TMEM79/MATTRIN defines a pathway for Frizzled regulation and is required for Xenopus embryogenesis
- F. M. Kirby Neurobiology Center, Boston Children’s Hospital, Department of Neurology, Harvard Medical School, United States
- Instituto de Ciências Biomédicas, Universidade Federal do Rio de Janeiro, Brazil
Figures
Figure 1 with 1 supplement
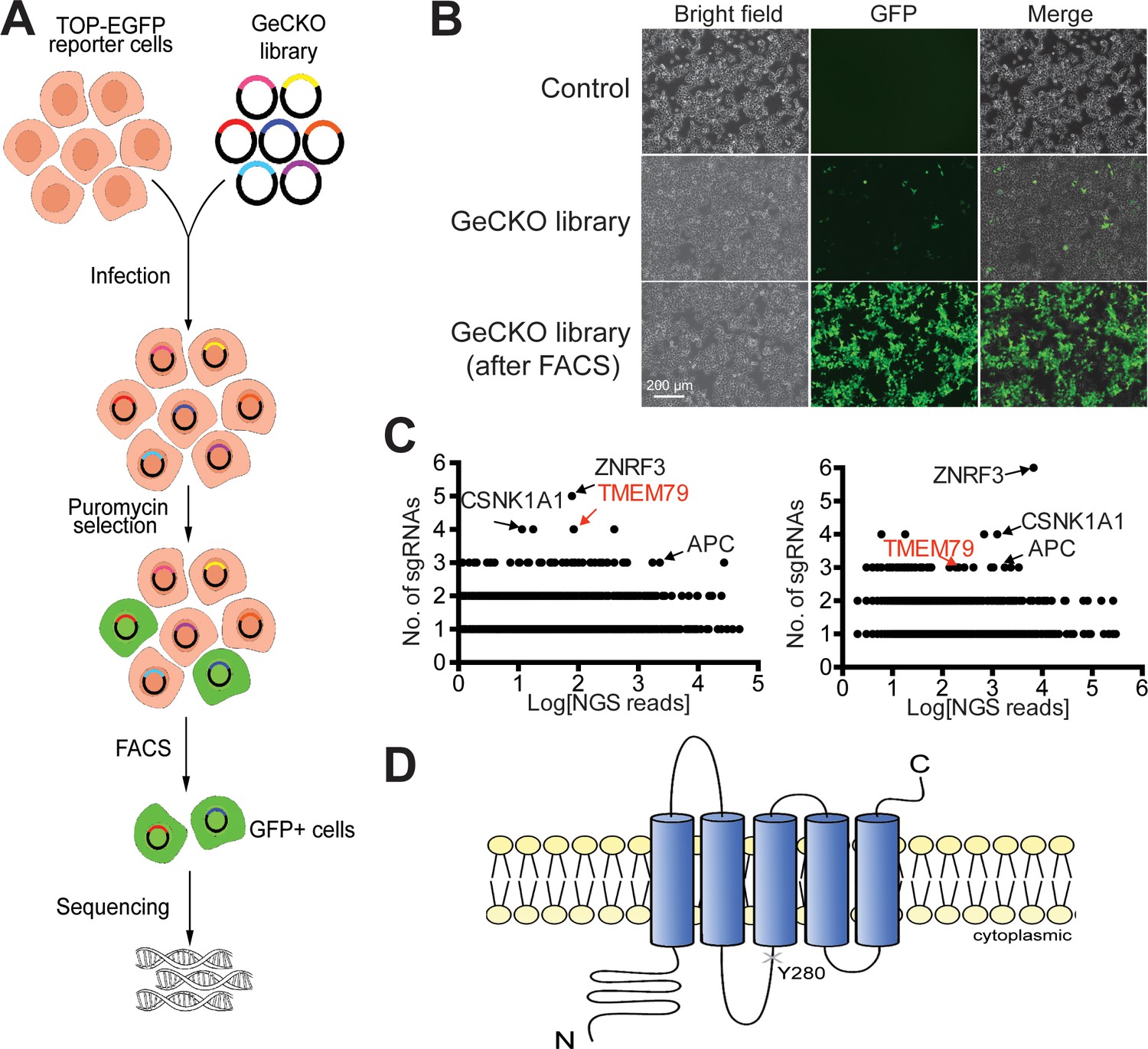
Identification of TMEM79 as an inhibitor of Wnt/β-catenin signaling through genome-wide loss-of-function screening using the CRISPR-Cas9 system.
(A) An outline of the screening strategy for negative components of Wnt/β-catenin signaling using a HEK293T-TOP-EGFP reporter line. (B) Representative images of the reporter cells with or without infection of the GeCKO library. GFP+ cells were enriched by FACS. Scale bar, 200 µm. (C) Scatter plots showing candidate genes corresponding to sgRNAs enriched. Results of two independent screens are shown. Each dot represents a candidate gene. Y-axis: number of unique sgRNAs for each gene (six sgRNAs per gene in the GeCKO library). X-axis: sgRNA NGS (next generation sequencing) reads for each gene. (D) Schematic diagram of TMEM79 protein (predicted). Y280*, a nonsense mutation at tyrosine 280 coded by the matted allele in mice.
-
Figure 1—source data 1
Raw data for Figure 1.
- https://cdn.elifesciences.org/articles/56793/elife-56793-fig1-data1-v2.xlsx
Figure 1—figure supplement 1
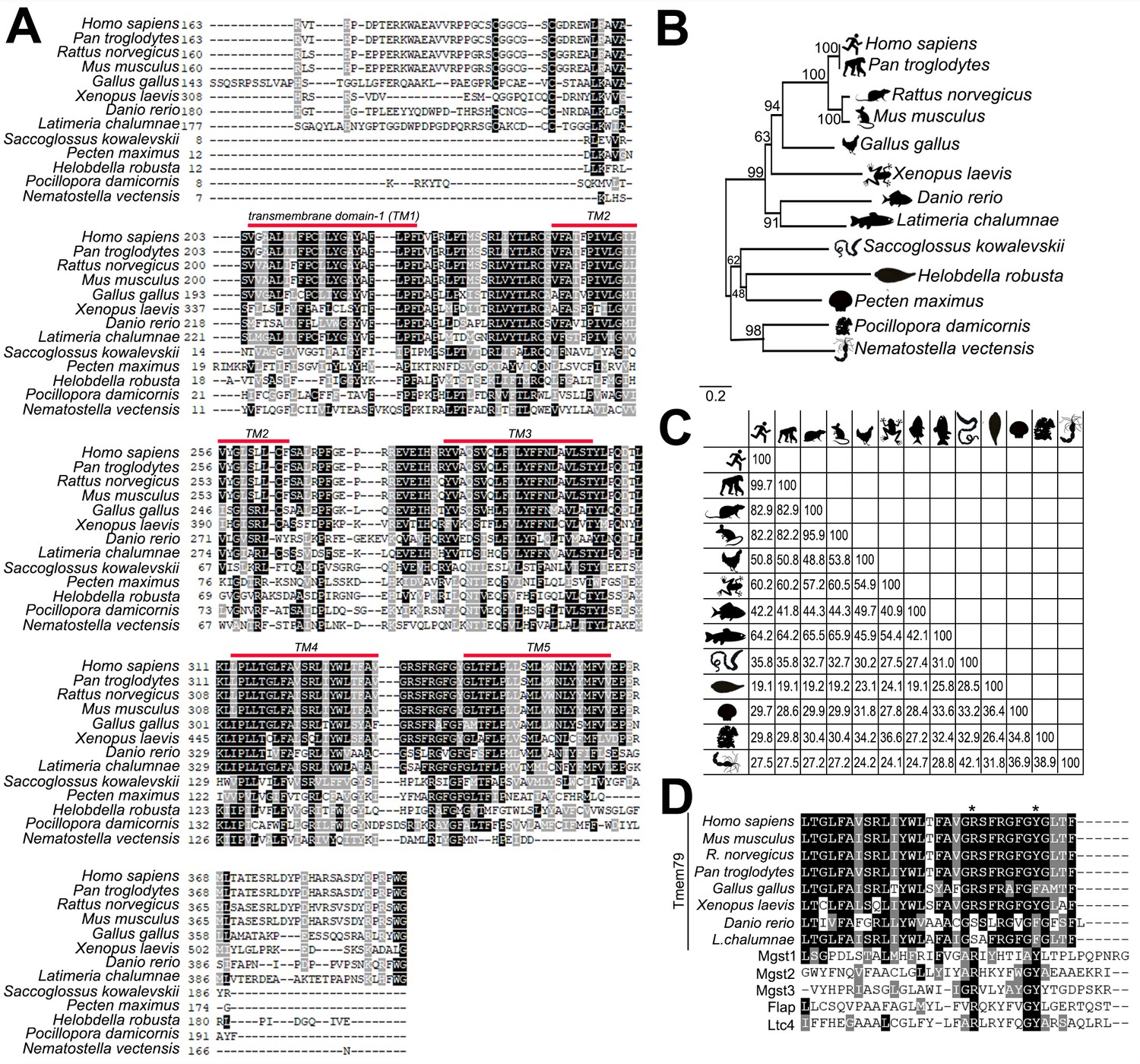
Evolutionary comparison of Tmem79 proteins in vertebrates, protostomes, and cnidarians.
(A) ClustalW alignment of TMEM79/Tmem79 proteins from human (Homo sapiens), chimpanzee (Pan troglodytes), rat (Rattus norvegicus), mouse (Mus musculus), chicken (Gallus gallus), frog (Xenopus laevis), zebra fish (Danio rerio), coelacanth (Latimeria chalumnae), hemicordate (Saccoglossus kowalevskii), great scallop (Pecten maximus), leech (Helobdella robusta), cauliflower coral (Pocillopora damicornis), and sea anemone (Nematostella vectensis). Black and grey represent identical and conservative amino acid residues, respectively. Note that the amino terminus before the transmembrane domain 1 (TM1) is poorly conserved and shown only partially. The predicted Tmem79 of Nematostella vectensis appears to lack the 5th transmembrane domain (TM5) that is present in other species listed. Accession numbers for the Tmem79 sequences in the alignment are listed in Materials and methods section. (B) The phylogenetic tree showing homology among Tmem79 proteins from species presented in A based on Neighbor-Joining phylogenetic methods. Distances in the cladogram are derived from amino acid substitutions. (C) Orthologous comparisons of identity of Tmem79 proteins among species listed in A using ClustalW alignment. (D) ClustalW alignment of vertebrate TMEM79/Tmem79 proteins from human, chimpanzee, rat, mouse, chicken, frog, zebra fish, and coelacanth with mouse MAPEG family members Mgst1, Mgst2, Mgst3, Flap and Ltc4s (Emrick et al., 2018; Saunders et al., 2013). Asterisk indicates two conserved residues within the region of limited sequence similarity corresponding to mouse Tmem79 R332 and Y339.
Figure 2 with 1 supplement
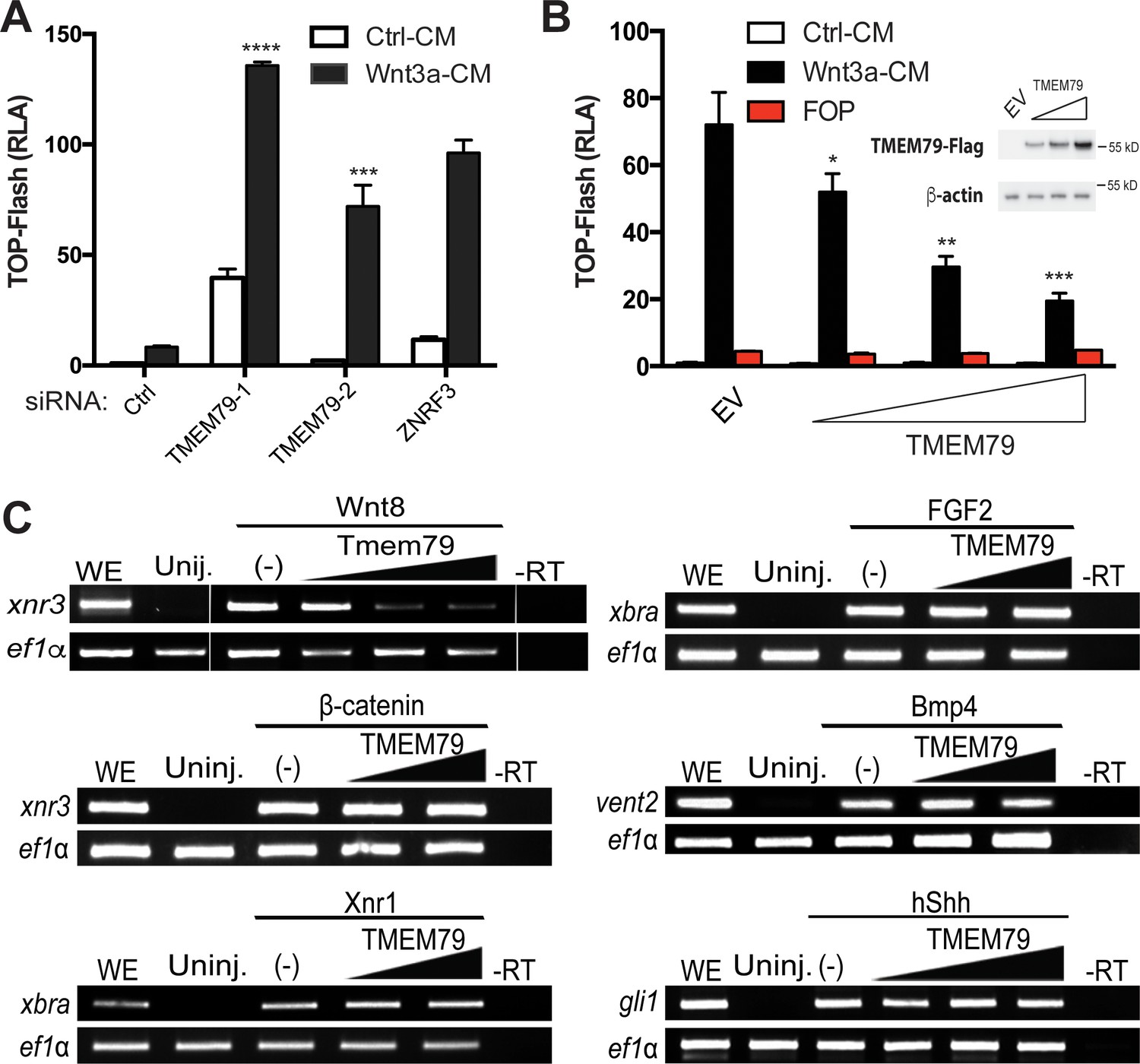
TMEM79 is a specific inhibitor of Wnt/β-catenin signaling.
(A) siRNA depletion of TMEM79 enhances top-flash reading with or without Wnt3a. A ZNRF3 siRNA serves as a positive control. CM, conditioned medium. TOP-Flash is quantified using RLA (relative luciferase activity), with the control set as 1. (B) TMEM79 dose-dependently suppresses top-flash. FOP, a negative control luciferase reporter. EV, empty vector. Inset: immunoblot of TMEM79 proteins, with β-actin as loading control. (C) In animal pole explants of Xenopus embryos, TMEM79 inhibits expression of xnr3 induced by Wnt8 but not by β-catenin. TMEM79 does not inhibit expression of target genes (Xbra, vent2, and Gli1) induced by Nodal, FGF, BMP, or Shh, respectively. Ef1α: RT-PCR/loading control; WE, whole embryo; Uninj, uninjected embryo; -RT: no reverse transcriptase. In this and all other figures, gradient bars symbolize different doses of DNA in transfection or mRNA in embryo injection; error bars represent SEM from at least three independent experiments; *p<0.05, **p<0.01, and ***p<0.001, ****p<0.0001 are based on student’s t-tests.
-
Figure 2—source data 1
Raw data for Figure 2.
- https://cdn.elifesciences.org/articles/56793/elife-56793-fig2-data1-v2.xlsx
Figure 2—figure supplement 1
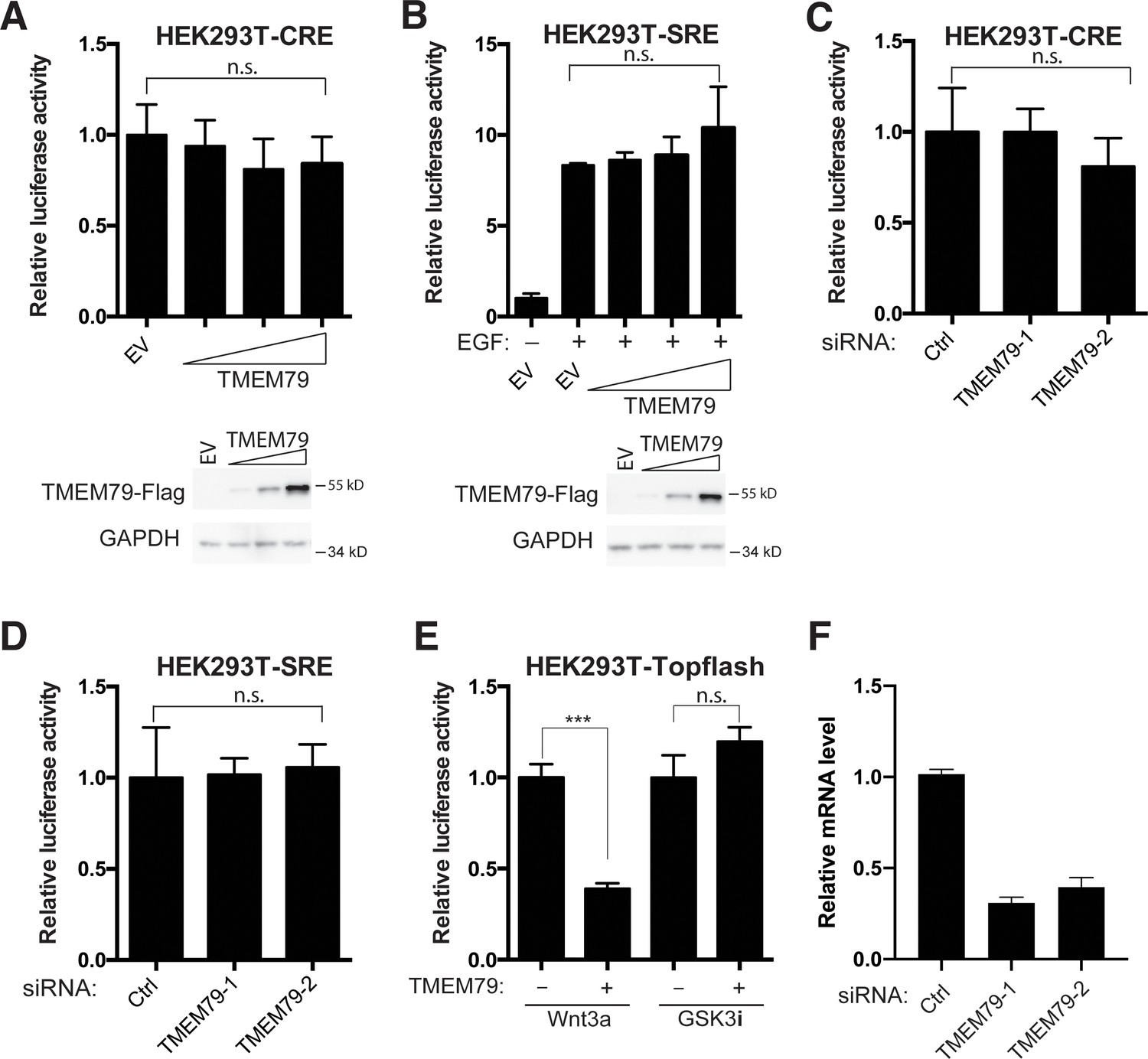
TMEM79 is a specific inhibitor for Wnt/β-catenin signaling.
(A and B) TMEM79 affects neither the cAMP/PKA pathway, assayed by a CRE (cAMP responsive element)-driven reporter, or the EGF/EGFR pathway, assayed by an SRE (serum response element)-(driven reporter). (C and D) Depletion of TMEM79 by siRNAs affects neither the CRE- or SRE-driven reporter. (E) TMEM79 inhibits the TOP-Flash reporter induced by Wnt3a but not that induced by a GSK3 inhibitor (GSK3i). (F) Relative mRNA levels of TMEM79 in HEK293 cells transfected with indicated siRNA.
-
Figure 2—figure supplement 1—source data 1
Raw data for Figure 2—figure supplement 1.
- https://cdn.elifesciences.org/articles/56793/elife-56793-fig2-figsupp1-data1-v2.xlsx
Figure 3 with 1 supplement

TMEM79 down-regulates the Frizzled (FZD) protein level at the cell surface independent of ZNRF3/RNF43.
(A) Porcupine inhibitor IWP-2 abolishes Top-Flash activities induced by a TMEM79 siRNA. (B) Levels of phospho-LRP6 and phospho-DVL2 are elevated in T79KO and ZRKO cells, and are reduced/reversed by IWP-2, as detected by a phospho-LRP6 (S1490) antibody and the upper DVL2 protein band, respectively. (C) Endogenous FZD5 levels are elevated in T79KO and ZRKO cells comparably. FZD null cells (knockout of FZD1-10) (Eubelen et al., 2018) serve as control for immunoblotting specificity. (D) TMEM79 and ZNRF3 inhibit Wnt signaling independent of each other. (E) siRNA depletion of ZNRF3 or TMEM79 further elevates TOP-Flash activities in T79KO or ZRKO cells. (F) Co-IP of TMEM79 (Flag-tagged) with endogenous FZD5 but not LRP6. (G) TMEM79 is co-IPed with each FZD (FZD1 to 8) but not SMO, and with FZD9 and FZD10 but not SMO (Figure 3—figure supplement 1F). (H) TMEM79 reduces the surface level of FZD5 as indicated by cell surface protein biotinylation assay. Surface proteins were labeled by membrane-impermeable biotin and isolated by streptavidin agarose (strep). Either TMEM79 or ZNRF3 reduces the upper band of FZD5 (the matured and biotinylated form) in input and streptavidin-precipitates, suggesting that TMEM79 or ZNRF3 diminishes surface level of FZD5. Note surface-biotinylation of ZNRF3 but not TMEM79, indicating that TMEM79 is not at the plasma membrane. (I) TMEM79 co-IPs the immature form of FZD5. (J) FZD5-K0 is resistant to down-regulation by TMEM79 or ZNRF3.
-
Figure 3—source data 1
Raw data for Figure 3.
- https://cdn.elifesciences.org/articles/56793/elife-56793-fig3-data1-v2.xlsx
Figure 3—figure supplement 1
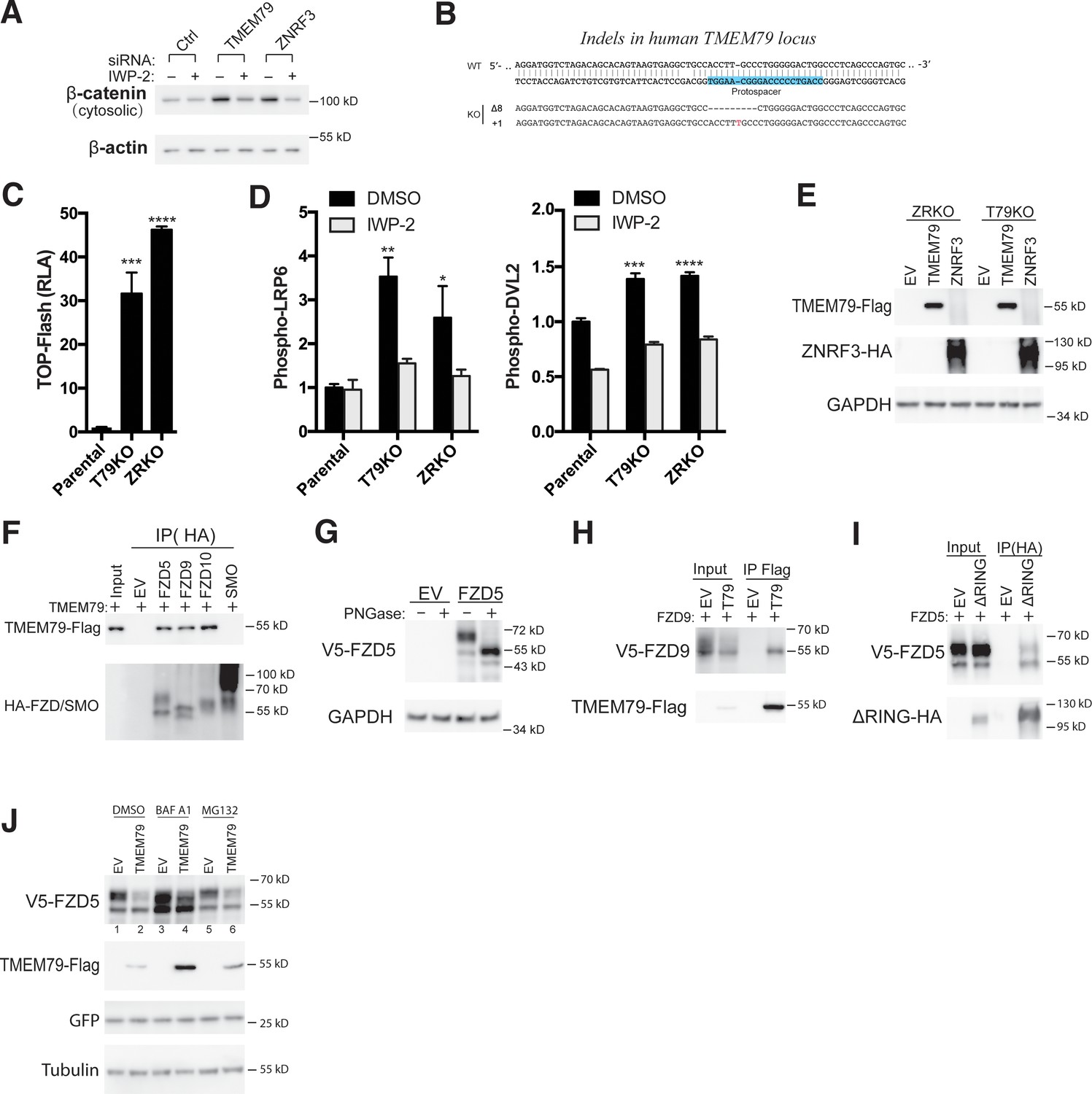
TMEM79 inhibits Wnt/β-catenin signaling by reducing surface level of frizzled receptor.
(A) Depletion of TMEM79 or ZNRF3 via an siRNA in HEK293T cells induces cytosolic β-catenin accumulation, which is prevented by IWP-2. (B) Genomic DNA sequencing showing indel (frame shift) mutations of TMEM79 alleles in the T79KO clone picked/used. Two clones were picked and tested with indistinguishable results (data not shown). (C) Top-flash reporter is activated comparably and strongly in T79KO cells and ZRKO cells. (D) Figure 3B quantification. Phospho-LPR6 (pS1490) and phospho-DVL2 (the upper band) levels are measured and normalized to total LRP6 and DVL2 levels, respectively. (E) Overexpression of TMEM79 and ZNRF3 proteins in ZRKO and T79KO cells, respectively. (F) TMEM79 is co-IPed with FZD5 (positive control), FZD9, and FZD10, but not SMO (negative control). (G) Mature FZD5 (the upper band) is de-glycosylated by PNGase and converted to the lower band as un-glycosylated immature FZD5. (H) TMEM79 co-IPs preferentially the lower band of FZD9, likely the immature form, and reduces the level of upper bands of FZD9, which appear to be heterogenous, possibly due to heterogeneity of glycosylation. This is consistent with FZD9 maturation being inhibited by TMEM79. Other FZDs when overexpressed do not exhibit distinct bands of separable electrophoretic mobilities (data not shown) as do FZD5 most overtly and FZD9 to a lesser degree. (I) ZNRF3ΔRING, a mutant with the RING domain deletion, co-IPs mature FZD5 and immature FZD5 (possibly during ZNRF3ΔRING’s transit through the ER to PM). Because the full-length ZNRF3 reduces FZD levels, we employed ZNRF3ΔRING, which does not have the E3 ligase function, to avoid fluctuations of the FZD5 level in the co-IP assay. (J) Lysosome inhibitor bafilomycin A1 (BAF A1), but not proteasome inhibitor MG132, inhibits TMEM79-induced degradation of immature FZD5. BAF A1 increases both mature and immature forms of FZD5 (compare lanes 1 and 3). BAF A1 blocks TMEM79-induced degradation of immature FZD5, but does not increase mature FZD5 (compare lanes 3 and 4), consistent with the notion that TMEM79 prevents FZD maturation to the PM by shuttling FZD5 to the lysosome. MG132 does not affect levels of immature and mature FZD5 in the absence (compare lanes 1 and 5) or presence of TMEM79 (compare lanes 2 and 6). Thus it appears that ZNRF3 and RNF43 promote degradation of mature FZD5 through the lysosome (Koo et al., 2012), whereas TMEM79 promotes degradation of immature FZD5 through the lysosome.
-
Figure 3—figure supplement 1—source data 1
Raw data for Figure 3—figure supplement 1.
- https://cdn.elifesciences.org/articles/56793/elife-56793-fig3-figsupp1-data1-v2.xlsx
Figure 4 with 1 supplement
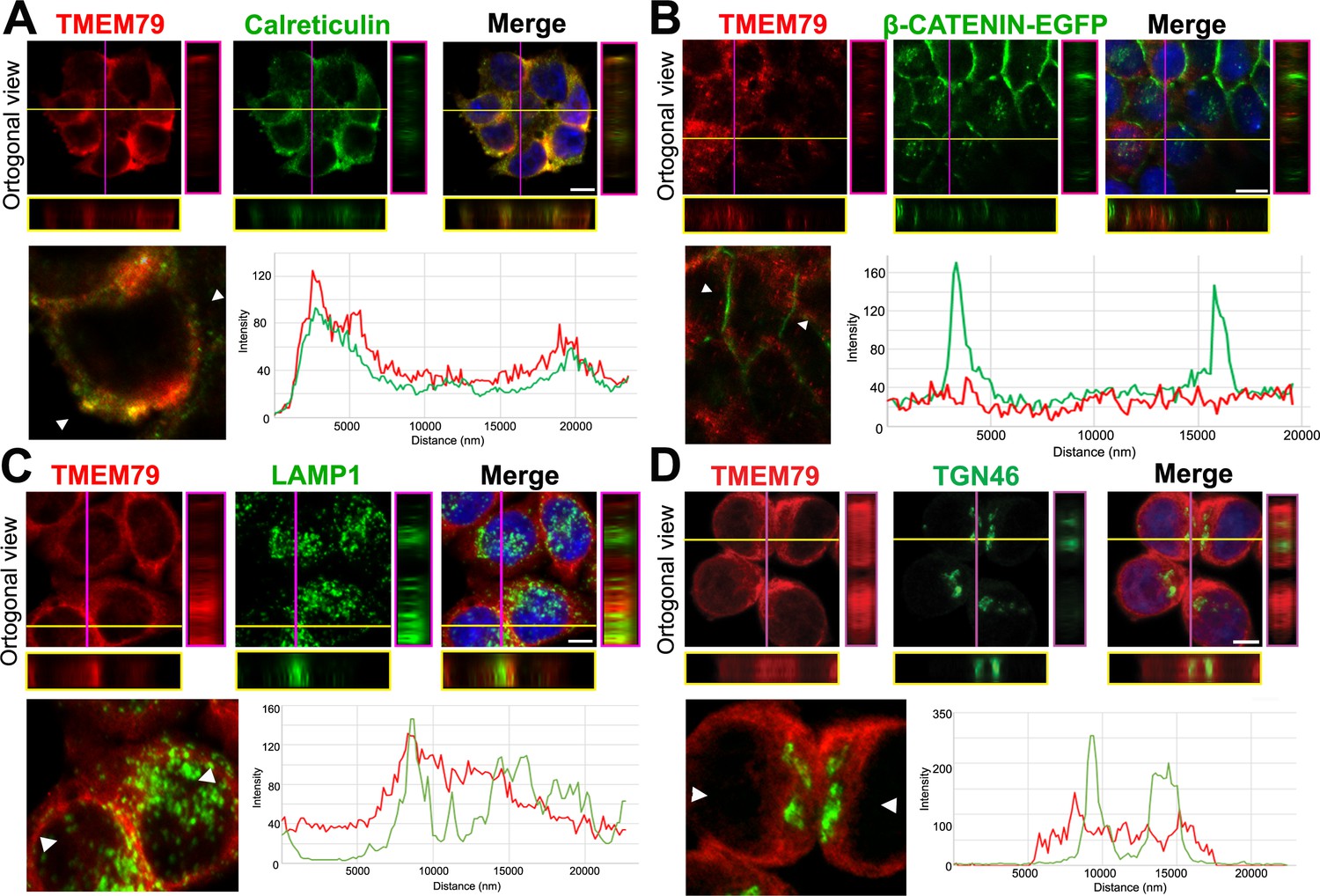
TMEM79 is likely an ER-resident protein.
(A to D) Immunofluorescence microscopy of endogenous TMEM79 (red) with ER, PM, lysosome, and Golgi markers (green), respectively, with the nucleus labeled by DAPI (blue). The z-axis is shown on right and underneath. The white scale bar represents 10 µm. Fluorescence intensity of TMEM79 and the respective marker is measured between the two arrowheads in the lower left panel and plotted at lower right. (A) TMEM79 is localized at the ER. TMEM79 and calreticulin (an ER marker) overlap extensively in images and intensity traces. (B) TMEM79 is not present at the PM, which is visualized by β-catenin-EGFP (produced from the endogenous β-catenin gene with an in-frame knock-in of the EGFP gene, see Materials and methods). Note that β-catenin is predominantly localized at the PM in epithelial cells. (C) Some TMEM79 is observed at the lysosome. TMEM79 and LAMP1 (lysosomal-associated membrane protein 1) overlap occasionally. (D) TMEM79 is hardly observed at the Trans Golgi and exhibits little or minor overlap with TGN46 (trans-Golgi marker 46).
Figure 4—figure supplement 1
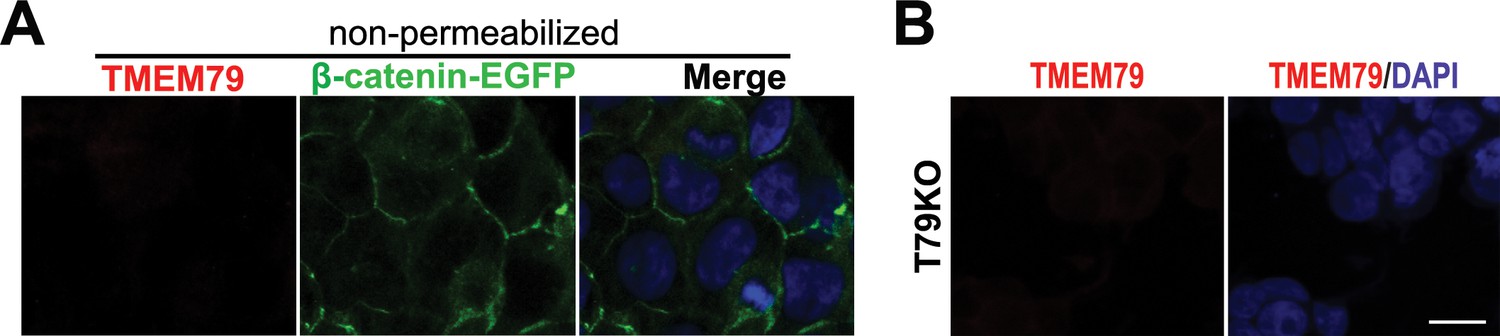
TMEM79 is an ER-resident protein.
(A) TMEM79 is not detectable by the TMEM79 antibody without cell permeabilization. Note that the antibody (Key Resources Table) recognizes the TMEM79 C-terminus (Figure 1D), which is either in the ER lumen, or extracellular if TMEM79 were to be on the PM. This result is consistent with TMEM79 being an ER- but not PM-resident protein. (B) T79KO cells lack TMEM79 staining completely, demonstrating specificity of the TMEM79 antibody for immunostaining. Scale bar = 10 μm.
Figure 5 with 1 supplement
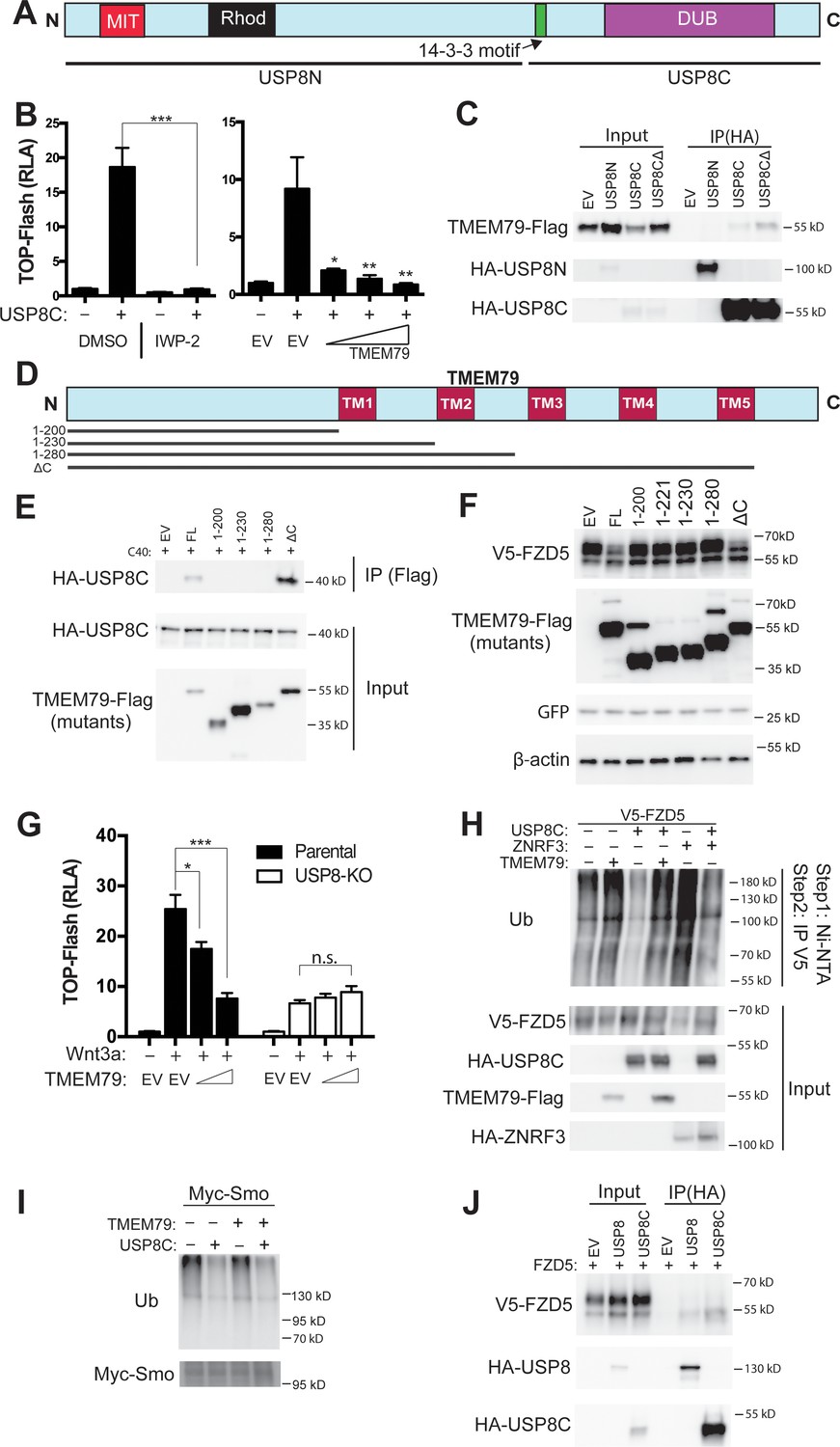
TMEM79 inhibits Frizzled (FZD) deubiquitination by USP8.
(A) Schematic diagram of USP8. USP8N, USP8 N terminal fragment; USP8C, USP8 C terminal fragment. Known domains including DUB and a 14-3-3-binding motif are depicted. (B) Activation of Top-Flash by USP8C is inhibited by IWP-2 or by TMEM79. (C) TMEM79 is co-IPed by USP8C or USP8CΔ (with the 14-3-3 binding motif deleted), but not by USP8N, suggesting that USP8C (but not its 14-3-3 binding motif) is involved in TMEM79 interaction. (D) Schematic diagram of TMEM79 and its deletion mutants. Five transmembrane (TM) regions are depicted. (E) TMEM79 (FL, full length) or TMEM79-ΔC, but none of the other TMEM79 mutants, co-IPs USP8C. (F) TMEM79 or TMEM79-ΔC, but none of the other TMEM79 mutants, down-regulates the level of mature FZD5. (G) The Wnt response in USP8-KO cells is reduced and is not inhibited by TMEM79. (H and I) TMEM79 inhibits USP8C deubiquitination of FZD5 (H), but not SMO (I). HEK293T cells were cotransfected with V5-FZD5 and His-ubiquitin, and in combination with USP8C, TMEM79 or ZNRF3 as indicated. Ubiquitinated proteins were precipitated by Ni-NTA (nickel-nitrilotriacetic acid–agarose beads for His tag pull-down), followed by wash in 7M urea to fully disassociate all protein complexes. Ubiquitinated V5-FZD5 was then precipitated by anti-V5 agarose beads. These multiple steps ensured detection of ubiquitinated V5-FZD5 only, but not V5-FZD5-associated proteins that are also ubiquitinated. Levels of ubiquitinated FZD5 were increased by TMEM79 or ZNRF3 co-expression whereas USP8C deubiquitinated FZD5. TMEM79 inhibited USP8C deubiquitination of FZD5 and restored FZD5 ubiquitination (H). USP8C also deubiquitinated SMO, but TMEM79 had no effect on USP8C deubiquitination of SMO (I). (J) USP8 or USP8C preferentially co-IPs the immature form of FZD5 (the lower band).
-
Figure 5—source data 1
Raw data for Figure 5.
- https://cdn.elifesciences.org/articles/56793/elife-56793-fig5-data1-v2.xlsx
Figure 5—figure supplement 1
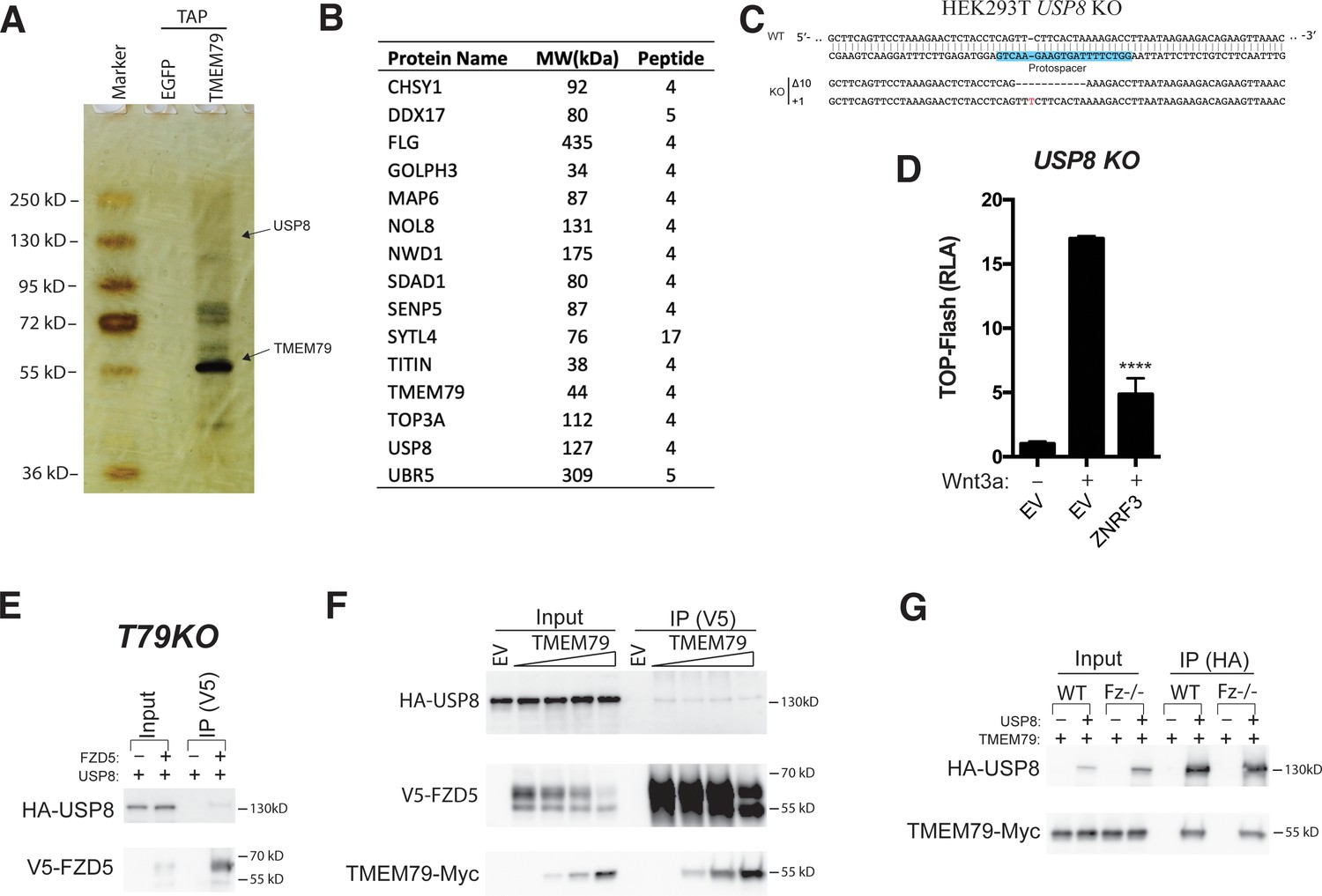
Tandem affinity purification-coupled mass spectrometry identifies USP8 as a TMEM79 binding partner.
(A) Tandem affinity purified proteins were resolved by SDS-PAGE and visualized by silver staining. The same tandem IP procedure of EGFP-expressing cell extracts serves as a negative control. The TMEM79 bait protein is marked, as is the gel fraction that corresponds to the molecular weight of USP8. (B) List of candidate proteins from tandem affinity purification-coupled mass spectrometry. Note that only a few visible fractions/bands in A were excised and pooled together for mass spectrometry analyses (see Materials and methods). The list is therefore not intended to be a comprehensive representation of all candidate TMEM79-associated proteins. (C) Genomic DNA sequencing showing indel (frame shift) mutations of USP8 alleles in a USP8 KO clone picked/used. Two clones were picked and tested with indistinguishable results (data not shown). (D) ZNRF3 inhibits top-flash reporter activity effectively in USP8 KO cells. (E) FZD5 co-IPs USP8 in the absence of TMEM79 in T79KO cells. (F) Overexpression of TMEM79 does not affect FZD5 co-IP of USP8. This experiment largely rules out a scenario that TMEM79 inhibits USP8 deubiquitination of FZD5 by competing with FZD5 for USP8. TMEM79 is also co-IPed by FZD5. (G) USP8 co-IPs TMEM79 in in FZD null cells.
-
Figure 5—figure supplement 1—source data 1
Raw data for Figure 5—figure supplement 1.
- https://cdn.elifesciences.org/articles/56793/elife-56793-fig5-figsupp1-data1-v2.xlsx
Figure 6 with 1 supplement
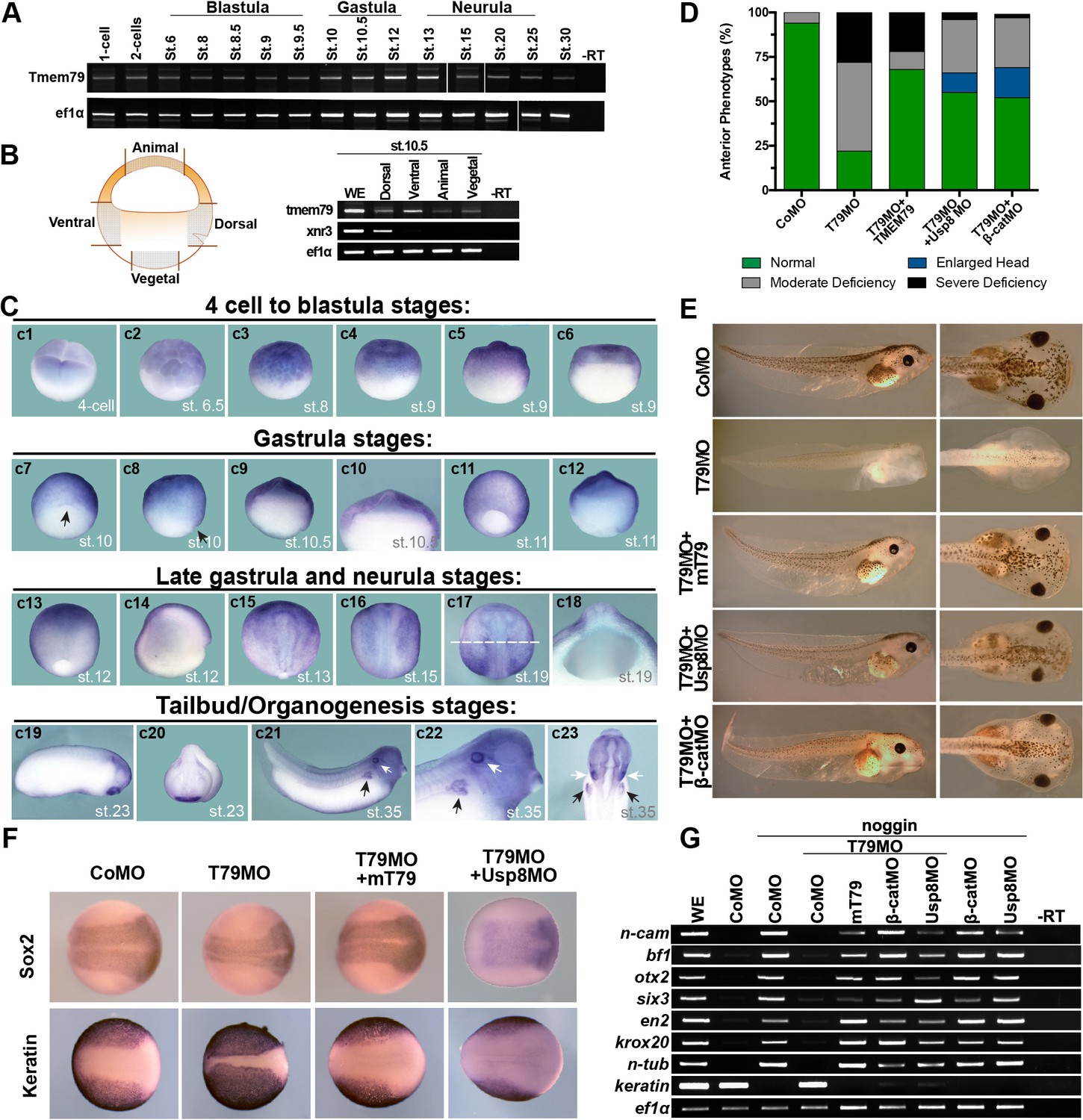
Xenopus Tmem79 is required in naive ectoderm for anterior and neural development.
(A) Tmem79 is expressed throughout Xenopus embryogenesis from 1 cell (fertilized egg) to stage (st) 30, spanning blastula, gastrula, neurula and tailbud/organogenesis stages, as assayed by RT-PCR. (B) Tmem79 is expressed in dissected animal, vegetal, dorsal, and ventral regions (as schemed in drawing) of early gastrula embryos (stage 10.5). xnr3 is a control for dorsal. (C) Tmem79 expression during Xenopus embryogenesis by whole-mount in situ hybridization. An apparent lack of hybridization signals in vegetal pole and derived tissues is likely technical due to yolk. Top row (c1 to c6) Tmem79 is expressed in the animal pole at the four-cell stage and in naive ectoderm at blastula stages: (c1) Dorsal-animal view; (c2 to c4) Lateral-animal views; (c5 and c6) Lateral and bisected views. Second row (c7 to c12) Tmem79 is expressed broadly in naive ectoderm at early gastrula stages: (c7 and c8) dorsal-vegetal and lateral views with the arrow pointing to the dorsal; (c9) bisected view with an enlarged view (c10) showing expression in naive ectoderm; (c11 and c12) lateral and bisected views. Anterior to top in c11. Third row (c13 to c18) Tmem79 shows dynamic expression from late gastrula to neurula stages: (c13 and c14) lateral and bisected views showing broad expression including in neural plate. Anterior to top in c13 and left in c14. (c15 to c17) Dorsal views. Anterior to top. Tmem79 expression is downregulated in neural plate while remains strong in epidermis, as seen also in a cross-section (c18) along the dashed line in c17. (c18) an enlarged cross-section view with dorsal on top. Bottom row (c19 to c23) Tmem79 is expressed during organogenesis: (c19 and c20) Lateral and anterior views showing expression in the cement gland. (c21 and c22) A lateral view with an enlarged view (anterior to right) and (c23) a ventral view (anterior to top) showing expression in primitive kidney (black arrows) and otic vesicle (white arrows). (D and E) Dorsal-animal (naive ectoderm) injection of Tmem79MO causes anterior deficiency, which is rescued/over-rescued by co-injection of mouse Tmem79 mRNA or Usp8MO or β-cateninMO. CoMO, Control Morpholino. Anterior to right (E). Also see Supplementary file 1A. (F) Tmem79MO reduces neural plate formation and reciprocally expands epidermal differentiation, reflected by Sox2 and cytokeratin expression domains at Stage 16, respectively. These changes are rescued by Tmem79 mRNA or Usp8MO. Anterior to right. Also see Supplementary file 1C. (G) Neural induction by Noggin in animal pole explants is prevented by Tmem79MO, whose effect is rescued by Tmem79 mRNA or Usp8MO or β-cateninMO. Pan-neural (n-cam), anterior (bf1, otx2, and six3) and mid/hind (en2 and krox20) brain markers and a neuronal differentiation marker (n-tub) are assayed, as is an epidermal marker (keratin).
Figure 6—figure supplement 1
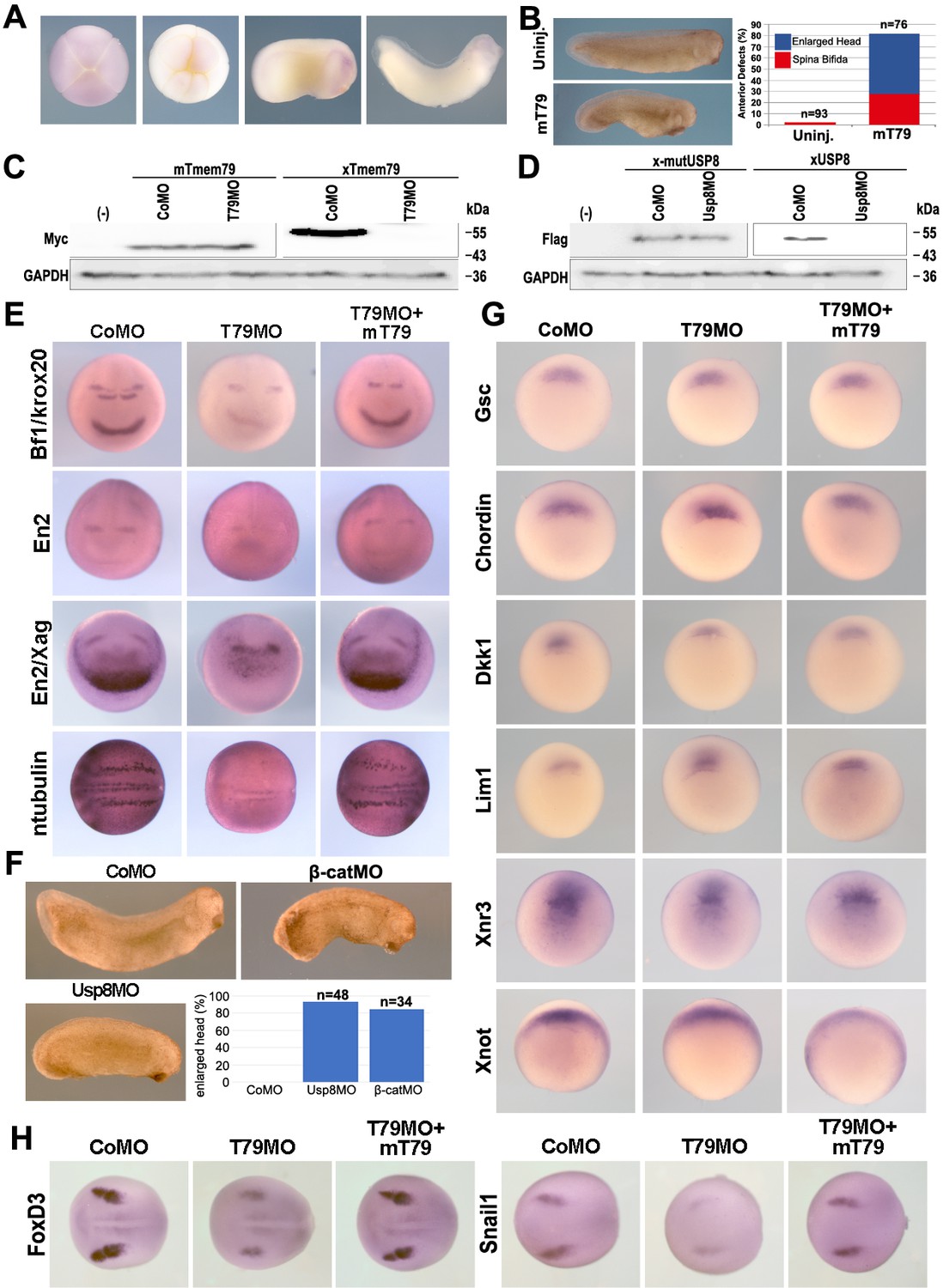
Xenopus tmem79 is required for anterior neural patterning and neural crest formation without affecting the head organizer.
(A) A Tmem79 sense probe as a negative control does not show any hybridization signal in embryos at different stages. (B) Dorsal injection of Tmem79 mRNA induces an enlarged head in >50% embryos and spina bifida in another about 30% embryos. Anterior to right. Also see Supplementary file 1G. (C) Tmem79MO inhibits protein synthesis from the Xenopus Tmem79 (xTmem79) mRNA, but not of the mouse Tmem79 (mT79) mRNA, which is used in rescue experiments. GAPDH is a protein loading control. (D) Usp8MO inhibits protein synthesis from the Xenopus Usp8 (xUsp8) mRNA, but not from a mutant Usp8 (x-mutUsp8) mRNA, in which several nucleotides in xUsp8 5’-UTR targeted by the MO have been altered. (E) Dorsal-animal injection of Tmem79 MO reduces expression of anterior and mid/hind brain markers Bf1, Krox20, and En2, an anterior (cement gland) marker Xag (each seen in the anterior view), and a neuronal marker n-tubulin (see in the dorsal view), examined at stage 16. Co-injection of mTmem79 with Tmem79MO rescues expression of these markers. Also see Supplementary file 1H. (F) Dorsal-animal injection of β-cateninMO or Usp8MO leads to enlarged head formation at the tailbud stage (anterior to right), supporting Usp8 acting positively in Wnt/β-catenin signaling in vivo. Also see Supplementary file 1I. (G) Dorsal-animal injection of Tmem79MO affects neither expression of head organizer genes (Goosecoid, Chordin, Dkk1, and Lim1) nor that of dorsal genes (xnr3 and xnot) at stages 10.5–11. Vegetal views with dorsal to top. Also see Supplementary file 1J. (H) Dorsal-animal injection of Tmem79MO reduces expression of neural crest markers FoxD3 and snail1 examined at stage 16. Also see Supplementary file 1K.
Figure 7
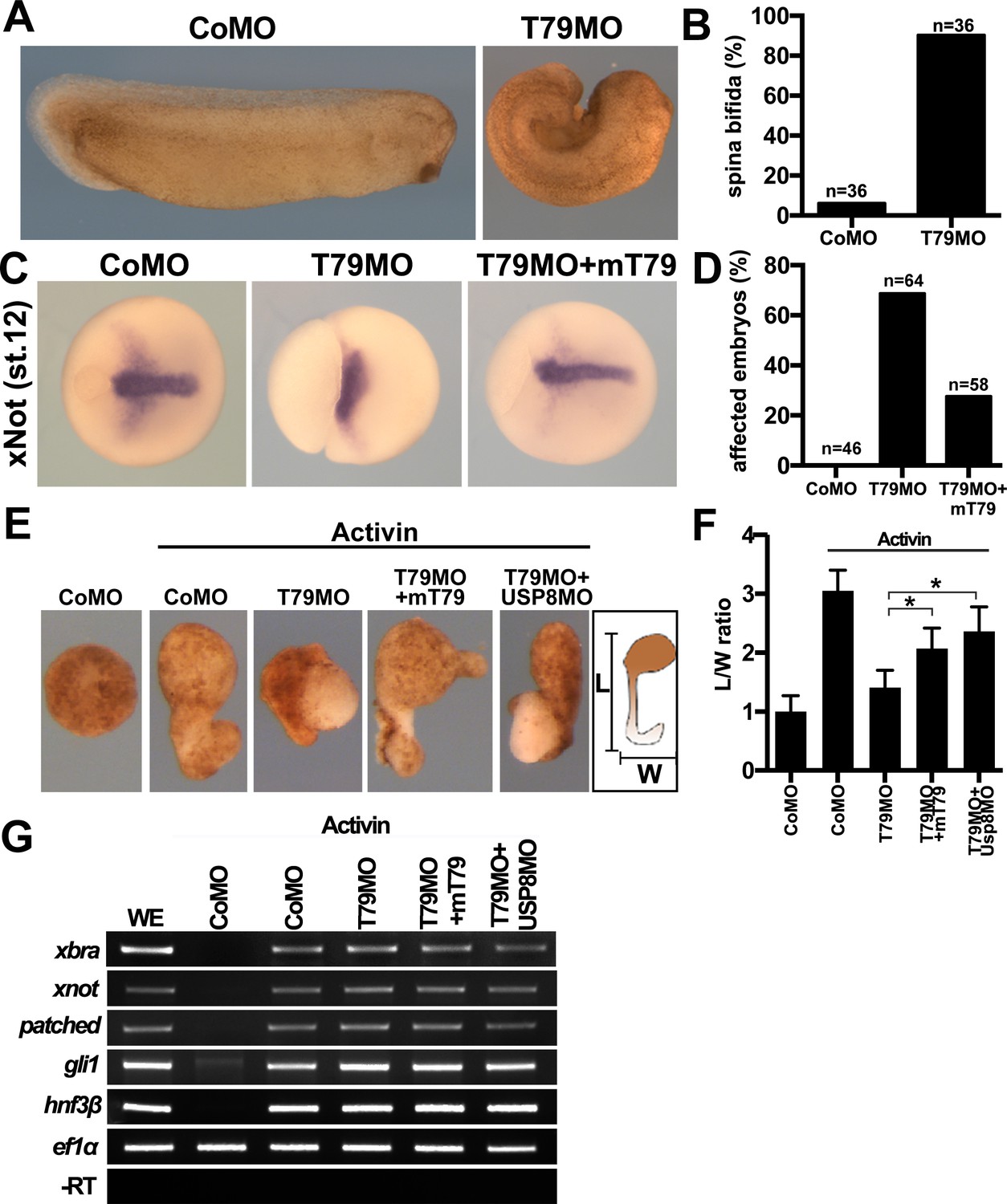
Xenopus Tmem79 regulates Frizzled (FZD)/PCP signaling and convergent-extension (CE) cell movements in axial mesoderm.
(A to D) Dorsal (axial mesoderm) injection of Tmem79MO causes spina bifida scored at stage 26 (A and B), and a lack of CE movements of axial mesoderm, visualized by a widened (not elongated) xNot-expressing axial tissue and an open blastopore at stage 12 (C and D). Axial mesoderm elongation is rescued by Tmem79 mRNA (C and D). Anterior to right. Also see Supplementary file 1D and E. (E to G) Axial mesoderm induced by activin in animal pole explants exhibits CE movements, visualized by explant elongation and quantified by a length/width (L/W) ratio (E and F). Control explants are round with a L/W ratio of 1 while explants (axial mesoderm induced by activin) elongate with a L/W ratio of 3 (E and F). Explants with Tmem79MO poorly elongate but elongation is rescued by Tmem79 mRNA or Usp8MO (E and F). Also see Supplementary file 1F. Note that Tmem79MO alone or together with Usp8MO does not affect activin-induced axial mesoderm gene expression (G).
Figure 8
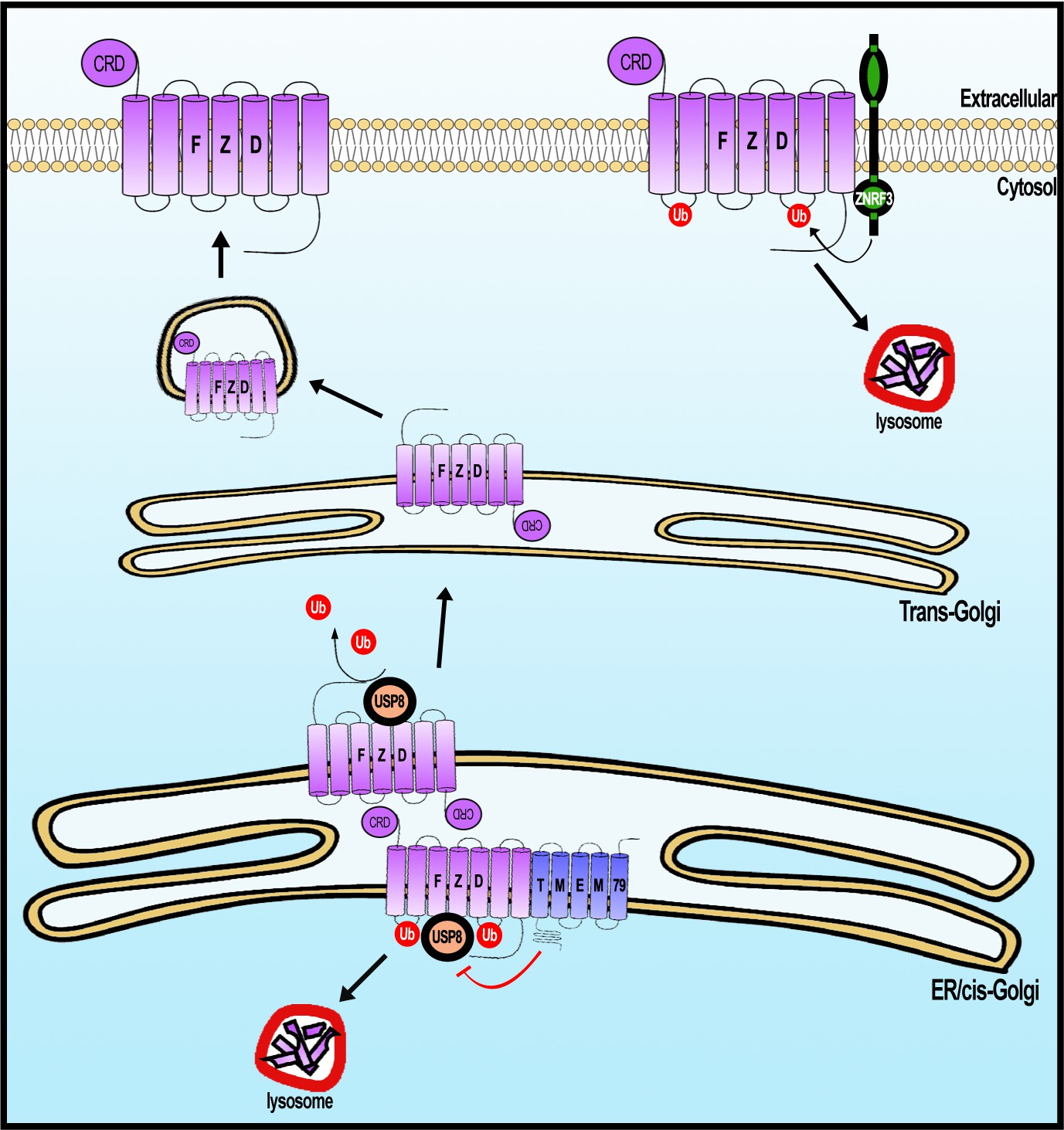
A working model of Frizzled (FZD) regulation by USP8 and TMEM79.
FZD is synthesized in the ER and is ubiquininated (by an unknown E3 ligase, not depicted). FZD deubiquitination by USP8 permits FZD to traffic to Golgi and PM ultimately. ER-resident TMEM79 acts by complexing with FZD and USP8 and inhibiting USP8 deubiquitination of FZD, thereby ensuring ubiquitinated FZD in the ER to traffic to the lysosome for degradation. For FZD that escapes TMEM79-mediated degradation and matures to the PM, ZNRF3 promotes FZD ubiquitination at the PM, leading to endosomal-lysosomal degradation of FZD. These two complementary mechanisms ensure a rate-limiting amount of FZD at the PM.
Figure 9
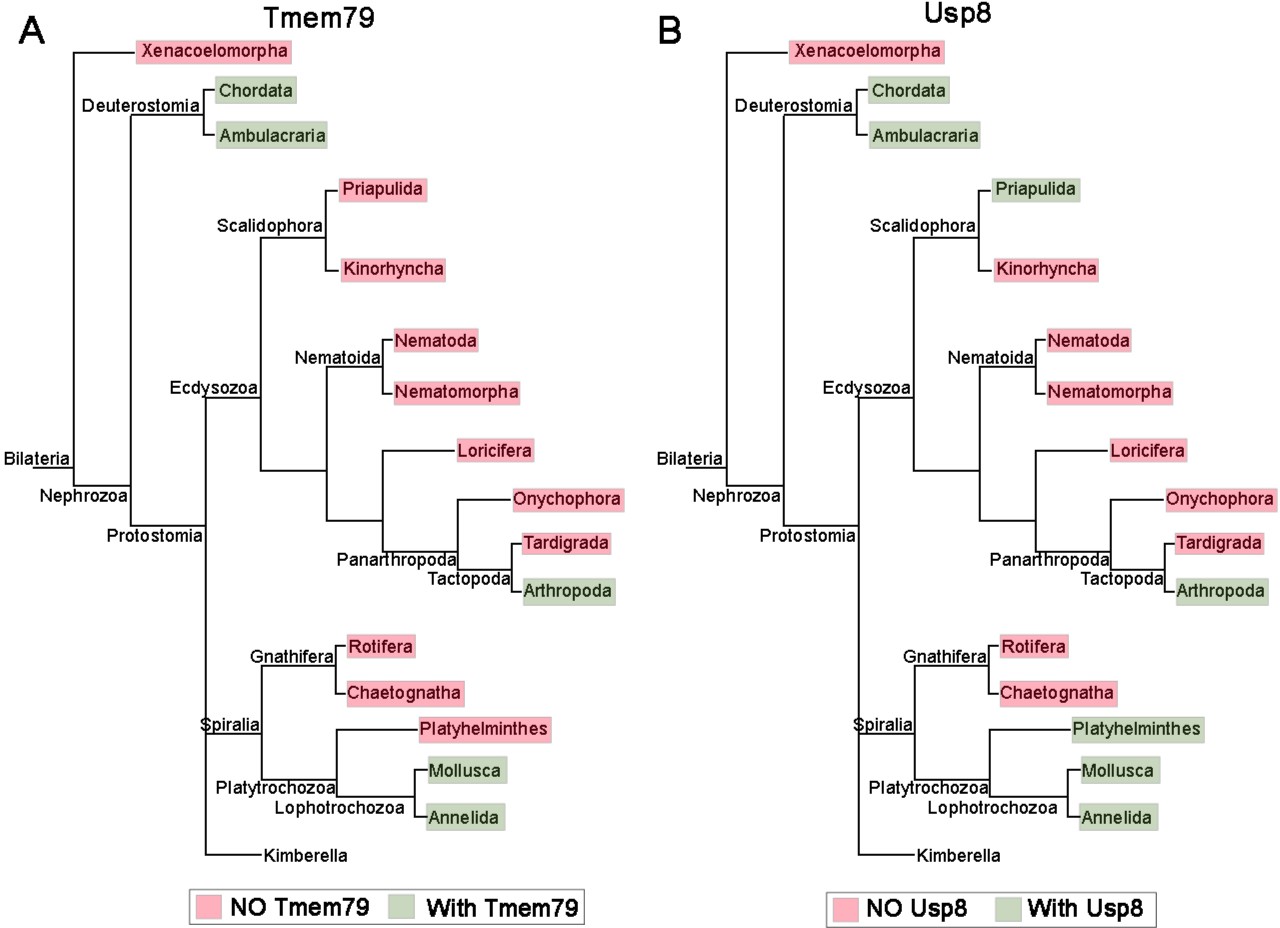
The phylogenetic preservation or loss in bilateria of Tmem79 (A) and Usp8 (B).
Tmem79 and Usp8 are present throughout deuterostomes while exhibit frequent and similar losses in protostomes. Note that both genes are found in cnidaria and thus was likely present in the common cnidarian and bilaterian ancestor (Figure 1—figure supplement 1). Preservation of Tmem79 in protostomes is always accompanied by preservation of Usp8, but not vice versa. One such example is in arthropods: Tmem79 is absent but Usp8 is present in Drosophila while the Acari has Tmem79 and Usp8. Neither gene is found and was probably lost in Xenacoelomorpha.
Tables
Appendix 1—key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene (M. musculus) | Tmem79 | Origene | MR206106 | |
| Strain, strain background (Escherichia coli) | E. coli NEB 5-alpha | New England Biolabs (NEB) | Cat#C2987H | |
| Strain, strain background (Xenopus laevis) | Xenopus laevis | Nasco | Cat# LM00715 | |
| Strain, strain background (Xenopus laevis) | Xenopus laevis | Nasco | Cat# LM00535 | |
| Cell line (Homo-sapiens) | Human: HEK293T | ATCC | Cat# CRL-11268 RRID:CVCL_1926 | |
| Cell line (Homo-sapiens) | Human: HEK293T-BAR/Renilla | He lab | See ‘dual luciferase assay’ | |
| Cell line (Homo-sapiens) | Human: HEK293T-ZRKO | Feng Cong PMID:25891077 | ||
| Cell line (Homo-sapiens) | Human: FZD null HEK293T | Benoit Vanhollebeke PMID:30026314 | ||
| Cell line (Homo-sapiens) | Human: HEK293T-β-catenin-EGFP knockin | This paper | He Lab; See‘Generation of HEK293T knockout (KO) and kncokin (KI) cell lines’ | |
| Transfected construct (human) | siRNA to TMEM79 | Sigma-Aldrich | See ‘RNA interference’ | |
| Antibody | β-catenin (Mouse monoclonal) | BD Transduction Laboratories | Cat# 610154 RRID:AB_397555 | WB (1:2000) |
| Antibody | β-tubulin (Mouse monoclonal) | Hybridoma Bank | E7 | WB (1:5000) |
| Antibody | GAPDH (Rabbit monoclonal) | Cell Signaling Technology (CST) | Cat# 2118 | WB (1:5000) |
| Antibody | HA tag (Rabbit monoclonal) | CST | Cat# 3724 | WB (1:1000) |
| Antibody | Flag tag (Rabbit monoclonal) | CST | Cat# 14793 | WB (1:1000) |
| Antibody | V5 tag (Rabbit monoclonal) | CST | Cat# 13202 | WB (1:1000) |
| Antibody | LRP6 (Rabbit monoclonal) | CST | Cat# 3395 | WB (1:1000) |
| Antibody | phospho-LRP6 (Ser1490) (Rabbit Polyclonal) | CST | Cat# 2568 | WB (1:1000) |
| Antibody | DVL2 (Rabbit monoclonal) | CST | Cat# 3224 | WB (1:1000) |
| Antibody | USP8 (Rabbit Polyclonal) | Thermo Fisher Scientific | Cat# PA5-27947 | WB (1:1000) |
| Antibody | EGFP (Mouse monoclonal) | Clontech | Cat# 632569 | WB (1:1000) |
| Antibody | TMEM79 (Rabbit Polyclonal) | Sigma-Aldrich | Cat# SAB1305998 | IF (1:200) |
| Antibody | TGN46 (Mouse monoclonal) | Sigma-Aldrich | Cat# SAB4200235 | IF (1:200) |
| Antibody | Lamp1 (Mouse monoclonal) | Santa Cruz | Cat# sc-20011 | IF (1:500) |
| Antibody | EEA1 (Mouse monoclonal) | Santa Cruz | Cat# sc-365652 | IF (1:500) |
| Antibody | PDI (Mouse monoclonal) | Thermo Fisher Scientific | Cat# MA3-019 | IF (1:200) |
| Antibody | Anti-Calreticulin antibody (Mouse monoclonal) | Abcam | Cat# ab22683 | IF (1:200) |
| Antibody | Alexa Fluor 488 (Goat Polyclonal) | Invitrogen | Cat# A-11001 | IF (1:500) |
| Antibody | Alexa Fluor 594 (Goat Polyclonal) | Invitrogen | Cat# A-11037 | IF (1:500) |
| Recombinant DNA reagent | pCS2-Tmem79-Myc-Flag (plasmid) | Origene | MR206106 | |
| Recombinant DNA reagent | pCS2-Tmem79-Myc (plasmid) | This paper | N/A | He lab; See ‘Plasmids’ |
| Recombinant DNA reagent | pCS2-Tmem79-Flag (plasmid) | This paper | N/A | He lab; See‘Plasmids’ |
| Recombinant DNA reagent | pCS2-V5-FZD5 (plasmid) | This paper | N/A | He lab; See ‘Plasmids’ |
| Recombinant DNA reagent | pCS2-V5-FZD5-K0 (plasmid) | This paper | N/A | He lab; See‘Plasmid’ |
| Recombinant DNA reagent | pCS2-2xHA-FZD1 (plasmid) | He Lab; PMID:17400545 | N/A | |
| Recombinant DNA reagent | pCS2-2xHA-FZD2 (plasmid) | He Lab; PMID:17400545 | N/A | |
| Recombinant DNA reagent | pCS2-2xHA-FZD3 (plasmid) | He Lab; PMID:17400545 | N/A | |
| Recombinant DNA reagent | pCS2-2xHA-FZD4 (plasmid) | He Lab; PMID:17400545 | N/A | |
| Recombinant DNA reagent | pCS2-2xHA -FZD5 (plasmid) | He Lab; PMID:17400545 | N/A | |
| Recombinant DNA reagent | pCS2-2xHA-FZD6 (plasmid) | He Lab; PMID:17400545 | N/A | |
| Recombinant DNA reagent | pCS2-2xHA-FZD7 (plasmid) | He Lab; PMID:17400545 | N/A | |
| Recombinant DNA reagent | pCS2-2xHA-FZD8 (plasmid) | He Lab; PMID:17400545 | N/A | |
| Recombinant DNA reagent | pRK5-FZD9 (plasmid) | Addgene | Plasmid #42261 | |
| Recombinant DNA reagent | pCS2-2xHA-FZD10 (plasmid) | He Lab; PMID:17400545 | N/A | |
| Sequence-based reagent | siRNA Universal Negative Control | Sigma-Aldrich | SIC001 | |
| Sequence-based reagent | siRNA to TMEM79 1# | Sigma-Aldrich | See ‘RNA interference’ | |
| Sequence-based reagent | siRNA to TMEM79 1# | Sigma-Aldrich | See ‘RNA interference’ | |
| Sequence-based reagent | Tmem79.S Morpholino | Gene Tools | See ‘Morpholino injection’ | |
| Sequence-based reagent | Tmem79.L Morpholino | Gene Tools | See ‘Morpholino injection’ | |
| Sequence-based reagent | Usp8 Morpholino | Gene Tools | See ‘Morpholino injection’ | |
| Peptide, recombinant protein | FLAG Peptide | Sigma-Aldrich | Cat# F3290 | |
| Peptide, recombinant protein | c-MYC Peptide | Sigma-Aldrich | Cat# M2435 | |
| Peptide, recombinant protein | Recombinant Human EGF | Peprotech | Cat# AF-100–15 | |
| Peptide, recombinant protein | bFGF2 | Prospecbio | Cat# CYT-218-a | |
| Peptide, recombinant protein | Recombinant Human SHH | Peprotech | Cat# 100–45 | |
| Peptide, recombinant protein | Recombinant Murine Noggin | Peprotech | Cat# 250–38 | |
| Commercial assay or kit | Fugene HD transfection reagent | Promega | Cat# E2312 | |
| Commercial assay or kit | Lipofectamine RNAiMAX | Thermo Fisher Scientific | Cat# 13778075 | |
| Commercial assay or kit | Dual-Luciferase Reporter Assay System | Promega | Cat# E1910 | |
| Commercial assay or kit | RNeasy Mini Kit | Qiagen | Cat# 74106 | |
| Commercial assay or kit | SilverQuest Silver Staining Kit | Thermo Fisher Scientific | Cat# LC6070 | |
| Commercial assay or kit | SuperScript IV Reverse Transcriptase | Thermo Fisher Scientific | Cat# 18090200 | |
| Commercial assay or kit | Taq DNA Polymerase, recombinant | Thermo Fisher Scientific | Cat# 10342020 | |
| Commercial assay or kit | DIG RNA Labeling Mix | S Sigma-Aldrich | Cat# 11277073910 | |
| Commercial assay or kit | Zero Blunt TOPO PCR Cloning Kit | Thermo Fisher Scientific | Cat# K2875J10 | |
| Chemical compound, drug | CHIR-99021 | Selleckchem | Cat# S1263 | |
| Chemical compound, drug | IWP-2 | Sigma-Aldrich | Cat# I0536 | |
| Chemical compound, drug | Bafilomycin A1 | Sigma-Aldrich | Cat# B1793 | |
| Chemical compound, drug | MG132 | Sigma-Aldrich | Cat# S2619 | |
| Chemical compound, drug | Digitonin | Sigma-Aldrich | Cat# D141 | |
| Chemical compound, drug | Sulfo-NHS-SS-Biotin | Thermo Fisher Scientific | Cat# 21328 | |
| Software, algorithm | Fiji | PMID:22743772 | RRID:SCR_002285 | |
| Software, algorithm | Mageck | Li et al., 2014 PMID:25476604 | ||
| Other | ANTI-FLAG M2 Affinity Gel | Sigma-Aldrich | Cat# A2220 | |
| Other | Anti-V5 Agarose Affinity Gel | Sigma-Aldrich | Cat# A7345 | |
| Other | Anti-c-Myc Agarose Gel | Pierce | Cat# 20168 | |
| Other | Anti-HA Agarose | Pierce | Cat# 26181 | |
| Other | Ni-NTA Agarose | Qiagen | Cat# 30230 |
Additional files
-
Supplementary file 1
Supplementary File 1A.
Number of embryos examined for anterior phenotypes in Figure 6D. Supplementary File 1B. Number of embryos examined for pigmentation phenotype in Figure 6D. Supplementary File 1C. Number of embryos examined for neural versus epidermal marker expression in Figure 6F. Supplementary File 1D. Number of embryos examined for spina bifida in Figure 7B. Supplementary File 1E. Number of embryos examined for axial elongation defect in Figure 7D. Supplementary File 1F. Number of animal pole explants examined for axial mesoderm elongation in Figure 7F. Supplementary File 1G. Number of embryos examined for enlarged head or spina bifida in Figure 6—figure supplement 1B. Supplementary File 1H. Number of embryos examined for anterior marker expression in Figure 6—figure supplement 1G. Supplementary File 1I. Number of embryos examined for anterior phenotypes in Figure 6—figure supplement 1F. Supplementary File 1J. Number of embryos examined for dorsal/organizer marker expression in Figure 6—figure supplement 1G. Supplementary File 1K. Number of embryos examined for neural crest marker expression in Figure 6—figure supplement 1H.
- https://cdn.elifesciences.org/articles/56793/elife-56793-supp1-v2.docx
-
Transparent reporting form
- https://cdn.elifesciences.org/articles/56793/elife-56793-transrepform-v2.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
TMEM79/MATTRIN defines a pathway for Frizzled regulation and is required for Xenopus embryogenesis
eLife 9:e56793.
https://doi.org/10.7554/eLife.56793




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}