Comment on 'Naked mole-rat mortality rates defy Gompertzian laws by not increasing with age'
- University of Duisburg-Essen, Germany
- University Hospital, University of Duisburg-Essen, Germany
- Jena University Hospital, Germany
- Moscow Society of Naturalists, Russia
- Leibniz Institute on Aging – Fritz Lipmann Institute, Germany
- Leibniz Institute for Zoo and Wildlife Research, Germany
- Czech University of Life Sciences, Czech Republic
- Ulm University, Germany
Abstract
Ruby et al. recently analyzed historical lifespan data on more than 3200 naked mole-rats, collected over a total observation period of about 38 years (Ruby et al., 2018). They report that mortality hazards do not seem to increase across the full range of their so-far-observed lifespan, and conclude that this defiance of Gompertz's law ‘uniquely identifies the naked mole-rat as a non-aging mammal’. Here, we explain why we believe this conclusion is premature.
https://doi.org/10.7554/eLife.45415.001Introduction
The historical data set analyzed by Ruby et al. (2018) is strongly skewed toward animals born in 2008 or later (Figure 1); thus, the observation time for most (87%) of the animals is no longer than 8 years. The reason for this skew – a massive expansion of the colony after 2007 – has been diligently explained by Ruby et al. However, one fundamental consequence of this skew is that the true informative value of the dataset regarding the key claim – that naked mole-rats do not age across the full range of their potential lifespan – is much smaller than that intuitively perceived by the large number of data points (>3,000) in their data set.
Figure 1
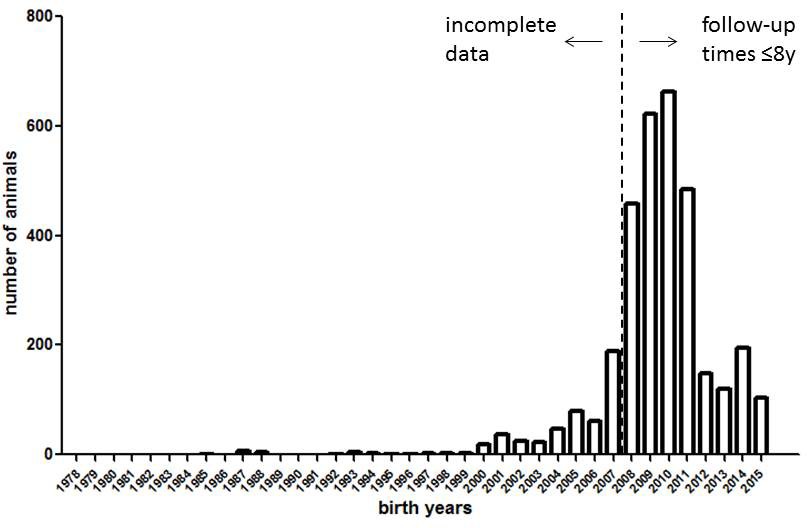
Histogram of birth years in the naked mole-rat dataset (3299 data points) underlying Figure 1 of Ruby et al. (2018).
https://doi.org/10.7554/eLife.45415.002Eight years correspond to approximately 25% of the maximum lifespan of the species as recorded to date (>33 years). In these young age cohorts, demographic aging (increasing mortality rates) is not expected to occur: as Ruby et al. correctly point out, Gompertz mortality acceleration typically has its onset in the second half of the potential lifespan of a mammalian species (see also Jones et al., 2014). Therefore, looking especially at older age cohorts is crucial for addressing the question of whether a species exhibits Gompertzian aging or not. If naked mole-rats were typical mammals, one would expect mortality acceleration to become apparent no earlier than at approximately 17 years (~50% of their maximum lifespan as determined to date), or even later if the mole-rats could indeed live considerably longer than has been recorded to date (as suggested by Ruby et al.). However, only 23 individuals (i.e. fewer than 1%) were observed for 18 years or longer in the most stringent analysis by Ruby et al. (their Figure 1), and all of them were subsequently right-censored. Consequently, Ruby et al. limit their conclusions to the first 18 years in some passages (‘The mortality hazard of naked mole-rats failed to increase for at least 18 years’), or the first 12 years in other passages (‘Our analyses […] confidently revealed a lack of demographic aging up to at least 4400 days of life (~12 years).’). Although we agree with this approach, we also believe that other statements (such as ‘this mouse-sized rodent exhibited no increase in mortality hazard, that is, no Gompertzian aging, across its full, as-yet-observed, multi-decade life-span’) are exaggerated at this point, because ‘full, as-yet-observed’ means more than 33 years.
Because of the small number of data points for mole-rats older than 18 years, Ruby et al. calculated one single hazard estimate for the entire final age group (>18 years). The fact that this hazard estimate was not higher than the hazard estimates for the preceding age groups is surely a strong argument for the interpretation put forward by Ruby et al. However, the decisive question of whether mortality hazards increase toward the end of that age group or not must, by definition, remain unanswered by this approach. Considering also the decrease in survival at approximately 29 years in Figure 2A of Ruby et al., and the unavoidably high statistical uncertainty toward the end of that final age group, we believe that the chance of not-yet-detected aging in later years – which has been acknowledged in principle by Ruby et al. – is too high to allow us to state at this stage that naked mole-rats are ‘non-aging mammals’.
Results and discussion
Bias due to missing death records
Working with the raw data of Ruby et al. revealed that for the observation period of July 2, 1978, through January 30, 2008, that is for nearly 30 years (~76% of the entire observation period), no deaths were reported. Because it is extremely unlikely that no animal died over such a long period of time, we contacted Ruby et al. and expressed our concern that the data set could be biased due to missing death records before 2008. They responded that indeed only animals that had survived after January 30, 2008, and those born after that date were included in the study. They further argued that additional (unpublished) left-censorship-analysis performed by them had satisfactorily controlled for the missing death records between 1978 and 2008, and that their conclusion about non-increasing hazard had not been modified by this alternative analytical approach (Ruby et al., personal communication).
We have not seen this analysis yet, but we doubt that such an approach could truly rescue the situation. Even methods for addressing left censoring require information on the number of missing animals. For the reader, it is hard to evaluate whether this information is available or not, and how much information can still be derived from it. In their supplemental data, Ruby et al. list 250 animals which could not be included in the study due to insufficient resolution (amongst them 37 animals labelled ‘death date unknown’), but the reader cannot determine whether these animals represent all missing animals that died between 1978 and January 2008, which age categories they might belong to and so on.
In our view, these aspects need to be addressed because selective reporting of death data can clearly bias all relevant hazard and Kaplan–Meier estimates (underestimation of hazard rates, overestimation of median survival). Since death events at ages up to 29 years may be missing, while no deaths occurring later than 6529 days (~18 years) are recorded in the most stringent analysis by Ruby et al. (their Figure 1A), the bias could be relevant, especially with regard to hazard estimates for older age groups.
We tried to exemplify the potential bias by modeling own lifespan data for related Fukomys mole-rats (Figure 2A) and using this dataset to ‘simulate’ the loss of death data over the first 75% (1984–2010) of the total observation period (1984–2018). The result of our simulation is depicted in Figure 2B. Not surprisingly, the slope of the survival curve decreases, median survival increases by more than 30%, and the estimated constant mortality hazard per day decreases by more than 25%, from 2.6 × 10−4 per day (Figure 3A) to 1.9 × 10−4 per day (Figure 3B). Although this is just a single example and both data sets may quantitatively react differently to the introduced bias, these results indicate that the ‘consistent estimate for mortality hazard (8 × 10−5 per day)’ for the naked mole-rat (Ruby et al., 2018) is probably underestimated. Hazards may remain constant with age, but even this possibility remains, in our view, unanswered to date.
Figure 2
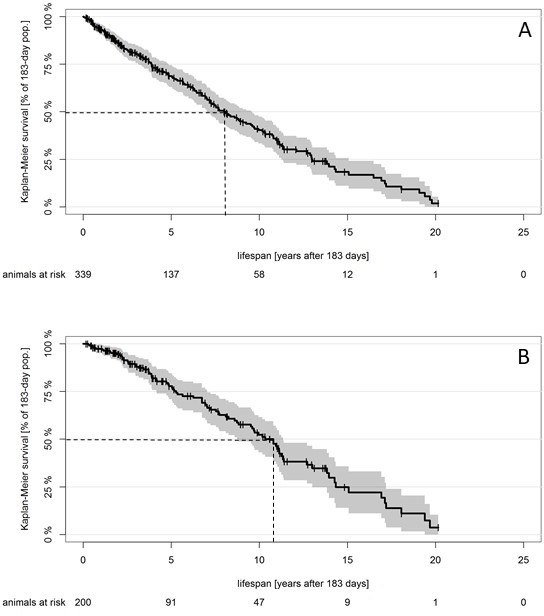
Survival curve for small Zambian mole-rats.
Kaplan–Meier survival curve for small Zambian mole-rats (Fukomys anselli and Fukomys anselli x kafuensis) that reach the same age as that used as a starting point in Ruby et al. (2018); 95% confidence intervals for the Kaplan–Maier curve and animals at risk are included, as suggested by Pocock et al. (2002). Dotted lines represent median survival after onset of the study at 0.5 years. (A) Original data from 339 animals; median survival after 0.5 years, 7.99 years (95% CI, 7.04–9.60 years). (B) Biased data presentation: animals that died before 2010 have been (artificially) deleted from the dataset. Median survival after 0.5 years, 10.80 years (95% CI, 8.82–11.39 years).
-
Figure 2—source code 1
This script reads the data from Figure 2—source data 1 to create the reported Kaplan–Meier estimators (Figure 2A,B).
- https://doi.org/10.7554/eLife.45415.004
-
Figure 2—source data 1
This xlxs-file contains the lifespan data for small Zambian Fukomys-mole rats that underlie Figure 2A,B.
Each row in the file contains data for one individual animal. Columns inform about individual animal IDs, birth date/year, sex, death date, the date on which data for that animal was compiled ('DataDate'), censorship, lifespan (in days) and additional notes whenever specific circumstances for that specific animal had to be reported. 'DeathDate' is empty if the animal was still alive at the time of data compilation ('DataDate'). In such cases, 'DataDate' was used as the date-of-censorship.
- https://doi.org/10.7554/eLife.45415.005
Homologies to other social mole-rats
Of note, we believe that Figure 3 of Ruby et al. contains an important finding because it shows for the first time that, under laboratory conditions, naked mole-rat breeders appear to live longer than helpers. Until recently, only data from the wild suggested this difference in life expectancy for naked mole-rats (Braude, 1991; Buffenstein, 2008). If corroborated by a stricter, unbiased analysis, this pattern would show a striking similarity to the divergent survival trajectories of breeders and non-breeders in several Fukomys species (Dammann and Burda, 2006; Dammann et al., 2011; Schmidt et al., 2013) and thus would support the hypothesis that many if not all social mole-rat species may have evolved life-prolonging mechanisms associated with sexual activity, breeding, or both, despite the classic trade-off between reproduction and somatic maintenance.
Conclusions
In summary, we argue that the lifespan data of Ruby et al. (2018) have to be used and interpreted with caution because they are skewed and biased. Even if naked mole-rats defy the Gompertz–Makeham timeline over large portions of their documented lifespan, they may nonetheless age afterwards (see, for example, Edrey et al., 2011 for signs of age-associated pathologies in naked mole-rats older than 28 years). In fact, the scientific literature regarding naked mole-rats describes a number of aging phenotypes for these mammals (see Edrey et al., 2011; Beltrán-Sánchez and Finch, 2018; Finch, 2009; Heinze et al., 2018 and references therein); these phenotypes should be taken into account in discussions concerning whether naked mole-rats are non-aging mammals.
Data availability
All data generated or analysed during this study are included in the manuscript and supporting files. Source data files have been provided for Figure 2.
References
-
ThesisThe Behavior and Demographics of the Naked Mole-Rat, Heterocephalus glaberUniversity of Michigan, Ann Arbor.
-
Negligible senescence in the longest living rodent, the naked mole-rat: insights from a successfully aging speciesJournal of Comparative Physiology B 178:439–445.https://doi.org/10.1007/s00360-007-0237-5
-
Sexual activity and reproduction delay ageing in a mammalCurrent Biology 16:R117–R118.https://doi.org/10.1016/j.cub.2006.02.012
Article and author information
Author details
Thomas Hildebrandt

Funding
Deutsche Forschungsgemeinschaft (DA 992/3-1)
- Philip Dammann
Deutsche Forschungsgemeinschaft (PL 173/8-1)
- Matthias Platzer
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Copyright
© 2019, Dammann et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 2,180
- views
-
- 144
- downloads
-
- 22
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 22
- citations for umbrella DOI https://doi.org/10.7554/eLife.45415
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Comment on 'Naked mole-rat mortality rates defy Gompertzian laws by not increasing with age'
eLife 8:e45415.
https://doi.org/10.7554/eLife.45415
Further reading
-
- Ecology
The longest-lived rodent, the naked mole-rat (Heterocephalus glaber), has a reported maximum lifespan of >30 years and exhibits delayed and/or attenuated age-associated physiological declines. We questioned whether these mouse-sized, eusocial rodents conform to Gompertzian mortality laws by experiencing an exponentially increasing risk of death as they get older. We compiled and analyzed a large compendium of historical naked mole-rat lifespan data with >3000 data points. Kaplan-Meier analyses revealed a substantial portion of the population to have survived at 30 years of age. Moreover, unlike all other mammals studied to date, and regardless of sex or breeding-status, the age-specific hazard of mortality did not increase with age, even at ages 25-fold past their time to reproductive maturity. This absence of hazard increase with age, in defiance of Gompertz’s law, uniquely identifies the naked mole-rat as a non-aging mammal, confirming its status as an exceptional model for biogerontology.




{kind=link}
{kind=link}