Regulating G protein-coupled receptors by topological inversion
- University of Texas Southwestern Medical Center, United States
Figures
Figure 1 with 1 supplement
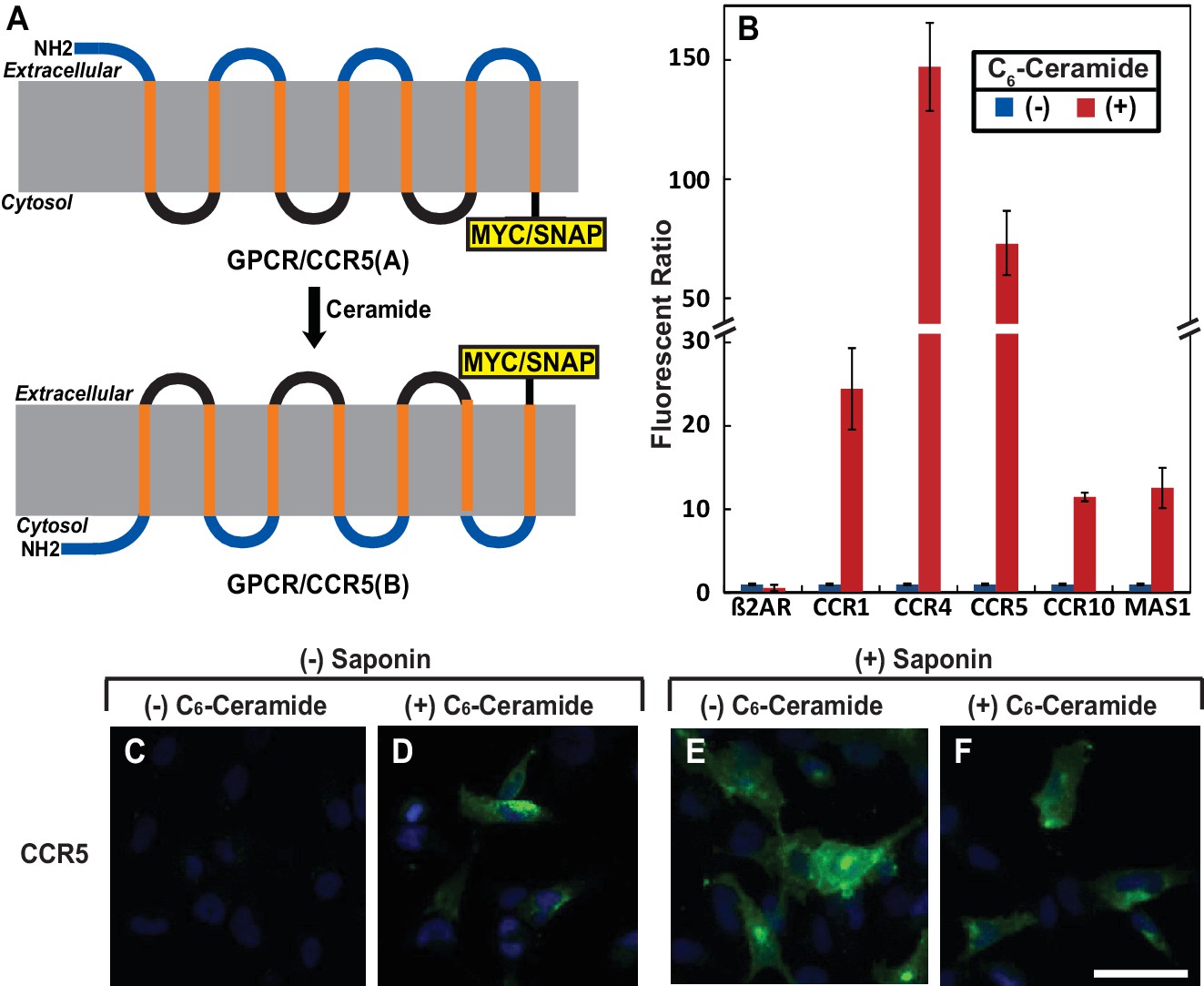
Ceramide alters localization of C-terminus of GPCRs containing the GXXXN motif.
(A) Schematic illustration of topological inversion of C-terminally tagged GPCRs. (B) HEK-293 cells transfected with a plasmid encoding the indicated GPCR fused with a SNAP-tag at the C-terminus were treated with 8 μM C6-ceramide for 16 hr, and labeled with a cell permeable or impermeable SNAP-tag substrate. The ratio of fluorescent signal generated from cell impermeable versus that from cell permeable substrate was reported, with the value from cells untreated with C6-ceramide set at 1. Results were reported as Mean ±S.E. from triplicate incubations of a typical experiment. Similar results were obtained from two other independent experiments. (C–F) SV589 cells transfected with pCCR5-Myc were treated with C6-ceramide as described in B, and subjected to immunofluorescent microscopy analysis with anti-Myc in the absence (C, D) or presence (E, F) of saponin-mediated cell permeabilization. Scale bar = 50 μm.
Figure 1—figure supplement 1
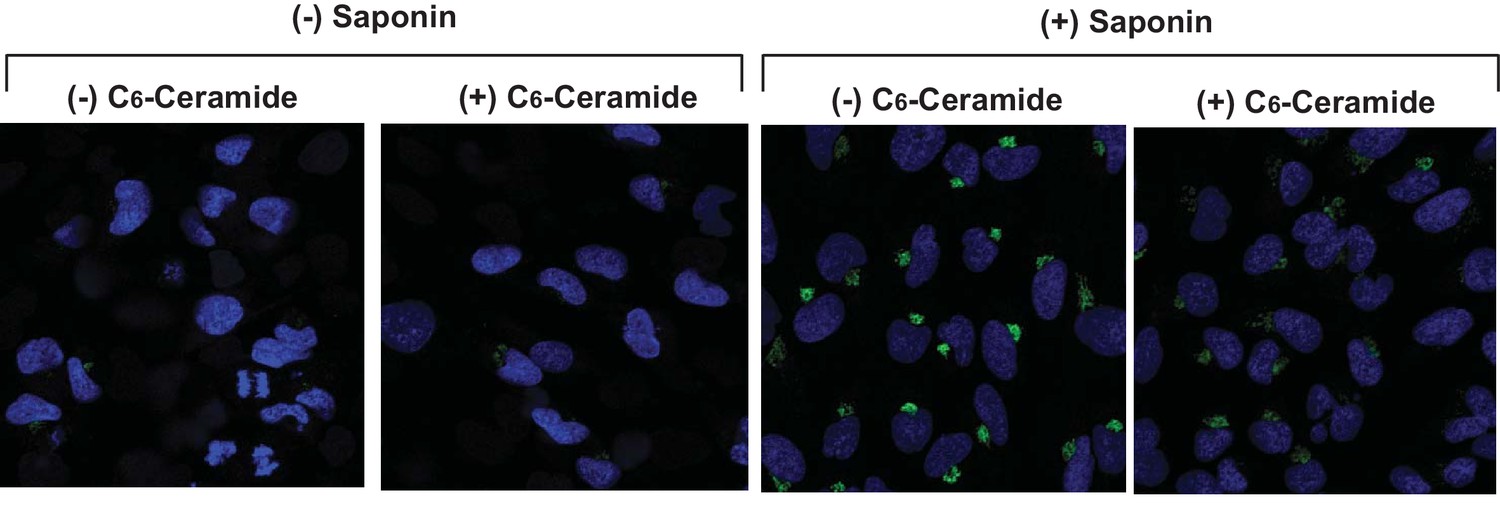
Ceramide does not increase cell permeability.
SV589 cells were treated with or without C6-ceramide and detected for expression of Giantin, a Golgi marker, by immunofluorescent microscopy in the absence or presence of saponin as described in Figure 1C-F.
Figure 2 with 2 supplements
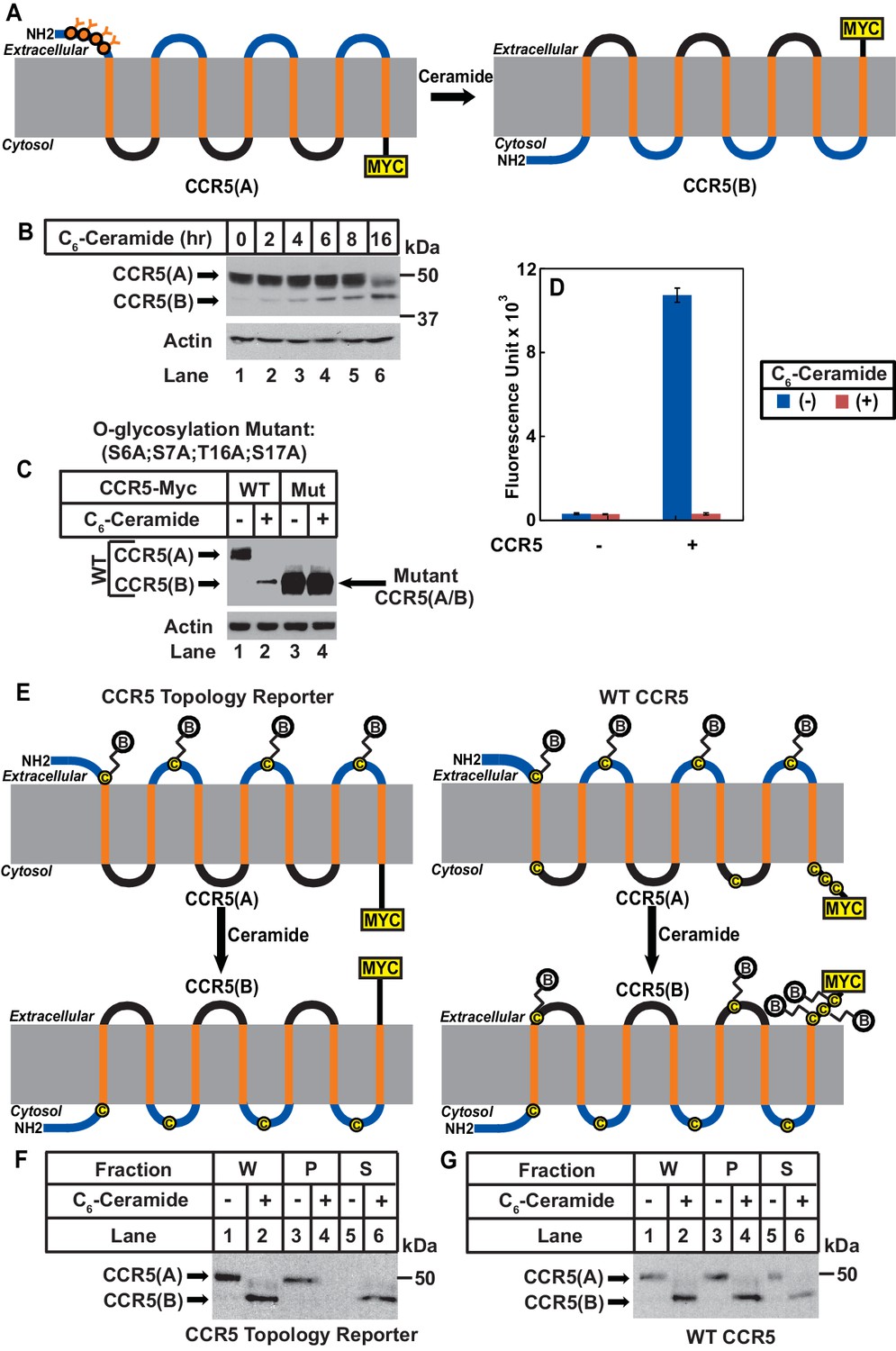
Ceramide stimulates RAT of CCR5.
(A) Schematic illustration of the effect of RAT on O-linked glycosylation of CCR5. (B) SV589 cells transfected with pCCR5-myc were treated with 8 μM C6-ceramide for the indicated time followed by immunoblot analysis with anti-Myc. (C) SV589 cells transfected with the indicated plasmid were treated with or without 8 μM C6-ceramide for 16 hr followed by immunoblot analysis with anti-myc. (D) HEK293 cells transfected with pSNAP-CCR5 were treated with 8 μM C6-ceramide for 16 hr, labeled with a cell-impermeable fluorescent substrate for SNAP-tag, and quantified the labeling reaction through a fluorimeter. Results are reported as mean ±S.E. from triplicate incubations of a typical experiment. (E) Schematic illustration of the effect of RAT on cell surface labeling of extracellular cysteine residues in wildtype and the topology reporter CCR5. (F and G) SV589 cells transfected with Myc-tagged wildtype or topology reporter CCR5 were treated with 8 μM C6-ceramide for 16 hr. After cell surface labeling of extracellular cysteine residues with biotin, cell lysates were precipitated with streptavidin beads. Equal fractions of whole cell lysate (W), pellet (P) and supernatant (S) were subjected to immunoblot analysis with anti-Myc.
Figure 2—figure supplement 1
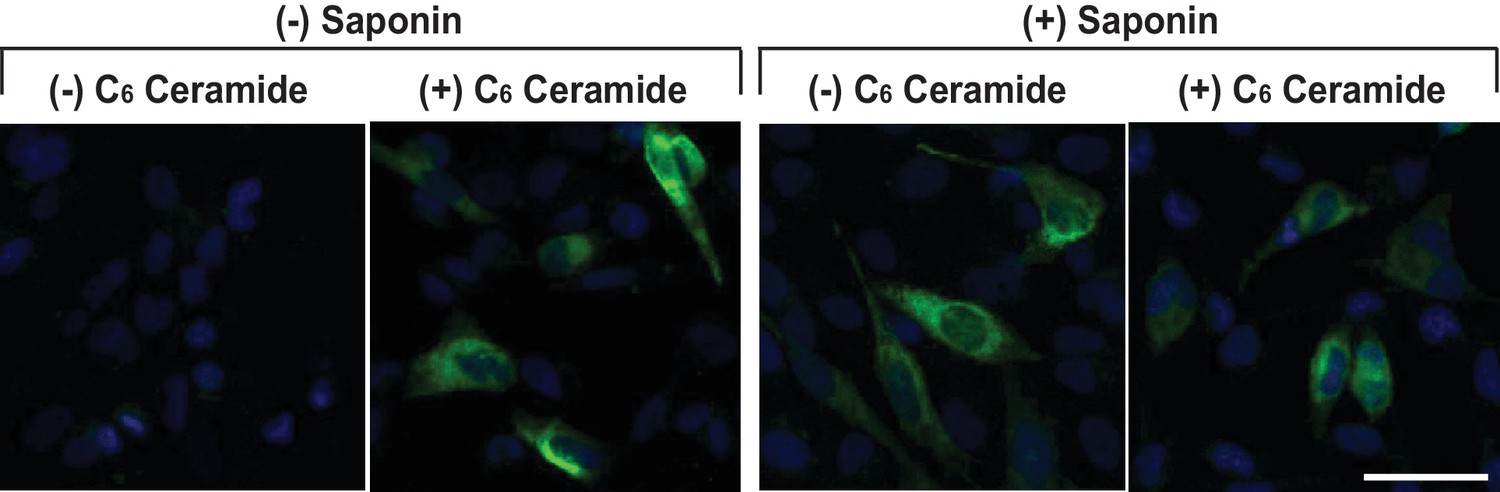
Mutations bocking glycosylation do not affect RAT of CCR5.
Localization of the Myc-tag fused at the C-terminus of CCR5(S6A, S7A, T16A, S17A) was analyzed as described in Figure 1. Scale bar = 50 μm.
Figure 2—figure supplement 2
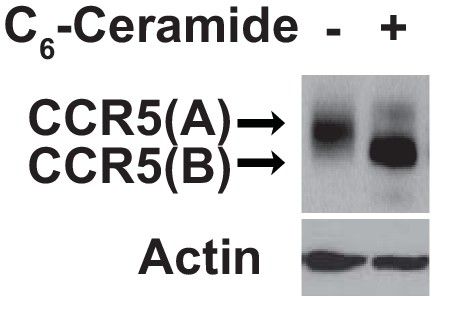
Ceramide treatment does not affect expression of SNAP-CCR5.
HEK293 cells were setup, transfected, and treated as described in Figure 2D. The amount of transfected SNAP-tagged CCR5 was analyzed by immunoblot analysis with anti-SNAP.
Figure 3 with 3 supplements

LPS induces RAT of CCR5 in primary mouse macrophages.
(A–I) Mouse macrophages were treated with or without 100 ng/ml LPS in the absence (B–E) or presence of 30 μM FB1 (F–I) for 16 hr. (A) The amount of dihydroceramide with the indicated amide-linked acyl chains in the cells was determined through LC-MS measurement. Results are reported as mean ±S.E. from three independent experiments. (B–I) Macrophages were subjected to immunofluorescent microscopy analysis with an antibody against the seoncd extracellular loop of CCR5(A) in the absence or presence of saponin-mediated cell permeabilization. Scale bar = 10 μm. (J and K) Macrophages treated without (J) or with (K) 100 ng/ml LPS for 24 hr were subjected to immuno-gold EM analysis with an antibody against the N-terminal domain of CCR5. Scale bar = 200 nm. (L) The number of intracellular and extracellular-localized CCR5 labeled by gold clusters per cell was quantified from macrophages treated with (n = 20) and without LPS (n = 28). The results are reported as mean ±S.D. This number should be smaller than that of CCR5 molecules, as it did not include CCR5 labeled by a single gold particle, the specificity of which was difficult to determine.
Figure 3—figure supplement 1
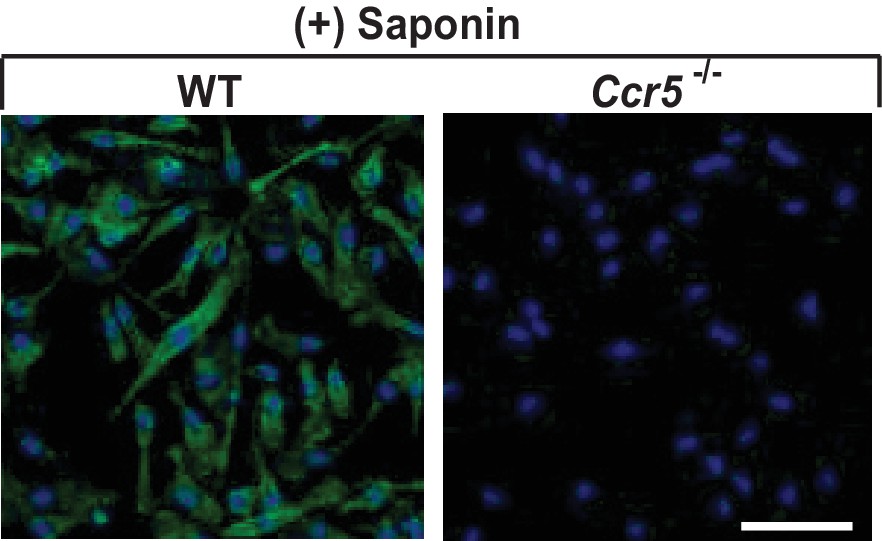
The specificity of CCR5 immunofluorescent microscopy.
Permeabilized mouse macrophages from wildtype (WT) or Ccr5 mice were subjected to immunofluorescent microscopy analysis with an antibody against the second extracellular loop of CCR5(A). Scale bar = 10 μm.
Figure 3—figure supplement 2
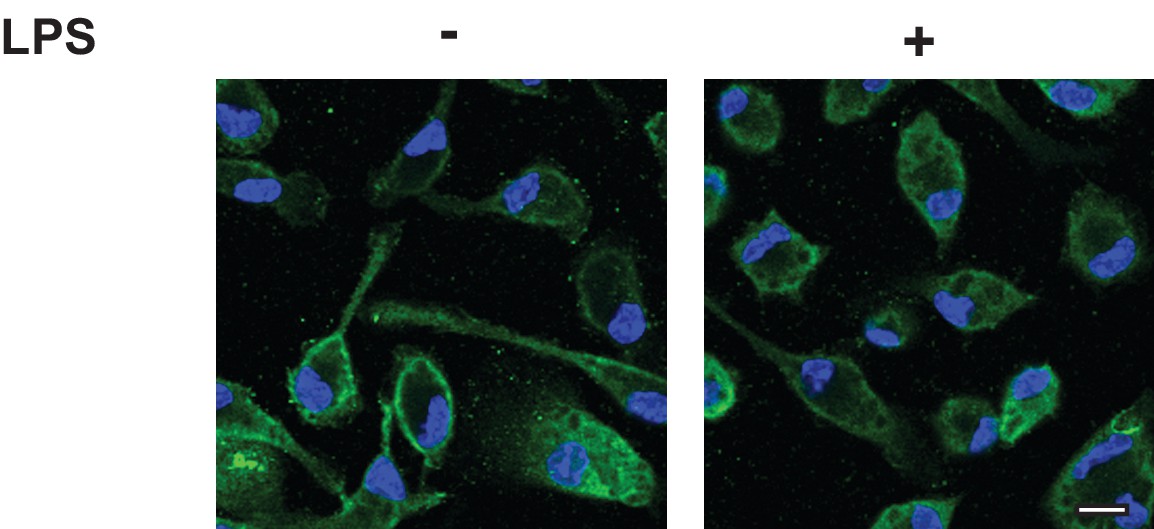
CCR5 is localized on cell surface regardless of LPS treatment.
Lighter exposure of images shown in Figure 3D and E. Scale bar = 10 µm.
Figure 3—figure supplement 3
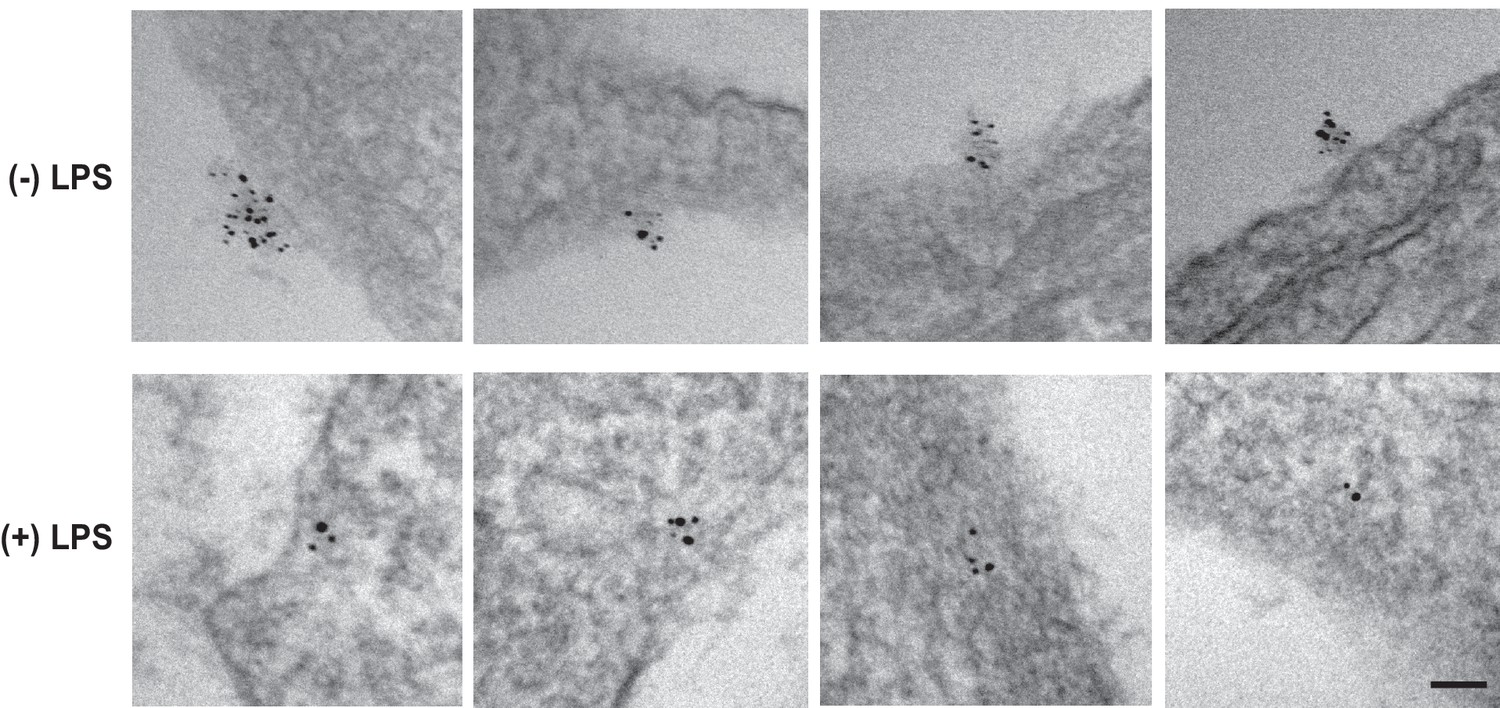
LPS stimulates RAT of CCR5 in macrophages.
CCR5 was detected in macrophages through EM as described in Figure 3J and K. Scale bar = 100 nm.
Figure 4
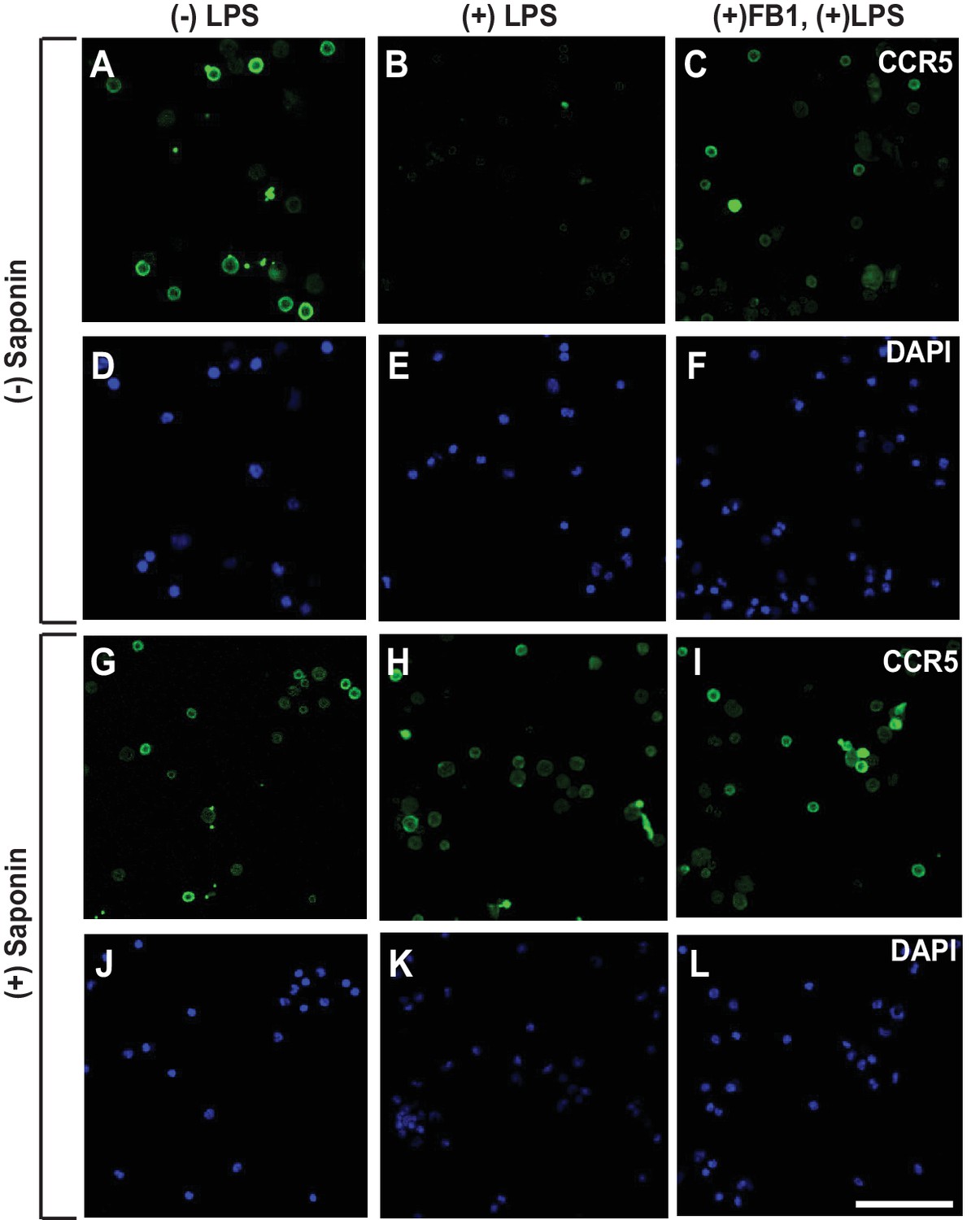
LPS injected intraperitoneally induces RAT of CCR5 in macrophages.
Macrophages isolated from peritoneal cavity of mice injected intraperitoneally with 1 mg/kg LPS and 1 mg/kg FB1 for 16 hr were analyzed as described in Figure 3B–I. Scale bar = 20 μm.
Figure 5
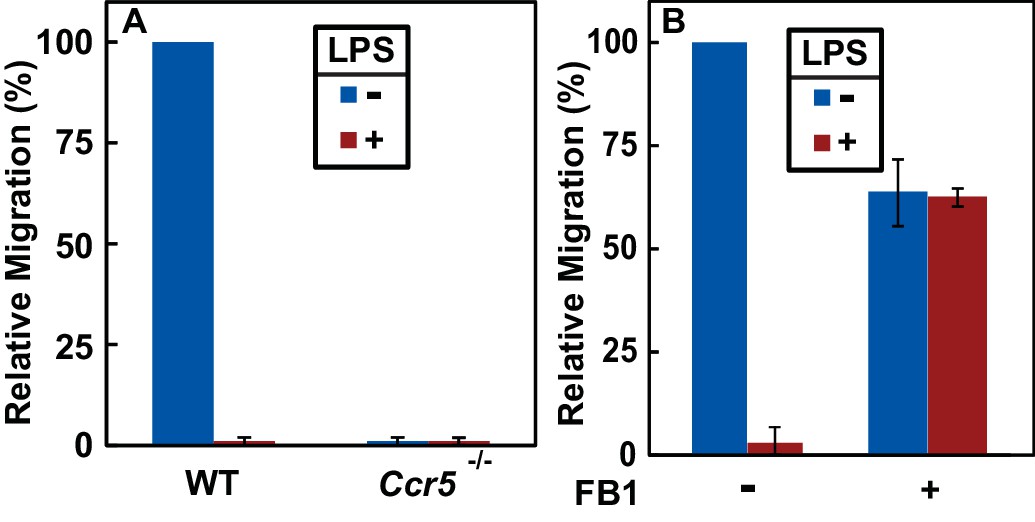
RAT of CCR5 blocks migration of macrophages toward CCL5.
(A) Migration of macrophages from mice with indicated genotype was assayed in transwell plates for 24 hr with or without 100 ng/ml LPS in the upper wells and 100 nM CCL5 as the chemoattractant in lower wells. Signals generated in the absence of CCL5 were subtracted to normalize for random migration. (B) Migration assays were performed as described in (A) with wildtype macrophages incubated with LPS and 30 μM FB1 in upper wells as indicated. (A and B) Results were reported as mean ±S.E. from three independent experiments, with the value obtained from untreated WT macrophages set at 100%.
Figure 6

A speculative model illustrating LPS-induced topological inversion of CCR5 through RAT.
Upon infection, cells surrounding the bacterial infected sites secret chemokines including CCL5. Through its interaction with CCR5 expressed in unprimed macrophages that adopts a topology consistent with that of a GPCR (CCR5(A)), CCL5 attracts migration of macrophages toward the infection site. Upon encounter with bacterial byproduct LPS, the increased production of dihydroceramide in macrophages causes RAT of CCR5, leading to expression of CCR5 with an inverted topology (CCR5(B)). This topological inversion prevents further migration of macrophages toward CCL5.
Tables
Table 1
GXXXN motif presented in the first transmembrane helix of selected GPCRs.
The GXXXN motif present in the first transmembrane helix of human TM4SF20, chemokine receptors and MAS1 is highlighted. The amino acid numbers of the aligned residues are indicated.
| GXXXN | |
| TM4SF20(14–34) | SLLVLLLLGVVLNAIPLIVSL |
| CCR1(40–60) | LYSLVFVIGLVGNILVVLVLV |
| CCR2(48–68) | LYSLVFIFGFVGNMLVVLILI |
| CCR3(58–78) | LYSLVFTVGLLGNVVVVMILI |
| CCR4(45–65) | LYSLVFVFGLLGNSVVVLVLF |
| CCR5(36–56) | LYSLVFIFGFVGNMLVILILI |
| CCR6(52–72) | AYSLICVFGLLGNILVVITFA |
| CCR7(64–84) | MYSIICFVGLLGNGLVVLTYI |
| CCR9(54–74) | LYWLVFIVGALGNSLVILVYW |
| CCR10(47–67) | VSLTVAALGLAGNGLVLATHL |
| CXCR3(59–79) | LYSLLFLLGLLGNGAVAAVLL |
| CXCR4(48–68) | IYSIIFLTGIVGNGLVILVMG |
| CXCR5(57–77) | AYSLIFLLGVIGNVLVLVILE |
| CXCR6(37–57) | MYLVVFVCGLVGNSLVLVISI |
| MAS1(38–58) | VIMSISPVGFVENGILLWFLC |
-
Table 1—source data 1
Proteins containing a GXXXN motif in their first transmembrnae helix.
- https://doi.org/10.7554/eLife.40234.005
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene () | NA | NA | ||
| Strain, strain background () | C57Bl/6 (Mus musculus, male and female) | UTSW Breeding Core | NA | |
| Strain, strain background () | Ccr5-/-(Mouse, male and female) | Jackson Laboratories | Stock #: 005427 | |
| Genetic reagent () | NA | |||
| Cell line () | HEK-293 cells (Homo Sapiens) | ATCC | ATCC CRL-3216 | |
| Cell line () | SV589 cells (human) | NIGMS Human Genetic Mutant Cell Repository | NA | Discontinued for distribution |
| Transfected construct () | pCCR5-Myc (human) | This paper | NCBI Reference Sequence: NM_000579.3 | Encodes full length human CCR5 followed by five tandem repeats of the Myc epitope tag. |
| Transfected construct () | pβ2AR-SNAP; pCCR1-SNAP; pCCR4-SNAP; pCCR5-SNAP; pCCR10-SNAP; and pMAS1-SNAP (human) | This paper | NCBI Reference Sequence for β2AR, CCR1, CCR4, CCR5, CCR10 and MAS1 are NM_000024.5, NM_001295.3, NM_005508.4, NM_000579.3, NM_016602.3 and NM_002377.3, respectively. | Encode indicated full length human GPCRs followed by a C-terminal SNAP-tag |
| Transfected construct () | pSNAP-CCR5 (human) | Cisbio | Cat#PSNAPCCR5 | |
| Biological sample () | NA | |||
| Antibody | IgG-9E10 | ATCC | ATCC CRL-1729 | 0.5 µg/ml for immunoblot analysis, 3 µg/ml for immunofluorescent microscopy |
| Antibody | Human CCR5 Antibody, 45531 | R and D Systems | Cat#MAB182-100 | 1 µg/ml |
| Antibody | Actin Antibody | Sigma Aldrich | Cat#A2066-100UL | 1:10,000 dilution |
| Antibody | Anti-SNAP-tag Antibody | New England Biolabs | Cat#P9310S | 1:1000 dilution |
| Recombinant DNA reagent | NA | |||
| Sequence-based reagent | NA | |||
| Peptide, recombinant protein | Recombinant Human CCL5/RANTES Protein | R and D Systems | Cat#278-RN-050 | |
| Commercial assay or kit | CLIP-Surface Starter Kit | New England Biolabs | Cat#E9230S | |
| Commercial assay or kit | SNAP-Lumi4-Tb | Cisbio | Cat#SSNPTBC | |
| Commercial assay or kit | Pierce Cell Surface Protein Isolation Kit | Thermo Fischer Scientific | Cat#89881 | |
| Commercial assay or kit | CytoSelect 96-Well Cell Migration Assay | Cell Biolabs, Inc. | CBA-105 | |
| Chemical compound, drug | Xtreme Gene HP DNA Transfection Reagent | Sigma Aldrich | Cat#6366244001 | |
| Chemical compound, drug | Saponin from quillaja bark | Sigma Aldrich | Cat#S4521-25G | |
| Chemical compound, drug | Lipopolysaccharides from Escherichia coli 0111:B4 | Sigma Aldrich | Cat#L3024-5MG | |
| Chemical compound, drug | Fumonisin B1 from Fusarium moniliforme | Sigma Aldrich | Cat#F1147-1MG | |
| Chemical compound, drug | N-Hexanoyl-D-sphingosine (C6-Ceramide) | Sigma Aldrich | Cat#H6524-1MG | |
| Software, algorithm | BLAST, blastp suite | NCBI | NA | |
| Other | NA |
Additional files
-
Transparent reporting form
- https://doi.org/10.7554/eLife.40234.016
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Regulating G protein-coupled receptors by topological inversion
eLife 8:e40234.
https://doi.org/10.7554/eLife.40234




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}