Evolution of (p)ppGpp-HPRT regulation through diversification of an allosteric oligomeric interaction
- University of Wisconsin, United States
Abstract
The alarmone (p)ppGpp regulates diverse targets, yet its target specificity and evolution remain poorly understood. Here, we elucidate the mechanism by which basal (p)ppGpp inhibits the purine salvage enzyme HPRT by sharing a conserved motif with its substrate PRPP. Intriguingly, HPRT regulation by (p)ppGpp varies across organisms and correlates with HPRT oligomeric forms. (p)ppGpp-sensitive HPRT exists as a PRPP-bound dimer or an apo- and (p)ppGpp-bound tetramer, where a dimer-dimer interface triggers allosteric structural rearrangements to enhance (p)ppGpp inhibition. Loss of this oligomeric interface results in weakened (p)ppGpp regulation. Our results reveal an evolutionary principle whereby protein oligomerization allows evolutionary change to accumulate away from a conserved binding pocket to allosterically alter specificity of ligand interaction. This principle also explains how another (p)ppGpp target GMK is variably regulated across species. Since most ligands bind near protein interfaces, we propose that this principle extends to many other protein–ligand interactions.
https://doi.org/10.7554/eLife.47534.001Introduction
Regulation by signaling ligands is a universal mechanism for adaptation and homeostasis (Chubukov et al., 2014; Traut, 2008). How proteins evolve to be differentially regulated by their signaling ligands is under ongoing investigation (Najmanovich, 2017; Taute et al., 2014). A key signaling ligand in bacteria is the nucleotide (p)ppGpp. (p)ppGpp directly binds and regulates diverse protein targets at positions ranging from protein active sites to protein–protein interfaces (Corrigan et al., 2016; Gourse et al., 2018; Liu et al., 2015a; Sherlock et al., 2018; Wang et al., 2019; Zhang et al., 2018). However, how these diverse themes evolve across bacterial species have not been systematically elucidated. In addition, while most of the (p)ppGpp regulation is studied for high (p)ppGpp concentrations upon stress and starvation (Cashel et al., 1996; Gourse et al., 2018; Liu et al., 2015a), (p)ppGpp at basal levels also has important protective roles (Gaca et al., 2013; Gaca et al., 2015a; Kriel et al., 2012; Potrykus et al., 2011; Puszynska and O'Shea, 2017). However, molecular targets and mechanisms of basal (p)ppGpp regulation have not been delineated.
Here, we examined the specificity of (p)ppGpp binding by characterizing (p)ppGpp regulation of the enzyme hypoxanthine phosphoribosyltransferase (HPRT), one of the earliest identified targets of (p)ppGpp (Hochstadt-Ozer and Cashel, 1972; Kriel et al., 2012). Using biochemical, structural and evolutionary analyses of HPRT homologs across diverse bacterial phyla, we identify a conserved (p)ppGpp binding motif and identified a HPRT dimer–dimer interaction that allosterically positions a flexible loop to promote strong (p)ppGpp binding. The bacterial HPRT homologs lacking this dimer–dimer interaction are largely refractory to (p)ppGpp regulation. We propose that changes in protein oligomerization enable evolution of regulation by (p)ppGpp, despite evolutionary constraint imposed on the direct (p)ppGpp-binding site. We examined this principle using another (p)ppGpp target GMK and discuss its broad implications in evolutionary diversification of signaling ligand specificity.
Results
Regulation of HPRT activity by basal levels of (p)ppGpp is important for GTP homeostasis
HPRT is a purine salvage enzyme that converts purine bases and PRPP to GMP and IMP, which are precursors of the essential nucleotide GTP (Figure 1A, upper panel). HPRT activity was previously found to be inhibited by (p)ppGpp in several bacteria including Bacillus subtilis (Gaca et al., 2015b; Hochstadt-Ozer and Cashel, 1972; Kriel et al., 2012). In B. subtilis, (p)ppGpp ranges from <30 μM at basal concentrations to millimolar levels upon starvation. (p)ppGpp inhibits B. subtilis HPRT activity at an IC50 of ≈10 μM, allowing (p)ppGpp to regulate purine salvage at its basal concentrations (Figure 1B). The absence of (p)ppGpp in B. subtilis results in an uncontrolled GTP increase and toxicity upon guanine addition (Kriel et al., 2012) (Figure 1A, middle panel).
Figure 1
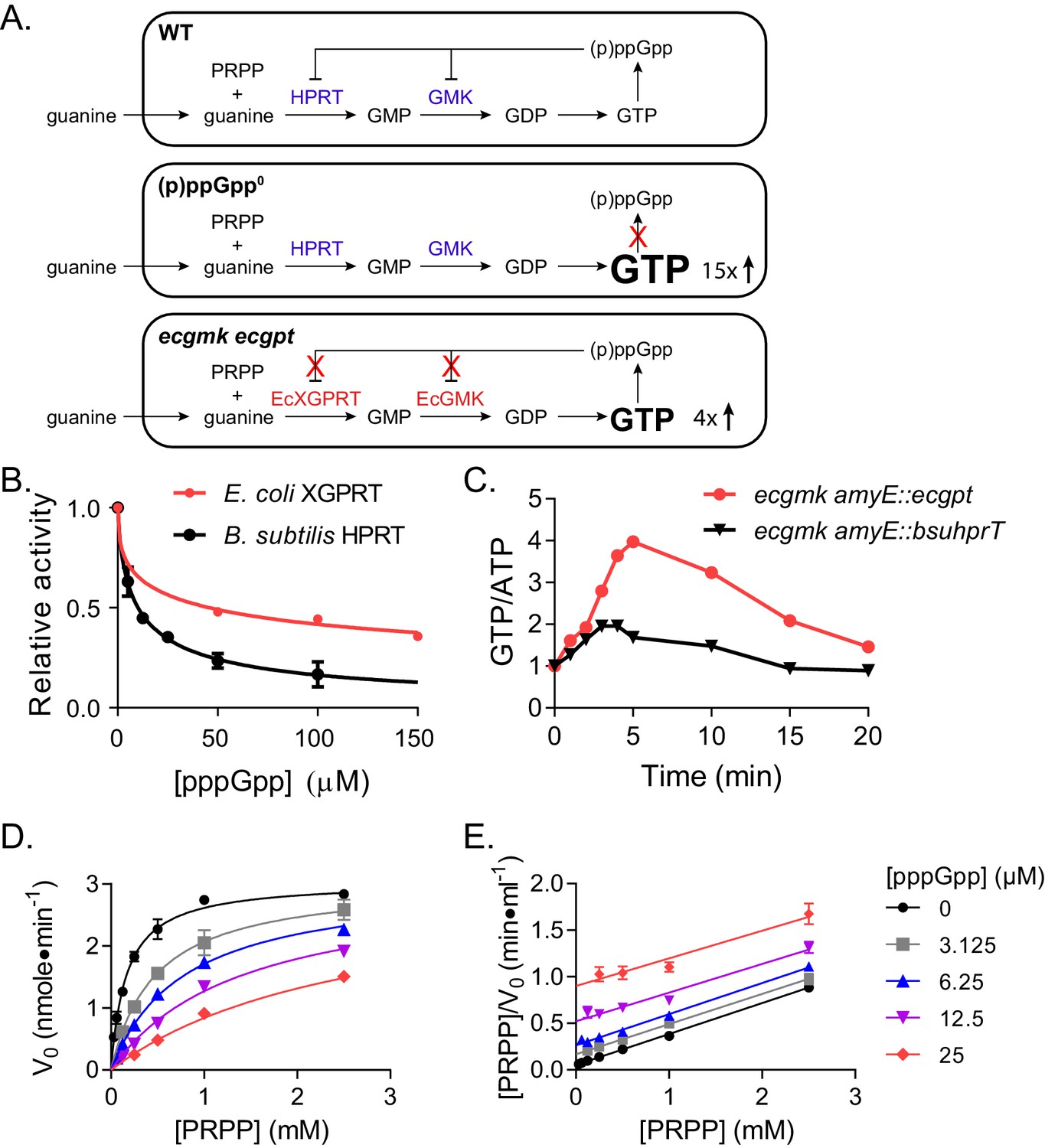
Regulation of HPRT by basal levels of (p)ppGpp is important for GTP homeostasis.
(A) Pathways showing the effect of extracellular guanine on B. subtilis GTP homeostasis. In WT, (p)ppGpp regulates HPRT and GMK. (p)ppGpp0 cannot produce (p)ppGpp (see B) and ecgmk ecgpt has enzymes resistant to (p)ppGpp regulation (see F). (B) E. coli XGPRT more weakly inhibited by pppGpp than B. subtilis HPRT. The IC50 for E. coli XGPRT is 45 µM compared to 10 µM for B. subtilis HPRT. Error bars represent SEM of triplicate. (C) Expression of E. coli XGPRT leads to imbalanced GTP/ATP homeostasis in B. subtilis. GTP/ATP ratio in B. subtilis treated with 1 mM guanosine as determined by thin layer chromatography of 32P-labeled cells. Time is minutes after guanosine treatment. In ecgmk, (p)ppGpp-insensitive E. coli gmk replaces B. subtilis gmk at its endogenous locus. ecgmk amyE::bsuhprT and ecgmk amyE::ecgpt express B. subtilis hprT and E. coli gpt, respectively, from an IPTG-dependent promoter at an exogenous locus. All strains grown with 1 mM IPTG. (D) Initial velocities of B. subtilis HPRT at varied PRPP and pppGpp concentrations. Data are fitted to a global competitive inhibition equation (r2 = 0.975), and Ki = 1.7 µM. (E) Data from (D) in a Hanes–Woolf transformation. Parallel lines indicate equal maximum velocities for each pppGpp concentration. Error bars represent SEM of at least three replicates.
To demonstrate the physiological relevance of basal (p)ppGpp regulation of HPRT, we introduced the E. coli enzyme XGPRT (ecgpt) into B. subtilis. E. coli XGPRT produces GMP similarly to B. subtilis HPRT but is only modestly inhibited by (p)ppGpp at induced concentrations (Figure 1B). To visualize the effect of this regulation on GTP homeostasis, we also introduced a (p)ppGpp-resistant E. coli GMK to replace the (p)ppGpp-sensitive B. subtilis GMK, allowing the subsequent conversion of GMP to GDP to bypass (p)ppGpp regulation (Figure 1A, bottom panel). When these strains were treated with guanosine, GTP levels increased far more in cells with the modestly regulated ecgpt than in cells with the basal (p)ppGpp-regulated B. subtilis hprT (Figure 1C), demonstrating that basal regulation of HPRT by (p)ppGpp is important for preventing strong fluctuations in GTP.
We performed Michaelis Menten kinetics on B. subtilis HPRT and found (p)ppGpp is a strong competitive inhibitor of its substrate PRPP (Figure 1D–E). The Ki for pppGpp is 1.7 μM, while the Km for PRPP is 166 μM, allowing (p)ppGpp at a basal level to effectively compete with physiological concentrations (0.1–1 mM) of PRPP (Bennett et al., 2009; Hove-Jensen et al., 2017) and keep GTP under surveillance not just during starvation, but also during unstarved growth.
Conservation and variation of (p)ppGpp regulation of HPRT across species
It has been shown for the (p)ppGpp targets RNA polymerase and guanylate kinase that conservation of their binding to (p)ppGpp is limited within distinct bacterial phyla (Liu et al., 2015b; Ross et al., 2016). We therefore tested the conservation of HPRT regulation by conducting a broad biochemical survey of HPRT enzymes from free-living bacteria, human commensals, and pathogens. We revealed (p)ppGpp can bind and inhibit activities of HPRTs from every phylum of bacteria we tested, including Actinobacteria, Bacteroidetes, Firmicutes, Deinococcus-Thermus, and Proteobacteria, and even eukaryotic HPRTs (Figure 2). To do so, we purified 32 recombinant HPRTs and tested their enzymatic activities and found that most of them were strongly inhibited by ppGpp and pppGpp at 25 μM (Figure 2A–B and Table 1). Considering that (p)ppGpp is induced to millimolar concentrations during starvation and its uninduced levels are ≈10–30 μM during exponential growth in B. subtilis and E. coli (Cashel et al., 1996; Kriel et al., 2012), these results suggest significant inhibition of HPRT under physiological, basal levels of (p)ppGpp.
Figure 2 with 1 supplement see all

Conservation and variation in (p)ppGpp regulation of HPRTs across species.
(A–B) (p)ppGpp inhibits HPRTs across species. Representative phylogenetic tree constructed from 16S rRNA sequences of species shown with eukaryotic outgroup hidden. Branch length scale represents 0.5 substitutions per site. See Materials and methods for tree construction. (A) Relative activities of HPRTs with 25 μM ppGpp. (B) Relative activities of HPRTs with 25 μM pppGpp. Error bars represent SEM of triplicates. (C–D) pppGpp binds HPRTs across species. (C) Representative DRaCALA between B. anthracis Hpt-1 in cell lysate serially diluted 1:2 and 32P-labeled pppGpp. The central signal is proportional to pppGpp – HPRT interaction. Binding isotherm from isothermal titration calorimetry between B. anthracis Hpt-1 and pppGpp. See Figure 2—figure supplement 1 for energy isotherm and parameters. (D) The Kd between pppGpp and HPRTs obtained with DRaCALA from serially diluted cell lysates containing overexpressed HPRTs (see Materials and methods). Error bars represent SEM derived from one binding curve. P. phosphoreum and C. burnetii HPRTs with * have affinities too weak to calculate but are estimated to be >15 μM (see Figure 2—figure supplement 1). (E) Relative activities of HPRTs with increasing concentrations of pppGpp. See Table 1 for partial datasets for HPRTs from M. tuberculosis, C. crescentus, S. meliloti, R. torques, S. mutans, and C. gilvus.
Table 1
(p)ppGpp binding and inhibition of HPRT homologs.
https://doi.org/10.7554/eLife.47534.005| Organism | Relative activityd | Kd (pppGpp) (μM) | IC50 (pppGpp) (μM) | |
|---|---|---|---|---|
| 25 μM ppGpp | 25 μM pppGpp | |||
| Actinobacteria | ||||
| Streptomyces coelicolor | −0.01 ± 0.01 | 0.12 ± 0.01 | 1.77 ± 0.31 | 3.8 ± 0.2 |
| Collinsella aerofaciens | 0.31 ± 0.04 | 0.42 ± 0.05 | 1.46 ± 0.56 | |
| Cellulomonas gilvus | NAb | NAb | 0.6 ± 0.11 | |
| Mycobacterium tuberculosis | NAa | NAa | NAc | |
| Bacteroidetes | ||||
| Bacteroides intestinalis | 0.49 ± 0.06 | 0.05 ± 0.01 | 0.11 ± 0.03 | |
| Bacteroides caccae | 0.34 ± 0.03 | 0.01 ± 0.02 | 0.11 ± 0.02 | |
| Bacteroides ovatus | 0.43 ± 0.04 | −0.03 ± 0.02 | 0.15 ± 0.04 | |
| Bacteroides uniformis | 0.4 ± 0.01 | 0.01 ± 0.01 | 0.1 ± 0.04 | |
| Bacteroides thetaiotaomicron | 0.41 ± 0 | 0 ± 0.01 | 0.11 ± 0.04 | |
| Bacteroides finegoldii | 0.46 ± 0.03 | −0.01 ± 0.01 | 0.15 ± 0.06 | |
| Firmicutes | ||||
| Eubacterium ventriosum | −0.03 ± 0.03 | −0.07 ± 0.03 | 0.66 ± 0.1 | |
| Ruminococcus lactaris | −0.01 ± 0 | 0 ± 0 | 0.11 ± 0.03 | |
| Ruminococcus torques | NAb | NAb | 0.22 ± 0.07 | |
| Clostridium leptum | −0.01 ± 0.01 | 0 ± 0 | 3.29 ± 0.94 | |
| Eubacterium eligens | −0.03 ± 0.03 | 0.02 ± 0.01 | 0.79 ± 0.18 | |
| Blautia hansenii | 0.07 ± 0 | 0.14 ± 0.01 | 0.83 ± 0.14 | |
| Eubacterium hallii | 0.17 ± 0.05 | −0.06 ± 0.05 | 0.07 ± 0.02 | |
| Clostridium spiroforme | 0.07 ± 0.01 | 0.17 ± 0 | 9.54 ± 1.17 | |
| Listeria monocytogenes | 0.13 ± 0 | 0.32 ± 0.02 | 0.32 ± 0.06 | |
| Bacillus subtilis | 0.08 ± 0.01 | 0.41 ± 0.01 | 0.75 ± 0.07 | 10.1 ± 1.3 |
| Staphylococcus aureus | 0.31 ± 0.01 | 0.48 ± 0.01 | 2.83 ± 0.38 | 10.6 ± 0.3 |
| Streptococcus mutans | NAb | NAb | 5.12 ± 1.45 | |
| Bacillus anthracis 1 | 0.25 ± 0.01 | 0.5 ± 0.01 | 1.54 ± 0.18 | 29.5 ± 1.2 |
| Enterococcus faecalis | 0.31 ± 0.01 | 0.49 ± 0.02 | 1.12 ± 0.11 | |
| Holdemania filiformis | 0.66 ± 0.01 | 0.43 ± 0 | 0.17 ± 0.11 | |
| Bacillus anthracis 2 | 0.81 ± 0.04 | 0.7 ± 0.05 | 6.18 ± 1.77 | |
| Deinococcus-Thermus | ||||
| Thermus thermophilus | 0.52 ± 0.07 | −0.01 ± 0.01 | 0.04 ± 0.01 | |
| Proteobacteria | ||||
| Caulobacter crescentus | 0.33 ± 0.05 | 0.47 ± 0.07 | 0.24 ± 0.09 | |
| Escherichia coli | 0.33 ± 0.01 | 0.48 ± 0.02 | 3.38 ± 1.47 | 26.2 ± 2.0 |
| Klebsiella pneumoniae | 0.47 ± 0.03 | 0.49 ± 0.01 | 3.38 ± 0.89 | |
| Sinorhizobium meliloti | NAb | NAb | 3.08 ± 0.92 | |
| Yersinia enterocolitica | 0.7 ± 0.01 | 0.7 ± 0.03 | 4.44 ± 1.25 | |
| Photobacterium phosphoreum | 0.97 ± 0.01 | 0.96 ± 0.03 | NAc | >100 |
| Pseudomonas syringae | 0.6 ± 0.01 | 0.6 ± 0.01 | 5.85 ± 1.76 | |
| Pseudomonas aeruginosa | 0.84 ± 0.01 | 0.83 ± 0.02 | 5.28 ± 1.52 | |
| Legionella pneumophila | 0.78 ± 0.01 | 0.69 ± 0.03 | 1.72 ± 0.38 | >100 |
| Coxiella burnetii | 0.79 ± 0.14 | 0.72 ± 0.16 | NAc | >100 |
| Neisseria meningitidis | 0.9 ± 0.02 | 0.8 ± 0.03 | 1.64 ± 0.55 | |
| Eukarya | ||||
| Saccharomyces cerevisiae | 0.64 ± 0.02 | 0.96 ± 0.01 | 8.08 ± 8.58 | |
| Caenorhabditis elegans | NAb | NAb | 12.77 ± 7.68 | 29.6 ± 2.8 |
| Homo sapiens | NA | NA | NA | 66.6 ± 5.4 |
-
± standard error of the mean.
NA = no data obtained.
-
aProtein not purified.
bProtein did not have activity.
-
cInteraction with pppGpp too weak to estimate Kd.
dRelative to activity without ppGpp or pppGpp.
We also quantified the HPRT homologs’ binding affinity (Kd) for pppGpp with differential radial capillary action of ligand assay (DRaCALA) (Figure 2C–D) (Roelofs et al., 2011), results of which were highly comparable with well-established quantitative methods such as isothermal titration calorimetry (Figure 2C and Figure 2—figure supplement 1). Using this method, we found that most HPRTs bind pppGpp with Kd values ranging from ≈0.1 to ≈10 μM (Figure 2D and Table 1).
Closer inspection revealed an additional quantitative difference in regulatory potency among these HPRTs. HPRTs from human microbiota commensals, including all species from Bacteroidetes along with the Firmicutes Eubacterium spp. and Ruminococcus spp., manifested the tightest interactions with pppGpp (Kd ≈ 0.1–1 μM) (Figure 2D and Table 1). This suggests a relationship between the intestinal environment and regulation of this purine salvage enzyme by (p)ppGpp, which may serve to buffer the intracellular environment against fluctuating extracellular purine concentrations. Beyond microbiota, HPRTs from soil-dwelling bacteria (e.g. Streptomyces coelicolor, B. subtilis) and pathogens (e.g. Bacillus anthracis, Staphylococcus aureus, E. coli), occupying the phyla Actinobacteria, Firmicutes and Proteobacteria, were also inhibited by (p)ppGpp with Kd values below 10 μM.
Interestingly, a few HPRTs were only weakly inhibited by 25 μM (p)ppGpp (Figure 2A–B). These weakly regulated HPRTs all clustered in β- and γ-Proteobacteria (e.g. Pseudomonas aeruginosa and Neisseria meningitidis), with the exception of Mycobacterium tuberculosis (Figure 2—figure supplement 1C). To test whether the HPRTs weakly inhibited at basal levels of (p)ppGpp (25 μM) can be inhibited by higher levels of (p)ppGpp, we examined select HPRT homologs at a range of pppGpp concentrations and at physiological substrate concentrations (1 mM PRPP and 50 μM guanine) (Bennett et al., 2009) (Figure 2E). Most bacterial HPRTs (e.g. B. subtilis, E. coli, S. aureus) were sensitive to basal pppGpp. However, several bacterial HPRTs that were weakly inhibited by 25 μM pppGpp (e.g. C. burnetii, L. pneumophila) are also almost refractory to high concentration of pppGpp. In fact, they are less inhibited by pppGpp than eukaryotic HPRTs from human and C. elegans, for which (p)ppGpp is not a physiological regulatory ligand. Therefore, we can broadly classify bacterial HPRTs by their sensitivity to (p)ppGpp: those that are sensitive to basal (p)ppGpp and a few that are resistant to basal (p)ppGpp, although may still be mildly sensitive to induced (p)ppGpp.
In addition to differences between HPRT homologs, we also noticed a strong, species-dependent difference between pppGpp and ppGpp inhibition. All HPRTs in Bacteroidetes were potently inhibited by pppGpp but only weakly by ppGpp (Figure 2A–B). In contrast, nearly all other HPRTs displayed stronger inhibition by ppGpp than pppGpp. It is noted that Bacteroides spp., whose HPRTs are more sensitive to pppGpp than ppGpp, contain a GppA homolog, and they make mostly ppGpp in vivo (Glass et al., 1979). On the other hand, Firmicutes, whose HPRTs are more sensitive to ppGpp than pppGpp, lack GppA homologs and produce predominantly pppGpp. Therefore, although pppGpp and ppGpp are often regarded as similar, they can have marked differences for certain cellular targets.
(p)ppGpp binds the conserved active site of HPRT and closely mimics substrate binding
To examine the molecular determinants underlying (p)ppGpp regulation of HPRT, we turned to Hpt-1 from the pathogenic bacterium B. anthracis, which pppGpp competitively inhibits similarly to B. subtilis HPRT (Figure 3—figure supplement 1). We crystallized Hpt-1 with and without ppGpp (Figure 3A and Table 2), and for comparison, we also crystallized Hpt-1 with its two substrates, PRPP and the non-reactive guanine analog 9-deazaguanine (Figure 3B and Table 2) (Héroux et al., 2000). Apo HPRT diffracted to 2.06 Å resolution with four molecules in the asymmetric unit and was nearly identical to a deposited apo structure of B. anthracis Hpt-1 (PDB ID 3H83) (Figure 3—figure supplement 2). The HPRT-substrates structure diffracted to 1.64 Å containing two molecules in the asymmetric unit (Figure 3—figure supplement 2), and PRPP and 9-deazaguanine were coordinated by two Mg2+ ions with each monomer (Figure 3B and Figure 3—figure supplement 3). The HPRT-ppGpp complex diffracted to 2.1 Å resolution with two molecules in the asymmetric unit (Figure 3—figure supplement 2), and each monomer contained one ppGpp coordinated with Mg2+, in agreement with near 1:1 stoichiometry measured via ITC (Figure 2—figure supplement 1 and Figure 3—figure supplement 3). These crystals formed in drops containing pppGpp, but there was insufficient density to completely model the 5′ γ-phosphate (Figure 3—figure supplement 4). With LC-MS/MS, we verified that the pppGpp was not contaminated with ppGpp. Although it is possible that the γ-phosphate was hydrolyzed during crystallization, the presence of waters and unassigned density around the 5′ phosphates allow for the possibility that the γ-phosphate is present but dynamic.
Figure 3 with 8 supplements see all
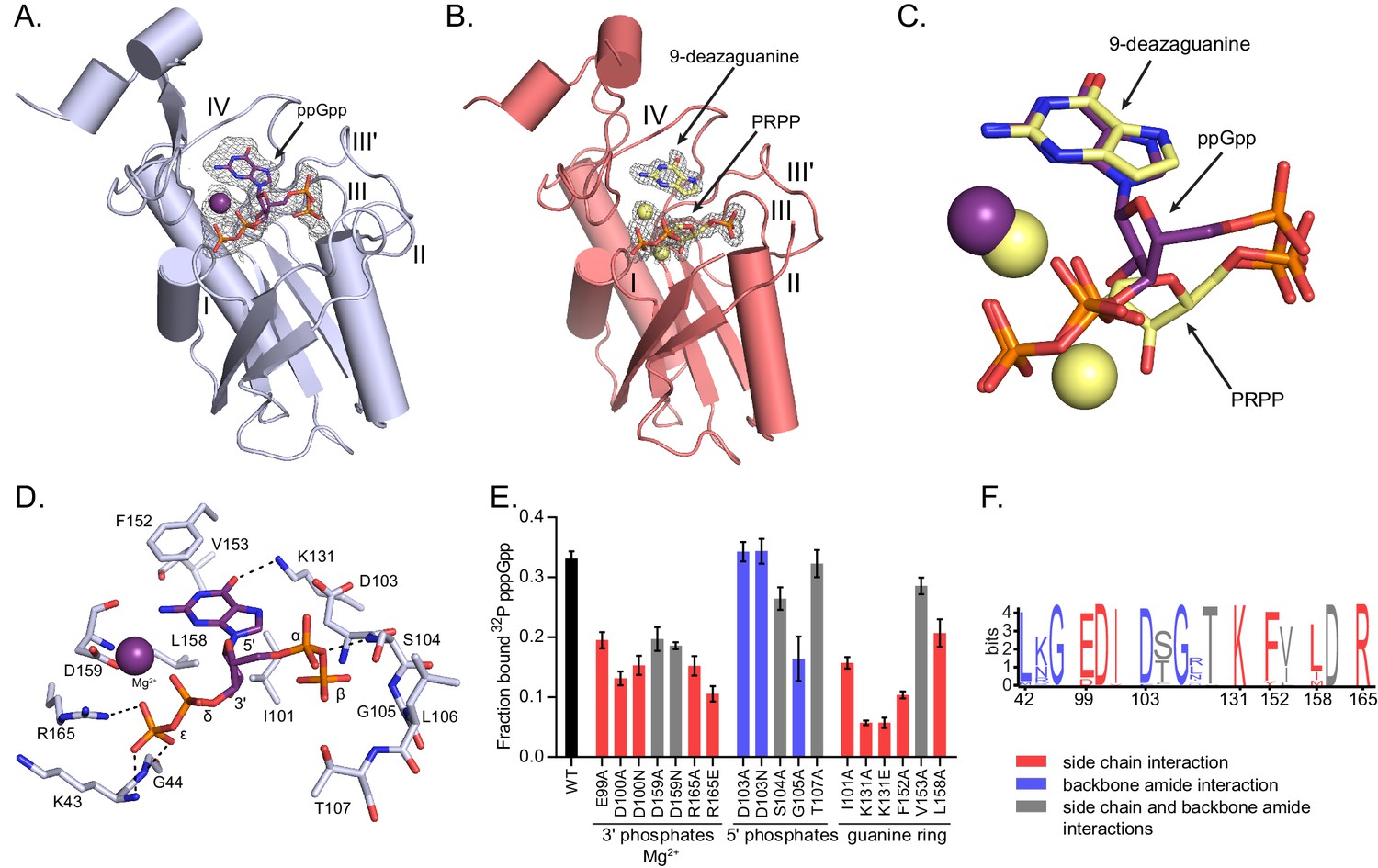
(p)ppGpp binds the HPRT active site and shares an almost identical binding pocket with substrates.
(A) B. anthracis Hpt-1 crystallized with ppGpp. Five loops (I, II, III, III′, IV) form the HPRT active site (Sinha and Smith, 2001). ppGpp and Mg2+ shown with omit electron density contoured at 2σ. Figure 3—figure supplement 1 shows that pppGpp competitively inhibits Hpt-1. Figure 3—figure supplement 4 shows omit electron density for the two ppGpp molecules crystallized in the asymmetric unit. (B) B. anthracis Hpt-1 crystallized with 9-deazaguanine, PRPP, and two Mg2+. Ligands shown with omit electron density contoured at 2.5σ. Residues Tyr70 – Ser76 in loop II were not resolved. See Figure 3—figure supplement 2 for asymmetric units of Hpt-1 crystallized with ppGpp, substrates, and sulfates. See Figure 3—figure supplement 3 for coordination of Mg2+ with ppGpp and PRPP. (C) Overlay of substrates (PRPP and 9-deazaguanine; yellow) and inhibitor (ppGpp; purple) bound to HPRT. Spheres represent Mg2+ and are colored according to their coordinating ligand. See Figure 3—figure supplement 6 for binding pocket comparison. (D) The ppGpp binding pocket on HPRT. Black dotted lines indicate select hydrogen bonds. The peptide backbone is shown for residues where the interactions are relevant. See Figure 3—figure supplement 5 for complete interaction maps. (E) DRaCALA of B. subtilis HPRT variants binding to 32P-labeled pppGpp, determined with cell lysates containing overexpressed HPRT variants. Disruption of side chain interactions weakened binding. For E and F, the residues are colored according to their interaction with ppGpp. Red = side chain interaction, blue = backbone amide interaction, and gray = both side chain and backbone interactions. Error bars represent SEM of three replicates. (F) Sequence frequency logo of the (p)ppGpp binding pocket from 99 bacterial HPRTs. Logo created using WebLogo from UC-Berkeley. See Figure 3—figure supplement 8 for binding residues from select bacterial and eukaryotic HPRTs.
Table 2
X-ray data collection and structure determination statistics.
https://doi.org/10.7554/eLife.47534.015| Data collection | |||
|---|---|---|---|
| Structure | HPRT with sulfates (PDB 6D9Q) | HPRT with substrates (PDB 6D9R) | HPRT with ppGpp (PDB 6D9S) |
| Wavelength | 0.9786 | 0.97872 | 0.9786 |
| Resolution range (highest resolution bin) (Å) | 41.3–2.06 (2.13–2.06) | 50–1.64 (1.67–1.64) | 40.2–2.11 (2.18–2.11) |
| Space group | P 31 2 1 | P 32 2 1 | P 31 2 1 |
| Unit cell | |||
| a, b, c (Å) | 82.597, 82.597, 242.416 | 113.758, 113.758, 56.731 | 82.61, 82.61, 174.92 |
| α, β, γ (°) | 90, 90, 120 | 90, 90, 120 | 90, 90, 120 |
| Completeness (%) | 99.16 (95.52) | 99.8 (99.0) | 98.92 (94.51) |
| Unique reflections | 60264 | 51750 | 40413 |
| Redundancy | 10.8 (6.4) | 21.5 (14.4) | 8.1 (6.2) |
| I/σI | 24.8 (1.74) | 31.09 (2.93) | 16.41 (1.22) |
| Rmeas | 0.124 | 0.137 (1.376) | 0.164 |
| Rpim | 0.037 | 0.029 (0.35) | 0.06 |
| CC 1/2 | (0.791) | (0.81) | (0.724) |
| Refinement | |||
| Resolution range (highest resolution bin) (Å) | 41.3–2.06 (2.11–2.06) | 37.19–1.64 (1.70–1.64) | 40.2–2.11 (2.16–2.11) |
| Rwork/Rfree a (%) | 18.2/21.7 | 16.5/19.6 | 20.2/24.4 |
| r.m.s.b deviations | |||
| Bonds (Å) | 0.004 | 0.01 | 0.003 |
| Angles (Å) | 1.03 | 1.41 | 0.812 |
| Ramachandran statistics (%) | |||
| Favored | 97.90 | 98.52 | 96.63 |
| Allowed | 2.10 | 1.48 | 3.37 |
| Disallowed | 0.00 | 0.00 | 0.00 |
| Rotamer outliers (%) | 0.78 | 0.30 | 0.63 |
| No. of atoms | |||
| Macromolecules | 5742 | 2881 | 2837 |
| Ligands | 46 | 161 | 116 |
| Solvent | 440 | 317 | 118 |
| B factor (Å2) | |||
| Macromolecules | 50.63 | 24.06 | 76.03 |
| Ligands | 68.83 | 37.76 | 161.17 |
| Solvent | 52.15 | 35.53 | 70.22 |
-
aRwork/Rfree = Σ||Fobs|-|Fcalc||/|Fobs|, where the working the free R factors are calculated by using the working and free reflection sets, respectively. The free R reflections were held aside throughout refinement. bRoot mean square. See Table 2—source datas 1–3 for PDB validation reports.
-
Table 2—source data 1
Validation report for PDB ID 6D9Q.
- https://doi.org/10.7554/eLife.47534.016
-
Table 2—source data 2
Validation report for PDB ID 6D9R.
- https://doi.org/10.7554/eLife.47534.017
-
Table 2—source data 3
Validation report for PDB ID 6D9S.
- https://doi.org/10.7554/eLife.47534.018
Comparison between the HPRT-ppGpp and HPRT-substrates structures revealed that ppGpp binds the HPRT active site and closely mimics the conformation of the two substrates (Figure 3C and Figure 3—figure supplements 5 and 6). The purine ring of ppGpp overlaps with the purine base substrate, and the two phosphate arms of ppGpp and PRPP both spread across the active site between loop I and loop III (Figure 3C and Figure 3—figure supplement 6). Generally speaking, in ppGpp–protein interactions, the phosphates of ppGpp are either elongated or compacted in a ring-like conformation (Steinchen and Bange, 2016). With the phosphates of ppGpp spread across the binding pocket, the HPRT-ppGpp interaction represents an elongated conformation (Figure 3A).
The (p)ppGpp-binding site in HPRT manifests a novel (p)ppGpp–protein interaction. The 5′ phosphates and ribose of ppGpp interact with loop III (EDIIDSGLT), a well-characterized PRPP binding motif (Sinha and Smith, 2001), through side chain interactions with Glu99 and Asp100 and backbone amide interactions with Asp103 – Thr107 (Figure 3D and Figure 3—figure supplement 7). The 3′ phosphates of ppGpp are coordinated by backbone amides of loop I, the side chain of Arg165, and the Mg2+ ion (Figure 3D and Figure 3—figure supplement 7). The guanine ring of ppGpp is surrounded by a hydrophobic cleft formed by Ile101, Phe152, and Leu158, and Lys131 hydrogen bonds the guanine’s exocyclic oxygen (Figure 3D and Figure 3—figure supplement 7). We validated the (p)ppGpp-binding residues by mutation analyses which showed that altering residues with side chain interactions with (p)ppGpp greatly weakened pppGpp binding (Figure 3E). In sum, this binding pocket illustrates the first example of a (p)ppGpp binding motif that shares the PRPP-binding motif. Since many proteins bind PRPP (Hove-Jensen et al., 2017), this (p)ppGpp motif may represent a new class of (p)ppGpp targets.
A frequency logo of the binding pocket from 99 bacterial HPRTs shows that most (p)ppGpp-interacting residues are highly conserved across bacteria (Figure 3F and Figure 3—figure supplement 8), and nearly all binding residues are also conserved in the eukaryotic HPRTs (Figure 3—figure supplement 8). The strong conservation across species is not surprising given the close overlap between (p)ppGpp and substrates in the HPRT structures. Since all residues involved in binding ppGpp are also involved in binding substrates, altering the site to affect inhibitor binding would also impact enzyme activity.
The nearly identical recognition of (p)ppGpp and substrates, along with the conservation of the active site, raised the following question: how can some HPRTs be strongly inhibited by basal levels of (p)ppGpp and other HPRTs be almost refractory to (p)ppGpp control despite sharing a conserved binding pocket?
(p)ppGpp prevents PRPP-induced dissociation of HPRT dimer-of-dimers
We noticed a significant difference between the ppGpp- and substrates-bound tertiary structures in the conformation of a flexible loop. This loop, also called loop II, is common to all HPRTs and covers the active site during catalysis (Shi et al., 1999). In the HPRT-ppGpp complex, one side of loop II faces the active site while the other side is a critical part of the interface between two HPRT dimers (Figure 4A). We applied PISA analysis (‘Protein interfaces, surfaces, and assemblies’ service at European Bioinformatics Institute) to predict that this dimer-dimer interaction results in a tetrameric HPRT (Krissinel and Henrick, 2007). In the substrates-bound state, loop II is instead shifted ≈4 Å toward the active site from its position in the dimer-dimer interface (Figure 4A). Loop II also pulls its flanking dimer interface components β3 and α3 toward the active site (Figure 4A). While crystal contacts likely prevent loop II from completely moving over the active site (Figure 4—figure supplement 1), PISA analysis predicts that these changes abolish the dimer-dimer interaction. Using size-exclusion chromatography, we confirmed that apo B. subtilis HPRT is a tetramer (Figure 4B). In contrast, when PRPP, the first substrate to bind HPRT (Yuan et al., 1992), is added at a high concentration (500 μM) in the mobile phase, HPRT tetramers dissociate to dimers (Figure 4B and Figure 4—figure supplements 2, 3 and 4).
Figure 4 with 7 supplements see all
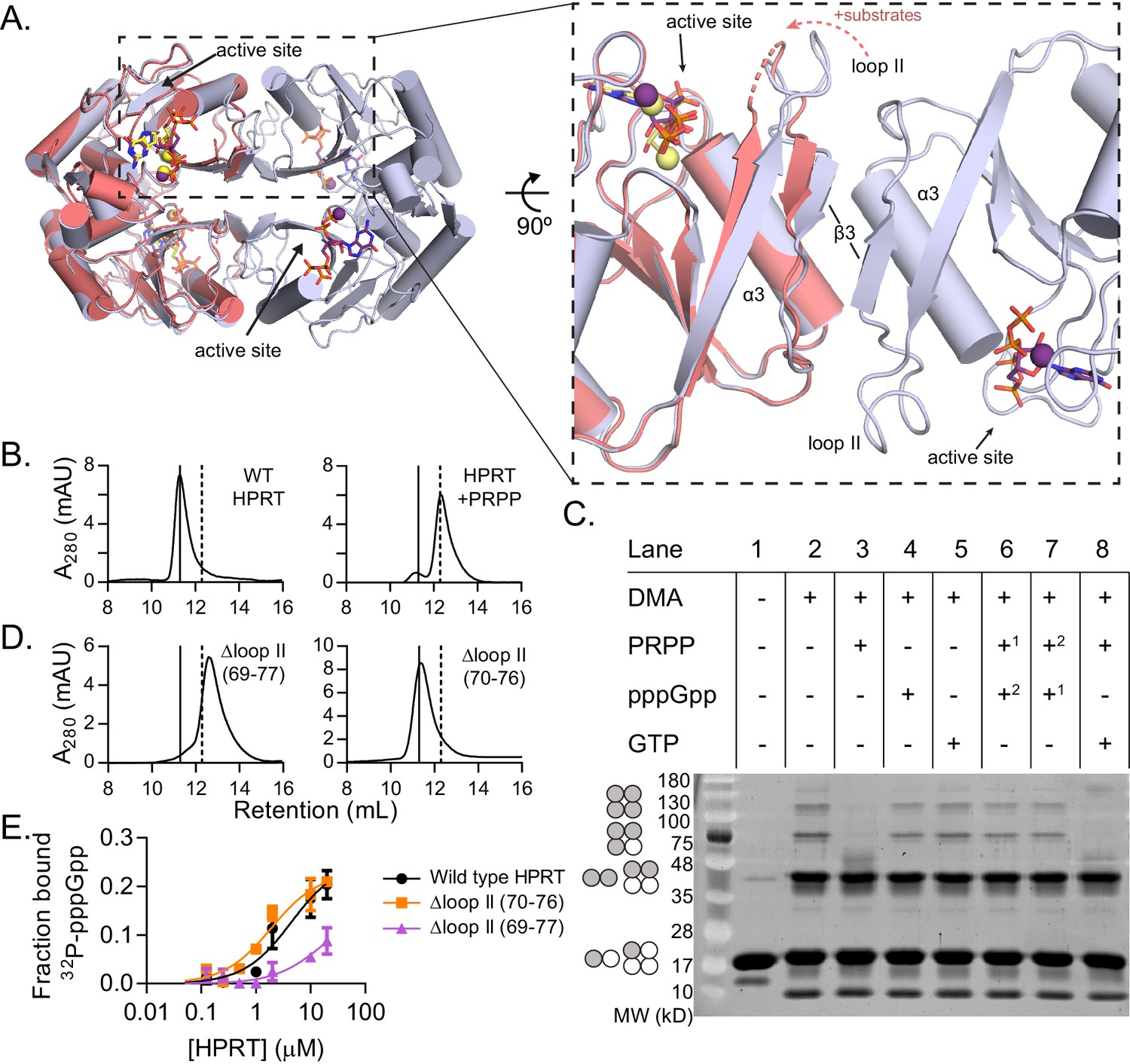
(p)ppGpp counteracts substrate-induced HPRT dimerization and HPRT tetramerization potentiates (p)ppGpp binding.
(A) Overlay of HPRT tetramer crystallized with ppGpp (silver) and HPRT dimer crystallized with substrates (salmon). ppGpp is purple and the substrates are yellow. Inset, View of the dimer–dimer interface within an HPRT tetramer. Secondary structure components at the interface are labeled β3, α3, and loop II. Figure 4—figure supplement 1 shows that changes in loop II are not induced by crystal contacts. (B) Size-exclusion chromatography of B. subtilis HPRT without ligand (left) and with PRPP (right). PRPP addition shifts the oligomeric state from tetramer to dimer. B. anthracis Hpt-1 is also a dimer with PRPP (see Figure 4—figure supplement 2). In B and D, solid line shows B. subtilis HPRT tetramer peak and dotted line shows dimer peak. PRPP must be in mobile phase to cause the shift (Figure 4—figure supplement 3), and the purine base does not affect the oligomeric state with PRPP (Figure 4—figure supplement 4). For molecular weight standards, see Figure 4—figure supplement 7. (C) B. subtilis HPRT crosslinked with dimethyl adipimidate (DMA). Crosslinked multimers were separated on SDS–PAGE. Predicted multimers are shown on the left. Shaded circles represent predicted to be crosslinked monomers, and white circles represent incomplete crosslinking. 1, 2 indicates order of incubation. GTP competition with PRPP did not recapitulate pppGpp blockage of PRPP-induced dimer–dimer dissociation which could be due to difference in affinity, whereby pppGpp binds strongly enough to outcompete PRPP but GTP does not. See Figure 4—figure supplement 5 for effect of crosslinked tetramer (Y117C) on HPRT activity and PRPP competition with pppGpp. See Figure 4—figure supplement 6 for ITC with PRPP. (D) Size-exclusion chromatography of B. subtilis HPRT with a partial loop II deletion (∆70–76) or a complete loop II deletion (∆ 69–77). The complete deletion results in dimerization of the protein. (E) Binding curves between 32P-labeled pppGpp and B. subtilis HPRT variants obtained with DRaCALA. The tetrameric Δloop II (70-76) variant (orange squares) binds as well as wild type HPRT. The dimeric Δloop II (69-77) variant (purple triangles) displays weaker binding. Error bars represent SEM of three replicates.
We next interrogated the effect of (p)ppGpp on the oligomeric state of HPRT using the protein crosslinker dimethyl adipimidate (DMA). Crosslinked apo HPRT was resolved by SDS–PAGE as bands corresponding to monomers, dimers, trimers, or tetramers (Figure 4C, lane 2). All but the tetramer is likely formed by incomplete crosslinking, since dynamic light scattering showed apo B. anthracis Hpt-1 to be a homogeneous population with a hydrodynamic radius (RH) consistent with a tetramer in solution (Table 3). HPRT with pppGpp remained a tetramer according to both crosslinking and dynamic light scattering experiments (Figure 4C and Table 3). Incubation of HPRT with PRPP resulted in loss of trimer and tetramer bands in SDS–PAGE (Figure 4C, lane 3) and a corresponding shift in tetramer to homogeneous dimer with dynamic light scattering (Table 3), confirming that PRPP-bound HPRT is a dimer.
Table 3
Dynamic light scattering of B. anthracis Hpt-1 with pppGpp and PRPP.
https://doi.org/10.7554/eLife.47534.027| Sample | RHa (nm) | Polydb | % Polyd | Est. MW (kDa) |
|---|---|---|---|---|
| HPRT | 4.1 | 0.9 | 23.1 | 90 |
| HPRT + PRPP | 3.1 | 0.7 | 22.5 | 47 |
| HPRT + pppGpp | 4.1 | 1.0 | 24.9 | 91 |
| HPRT + both | 4.2 | 1.2 | 27.8 | 97 |
-
PRPP = phosphoribosyl pyrophosphate.
aRH = hydrodynamic radius.
-
bPolyd = polydispersity.
Importantly, with both PRPP and pppGpp present, HPRT was tetrameric (Figure 4C, lanes 6 and 7 and Table 3), indicating that pppGpp prevents PRPP-induced dimer–dimer dissociation. Thus, pppGpp appears to selectively stabilize HPRT tetramers against PRPP-induced dissociation.
These data also suggest that preventing HPRT from dimer–dimer dissociation and keeping it as a tetramer will disfavor PRPP binding. Indeed, a cysteine substitution across the dimer–dimer interface (Y117C) forcing HPRT to stay as a tetramer via disulfide crosslink results in lower enzymatic activity, and PRPP cannot outcompete pppGpp for binding the tetrameric HPRT variant (Figure 4—figure supplement 5).
Taken together, these data reveal that HPRT binds PRPP as catalytically active dimers, whereas (p)ppGpp maintains HPRT as tetramers.
Dimer–dimer interaction allosterically positions loop II for potentiated (p)ppGpp binding
Given HPRT’s oligomerization, we performed ITC experiments with pppGpp (Figure 2—figure supplement 1) and PRPP (Figure 4—figure supplement 6) to examine ligand binding cooperativity. In the conditions we tested, within physiological ranges of pppGpp and PRPP, we did not observe significant cooperativity. This is expected as cooperativity has not been observed in native bacterial and human HPRTs and has been reported only in mutant human HPRTs (Balendiran et al., 1999; Guddat et al., 2002; Lightfoot et al., 1994; Patta et al., 2015).
To understand the importance of HPRT’s differential oligomeric state with substrates and (p)ppGpp, we performed interface mutational analyses, and we found that, unexpectedly, the dimer–dimer interaction is critical for HPRT’s regulation by (p)ppGpp. We disrupted the dimer–dimer interface of B. subtilis HPRT by constructing two loop II deletion variants of different lengths that resulted in different apo oligomeric states (Figure 4D). While the tetrameric Δloop II (70-76) bound pppGpp as tightly as wild-type HPRT, the dimeric Δloop II (69-77) displayed strongly ablated binding to pppGpp (Kd too weak to estimate; Figure 4E).
Since an engineered dimeric HPRT variant has weakened binding to (p)ppGpp, we next examined the oligomeric state of naturally occurring HPRT homologs with weakened inhibition by (p)ppGpp (Figure 2). Strikingly, in nearly all cases, the (p)ppGpp-insensitive HPRTs were also constitutive dimers (Figure 5A). These homologs have normal enzymatic activity (Table 4) and only differ in their ability to be regulated by (p)ppGpp. Our results suggest that tetrameric HPRT, but not dimeric HPRT, allows (p)ppGpp at basal levels to bind to and inhibit its activity.
Figure 5 with 3 supplements see all
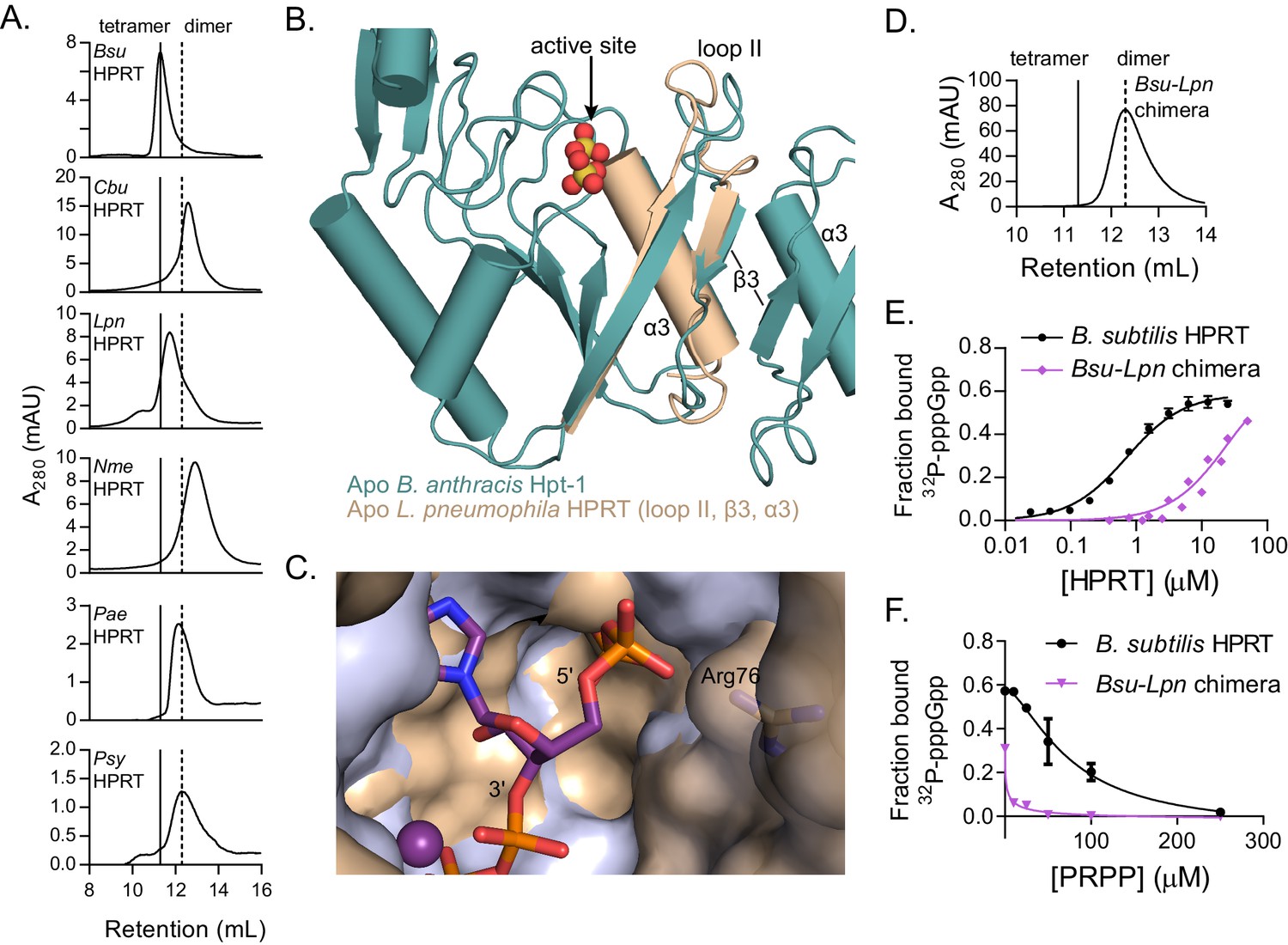
Dimer–dimer interaction holds loop II away from the (p)ppGpp binding pocket to potentiate (p)ppGpp binding.
(A) Size-exclusion chromatograms show that C. burnetii, L. pneumophila, N. meningitidis, P. aeruginosa, and P. syringae HPRTs are non-tetrameric without ligands. Vertical lines represent B. subtilis HPRT tetramer and dimer peaks. (B) Overlay of apo B. anthracis Hpt-1 (teal) and the L. pneumophila HPRT interface (wheat; PDB ID 5ESW). In L. pneumophila HPRT, loop II and β3 are further from the dimer-dimer interface and closer to the active site. See Figure 5—figure supplement 1 for overlay with substrates-bound HPRT. (C) Overlay of B. anthracis Hpt-1 – ppGpp (silver; ppGpp in purple) and L. pneumophila HPRT (wheat; PDB ID 5ESX) shows that the 5′ phosphate binding pocket is compressed by the conformation of loop II in the dimeric HPRT. Arg76 (side chain shown as sticks) in L. pneumophila HPRT forms part of the compressed pocket. Asp109 in L. pneumophila HPRT is hidden due to poor electron density. (D) Size-exclusion chromatogram of the dimeric Bsu-Lpn chimera. The chimera has 21 residues at the B. subtilis HPRT interface replaced with L. pneumophila HPRT residues (see Figure 5—figure supplement 2). (E) DRaCALA shows that the Bsu-Lpn chimera binds 32P-labeled pppGpp more weakly. Error bars represent SEM of three replicates. Bsu-Lpn chimera performed in duplicate. Figure 5—figure supplement 3 shows that stability of dimeric HPRTs is not compromised. (F) Competition between PRPP and 32P-labeled pppGpp shows that the dimeric Bsu-Lpn chimera is more sensitive to PRPP than the tetrameric wild type. Error bars are range of duplicate.
Table 4
HPRT kinetic parameters.
https://doi.org/10.7554/eLife.47534.032| HPRT | Km (guanine) (μM) | Km (PRPP) (μM) | kcat (s−1) |
|---|---|---|---|
| B. anthracis 1 | 5.2 ± 1.8 | 111.6 ± 16.8 | 22 |
| B. subtilis | -- | 165.6 ± 11.9 | 25.4 |
| C. burnetii | 17.6 ± 2.0 | 44 ± 3.3 | 12.9 |
| P. phosphoreum | -- | 81 ± 12.5a | 30.4a |
| N. meningitidis | 68 ± 7.0 | 103 ± 12 | 9.6 |
| E. colib | -- | 192 ± 7.0a | 59a |
| L. pneumophilac | 10.5 ± 1.1 | 60 ± 9 | -- |
| M. tuberculosisd | 10 ± 1 | 650 ± 70 | 0.193 |
-
PRPP = phosphoribosyl pyrophosphate.
aobtained with hypoxanthine as the purine base.
-
± standard error.
The structural basis of how the dimer–dimer interaction promotes (p)ppGpp binding in tetrameric HPRTs can be seen from comparative structures of apo B. anthracis Hpt-1 and apo L. pneumophila HPRT (PDB ID 5ESW) (Zhang et al., 2016). Loop II in dimeric L. pneumophila HPRT is positioned closer to the active site (Figure 5B), mimicking substrates-bound HPRT even in the absence of substrates (Figure 5—figure supplement 1), and compresses the cavity surrounding the 5′ phosphates of ppGpp (Figure 5C). This conformation does not fully accommodate (p)ppGpp, but it should accommodate PRPP since its 5′ monophosphate fits in the cavity compressed by loop II (Figure 5—figure supplement 1). In contrast, in tetrameric HPRTs, loop II is pulled away from the active site by the dimer–dimer interaction and is positioned for optimal (p)ppGpp binding. There are many examples of the interdependence of protein oligomerization and ligand binding (Traut, 1994). By discovering and comparing two different evolved oligomeric states of HPRT, we have been able to demonstrate how a structural rearrangement through oligomerization allosterically affects ligand binding to a non-interface pocket.
To identify the determinant for the oligomeric state of naturally occurring HPRTs, we constructed a chimera of B. subtilis HPRT with 21 dimer–dimer interface residues replaced with their corresponding L. pneumophila HPRT residues (Figure 5—figure supplement 2). The chimera is a stable dimer (Figure 5D and Figure 5—figure supplement 3), indicating that the determinants for oligomerization lie in the dimer-dimer interface residues. The chimera also had a > 20 fold lower affinity for pppGpp than the tetramer (Kd ≈ 24 μM versus ≈ 1 μM) (Figure 5E), and PRPP more potently competed with pppGpp binding to the dimeric chimera than to tetrameric WT (Figure 5F), suggesting that there may be an evolved linkage between the dimer–dimer interface of HPRT tetramers and (p)ppGpp regulation.
Dimer-dimer interface coevolved with strong (p)ppGpp binding across species
The relationship between HPRT oligomeric state and sensitivity to (p)ppGpp prompted us to examine whether HPRT oligomerization has coevolved with (p)ppGpp regulation. To test this, we turned to ancestral protein sequence reconstruction to infer the evolution of HPRT (Hochberg and Thornton, 2017). With a phylogenetic tree derived from an alignment of 430 bacterial and eukyarotic HPRTs, we used maximum likelihood to infer the most likely ancestral HPRT protein sequences based on the phylogeny of the extant HPRT sequences (Yang et al., 1995) (Figure 6A). From the first ancestral HPRT (Anc1), the phylogenetic tree bifurcated into two broad lineages. One lineage contained the known dimeric HPRTs with weakened (p)ppGpp inhibition (Figure 6A) (Hug et al., 2016). The second lineage contained the vast majority of HPRTs, including (p)ppGpp-sensitive HPRTs in the Bacteroidetes, Firmicutes, and γ-proteobacteria. There are three interface residues conserved only in the dimeric lineage (Trp82, Pro86, and Ala110) (Figure 6B). Varying these residues in B. subtilis HPRT produces non-tetrameric variants (Figure 6—figure supplement 1). Notably, Anc1 HPRT was likely tetrameric, since it lacks the residues found in the dimeric lineage (Figure 6B).
Figure 6 with 2 supplements see all
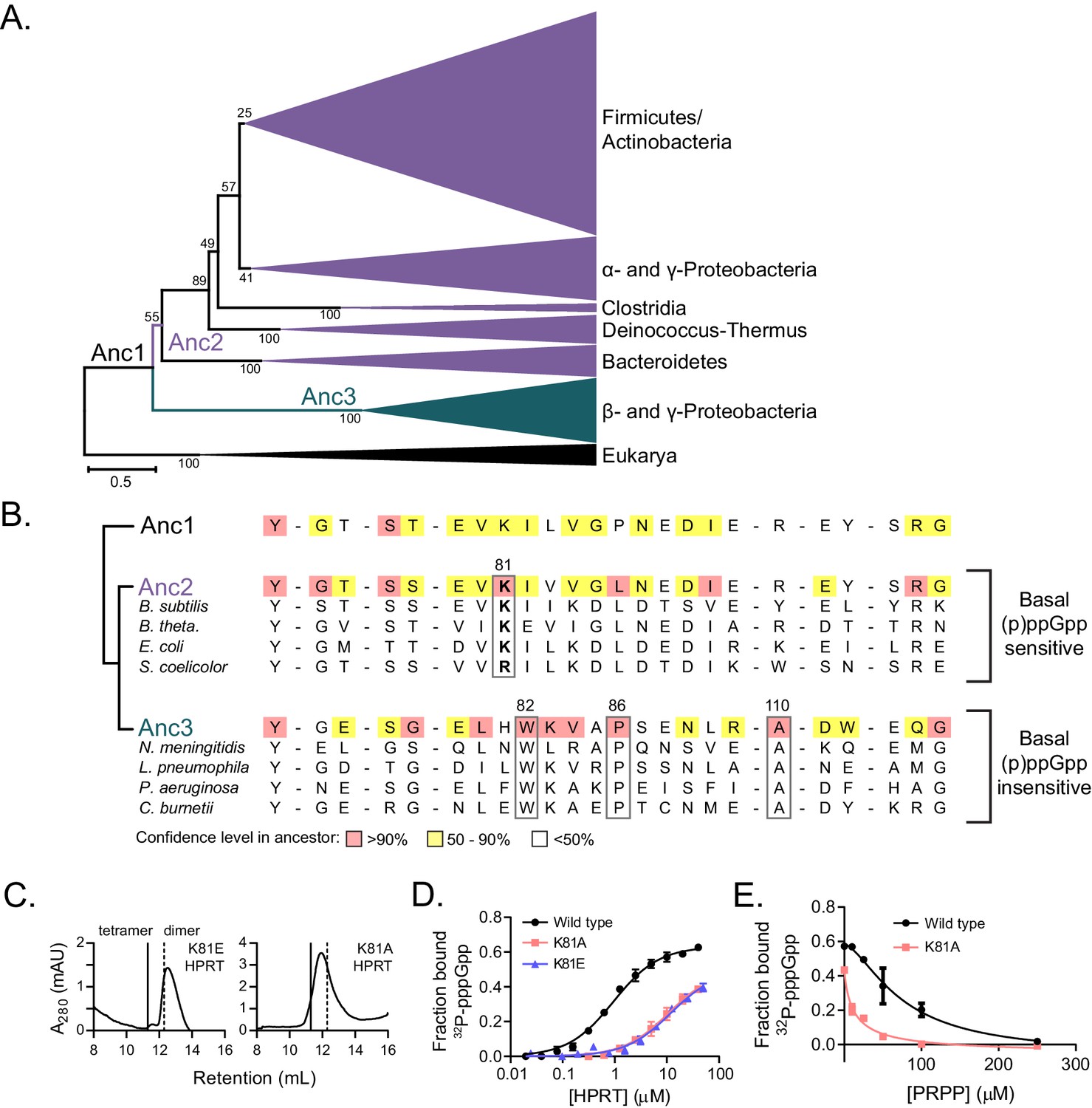
Coevolution of the HPRT dimer-dimer interface and regulation by basal levels of (p)ppGpp.
(A) A maximum likelihood phylogenetic tree of 430 HPRT amino acid sequences rooted on eukaryotic HPRTs. Bootstrap values from 100 replicates are shown. Anc1-3 refer to ancestral HPRTs. The tree reveals two main lineages: one with dimer-dimer interaction motifs (Anc2; purple) and one lacking the interaction motifs (Anc3; blue). Branch length scale indicates 0.5 substitutions per site. See Materials and methods for tree construction. See Source Data one for the full alignment of HPRTs. (B) Alignment of HPRT dimer-dimer interface from Anc1, Anc2, Anc3, and select extant HPRTs. Nearly all (p)ppGpp-regulated HPRTs share Lys81 found in Anc2. Dimeric HPRTs insensitive to basal (p)ppGpp lack Lys81, but share Trp82, Pro86, and Ala110 (B. anthracis Hpt-1 numbering). Figure 6—figure supplement 1 shows the role of these residues at the interface. For the ancestors, red indicates high (>90%), yellow moderate (50–90%), and no color low (<50%) confidence level in the residue identity. (C) B. subtilis K81E and K81A HPRTs have weakened tetramerization according to size-exclusion chromatography. Figure 6—figure supplement 2 shows Lys81 at the dimer–dimer interface. (D) Binding of 32P-labeled pppGpp to wild type (black circles), K81A (red squares), and K81E (blue triangles) HPRTs using DRaCALA. Error bars represent SEM of three replicates. (E) PRPP competition with 32P-labeled pppGpp for binding wild type (black circles) and K81A HPRTs (red squares). K81A is more sensitive to PRPP competition than the tetrameric wild type. Error bars are range of duplicate.
-
Figure 6—source data 1
Protein alignment of HPRTs.
- https://doi.org/10.7554/eLife.47534.036
A tracing of the dimer-dimer interface pinpointed residues that coevolve with (p)ppGpp regulation. We followed the change in interface residues from Anc1 to Anc2, the ancestor common to the (p)ppGpp-inhibited HPRTs (Figure 6A). One residue in particular, Lys81, increased in confidence between Anc1 and Anc2 (Figure 6B). Lys81 is part of a conserved β-strand and loop motif (residues 81–87) that interact with one another across the dimer–dimer interface, so their evolution is likely important for HPRT tetramerization (Figure 6—figure supplement 2). Lys81 is highly conserved in (p)ppGpp-regulated HPRTs but not in (p)ppGpp-insensitive HPRTs (Figure 6B), suggesting that (p)ppGpp regulation is associated with the evolution of this residue. Therefore, we constructed Lys81 variants with weakened dimer–dimer interfaces (Figure 6C). One variant with a charge reversal (K81E) resulted in a mostly dimeric HPRT and a less disruptive K81A HPRT exhibited a rapid equilibrium between tetramer and dimer (Figure 6C). Importantly, pppGpp bound less well to both K81A and K81E HPRT variants relative to wild-type HPRT (Figure 6D), and PRPP more potently competed with pppGpp binding to the K81A variant relative to the tetrameric wild type (Figure 6E). We conclude that the HPRT dimer-dimer interface has coevolved with strong (p)ppGpp regulation by sequestering loop II at the dimer–dimer interface and opening the active site for (p)ppGpp binding (Figure 7), whereas evolution of interface residues associated with losing this dimer-dimer interaction weakens (p)ppGpp regulation.
Figure 7
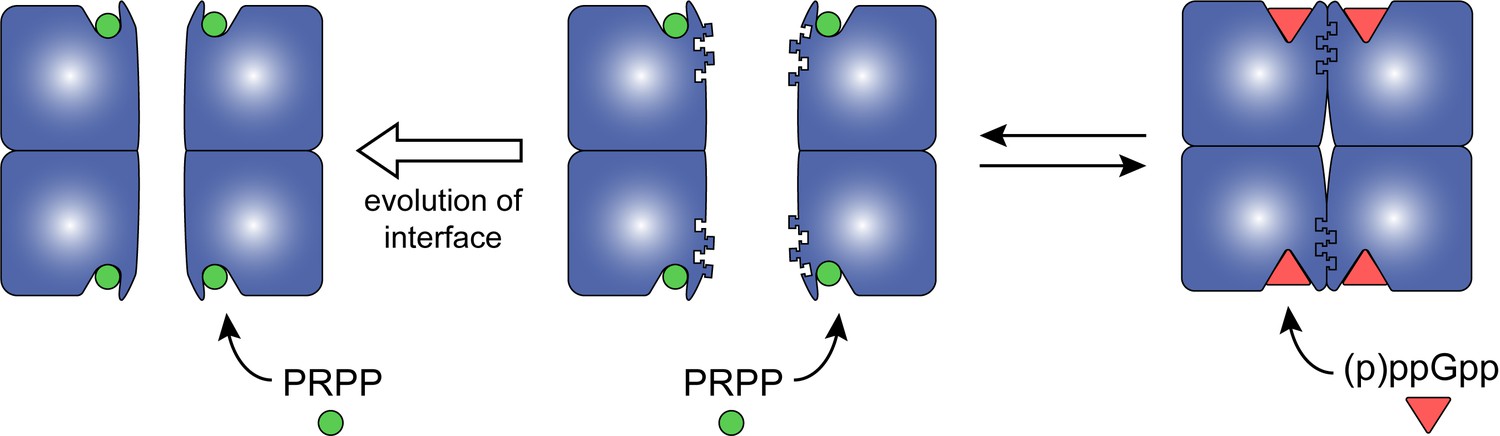
Model for how evolution of protein oligomerization affects ligand-mediated regulation.
For an enzyme such as HPRT, where the inhibitor (p)ppGpp binds to nearly identical sites as the substrates (PRPP), evolutionary plasticity of inhibition can be mediated through subunit oligomerization. Dimeric HPRTs present a smaller binding pocket that preferentially bind to the substrate than to (p)ppGpp. On the other hand, HPRTs with a dimer–dimer interface motif would allosterically open the pocket, thus favoring (p)ppGpp binding and inhibition than substrate-binding and activity.
Protein–protein interface allosterically alters the binding of (p)ppGpp to guanylate kinase
Intrigued by finding the determinants of (p)ppGpp regulation of HPRT, we turned to guanylate kinase (GMK), another (p)ppGpp-regulated enzyme (Kriel et al., 2012; Liu et al., 2015b). We previously found that GMK is inhibited by (p)ppGpp in multiple phyla of bacteria but not in Proteobacteria despite the fact (p)ppGpp binds the conserved active site (Liu et al., 2015b). This puzzle may now be explained by a protein–protein interaction affecting (p)ppGpp binding: GMK is a dimer. In the (p)ppGpp-sensitive GMK, a lid domain opens the active site for (p)ppGpp binding through pulling of the lid by the C-terminal helix of the adjoining monomer in a GMK dimer (Figure 8A). In the (p)ppGpp-insensitive E. coli GMK, the lid domain is closed, with the C-terminal helix from the adjoining monomer perhaps responsible for stabilizing the closed position (Figure 8A). We had previously swapped the lid domains to test their role in (p)ppGpp inhibition, but this had led to inactive enzymes (Liu et al., 2015b). To identify whether the C-terminal tail is holding the lid domain open or closed, we deleted 10 residues from the C-terminus in both the (p)ppGpp-sensitive B. subtilis GMK and the (p)ppGpp-resistant E. coli GMK with striking observations (Figure 8B C). Remarkably, while these residues are away from the (p)ppGpp binding pocket of the adjoining monomer, their deletion led to weakened binding between B. subtilis GMK and pppGpp and reduced inhibitory effect on its enzymatic activity. On the other hand, deleting the C-terminus of the (p)ppGpp-resistant E. coli GMK allowed a weak binding between E. coli GMK and pppGpp, and rendered E. coli GMK sensitive to pppGpp inhibition. This demonstrates that the allosteric interaction between the lid domain and C-terminus of two adjoining GMK monomers is the determining factor for (p)ppGpp specificity.
Figure 8

Protein–protein interface allosterically alters the binding of (p)ppGpp to guanylate kinase.
(A) In S. aureus GMK (left; PDB ID 4QRH), an open lid domain (red) allows (p)ppGpp binding at the active site, but in E. coli GMK (right; PDB ID 2ANB), the closed lid domain occludes (p)ppGpp binding at the active site (bound to GMP). A C-terminal tail in the second monomer of a GMK dimer interacts with the lid domain in both proteins. Nineteen residues in the lid domain and six residues in the C-terminal tail of S. aureus GMK are unresolved. (B) DRaCALA of full-length and ∆C-terminal tail (∆CT tail) B. subtilis and E. coli GMKs. Error bars are mean of duplicate. (C) Inhibition of GMKs from (D) relative to uninhibited activity. Error bars are SEM of triplicate. (D) Maximum likelihood phylogenetic tree of 41 GMKs rooted on eukaryotic GMKs as the outgroup. Bootstrap values from 1000 replicates are shown. Anc1-3 refer to ancestral HPRTs. See Materials and methods for tree construction. See Source Data one for the full alignment of GMKs. (E) Alignment of the lid domain and C-terminal tail of ancestral and select extant GMKs. Boxed residues are part of the lid domain - C-terminal tail interaction.
-
Figure 8—source data 1
Protein alignment of GMKs.
- https://doi.org/10.7554/eLife.47534.038
We constructed a phylogenetic tree of 41 GMKs (Figure 8D). As with HPRT, the tree showed that the (p)ppGpp-insensitive and (p)ppGpp-sensitive GMKs bifurcated from the last common ancestor. Importantly, these (p)ppGpp-insensitive and (p)ppGpp-sensitive GMKs use different residues to form the interaction between the lid domain and the C-terminal helix (Figure 8E). (p)ppGpp-insensitive GMKs have a hydrophobic interaction between the lid domain and C-terminal tail, whereas (p)ppGpp-sensitive GMKs contain more charged residues at this interface. Ancestral protein reconstruction suggests that the last common ancestor (Anc1) is more similar to the (p)ppGpp-sensitive GMKs (Anc3) (Figure 8E). Altogether, this suggests that the protein-protein interface of GMK has also evolved to affect (p)ppGpp binding.
Discussion
(p)ppGpp is a stress-induced signaling molecule in bacteria that is also critical for cellular fitness and homeostasis even at basal levels. However, (p)ppGpp targets and their evolution in different bacteria remain poorly understood. Here, we have described a mechanism explaining how basal levels of (p)ppGpp potently regulate the activity of the housekeeping enzyme HPRT through a novel binding site that may represent a new class of (p)ppGpp effectors. Intriguingly, this site overlaps completely with the active site and is conserved among bacteria, yet differential regulation by (p)ppGpp can be achieved through variation of an allosteric component: the interaction between dimeric subunits of the HPRT tetramer. This interaction tethers a flexible loop at the interface and away from the active site, allowing the binding pocket to accomodate (p)ppGpp. Lack of the dimer–dimer interaction causes HPRT to occlude (p)ppGpp and favor substrate binding. This dimer–dimer interaction is due to an interface motif that appears to have co-evolved with (p)ppGpp binding in the majority of bacterial HPRTs sensitive to (p)ppGpp, whereas dimeric HPRTs without the motif are resistant to (p)ppGpp. We conclude that evolution of the dimer-dimer interface in tetrameric HPRTs has potentiated (p)ppGpp binding to enable basal (p)ppGpp modulation of metabolism, thus increasing bacterial fitness in fluctuating environments.
A novel, high-affinity (p)ppGpp binding motif that shares the PRPP motif
Our HPRT-ppGpp structure revealed a binding motif distinct from known (p)ppGpp-protein interactions. There are over 50 known (p)ppGpp targets across bacteria, but identifying motifs associated with (p)ppGpp binding has been difficult (Corrigan et al., 2016; Wang et al., 2019; Zhang et al., 2018). In a few cases, (p)ppGpp binds allosteric sites at protein interfaces (Kanjee et al., 2011; Ross et al., 2013; Ross et al., 2016; Steinchen et al., 2015; Wang et al., 2019). For many other targets, (p)ppGpp binds at a GTP binding site, leading to overlapping (p)ppGpp and GTP binding motifs (Fan et al., 2015; Kihira et al., 2012; Liu et al., 2015b; Pausch et al., 2018; Rymer et al., 2012). In the case of HPRT, however, the (p)ppGpp binding site is not at an interface between two proteins, nor is it a GTP binding site. Instead, it shares a well-characterized motif associated with PRPP binding (EDIIDSGLT in B. anthracis Hpt-1) (Sinha and Smith, 2001). Other PRPP-binding proteins, including UPRT and APRT, have been shown to bind (p)ppGpp (Wang et al., 2019; Zhang et al., 2018), and it is likely that (p)ppGpp also binds to their PRPP motif. Identification of this motif may provide a new class of (p)ppGpp-binding proteins, allowing us to predict additional targets.
(p)ppGpp interacts with proteins in two main conformations: elongated, with the phosphate arms extended away from one another in a T shape, and ring-like, with the phosphate arms near one another in a Y shape. The compact, ring-like conformation has been associated with higher affinity interactions than the elongated conformation (Steinchen and Bange, 2016). However, in the HPRT–ppGpp interaction, ppGpp takes an elongated conformation, but exhibits strikingly tight affinities as high as Kd ≈ 0.1 μM for some species (Figure 2D and Table 1). This high affinity may be due to extensive backbone amide interactions with both phosphate arms as well as (p)ppGpp’s close mimicry of substrate binding (Figure 3D). It is likely that higher affinity interactions with the elongated (p)ppGpp conformation will become more common as additional targets are characterized.
HPRT tetramerization enables basal (p)ppGpp inhibition
Our data show that the dimer–dimer interface of HPRT potentiates (p)ppGpp regulation by allosterically promoting a conformation conducive to ligand binding (Figure 7). HPRT’s tetramerization harnesses the flexible loop II to open the binding pocket (Figure 4A) for (p)ppGpp to bind with high affinity. Loop II’s influence on (p)ppGpp binding may be the reason that it has evolved to be part of the interface of bacterial HPRTs, whereas in eukaryotic (e.g. human) HPRTs, which likely do not interact with (p)ppGpp in nature, loop II is on the outside of the oligomer facing solvent (Eng et al., 2015).
Our model can also explain the evolutionary significance of bacterial HPRTs functioning as dimers without dissociating to monomers. Strength of protein–protein interactions has been correlated with increased buried surface area at the interface (Nooren and Thornton, 2003). In a hypothetical monomer–monomer interaction, the smaller loop II interface may be too weak and too transient to keep loop II away from the active site. In the interaction between two homodimers, the loop II interface is duplicated, which increases the surface area and provides anchor points to hold loop II at the interface and away from the (p)ppGpp binding pocket (Figure 7).
HPRT tetramerization also affects a key component of basal (p)ppGpp’s ability to regulate HPRT: its competition with PRPP. Most bacterial HPRTs have a Km for PRPP in the range of 50–200 μM (Guddat et al., 2002; Zhang et al., 2016) (Table 4). Tetrameric HPRTs that bind (p)ppGpp with high affinity also tend to have higher Km values for PRPP, suggesting that they are optimized for (p)ppGpp binding rather than PRPP binding (Table 4). By forcing HPRT into a dimeric state independent of ligands, either through natural evolution in the case of the HPRT homologs or through mutating the dimer–dimer interface sequence, HPRT becomes both refractory to (p)ppGpp binding (Figure 5) and also preferably binds PRPP (Figures 5F and 6E, Table 4). The overall effect is an HPRT that is resistant to inhibition by basal levels of (p)ppGpp.
HPRT regulation allows maintenance of cellular homeostasis by basal levels of (p)ppGpp
Our characterization of HPRT explains how bacteria can be regulated by basal levels of (p)ppGpp. While (p)ppGpp is mostly chacterized as a regulator of gene expression that functions in starvation-induced concentrations, basal concentrations of (p)ppGpp are known to be responsible for sustaining antibiotic tolerance and virulence in E. faecalis (Gaca et al., 2013), maintaining cyanobacterial light/dark cycles (Puszynska and O'Shea, 2017), influencing rRNA expression and growth rate in E. coli (Potrykus et al., 2011), and regulating GTP synthesis in Firmicutes (Gaca et al., 2013; Kriel et al., 2012). However, the principles that determine how basal and induced (p)ppGpp regulate different cellular targets remained unclear. In the single species B. subtilis, for example, (p)ppGpp interacts with DNA primase (Ki, ppGpp = 250 μM), IMP dehydrogenase (Ki, ppGpp = 50 μM), and guanylate kinase (Ki, pppGpp = 14 μM), allowing them to be inhibited in vivo at induced (p)ppGpp levels (Liu et al., 2015b; Pao and Dyes, 1981; Wang et al., 2007). In contrast, (p)ppGpp’s interaction with B. subtilis HPRT is far stronger (Ki = 1.7 μM; Figure 1D), potentially allowing (p)ppGpp to modulate its activity at basal levels throughout growth. Our data suggest that the dimer-dimer interaction is a mechanism that strengthens (p)ppGpp binding affinity, but also favors the inhibitor over the substrate PRPP. Physiological PRPP levels in bacteria are roughly 0.1–1 mM (Bennett et al., 2009; Berlin and Stadtman, 1966; Jensen, 1983). At 1 mM PRPP, the high end of physiological levels, (p)ppGpp at a basal level (25 μM) can still strongly inhibit tetrameric HPRT activity but not the constitutive dimeric homologs (Figure 2E). Thus tetramerization allows basal (p)ppGpp to regulate HPRT and thus constantly regulate purine nucleotide synthesis via the salvage pathway.
The basal regulation of HPRT may be important for fitness in environmental niche. For example, for bacteria in the microbiota whose HPRTs were potently inhibited by (p)ppGpp, it may robustly maintain intracellular metabolism despite fluctuations in exogenous purines that could depend on the purine content of the diet (Choi et al., 2004; Kaneko et al., 2014; Zgaga et al., 2012).
Protein oligomerization allosterically alters ligand specificity
The mechanism we characterized for HPRT and GMK may represent a more broadly applicable principle by which evolution of protein oligomerization changes the conformation of a ligand binding pocket to alter ligand specificity. Oligomerization of proteins into homomers can provide multiple adaptive advantages, including mechanisms of allosteric regulation by a ligand binding to a site other than the enzyme's active, enabling cooperative substrate binding, promoting protein stability, providing complete active sites or ligand binding sites at oligomeric interfaces, maintaining proximity of signal transduction, and serving cytoskeletal or other structural roles (Ali and Imperiali, 2005; Bergendahl and Marsh, 2017; Matthews and Sunde, 2012; Perutz, 1989; Perica et al., 2012; Traut, 2008; Traut, 1994). For HPRT, instead, the advantage of tetramerization is strengthening (p)ppGpp specificity at the active site for enhanced competitive regulation by basal levels of this nucleotide.
Similarly, we show oligomeric interaction also plays a major role in altering specificity for the (p)ppGpp target guanylate kinase (GMK). GMK is inhibited by (p)ppGpp in multiple phyla of bacteria but not in Proteobacteria despite the fact (p)ppGpp binds the conserved active site (Liu et al., 2015b). We now found that this is because GMK is a dimer and the monomer–monomer interaction in GMK allosterically affects the lid domain conformation and (p)ppGpp binding by occluding (p)ppGpp binding in E. coli and promoting (p)ppGpp binding in B. subtilis. By deleting the C-terminal helix from the adjoining monomer that is responsible for stabilizing the lid domain position, we can, for the first time, remove the differential specificity to (p)ppGpp between E. coli and B. subtilis GMK homologs while keeping the enzymes active (Figure 8).
It is striking to note that a majority of ligands bind within 6 Å of a protein–protein interface (Gao and Skolnick, 2012). Thus, the allosteric effect of oligomeric or other protein-protein interactions on ligand binding likely extends to many other regulations by signaling ligands.
Coevolution of ligand binding and protein oligomerization
What is the evolutionary advantage of regulating ligand-binding through protein-protein interactions? One potential advantage is that it provides evolutionary flexibility for ligand binding. Evolving different residues within ligand binding sites can alter ligand specificity, but active sites that bind substrates and inhibitors are under functional and evolutionary constraints to maintain enzymatic activity (Echave et al., 2016; Huang et al., 2015). On the other hand, protein–protein interfaces like the dimer–dimer interface of HPRT and the monomer–monomer interface of GMK are more evolutionarily flexible, particularly when they are not obligate interactions, as they allow mutations to accumulate away from a conserved active site (Echave et al., 2016; Mintseris and Weng, 2005). This suggests that changing oligomeric states could be an evolutionarily flexible mechanism for altering ligand specificity (Figure 7). Indeed, the coevolution of (p)ppGpp regulation and the HPRT and GMK protein–protein interactions has provided us with examples of how protein oligomerization and ligand binding can coevolve, demonstrating that organisms have already adopted this strategy.
In addition to (p)ppGpp, many small molecules serve as signaling ligands that regulate protein activities (Gao and Skolnick, 2012; Najmanovich, 2017). Our results suggest that ligand binding, even at non-interface binding pockets, influence evolutionary diversification of protein oligomers potentially through purifying selection of conformations that favor protein–ligand interactions. While homomeric evolution of some proteins has been implicated as physicochemical or stochastic processes (Abrusán and Marsh, 2018; André et al., 2008; Lukatsky et al., 2007; Lynch, 2013), our data provide evidence for ligand binding as an adaptive advantage driving the evolutionary diversification of protein homomers. Given the proximity of ligand binding sites to protein interfaces (Gao and Skolnick, 2012), and since it is easier to evolve protein–protein interactions (Perica et al., 2012) rather than evolving new sites for allosteric regulation, such an adaptive benefit is likely to exist more broadly beyond (p)ppGpp in other protein–ligand interactions.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (Bacillus subtilis) | gmk::ecgmk | (Liu et al., 2015b) | N/A | |
| Strain, strain background (B. subtilis) | gmk::ecgmk amyE::Phyperspac-ecgpt | This work | N/A | |
| Strain, strain background (B. subtilis) | gmk::ecgmk amyE::Phyperspac-bshprT | This work | N/A | |
| Recombinant DNA reagent | pLIC-trPC-HA (plasmid) | (Stols et al., 2002) | N/A | |
| Recombinant DNA reagent | pHM1381 (plasmid) | (Mechold et al., 2002) | N/A | |
| Recombinant DNA reagent | pDR90 (plasmid) | (González-Pastor et al., 2003) | N/A | |
| Peptide, recombinant protein | human HPRT1 | Novoprotein | Novoprotein #C294 | |
| Chemical compound, drug | 5-phospho-D -ribose 1-diphosphate pentasodium salt (PRPP) | MilliporeSigma | MilliporeSigma # P8296 | |
| Chemical compound, drug | dimethyl adipimidate (DMA) | ThermoFisher | ThermoFisher # 5001417627 | |
| Chemical compound, drug | 9-deazaguanine | Santa Cruz Biotechnology | Santa Cruz #sc-217528 | |
| Chemical compound, drug | SYPRO Ruby | Bio-Rad | Bio-Rad #1703125 | |
| Chemical compound, drug | SYPRO Orange | MilliporeSigma | MilliporeSigma #S5692 | |
| Chemical compound, drug | guanine | MilliporeSigma | MilliporeSigma #G6779 | |
| Software, algorithm | GraphPad Prism v. 5.02 (software) | https://www.graphpad.com/ | RRID:SCR_002798 | |
| Software, algorithm | ImageQuant (software) | https://www.gelifesciences.com | RRID:SCR_014246 | |
| Software, algorithm | MicroCal Origin 5.0 (software) | https://www.malvernpanalytical.com | RRID:SCR_002815 | |
| Software, algorithm | MEGA X (software) | https://www.megasoftware.net/ | RRID:SCR_000667 | |
| Software, algorithm | Phenix (software) | https://www.phenix-online.org/ | RRID:SCR_014224 | |
| Software, algorithm | Coot (software) | https://www2.mrc-lmb.cam.ac.uk/personal/pemsley/coot/ | RRID:SCR_014222 | |
| Software, algorithm | Dynamics v. 5.25.44 (software) | https://www.wyatt.com/products/software/dynamics.html | N/A |
Strain and plasmid construction and mutagenesis
Request a detailed protocolFor purification and DRaCALA analyses, hprT coding sequences were cloned into the pLIC-trPC-HA vector (pJW269) using the ligation independent cloning (LIC) protocol (Eschenfeldt et al., 2009; Stols et al., 2002) and was scaled up to include more hprT sequences when necessary as described (Abdullah et al., 2009). The hprT sequences were identified in each organism by enzyme classification number (EC 2.4.2.8), by BLAST using B. subtilis or E. coli hprTs, or by database annotation (UniProt, EnsemblBacteria, KEGG Genome). Bacterial hprT homologs were amplified from genomic DNA, cloned using LIC, and transformed into E. coli BL21(DE3) (see Supplementary file 1 for primers and plasmids). Amino acid substitutions were performed either with QuikChange XL Site-Directed Mutagenesis (Agilent Technologies) or with a megaprimer site-directed mutagenesis protocol (Kirsch and Joly, 1998). Loop II deletions were made using a protocol as described (Hansson et al., 2008). pLIC-trPC-HA inserts were amplified and sequenced for confirmation using oJW1124 and oJW492.
To create strains overexpressing protein in B. subtilis gmk::ecgmk, B. subtilis hprT and E. coli gpt were amplified with oJW1569/1570 and oJW1550/1551, respectively, prior to restriction cloning into pDR90 using HindIII and SphI (New England Biolabs). Plasmids were linearized with XhoI and transformed into JDW2108 (B. subtilis gmk::ecgmk) by growing JDW2108 in 1x modified competence medium (Spizizen, 1958) until turbid (≈6 hr) prior to adding linearized plasmid and growing an additional 2 hr. Transformants were selected on 80 μg/ml spectinomycin (MilliporeSigma) and recombination into amyE was confirmed by patching transformants on LB-starch plates and checking for lack of starch utilization.
Protein purification
Request a detailed protocolHPRTs were recombinantly expressed in E. coli BL21(DE3) (NEB) from a pLIC-trPC-HA plasmid with the gene inserted downstream of a sequence encoding a 6X histidine tag and a tobacco etch virus (TEV) protease recognition site. Seed cultures grown to mid-log phase were diluted 1:50 into batch culture of LB supplemented with 100 μg/mL carbenicillin (Research Products International). Protein synthesis was induced at OD600 ≈ 0.8 with 1 mM isopropyl β-D-1 thiogalactopyranoside (IPTG) (RPI) for 4 hr. Cells were pelleted and stored at −80° C until purification. GMKs and E. coli XGPRT were expressed in an identical manner.
For large-scale purifications, cells were resuspended in Lysis Buffer (50 mM Tris-HCl pH 7.5, 500 mM NaCl, 10 mM imidazole) and lysed with a French press. The lysate was centrifuged to obtain the soluble fraction, which was filtered through 0.45 μm filters. The sample was put over a HisTrap FF column (GE Healthcare) on an AktaPure FPLC (GE Healthcare), the column was washed with 20 column volumes of Lysis Buffer, and the protein was eluted with a gradient of increasing Elution Buffer (50 mM Tris-HCl pH 7.5, 500 mM NaCl, 250 mM imidazole). Recombinant protein fractions were dialyzed with 10 mM Tris-HCl (pH 7.5), 100 mM NaCl, 1 mM DTT, and 10% glycerol prior to concentrating and flash-freezing for storage. For small-scale purifications of HPRTs, GMKs, and XGPRT, Ni-NTA spin columns were used according to manufacturer’s instructions (Qiagen). Lysis buffer was 100 mM sodium phosphate (pH 8.0), 500 mM NaCl, and 10 mM imidazole. Wash and elution buffers were the same as the lysis buffer except with 20 mM and 500 mM imidazole, respectively. Protein purity was determined using SDS-PAGE, and concentrations were measured using the Bradford assay (Bio-Rad) or using A280 with extinction coefficients calculated by ProtParam (SIB ExPASy Bioinformatics Resource Portal).
To purify 35 HPRT homologs for activity assays (see Figure 2), a 96-well Capturem His-tagged purification kit (Clontech) was used according to the manufacturer’s instructions. The following buffers were used: lysis [xTractor (Clontech) + 1 μg/mL DNase I], wash [20 mM Na3PO4 pH 7.6, 200 mM NaCl, 10 mM imidazole], elution [20 mM Na3PO4 pH 7.6, 500 mM NaCl, 500 mM imidazole]. Following purification, the buffer was exchanged to 10 mM HEPES pH 8, 150 mM NaCl, 10 mM MgCl2, 1 mM DTT using Zeba spin desalting plates (Thermo Scientific) according to the manufacturer’s instructions. The Bradford assay was used to measure protein concentration, and the proteins were aliquoted at 8 μM and flash-frozen with liquid nitrogen.
For crystallography with ppGpp, recombinant B. anthracis Hpt-1 was purified as described above followed by dialysis with His-tagged TEV protease in 10 mM Tris-HCl (pH 7.5), 100 mM NaCl, and 1 mM DTT. The dialyzed protein was incubated with Ni-NTA beads for 30 min, the beads were centrifuged and the supernatant was run over a HiPrep Sephacryl 16/60 S-100 HR column (GE Healthcare) in 10 mM Tris-HCl pH 8.3, 250 mM NaCl, and 1 mM DTT. Relevant fractions were concentrated to ≈10 mg/mL. For crystallography with substrates, the same protocol was followed but with 10 mM Tris-HCl pH 8 and 100 mM NaCl as the size exclusion buffer.
Enzyme inhibition assays
Request a detailed protocolHPRT activity assays were performed as described previously (Biazus et al., 2009; Kriel et al., 2012; Xu et al., 1997). The standard assay was performed at 25°C and contained 100 mM Tris-HCl (pH 7.4), 12 mM MgCl2, 1 mM PRPP (MilliporeSigma), 50 μM guanine or hypoxanthine (MilliporeSigma), and 20 nM HPRT (measured by monomer). Reactions were initiated with the purine base and monitored in a spectrophotometer (Shimadzu UV-2401PC) at 257.5 nm or 245 nm for conversion of guanine to GMP or hypoxanthine to IMP, respectively. A difference in extinction coefficients of 5900 M−1cm−1 was used for GMP and guanine and 1900 M−1cm−1 for IMP and hypoxanthine. For inhibition curves, assays were performed at the substrate concentrations listed above and at variable pppGpp concentrations. (p)ppGpp was synthesized as described (Liu et al., 2015b). Initial velocities of the inhibited reactions were normalized to the uninhibited initial velocity prior to fitting to the equation Y = 1/(1 + (x / IC50)s) to calculate IC50. GMK activity assays were performed as previously described (Liu et al., 2015b), except an enzyme concentration of 20 nM and a substrate concentration of 100 μM GMP were used. Due to lower activity of the E. coli ΔCT-tail variant, assays were performed at 200 nM enzyme rather than 20 nM enzyme for E. coli GMK.
To test inhibition of HPRT homologs (see Figure 1), reactions were performed in a Synergy Two microplate reader (BioTek). The assay was performed at 25°C with 50 μM hypoxanthine, 1 mM PRPP, and 100 nM HPRT (measured by monomer) in the same reaction buffer as above. For Coxiella burnetii HPRT, 50 μM guanine was used as the substrate since its activity was very low with hypoxanthine. Reactions were performed in triplicate without (p)ppGpp, with 25 μM ppGpp, and with 25 μM pppGpp. Reaction rates from the first-order kinetic curves were determined using R (v 3.4.3).
For kinetic studies, pppGpp concentrations were varied between 3.125 and 50 μM and PRPP concentrations were varied between 0.03125 and 2.5 mM. Data were analyzed and fitted to a global Michaelis-Menten competitive inhibition model using GraphPad Prism v5.02. PRPP provided by MilliporeSigma was listed as ≥75% pure. For the purposes of kinetic experiments, the purity was assumed to be 100%.
Isothermal titration calorimetry
Request a detailed protocolExperiments were performed using the MicroCal iTC200 (GE Healthcare). B. anthracis Hpt-1 was dialyzed into the ITC buffer (10 mM HEPES pH 8, 150 mM NaCl, 10 mM MgCl2) with three buffer changes at 4°C. The concentration of protein was calculated using a molar extinction coefficient of 16390 M−1cm−1 and A280(true) (A280 – (1.96 × A330) (Pace et al., 1995). The experiments were performed at 25°C with 45.5 μM HPRT (measured by monomer), a reference power of 6 μCal/s, and a stirring speed of 1000 RPM. pppGpp was solubilized in dialysate from protein dialysis and its concentration was measured using a molar extinction coefficient of 13,700 M−1cm−1 and A253. pppGpp (500 μM) was titrated into B. anthracis Hpt-1 with the following: 1 × 1 μL (discarded), 19 × 2 μL. ITC between PRPP and Hpt-1 were performed in identical conditions. PRPP was solubilized in protein dialysate at a concentration of 1 mM. For data analysis, PRPP was assumed to be at a concentration of 820 μM, since the PRPP lot analysis from MilliporeSigma listed the PRPP as 82% pure. Data analysis and one-site binding modeling were performed using MicroCal Origin 5.0 software.
X-ray crystallography
Request a detailed protocolProteins were prepared for crystallography as described above. For crystals formed with pppGpp, B. anthracis Hpt-1 was concentrated to ≈10 mg/mL in 10 mM Tris pH 8.3, 250 mM NaCl, and 1 mM DTT. The pppGpp ligand was resuspended in ddH2O. MgCl2 was added to the protein at a final concentration of 1 mM and crystals were formed using hanging drop vapor diffusion with 900 μL of reservoir liquid in each well. Each drop contained 0.9 μL protein, 0.9 μL reservoir liquid, and 0.2 μL pppGpp (final concentration, 1.5 mM pppGpp). Crystals formed in 0.2 M ammonium tartrate dibasic pH 6.6% and 20% PEG 3350 in 3–6 months. Identical crystals formed in replicated conditions in 1–2 weeks. Crystals were soaked in reservoir liquid with 25% glycerol prior flash freezing in liquid nitrogen. Apo protein crystals formed within 1–2 days in multiple conditions with high sulfate concentrations. Using protein preparations from above, apo crystals formed in 0.01 M CoCl2, 0.1 M MES monohydrate pH 6.5, 1.8 M ammonium sulfate, and crystals were soaked in reservoir liquid with 25% glycerol for cryoprotection prior to freezing.
For crystals with substrates, B. anthracis Hpt-1 was concentrated to ≈10 mg/mL in 10 mM Tris pH 8 and 100 mM NaCl. Additives were diluted in the protein solution at final concentrations of 10 mM MgCl2, 2 mM PRPP, and 1 mM 9-deazaguanine (Santa Cruz Biotechnology) prior to crystallization. PRPP was resuspended in ddH2O and 9-deazaguanine was resuspended in 100% DMSO. Drops for hanging drop vapor diffusion comprised 1 μL crystal condition and 1 μL protein/ligand mixture. Crystals formed within 3 days in 0.2 M ammonium acetate, 0.1 M sodium acetate trihydrate pH 4.6, 30% PEG 4000. Reservoir solution with 25% ethylene glycol was added to the drops for cryoprotection prior to flash freezing in liquid nitrogen.
Diffraction data was collected at the Life-Science Collaborative Access Team (LS-CAT), beamline 21-ID-F (Hpt-1 with sulfates), 21-ID-G (Hpt-1 with ppGpp), and 21-ID-D (Hpt-1 with substrates) at the Advanced Photon Source (APS) at Argonne National Labs (Argonne, IL). Data was indexed and scaled using HKL2000 (Otwinowski and Minor, 1997). Phasing for Hpt-1 with sulfates and ppGpp was determined by molecular replacement with PDB ID 3H83 as a search model using Phenix (Adams et al., 2010). Phasing for Hpt-1 with substrates was determined by molecular replacement with Hpt-1-ppGpp as a search model using Phenix. Iterative model building with Coot and refinement with Phenix produced final models (Emsley and Cowtan, 2004).
Predicted oligomeric states of crystallographic assemblies were determined by PISA (‘Protein interfaces, surfaces, and assemblies’ service PISA at the European Bioinformatics Institute) (Krissinel and Henrick, 2007).
Size exclusion chromatography
Request a detailed protocolSize exclusion chromatography was performed using AktaPure and a Superose 12 10/300 GL column (GE Healthcare, Inc). A buffer consisting of 10 mM HEPES pH 8.0, 100 mM NaCl, and 10 mM MgCl2 was used to run ≈10–20 μM (0.2–0.5 mg/mL) protein over the column at 0.1–0.25 mL/min. Additional 1 mM DTT was used for proteins containing cysteines. For gel filtration with ligands, 500 μM PRPP and 100 μM 9-deazaguanine were included in the mobile phase buffer where necessary. A gel filtration standard (Bio-Rad) was used to establish molecular weight, and bovine serum albumin (BSA) was included as an additional marker.
Dimethyl adipimidate crosslinking
Request a detailed protocolCrosslinking was performed with ≈10 μM B. subtilis HPRT (measured by monomer), 20 mM dimethyl adipimidate (DMA) (Thermo Scientific), and 500 μM ligand. DMA was suspended in 25 mM HEPES pH 8.0, 100 mM NaCl, 10 mM MgCl2, and 10% glycerol, and the solution was buffered to pH 8.5. HPRT was dialyzed into 25 mM HEPES pH 7.5, 100 mM NaCl, and 10% glycerol. Ligands were incubated with protein for 10 min followed by a 15-min incubation with DMA at room temperature. Reactions were terminated with addition of 2X Laemmli buffer (Bio-Rad) for immediate analysis with SDS-PAGE (10% polyacrylamide gel). Gels were stained with SYPRO Ruby (Bio-Rad) according to the manufacturer’s protocol and imaged using a Typhoon FLA9000 (GE Healthcare).
Dynamic light scattering
Request a detailed protocolDynamic light scattering was performed using DynaPro99 (Protein Solutions/Wyatt Technologies). Readings were from a 20 μL solution (10 mM Tris-HCl pH 8, 100 mM NaCl, 10 mM MgCl2, 6.5% glycerol) with 4 mg/mL (≈175 μM) B. anthracis Hpt-1 and 2 mM ligands. Data were analyzed using Dynamics version 5.25.44 software.
DRaCALA
Request a detailed protocolDRaCALA was performed with pure protein and radioactive ligand as described (Roelofs et al., 2011). [5′ α-32P] pppGpp was synthesized according to modified protocols of non-radioactive and radioactive pppGpp syntheses (Corrigan et al., 2016; Hogg et al., 2004; Mechold et al., 2002). The reaction contained 25 mM bis-Tris propane (pH 9.0), 15 mM MgCl2, 0.5 mM DTT, 2 mM ATP, 2 μM RelSeq (1-385), and 37.5 μCi [α-32P] GTP (Perkin Elmer). The reaction was incubated at 37°C for 1 hr. The reaction was diluted in 0.5 mL of Buffer A (0.1 mM LiCl, 0.5 mM EDTA, 25 mM Tris-HCl pH 7.5) prior to adding to a 1 mL HiTrap QFF strong anion exchange column (GE Healthcare) equilibrated with 10 column volumes (CV) of Buffer A. The column was washed with 10 CV of Buffer A followed by an additional wash with 10 CV of 83% Buffer A + 17% Buffer B (Buffer B: 1 M LiCl, 0.5 mM EDTA, 25 mM Tris-HCl pH 7.5). 32P-pppGpp was eluted with a mixture of 50% Buffer A + 50% Buffer B. Fractions of 1 mL were collected from the elution.
DRaCALA reactions (20 μL) using purified HPRT contained 100 mM Tris-HCl (pH 7.4), 12 mM MgCl2, protein diluted to appropriate concentrations in 20 mM HEPES pH 8 and 100 mM NaCl, and 32P-pppGpp (1:100 final dilution of first elution fraction from 32P-pppGpp purification). DRaCALA reactions using GMK contained 100 mM Tris-HCl (pH 7.4), 100 mM KCl, and 10 mM MgCl2. Reactions with B. subtilis GMK and ΔCT-tail were performed with 20 μM protein and reactions with E. coli GMK and ΔCT-tail were performed with 40 μM protein. All protein concentrations were measured by monomer. For competition experiments between 32P-labeled pppGpp and PRPP, non-radioactive PRPP was resuspended in ddH2O and added at the concentrations specified. Protein was dialyzed or diluted into buffer lacking glycerol, as glycerol interferes with diffusion of the aqueous phase. Reactions were incubated at room temperature for 10 min. Two microliters from each reaction were spotted in duplicate on Protran BA85 nitrocellulose (GE Healthcare) via pipette or a replicator pinning tool (VP 408FP6S2; V and P Scientific, Inc). Spots were allowed to dry and radioactivity was detected with phosphorimaging (Typhoon FLA9000). Fraction bound of 32P-pppGpp was calculated as described (Roelofs et al., 2011). Data were analyzed in GraphPad Prism v5.02 and fitted to the equation Y = (Bmax ×Kd) / (Kd + X) (Roelofs et al., 2011). PRPP competition curves were fitted with a four-parameter logistical (4PL) model.
DRaCALA using cell lysates was adapted from a previous protocol (Roelofs et al., 2015). One milliliter of cells containing overexpressed recombinant HPRT was pelleted and resuspended in 100 μL of binding buffer (20 mM Tris-HCl pH 8, 100 mM NaCl, 12 mM MgCl2, 0.1 mM DTT) supplemented with 1 μM PMSF, 250 μg/mL lysozyme, and 10 μg/mL DNase I. Cells were lysed with three freeze/thaw cycles. In a 20 μL DRaCALA reaction, 10 μL of cell lysate was added to binding buffer and 32P-pppGpp. Reactions were performed and analyzed as above. For measuring binding affinity (Kd) of proteins in cell lysates, recombinant protein expression level in cell lysates was determined by comparing expression to a standard of purified B. subtilis HPRT co-resolved with SDS-PAGE.
Differential scanning fluorimetry
Request a detailed protocolDifferential scanning fluorimetry was performed using 10 μM protein (measured by monomer) in a buffer containing 20 mM HEPES pH 8, 100 mM NaCl, 10 mM MgCl2, 1 mM DTT, and 10X SYPRO orange dye (diluted from 5000X stock; MilliporeSigma). Proteins were mixed in an optically clear quantitative PCR (qPCR) 96-well plate and sealed with plastic film. Relative fluorescence intensity was monitored in a Bio-Rad qPCR machine using FRET detection over a temperature increase of 1°C/min from 25°C to 90°C. Data were analyzed using GraphPad Prism.
Thin layer chromatography
Request a detailed protocolThin layer chromatography was performed as described previously (Bittner et al., 2014; Schneider et al., 2003). Cells were grown in a low-phosphate medium with casamino acids (S7 defined medium [Vasantha and Freese, 1980] supplemented with 0.1% glutamate, 1% glucose, and 0.5% casamino acids; low-phosphate medium contained one-tenth the phosphate concentration as S7 defined medium). JDW2121 and JDW2128 were grown with 1 mM IPTG to induce expression of exogenous phosphoribosyltransferases. Cells were labeled with 50 μCi/ml 32P orthophosphate (900 mCi/mmol; PerkinElmer) at an OD ≈ 0.02–0.05 and grown to an OD ≈ 0.15–0.2 prior to treating with 1 mM guanosine (MilliporeSigma). Guanosine stock was 100 mM diluted in 100% DMSO. At given timepoints following guanosine addition, nucleotides were extracted by adding 100 μl of sample to 20 μl 2 N formic acid and incubating on ice for at least 20 min. Samples were centrifuged at max speed for 15 min and supernatant (≈60 μl) was transferred to a new tube. Samples were spotted on PEI cellulose plates (MilliporeSigma) and developed in 1.5 M KH2PO4 pH 3.4. Plates were exposed to a phosphor screen and scanned on a Typhoon scanner. Nucleotide levels were quantified by subtracting background signal in each lane and expressing the level as a ratio to the untreated sample (t = 0).
Protein sequence analysis and ancestral sequence reconstruction
Request a detailed protocolTo construct a 16S rRNA tree, sequences were obtained from the Ribosomal Database Project (Cole et al., 2014) or from NCBI. Sequences were aligned with ClustalW in MEGA X. MEGA X was used to identify the best substitution model for evolution as the General Time Reversible model with gamma distributed substitution rates. The maximum likelihood phylogenetic tree was constructed with this model in MEGA X with 1000 bootstrap replicates. Eukaryotic 18S rRNA sequences for H. sapiens, C. elegans, and S. cerevisiae were used as the outgroup.
For HPRT phylogenetic analysis, protein sequences were obtained in UniProt either by searching for HPRT’s enzyme classification number (EC 2.4.2.8) in a given organism or by using NCBI’s and UniProt’s BLAST algorithms to find HPRTs similar (>50% identity) to a model organism representing different clades (e.g. B. subtilis, E. coli, S. aureus, S. coelicolor, B. thetaiotaomicron, L. pneumophila, P. aeruginosa, T. gondii, C. elegans, H. sapiens). Protein sequences were aligned using MUSCLE in MEGA X with a gap-opening penalty of −3.2 and a gap-extending penalty of −0.2. See Figure 6—source data 1 for the complete alignment. The most appropriate evolutionary model for amino acid substitution was identified using ProtTest 3 (Darriba et al., 2011) as the Le and Gascuel model (Le and Gascuel, 2008) with gamma distributed substitution rates (four categories) and invariant sites. The maximum likelihood phylogenetic tree was constructed using this substitution model in MEGA X with 100 bootstrap replicates and a nearest neighbor interchange topology search. The clade containing eukaryotic HPRTs was used as the outgroup. Ancestral HPRT sequences were reconstructed using a marginal probability reconstruction in MEGA X with the phylogenetic tree and the same substitution model used to obtain the tree (Yang et al., 1995).
For GMK phylogenetic analysis, protein sequences were obtained from UniProt by searching for EC 2.7.4.8 in the organisms studied in Liu et al. (2015b). Related organisms were identified with UniProt’s BLAST algorithm. Protein sequences were aligned using MUSCLE in MEGA X with default settings. See Figure 8—source data 1 for the complete alignment. ProtTest3 identified the most appropriate substitution model as the Le and Gascuel model (Le and Gascuel, 2008) with gamma distributed substitution rates (four categories) and invariant sites. The phylogenetic tree and ancestral reconstruction were performed as they were with HPRT.
Data availability
Diffraction data have been deposited in PDB under the accession codes 6D9Q (https://www.rcsb.org/structure/6d9q), 6D9R (https://www.rcsb.org/structure/6d9r), and 6D9S (https://www.rcsb.org/structure/6D9S). All data generated or analysed during this study are included in the manuscript and supporting files. Source data files have been provided for Table 2.
-
Protein Data BankID 6D9Q. The sulfate-bound crystal structure of HPRT (hypoxanthine phosphoribosyltransferase).
-
Protein Data BankID 6D9R. The substrate-bound crystal structure of HPRT (hypoxanthine phosphoribosyltransferase).
-
Protein Data BankID 6D9S. The (p)ppGpp-bound crystal structure of HPRT (hypoxanthine phosphoribosyltransferase).
-
Protein Data BankID 5ESW. Crystal structures of Apo and GMP bound hypoxanthine-guanine phosphoribosyltransferase from Legionella pneumophila and the implications in gouty arthritis.
-
Protein Data BankID 5ESX. Crystal structures of Apo and GMP bound hypoxanthine-guanine phosphoribosyltransferase from Legionella pneumophila and the implications in gouty arthritis.
-
Protein Data BankID 3H83. 2.06 Angstrom resolution structure of a hypoxanthine-guanine phosphoribosyltransferase (hpt-1) from Bacillus anthracis str. 'Ames Ancestor'.
References
-
"System 48" high-throughput cloning and protein expression analysisMethods in Molecular Biology 498:117–127.https://doi.org/10.1007/978-1-59745-196-3_8
-
PHENIX : a comprehensive Python-based system for macromolecular structure solutionActa Crystallographica Section D Biological Crystallography 66:213–221.https://doi.org/10.1107/S0907444909052925
-
Protein oligomerization: how and whyBioorganic & Medicinal Chemistry 13:5013–5020.https://doi.org/10.1016/j.bmc.2005.05.037
-
Absolute metabolite concentrations and implied enzyme active site occupancy in Escherichia coliNature Chemical Biology 5:593–599.https://doi.org/10.1038/nchembio.186
-
A possible role of purine nucleotide pyrophorylases in the regulation of purine uptake by Bacillus subtilisThe Journal of Biological Chemistry 241:2679–2686.
-
Hypoxanthine-guanine phosphoribosyltransferase from Mycobacterium tuberculosis H37Rv: cloning, expression, and biochemical characterizationProtein Expression and Purification 66:185–190.https://doi.org/10.1016/j.pep.2009.04.001
-
BookThe Stringent ResponseIn: Neidhardt FC, Curtiss R, Ingraham JL, Lin ECC, Low KB, Magasanik B, Reznikoff W, Riley M, Schaechter M, Umbarger HE, editors. Escherichia Coli and Salmonella Cellular and Molecular Biology. American Society for Microbiology. pp. 1458–1496.
-
Purine-rich foods, dairy and protein intake, and the risk of gout in menNew England Journal of Medicine 350:1093–1103.https://doi.org/10.1056/NEJMoa035700
-
Coordination of microbial metabolismNature Reviews Microbiology 12:327–340.https://doi.org/10.1038/nrmicro3238
-
Ribosomal database project: data and tools for high throughput rRNA analysisNucleic Acids Research 42:D633–D642.https://doi.org/10.1093/nar/gkt1244
-
Causes of evolutionary rate variation among protein sitesNature Reviews Genetics 17:109–121.https://doi.org/10.1038/nrg.2015.18
-
Coot: model-building tools for molecular graphicsActa Crystallographica. Section D, Biological Crystallography 60:2126–2132.https://doi.org/10.1107/S0907444904019158
-
First crystal structures of Mycobacterium tuberculosis 6-Oxopurine phosphoribosyltransferase: complexes with GMP and pyrophosphate and with acyclic nucleoside phosphonates whose prodrugs have antituberculosis activityJournal of Medicinal Chemistry 58:4822–4838.https://doi.org/10.1021/acs.jmedchem.5b00611
-
A family of LIC vectors for high-throughput cloning and purification of proteinsMethods in Molecular Biology 498:105–115.https://doi.org/10.1007/978-1-59745-196-3_7
-
Structural and functional analysis of BipA, a regulator of virulence in enteropathogenic Escherichia coliJournal of Biological Chemistry 290:20856–20864.https://doi.org/10.1074/jbc.M115.659136
-
Many means to a common end: the intricacies of (p)ppGpp metabolism and its control of bacterial homeostasisJournal of Bacteriology 197:1146–1156.https://doi.org/10.1128/JB.02577-14
-
Synthesis of Guanosine Tetra- and pentaphosphates by the obligately anaerobic bacterium Bacteroides thetaiotaomicron in response to molecular oxygenJournal of Bacteriology 137:956–962.
-
Transcriptional responses to ppGpp and DksAAnnual Review of Microbiology 72:163–184.https://doi.org/10.1146/annurev-micro-090817-062444
-
PCR-mediated deletion of plasmid DNAAnalytical Biochemistry 375:373–375.https://doi.org/10.1016/j.ab.2007.12.005
-
Reconstructing ancient proteins to understand the causes of structure and functionAnnual Review of Biophysics 46:247–269.https://doi.org/10.1146/annurev-biophys-070816-033631
-
The regulation of purine utilization in Bacteria. V. inhibition of purine phosphoribosyltransferase activities and purine uptake in isolated membrane vesicles by Guanosine tetraphosphateThe Journal of Biological Chemistry 247:7067–7072.
-
Phosphoribosyl diphosphate (PRPP): Biosynthesis, enzymology, utilization, and metabolic significanceMicrobiology and Molecular Biology Reviews 81:e00040.https://doi.org/10.1128/MMBR.00040-16
-
The conservation profile of a protein bears the imprint of the molecule that is evolutionarily coupled to the proteinProteins: Structure, Function, and Bioinformatics 83:1407–1413.https://doi.org/10.1002/prot.24809
-
A new view of the tree of lifeNature Microbiology 1:1–6.https://doi.org/10.1038/nmicrobiol.2016.48
-
BookMetabolism of 5-phosphoribosyl 1-pyrophosphate (PRPP) in Escherichia coli and Salmonella typhimuriumIn: Munch-Petersen A, editors. Metabolism of Nucleotides, Nucleosides and Nucleobases in Microorganisms. London: Academic Press. pp. 1–25.
-
Total purine and purine base content of common foodstuffs for facilitating nutritional therapy for gout and hyperuricemiaBiological and Pharmaceutical Bulletin 37:709–721.https://doi.org/10.1248/bpb.b13-00967
-
Inference of macromolecular assemblies from crystalline stateJournal of Molecular Biology 372:774–797.https://doi.org/10.1016/j.jmb.2007.05.022
-
An improved general amino acid replacement matrixMolecular Biology and Evolution 25:1307–1320.https://doi.org/10.1093/molbev/msn067
-
Diversity in (p)ppGpp metabolism and effectorsCurrent Opinion in Microbiology 24:72–79.https://doi.org/10.1016/j.mib.2015.01.012
-
Structural similarity enhances interaction propensity of proteinsJournal of Molecular Biology 365:1596–1606.https://doi.org/10.1016/j.jmb.2006.11.020
-
Protein Dimerization and Oligomerization in Biology1–18, Dimers, oligomers, everywhere, Protein Dimerization and Oligomerization in Biology, New York, Springer.
-
Evolutionary studies of ligand binding sites in proteinsCurrent Opinion in Structural Biology 45:85–90.https://doi.org/10.1016/j.sbi.2016.11.024
-
Structural characterisation and functional significance of transient protein-protein interactionsJournal of Molecular Biology 325:991–1018.https://doi.org/10.1016/S0022-2836(02)01281-0
-
Processing of X-ray diffraction data collected in oscillation modeMethods in Enzymology 276:307–326.https://doi.org/10.1016/S0076-6879(97)76066-X
-
Effect of unusual guanosine nucleotides on the activities of some Escherichia coli cellular enzymesBiochimica Et Biophysica Acta (BBA) - General Subjects 677:358–362.https://doi.org/10.1016/0304-4165(81)90247-6
-
Structural basis for (p)ppGpp-mediated inhibition of the GTPase RbgAJournal of Biological Chemistry 293:19699–19709.https://doi.org/10.1074/jbc.RA118.003070
-
The emergence of protein complexes: quaternary structure, dynamics and allostery. Colworth medal lectureBiochemical Society Transactions 40:475–491.https://doi.org/10.1042/BST20120056
-
Mechanisms of cooperativity and allosteric regulation in proteinsQuarterly Reviews of Biophysics 22:139–237.https://doi.org/10.1017/S0033583500003826
-
ppGpp is the major source of growth rate control in E. coliEnvironmental Microbiology 13:563–575.https://doi.org/10.1111/j.1462-2920.2010.02357.x
-
The PRT protein familyCurrent Opinion in Structural Biology 11:733–739.https://doi.org/10.1016/S0959-440X(01)00274-3
-
The magic dance of the alarmones (p)ppGppMolecular Microbiology 101:531–544.https://doi.org/10.1111/mmi.13412
-
A new vector for high-throughput, ligation-independent cloning encoding a tobacco etch virus protease cleavage siteProtein Expression and Purification 25:8–15.https://doi.org/10.1006/prep.2001.1603
-
Dissociation of enzyme oligomers: a mechanism for allosteric regulationCritical Reviews in Biochemistry and Molecular Biology 29:125–163.https://doi.org/10.3109/10409239409086799
-
Enzyme changes during Bacillus subtilis sporulation caused by deprivation of guanine nucleotidesJournal of Bacteriology 144:1119–1125.
-
Affinity-based capture and identification of protein effectors of the growth regulator ppGppNature Chemical Biology 15:141–150.https://doi.org/10.1038/s41589-018-0183-4
-
A new method of inference of ancestral nucleotide and amino acid sequencesGenetics 141:1641–1650.
Article and author information
Author details
Kenneth A Satyshur

Funding
National Institute of General Medical Sciences (R35 GM127088)
- Jue D Wang
Howard Hughes Medical Institute (Faculty Scholar)
- Jue D Wang
National Science Foundation (GRFP DGE-1256259)
- Brent W Anderson
National Institutes of Health (R01 GM084003)
- Jue D Wang
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank Esra Gumuser and Matthew Macgilvray for technical help, and members of the Wang lab for feedback on this manuscript. We thank Federico Rey, Bob Kerby, JD Sauer, Dan Pensinger, Helen Blackwell, and Kayleigh Nyffeler for assistance in obtaining bacterial genomic DNA. We thank Katrina Forest for assistance with dynamic light scattering. This research used resources of the Advanced Photon Source, a U.S. Department of Energy (DOE) Office of Science User Facility operated for the DOE Office of Science by Argonne National Laboratory under Contract No. DE-AC02-06CH11357. Use of Life Sciences Collaborative Access Team beamline 21-ID-D was supported by the Michigan Economic Development Corporation and the Michigan Technology Tri-Corridor (Grant 085P1000817). This work was funded by NIH R35 GM127088 and R01 GM084003 to JDW, and BWA was supported by NSF GRFP DGE-1256259.
Copyright
© 2019, Anderson et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 2,005
- views
-
- 290
- downloads
-
- 37
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 37
- citations for umbrella DOI https://doi.org/10.7554/eLife.47534
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Evolution of (p)ppGpp-HPRT regulation through diversification of an allosteric oligomeric interaction
eLife 8:e47534.
https://doi.org/10.7554/eLife.47534




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}