Lateral interactions between protofilaments of the bacterial tubulin homolog FtsZ are essential for cell division
- Zhejiang University, China
- Zheijiang University, China
- Peking University, China
- Stanford University, United States
- Stanford University School of Medicine, United States
- Chan Zuckerberg Biohub, United States
Figures
Figure 1 with 1 supplement
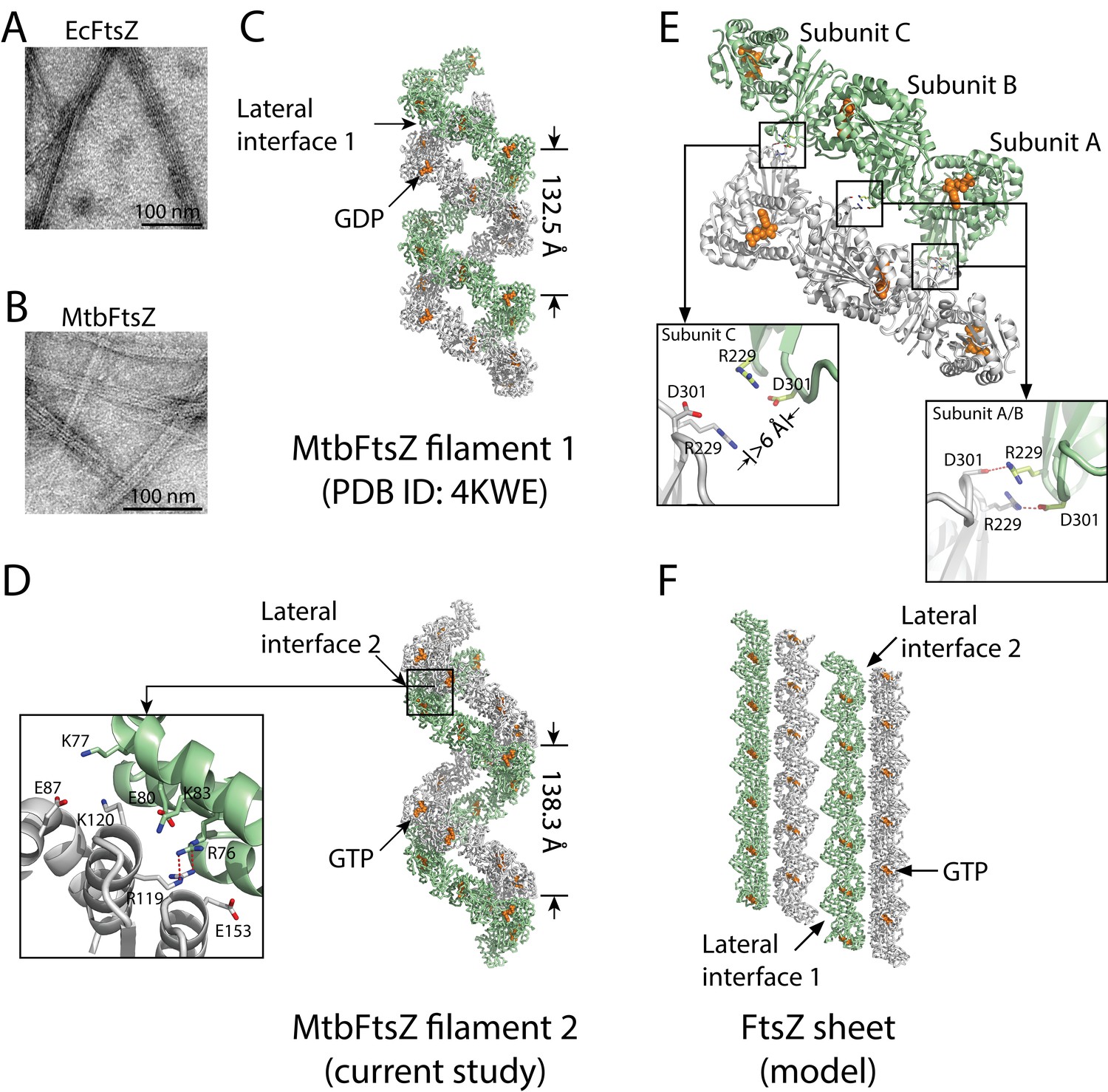
Structures of double-stranded MtbFtsZ-GDP and MtbFtsZ -GTP protofilaments reveal lateral contacts across FtsZ protofilaments.
(A, B) Electron micrographs of protofilament bundles of EcFtsZ-GTP (A) and MtbFtsZ-GTP (B). Both were polymerized with the addition of 0.6 mg/mL DEAE-Dextran, and in the presence of 2 mM GTP. (C, D) Cartoon representations of double-stranded MtbFtsZ-GDP (C; PDB ID: 4KWE) and MtbFtsZ-GTP (D; this study) protofilaments containing a total of 24 subunits. The helices have a pitch of 132.5 Å for MtbFtsZ-GDP (C) and 138.3 Å for MtbFtsZ-GTP (D) protofilaments. Each structure reveals unique lateral interactions across the protofilaments. Inset: atomic details of the lateral interface of the double-stranded MtbFtsZ-GTP protofilaments. (E) Molecular details of the lateral interface of the double-stranded MtbFtsZ-GDP protofilaments shown in (C). Inset: atomic details of the lateral interactions. (F) A structural model for sheet-like bundles of FtsZ protofilaments. Ribbon representation of four straight FtsZ-GTP protofilaments (each containing six subunits, arranged in an antiparallel fashion).
Figure 1—figure supplement 1

Multiple sequence alignment of FtsZ and secondary structure elements.
Amino acid sequence alignment of FtsZ from Mycobacterium tuberculosis (MtbFtsZ), Staphylococcus aureus (SaFtsZ), Escherichia coli (EcFtsZ), Methanococcus jannaschii (MjFtsZ), Aquifex aeolicus (AaFtsZ), Pseudomonas aeruginosa (PaFtsZ), and Bacillus subtilis (BsFtsZ). The secondary structures of α-helices, β-strands, and loops in MtbFtsZ are shown as cylinders, arrows, and lines, respectively. Residues from the observed lateral interfaces in EcFtsZ are highlighted in red (interface 1) and green (interface 2). These residues were subjected to mutagenesis in EcFtsZ in the present study.
Figure 2
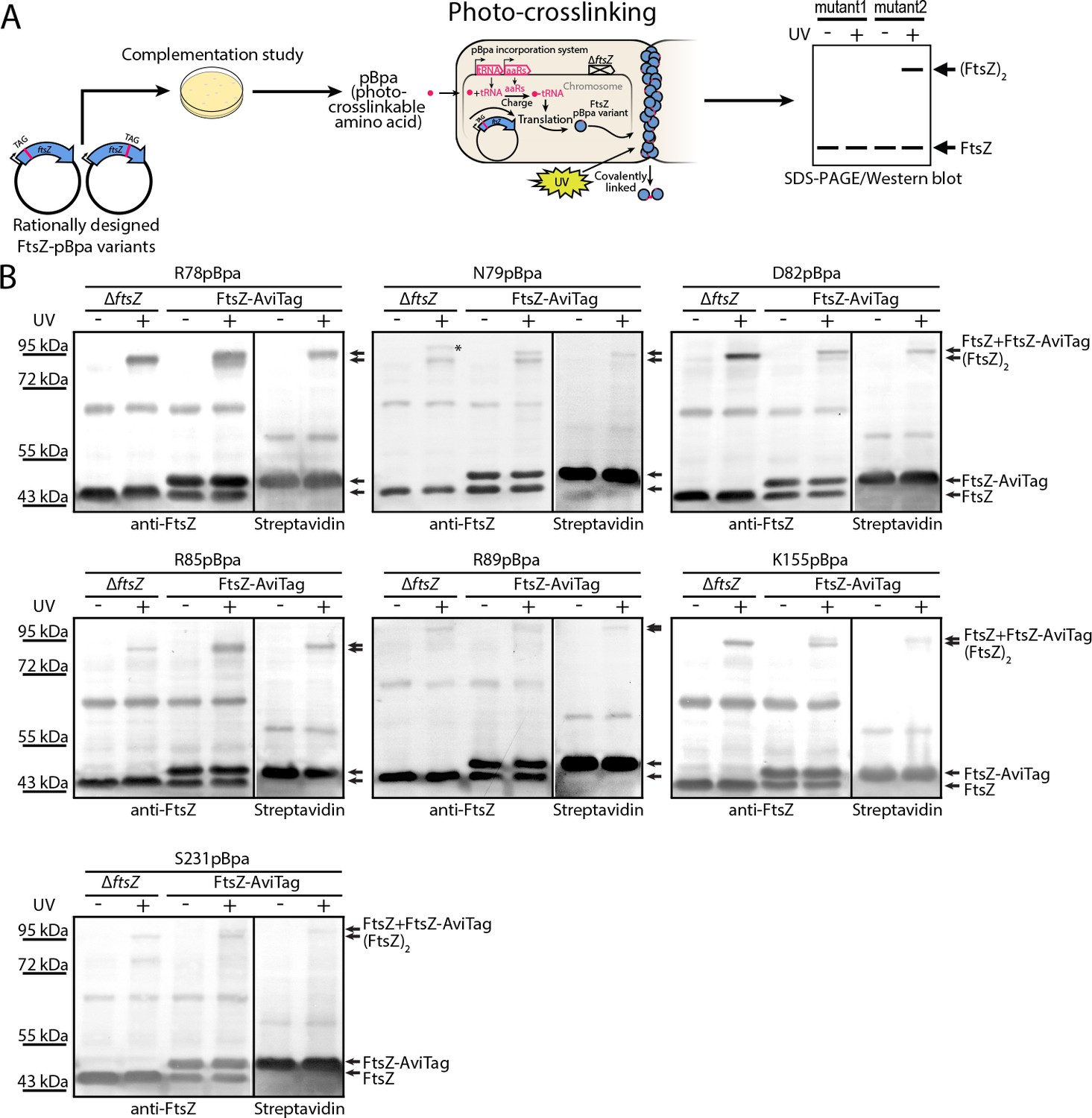
In vivo photocrosslinking characterization of EcFtsZ variants in which pBpa was introduced at specific sites validates inter-protofilament lateral interactions.
(A) Schematic illustrating the rationale of in vivo photocrosslinking analysis via incorporation of the unnatural amino acid pBpa at specific residue positions of FtsZ. (B) Results of blotting analyses of photocrosslinked products with lysates of E. coli cells (LY928-ΔftsZ or LY928-FtsZ-AviTag) expressing the indicated pBpa variant of FtsZ, using antibodies against EcFtsZ (left) or using alkaline phosphatase-conjugated streptavidin (right). Indicated on the right of each gel are the positions of the non-photocrosslinked monomers and the photocrosslinked dimers of the pBpa variant of FtsZ with or without the AviTag. The asterisk indicates the position of a crosslinked product of FtsZ and another as yet unidentified protein.
Figure 3

In vivo photocrosslinking characterization of randomly generated EcFtsZ pBpa variants.
(A) Schematic illustrating the random in vivo photocrosslinking screening strategy for unbiased identification of amino acid positions mediating subunit interactions. (B) Immunoblotting analysis of the photocrosslinked products in lysates of E. coli LY928-ΔftsZ cells that were transformed with the library of plasmids within which the in-frame TAG amber codon was randomly inserted throughout the ftsZ gene, as probed with antibodies against EcFtsZ.
Figure 4 with 2 supplements
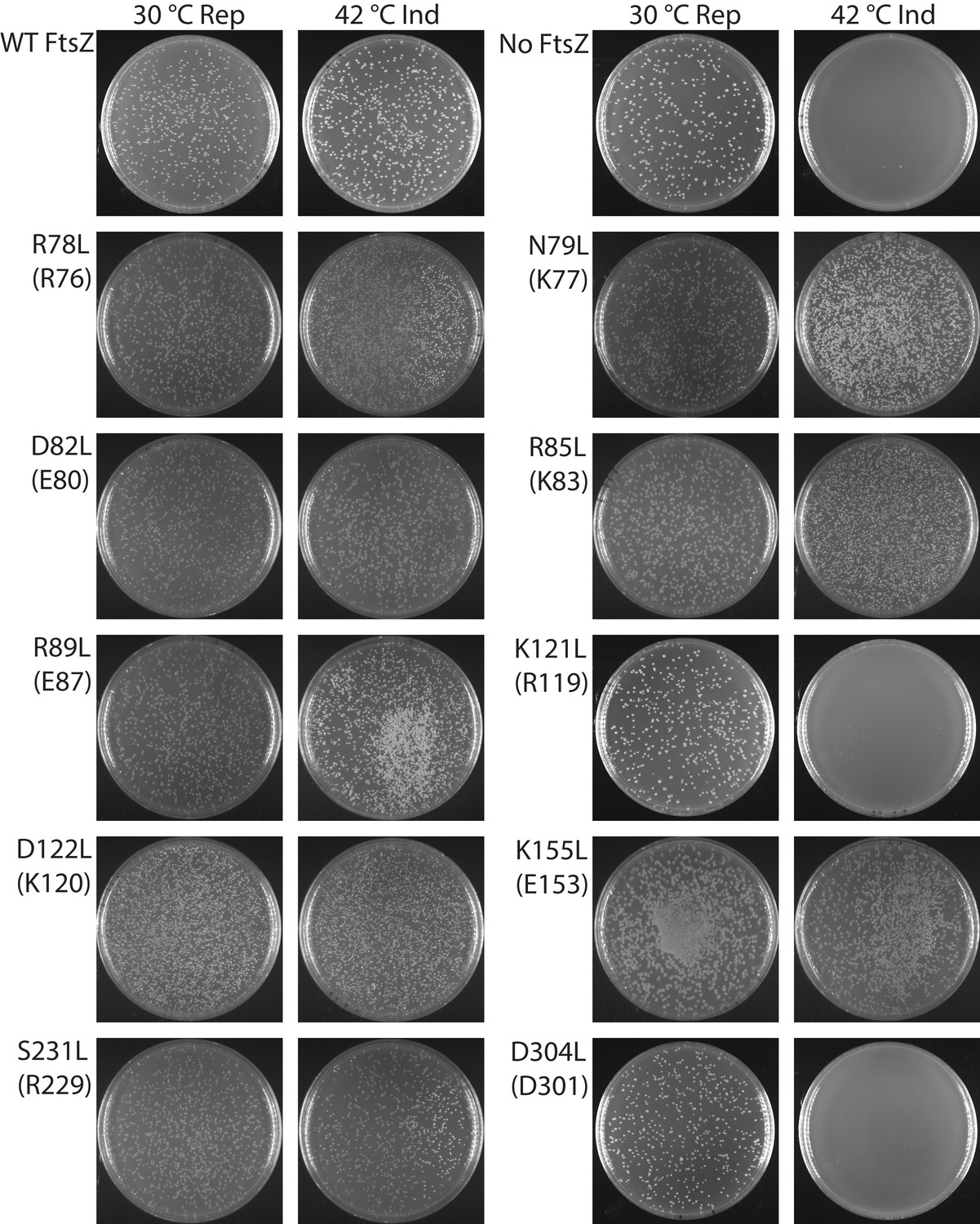
Complementation characterization of EcFtsZ mutants at inter-protofilament lateral interfaces.
Ten EcFtsZ mutants were selected based on our crystallographic observations. Mutations were introduced by altering hydrophilic residues to hydrophobic leucine. The division phenotype was characterized using a ΔftsZ strain expressing the EcFtsZ mutants, with conditional expression of wild-type FtsZ from a plasmid at 30°C but not 42°C. ‘Rep’ and ‘Ind’ indicate repression and induction media, respectively. For each mutant, the complementation assay was repeated three times.
Figure 4—figure supplement 1
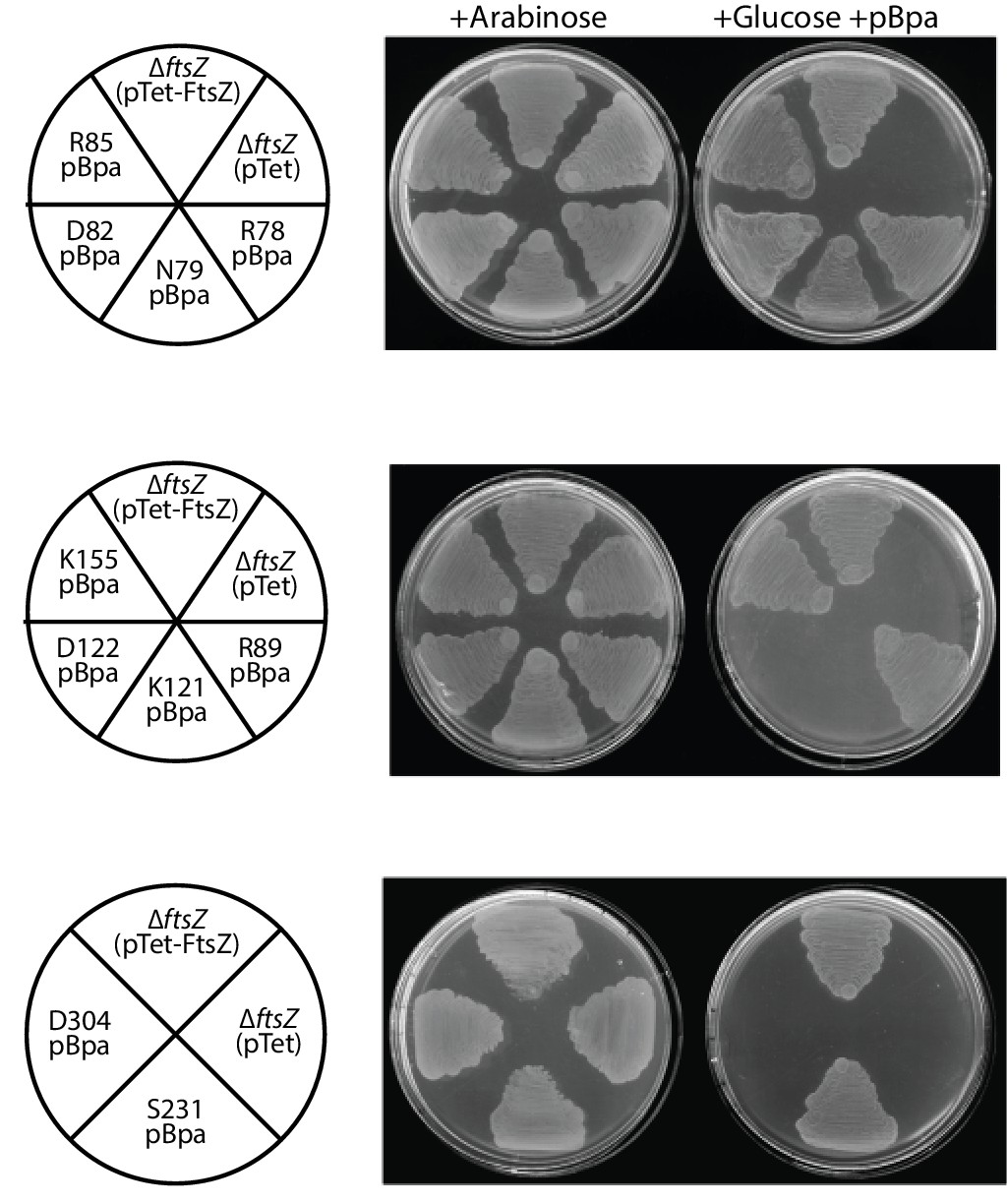
Complementation characterization of pBpa-incorporated EcFtsZ variants.
Ten replacements of EcFtsZ were selected based on our crystallographic observations. pBpa-incorporated variants were created by altering the corresponding sense codon to an amber TAG stop codon. The FtsZ expression plasmid pTet-FtsZ carrying an in-frame amber mutation was used to transform E. coli LY928-ΔftsZ (pJSB100) cells. Cell division, and thus cell growth, could only occur when an FtsZ variant was functional and in the addition of glucose and pBpa.
Figure 4—figure supplement 2

Immunoblotting analysis of three pBpa variants of EcFtsZ that failed to complement the ftsZ conditional-null strain shows absence of crosslinked dimers.
The variants were expressed in LY928-FtsZ-AviTag cells and crosslinked by UV irradiation. Lysate from cells expressing the R78pBpa variant of FtsZ was analyzed as a positive control for photocrosslinked FtsZ dimers.
Figure 5

Complementation characterization of EcFtsZ mutants.
Mutations were introduced by altering D304 and K121 to different hydrophobic residues. In addition to the D304L mutation, S231 was replaced several other amino acids. The division phenotype was characterized using a ΔftsZ strain with expression of the EcFtsZ mutants, and conditional expression of wild-type FtsZ from a plasmid at 30°C but not 42°C. ‘Rep’ and ‘Ind’ indicate repression and induction media, respectively. For each mutant, the complementation assay was repeated three times.
Figure 6
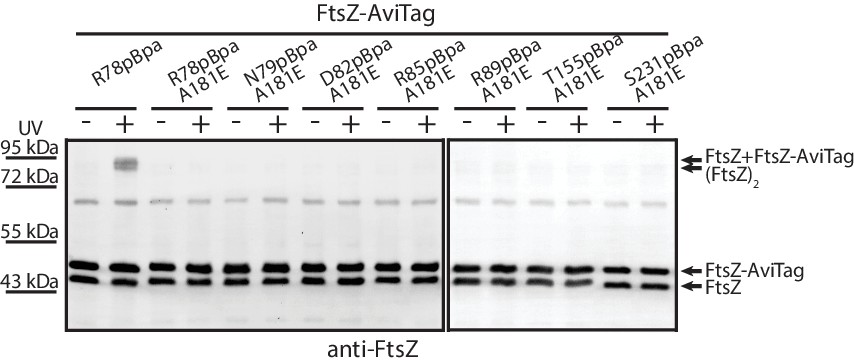
Protofilament formation is required for FtsZ lateral interactions to occur.
Streptavidin blotting analyses of photocrosslinked products of EcFtsZ variants in which pBpa was incorporated at the lateral interface in addition to replacement of alanine 181 with glutamate, which is known to disrupt protofilament formation. The R78pBpa variant (with no disrupting replacement at position 181) was analyzed as a positive control.
Figure 7 with 2 supplements
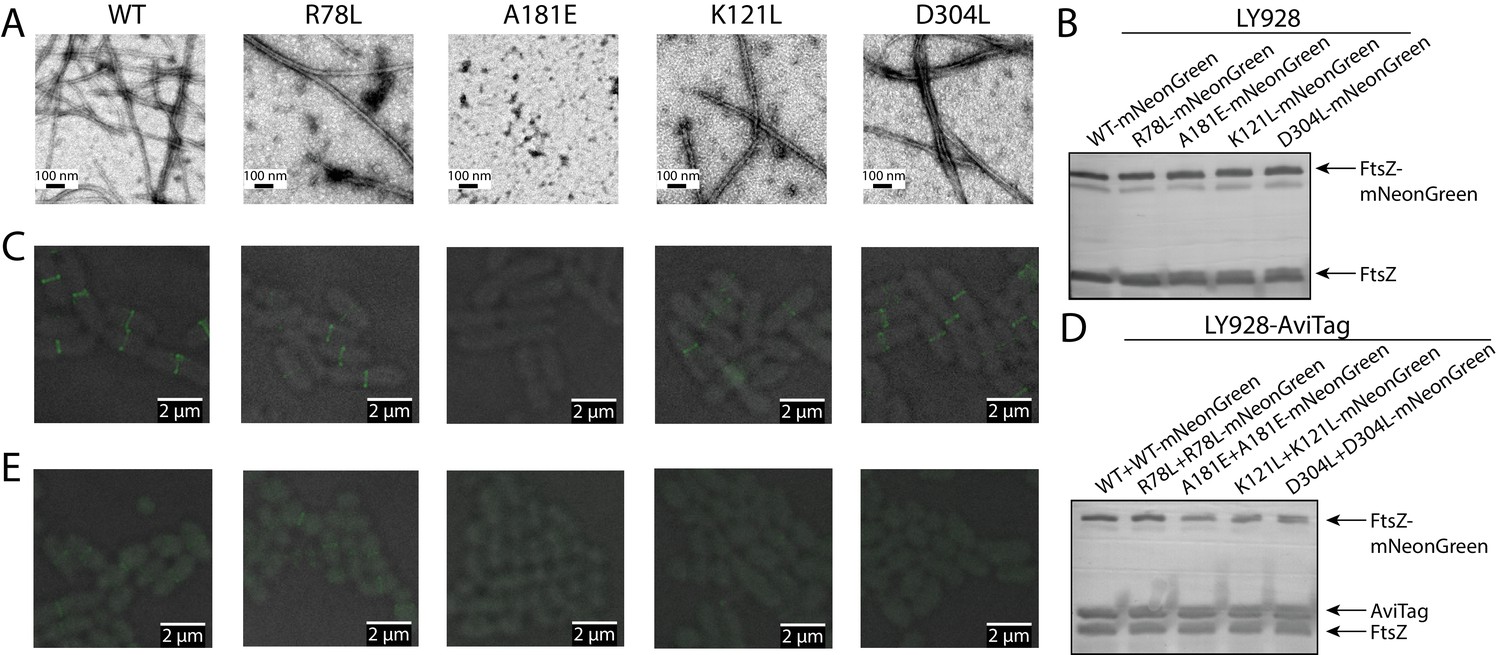
Disruption of the integration of lateral interaction-impaired protofilaments into the Z-ring.
(A) Electron microscopy analysis of GTP-dependent polymerization (with the addition of 0.6 mg/ml DEAE-Dextran) of wild-type or mutated EcFtsZ in which all mutants except A181E form protofilaments similar to wild-type. (B) Western blot analysis of levels of mNeonGreen fusions to wild-type EcFtsZ and mutants. The cellular proportions of mutant FtsZ were ~40%. (C) Fluorescence microscopy demonstrates that low-level expression of laterally disruptive EcFtsZ mutants fused to mNeonGreen does not affect protofilament integration into the Z-ring. (D) Western blot analysis of levels of mNeonGreen fusions to wild-type EcFtsZ and mutants, as well as the AviTagged version. The cellular proportions of mutant FtsZ (mNeonGreen-tagged and untagged) were ~60%. (E) Dominant expression of disruptive EcFtsZ mutants fused to mNeonGreen hinders protofilament integration into the Z-ring, unlike wild-type or the non-disruptive interfacial mutant EcFtsZ R78L.
Figure 7—figure supplement 1
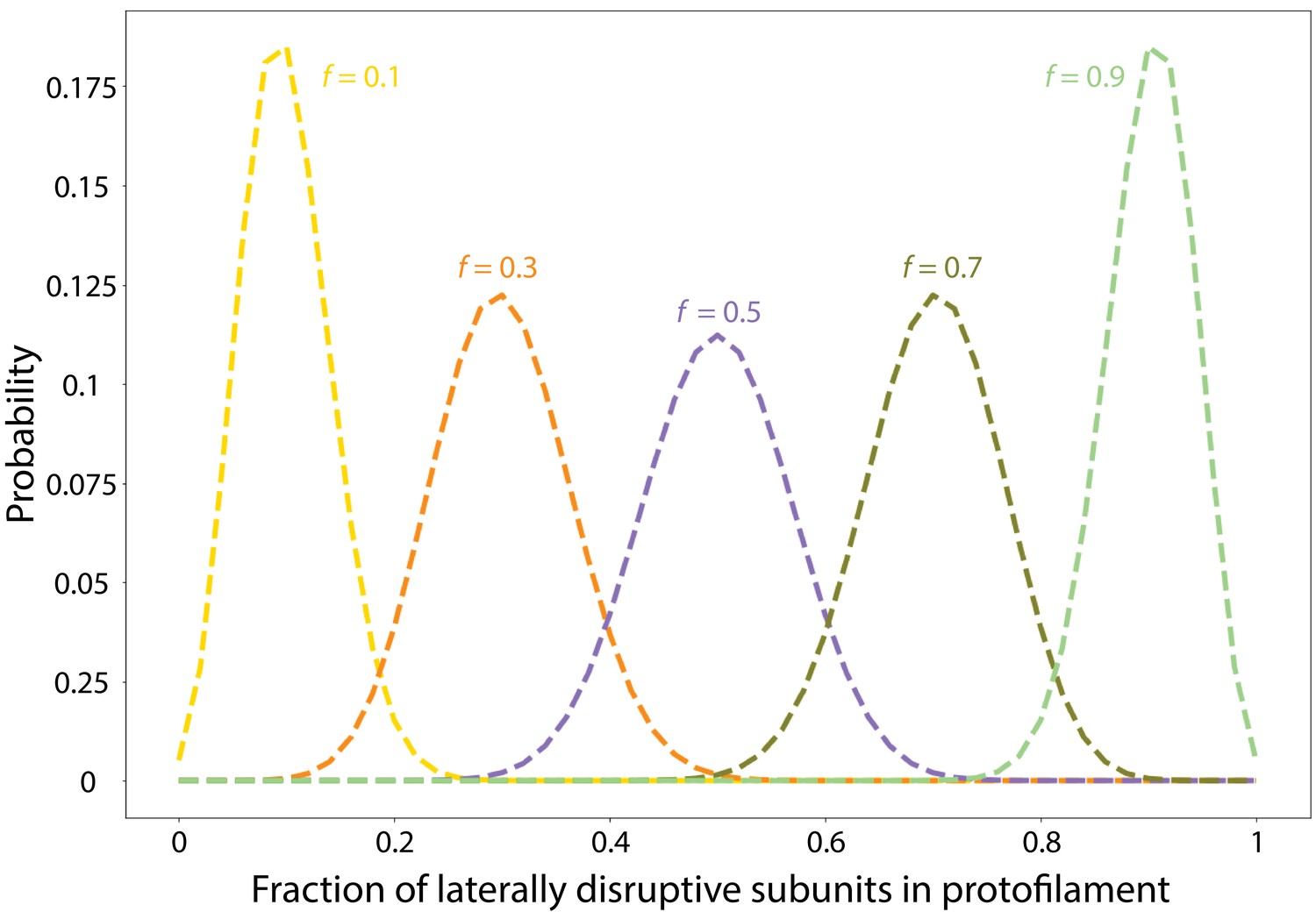
Laterally disruptive FtsZ subunits incorporate into FtsZ protofilaments following a binomial distribution.
Shown are the results of a model in which we assume that all FtsZ protofilaments are 50 subunits long and compute the fraction X of each protofilament that is composed of laterally disruptive subunits, as a function of the overall proportion f of laterally disruptive subunits in a cell. The probability density function is the Binomial distribution .
Figure 7—figure supplement 2
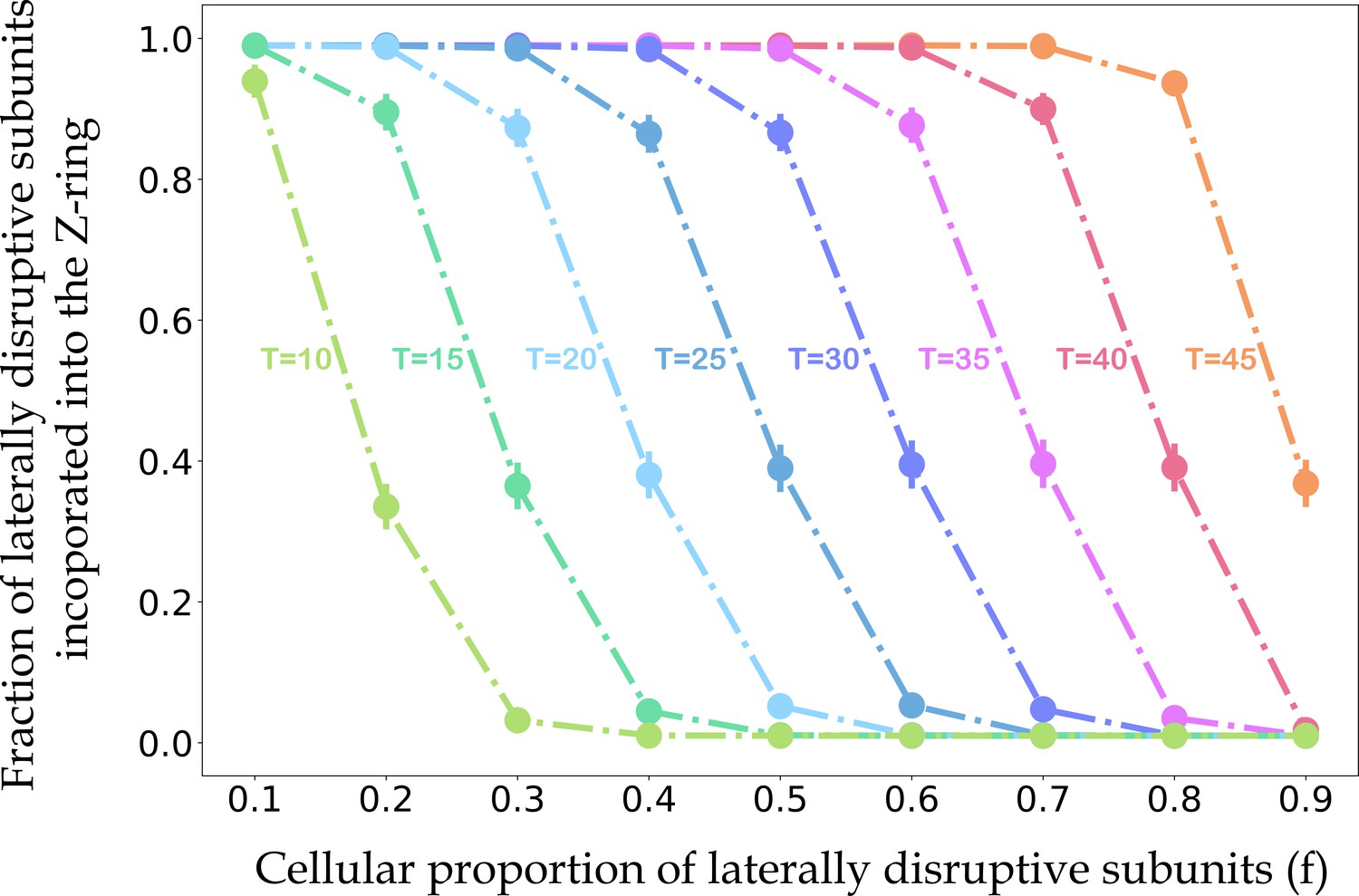
The relationships between the fraction of laterally disruptive subunits incorporated into the Z-ring, the cellular proportion of disruptive subunits, and the maximum number of disruptive subunits that still allow for efficient Z-ring lateral association.
For each value of the cellular proportion of disruptive subunits (f) and threshold for efficient Z-ring incorporation (T), we performed 10,000 independent simulations. In each simulation, we generated a vector X = (x1, x2,. .., x200), where xi represents the number of laterally disruptive subunits in the ith protofilament, selected from the Binomial distribution with mean 50f (Figure 7—figure supplement 1). For a protofilament with more or less laterally disruptive subunits than the threshold T, we set the probability of Z-ring incorporation to 0.01 or 0.99, respectively. The Boolean vector V represents the incorporation state of each protofilament. We then calculated the percentage of laterally disruptive subunits incorporated into the Z-ring as . Circles indicate the mean percentages of laterally disruptive subunits in the Z-ring across simulations with a given value of f and T, and solid vertical lines represent the standard deviations. As the total proportion of laterally disruptive subunits increases, the proportion of laterally disruptive subunits incorporated into the Z-ring decreases, consistent with our experimental results showing a faint or non-existent Z-ring with dominant expression of laterally disruptive mutants (Figure 7E).
Tables
Table 1
X-ray data and refinement statistics.
https://doi.org/10.7554/eLife.35578.005| Data set | |
|---|---|
| Space group Unit cell Resolution (Å) Measured reflections Unique reflections Redundancy Completeness (%, highest shell) Mean I/σI (highest shell) Rsym (%, highest shell) | P6522 a = 100.5 Å, c = 138.3 Å 2.7 167,791 11,805 14.2 99.3 (99.5) 44.1 (1.6) 10.7 (100) |
| Refinement | |
| Resolution (Å) Number of reflections |F| > 0 σF R-factor/R-free (%) Number of protein atoms Number of GTP molecules Number of water molecules rmsd bond lengths (Å) rmsd bond angles (°) | 2.7 10,593 21.1/25.3 2203 1 0 0.008 0.97 |
Table 2
Complementation effects of inter-protofilament interface mutants.
https://doi.org/10.7554/eLife.35578.011| Corresponding amino acid in MtbFtsZ | Complementation on plates | Complementation in liquid culture | |
|---|---|---|---|
| Wildtype | Yes | Yes | |
| R78L | R76 | Yes | Yes |
| N79L | K77 | Yes | Yes |
| D82L | E80 | Yes | Yes |
| R85L | K83 | Yes | Yes |
| R89L | E87 | Yes | Yes |
| K121L | R119 | No | No |
| D122L | K120 | Yes | Yes |
| K155L | E153 | Yes | Yes |
| S231L | R229 | Yes | Yes |
| D304L | D301 | No | No |
| N78pBpa | Yes | Yes | |
| N79pBpa | Yes | Yes | |
| D82pBpa | Yes | Yes | |
| R85pBpa | Yes | Yes | |
| R89pBpa | Yes | Yes | |
| K121pBpa | No | No | |
| D122pBpa | No | No | |
| K155pBpa | Yes | Yes | |
| S231pBpa | Yes | Yes | |
| D304pBpa | No | No |
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene (Escherichia coli) | FtsZ | NA | Uniprot ID: P0A9A6 | |
| Gene (Mycobacterium tuberculosis) | FtsZ | NA | Uniprot ID: P9WN95 | |
| Gene (Branchiostoma lanceolatum) | mNeonGreen | PMID:23524392 | Uniprot ID: A0A1S4NYF2 | |
| Stain, strain background (Escherichia coli) | W3110 ftsZ::kan recA56 ΔftsZ | PMID:2045370 | JKD7-1/pKD3, ftsZ conditional null strain with pKD3 rescue plasmid | |
| Stain, strain background (Escherichia coli) | BW25113 ΔinsH11:: pBpa-tRNA synthetase /tRNA-pBpa | PMID:27298319 | ||
| Stain, strain background (Escherichia coli) | BW25113 ΔinsH11:: pBpa-tRNA synthetase /tRNA-pBpa ΔftsZ | This paper | ftsZ conditional null strain with pBpa incorporation system | |
| Stain, strain background (Escherichia coli) | BW25113 ΔinsH11::pBpa- tRNA synthetase/tRNA- pBpa ftsZ::ftsZ-AviTag | This paper | The endogenous ftsZ gene was modified to encode FtsZ linked with an AviTag at the C-terminus | |
| Antibody | Rabbit anti-EcFtsZ polyclonal antibody | This paper | The antibody was harvested by immuning rabbit with the purified EcFtsZ protein. Dilution: 1:10000, for western blot | |
| Antibody | Alkaline phosphatase- conjugated streptavidin | Beyotime | A0312 | Dilution: 1:5000, for western blot |
| Recombinant DNA reagent | pJSB100 | PMID:12896999 | pJSB2-FtsZ | |
| Recombinant DNA reagent | pTet-FtsZ | This paper | bla pBR322 P tet::ftsZ, P tet incates Tet-on/Tet-off promoter and the expression of FtsZ in this plasmid via leaky expression | |
| Recombinant DNA reagent | pTac-L3-FtsZ-mNeonGreen | This paper | bla pBR322 P L3::ftsZ-mNeonGreen, P L3 incates a synthetic constitutive promoter selected from Anderson promoter collection | |
| Recombinant DNA reagent | pTac-0.16-FtsZ-TAG- mNeonGreen | This paper | bla pBR322 P0.16::ftsZ-TAG- mNeonGreen, P0.16 indicates a synthetic constitutive promoter selected from Anderson promoter collection and TAG amber condon was inserted into FtsZ-TAG- mNeonGreen fusion protein | |
| Chemical compound, drug | p-benzoyl-L-phenylalanine (pBpa) | Bachem | F-2800 |
Additional files
-
Transparent reporting form
- https://doi.org/10.7554/eLife.35578.017
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Lateral interactions between protofilaments of the bacterial tubulin homolog FtsZ are essential for cell division
eLife 7:e35578.
https://doi.org/10.7554/eLife.35578




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}