TRPV4 is the temperature-sensitive ion channel of human sperm
- University of California, Berkeley, United States
- RWTH Aachen University, Germany
Figures
Figure 1 with 2 supplements
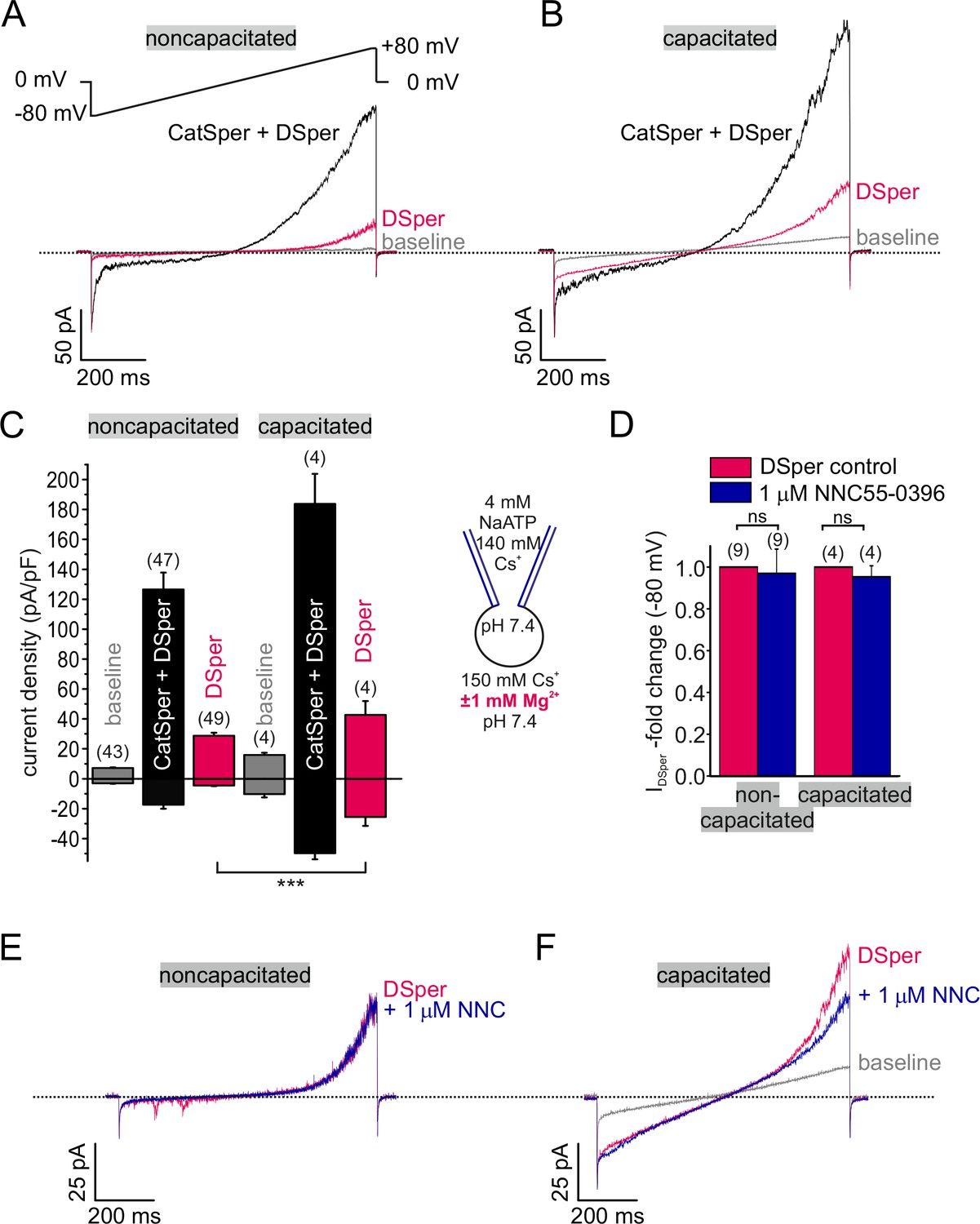
Electrophysiological recordings reveal a novel non-CatSper conductance.
(A-B) Original current traces from representative whole-cell patch-clamp recordings from noncapacitated (A) and capacitated (B) human spermatozoa. Inward- and outward currents were elicited with voltage ramps as depicted in (A). Under divalent free conditions (black traces), typical CatSper monovalent caesium currents can be recorded. In the presence of 1 mM Mg2+ (red traces), an outward rectifying ‘DSper’ current component remains. Hence, the black traces represent a mixture of both CatSper and DSper monovalent Cs+ currents, while the red traces show pure Cs+ currents through DSper. (C) Quantification of current densities for all three conditions in (A-B). DSper currents are potentiated upon capacitation (noncapacitated cells: -4.50 ± 0.41 pA/pF, capacitated cells: -25.58 ± 5.88 pA/pF for inward currents recorded at -80 mV; noncapacitated cells: 28.80 ± 1.93 pA-pF, capacitated cells: 42.67 ± 9.27 pA/pF for outward currents recorded at +80 mV). Data are mean ± S.E.M., with (n) representing the number of individual sperm cells tested. Statistical significance (unpaired t-test) was indicated by: ***p ≤0.0005. Data was collected from 3 human donors, no variations between human donors were noticed. Quantification of normalized DSper inward currents (D) and original current traces (E-F) in presence and absence of the CatSper inhibitor NNC 55-0396 demonstrate the absence of the inhibition.
-
Figure 1—source data 1
The source data for the inward and outward monovalent and divalent DSper current densities.
(A) Inward and Outward monovalent DSper current densities (pA/pF) recorded at -80mV and +80mV, respectively. (B) Divalent CatSper inward current densities (pA/pF).
- https://doi.org/10.7554/eLife.35853.005
Figure 1—figure supplement 1
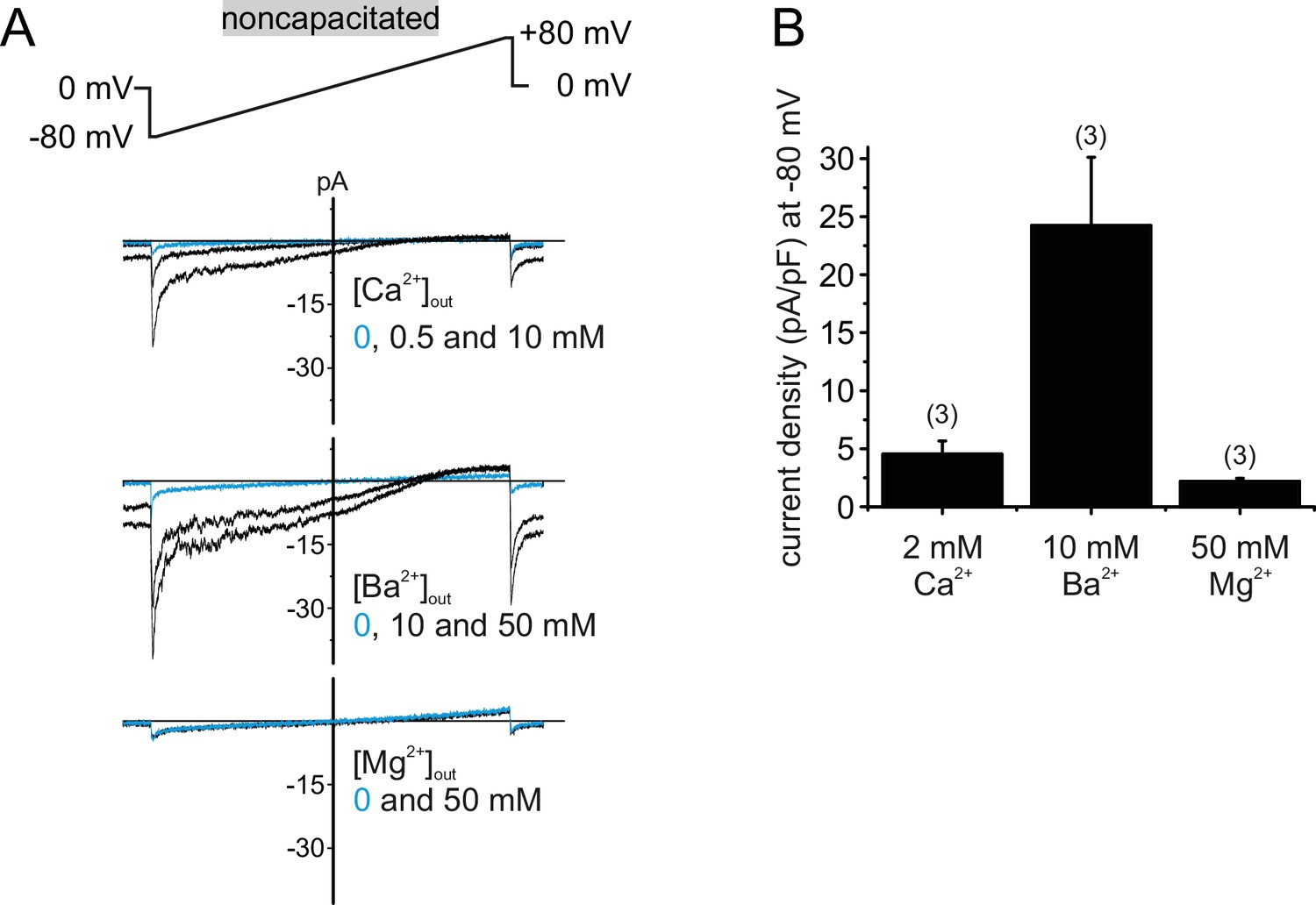
Human CatSper conducts Ca2+ and Ba2+ but not Mg2+.
(A) Original current traces from whole-cell voltage-clamp recordings of noncapacitated human spermatozoa. Inward- and outward currents were elicited with voltage ramps as depicted. Pipette solution was: 140 mM NMDG, 100 mM Hepes, 5 mM EGTA, 5 mM EDTA, 330 mOsmol, pH 7.3, composition of bath solution was: 500 nM progesterone, 100 mM Hepes, 130 mM NMDG, plus X mM Ca2+, Ba2+ or Mg2+ as depicted, 317 mOsmol, pH 7.4. When the major permeable extracellular cation was Ca2+ or Ba2+, negative membrane potentials induced concentration-dependent inward currents. In the presence of Mg2+, CatSper currents remained at baseline level (0 mM), indicating that human CatSper is not permeable for Mg2+. (B) Quantification of current densities (pA/pF) for either Ca2+, Ba2+ or Mg2+ inward currents through CatSper.
Figure 1—figure supplement 2
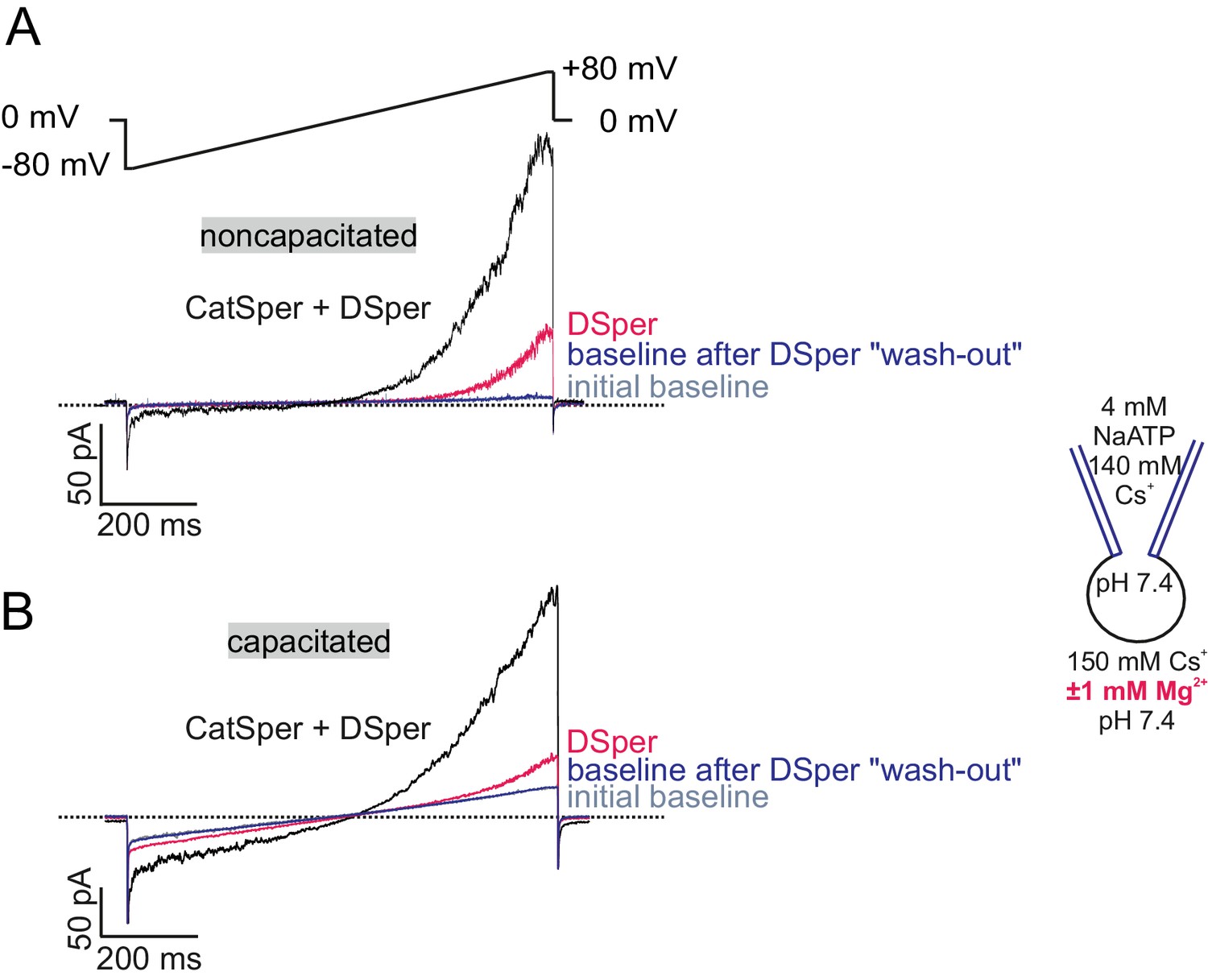
DSper currents are recorded under stable conditions.
(A–B) Original current traces from representative whole-cell patch-clamp recordings of noncapacitated (A) and capacitated (B) human spermatozoa. Inward- and outward currents were elicited with voltage ramps as depicted in (A). Represented are three conditions – baseline (in HS solution), CatSper + DSper currents and isolated DSper currents. Whole-cell currents returned to their initial baseline level after returning to HS solution, indicating that the recorded DSper currents are not a remnant of an increased leak-current.
Figure 2
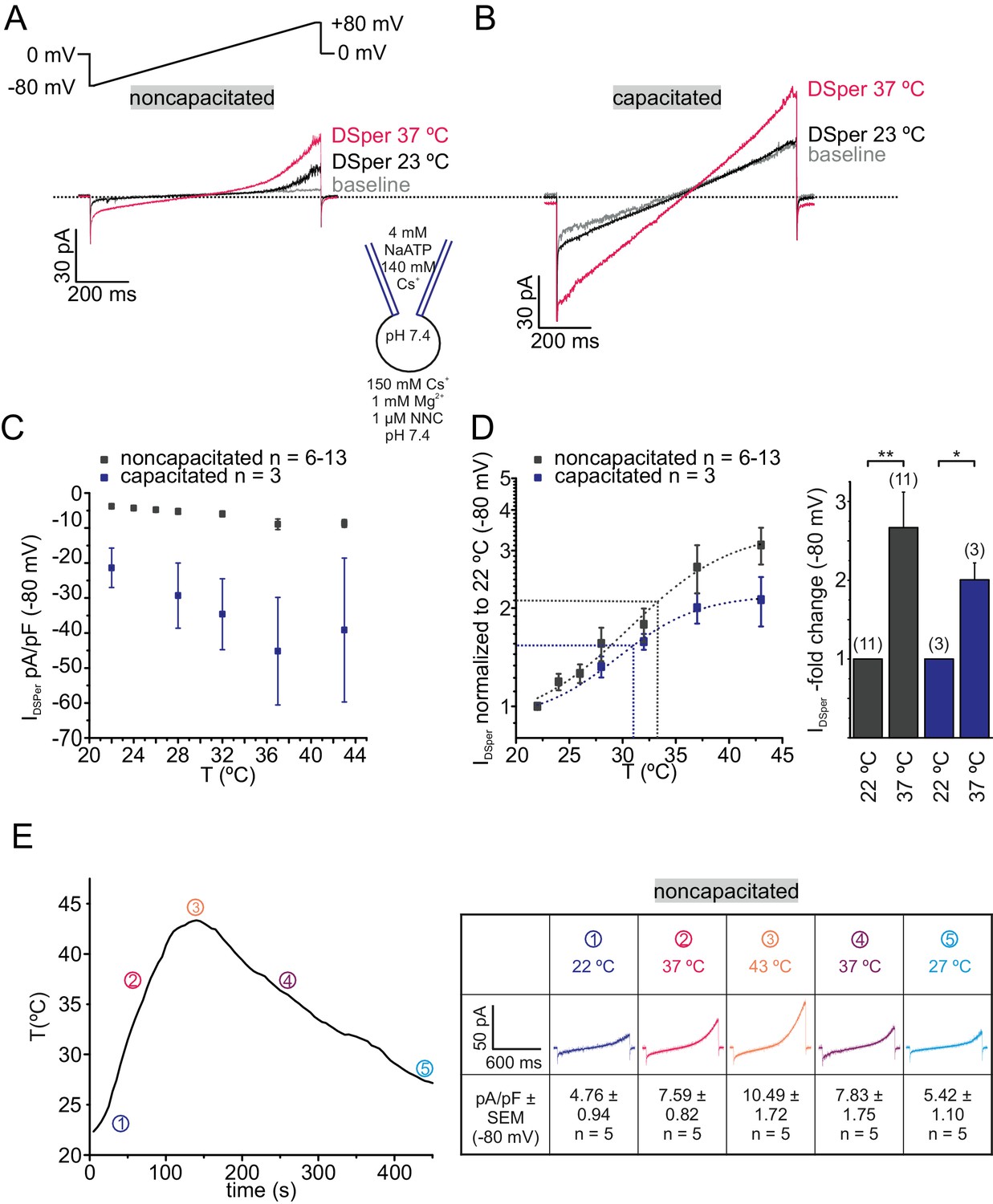
DSper is activated by warm temperatures.
(A–B) Representative current traces from whole-cell patch-clamp recordings from noncapacitated (A) and capacitated (B) human spermatozoa challenged with a rise in temperature from 23°C to 37°C. Both DSper inward- and outward currents are increased at warmer temperatures. (C) Quantification of DSper inward current densities as a function of bath temperature (in °C). Noncapacitated (grey squares) as well as capacitated cells (blue squares) show increased current densities when stimulated with increasing bath temperatures. (D) Data of (C) normalized to room temperature (22°C). Half maximal activation at T1/2 = 34°C (noncapacitated) and T1/2 = 31°C (capacitated) indicated by the dotted lines. The data were fitted with Boltzmann equation to estimate the temperature at which DSper currents have half-maximal activation. Right panel: statistical significance (unpaired t-test) was indicated by: *p≤0.05, **p≤0.005 for capacitated (blue bars) and noncapcitated (black bars) human sperm. (E) The bath temperatures as a function of time and corresponding DSper currents. Inset shows representative traces indicating that the temperature-induced potentiation effect was reversible. Data are mean ± S.E.M., with (n) representing the number of individual sperm cells tested obtained from three human donors.
-
Figure 2—source data 1
DSper inward current densities as a function of the bath temperature.
- https://doi.org/10.7554/eLife.35853.007
Figure 3

DSper conducts sodium ions.
(A) Representative current traces from the whole-cell patch-clamp recordings of noncapacitated human spermatozoa. Inward and outward currents were elicited with the voltage ramps as depicted. To record DSper currents, extracellular Cs+ was substituted with the same concentration of Na+. Representative current traces (B) and quantification of the normalized DSper inward currents (C) before and after stimulation with 1 μM NNC suggest that CatSper channel does not contribute to the recorded sodium inward conductance. (D–E) Representative current traces in (D) and quantification of the inward currents normalized to 22°C (E) at increasing bath temperatures. A similar temperature-induced potentiation effect of DSper sodium inward currents was observed as for caesium currents. Recordings were performed in the presence of NNC to exclude any CatSper contribution. Half maximal activation was achieved at T1/2sodium = 34°C (dotted line). The data were fitted with the Boltzmann equation. Statistical significance (unpaired t-test) is indicated by: **p≤0.005. Data are mean ± S.E.M., with (n) representing the number of individual sperm cells tested obtained from two human donors.
-
Figure 3—source data 1
The source data for the inward sodium currents via DSper.
- https://doi.org/10.7554/eLife.35853.009
Figure 4 with 1 supplement
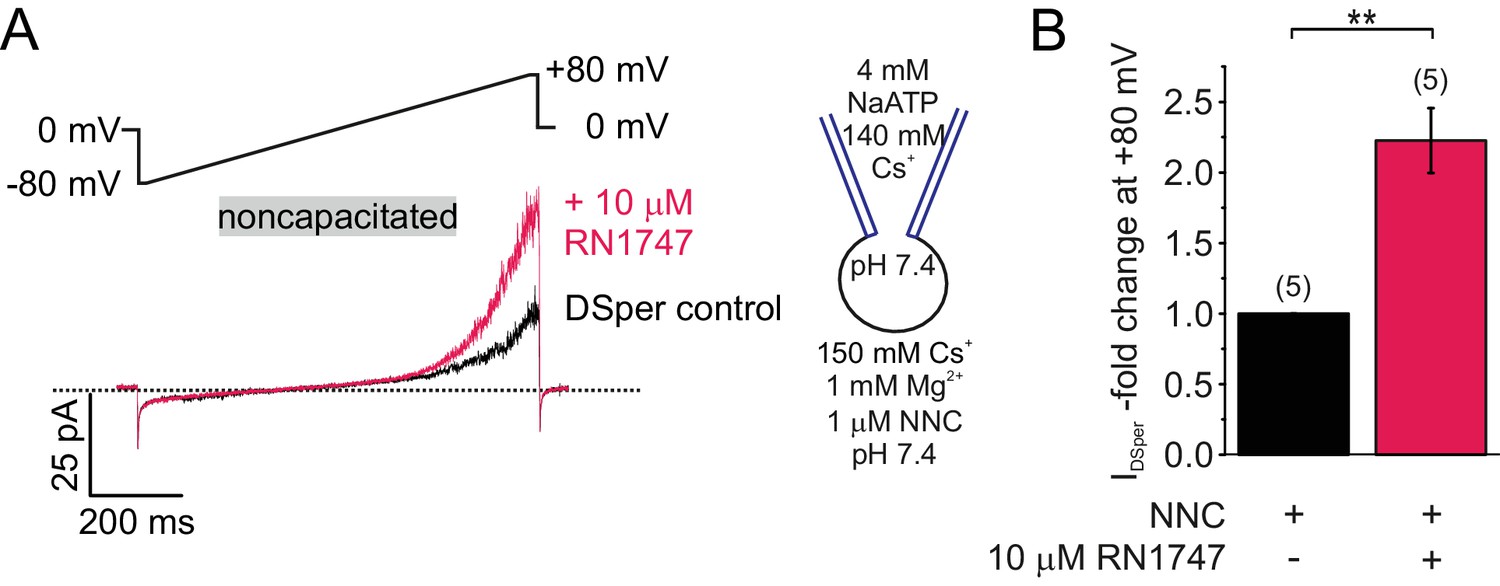
DSper is activated by the TRPV4 agonist RN1747.
(A) Representative whole-cell patch-clamp recordings of noncapacitated human spermatozoa. Inward- and outward currents were elicited with voltage ramps as depicted. DSper monovalent caesium currents (black trace) are increased after stimulation with 10 μM RN1747 (red trace). Both recordings were performed in the presence of 1 μM NNC. (B) Quantification of normalized DSper outward currents under control conditions and after stimulation with RN1747. A significant gain upon stimulation with the TRPV4 agonist (factor 2.22 ± 0.23, **p=0.0007, unpaired t-test, n = 6) is observable. No variation between human donors were noticed.
-
Figure 4—source data 1
The source data for DSper regulation of TRP channel agonists and inhibitors.
(A) DSper inward currents potentiated by RN1747. (B) DSper currents after stimulation with Capsaicin. (C) Calcium imaging (stimulation with 10 μM capsaicin after 20 s). (D) Calcium imaging (stimulation with 500 μM carvacrol after 20 s). (E) Pregnonolone sulfate effect on IDSper.
- https://doi.org/10.7554/eLife.35853.012
Figure 4—figure supplement 1
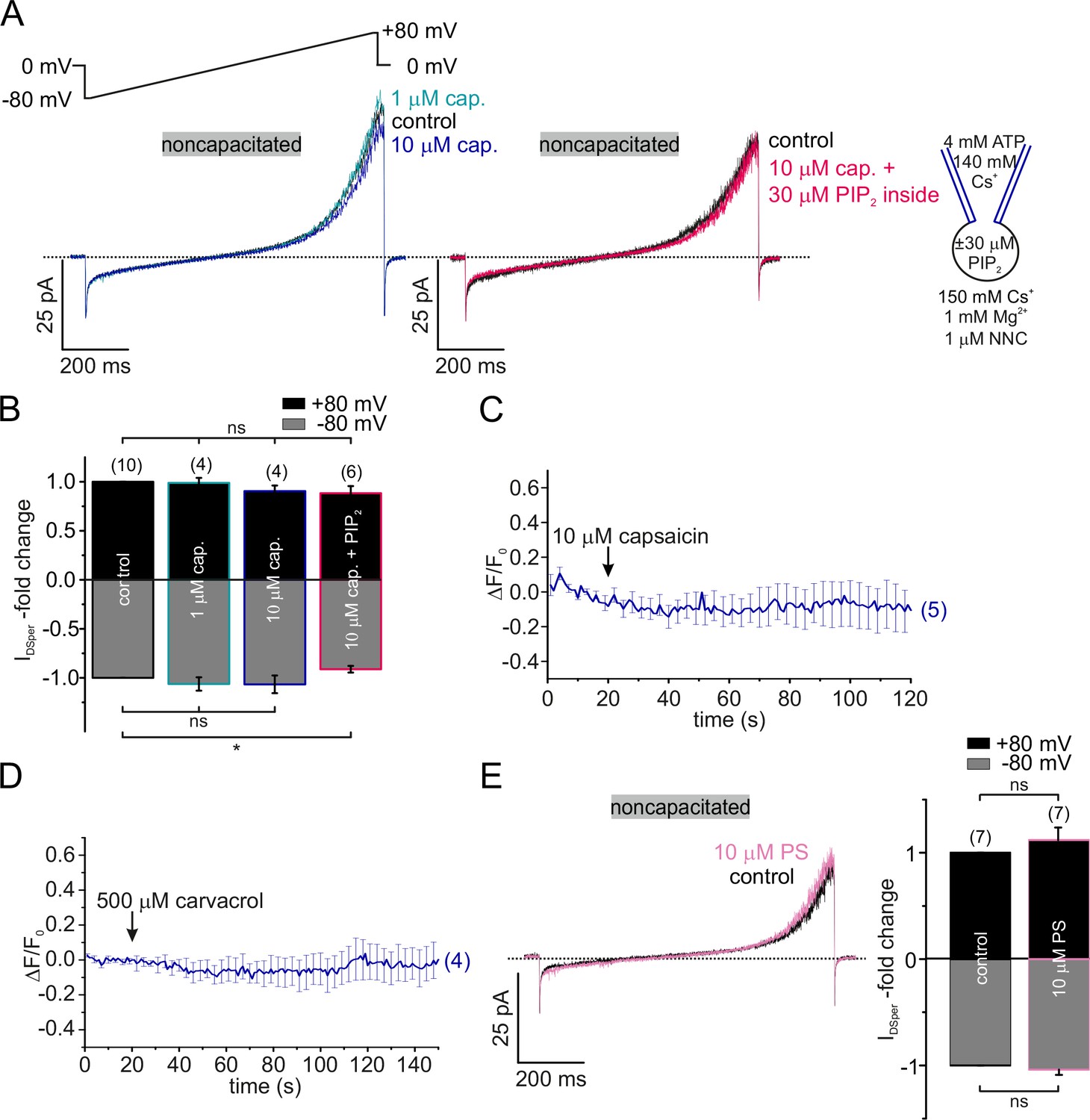
TRPV1, TRPV3 and TRPM3 are not functionally expressed in human spermatozoa.
(A) Original current traces from representative whole-cell patch-clamp recordings of noncapacitated human spermatozoa. Inward-and outward currents were elicited with voltage ramps as depicted. Stimulation with two different concentrations (1 and 10 μM) of the specific TRPV1 agonist capsaicin did not induce any significant effect on DSper inward- or outward currents (left). Addition of 30 μM PIP2 to the pipette solution (right) did not generate a capsaicin-induced potentiation of DSper activity either. Quantification of normalized DSper currents for all four conditions is depicted in (B). (C) Single-cell calcium imaging confirmed electrophysiological findings. Application of 10 μM capsaicin had no effect on cytosolic calcium levels. (D) Single-cell calcium imaging approach did not reveal any notable effect of the TRPV3 specific agonist carvacrol (500 μM). (E) (Left) Original current traces from representative whole-cell patch-clamp recordings of noncapacitated human spermatozoa. Stimulation with 10 μM PS did not have any effect on DSper inward and outward currents as indicated by the pink trace. (Right) Quantification of DSper current amplitudes normalized to control conditions reveals no statistical significance of PS stimulation. Data are mean ± S.E.M., with (n) representing the number of individual sperm cells tested obtained from four donors.
Figure 5 with 2 supplements
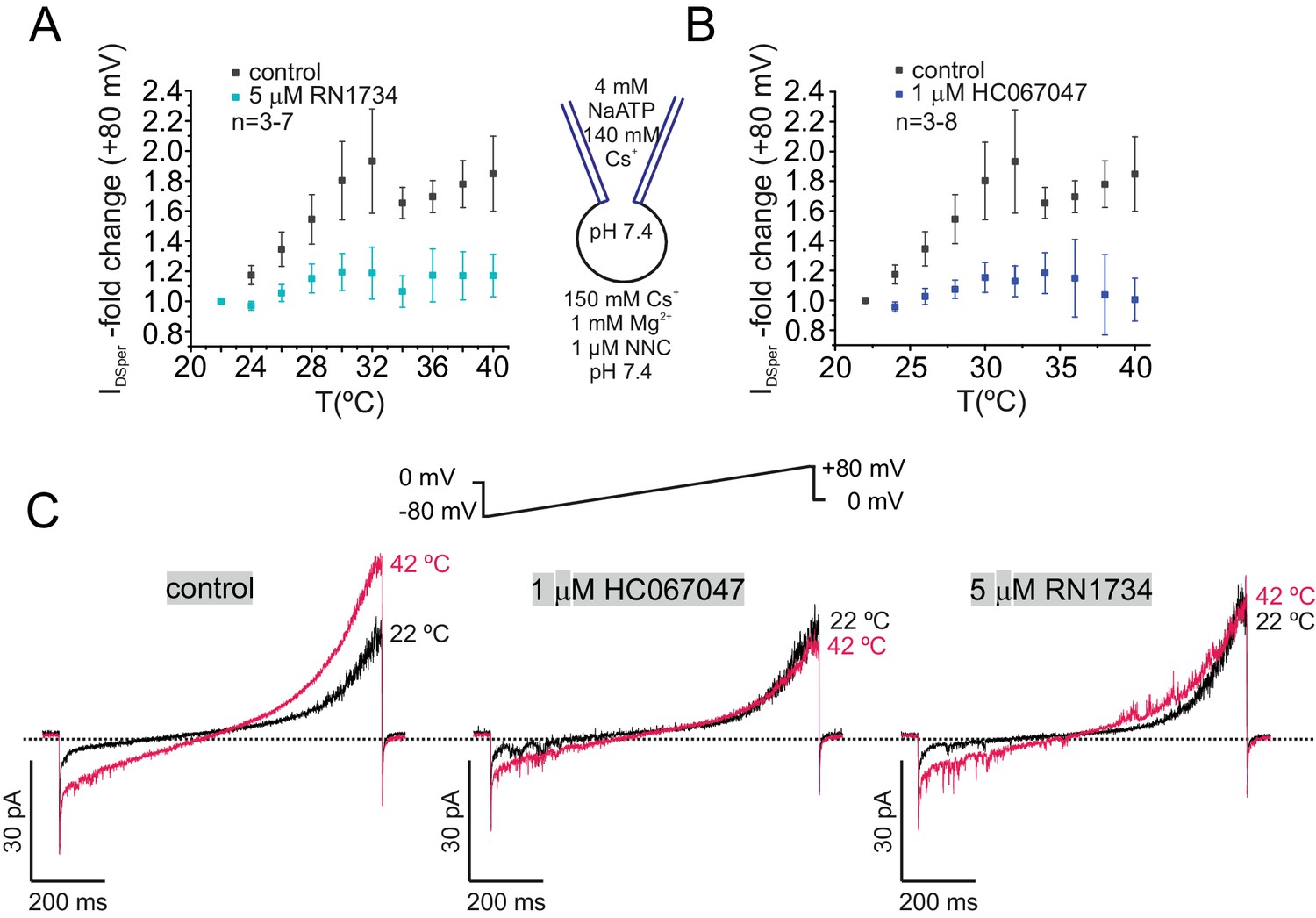
TRPV4 inhibitors RN1734 and HC067047 antagonize human DSper activity.
(A–B) Quantification of normalized DSper outward currents as a function of bath temperature (in °C) in the absence (grey squares) and the presence (turquoise/blue squares) of TRPV4 inhibitors RN1734 (A) and HC067047 (B), respectively. Both inhibitors greatly reduce DSper’s temperature-sensitivity. Representative current traces of whole-cell voltage clamp recordings from noncapacitated spermatozoa for all three conditions are depicted in (C). Representative current traces indicate that a rise in temperature from 22°C to 42°C considerably potentiated DSper inward and outward currents under control conditions but not in presence of the TRPV4 antagonists HC067047 and RN1734. No variations between human donors were noticed.
-
Figure 5—source data 1
The source data for the experiments shown on Figure 5 and its Figure 5—figure supplement 1.
(A) DSper’s temperature-sensitivity in presence of TRPV4 specific inhibitors RN1734 and HC067047. (B) EtOH vehicle control.
- https://doi.org/10.7554/eLife.35853.016
Figure 5—figure supplement 1
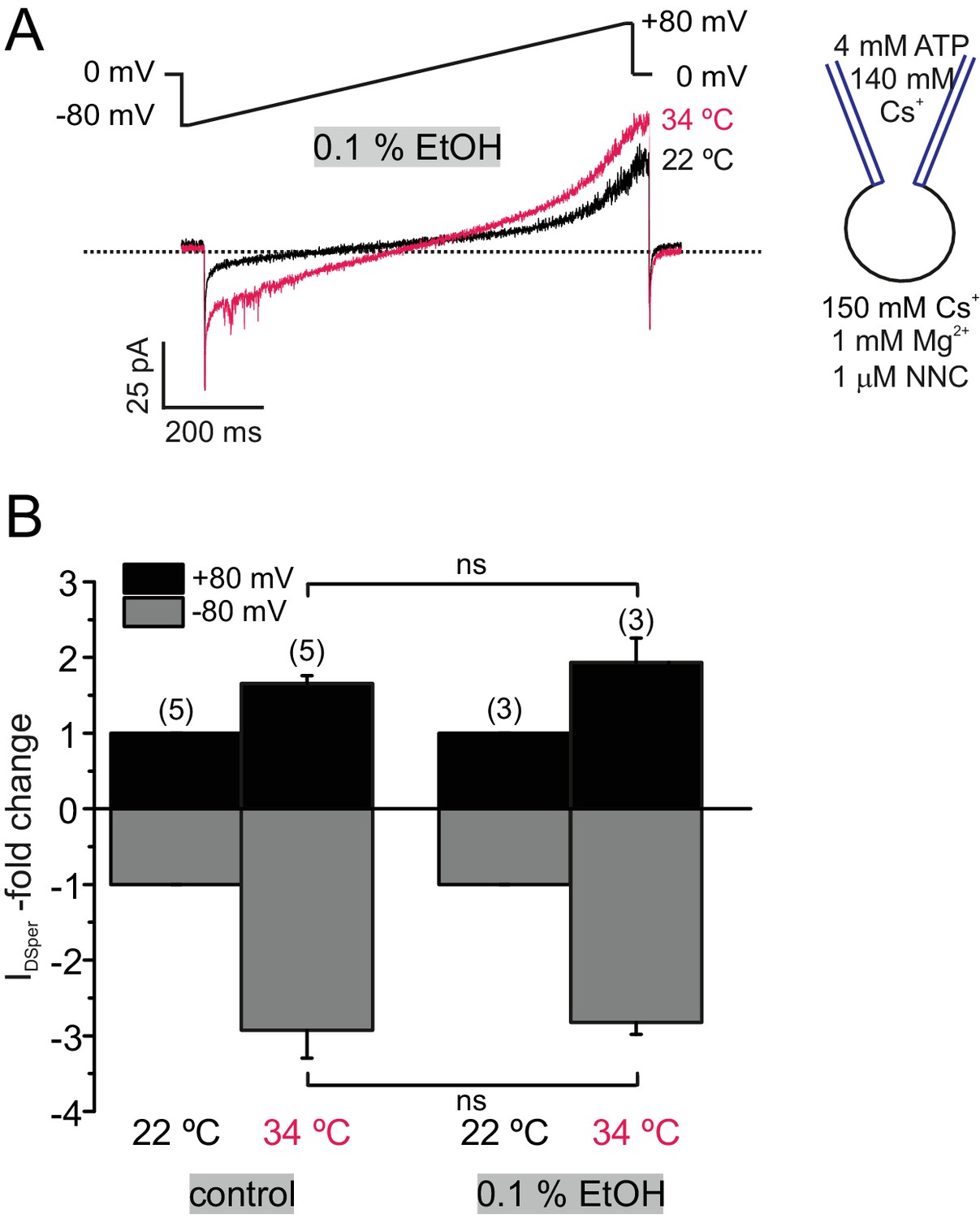
Ethanol does not affect DSper temperature activation.
(A) Original current traces from representative whole-cell patch-clamp recordings of noncapacitated human spermatozoa exposed to 22 and 34°C and an extracellular ethanol (EtOH) concentration of 0.1%. Inward and outward currents were elicited with voltage ramps as depicted. Rise in the temperature resulted in an increase in DSper inward- and outward currents similar to control conditions, indicating no effect by 0.1% EtOH as a solvent. (B) indicates the absence of inhibition by 0.1% extracellular EtOH as quantified normalized DSper inward and outward currents at 22 and 34°C, respectively.
Figure 5—figure supplement 2
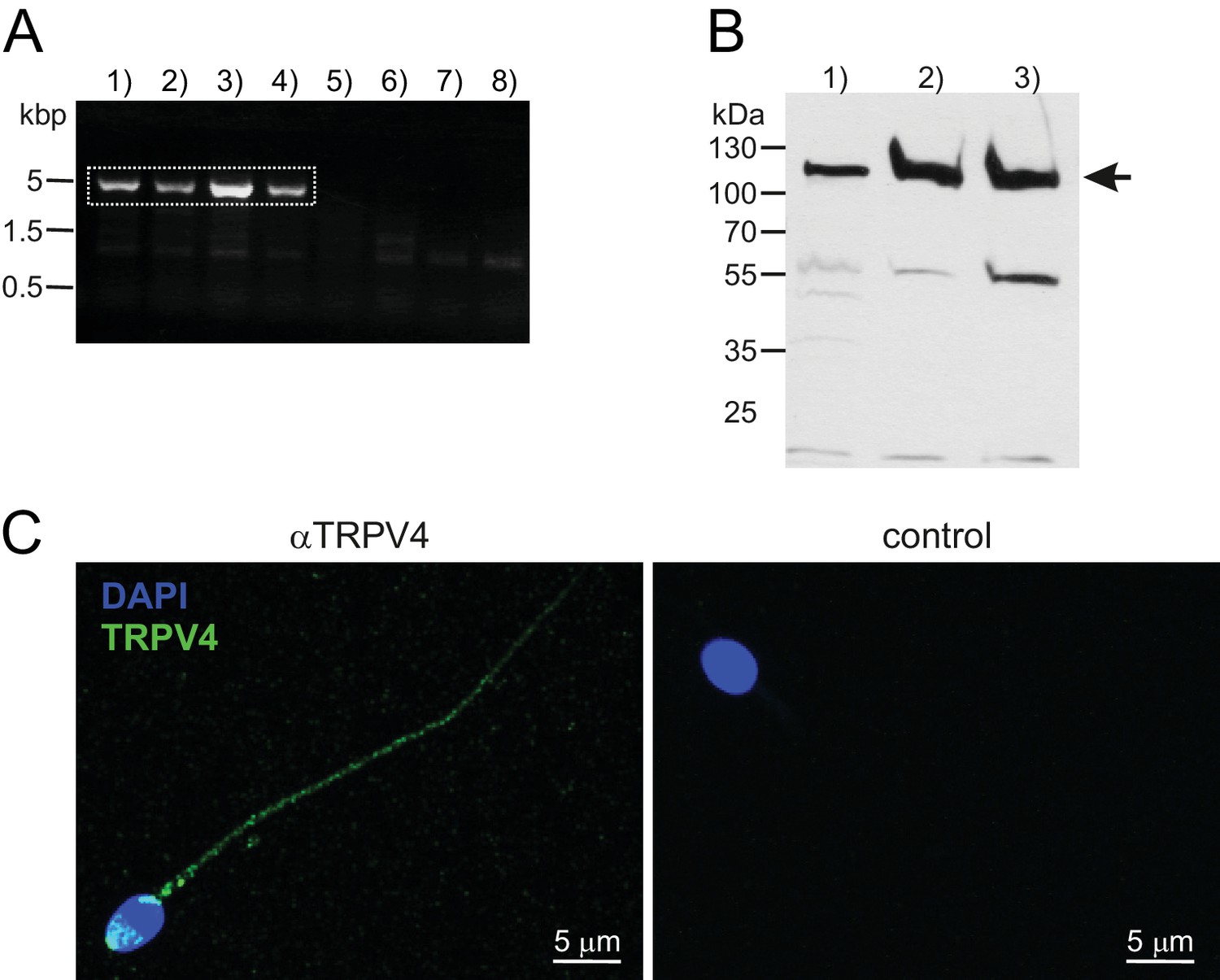
TRPV4 can be detected on the protein and mRNA levels.
(A) RT-PCR using a full-length TRPV4 primer pair. cDNA was obtained from mRNA isolated from swim-up purified noncapacitated human spermatozoa and PCR conditions were as follows: 1)–4) varying annealing temperatures (52–60°C), 5)–6) negative control in absence of the reverse transcriptase enzyme (Ta = 50, 56°C), 7)–8) no template control (Ta = 56°C). Dotted square marks bands that were selected for gene product sequencing. (B) Western blotting confirms the presence of the TRPV4 protein in (1) human testicular tissue (2) capacitated and (3) noncapacitated human spermatozoa. Immunopositive bands can be detected in all three samples at approximately 115 kDa. (C) Confocal fluorescence image of an immunostaining against TRPV4. Noncapacitated human spermatozoa were labeled with an anti-TRPV4 selective antibody and a Dylight488-conjugated secondary antibody. Nuclear dye DAPI locates the sperm head. Immunopositive fluorescent signals were detected in the sperm flagellum and the acrosome region (left). Specificity of the secondary antibody was confirmed by immunostaining in absence of the primary antibody (negative control, right).
Figure 6
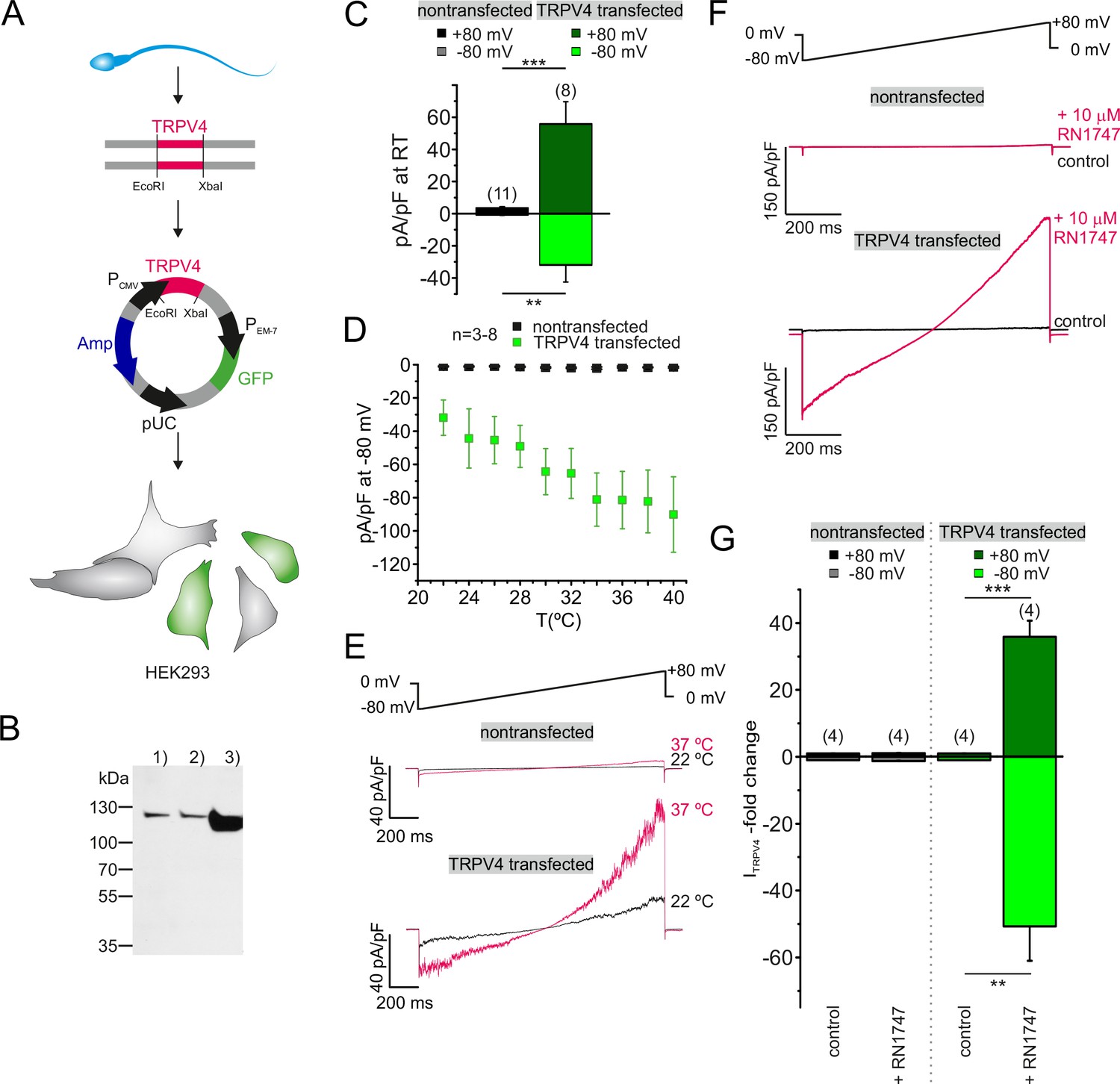
Sperm TRPV4 assembles into a functional channel when recombinantly expressed in HEK293 cells.
(A) Schematic representation of experimental approach: Full-length TRPV4 (isoform A) was cloned from human sperm mRNA extracts and recombinantly expressed in HEK293 cells, utilizing a pTracer CMV vector (Invitrogen). The GFP containing bicistronic vector allowed identification of green fluorescent transfected cells. (B) Western blotting results are shown for (1) nontransfected HEK293 cells, (2) cells transfected with the empty vector and (3) HEK293 cells transfected with the TRPV4-containing vector. An intense immunopositive band can be detected in line 3) at approx. 115 kDa corresponding to the expected size for TRPV4. Weak bands in 1) and 2) suggest endogenous expression of TRPV4 in HEK293 cells. (C) Electrophysiological characterization reveals a significant increase in the basic activity of TRPV4 transfected cells (at room temperature, 22°C). Inward and outward current densities were recorded as described before (ramp recordings −80 to +80 mV). Statistical significance (unpaired t-test) is indicated by: **p≤0.005, ***p≤0.0005. (D) Quantification of the inward current densities for nontransfected vs. TRPV4 transfected cells as a function of bath temperature (in °C) and representative current traces of whole-cell voltage clamp recordings for both cell populations in (E). Temperature-induced potentiation of inward and outward currents in TRPV4 transfected cells suggests that TRPV4 cloned from human sperm assembles into a functional protein in the heterologous expression system. (F) Whole-cell voltage clamp recordings reveal a strong potentiation of the inward and outward currents upon stimulation with the TRPV4 agonist RN1747 in TRPV4- transfected but not in nontransfected cells. Inward and outward currents were elicited via voltage ramps as depicted above. (G) Quantification of both inward and outward currents normalized to control conditions indicates statistical significance (**p≤0.005, ***p≤0.0005) for TRPV4 transfected cells.
-
Figure 6—source data 1
Recombinantly expressed TRPV4, initially isolated from human sperm mRNA pool, exhibits typical TRPV4-like behavior when expressed in HEK293 cells.
- https://doi.org/10.7554/eLife.35853.018
Figure 7
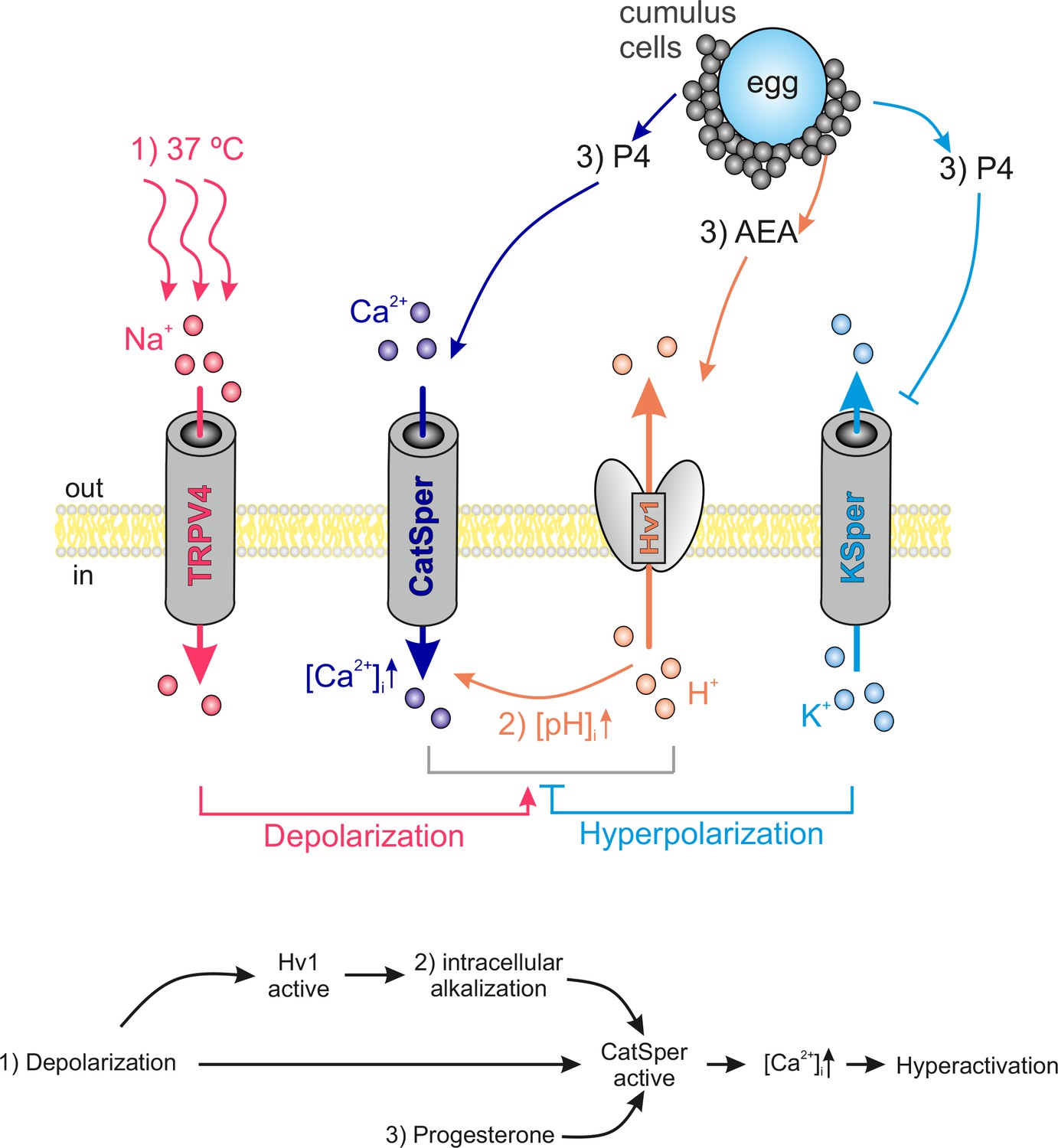
Interdependency of ion channel complexes in the human sperm flagellum.
Transition into hyperactivated motility is triggered by a CatSper-mediated rise in the cytosolic calcium levels. Proper CatSper function requires three concurrent activation mechanisms: (1) membrane depolarization, (2) intracellular alkalization via Hv1-mediated proton extrusion, and (3) abundance of progesterone. In our proposed model the sperm’s sodium channel TRPV4 is activated by warm temperatures (37°C at the site of fertilization). TRPV4-mediated sodium influx induces: (1) membrane depolarization, which in turn activates both Hv1 and CatSper. Hv1 then extrudes protons out of the sperm, thereby leading to (2) intracellular alkalization and further activation of CatSper. Cumulus cells surrounding the egg secrete (3) P4 and AEA. P4 releases CatSper inhibition and high P4 concentrations block KSper-mediated hyperpolarization. AEA was shown to potentiate Hv1. The resulting opening of CatSper generates a Ca2+ influx that serves as the trigger for hyperactivation.
Author response image 1
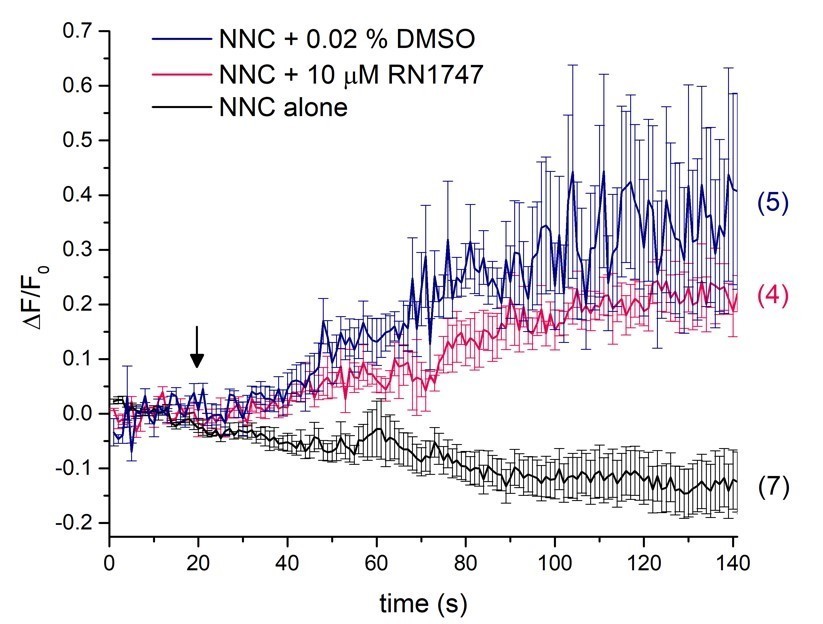
Single-cell calcium imaging of the sperm flagellum.
Arrow indicates application of either RN1747 dissolved in DMSO (red) or the corresponding concentration of DMSO as a vehicle control (blue). The CatSper inhibitor NNC was present during the whole recording period and did not induce a rise in cytosolic calcium levels as indicated by the black trace.
Tables
Key resources table
| Reagent type | Designation | Source | Identifier | Additional information |
|---|---|---|---|---|
| Cell line | HEK293 | ATCC | ATCC cat# CRL-1573 RRID:CVRL_0045 | |
| Compound | RN1747 | Tocris | 3745 | TRPV4 agonist |
| Compound | capsaicin | Cayman | 404-86-4 | TRPV1 agonist |
| Compound | carvacrol | Sigma | 499-75-2 | TRPV3 agonist |
| Compound | Pregnenolone sulfate | Sigma | P162 | TRPM3 agonist |
| Compound | HC067047 | Tocris | 4100 | TRPV4 antagonist |
| Compound | RN1734 | Tocris | 3746 | TRPV4 antagonist |
| Compound | NNC 55–0396 | R and D systems | 2268 | CatSper antagonist |
| Compound | Fluo-4/AM | Invitrogen | F14201 | Ca2+ indicator |
| Compound | Lipofectamine | Invitrogen | 11668019 | Transfection reagent |
| Antibody | α-TRPV4 | Alomone | Alomone cat# ACC-034 RRID:AB_2040264 | polyclonal; host: rabbit; unconjugated |
Table 1
Fitting parameters.
https://doi.org/10.7554/eLife.35853.020| Permeant cation | Cell type | A1 | A2 | x0 | Dx |
|---|---|---|---|---|---|
| Cesium | noncapacitated | 0.87314 | 3.45451 | 33.8 | 6.4 |
| Cesium | capacitated | 0.90892 | 2.19096 | 31.2 | 3.7 |
| Sodium | noncapacitated | 0.84686 | 5.19918 | 34.1 | 3.6 |
Additional files
-
Transparent reporting form
- https://doi.org/10.7554/eLife.35853.021
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
TRPV4 is the temperature-sensitive ion channel of human sperm
eLife 7:e35853.
https://doi.org/10.7554/eLife.35853




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}