Concerted IL-25R and IL-4Rα signaling drive innate type 2 effector immunity for optimal helminth expulsion
- Cardiff University, United Kingdom
- University of Cape Town, South Africa
- University of Glasgow, United Kingdom
- University of Edinburgh, United Kingdom
- MRC Laboratory of Molecular Biology, United Kingdom
Figures
Figure 1

IL-25R signaling is required for expulsion of H.
polygyrus from BALB/c mice. H. polygyrus-infected BALB/c or Il17rb-deficient (IL25R–/–) BALB/c mice were analyzed at day 14 post-infection for fecal egg counts (A) and intestinal adult worm burden (B) or day 28 post-infection for fecal egg counts (C) and intestinal adult worm burden (D). MLN cells underwent intracellular cytokine staining (ICCS) to compare the number of Lin-ICOS+innate lymphoid cells (ILCs) (E) and IL-13+ ILC2s (F) by flow cytometry in the different naïve and day 14 infected genotypes. Peritoneal lavage cells (PL) were stained with Siglec-F, CD11b and Ly6G/C to compare the number of Siglec-F-CD11b+Ly6G/C+ monocytes (G) in the different naïve and day 14 infected genotypes. Percentages of eosinophils (H) and RELMα+ alternatively-activated macrophages (Ι) were also determined over a 4 week infection timecourse. ELISA of supernatants from MLN cells incubated with media or 1 μg HES for 72 hr was performed to compare IL-4 (J), IL-5 (K) and IL-13 (L) production in the different naïve and day 14 infected genotypes. ICCS of MLN allowed a comparison of the proportion of IL-4+, IL-5+ and IL-13+ CD4+ T cells by flow cytometry in the different naïve and day 14 infected genotypes (M–O). Results shown are one representative of three experiments with n ≥ 4 mice/group (A–D), pooled data from two experiments with n ≥ 4 mice/group (C–F, M–O) or one representative of two experiments with n ≥ 4 mice/group (G–L). Data were analysed by unpaired t test or one way ANOVA, where *=p≤0.05,**=p≤0.01,***=p≤0.001 and ns = not significant. Error bars represent Standard Error of the Mean.
Figure 2
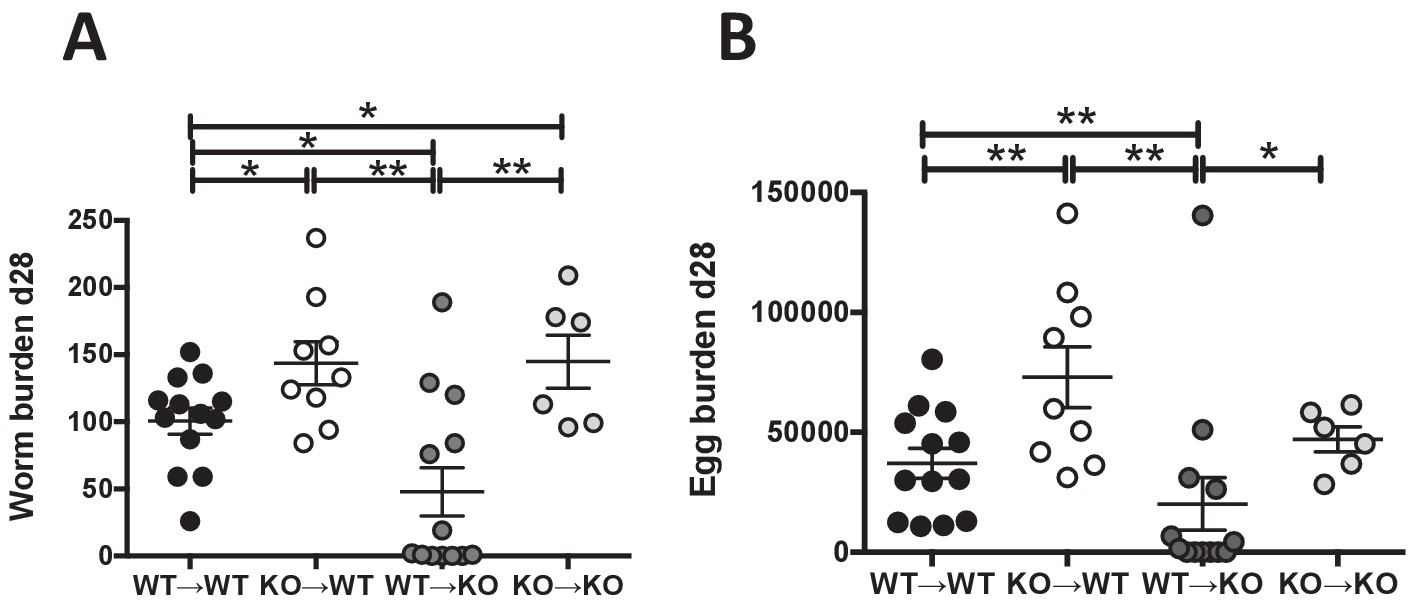
IL-25R signaling is required within the hematopoietic compartment for efficient expulsion.
Bone marrow chimeras generated from BALB/c (WT) or Il17rb-deficient (KO) donor and BALB/c (WT) or Il17rb-deficient (KO) recipient mice were infected with H. polygyrus and intestinal adult worm burden (A) and fecal egg burden (B) performed at day 28 post-infection. Results shown are pooled data from two experiments performed with n ≥ 3 mice/group, and data from all individual mice are presented. Data were analysed by unpaired t test, where *=p≤0.05,**=p≤0.01,***=p≤0.001 and ns = not significant. Error bars represent Standard Error of the Mean.
Figure 3
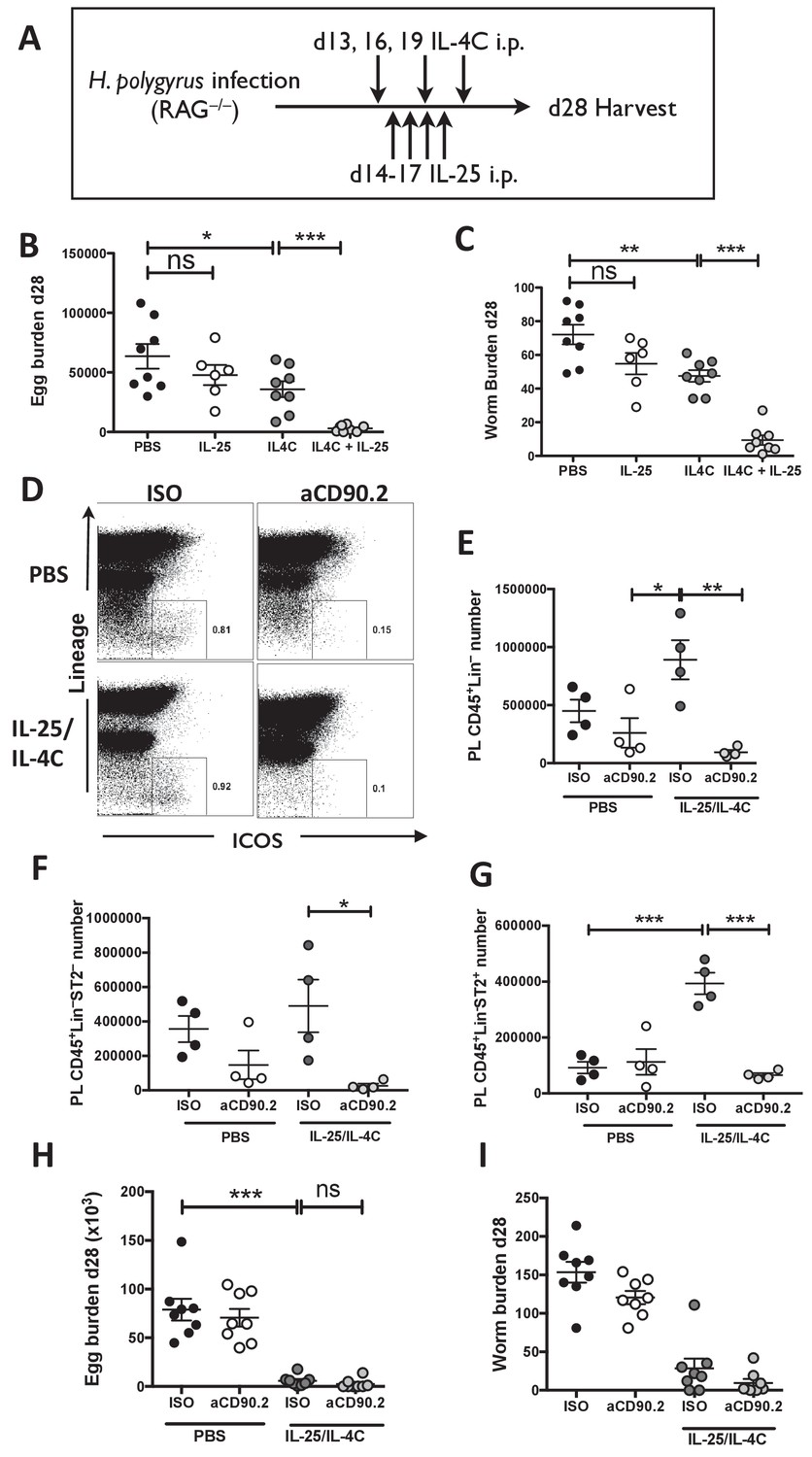
IL-25R signaling synergises with IL-4Rα within the innate immune compartment to facilitate efficient worm expulsion.
H. polygyrus-infected Rag1-/- mice (RAG-/-) were given 0.4 µg recombinant IL-25 i.p. days 14–17 (late) post-infection with or without a complex of 5 μg rIL-4 and 25 μg anti-IL-4 (IL-4C) on days 13, 16 and 19 post-infection, according the schedule shown in (A). Mice were analyzed at 28 days post-infection for fecal egg burden (B) and intestinal adult worm burden (C). H. polygyrus-infected Rag1-/- mice were given IL-25 and IL-4C according to the same schedule, as well as 200 µg of anti-CD90.2/Th1.2 antibody or rat IgG2b control (days 12, 15, 18 and 21). The peritoneal lavage was analyzed at 28 days post-infection for CD45+lin- (CD3, CD5, CD8α, CD11c, CD19, DX5, F4/80, GR-1, TCRβ, CD11b), ICOS and ST2 staining by flow cytometry as shown (D) and the number of CD45+lin– (E), CD45+lin–ST2– (F) and CD45+lin–ST2+ (G) ILCs was determined. Mice were analysed at 28 days post-infection for fecal egg burden (H) and intestinal adult worm burden (I). Results shown are one representative of two experiments with n = 4 mice/group (D–G), or pooled data from two experiments with n ≥ 3 mice/group (B,C,H,I). Data were analysed by unpaired t test, where *=p≤0.05,**=p≤0.01,***=p≤0.001 and ns = not significant. Error bars represent Standard Error of the Mean.
Figure 4
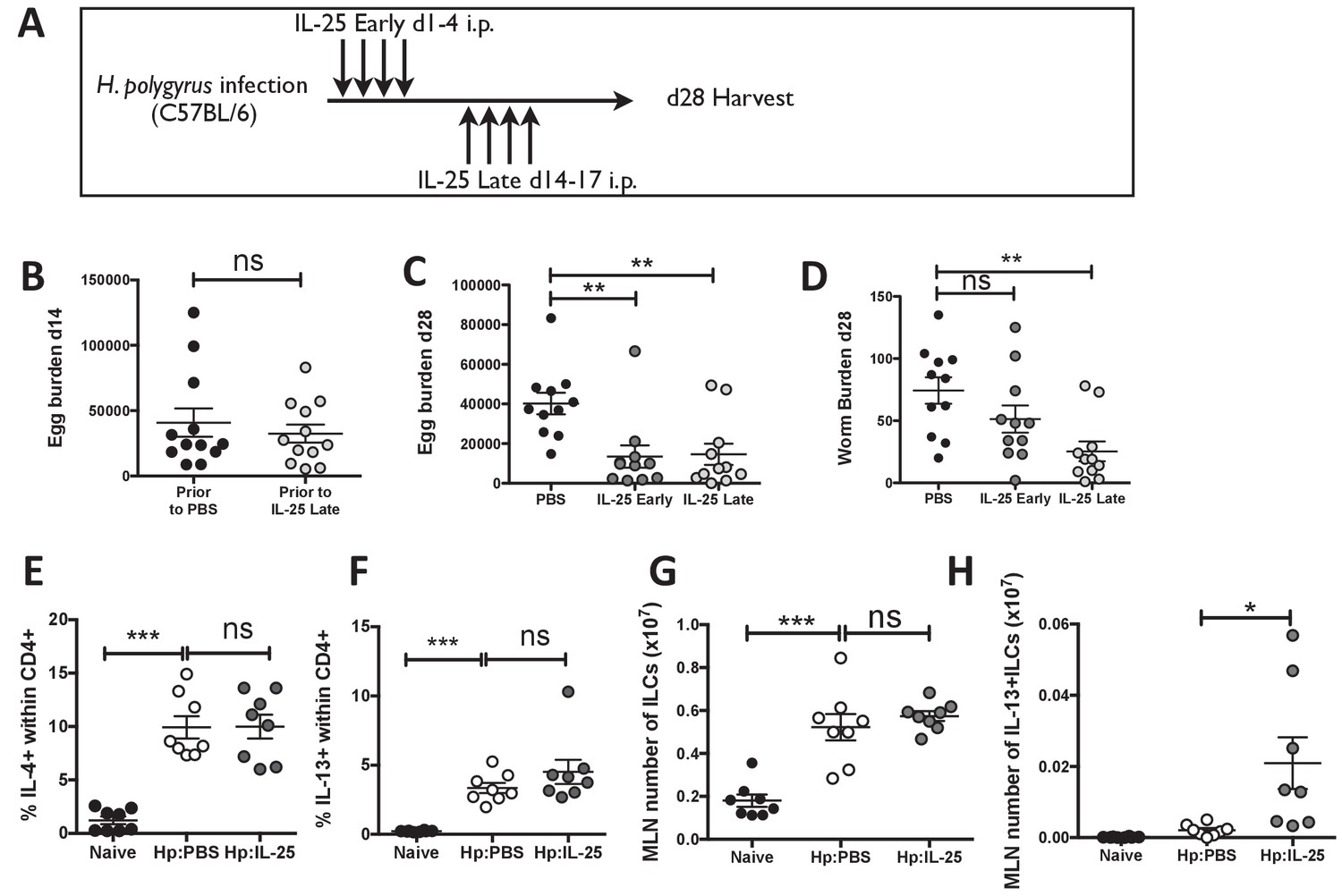
IL-25 induces protective immune responses in the late stage of infection.
H. polygyrus-infected C57BL/6 mice were given 0.4 μg recombinant IL-25 i.p. at day 1–4 (early) or day 14–17 (late) post-infection according to the schedule shown in (A). Before administration of IL-25, intestinal egg burden was analysed in two groups at day 14 post-infection (B). Mice were then analysed at day 28 post-infection for fecal egg counts (C) and intestinal adult worm burden (D) following administration of IL-25 early or late. H. polygyrus-infected C57BL/6 mice were given 0.4 μg recombinant IL-25 i.p. at day 14–17 (late) post-infection. Day 18 post-infection, MLN cells underwent ICCS to compare the proportion of IL-4+CD4+ (E) and IL-13+CD4+ (F) T cells, as well as the number of ILCs (G) and Lin-IL-13 + ILC2 s (H) by flow cytometry. Results shown are data pooled from three experiments with n ≥ 3 mice/group (B–D), or are representative of two experiments with n = 4 mice/group (E,F) or pooled from two experiments with n = 4 mice/group (G, H). Data were analysed by unpaired t test, where *=p≤0.05,**=p≤0.01,***=p≤0.001 and ns = not significant. Error bars represent Standard Error of the Mean.
Figure 5
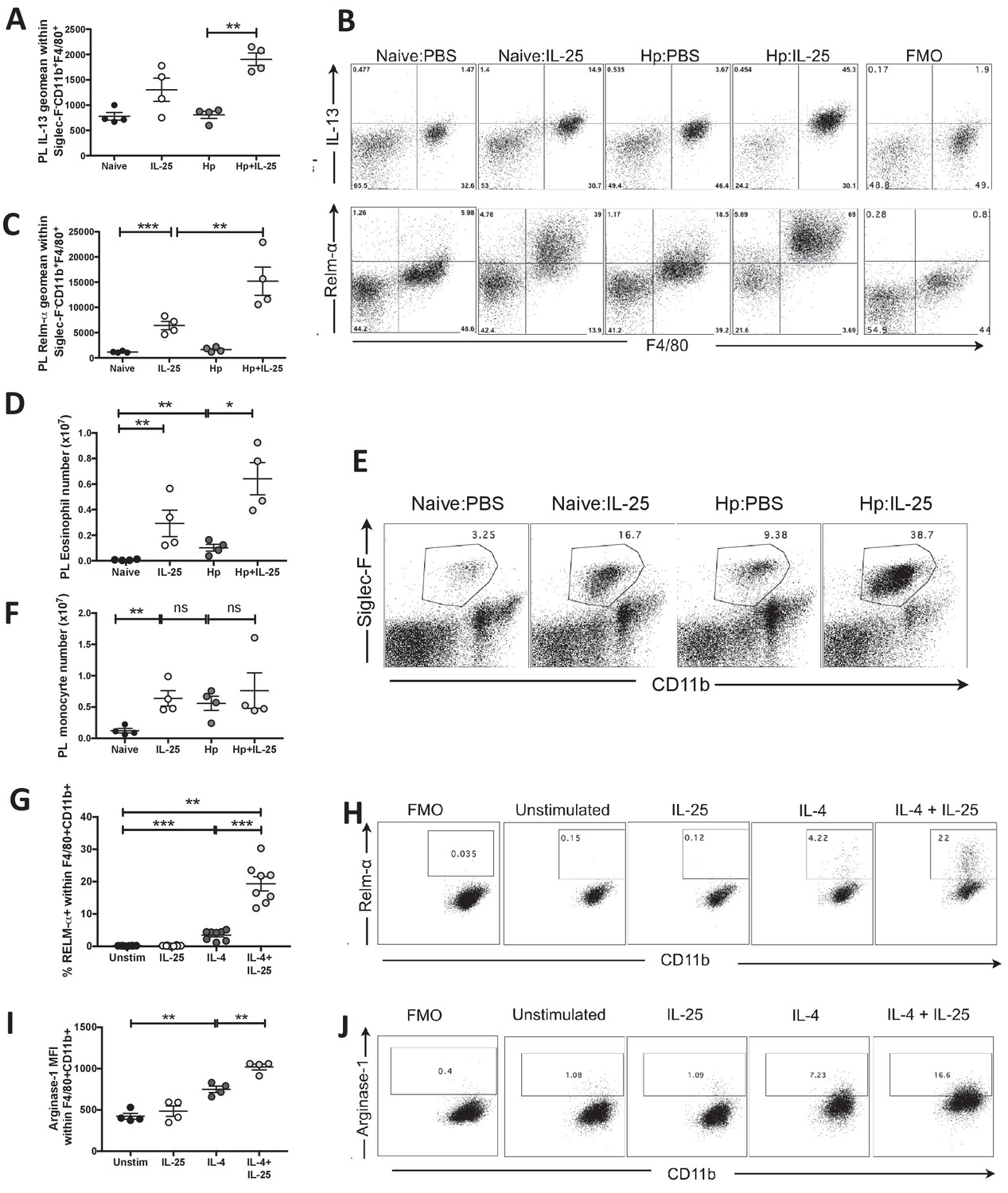
IL-25 drives alternative activation and IL-13 expression of macrophages.
H. polygyrus-infected C57BL/6 mice were given 0.4 μg recombinant IL-25 i.p. at day 14–17 (late) post-infection. At day 18 post-infection, PL cells were taken and stimulated with 10 μg Brefeldin A to determine the expression intensity (geomean) of IL-13 (A) or were unstimulated to determine the expression intensity (geomean) of RELM-α (B, C) within Siglec-F–CD11b+F4/80+ monocytes by flow cytometry, as shown by the example flow cytometry plots in B and compared to fluorescence minus one (FMO) samples stained from the Hp:IL-25 group. Total Siglec-F+CD11b+ eosinophil (D) numbers were calculated from populations shown in the example flow cytometry plots (E), numbers of Siglec-F–CD11b+ (F) monocytes. Bone marrow-derived macrophages from C57BL/6 mice were generated in vitro and stimulated with 10 ng/ml IL-4, 200 ng/ml IL-25 or a combination of both for 16 hr before analysis of the percentage of RELM-α expression (G, H) and the mean fluorescence intensity (geometric mean) of Arginase-1 expression (I, J) within CD11b+F4/80+ cells by flow cytometry. Results shown are one representative of 2 experiments with n = 4 mice/group (A–F, I, J) or pooled from two experiments with n = 4 replicates/group (G, H). Data were analysed by unpaired t test, where *=p≤0.05,**=p≤0.01,***=p≤0.001 and ns = not significant. Error bars represent Standard Error of the Mean.
Figure 6
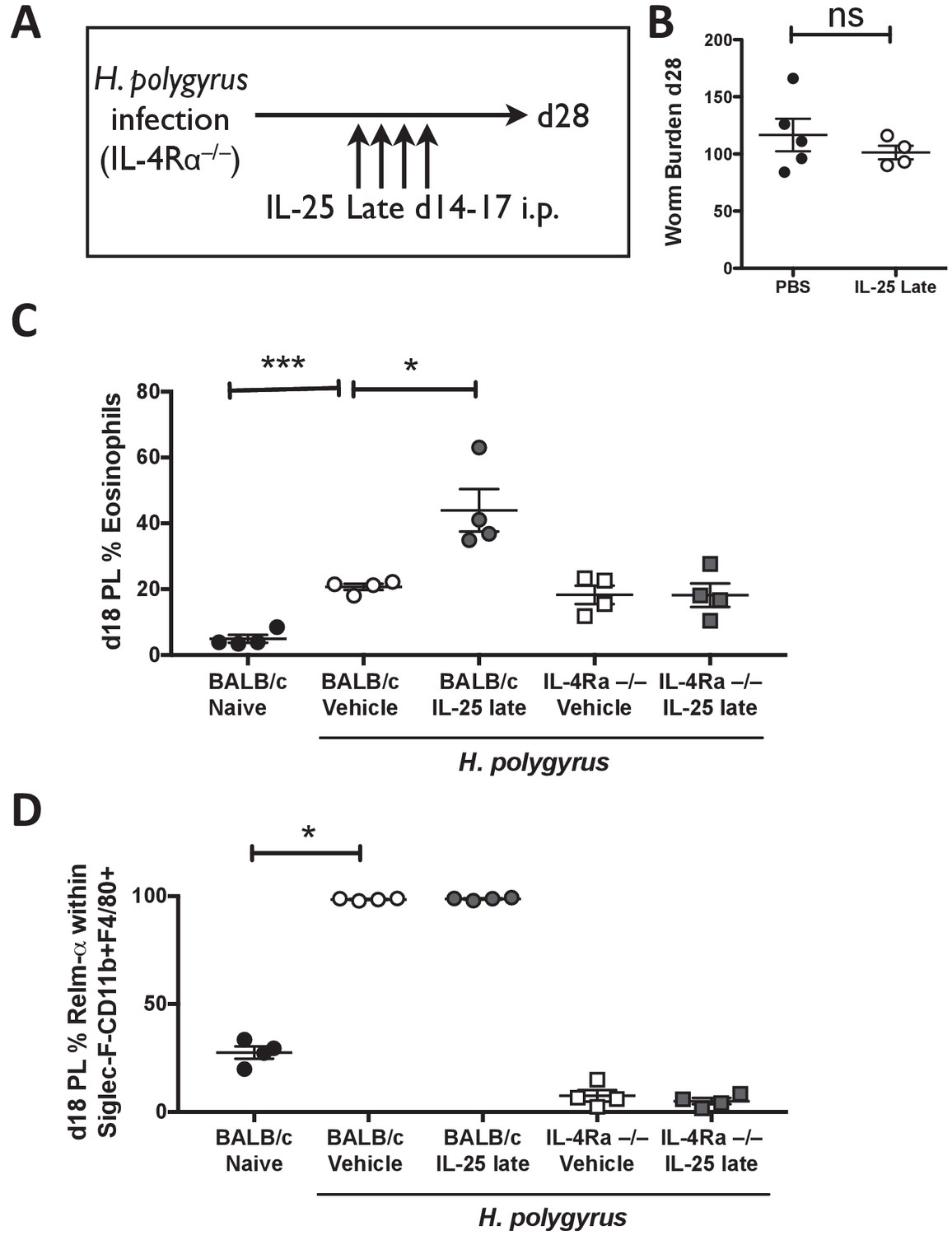
IL-4Rα expression is required for activation of the immune system by IL-25.
H. polygyrus-infected Il4ra-/- mice (IL-4R-/-) were given 0.4 μg recombinant IL-25 i.p. days 14–17 (late) post-infection (A). Mice were analyzed at day 28 post-infection for intestinal adult worm burden (B). H. polygyrus-infected BALB/c and Il4ra-/- mice (IL-4R-/-) were given 0.4 μg recombinant IL-25 i.p. days 14–17 (late) post-infection. Day 18 post-infection, PL cells were taken to determine the percentage of eosinophils (C) and percentage of RELM-α expression within Siglec-F-CD11b+F4/80+ macrophages by flow cytometry (D). Results shown are one representative of 2 experiments with n ≥ 4 mice/group (A–D). Data were analysed by unpaired t test, where *=p≤0.05,**=p≤0.01,***=p≤0.001 and ns = not significant. Error bars represent Standard Error of the Mean.
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers |
|---|---|---|---|
| Mus musculus, BALB/c | Il17rb-/- (IL-25R-/-) | Neill et al. (2010) Nuocytes represent a new innate effector leukocyte that mediates type-2 immunity. Nature 464:1367–1370 | |
| Mus musculus, C57BL/6 | Rag1-/- | Mombaerts,P., Iacomini,J., Johnson, R.S., Herrup, K., Tonegawa,S., and Papaioannou, V. E. 1992. Cell. Vol 68: 869–877. PMID: 1547488 | |
| Mus musculus, C57BL/6 | Il4ra-/- | Noben-Trauth N, Shultz LD, Brombacher F, Urban JF, Jr., Gu H, Paul WE. 1997. An interleukin 4 (IL-4)-independent pathway for CD4 + T cell IL-4 production is revealed in IL-4 receptor-deficient mice. Proc Natl Acad Sci USA 94:10838–10843. | |
| Parasite | Heligmosomoides polygyrus bakeri | Johnston et al., 2015. Cultivation of Heligmosomoides polygyrus: an immunomodulatory nematode parasite and its secreted products. Journal of Visualized Experiments 98:e52412. | |
| Antibody | Anti-IL-4 | BioXCell | Clone 11B11 |
| Antibody | Anti-IL-13 | BioLegend | JES10-5A2 |
| Antibody | Anti-CD90.2 | BioXCell | 30H12 |
| Antibody | Anti-SiglecF | BD Pharmingen | E50-2440 |
| Antibody | Anti-CD11b | BioLegend | M1/70 |
| Antibody | Anti-F4/80 | BioLegend | BM8 |
| Antibody | Anti-Ly6G | BioLegend | 1A8 |
| Antibody | Anti-Ly6C | BioLegend | AL-21 |
| Antibody | Anti-RELMα | R and D Systems | 22603 |
| Antibody | Anti-Arginase-1 (Polyclonal) | R and D Systems | IC5868P |
| Antibody | Anti-Lineage | BioLegend | 17A2,RB6-8C5,RA3-6B2,Ter-119,M1/70 |
| Peptide, recombinant protein | IL-4 | Peprotech | 214–14 |
| Peptide, recombinant protein | IL-25 | BioLegend | 587302 |
| Commercial assay or kit | Foxp3 Staining Kit | eBioScience | 88–8118 |
| Other | Control IgG2b immunoglobulin | BioXcell | LTF-2 |
| Other | Anti-CD90.2 Microbeads | Miltenyi | 130-049-101 |
Additional files
-
Transparent reporting form
- https://doi.org/10.7554/eLife.38269.008
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Concerted IL-25R and IL-4Rα signaling drive innate type 2 effector immunity for optimal helminth expulsion
eLife 7:e38269.
https://doi.org/10.7554/eLife.38269




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}