Characterization of convergent thickening, a major convergence force producing morphogenic movement in amphibians
- Department of Biology, University of Virginia, United States
- Department of Cell Biology, University of Virginia, School of Medicine, United States
- Department of Cell and Systems Biology, University of Toronto, Canada
- Centre for Craniofacial and Regenerative Biology, King's College London, United Kingdom
- Aduro Biotech, United States
- Independent researcher, United States
Figures
Figure 1 with 1 supplement
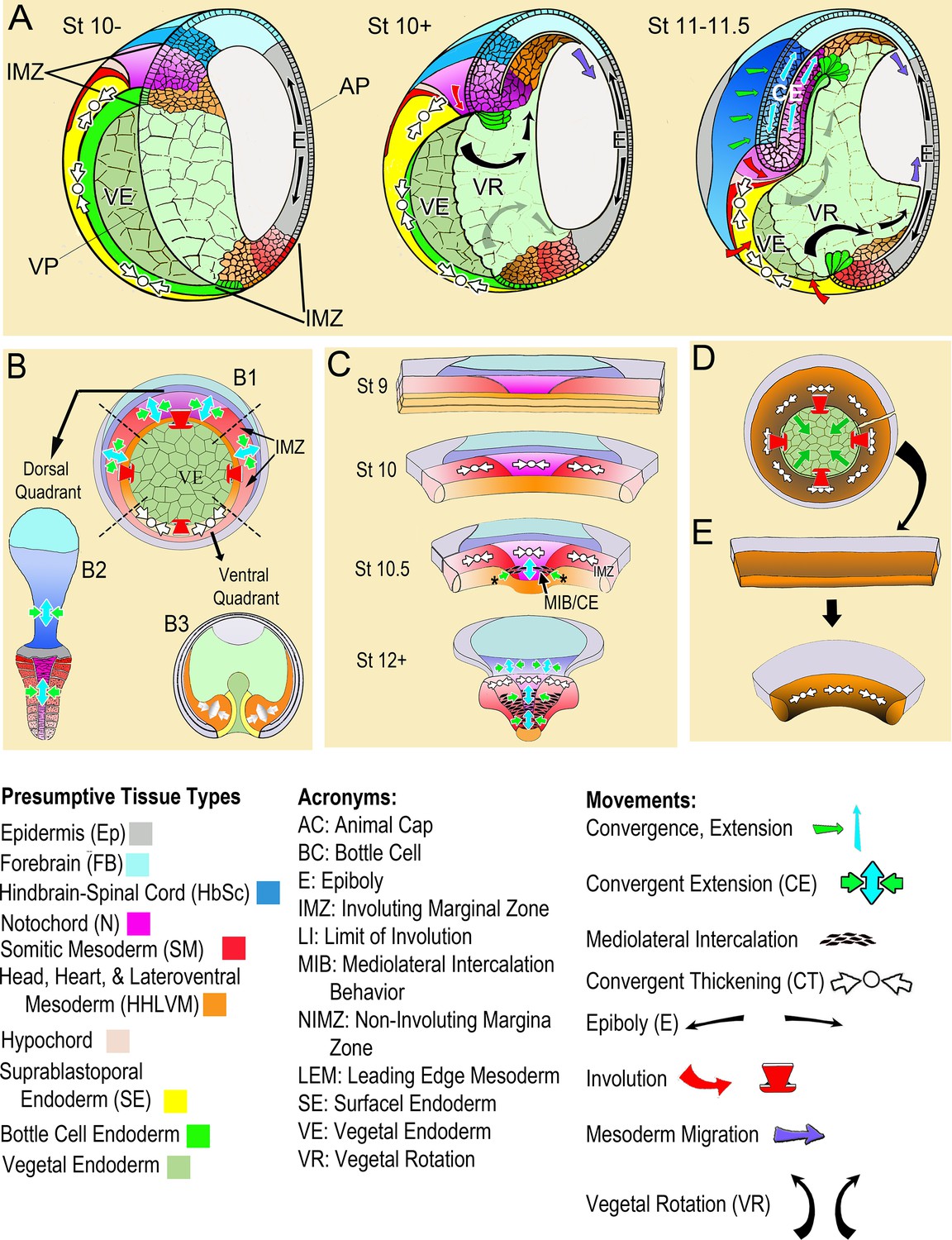
Morphogenic movements in the embryo and explants.
(A) The behavior of the involuting marginal zone (IMZ) in blastopore closure is shown in the pre-gastrula embryo (St 10-) and at subsequent stages of gastrulation (St. 10+, 11–11.5) in mid-sagittal cut-aways rotated 45°, within the context of the whole embryo and the other morphogenic movements. This includes the animal and vegetal poles (AP, VP), the involuting marginal zone (IMZ), the vegetal endoderm (VE), and the morphogenic movements convergent thickening (CT, white arrows), epiboly (E, black arrows within epidermis), vegetal rotation (VR, gray arrows/black arrows within VE), mesendoderm migration (purple arrows), involution (red arrows), and convergent extension (CE, green (C) and blue (E) arrows within neural and mesodermal tissues). See legend of tissue colors, acronyms and movements at the bottom of the figure. The IMZ is a ring of several layers of deep cells, plus a superficial epithelium, that lies above and around the VE prior to gastrulation (St 10-). Deep IMZ cells are presumptive mesoderm whereas the superficial cells are primarily presumptive endoderm, with some presumptive mesoderm dorsally (Shook et al., 2004). After involution, the early involuting tissues originating from the lower (vegetal) region of the IMZ, including the presumptive head, heart, and lateroventral mesoderm (orange), migrate across the blastocoel roof (St 10+, 11–11.5, purple arrows), whereas tissues originating from the upper (animal) region of the IMZ, the presumptive notochordal (magenta) and somitic (red) mesoderm engage in CE (green and blue arrows), as does the overlying neural tissue (dark blue). Only the most recently involuted notochordal and somitic tissue, lying just around the inside of the blastopore directly generate convergence forces closing the blastopore by CE. The non-involuting marginal zone (NIMZ) is the ring of tissue above the IMZ that bounds the blastopore at the end of gastrulation, with its vegetal edge being the ‘Limit of Involution’ (LI; see Figure 1—figure supplement 1B). The dorsal and dorsolateral sectors of the NIMZ contains the posterior neural tissues (Figure 1A St 11–11.5, darker blue cells), which will undergo CE in parallel to the underlying somitic and notochordal mesoderm, driven by an intercalation behavior similar to MIB (Keller et al., 1992; Elul et al., 1997; Elul and Keller, 2000; Ezin et al., 2003; Ezin et al., 2006). (B) A vegetal view of the early gastrula shown without the superficial, epithelial layer over the IMZ reveals the underlying deep presumptive mesoderm (B1). Sandwich explants of the dorsal quadrant of the Marginal Zone (IMZ plus NIMZ, B2), including tissues of presumptive notochordal (magenta), somitic (red), and neural (blue) fates undergo CE (green and blue symbols)(for detailed anatomy of explants, see Figure 1—figure supplement 1). In contrast, explants of the ventral quadrant (B3) develop only ventrolateral mesoderm (orange) when separated from the organizer (‘dorsal’) side of the embryo, round up and show a thickening of the mesodermal tissues, a movement called Convergent Thickening (CT, white symbols). Together, CE on the dorsal side and CT on the ventral side of the IMZ were previously thought to provide the convergence forces that close the blastopore in the intact embryo (Keller and Danilchik, 1988) (B1). (C) In the current work, a more detailed analysis of ‘giant’ sandwich explants of the entire Marginal Zone, and variants thereof (Figure 1—figure supplement 1A), reveals a revised view of the pattern of CT expression, wherein the entire IMZ uniformly expresses CT during early gastrulation (white symbols, St 10). At the midgastrula stage, expression of CT begins to transition into a progressive expression of CE (green and blue arrows, St 10.5), a post-involution progression from anterior to posterior, as described previously (Shih and Keller, 1992a), while CT continues in the pre-involution IMZ; CE also begins in the posterior neural tissue (St 10.5–12+). (D) Lastly, ventralized embryos, which lack the presumptive tissues of notochordal and somitic mesoderm and contain only presumptive ventrolateral tissues, involute their marginal zone (red arrows) and close their blastopore symmetrically (green arrows), using CT alone (white symbols), and giant explants of ventralized embryos (E), consisting entirely of ventral tissues, undergo uniform CT. For a complete description of these and other types of explants, see Materials and methods and Figure 1—figure supplement 1.
Figure 1—figure supplement 1
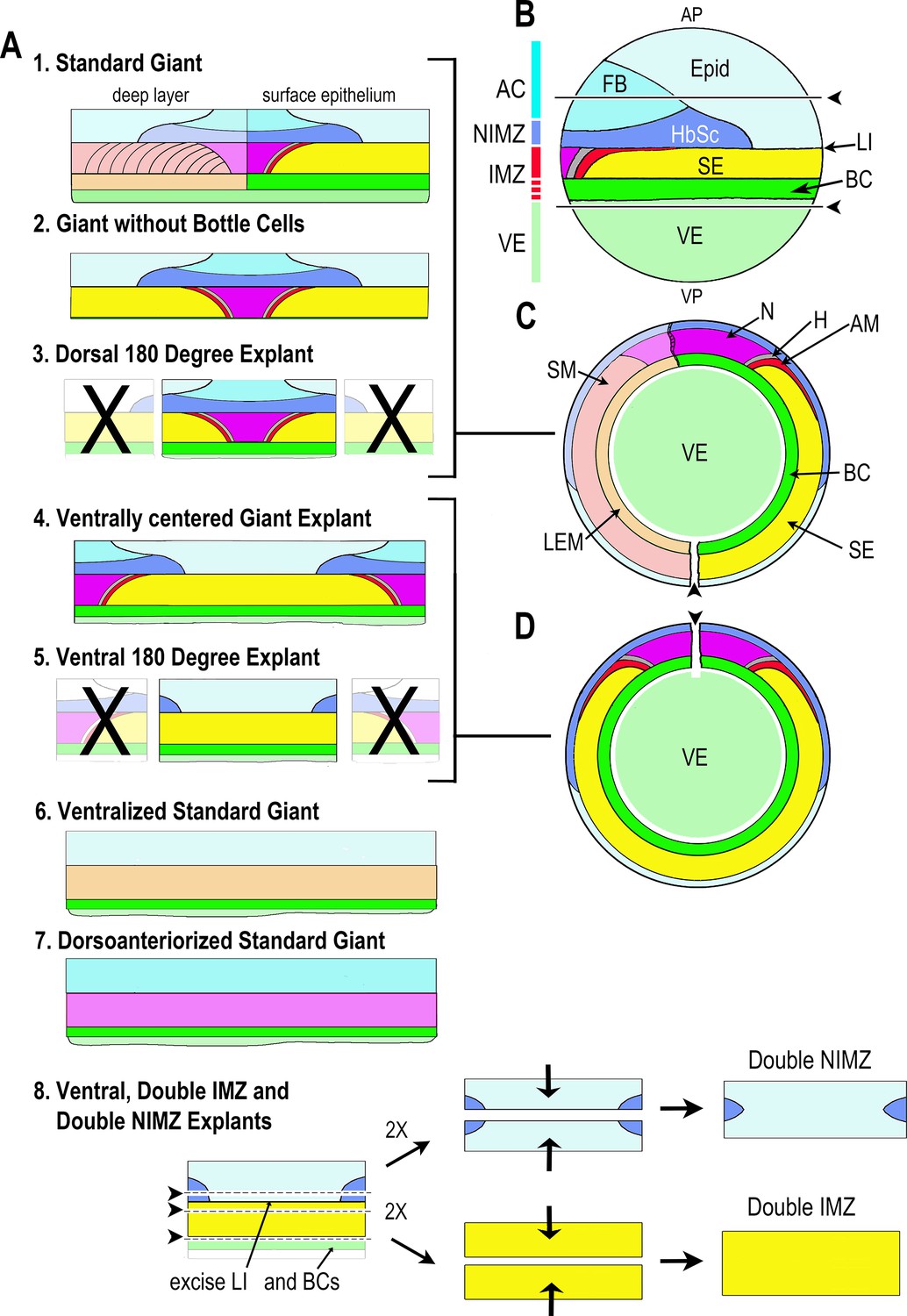
Description of explant construction.
Various types of sandwich explants that included some or all of the Marginal Zone were made at Stage 9–10.25 for use in this work (A). Lateral view of the pre-gastrulation embryo (B) (Stage 10-) with animal pole (AP) at the top and vegetal pole (VP) at the bottom, showing presumptive tissues and the ranges of the AC, NIMZ, IMZ, and VE regions (colored bar). AC and NIMZ are ectodermal, the deep layers of the IMZ are mesodermal, whereas the superficial layer is either mesodermal (dorsally) or endodermal (laterally and ventrally); see below for details. Upper and lower regions of the IMZ are indicated (solid vs. broken red). The Limit of Involution (LI) lies at the boundary between the NIMZ and the IMZ and divides the mesodermal and endodermal tissues that will be internalized (below the LI) from the ectodermal tissues that will remain outside (above the LI). Images of whole embryos showing tissue fates are shown in lateral (B) and in vegetal view, with the epithelial tissues removed to show the underlying deep tissues in the left half (C) and with the epithelial tissues left intact (D). Presumptive tissue fates in the superficial (epithelial) layer: Light blue = epidermis (Epid), darker blues = neurectoderm (Forebrain (FB), hindbrain-spinal cord (HbSc)); magenta = notochord (N), gray = hypochord (H), red = adaxial somitic mesoderm (AM); yellow = supra-blastoporal endoderm (SE), bright green = presumptive bottle cells (BC, also SE); pale green is vegetal endoderm (VE) or ‘sub-blastoporal’ endoderm. Presumptive fates in the deep layer of the Marginal Zone are shown in the left half of C and A1, and in the entire IMZ in A6,7: light orange = leading edge mesoderm (LEM, migratory mesoderm); light red = somitic mesoderm (SM); light magenta = notochordal mesoderm. Standard ”Giant” sandwich explants (A1) were made of the entire Marginal Zone with bottle cells and some vegetal endoderm included (Shook et al., 2004), the ‘standard’ version for this work. Embryos were first cut ventrally (pointer in C), then AC and VE tissue removed as indicated (above and below pointers and lines in B) and VE was removed from the inner face of the IMZ (see Figure 1A, St 10-). Two such explants were then pressed under cover glass to make a sandwich, inner face to inner face, to allow the epithelium to heal together around the edges. To make Giants without bottle cells (A2), all VE and some or all of the bottle cells were additionally removed from the explants prior to sandwiching (Poznanski et al., 1997). Ventrally centered giant sandwich explants (A4) were made as for standard giants, but initially cutting along the dorsal (pointer in D), rather than ventral midline. Dorsal 180° (D180) (A3), and Ventral 180° (V180) (A5) explants are made as standard or ventrally centered giants, with the ventral or dorsal 180°, respectively, of the marginal zone cut off. Ventralized (A6) or dorsoanteriorized (A7) giant explants were made from ventralized or dorsoanteriorized embryos, cut without reference to ‘dorsal’, as these embryos are symmetrical about the blastopore (Scharf and Gerhart, 1980; Kao and Elinson, 1988). Double ventral or dorsal, IMZ or NIMZ only sandwich explants (A8) were made by first constructing V180 explants with most of the presumptive bottle cells removed, allowing them to heal together at least 15 min, then cutting out the NIMZ and IMZ regions, avoiding the LI, and recombining two NIMZ- or two IMZ-only pieces with their newly cut posterior edges (those closest to the LI) juxtaposed and pressed together by two cover glass bits against the outer, anterior surfaces of the individual pieces. The epithelia at the cut edges heal together rapidly, preventing artifacts resulting from extended healing of the epithelium over the cut face. The doubled explants were allowed to heal for 30 min to 1 hr before being moved onto a cover glass platform in front of a 45° mirror in the culture dish, with the lateral edge of the explant facing the mirror, such that we could image both convergence (from above) and thickening (from the side). Convergence is reported from the point at which cover glass is lifted from the initial sandwiches, prior to cutting apart the IMZ and NIMZ portions of the explants. Thickness is reported from the time the two IMZ or NIMZ regions heal together. As a consequence of the explant construction process, our measures do not capture the initial ~25% convergence of the IMZ with respect to its initial mediolateral extent in the embryo or the initial ~50% thickening of the IMZ with respect to its initial thickness in the embryo. NIMZ explants show little or no change in thickness over the course of explant construction. See Materials and methods for general details of explant construction.
Figure 2 with 1 supplement
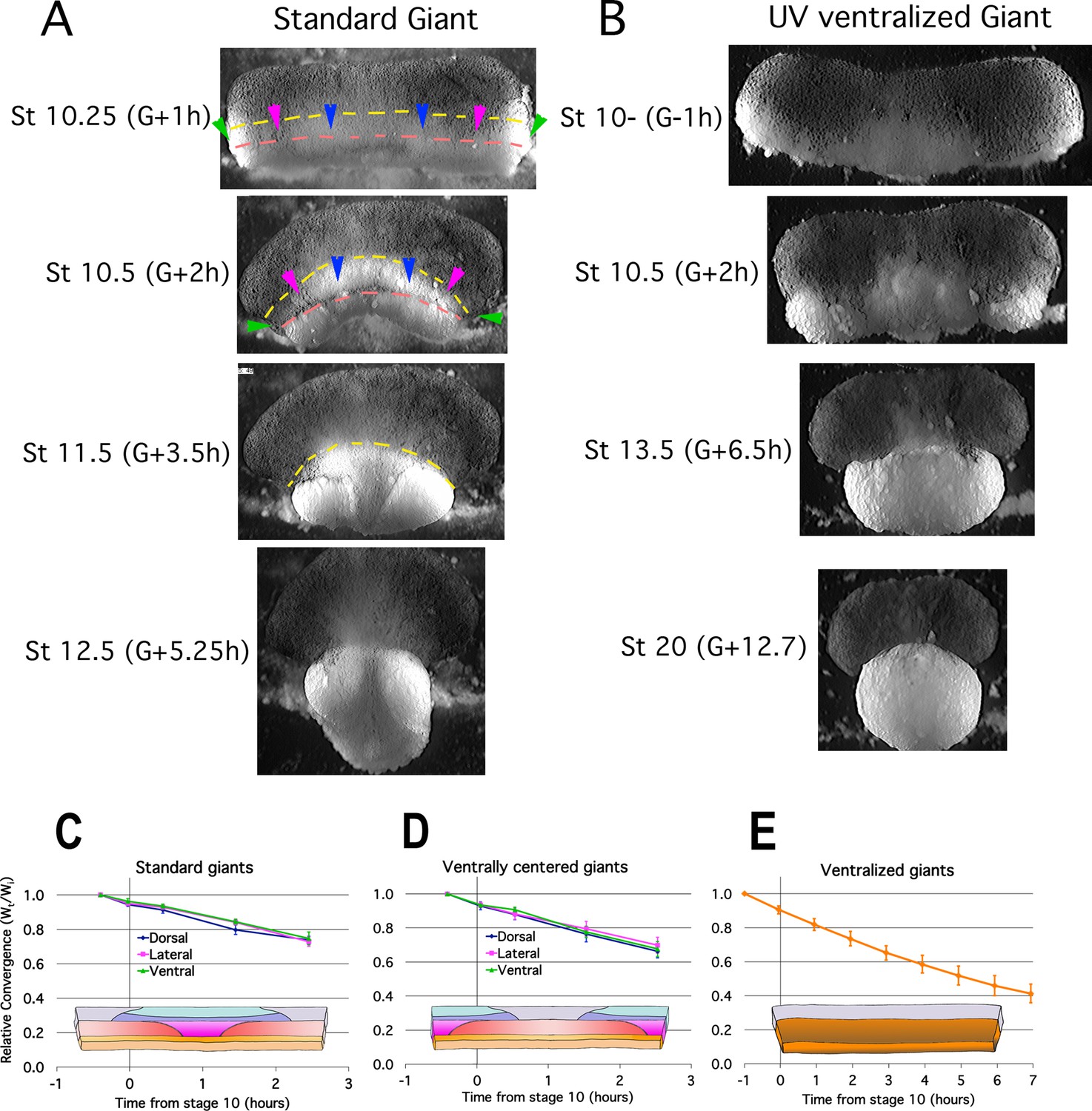
Early Convergence is uniform across regions.
Frames from time lapse movies of standard giant sandwich explants (A; Video 2*, left) or ventralized standard giants (B; Video 3*) after release from their constraining coverslips show uniform convergence of the IMZ. The upper limits of the upper and lower IMZ are indicated by dashed yellow and pink lines, respectively (A). To compare regional rates of convergence in such explants, the mediolateral extent of the dorsal (A, between blue pointers), lateral (between blue and magenta pointers), and ventral regions (between magenta and green pointers) of the IMZ were measured at times from Stage 10- and plotted with respect to initial width (Wt/Wi) in standard (dorsally centered) giants (n = 3–5 embryos per time point) (C) and ventrally centered giants (n = 4–5 embryos) (D). The convergence of the entire IMZ was also measured in giants from ventralized embryos (n = 3 embryos) (E); error bars = SEM. For C-E, the same explants were measured at sequential time points.
-
Figure 2—source data 1
Figure 2C, D source data: Relative live explant convergence vs. time.
- https://cdn.elifesciences.org/articles/57642/elife-57642-fig2-data1-v2.xlsx
Figure 2—figure supplement 1
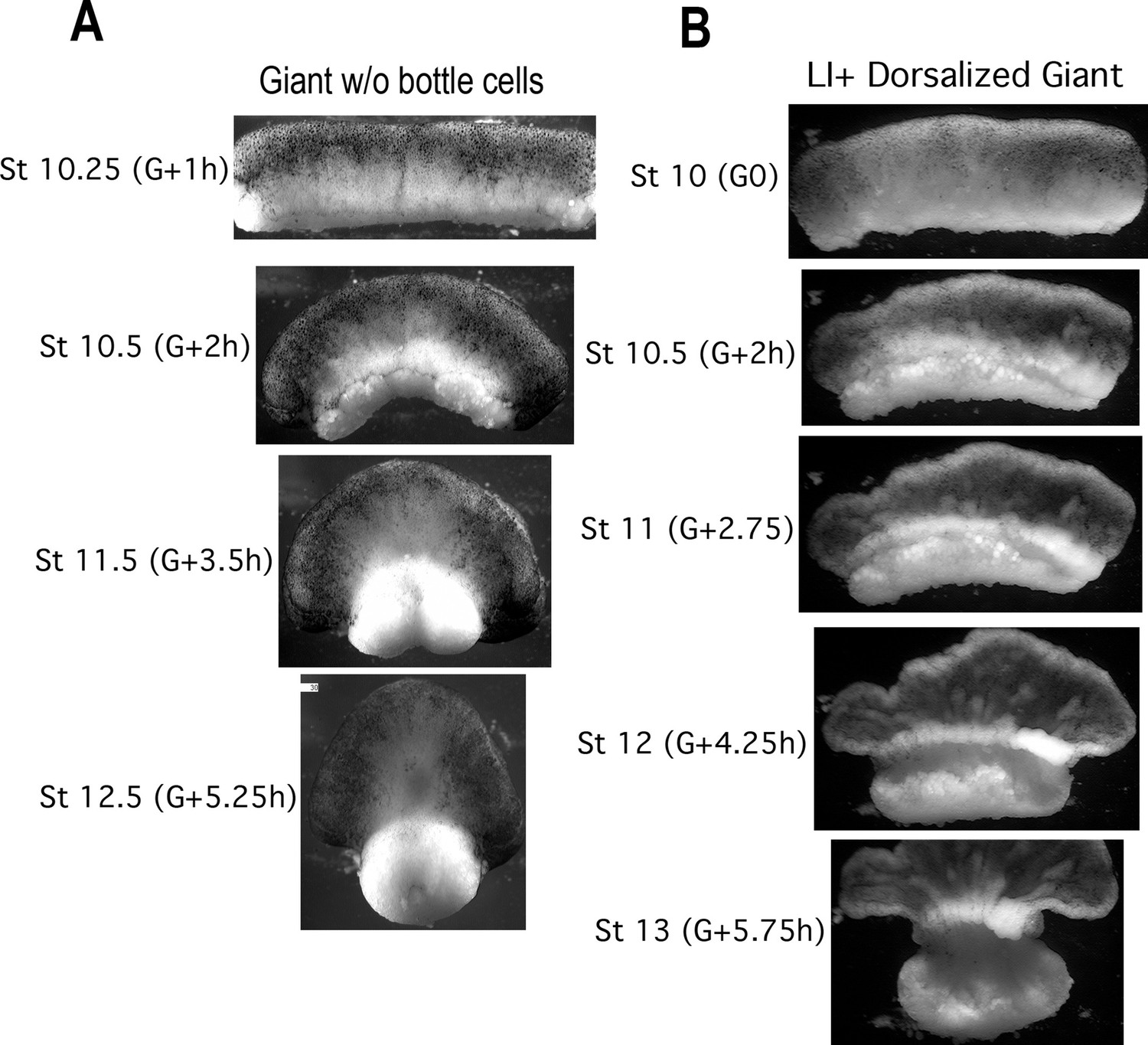
CT in giants without bottle cells and in Li+ Dorsoanteriorized giant sandwich explants.
Giants made without bottle cells (A, Video 2, right) also show isotropic convergence and thickening of the IMZ throughout gastrulation in control embryos (stages and hours from the onset of gastrulation (G + Xh) indicated). Giants made from dorsoanteriorized embryos (B, Video 5) show isotropic convergence and thickening of the IMZ through at least Stage 10.5, when they begin isotropic convergence driving anterior-posterior extension; a thickened mass remains in the more animal portion of the IMZ and feeds into the converging and extending tissue.
Figure 3 with 2 supplements
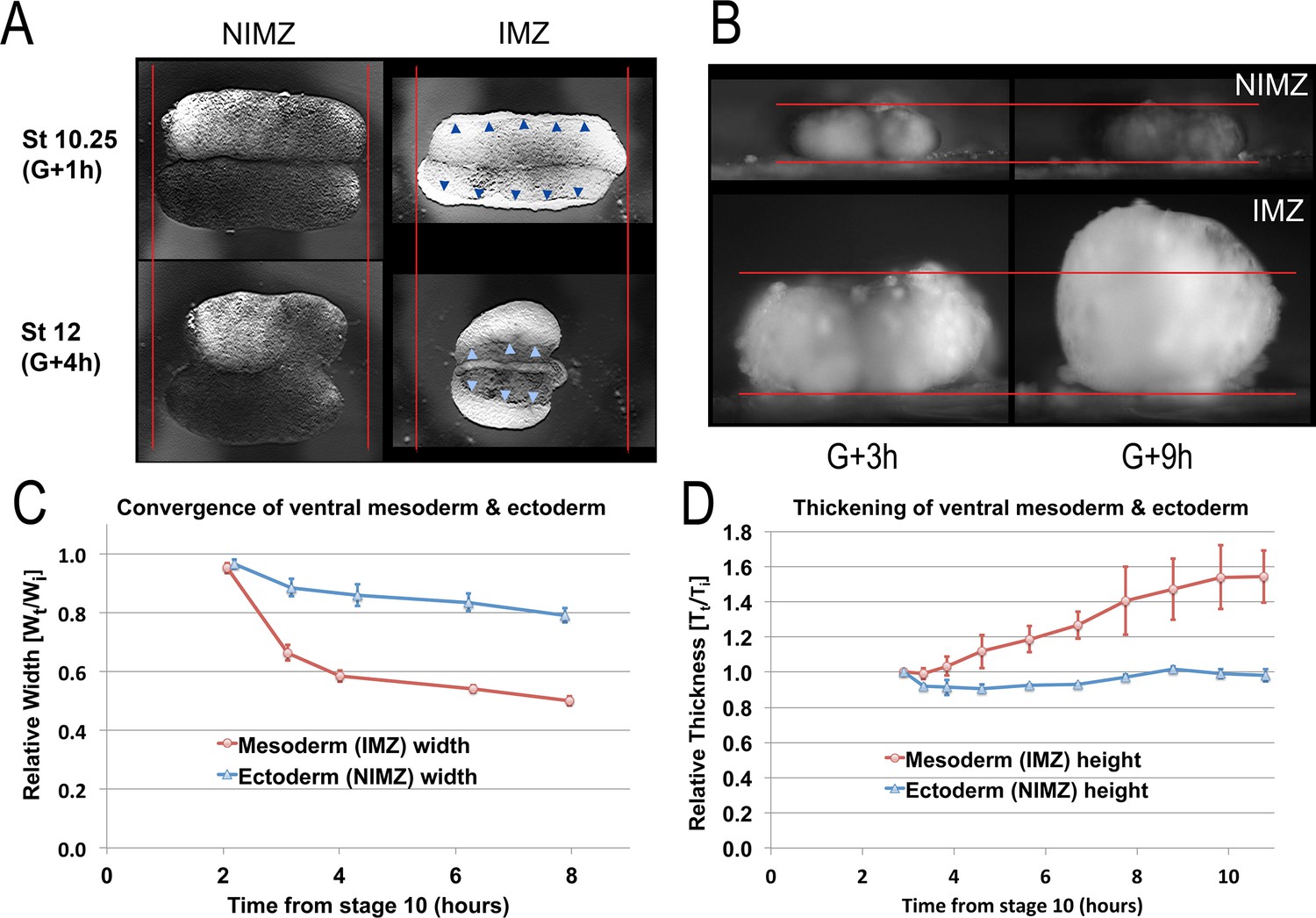
Quantitation of convergence and thickening in IMZ vs. NIMZ regions of explants.
Double ventral NIMZ and IMZ sandwich explants (see Figure 1—figure supplement 1A8) were used to examine the autonomous rates of convergence and thickening in each region, by imaging both from above (A, Video 9*) and from the side, viewing the lateral edge (B, Video 10*); pointers in A indicate the edge of the superficial epithelium on the IMZ explant, red lines in A and B indicate the initial width or height of the explants. Explant width (C; n = 6 explants per time point) and thickness (D; n = 2 explants per time point) were measured over time (Wt and Tt respectively; see Materials and methods for details) and plotted with respect to the initial value (Wi and Ti); error bars = SEM. Measures for C and D were made from the same explants at sequential time points.
-
Figure 3—source data 1
Figure 3C, D source data: Relative live explant thickness and width vs. time.
- https://cdn.elifesciences.org/articles/57642/elife-57642-fig3-data1-v2.xlsx
Figure 3—figure supplement 1
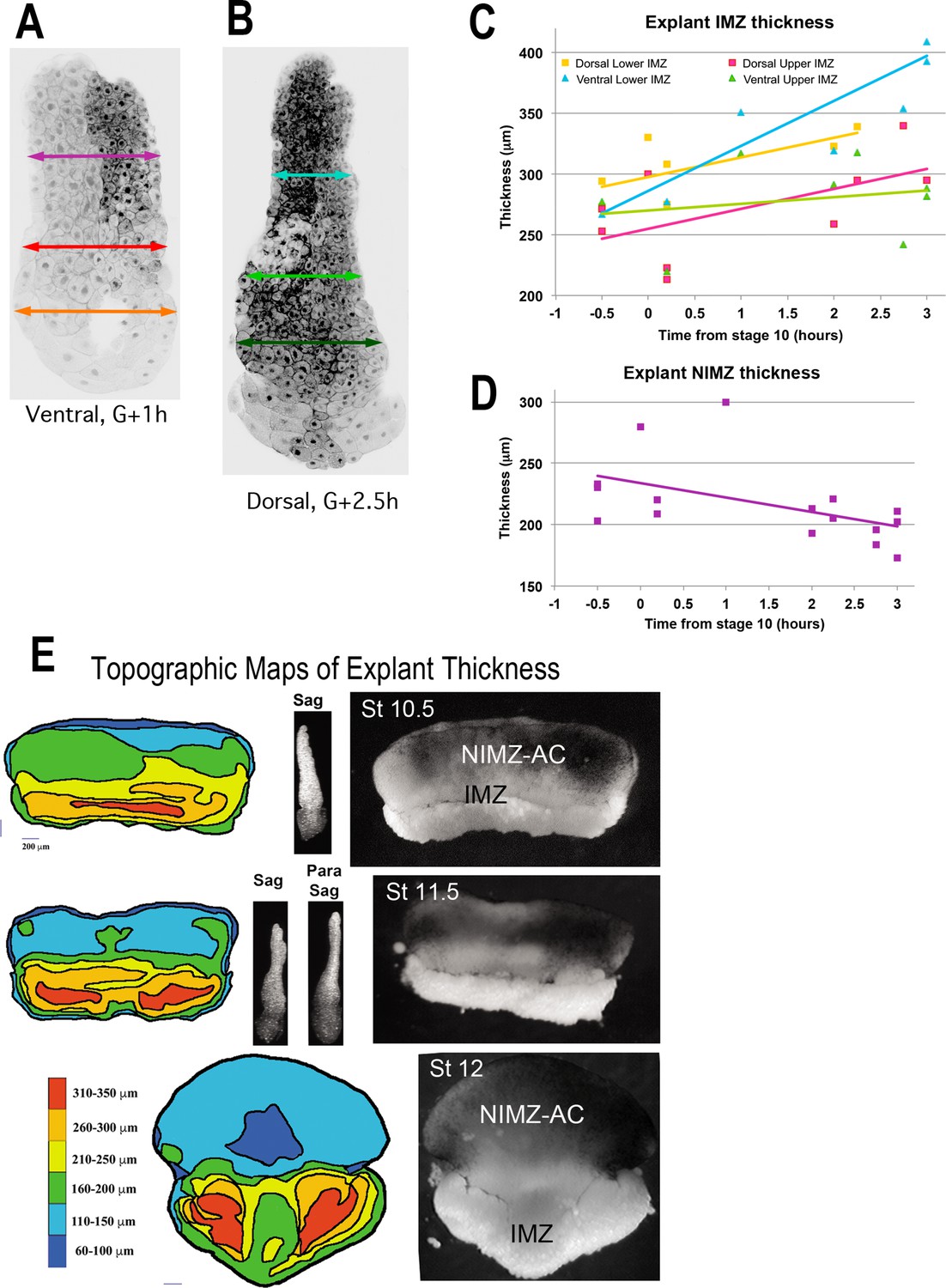
Giant sandwich explants were made from rhodamine dextran amine (RDA) labeled embryos (A,B) prior to the onset of gastrulation, allowed to heal under a cover glass for 30 minutes, then allowed to equilibrate for at least 1 hour after cover glass removal, prior to fixation.
Explants were then cut parasagittally in the dorsal and ventral regions and the resulting “chops” were imaged with LSCM parallel to the cut surface. The total thickness of the lower and upper IMZ and the NIMZ as in the examples shown (A, B), were measured for explants fixed at timepoints (C, D). This revealed that the lower portion of the ventral IMZ thickened the most during early gastrulation, whereas the upper portion of the ventral IMZ thickened the least; similarly, the lower dorsal IMZ remained thicker than the upper dorsal IMZ (C). During this period, the NIMZ of both thinned (D). For C and D, each time point represents a single measure; 1–2 measures were made from each of 5–7 different explants, fixed at various time point. These data were highly variable in these types of explants, especially in the IMZ where the subtle variations of explant construction come into play. To resolve the spatial pattern of thickening of the pre-involution IMZ in explants, we made standard giant explants at Stage 9.5 (G-1h) from RDA injected embryos, fixed them at mid-gastrula stages, and imaged them en face with LSCM to generate topographic maps of thickness (left column). Corresponding giant explants just prior to fixation (right column) and lateral views reconstructed from confocal stacks (center column). Explants just before onset of CE show a thickened IMZ (Stage 10.5, yellow and orange) and a thinner NIMZ (Stage 10.5, blue and green). The thickness at the vegetal edge of IMZ consists of the leading edge mesoderm and a small amount of attached VE (right, Stage 10.5–11.5, lighter tissue at vegetal edge; left, band contiguous with red), which rounds up after being exposed to the media by the retracting bottle cell epithelium. Beginning at Stage 11, the medial region of the IMZ begins thinning (Stage 11.5, 12), which reflects radial intercalation of the notochord and somitic tissues prior to MIB and CE (Wilson and Keller, 1991). The upper (animal) portion of the IMZ remains relatively thick across its top throughout gastrulation (left, Stage 11.5, upper orange and yellow band; Stage 12, medial yellow arc contiguous with upper orange laterally). The extreme thinning of the central region of NIMZ (left, Stage 12, dark blue), reflects the center part of the posterior neural ectoderm, which is undergoing CE at this point (see Keller and Danilchik, 1988).
-
Figure 3—figure supplement 1—source data 1
Figure 3—figure supplement 1C, D source data: Fixed explant thickness vs. time.
- https://cdn.elifesciences.org/articles/57642/elife-57642-fig3-figsupp1-data1-v2.xlsx
Figure 3—figure supplement 2
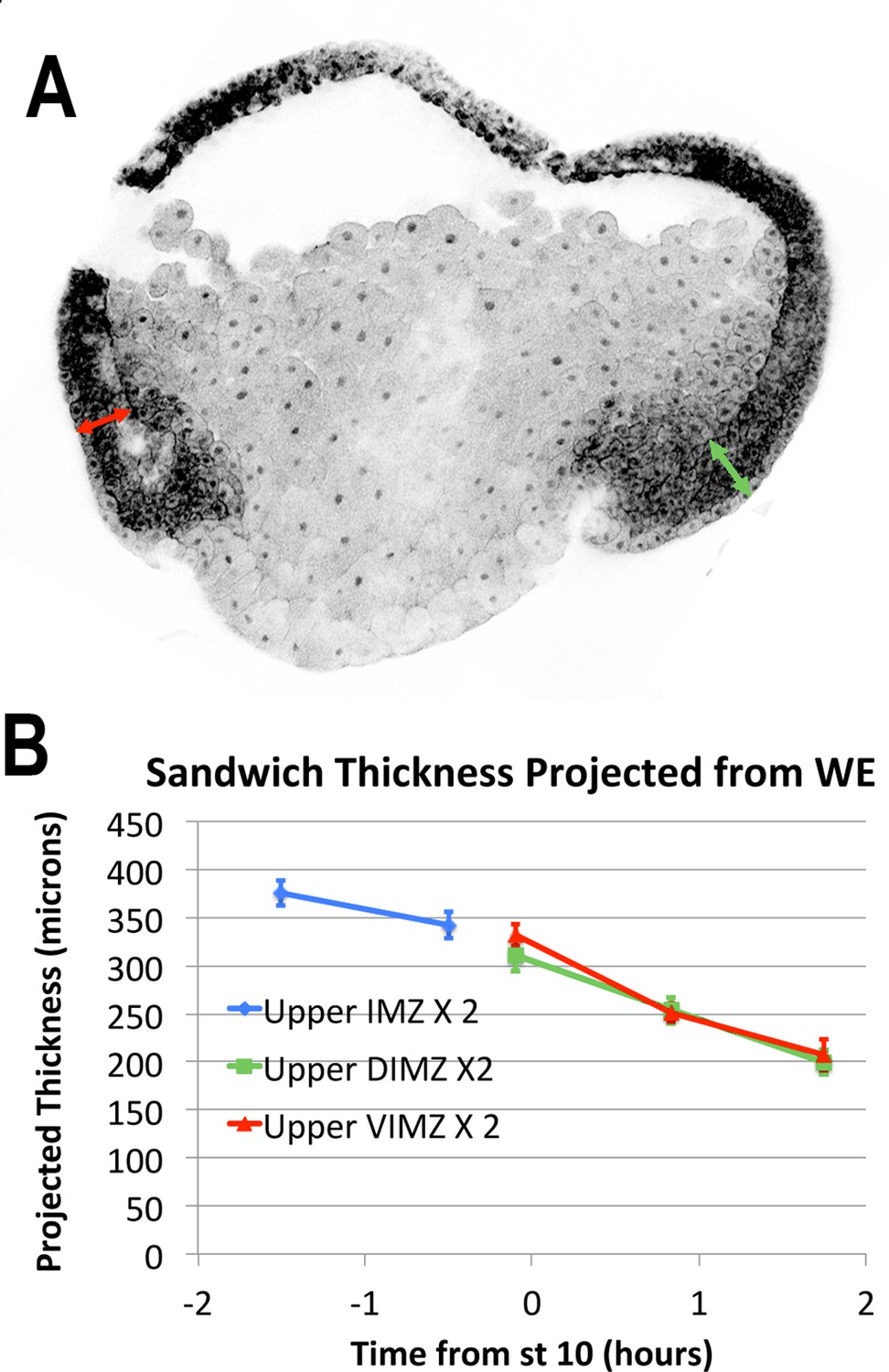
LSCM images of RDA injected embryos (A) were used to measure regional thickness of the IMZ and NIMZ, before and during early gastrulation.
In gastrula stage embryos, the thickness of the upper (more animal) IMZ was measured dorsally and ventrally (A, green and red double-headed arrows, respectively). The thickness of the lower IMZ in whole embryos after the onset of gastrulation is more ambiguous, so is not reported. In pre-gastrula embryos, upper IMZ thickness was measured on opposite sides and averaged (not shown). The expected thickness that would result from sandwich explants made from embryos at time points, assuming no change upon explantation, for purposes of comparison (B).
-
Figure 3—figure supplement 2—source data 1
Figure 3—figure supplement 2B source data: Projected sandwhich explant thickness.
- https://cdn.elifesciences.org/articles/57642/elife-57642-fig3-figsupp2-data1-v2.xlsx
Figure 4
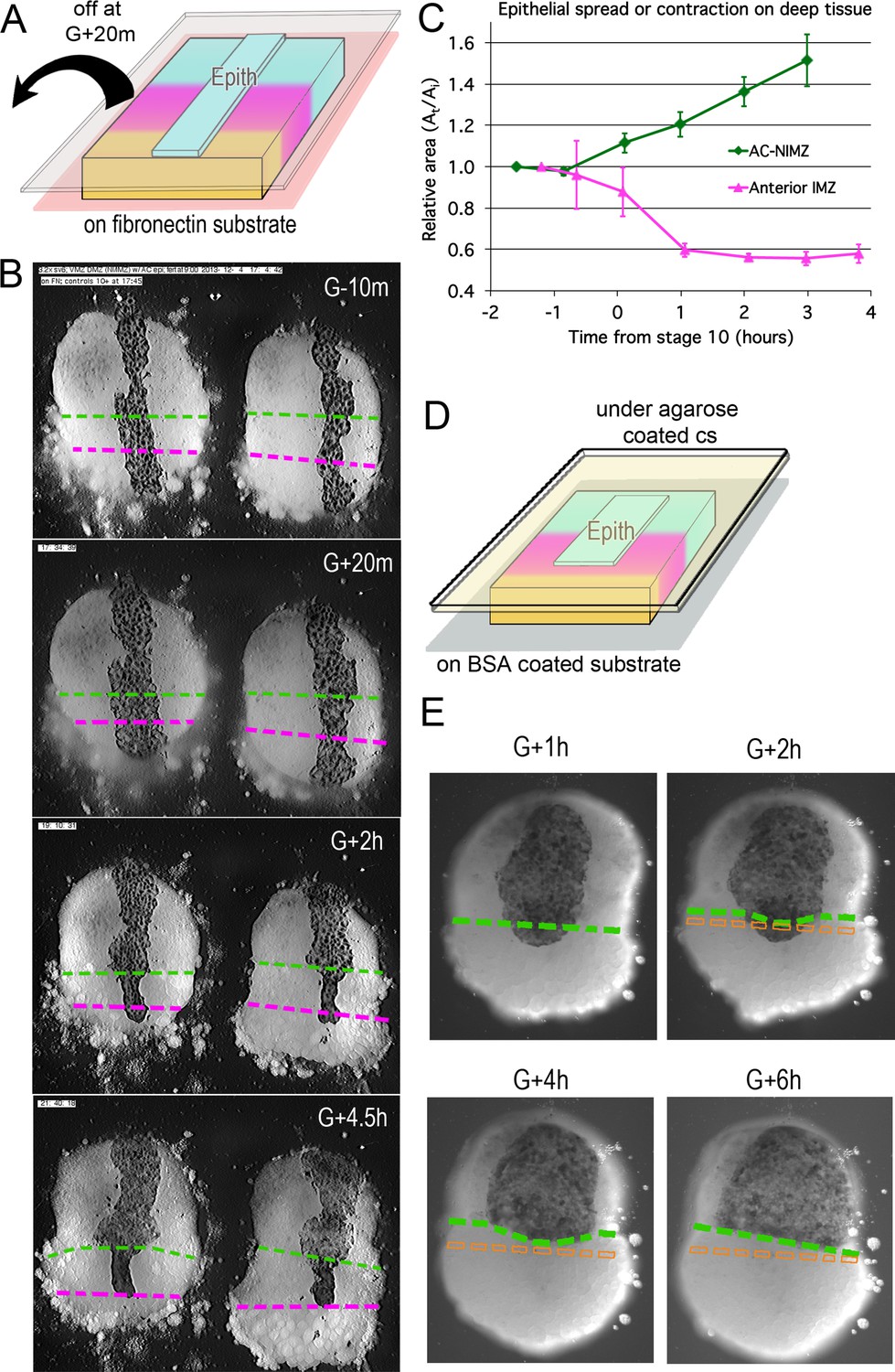
Epithelial affinity.
(A) Explants of deep IMZ plus NIMZ tissue were made prior to the onset of gastrulation and stabilized by adhering them to fibronectin-coated cover glass below the explant; strips of AC epithelium large enough to span the entire animal-vegetal extent of the deep tissue were subsequently grafted onto its surface and kept in place with cover glass above the explant. (B) In the examples shown (lower IMZ below magenta dashed line, upper IMZ between magenta and green, NIMZ & AC above green dashed line; boundaries between the three regions identified by cell, motility, size and gray cast), the movie (Video 11*, where these events can be most easily seen) begins 10 min prior to the onset of gastrulation (G-10m). The epithelium adhered and spread over all regions (G + 20 m), after which the upper cover glass was removed. Immediately afterward, epithelium over both NIMZ and IMZ regions contracted slightly, then that over the NIMZ re-spread, while that over the IMZ contracted more strongly and continuously, often to the point of becoming detached from the deep tissue, and in many cases retracted from the lower (vegetal) edge of the deep tissue (G + 2 h). Tissue over the IMZ remained contracted, while that over the NIMZ tended to spread (G + 4.5 h). (C) The relative area of spreading or contraction of the epithelial tissue over different deep tissue regions over time was quantitated, beginning after cover glass removal (each time point based on measures from 2 to 8 explants at sequential time points; error bars = SEM). (D) In a variation on this assay, explants of deep ventral IMZ plus NIMZ tissue were cultured on BSA coated cover glass (below) and patches of epithelium were lapped across the NIMZ/IMZ boundary and held apposed to the deep tissue with an agarose coated cover glass (above) throughout the experiment. In the example (E), the epithelium initially over the deep IMZ (below green line) first contracts animally (G + 2 h), eventually retracting across the boundary (G + 4 h), then respreads on the deep NIMZ (G + 6 h) (Video 12*, where these events can be most easily seen). During this time, upper deep IMZ cells (mesoderm) spread over deep NIMZ cells (ectoderm), such that the NIMZ/IMZ boundary on the surface of the deep tissue is shifted from its original position (dashed orange line). The deep NIMZ/IMZ boundary is based on differential cell size and motility, which are evident in the movie.
-
Figure 4—source data 1
Figure 4C source data: area of epithelia ove IMZ and NIMZ vs. time.
- https://cdn.elifesciences.org/articles/57642/elife-57642-fig4-data1-v2.xlsx
Figure 5 with 3 supplements
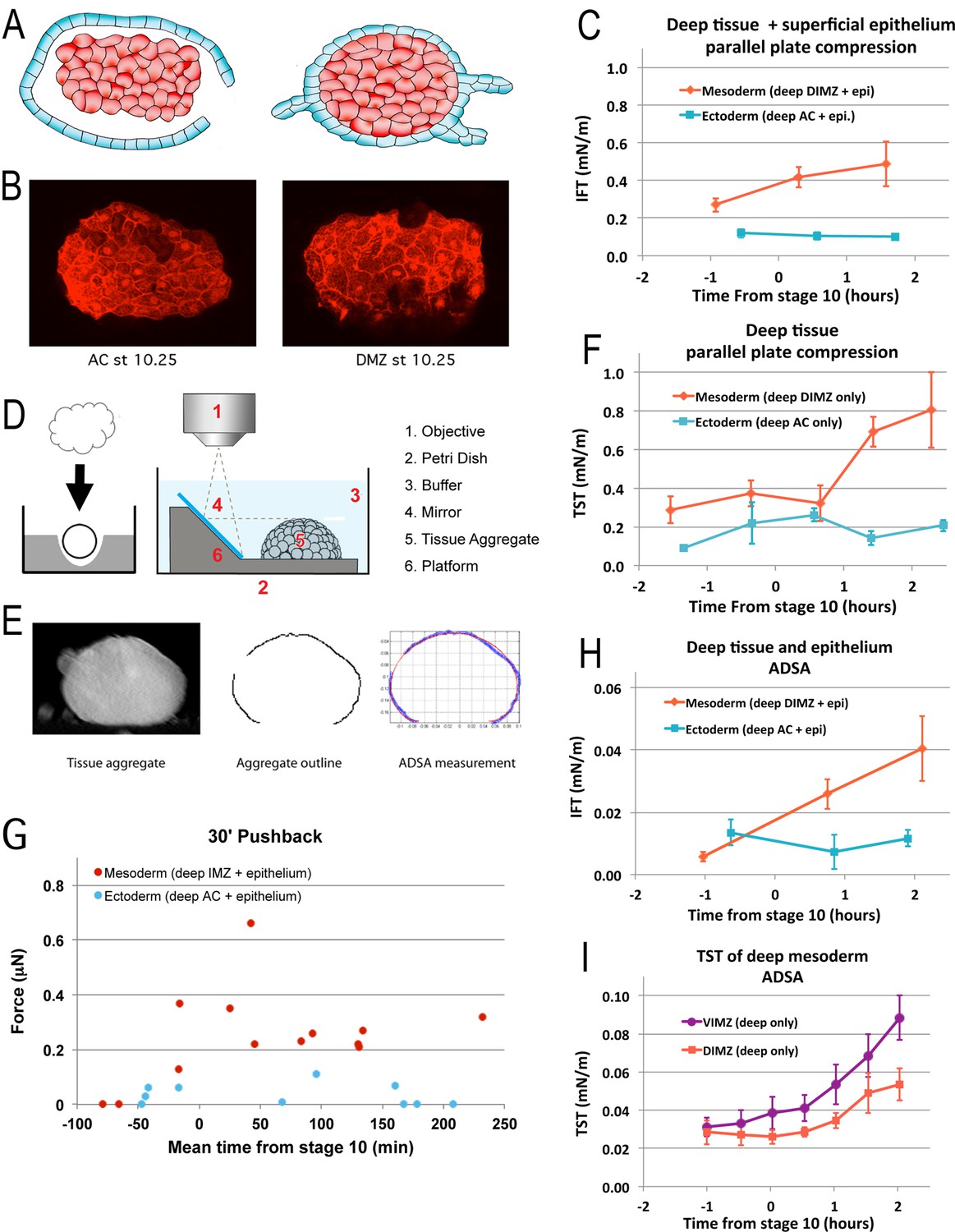
Tissue surface tension and interfacial tension rise in association with the onset of CT.
Aggregates of deep tissue wrapped in excess superficial epithelium (‘wontons’), (A), were used to measure the IFT between DIMZ (Mesoderm) or AC (Ectoderm) deep tissue and epithelium (C), assayed by parallel plate compression (using a MicroSquisher; see Materials and methods and Figure 5—figure supplement 1). Aggregates of deep mesodermal or ectodermal tissue, prepared by explanting deep tissue into an agarose well and allowing the explant to round up (D, left panel) were similarly used to measure TST (F). IFT and TST were determined using outline and analysis features of the MicroSquisher software, SquisherJoy (see Materials and methods and Figure 5—figure supplement 1). For IFT, the outline of the wrapped tissue was used as an approximation for the deep tissue. n’s = 4–9 for mesodermal and ectodermal IFT; n’s = 4–7 explants for mesodermal, 1–5 for ectodermal TST; error bars = SEM; the same explants were measured at sequential time points, roughly 1 hr apart. Axisymmetric drop shape analysis (ADSA)(David et al., 2009; Luu et al., 2011) of explants was also used to measure surface tensions. Wontons made with rhodamine dextran labeled deep tissues wrapped in unlabeled epithelium were allowed come to equilibrium and develop for a time (Video 13), then fixed, the wonton cut in half and LSCM used to capture the profile of each half (B). ADSA of the profile (E) was used to determine the IFT between the deep and superficial tissues (H); n’s = 3–8 explants per time point; error bars = SEM. To measure TST with ADSA, deep cell aggregates were prepared as above (D, left panel), then the profile of the aggregate under 1 g was imaged at sequential time points using a 45° mirror (D, right panel; Video 14*). ADSA was used on outlines of the aggregate to determine the TST of each aggregate over time (I); the mean of TSTs for dorsal and ventral IMZ tissue (DIMZ, VIMZ) was plotted over time (I); n’s = 4–6; error bars = SEM. The MicroSquisher was also used to determine the amount of compression force that wrapped deep tissue (IMZ wontons (Mesoderm) and AC wontons and sandwiches (Ectoderm)) could generate in 30 min (G); each point represents a single run.
-
Figure 5—source data 1
Figure 5C, F-I source data: IFT, TST or force vs. time.
- https://cdn.elifesciences.org/articles/57642/elife-57642-fig5-data1-v2.xlsx
Figure 5—figure supplement 1
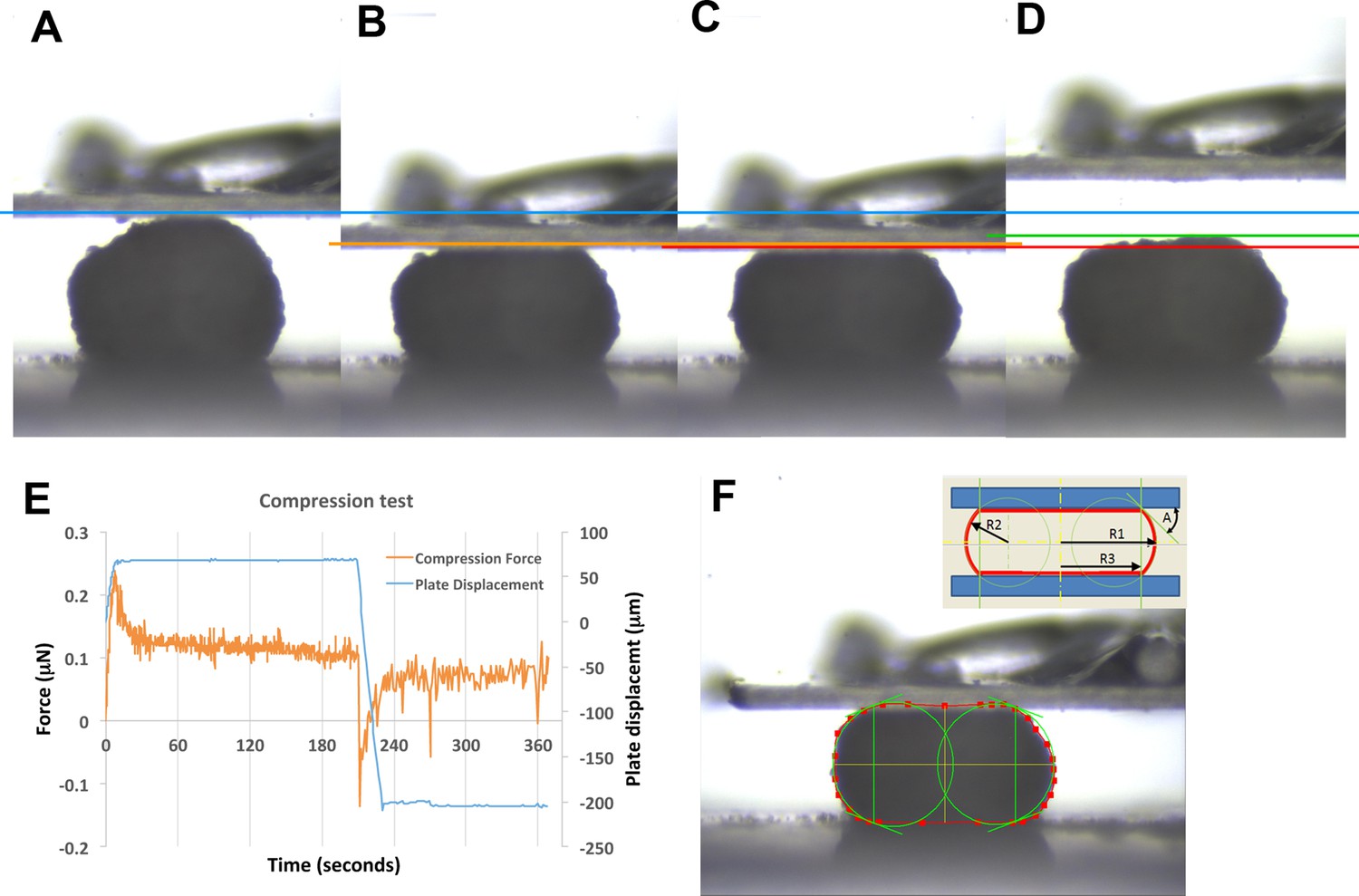
Example of parallel plate compression test.
Frames taken at 0, 10, 180, and 220 seconds (A, B, C, D, respectively). Blue line marks the top of the explant prior to compression (0 s, A). Orange line marks top of explant after initial compression is applied (10 s, B). Red line marks top of explant after 180 s of stress relaxation (C). Green line marks top of explant 10 s after plate is lifted off explant (220 s, D). Compression is applied to 20% strain over the first 10 s, then stabilized for an additional 10 s, after which active plate movement stops and the explant is allowed to undergo stress relaxation (E). The shape of the explant profile is measured at 180 s (F), giving the radii of curvature and the angle between explant and plate (F – inset; R1–R3, A); these are used to give the surface tension (see Materials and methods). Figure 5—figure supplement 1F inset is reproduced from image from CellScale MicroTester software, with permission from CellScale.
Figure 5—figure supplement 2
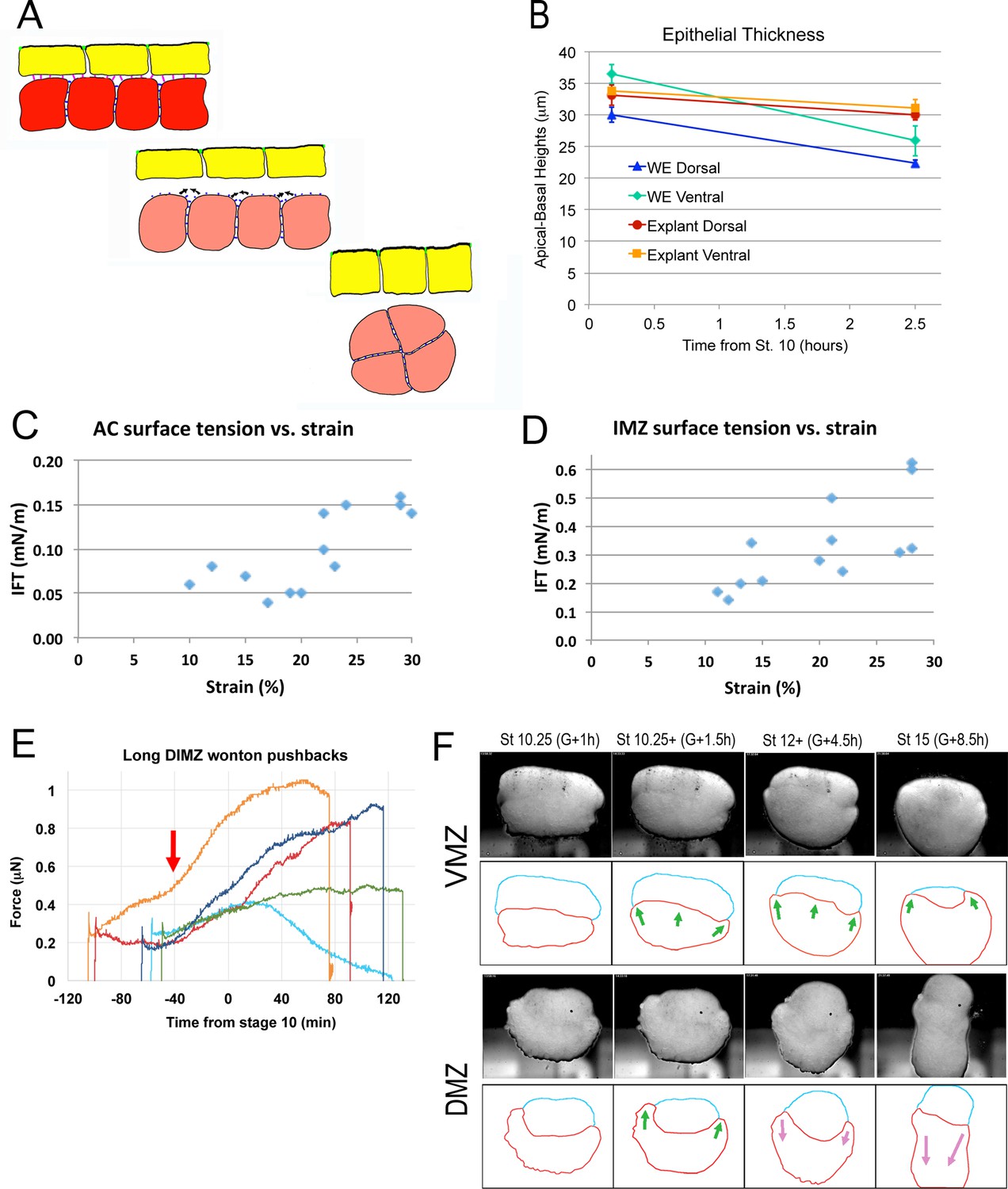
Test of epithelial thickness; test of surface tension vs. strain; test of IMZ pushback; behavior of deep IMZ plus NIMZ tissue explants.
(A) Model for the development of IFT between superficial (yellow) and deep layers (red-ish) of the IMZ; prior to Stage 10- (top), the superficial epithelium and deep tissue have high affinity for each other, so low IFT; beginning at Stage 10-, affinity decreases (middle), increasing IFT and leading the deep tissue to round up (bottom). One possibility is that the epithelium columnarizes (Bottom, yellow cells), to match the new, lower surface area of the deep tissue. (B) Thickness of the non-bottle cell epithelium of the IMZ in both whole embryos (WE) and explants declines during the first two hours of gastrulation (based on RDA-labeled explants and embryos as showing in Figure 3—figure supplement 1A,B, 2 A). The IFT of small sandwiches of both AC (C) and IMZ (D) over a range of strains. (E) DIMZ wontons held under the MicroSquisher platen, initially at 0.3 μN, develop additional compression force over three hours (e.g. Video 15*, on which the dark blue line is based). The mean time of force increase, G-41m, is indicated (red arrow). (F) Explants of about 90° of deep ventral and dorsal upper IMZ plus NIMZ tissue (VMZ and DMZ) under a cover glass (upper panels). The upper IMZ (red outline in lower panels) begins to engulf the NIMZ (blue outline) in both VMZ and DMZ explants (green arrows), but at Stage 10.5, IMZ in the DMZ explant reverses course and begins to converge and extend (magenta arrows; Video 16*, 21*; see also Video 17*).
-
Figure 5—figure supplement 2—source data 1
Figure 5—figure supplement 2C-D source data: Epithelial thickness vs. time; IMZ, AC surface tension vs. strain.
- https://cdn.elifesciences.org/articles/57642/elife-57642-fig5-figsupp2-data1-v2.xlsx
-
Figure 5—figure supplement 2—source data 2
Figure 5—figure supplement 2E source data: Long push-back force vs. time.
- https://cdn.elifesciences.org/articles/57642/elife-57642-fig5-figsupp2-data2-v2.xlsx
Figure 5—figure supplement 3
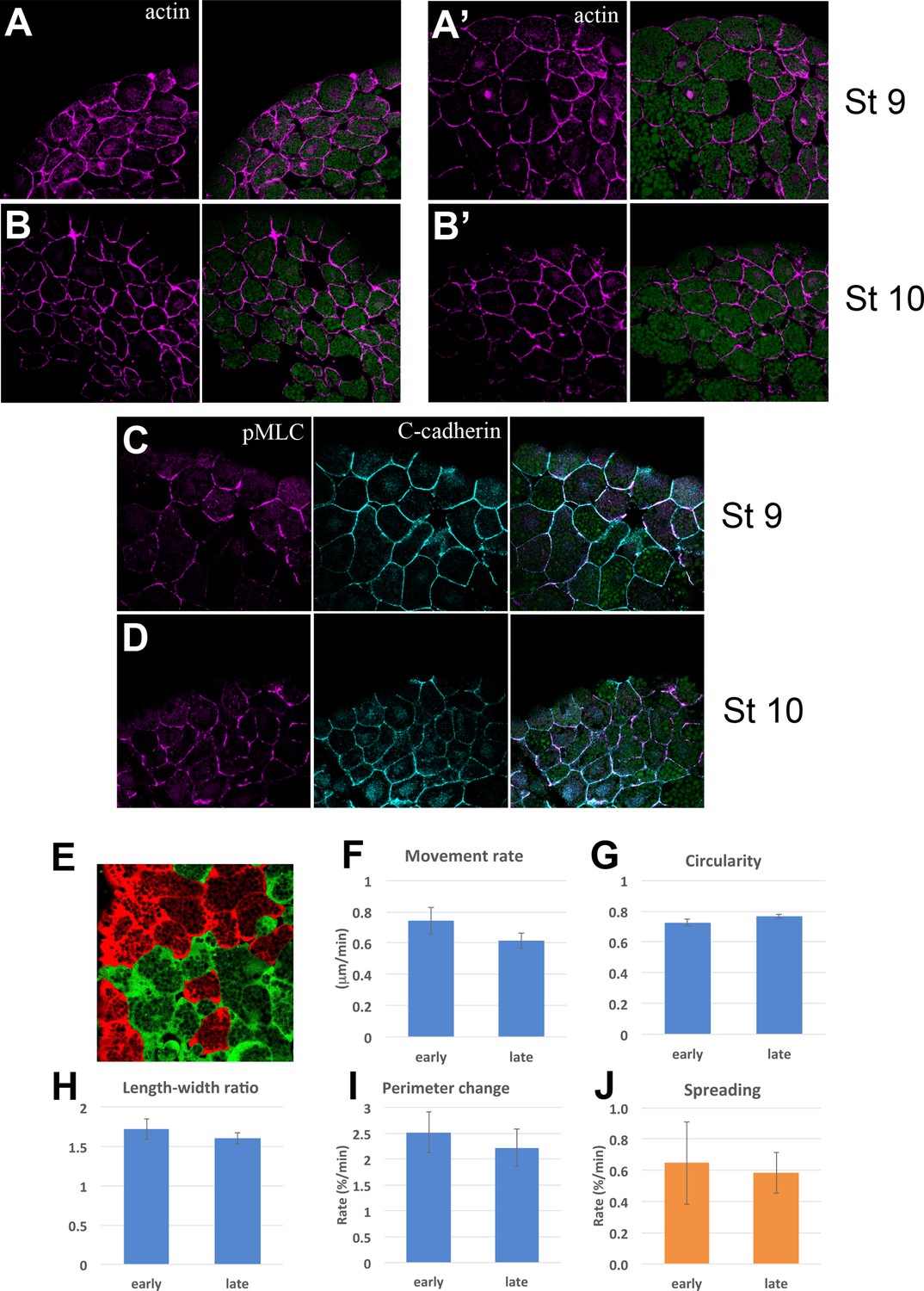
Cytoskeleton and Cell motility.
Actin (magenta) and yolk autofluorescence (green) in the upper and lower IMZ at Stages 9 (A, A’) and 10 (B, B’); vegetal is to the left. Loose cell organization is a by-product of the rapid fixation used to allow Phalloidin staining. pMLC (magenta) and C-cadherin (cyan) in the upper IMZ at Stages 9 (C) and 10 (D). Fluorescently labeled cells in deep cell explants of upper IMZ were imaged via LSCM (Video 19) and their outlines and centroids were tracked and analyzed. Mean movement rate of the centroid over 2–3 min intervals (E), mean Circularity ( = 4π*(area)/(perimeter)^2, such that 1 = a perfect circle) (F), mean Aspect ratio (length:width ratio of an ellipse fit to the cell outline) (G), and mean Perimeter Length change (H) show no significant differences between Stages 9–9.5 (‘early’) and Stage 10 (‘late’). Dissociated deep IMZ cells were plated on C-cadherin-Fc and imaged with DIC optics (Video 20) and their outline was tracked and analyzed at 5–16 min intervals. The rate of cell spreading was not significantly different at Stage 9–9.5 vs. Stage 10 (I). Error bars for E-I are standard error of the mean.
-
Figure 5—figure supplement 3—source data 1
Figure 5—figure supplement 3F-J source data: Cellular movement rate, circularity, length-width ratio, perimeter change rate and spreading rate.
- https://cdn.elifesciences.org/articles/57642/elife-57642-fig5-figsupp3-data1-v2.xlsx
Figure 6 with 2 supplements
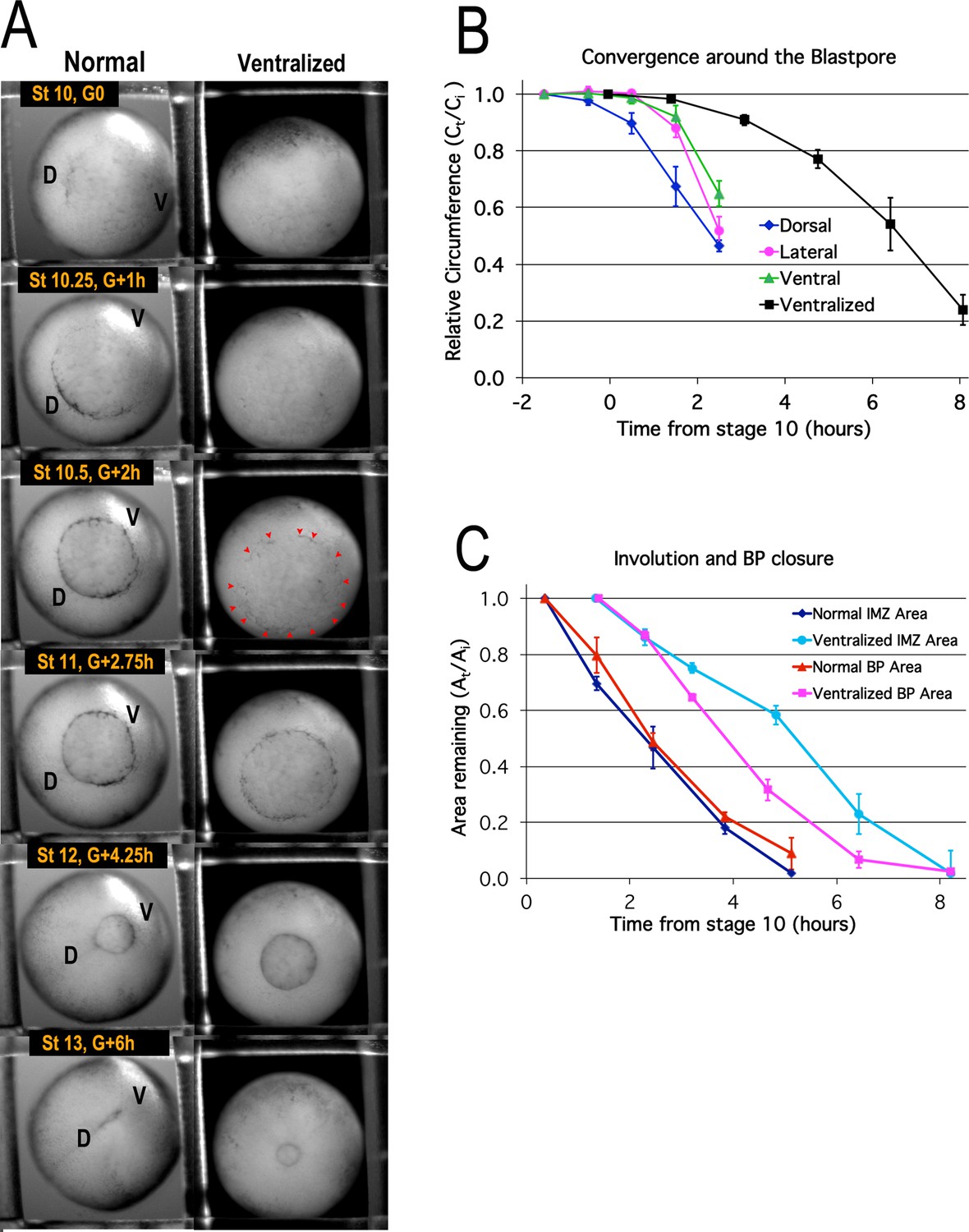
Blastopore closure.
Time lapse movie frames of normal (left) and ventralized embryos (right) are shown at the stages and times indicated (A, Video 1*). Dorsal (‘D’) and ventral (‘V’) sides of the embryo are indicated. Blastopore formation (blastopore pigment line formation) and subsequent blastopore closure (see Figure 6—figure supplement 1 for details) is delayed in the ventralized embryos, with formation of the blastopore usually occurring at control Stage 10.5 (red pointers), two hours after the onset of gastrulation at Stage 10 (G + 2 h). The blastopore of ventralized embryos closes symmetrically whereas the normal closure is biased toward the future ventral side (compare Stages 10.5 vs 12). Convergence (B) in the dorsal, lateral and ventral quadrants of the pre-involution IMZ of normal embryos (n = 3 or 4 embryos per time point; error bars = SEM within region, across embryos) and in the entire IMZ of ventralized embryos (n = 5 embryos per time point; error bars = SEM across embryos). Change in superficial area compared to its original size (At/Ai) of the pre-involution IMZ, a measure of its involution, and the area of the exposed vegetal endoderm, a measure of blastopore closure (C) (n = 3 normal embryos and 3 ventralized embryos; error bars = SEM across embryos). For both B and C, the same embryos were measured at sequential time points.
-
Figure 6—source data 1
Figure 6B,C source data: Relative blastopore circumference vs. time, relative blastopore and IMZ area vs. time.
- https://cdn.elifesciences.org/articles/57642/elife-57642-fig6-data1-v2.xlsx
Figure 6—figure supplement 1
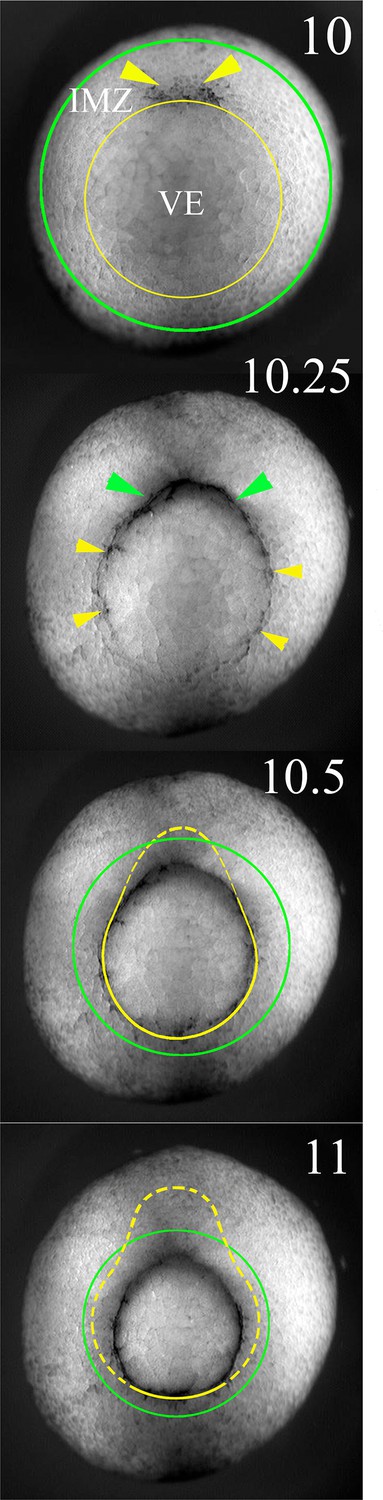
The lower (vegetal) and upper (animal) limits of the IMZ are indicated by yellow and green lines superimposed on to vegetal views of a Xenopus gastrula stage embryo at the indicated Nieuwkoop and Faber stages (Nieuwkoop and Faber, 1967).
Formation of the blastoporal pigment line, which defines the vegetal end of the IMZ, and initially defines the blastopore, is indicated by yellow pointers, and formation of a depression, the initiation of involution (and an invagination) of the IMZ is indicated by green pointers. Note that the vegetal boundary of the IMZ is carried inside (dashed yellow lines) and covered over by the blastoporal lips from dorsal to ventral as gastrulation proceeds; as this occurs, the lips define the blastopore.
Figure 6—figure supplement 2
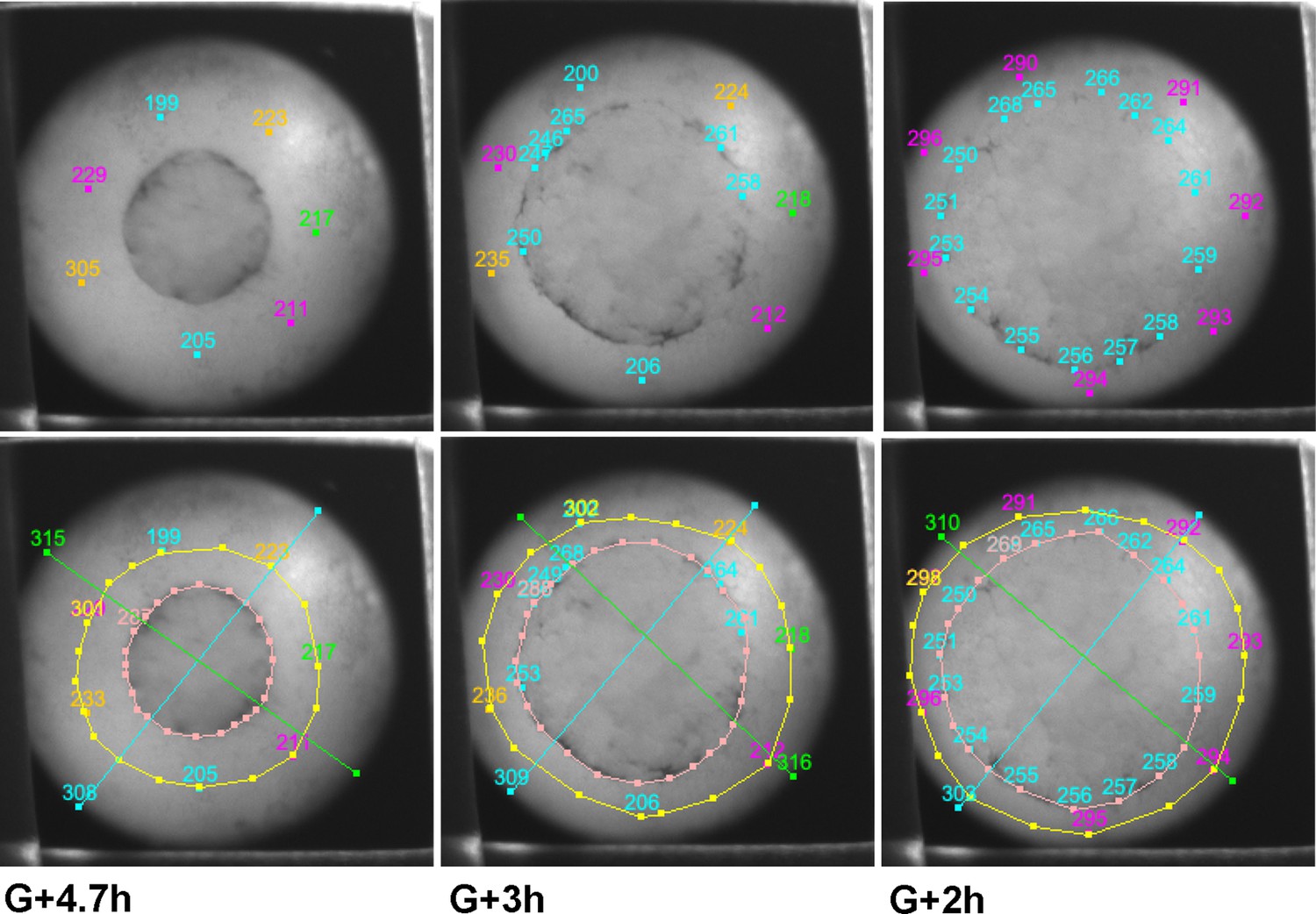
Examples of tracking of IMZ and blastopore area through the course of a time lapse movie using fiduciary points.
Points initially laying at the blastopore lip at late time points (~G + 8 h) are tracked backward to early gastrula stages (top row, outer points). Once bottle cells apices are revealed, the vegetal edge of the bottle field is also tracked backwards (top row, inner circle of blue points). At each time point, the blastoporal area (or prospective blastoporal area, at earlier stages) is determined (area inside pink line), as is the area inside the upper boundary of the IMZ (yellow line – a bit below the actual limit of involution). Areas are determined based on the spherical area of the embryo occupied by each region (see Materials and methods). IMZ area is determined as the area inside the upper IMZ boundary minus the blastoporal area ( = area inside the lower IMZ boundary). Areas are normalized to starting area for each embryo. The embryo shown is a ventralized embryo.
Figure 7 with 1 supplement
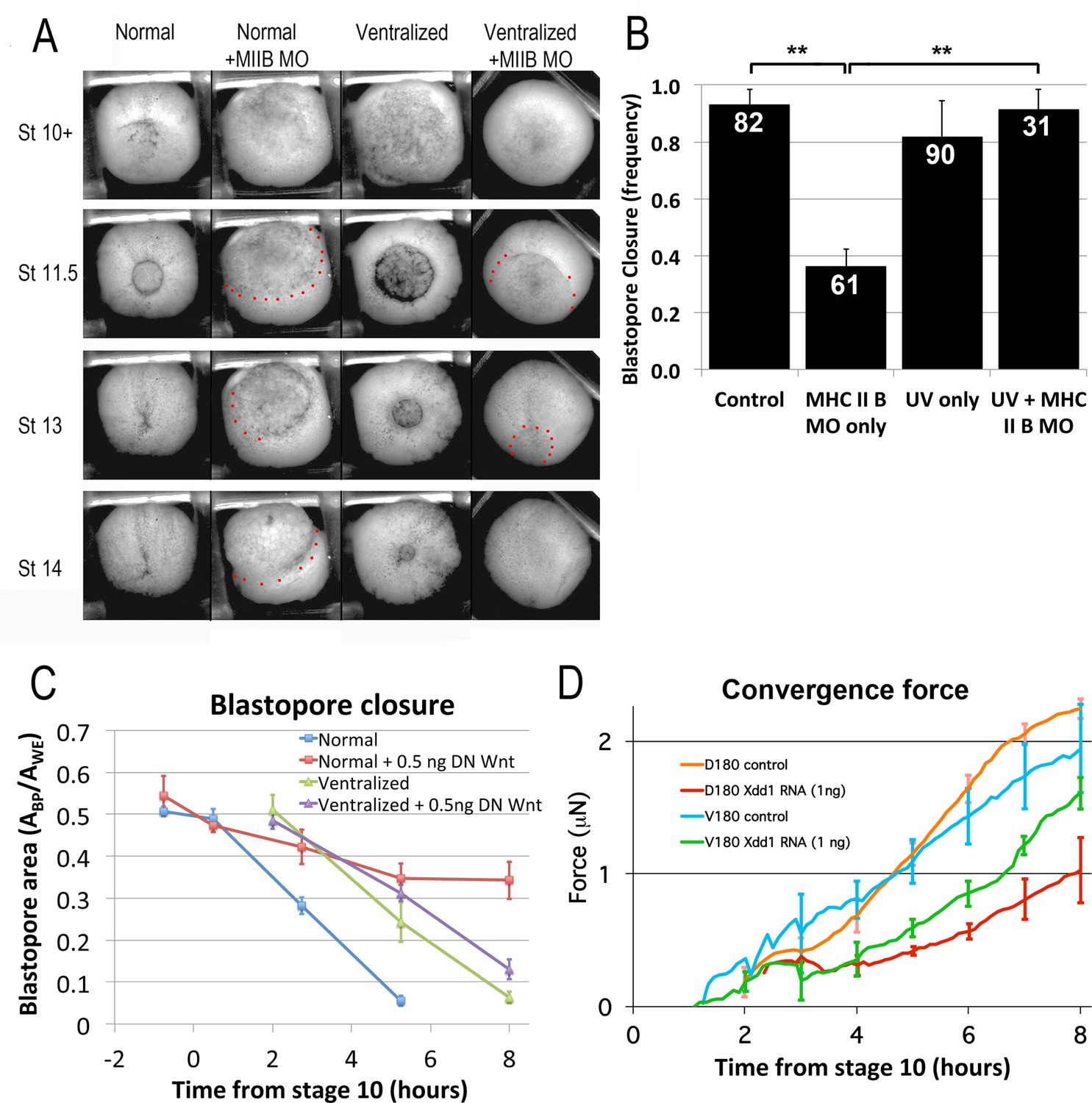
CT and CE depend on different molecular pathways.
(A) Representative stills of time lapse movies (Video 22*) comparing blastopore closure in normal and ventralized embryos, un-injected or injected with an MHC IIB morpholino. The region just outside the blastopore or forming blastopore are indicated by red dots in cases where it is difficult to see. (B) Frequency of blastopore closure by Stage 20 (G + 13 h effective success of blastopore closure) was scored for embryos from 4 different clutches of embryos (n’s indicated on chart); ** = p < 0.01. Comparison of the extent of blastopore closure over time in normal and ventralized embryos, either un-injected or injected with 0.5 ng dnWnt11 RNA (C; see for example Figure 7—figure supplement 1 and Video 23*), based on the projected area of the blastopore, Abp, divided by the projected area of the whole embryo, Awe, measured from time lapse movies (n = 4 embryos for each treatment). Force measurements (Shook et al., 2018) of D180 or V180 explants made from embryos, either un-injected or injected with 1 ng Xdd1 RNA (D; n’s = 3–4 explants per treatment). Errors bars in all cases = SEM.
-
Figure 7—source data 1
Figure 7B source data: frequency of successul blatopore closure.
- https://cdn.elifesciences.org/articles/57642/elife-57642-fig7-data1-v2.xlsx
-
Figure 7—source data 2
Figure 7C source data - extent of blastopore closure vs. time.
- https://cdn.elifesciences.org/articles/57642/elife-57642-fig7-data2-v2.xlsx
-
Figure 7—source data 3
Figure 7D source data: Convergent force of D180 and V180 explants.
- https://cdn.elifesciences.org/articles/57642/elife-57642-fig7-data3-v2.xls
Figure 7—figure supplement 1
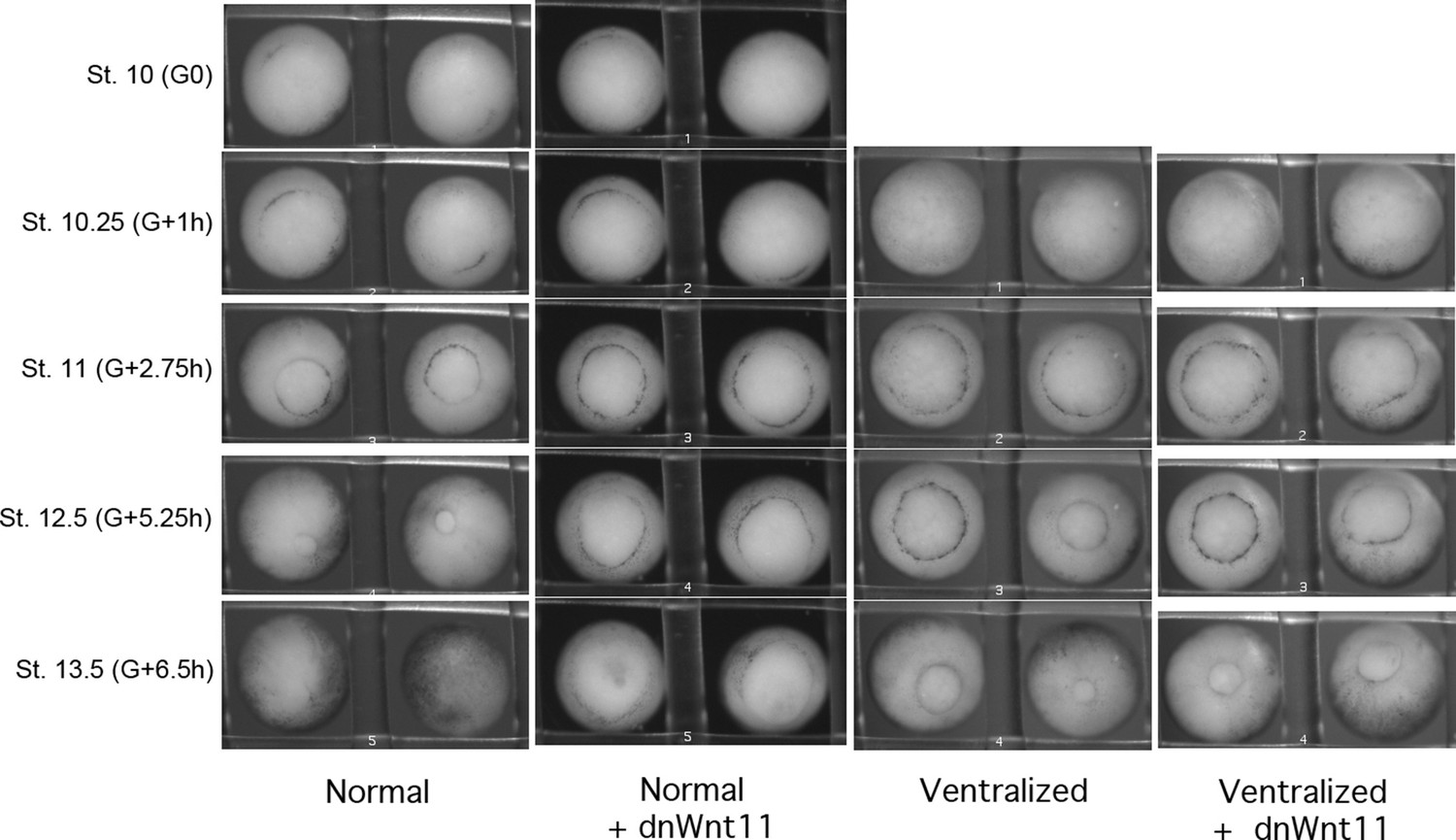
Time lapse movie stills of normal and ventralized embryos, +/- 0.5 ng dnWnt11 mRNA.
dnWnt11 causes a delay in blastopore closure in normal, but not in ventralized embryos.
Figure 8
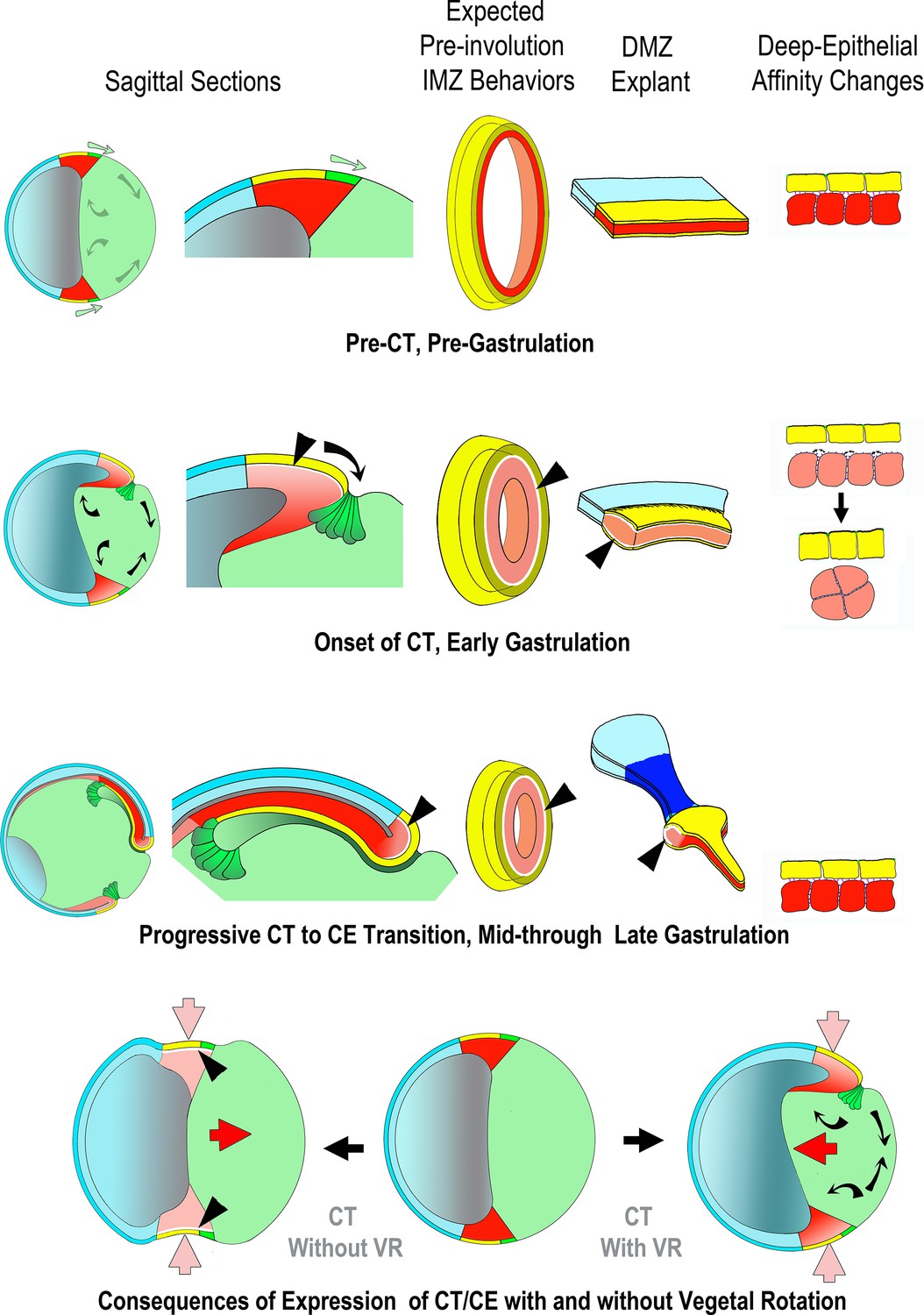
Proposed mechanism of CT, its function in the gastrula and in explants, its relationship to CE and its integration with the other movements of gastrulation.
Yellow/green = endoderm, pink/red = mesoderm, blue = ectoderm (See Figure 1 for details). Prior to the onset of gastrulation (top row), vegetal rotation (VR) (Sagittal Sections, gray arrows) has not yet occurred and the IMZ annulus (3rd column) remains stable, as does the IMZ explant (4th column), due to a stable deep-epithelial cell affinity (far right). With the onset of gastrulation and CT (2nd row), apical constriction of the lower IMZ epithelium (bottle cell formation – Sagittal Sections, dark green), VR, involution and invagination begin (Sagittal Sections, black arrows). Co-incident with bottle cell formation, the deep mesodermal tissue begins to lose its affinity with the overlying epithelium. During early gastrulation both tissue surface tension of the IMZ as a whole and its interfacial tension with an epithelium increase, resulting in a tendency for the IMZ to minimize its surface area by converging and thickening (2nd row, far right). This loss of affinity at the interface of deep and epithelial tissue (2nd and 3rd rows, shown by white spaces between epithelial and deep, at black arrowhead pointers) increases the IFT between deep and superficial layers, resulting in both convergence and thickening of the IMZ of the explants (CT, second row, bulge in DMZ explant). Increased IFT would have a tendency to thicken and decrease the circumference of the IMZ annulus in embryos (3rd column, 2nd row) were it not resisted by adjacent tissue, so instead generates a pre-involution tensile force around the blastopore and converges, but does not thicken (not shown). In explants, the weak affinity of the IMZ epithelium to the underlying deep region results in its retraction, initially from the leading edge mesoderm (not shown), whereas in the intact embryo, the IMZ epithelium is tightly attached at its vegetal edge to the forming bottle cells, which in turn are tightly attached to the adjacent vegetal endodermal tissues. Thus the IMZ epithelium is pulled along with the bottle cells as they move inside the blastopore (1st, 2nd column). During early involution, as the lower edge of the IMZ involutes, it transitions from expressing CT to expressing directed migration (2nd column, 2nd row), and during later involution, the upper IMZ transitions to expressing CE (2nd column, 3rd row). In the case of CE, this transition is accompanied by regaining a high deep-epithelial affinity (loss of white space, on involution, 2nd column, 2nd and 3rd rows). In explants, the later transition from CT to CE first occurs in tissue laying dorsally at the vegetal edge of the upper IMZ, and continues in an anterior to posterior progression (Shih and Keller, 1992a). The snout of tissue extending from the thickened collar of mesoderm in the explant reflects the onset of CE and is coincident with the progressive return of the affinity between the deep cells and the bottle cell epithelium (3rd row, 4th column). In the meantime, VR (black arrows, 1st column, 2nd row) has moved the vegetal endoderm inward at the onset of gastrulation such that convergence force generated by CT cooperates with VR to move the IMZ vegetally and push the vegetal endoderm inside (compare to 1st column, 1st row). VR, CT and CE also cooperate to stretch the ectodermal (NIMZ-AC) region by pulling it vegetally (1st column, 1st to 3rd rows). When CE begins at Stage 10.5, it has been positioned by the actions of CT and VR such that it further moves the IMZ vegetally and pushes the vegetal endoderm inside (4th row, right, red arrow), in contrast to converging in its initially more equatorial position, where it would tend to push the vegetal endoderm out, rather than in (4th row, left, red arrow).
Videos
Video 1
Blastopore closure in normal vs. ventralized embryos.
Top two embryos are normal, bottom two were UV ventralized. The two on the left are those shown in Figure 6A. Stage 10–20 (G + 0.25 h to G + 12.75 h); most of neurulation is occurring on the opposite side of the normal embryos. 3 min/frame. Scale bar = 1 mm.
Video 2
Normal giants with and without bottle cells.
Giant with bottle cells on left, without on right. Movies begins about Stage 10.25 (G + 30 m), just after release from cover glass. 3 min per frame. Total Elapsed time = 10:15. Time stamp on right movie is fast by 1:02 (h:mm). Scale bar = 1 mm.
Video 3
Early UV ventralized giant.
Movie begins at control Stage 9.5 (G-1h). 3 min per frame. Total elapsed time = 14:52. Scale bar = 500 μm.
Video 4
Ventral giant sandwich.
Made from normal embryos, cut through the dorsal midline. Only a few bottle cells included. NIMZ and IMZ regions converge equally through frame 10 (G-1h), after which IMZ converges more rapidly. Begins Stage 9+ (G-1.5h) through early neurulation (G + 7.45). 3 min per frame, 180 frames, 15 fps. Scale bar = 500 μm.
Video 5
Lithium dorsoanteriorized giant.
Movie begins at control Stage 10+ (G + 0.5 h). 3 min per frame. Elapsed time to penultimate frame = 15 h; final frame is at 17 hr. Scale bar = 1 mm.
Video 6
DMZ 120° sandwich explant showing bottle cell retraction and spreading.
Note also adjacent epithelial are pulled toward forming bottle cells. Starts Stage 9+ (G-1.75h) through early neurulation (G + 7.75 h). 3 min/frame. Scale bar = 500 μm.
Video 7
DMZ 180° sandwich explant from Stage 9.5–16 (G-1h to G + 9 h). 3 min/frame, scale bar = 500 μm.
Video 8
Ventral 180° sandwich explant.
Movie begins at control Stage 9.5 (G-0.9h). 3 min per frame. Elapsed time = 8 hr. Scale bar = 500 μm.
Video 9
Double NIMZ and double IMZ sandwich explants.
Movie begins at G + 1 h. 3 min/frame. Elapsed time 8.9 hr. Scale bar = 500 μm.
Video 10
Double ventral IMZ and NIMZ explants viewed from the side, facing lateral ends.
Movie begins at about G + 3 h. 5 min/frame. Elapsed time = 6 hr. Scale bar = 500 μm.
Video 11
Epithelial spreading/contraction assay.
Movie from which Figure 4B was made. Animal cap epithelium, recombined with deep MZ, ventral on the left, dorsal on the right, on FN coated substrate; vegetal end down. Stage 10–12 (G-0.2h to G + 4.6 h). Cover glass removed between frame 11 and 12. 3 min/frame. Scale bar = 500 μm.
Video 12
Epithelial retreat and respreading.
Movie from which Figure 4E was made. Animal cap epithelium, recombined with ventral deep MZ. Under cover glass the entire time. Note that deep IMZ cells spread animally over the deep NIMZ tissue between frames 1 and 30 (first 20% of the movie), most obviously at the lateral edges; in response the more vegetal superficial cells over the IMZ initially contract; during the next 20% of the movie, the superficial cells move animally and begin to respread over the NIMZ. Stage 10.25–15 (G + 1 h to G + 8.5 h). 3 min/frame. Scale bar = 100 μm.
Video 13
Wontons rounding up, viewed from the side, parallel to their long axis; out of focus top-down view below.
Wontons contain DMZ deep tissue on the left, AC deep tissue on the right. Movie begins at roughly G-1h. 3 min/frame. Elapsed time = 2 hr. Scale bar = 500 μm.
Video 14
Aggregates of deep VIMZ (left) and deep animal cap (right) cells rounding up, just after release from cover glass.
Movie begins about G + 1 h. 3 min per frame. Elapsed time = 2 hr. Scale bar = 500 μm.
Video 15
Example of a long duration pushback experiment.
0.3 μN compression force was applied to a DIMZ wonton (see Materials and methods) over 50 s, after which the plate was left in place to respond passively in order to measure the response of the explant over the next 3 hr. Capture rate = 1 s/frame for first 30 frames, then 100 s/frame for 110 frames, then 1 frame/s for 140 s. Playback = 15 fps. Scale bar = 250 μm.
Video 16
Naked ventral IMZ showing engulfment.
Stages 10.25 through 14 (G + 1h to 7.25h). IMZ cells engulf NIMZ; no reversal at Stage 10.5 (15:00). Under cover glass; imaged at 20 X on an Olympus IX70. 1.5 min per frame, 250 frames, 15 fps. Scale bar = 200 μm.
Video 17
High-resolution movie of deep ventral NIMZ (top) engulfment by deep ventral IMZ (bottom).
Stages 10.25 through 14 (G + 1h to 7.25h). Vegetal toward the bottom. Smaller, more animal IMZ cells show motility over NIMZ region. A small patch of epithelium (cells much less motile) appears on the right side about 1/3 of the way through the movie, moves with the NIMZ and is eventually covered by IMZ cells. Imaged surface is covered by a thin sheet of agarose, under cover glass. Made with a 20 X dipping lens on an Olympus AX70. 1.5 min per frame, 250 frames, 15 fps. Scale bar = 100 μm.
Video 18
Explant of deep upper DIMZ, imaged with low-angle illumination.
The LR axis is oriented up and down in the movie. Left image is around Stage 9.5 (G-1.5h to G-0.9h), right image is around Stage 10 (G-0.4h to G + 0.2 h). 1 min/frame. Scale Bar = 200 μm.
Video 19
Explant of deep IMZ from an embryo injected at 2 cells stage with Alexa-555 dextran in one cell and Alexa-488 dextran in the other, imaged at 60 X via LSCM at a depth of 3–5 μm.
Movie on the left is around Stage 9.5(G-64m to –49 m), the one on the right around Stage 10 (G-2m to +15 m). 30 s/frame. Scale bar = 20 μm.
Video 20
Dissociated cells transferred onto C-cadherin-Fc substrate, imaged using DIC optics on a Zeiss Axio Observer.
G-1.2h to G + 0.8 h. 20–30 s/frame, 6 fps playback. Scale bar = 20 μm.
Video 21
Naked dorsal IMZ showing engulfment and reversal.
Stages 10.25 through 14 (G + 1h to 7.25h). IMZ cells begin to engulf NIMZ, then, at Stage 10.5 (15:00, frame 43), stop and reverse course, eventually converging and extending. Under cover glass; imaged at 20 X on an Olympus IX70. 1.5 min per frame, 250 frames, 15 fps. Scale bar = 200 μm.
Video 22
Blastopore closure in normal and ventralized embryos, either uninjected or injected to 10 μMolar MHC II B MO.
From left to right: Normal; Ventralized; MHCIIB MO injected; Ventralized and MHCIIB MO injected. Moive starts at about Stage 10.5 and continues through neurulation. Scale bar = 1 mm.
Video 23
Blastopore closure in embryos untreated, injected with 0.5 ng dnWnt, ventralized or ventralized +injected with 0.5 ng dnWnt (left to right).
Movie starts at Stage 10- (G-0.7h) and continues through late neurulation (~Stage 18). 5 min per frame. Scale bar = 1 mm.
Video 24
Dorsal 180° sandwich explant.
Movie begins at G + 2.2 h. 3 min per frame. Elapsed time = 13.4 h. Scale bar = 1 mm.
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Antibody | pMLC 2 (Ser19)(Rabbit monoclonal) | Cell Signaling | Cat. #3,671RRID:AB_330248 | (1:100) |
| Antibody | 6B6(recognizes C-cadherin)(mouse monoclonal) | Developmental Studies Hybridoma Bank | AB_528113RRID:AB_528113 | (1:300) |
| Other | phalloidin-Alexa 555 | (Acti-stain, Cytoskelton.com) | 8,953 | Derivatized fluorescent dextran (1:100 at 4 C, 12–36 hours) |
| Sequence-based reagent | MHC IIB morpholino | Skoglund et al., 2008 | Morpholino5’CTTCCTGCCCTGGTCTCTGTGACAT3’ | |
| Sequence-based reagent | Xdd1 | Sokol, 1996 | RNA | |
| Sequence-based reagent | dnWnt11 | Tada and Smith, 2000 | RNA | |
| Software, algorithm | NIH Image 1.6 | Wayne Rasband, National Institutes of Health; available at http://rsb.info.nih.gov/nih-image/ | RRID:SCR_003073 | |
| Software, algorithm | Object Image | Norbert Vischer, University of Amsterdam; available at https://sils.fnwi.uva.nl/bcb/Object-Image/object-image.html | RRID:SCR_015720 | |
| Software, algorithm | Image J | http://rsb.info.nih.gov/ij/ | RRID:SCR_003070 | |
| Software, algorithm | Metamorph | https://www.moleculardevices.com/products/cellular-imaging-systems/acquisition-and-analysis-software/metamorph-microscopy#gref | RRID:SCR_002368 | |
| Software, algorithm | SquisherJoy | (CellScale, Waterloo, Canada) | Software for MicroSquisher; RRID:SCR_022034 | |
| Software, algorithm | Axisymmetric Drop Shape Analysis | del Rı́o and Neumann, 1997 | ADSA in MatlabRRID:SCR_001622 for Matlab | |
| Software, algorithm | curvature outline detection | https://www.mathworks.com/products/matlab.html | (Canny) in MATLABRRID:SCR_001622 for Matlab | |
| Software, algorithm | Surface tension determination in SquisherJoy | Based on Brodland et al., 2009; Mgharbel et al., 2009 |
Additional files
-
Transparent reporting form
- https://cdn.elifesciences.org/articles/57642/elife-57642-transrepform1-v2.docx
-
Source data 1
Convergence rates of NIMZ and IMZ in explants.
- https://cdn.elifesciences.org/articles/57642/elife-57642-data1-v2.xlsx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Characterization of convergent thickening, a major convergence force producing morphogenic movement in amphibians
eLife 11:e57642.
https://doi.org/10.7554/eLife.57642




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}