A novel role for lipoxin A4 in driving a lymph node–eye axis that controls autoimmunity to the neuroretina
- Vision Science Program, University of California, Berkeley, United States
- Laboratory of Immunology, National Eye Institute, National Institutes of Health, United States
- School of Optometry, University of California, Berkeley, United States
- Infectious Disease and Immunity Program, University of California, Berkeley, United States
Peer review process
This article was accepted for publication as part of eLife's original publishing model.
History
- Version of Record published
- Accepted Manuscript published
- Accepted
- Received
Decision letter
-
Tadatsugu TaniguchiSenior Editor; Institute of Industrial Science, The University of Tokyo, Japan
-
Lois SmithReviewing Editor; Boston Children's Hospital/Harvard Medical School, United States
-
Nicolas G BazanReviewer; Louisiana State University Health Sciences Center, United States
In the interests of transparency, eLife publishes the most substantive revision requests and the accompanying author responses.
Acceptance summary:
In experimental (mouse) autoimmune uveitis the authors found a basal level of the lipid mediator LXA4 in lymph nodes that decreases in the draining lymph node during adaptive immune responses. LXA4, a Specialized Pro-resolving Mediator, has a clinically relevant role in regulating acute inflammation and inflammatory resolution.
Decision letter after peer review:
Thank you for submitting your article "A novel role for lipoxin A4 in driving a lymph node-eye axis that controls autoimmunity to the neuroretina" for consideration by eLife. Your article has been reviewed by three peer reviewers, one of whom is a member of our Board of Reviewing Editors, and the evaluation has been overseen by Tadatsugu Taniguchi as the Senior Editor. The following individual involved in review of your submission has agreed to reveal their identity: Nicolas G Bazan (Reviewer #2).
The reviewers have discussed the reviews with one another and the Reviewing Editor has drafted this decision to help you prepare a revised submission. Please aim to submit the revised version within two months.
Summary:
Wei et al. conducted experiments to determine whether and how LXA4 regulates T effector cell function in the pathogenesis of autoimmune uveitis. They showed that LXA4 serum concentration may be increased in posterior uveitis patients, and the disease severity can be altered by modulating LXA4 concentration, i.e. ALOX5-deficient mice and LXA4 injection in the mouse experimental autoimmune uveitis (EAU) model.
These findings clearly provide evidence for the involvement of LXA4 signaling pathways in the pathogenesis of autoimmune uveitis. However, the study falls a little short to provide sufficient mechanistic insight how LXA4 regulates T effector cell function in the pathogenesis of EAU. The main criticism is that it is not clear where is the specific site of LXA4 action to regulate T cell function. They suggest that the resident LXA4 circuit in inguinal lymph node plays roles, but it is not clear if the LXA4 signaling is up-regulated or down-regulated in the inguinal lymph node during the EAU, since LXA4 concentration goes down while All/Fpr2 expression goes up (Figure 1C and 1E). After LXA4 i.p. injections, LXA4 concentration should be determined in eyes, inguinal lymph, and serum. 5-LOX is involved in the production of not only LXA4, but also leukotriene species. Also, the Cox pathway might be augmented in the compensatory manner in the 5-LOX-deficient background. So it is not clear whether the EAU phenotype observed in the ALOX4-/- mice can be attributed to LXA4 alone or not. In the adoptive transfer experiment of T cells, they transfer the T cells from the EAU mice. These T cells are already activated in the global ALOX5-/- environment, and do not support the conclusion that effect of LXA4 is T cell mediated.
As potential mechanisms, they suggest the regulations by LXA4 of chemokine receptors and sphingosine-1 receptor expression as well as glucose metabolism. These observations are descriptive, only based on the gene expression levels and glycolytic rate, not providing the direct regulation by LXA4.
Essential revisions:
1) Figure 1A.
It is important to see if the rodent findings translate to human inflammation / immune responses. So this figure is critical. Within the "other" uveitis group was there a difference between "pan uveitis" which also has posterior (choroidal) uveitis and "anterior uveitis"? Anterior uveitis alone may be a better control if you suggest that specific ocular tissue inflammation determines the level of LXA4 and thus pan uveitis should be added to the posterior group. If on the other hand you suggest the instigating insult is associated with LXA4 levels then more explanation about the choices of categories of human disease is needed particularly because the difference between groups you currently have defined is small. An unbiased approach (put all human data together and see which patient groups are associated with what LXA4 levels) would be the most telling. "Dot plots" should be done as in the rest of the paper as it would give more information.
2) There are great differences between mouse and human inflammation/immune responses (Seok et al., 2013). Please comment in detail in the Discussion and Introduction with respect to this work.
3) Figure 2.
Alox5-/- mice lack more than LXA4. What are the possible effects of other metabolic products that are missing in these mice?
What was the diet composition for the mice specifically regarding omega 3 and 6 LCPUFAs and was it maintained throughout as standard chow can vary batch to batch? A diet high in AA versus DHA would likely influence results.
Figure 2D The authors are presenting a single point for the electroretinography analysis, for which we have no indication on the light intensity used to get this b-wave response. In Materials and methods it is mentioned that the ERG was performed with controlled light stimuli (subsection “Electroretinography and fundus imaging”), meaning that more than one stimulus was performed, however this figure presents the response from only one intensity point. Moreover, the aforementioned subsection mentions a-wave, which are not apparent in this figure, nor discussed in Results. A complete ERG plot with increasing light stimuli showing a-wave and b-wave would be more appropriate.
4) Figure 3.
Figure 2G and Figure 3H Are the statistical analysis missing in these graphs or the differences are not significant? If the latter mention it in the legend.
Figure 3C What is the mouse strain background of EAU and EAU+LXA4 animals, because their RPE is not pigmented. This is not clear, because according to the subsection “Mice, EAU induction and LXA4 treatment”, C57BL/6J (pigmented animals) were used for the EAU induction and LXA4 treatment. Can the authors clarify the reason of the lack of pigmentation in the EAU and EAU+LXA4 pictures?
Figure 3D Can the authors improve the quality of their immunofluorescent pictures? For example, the DAPI staining is barely visible, which is essential for the identification of the layering of the retina and proper morphological localization of the labeled targets. Moreover, a thicker line for the scale bar to make it more visible would help improve this picture.
5) Mechanism.
As potential mechanisms, they suggest the regulations by LXA4 of chemokine receptors and sphingosine-1 receptor expression as well as glucose metabolism. These observations are descriptive, only based on the gene expression levels and glycolytic rate, not providing the direct regulation by LXA4.
To strengthen their conclusion, additional experiments listed below are needed.
1) Provide LXA4 concentrations in the eye, inguinal lymph node, and serum after i.p. injection of LXA4 during the EAU model.
2) Provide evidence or discussion to show that other lipid mediators (i.e. leukotrienes and prostaglandins) are not affected in the ALOX5-deficient mice.
3) Examine if LXA4 treatment of isolated T cells in vitro can impact the gene expression levels of chemokine receptors and S1pr1, like they did in the glycolysis analysis (Figure 6).
https://doi.org/10.7554/eLife.51102.sa1Author response
Essential revisions:
1) Figure 1A.
It is important to see if the rodent findings translate to human inflammation / immune responses. So this figure is critical. Within the "other" uveitis group was there a difference between "pan uveitis" which also has posterior (choroidal) uveitis and "anterior uveitis"? Anterior uveitis alone may be a better control if you suggest that specific ocular tissue inflammation determines the level of LXA4 and thus pan uveitis should be added to the posterior group. If on the other hand you suggest the instigating insult is associated with LXA4 levels then more explanation about the choices of categories of human disease is needed particularly because the difference between groups you currently have defined is small. An unbiased approach (put all human data together and see which patient groups are associated with what LXA4 levels) would be the most telling. "Dot plots" should be done as in the rest of the paper as it would give more information.
As the reviewers suggested we reanalyzed the human serum data. Detailed analysis of all uveitis groups identified patients with 13 distinct diagnoses. Only 1 patient had a diagnosis of anterior uveitis. All other uveitides can include posterior uveitis and/or immune responses as part of the pathogenesis. In total there were 4 uveitis variants with only 1 patient (edema, retinal detachment, anterior uveitis, photopsia), which were excluded from the new analysis. All other uveitides ranged from 2 to 23 patient serum samples per group. Decoding of the samples also revealed that our original dataset erroneously included a number of plasma (and not serum) samples, which skewed the mean of the uveitis groups. After excluding the plasma samples, the new analysis revealed an opposite result from our original finding, namely serum LXA4 levels in patients with posterior uveitis symptoms (n=78) were significantly lower (p=0.0092) than in healthy controls (n=41). Revised Figure 1A shows significantly lower serum levels of LXA4 when all posterior uveitis patients were compared to healthy controls. All individual diagnosis groups had means that were lower than the healthy controls, with the exception of Vogt-Koyanagi-Harada disease (VKH, n=9). Due to the variation in sample size in each group, differences between individual diagnosis groups and healthy controls did not reach significance, except for intermediate uveitis (n=4, p=0.04). We are fortunate to have access to these human samples, but it is a post-hoc analysis of all available serum samples and we cannot increase the number of samples for individual diagnosis groups to establish which type of uveitis or pathogenesis correlates with the most significant decrease in serum LXA4 levels. Our findings add to a growing body of evidence that has recently been published by the Serhan and Dalli research groups (1-4), which document dynamic and disease-specific changes in SPM serum or plasma levels in humans. A new section for Figure 1A (subsection “LXA4 is generated during autoimmune uveitis in a time- and site- dependent manner”) has been added to Results and corresponding discussion (Discussion, first paragraph) has been added.
2) There are great difference between mouse and human inflammation/ immune responses (Seok et al., 2013). Please comment in detail in the Discussion and Introduction with respect to this work.
We fully agree that there are differences in mouse and human inflammation and immune responses and that all findings from mouse models have to eventually be validated in humans. As requested by the reviewer we added the PNAS reference that compared genomic responses in mice and humans and added discussion to highlight the limitations of our mouse model (Discussion, seventh paragraph). It is important point out that despite limitations of mouse models, major breakthroughs in our understanding of human immunology and disease treatment were discovered in mouse and invertebrate models (e.g., 2011 and 2018 Nobel Prize in Physiology or Medicine).
3) Figure 2.
Alox5-/- mice lack more than LXA4. What are the possible effects of other metabolic products that are missing in these mice?
5-LOX is also the required enzyme for generation leukotrienes. LTB4 and peptido-leukotriene (LTC4/D4/E4) formation is impaired in Alox5-/- mice. LTB4 has established roles in regulation of innate immune cells and Th2 responses. However, in the Th1/17 -driven autoimmune uveitis model, peptido-leukotrienes were not consistently detected in tissues and LTB4 levels did not significantly change in lymph nodes or the eye during EAU development (new Figure 2—figure supplement 1), which is why our study focused on the 5-LOX metabolite LXA4. Our previous studies (5, 6), which investigated SPM formation in draining lymph nodes during the initiation of ocular surface immune responses, also did not detect significant levels or changes in LTB4. 5-LOX is also the required enzyme for the formation of several DHA-, DPA- and EPA-derived SPM; however, these were not consistently detected in lymph nodes or the eye with our LC-MS/MS method. New text has been added to the Results section (subsection “LXA4 is generated during autoimmune uveitis in a time- and site- dependent manner”, second paragraph) to justify our focus on LXA4 as the most relevant 5-LOX metabolite in the EAU model.
What was the diet composition for the mice specifically regarding omega 3 and 6 LCPUFAs and was it maintained throughout as standard chow can vary batch to batch? A diet high in AA versus DHA would likely influence results.
All animals were maintained on the NIH-31 Open Formula diet with 1.9% C18:2 ω6 linoleic acid and 0.2% C18:3 ω3 linolenic acid (Envigo, WI). Vendor, name and PUFA composition of the standard rodent chow have been added to the Materials and methods section. Mice were housed in the same animal room and experienced the same controlled environmental conditions. Control and experimental animals were on the same open formula NIH rodent diet from weaning through the duration of the experiment thus batch to batch variation in the diet would equally affect all experimental and control groups. We agree that batch to batch variation in rodent diets could potentially change the ratio of dietary intake of AA:DHA:EPA. However, in all our tissue lipidomic analyses AA, DHA and EPA are included as standard analytes and we did not notice any marked changes in the ratio of AA:DHA:AA among different sets of mouse experiments.
Figure 2D The authors are presenting a single point for the electroretinography analysis, for which we have no indication on the light intensity used to get this b-wave response. In Materials and methods it is mentioned that the ERG was performed with controlled light stimuli (subsection “Electroretinography and fundus imaging”), meaning that more than one stimulus was performed, however this figure presents the response from only one intensity point. Moreover, the aforementioned subsection mentions a-wave, which are not apparent in this figure, nor discussed in the Results. A complete ERG plot with increasing light stimuli showing a-wave and b-wave would be more appropriate.
Only one light intensity was used per condition. Light stimulus of 10cds/m2 was used for a-wave and b-wave. Details for light stimuli have been added to the Materials and methods section and a-wave data has been added to revised Figure 2D.
4) Figure 3.
Figure 2G and Figure 3H Are the statistical analysis missing in these graphs or the differences are not significant? If the latter mention it in the legend.
No statistical analyses were performed in these graphs because error bars depict n=3 technical replicates from pooled n=6-9 animals per experimental group. Using ANOVA, stimulation index of proliferation assays in 2G and 3H show statistical significance between EAU vs. EAU+ LXA4 groups, as well as WT vs. Alox5-/- groups (Author response image 1). Showing both stimulation index as well as cpm in the manuscript would be redundant, therefore we are appending the figures in the rebuttal only.
Author response image 1

Stimulation index of proliferation assays from A) vehicle vs LXA4 treated EAU-challenged mice, p= 0.0154 and B) WT vs Alox5-/- EAU-challenged mice, p= 0.0248.
Two-way ANOVA.
Figure 3C What is the mouse strain background of EAU and EAU+LXA4 animals, because their RPE is not pigmented. This is not clear, because according to the subsection “Mice, EAU induction and LXA4 treatment”, C57BL/6J (pigmented animals) were used for the EAU induction and LXA4 treatment. Can the authors clarify the reason of the lack of pigmentation in the EAU and EAU+LXA4 pictures?
The mouse strain for all experiments is C57BL6, lack of pigmentation was due to poor imaging from an old microscope. Slides are now digitally scanned in high resolution and images have been replaced.
Figure 3D Can the authors improve the quality of their immunofluorescent pictures? For example, the DAPI staining is barely visible, which is essential for the identification of the layering of the retina and proper morphological localization of the labeled targets. Moreover, a thicker line for the scale bar to make it more visible would help improve this picture.
Quality of immunofluorescent images has been improved. DAPI fluorescence was enhanced and scale bar thickened in images.
5) Mechanism.
As potential mechanisms, they suggest the regulations by LXA4 of chemokine receptors and sphingosine-1 receptor expression as well as glucose metabolism. These observations are descriptive, only based on the gene expression levels and glycolytic rate, not providing the direct regulation by LXA4.
To strengthen their conclusion, additional experiments listed below are needed.
1) Provide LXA4 concentrations in the eye, inguinal lymph node, and serum after i.p. injection of LXA4 during the EAU model.
It is not possible to capture changes in LXA4 tissue levels after a low dose (1 μg) injection of LXA4 at the time of tissue harvest since, increases in eye or lymph node LXA4 levels are likely rapid and of short-duration (minutes). LXA4, like all eicosanoids, is enzymatically inactivated in minutes when not bound to an endogenous carrier protein such as albumin. However, several research groups have established that tail vein, intraperitoneal, subcutaneous or subconjunctival injections of LXA4 or other SPMs (10 ng-1 mg) in mouse disease models induce long-lasting protective actions through changes in lymphocyte and PMN function and macrophage phenotype at distant tissue sites. We did measure LXA4 in inguinal lymph nodes in LXA4 or vehicle control treated mice on day 10 and day 16 in the pathogenesis of EAU (Author response image 2). Since the tissue was collected 2-3 hours after the last LXA4 treatment, as expected, LXA4 tissue level did not change. We added discussion (Discussion, second paragraph) to clarify that we cannot directly prove that LXA4 treatment is increasing LXA4 levels in the inguinal lymph nodes or the eye.
Author response image 2
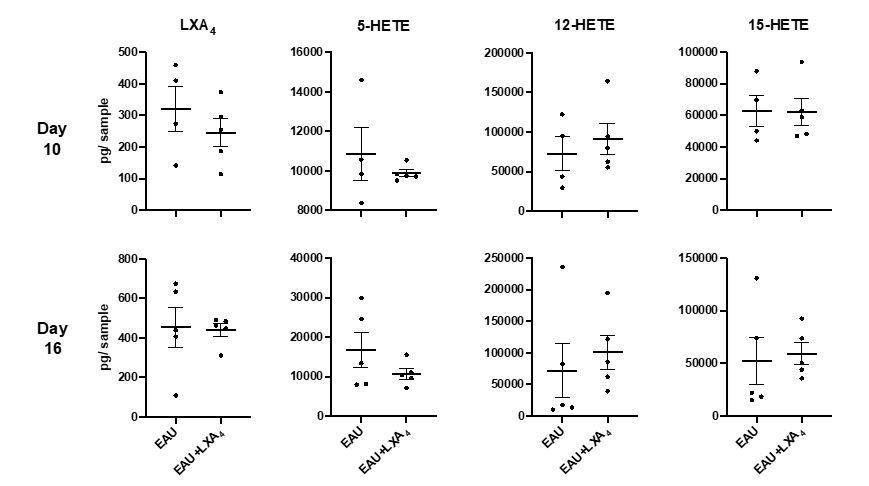
Quantification of LXA4 in EAU-challenged mice treated with LXA4 or vehicle control.
LXA4, 5-HETE, 12-HETE, 15-HETE in pg per sample in inguinal lymph nodes of EAU-challenged mice harvested on day 10 and day 16 post-immunization, quantified by LC-MS/MS. n=4-5 per group.
2) Provide evidence or discussion to show that other lipid mediators (i.e. leukotrienes and prostaglandins) are not affected in the ALOX5-deficient mice.
The Alox5-/- mouse line used in our experiments was generated in 1994 (7) and is an established Jackson Laboratory strain that has been used in numerous publications to study the 5-LOX pathway in disease models. Deficient LTB4, peptido-leukotrienes and LXA4 formation has been established for this mouse line. We are not aware of any publications that have reported marked compensatory upregulation of other eicosanoid pathways (COX, 12-LOX or 15-LOX) in this mouse line. In response to the reviewer’s concern, we performed lipidomic analysis (new Figure 2—figure supplement 1 and Results subsection “LXA4 limits development and progression of EAU”) but did not detect significant differences in basal inguinal lymph node levels of 12-HETE, 15-HETE, PGE2 or PGD2 in Alox5-/- mice. In addition, in WT mice LTB4 levels did not significantly change in lymph nodes or the eye in EAU (new Figure 1—figure supplement 1B). Therefore, we focused on LXA4, which is selectively down-regulated in inguinal lymph nodes during the development of an adaptive immune response.
3) Examine if LXA4 treatment of isolated T cells in vitro can impact the gene expression levels of chemokine receptors and S1pr1, like they did in the glycolysis analysis (Figure 6).
As requested, we have carried out additional experiments to examine the direct action of LXA4 with T cells in vitro. Flow plots shown in the new Figure 4F-G and Results (subsection “LXA4 modulates T cell trafficking in inguinal lymph nodes that drain the immunization site”) show that consistent with our hypothesis, LXA4 treatment of anti-CD3 and anti-CD28 stimulated T cells leads to increase in CCR7 and decrease in S1PR1 expression.
References:
1) Arnardottir, H. H., Dalli, J., Norling, L. V., Colas, R. A., Perretti, M., and Serhan, C. N. (2016) Resolvin D3 Is Dysregulated in Arthritis and Reduces Arthritic Inflammation. J Immunol 197, 2362-2368
2) Becares, N., Harmala, S., China, L., Colas, R. A., Maini, A. A., Bennet, K., Skene, S. S., Shabir, Z., Dalli, J., and O'Brien, A. (2019) Immune Regulatory Mediators in Plasma from Patients with Acute Decompensation are Associated With 3-month Mortality. Clin Gastroenterol Hepatol
3) Fosshaug, L. E., Colas, R. A., Anstensrud, A. K., Gregersen, I., Nymo, S., Sagen, E. L., Michelsen, A., Vinge, L. E., Oie, E., Gullestad, L., Halvorsen, B., Hansen, T. V., Aukrust, P., Dalli, J., and Yndestad, A. (2019) Early increase of specialized pro-resolving lipid mediators in patients with ST-elevation myocardial infarction. EBioMedicine 46, 264-273
4) Shivakoti, R., Dalli, J., Kadam, D., Gaikwad, S., Barthwal, M., Colas, R. A., Mazzacuva, F., Lokhande, R., Dharmshale, S., Bharadwaj, R., Kagal, A., Pradhan, N., Deshmukh, S., Atre, S., Sahasrabudhe, T., Kakrani, A., Kulkarni, V., Raskar, S., Suryavanshi, N., Chon, S., Gupte, A., Gupta, A., Gupte, N., Arriaga, M. B., Fukutani, K. F., Andrade, B. B., Golub, J. E., and Mave, V. (2019) Lipid mediators of inflammation and Resolution in individuals with tuberculosis and tuberculosis-Diabetes. Prostaglandins and other lipid mediators 147, 106398
5) Gao, Y., Min, K., Zhang, Y., Su, J., Greenwood, M., and Gronert, K. (2015) Female-Specific Downregulation of Tissue Polymorphonuclear Neutrophils Drives Impaired Regulatory T Cell and Amplified Effector T Cell Responses in Autoimmune Dry Eye Disease. J Immunol 195, 3086-3099
6) Gao, Y., Su, J., Zhang, Y., Chan, A., Sin, J. H., Wu, D., Min, K., and Gronert, K. (2018) Dietary DHA amplifies LXA4 circuits in tissues and lymph node PMN and is protective in immune-driven dry eye disease. Mucosal immunology 11, 1674-1683
7) Chen, X. S., Sheller, J. R., Johnson, E. N., and Funk, C. D. (1994) Role of leukotrienes revealed by targeted disruption of the 5-lipoxygenase gene. Nature 372, 179-182
https://doi.org/10.7554/eLife.51102.sa2Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
A novel role for lipoxin A4 in driving a lymph node–eye axis that controls autoimmunity to the neuroretina
eLife 9:e51102.
https://doi.org/10.7554/eLife.51102




{kind=link}
{kind=link}