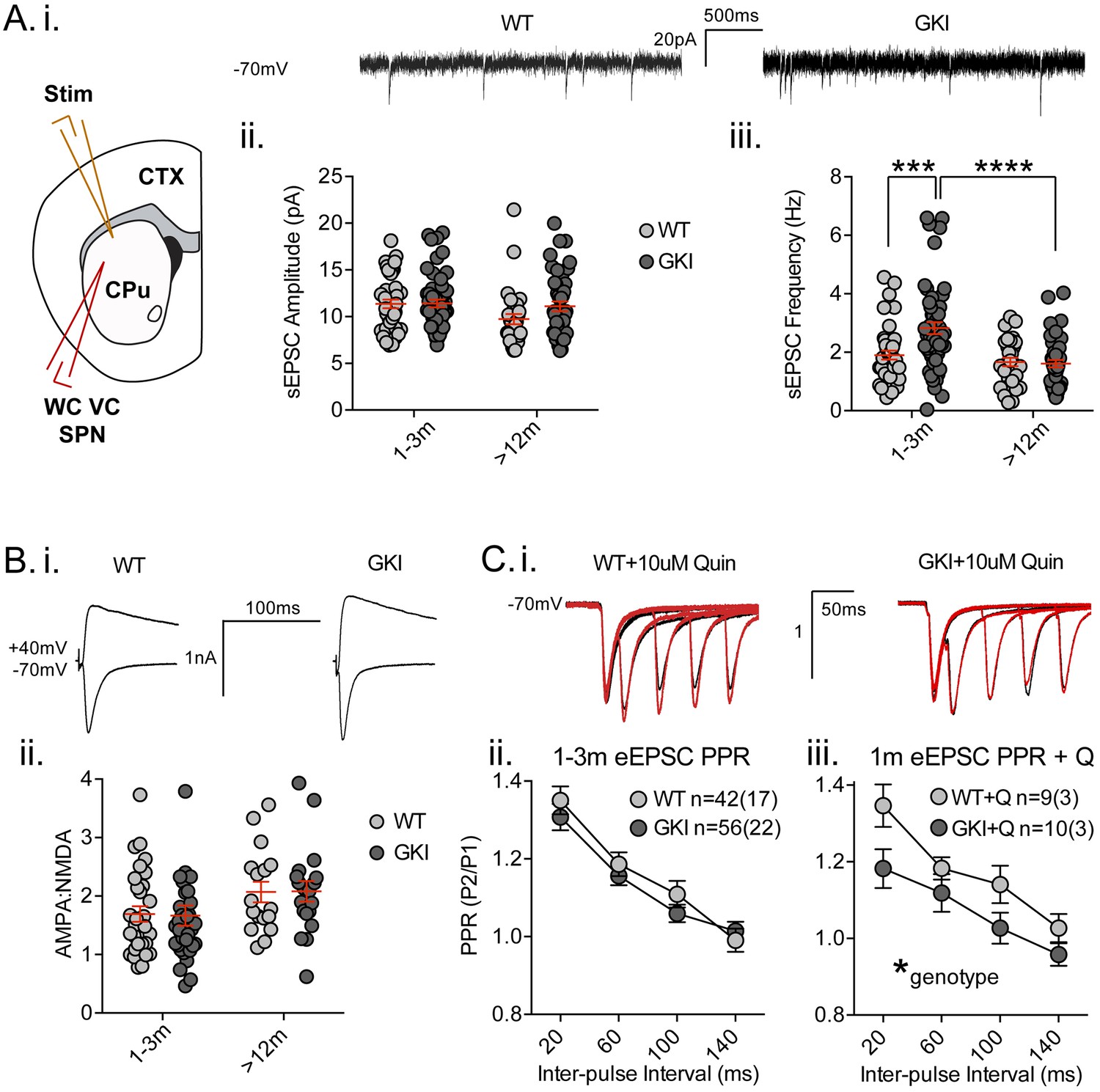
Increased glutamate transmission onto young GKI striatal projection neurons declines with age.
(A) Whole-cell voltage-clamp (WC VC) recording in striatal medium-sized spiny projection neurons (SPNs) of the dorsolateral striatum of acutely prepared brain slices. Recording electrodes targeted visually identified SPNs in the dorsolateral caudate putamen (CPu) of the striatum and stimulating electrodes were placed intrastriatally ~200 um dorsal of the recording site toward M1/S1 cortex (CTX) (i). Example traces of spontaneous activity (spontaneous excitatory post-synaptic currents: sEPSCs) in slices from 1m-old WT (left) and GKI (right) mice. There was a significant main effect of age upon mean sEPSC amplitude (ii, p=0.049), but no main genotype (p=0.15) or age-genotype interaction (p=0.18). In contrast, there were significant main age (0.0001) and genotype (p=0.013) effects and an age-genotype interaction effect (p=0.006) upon the frequency of sEPSCs. At 1–3 months, GKI SPNs exhibited a significant 48.4% increase in mean event frequency (post-test **p=0.0004), which significantly declined (by 57%) with age (post-test ****p=0.0001). In contrast, the 13% decline with age in WT SPNs was not significant (post-test p=0.29). (B) Representative traces of intrastriatally-evoked AMPA- and NMDA-receptor mediated EPSCs (eEPSCs) recorded at Vh −70 mV and + 40 mV, respectively (i). There was a significant main effect of age (ii; p=0.041) upon the ratio of AMPA to NMDA eEPSC amplitudes, but no main genotype (p=0.96) or age-genotype interaction effect (p=0.92). (C) Representative peak-normalized traces of evoked EPSCs (eEPSCs) in SPNs, resulting from intrastriatal paired-pulse stimuli (20–140 ms inter-pulse intervals; IPIs) prior to (i. black trace) and in the presence of 10 uM quinpirole (red trace; slices from 1 month WT and GKI slices). There were no significant genotype or genotype-IPI interaction effects upon paired-pulse ratios in pooled experiments from animals aged 1–3 months in standard conditions (ii). In the presence of the D2 agonist quinpirole (i) there was a significant main genotype effect (*p=0.042) in slices from 1 m old mice (iii).
Figure 2—figure supplement 1
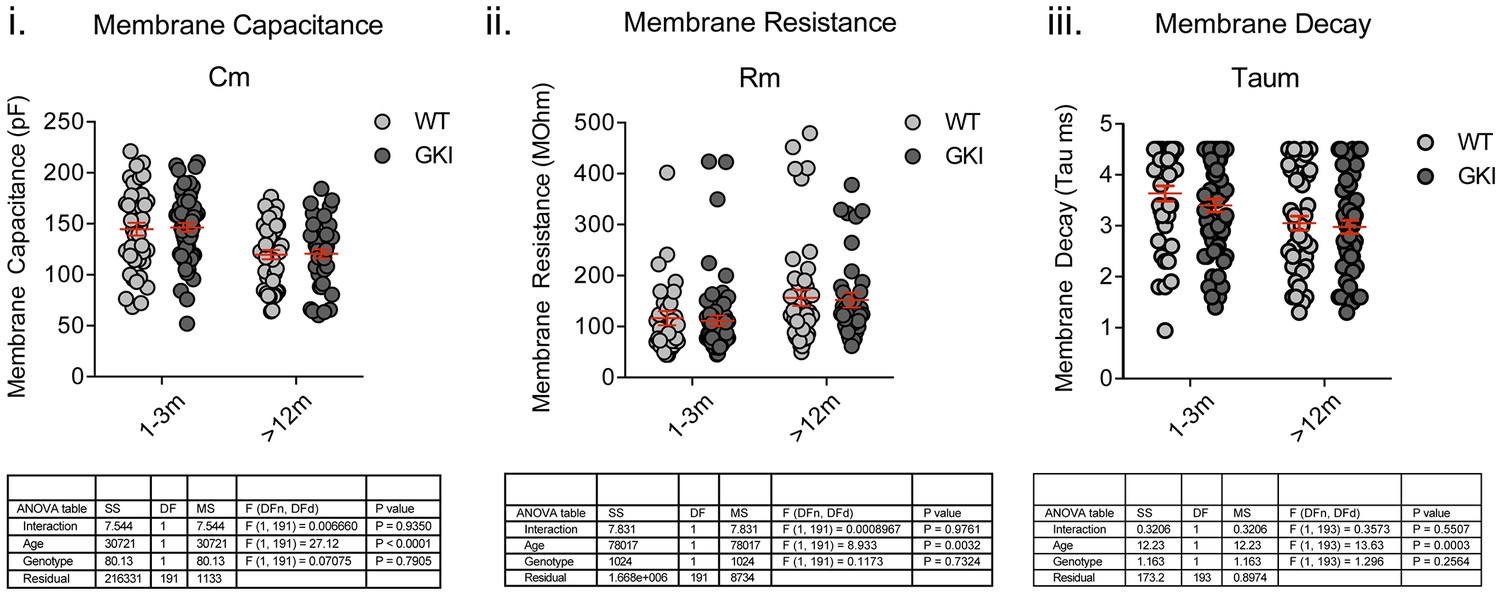
There are no genotype-dependent alterations to SPN intrinsic membrane properties.
Whole-cell recording of medium-sized spiny striatal projection neurons (SPNs) in membrane test mode and analysis of intrinsic membrane properties. While significant age-dependent changes in membrane capacitance (Cm), resistance (Rm) and decay time (Taum) were observed, there were no genotype or interaction effects. 2-way ANOVA detailed below.
Figure 2—figure supplement 2
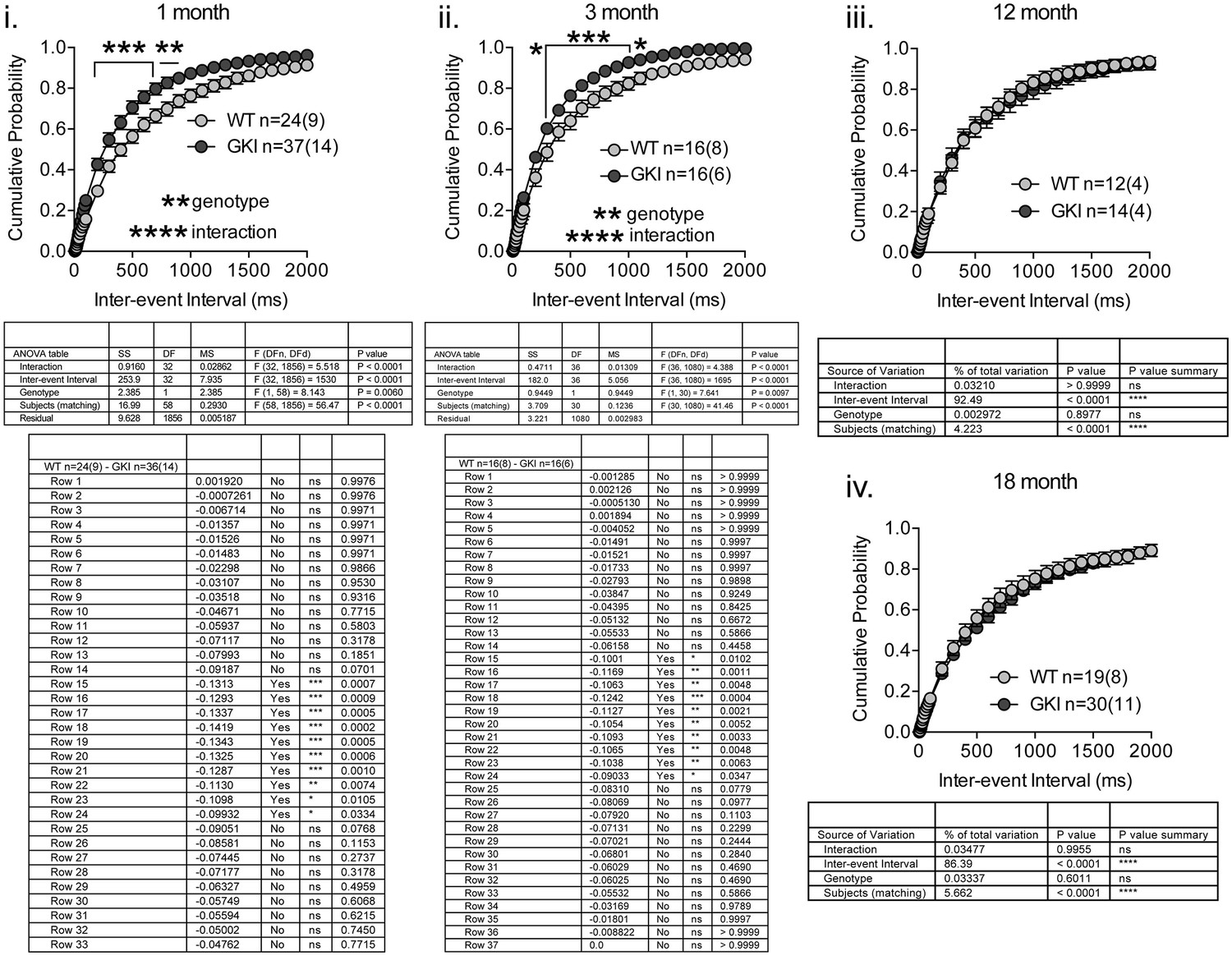
GKI SPN sEPSC frequency is similarly increased in 1 and 3 month slices, and similar to WT in 12 and 18 month slices.
Whole-cell recording in dorsolateral striatal SPNs and analysis of sEPSCs recorded at −70 mV. Higher sEPSC frequencies (as reflected by decreased inter-event intervals) underlie significant main genotype effects, to similar extents, in slices from 1 and 3 month animals, but there were no significant effects of genotype in 12 or 18 month slices. Details of 2-way RM-ANOVA and Holm-Sidak post-tests are shown below.
Figure 2—figure supplement 3
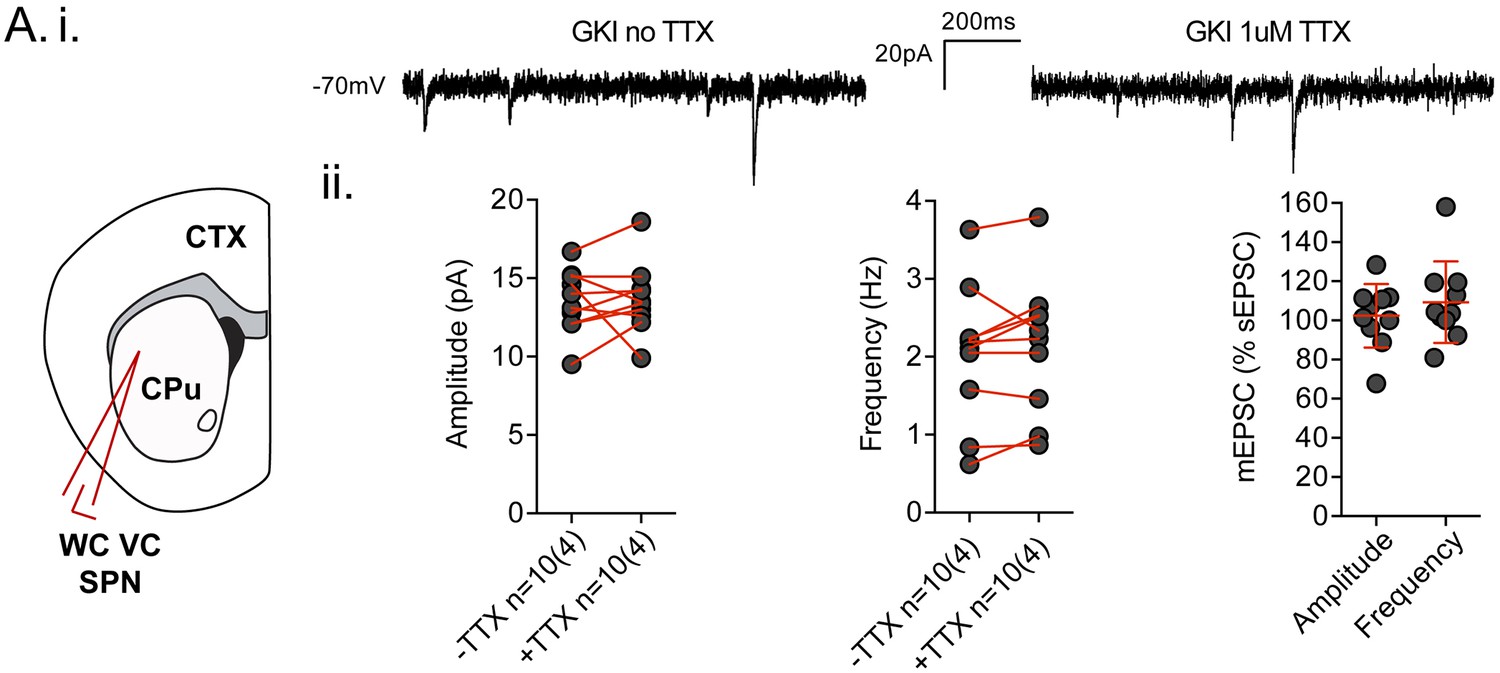
Spontaneous EPSCs recorded in dorsolateral striatum in the coronal slice preparation are TTX insensitive.
(A.i) Example traces of spontaneous EPSCs (sEPSCs) in whole-cell voltage-clamp (WCVC) recordings in striatal projection neurons (SPN) of the dorsolateral striatum, prior to, and in the presence of, tetrodotoxin (TTX; 1 uM) rendering any remaining responses miniature EPSCs (mEPSCs). (ii) There was no significant effect of TTX upon mean sEPSC amplitude (paired t-test p=0.82) or frequency (paired t-test p=0.29) within each cell. Expressed as a percentage of the initial sEPSC mean value, neither mEPSC amplitudes nor frequencies were significantly altered by TTX application (one-sample t-test against 100, p=0.7 and 0.2 respectively). The weak trend in frequency was towards increases in GKI, rather than the decreases predicted if action potential-dependent events were contributing to mean sEPSC frequency.
Figure 2—figure supplement 4
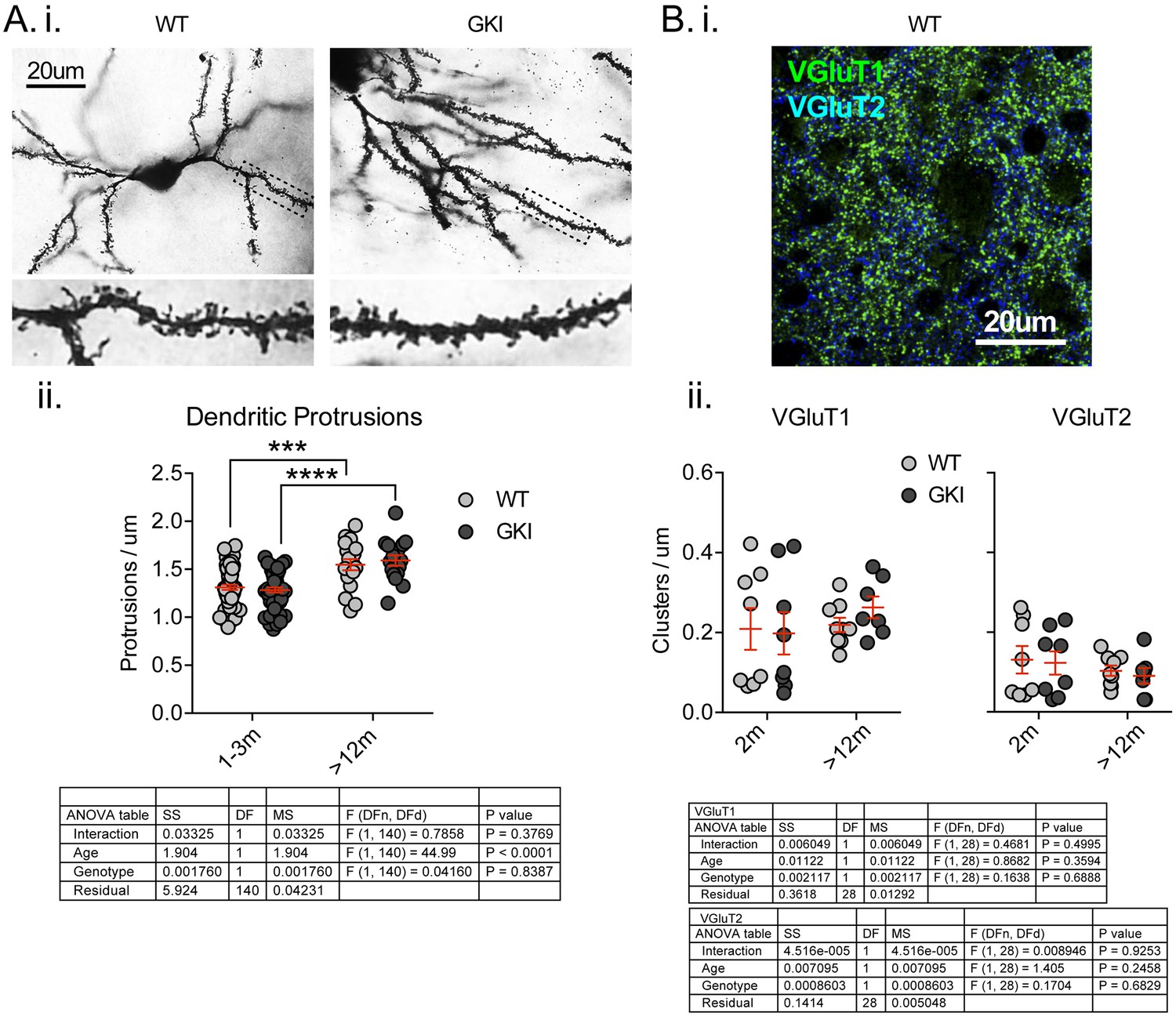
There are no differences in the number of postsynaptic specializations, or presynaptic excitatory nerve terminal markers in the striatum of young or aged GKI mice.
(A) Representative bright-field images of Golgi-impregnated medium-sized spiny projection neurons (SPNs) of the dorsolateral striatum from WT and GKI mice aged 1–3 months (i), with inserts showing a zoom of areas marked in main image by dashed lines. There were significant increases in the density of postsynaptic protrusions as animals aged between 1–3 and > 12 months, but there were no significant differences between WT and GKI dendritic protrusions at either age (WT n = 57(9) and 18(3), GKI n = 56(9) and 16(3), for 1–3 and >12, respectively). (B) Representative confocal micrograph of VGluT1 and VGluT2 immunofluorescence staining in 2 month-old WT dorsolateral striatum, demonstrating the expected pattern of a higher density of VGluT1 than VGluT2 glutamatergic terminals and very little overlap between VGluT1 and VGluT2 (i). There were no significant effects of age or genotype upon VGluT1 or VGluT2 cluster densities (ii; WT n = 8 (4) and 9 (4), GKI n = 8 (4) and 7 (4), for 2 m and >12 m, respectively). Details of 2-way ANOVA and Holm-Sidak post-tests are shown below.
Figure 2—figure supplement 5
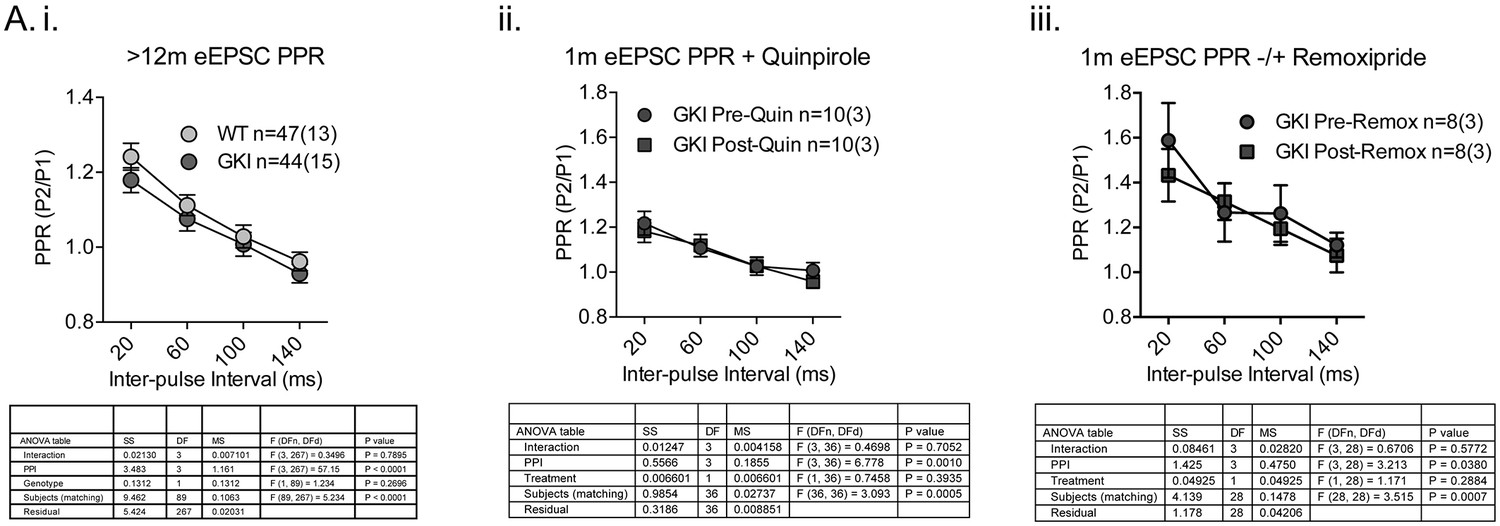
Paired-pulse facilitation profiles in the striatum of aged GKI mice are similar and GKI mutant PPRs are insensitive to dopamine agonism and antagonism.
Whole-cell patch clamp recording and eEPSC paired-pulse experiments. (i) There were no significant differences in glutamatergic paired-pulse ratio at > 12 months, similarly to the 1–3 month age point in the absence of quinpirole (Figure 2.C). At 1 month of age, when WT and GKI PPRs differ significantly in the presence of quinpirole (Figure 2.C), GKI PPRs were insensitive to both dopamine agonism (ii. 10 uM quinpirole) and antagonism (iii. 10 uM remoxipride). Details of 2-way ANOVA and Holm-Sidak post-tests are shown below.
Figure 2—figure supplement 6
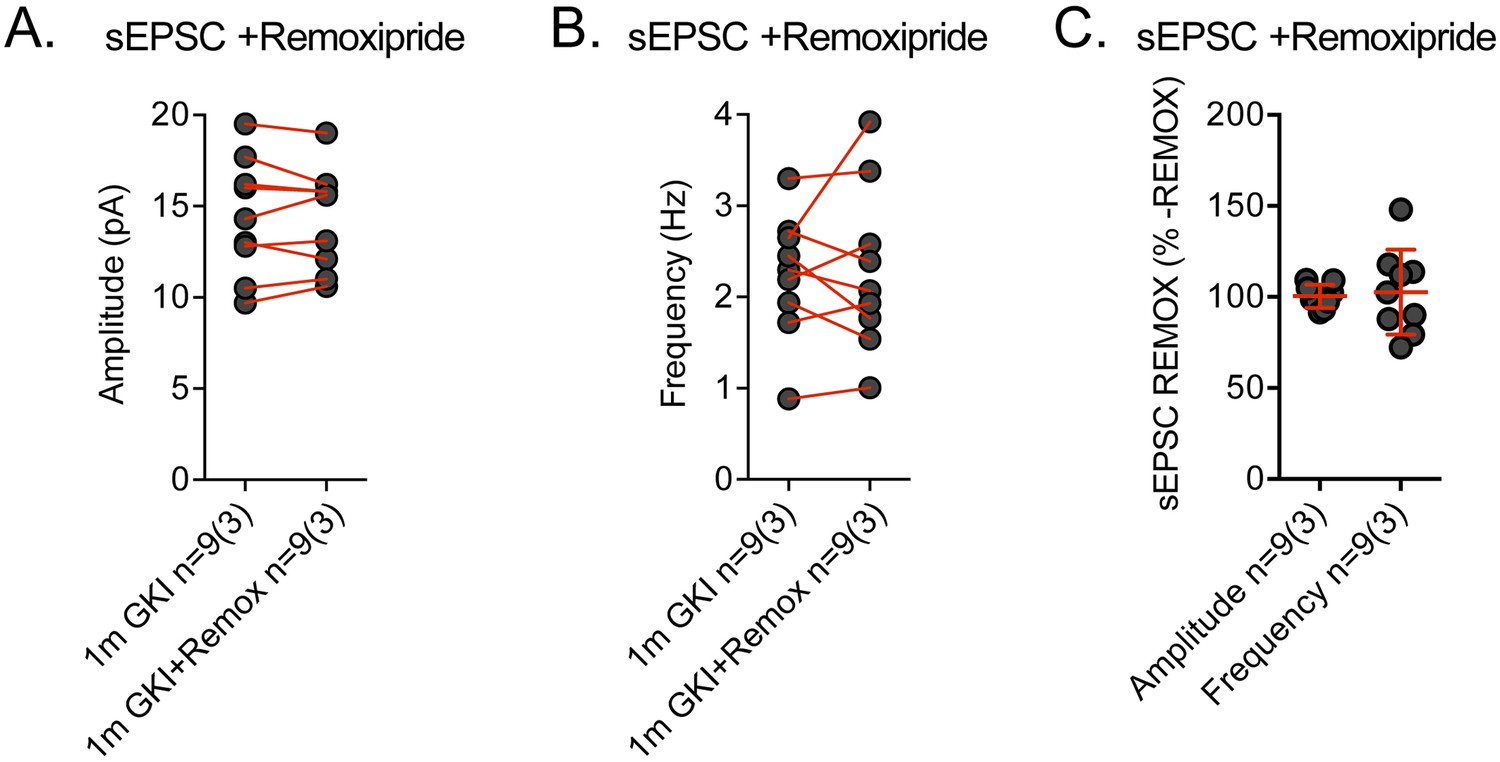
Spontaneous EPSCs recorded in GKI dorsolateral striatum SNPs are remoxipride insensitive.
Spontaneous EPSCs (sEPSCs) were recorded prior to, and in the presence of, remoxipride (Remox; 10 uM). (A) There was no significant effect of remoxipride upon mean sEPSC amplitude (paired t-test p=0.86) or frequency (paired t-test p=0.80) within each cell. Expressed as a percentage of the initial sEPSC mean value, neither amplitude nor frequency was significantly altered by remoxipride application (one-sample t-test against 100, p=0.84 and 0.74, respectively).
Figure 2—figure supplement 7

There were no differences in GKI sEPSC frequency between D1- and D2-dopamine receptor expressing SPNs, and no consistent effect of quinpirole upon eEPSC peak or PPR in either cell type.
Whole-cell patch clamp recording in D1 vs. D2-type SPNs in GKI slices. (A) There were no differences in sEPSC mean event frequency in either cell type at young (1–3 m) or old (>12 m) age points. (B) The D2 agonist quinpirole had no consistent modulatory effect upon D1 or D2 eEPSC peak amplitudes, or Paired-pulse ratios expressed as post-application values over pre-application (C). 2-way ANOVA and unpaired Student’s t-test values are shown.
Figure 3 with 1 supplement
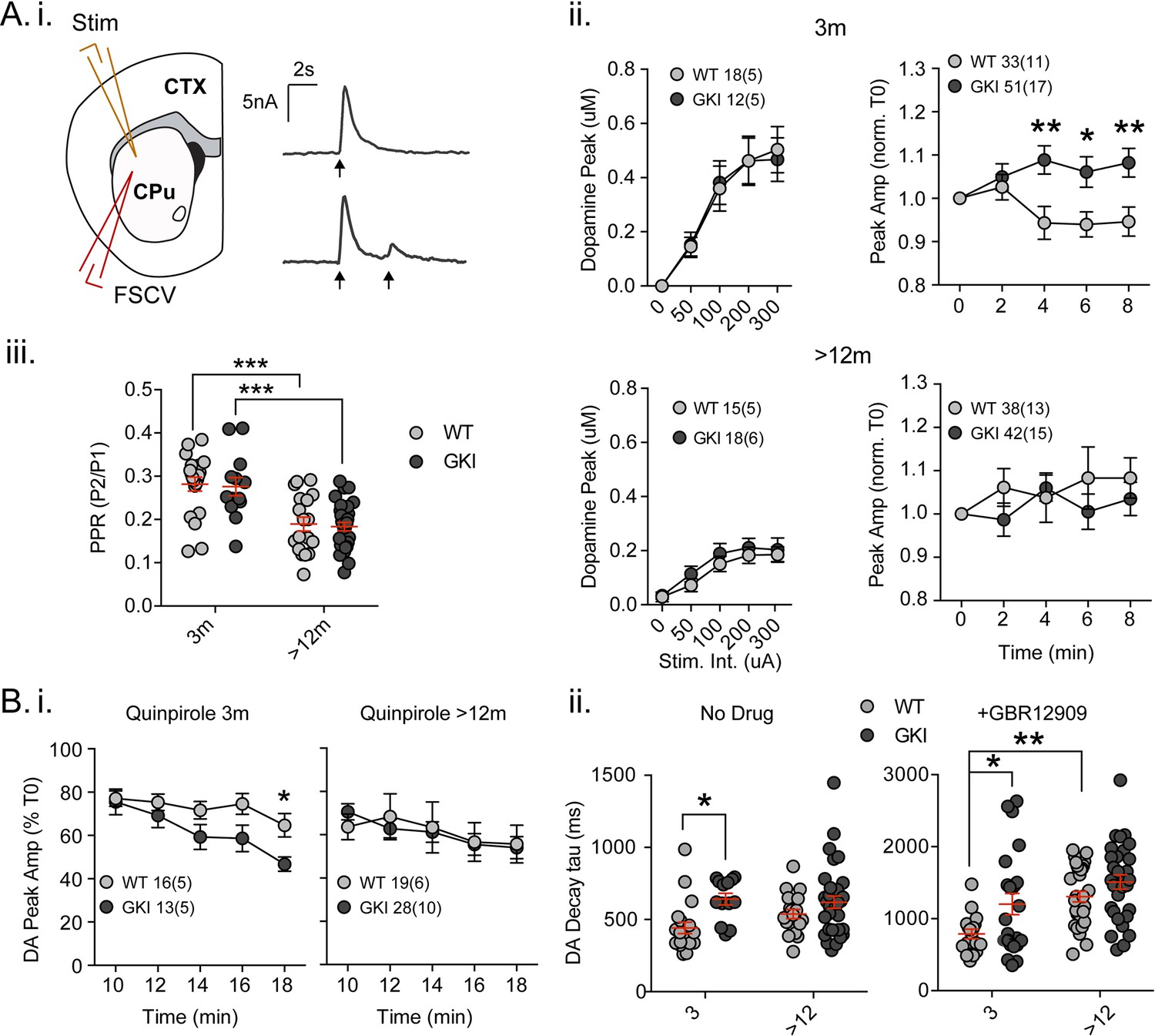
Increased nigrostriatal dopamine transmission in young GKI slices is lost with age, whereas early increases in dopamine extracellular lifetime are maintained in old GKI slices, and matched by age-related increases in WT slices.
Fast-scan cyclic voltammetry (FSCV) was conducted in the dorsolateral striatum of acute slices prepared from WT and GKI mice. (A) Dopamine release and reuptake transients were evoked by single or paired (4 s IPI) local stimuli (i). There was no significant genotype effect (see text) on peak dopamine release over a range of stimulus intensities in slices from 3 month or > 12 month old mice (ii). Upon repeated stimulation, there were significant (post-test *p=0.05, **p=0.01), elevations in dopamine peak release in GKI mice, relative to WT controls at 3 months (ii). In WT and GKI slices from > 12 month mice, peak dopamine release was more variable, there was no modulation induced by repeated stimulation, as in younger slices, nor were there genotype differences. There was a significant age-dependent increase in D2 autoreceptor mediated paired-pulse depression (iii; 3 month n = 20(5) and 13(5),>12 month n = 18(6) and 30(11) for WT and GKI, respectively, post-test ***p=0.001), reflected by reduced paired-pule ratios (PPR), but there was no main effect of genotype (p=0.7) or genotype-age interaction (p=0.99). (B) The D2 agonist quinpirole (10 uM) equally suppressed peak evoked dopamine release in WT and GKI slices from 3 month mice after a 10 min wash-in; however, continued exposure revealed a trend toward a main effect of genotype (p=0.07) on repeated release, and a significant genotype-time interaction (p=0.039, post-test *p=0.049 at 18 min post application). This effect was not observed in > 12 month slices (i). Evoked dopamine transient decay time revealed a significant main genotype effect (ii. No drug, 3 month n = 20(5) and 13(5),>12 month n = 19(6) and 33(11) for WT and GKI, respectively p=0.0025) due to significantly longer decay times (slower transients) in 3 month GKI slices (post-test *p=0.027). In > 12 month slices, WT decay times had increased to a value similar to 3 month GKI (ii. No drug). The presence of the presynaptic dopamine transporter (DAT) blocker GBR (10 uM, ii. + GBR12909) increased all transient decay times, as expected. There were significant main effects of age and genotype (3 month n = 17(6) and 23(8),>12 month n = 31(9) and 30(11), p=0.0002 and 0.0043 for WT and GKI, respectively) upon transient lifetime; in the absence of dopamine re-uptake, 3 GKI transients were still significantly slower than WT controls (post-test *p=0.014), and significant age-dependent increases in dopamine decay times were observed in WT slices (post-test *p=0.0024).
Figure 3—figure supplement 1
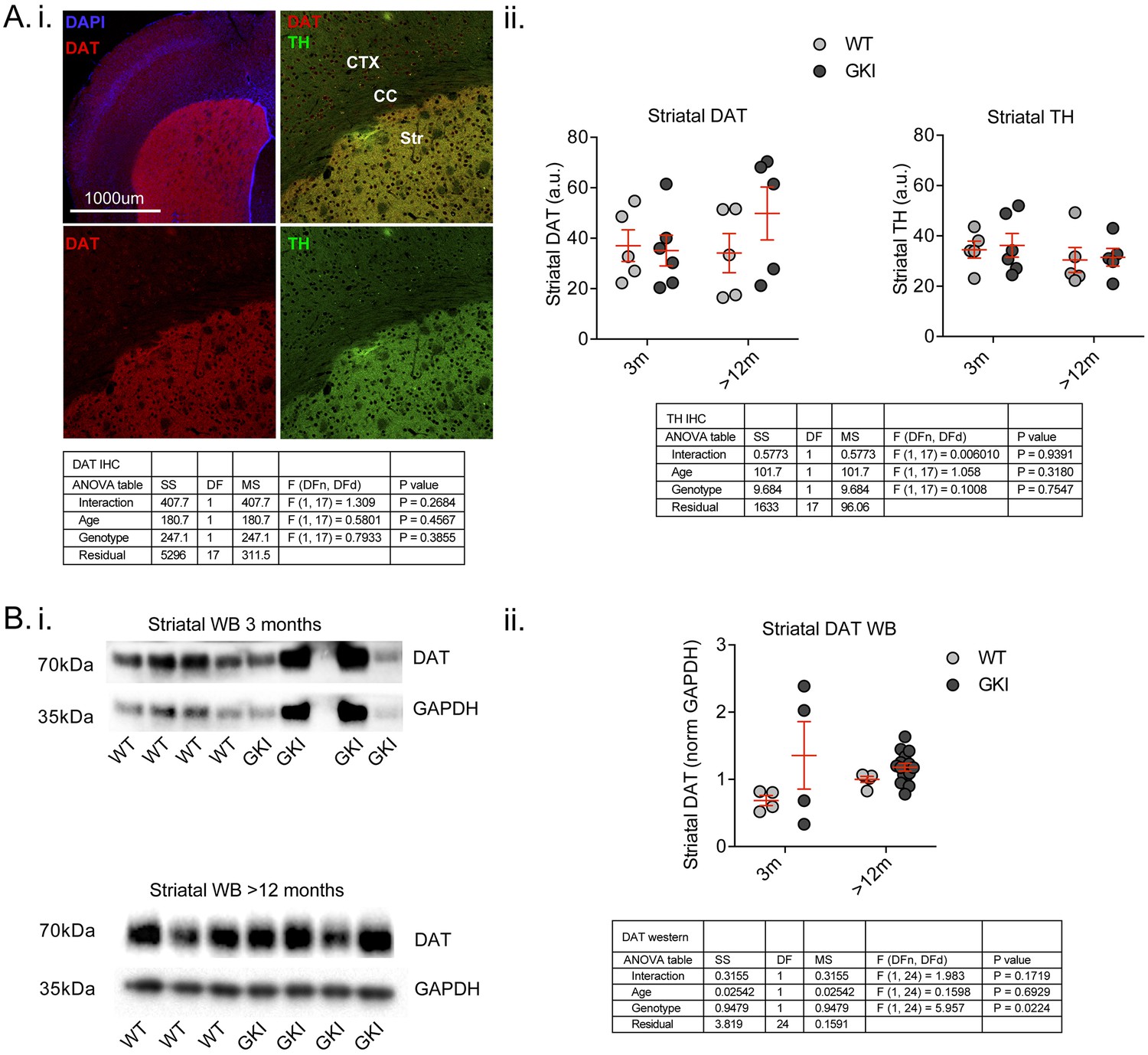
Early alterations in GKI dopamine release are not associated with reductions in nigrostriatal dopamine markers and persist despite genotype-dependent increases in DAT protein levels by western blot.
(A). Staining for presynaptic dopamine markers DAT and tyrosine hydroxylase (TH; i) showed highly specific enrichment in the striatum; however, there were no age or genotype differences (ii). Although variable, the data suggest there is no loss of either terminal marker in GKI striatum. (B) Striatal DAT levels were also assessed by western blot (i); although variable results in young GKI mice suggest no loss of DAT. ii) A significant main genotype effect (due to increased DAT levels in aged GKI mice) revealed increases in striatal DAT levels in the presence of endogenous G2019S. 2-way values are shown.
Additional files
-
Transparent reporting form
- https://doi.org/10.7554/eLife.28377.014
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Initial elevations in glutamate and dopamine neurotransmission decline with age, as does exploratory behavior, in LRRK2 G2019S knock-in mice
eLife 6:e28377.
https://doi.org/10.7554/eLife.28377




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}