EXOC1 plays an integral role in spermatogonia pseudopod elongation and spermatocyte stable syncytium formation in mice
- Master’s Program in Medical Sciences, Graduate School of Comprehensive Human Sciences, University of Tsukuba, Japan
- Laboratory Animal Resource Center, Trans-border Medical Research Center, University of Tsukuba, Japan
- School of Medical Sciences, University of Tsukuba, Japan
- Ph.D Program in Human Biology, School of Integrative and Global Majors, University of Tsukuba, Japan
- Division of Germ Cell Biology, National Institute for Basic Biology, National Institutes of Natural Sciences, Japan
- Department of Basic Biology, School of Life Science, Graduate University for Advanced Studies (Sokendai), Japan
- Department of Comparative and Experimental Medicine, Nagoya City University Graduate School of Medical Sciences, Japan
- Doctoral program in Biomedical Sciences, Graduate School of Comprehensive Human Sciences, University of Tsukuba, Japan
- Doctoral program in Medical Sciences, Graduate School of Comprehensive Human Sciences, University of Tsukuba, Japan
- Department of Stem Cells and Human Disease Models, Research Center for Animal Life Science, Shiga University of Medical Science, Japan
Figures
Figure 1 with 3 supplements
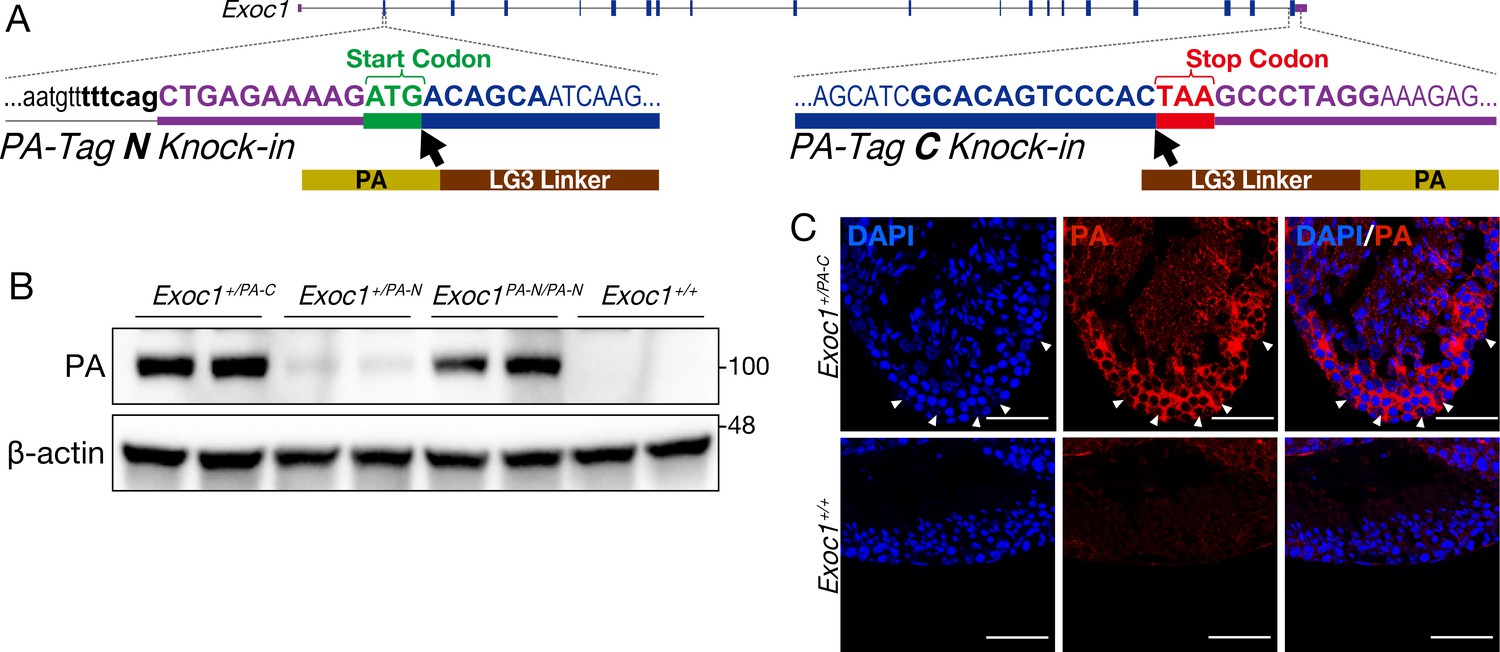
Confirmation of EXOC1 expression in testes using the PA-Tag knock-in mouse.
(A) Generation of PA-Tag knock-in mice. In the Exoc1PA-C allele, the LG3-linker connected PA-tag gene fragment was knocked-in just before the stop codon of Exoc1 using CRISPR-Cas9. In the Exoc1PA-N allele, the LG3-connected PA-tag gene fragment was knocked in just after the start codon of Exoc1 using CRISPR-Cas12a. Bold letters represent the CRISPR-Cas9 and Cas12a target sequence. (B) Western blotting of PA-Tag antibody demonstrated that each C- and N-terminal PA-tagged EXOC1 protein was expressed in the adult testes (n = 2 in each genotype). (C) Immunofluorescence with PA-Tag antibody. EXOC1 is observed in every cell in the adult testes. The arrowheads indicate Sertoli cells in which the nucleus is eurochromatin with a large nucleolus. Scale bars: 50 μm.
-
Figure 1—source data 1
Raw data of the in vivo western blot.
- https://cdn.elifesciences.org/articles/59759/elife-59759-fig1-data1-v1.pptx
Figure 1—figure supplement 1
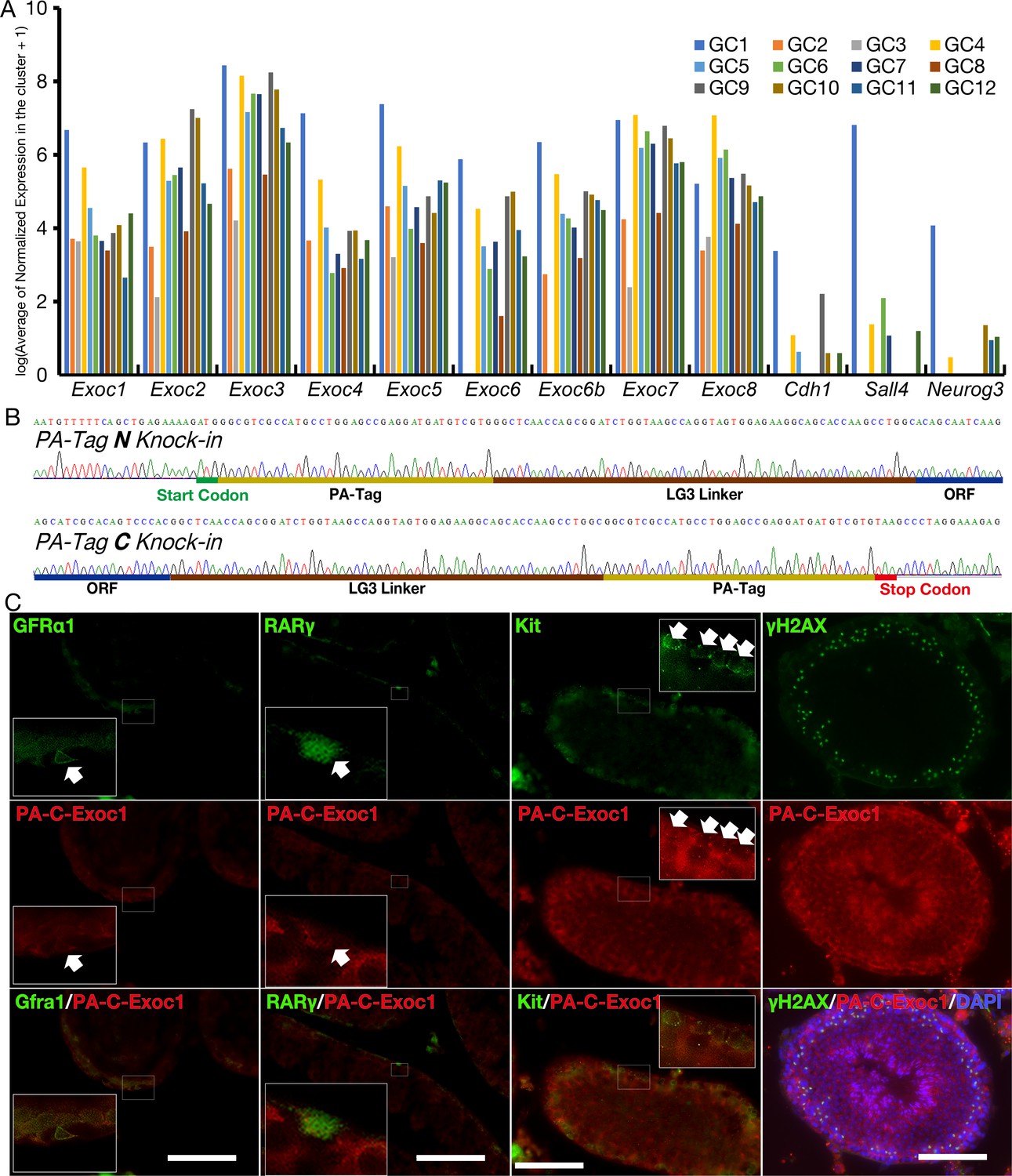
Expression of mRNA for each Exocyst subunit and EXOC1 protein in adult mouse male germ cells.
(A) The expression data of each Exocyst subunit were extracted from the open data source, GSE112393 (Green et al., 2018). GC1 is spermatogonia, GC2 and GC3 are preleptotene stages, GC4–GC8 are meiotic spermatocytes, GC9–GC11 are post-meiotic haploid round spermatids, and GC12 is elongating spermatids. All exocyst subunits are expressed in all differentiation stages. Cdh1, Sall4, and Neruog3 are known markers of spermatogonia and were used as positive controls. (B) Sanger sequencing of PA-Tag and LG3 Linker in Exoc1PA-N and Exoc1PA-C alleles. Intended Knock-in sequence was found in each allele. (C) Immunofluorescence of Exoc1+/PA-C adult testis using anti-PA-tag antibody. PA-tagged EXOC1 was observed in the adult male germ cells of interest in this study: undifferentiated spermatogonia (GFRa1+, Rarγ+), differentiating spermatogonia (Kit+), and spermatocyte (γH2AX+). Scale bars: 100 μm.
Figure 1—figure supplement 2
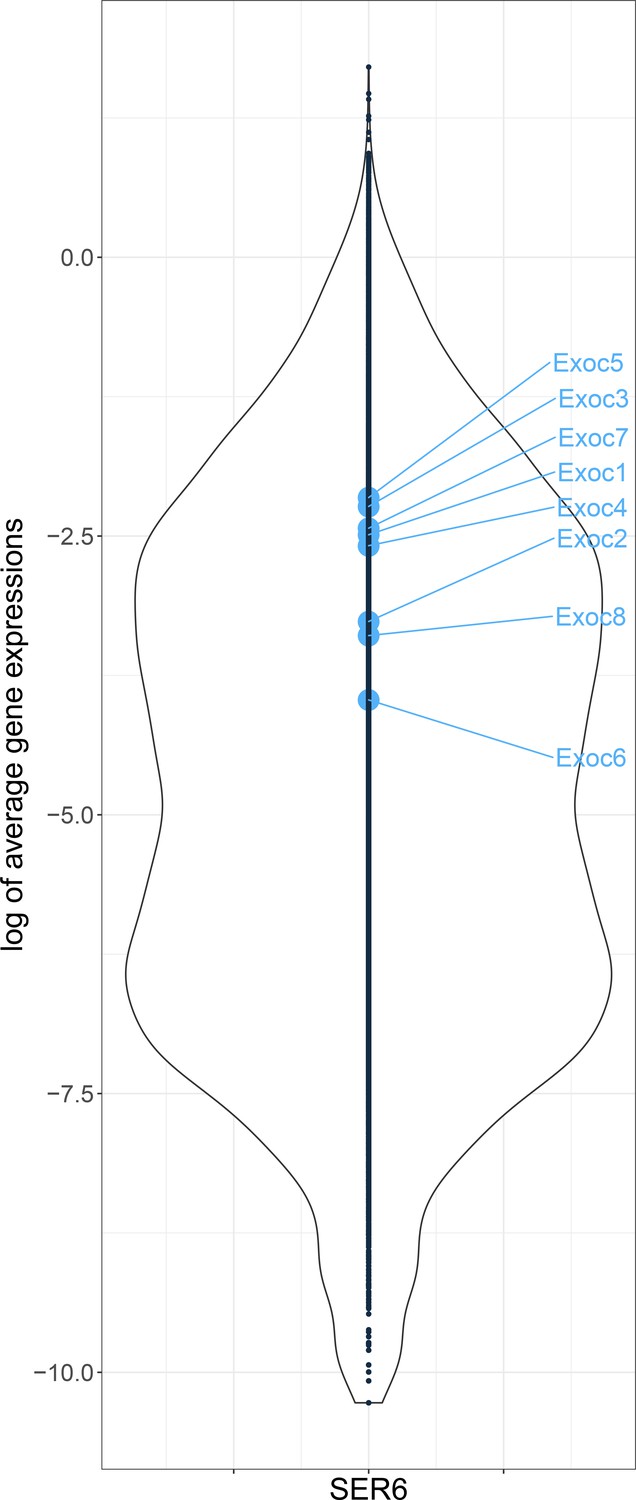
Expression of Exocyst subunits in Sertoli cells of adult mice.
The expression data of each Exocyst subunit in Sertoli cells (from 9 weeks mouse) was extracted from the open data source, GSM3069461 (Green et al., 2018). The y-axis is the expression level of all genes, and the x-axis is the number of genes at that expression level. All Exocyst subunits might be expressed slightly higher than average.
Figure 1—figure supplement 3
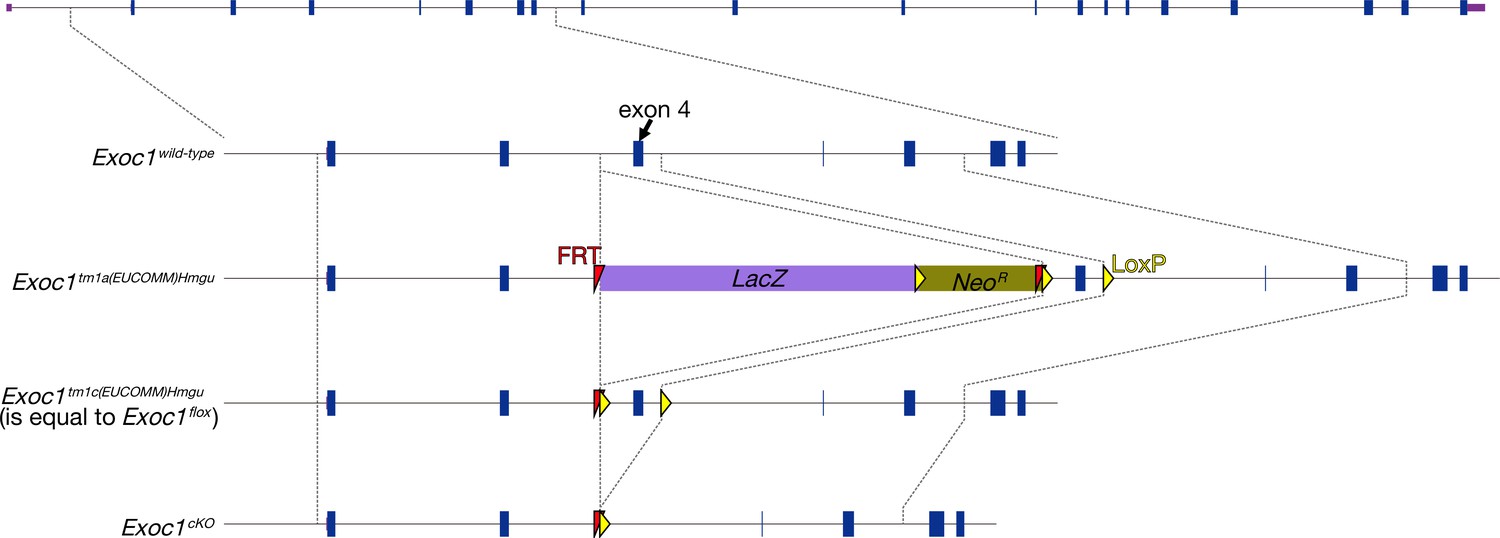
The production of Exoc1 flox mice.
The Exoc1 flox strain was from the Exoc1tm1a(EUCOMM)Hmgu mouse. The Exoc1tm1a(EUCOMM)Hmgu allele was changed to the Exoc1tm1c(EUCOMM)Hmgu (is equal to Exoc1flox) allele by mating with the Flpe expression mouse (B6;SJL-Tg(ACTFLPe)9205Dym/J) (Muranishi et al., 2011). Exon 4 of Exoc1 was floxed in this allele. Arrows indicate primers for detecting the flox allele.
Figure 2 with 2 supplements
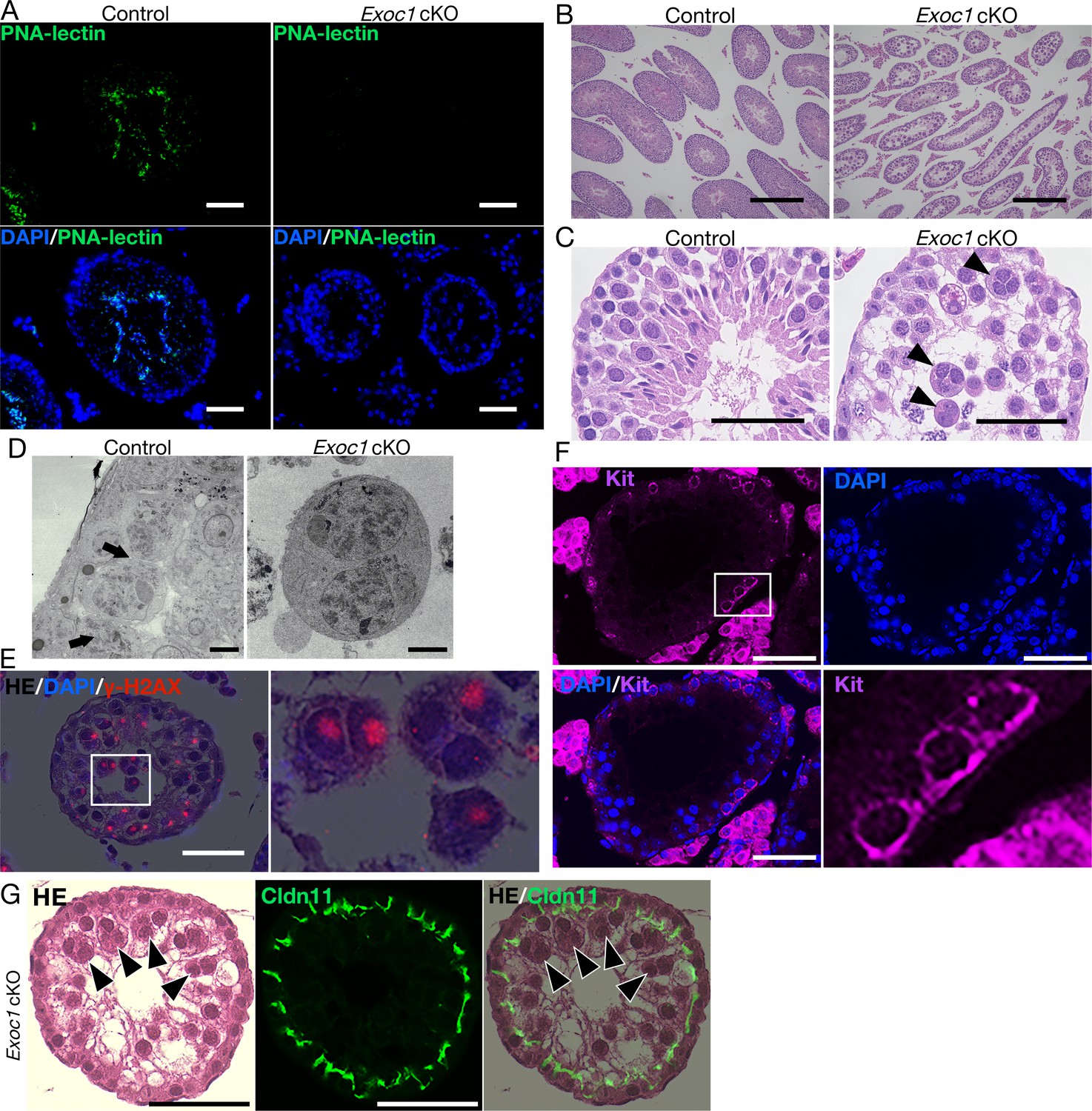
Impaired spermatogenesis in Exoc1 cKO mice.
(A) PNA-lectin staining of the Exoc1 adult cKO testis. Signals of PNA-lectin, an acrosomal marker was not observed in the Exoc1 cKO testis. Scale bars: 50 μm. (B) The macroscopic images for H and E staining. Normal spermatogenesis was not observed in almost all of the seminiferous tubules in the Exoc1 cKO adult testis. Scale bars: 300 μm. (C) The mesoscopic images for H and E staining of the Exoc1 cKO testis. Large and circular cells, appearing to be aggregates of syncytia (AGS), containing multiple nuclei were observed in the lumen of seminiferous tubules (arrowheads). Scale bars: 100 μm. Control: Exoc1flox/wt:: Nanos3+/Cre adult mice. (D) SEM observation of the Exoc1 cKO adult testis. Intercellular bridges (ICBs) (arrows) were found in the syncytia in the control (Exoc1flox/wt:: Nanos3+/Cre) testis. There were no ICB observed in the AGS of Exoc1 cKO. Scale bars: 5 μm. (E) The serial section overlay image of Exoc1 cKO adult testis. There were γ-H2AX (marker of spermatocyte) signals in the AGS nucleus. Scale bars: 50 μm. (F) A representative immunofluorescence image of an Exoc1 cKO seminiferous tubule. Kit+ syncytia, which are observed in differentiating spermatogonia, have ICB. Scale bars: 50 μm. (G) H and E staining and immunofluorescence with CLDN11. CLDN11-positive Sertoli cell tight junction (SCTJ) divides the space between the basal and the luminal compartment, and AGS are present within the luminal compartment (arrowheads). Scale bars: 50 μm.
Figure 2—figure supplement 1
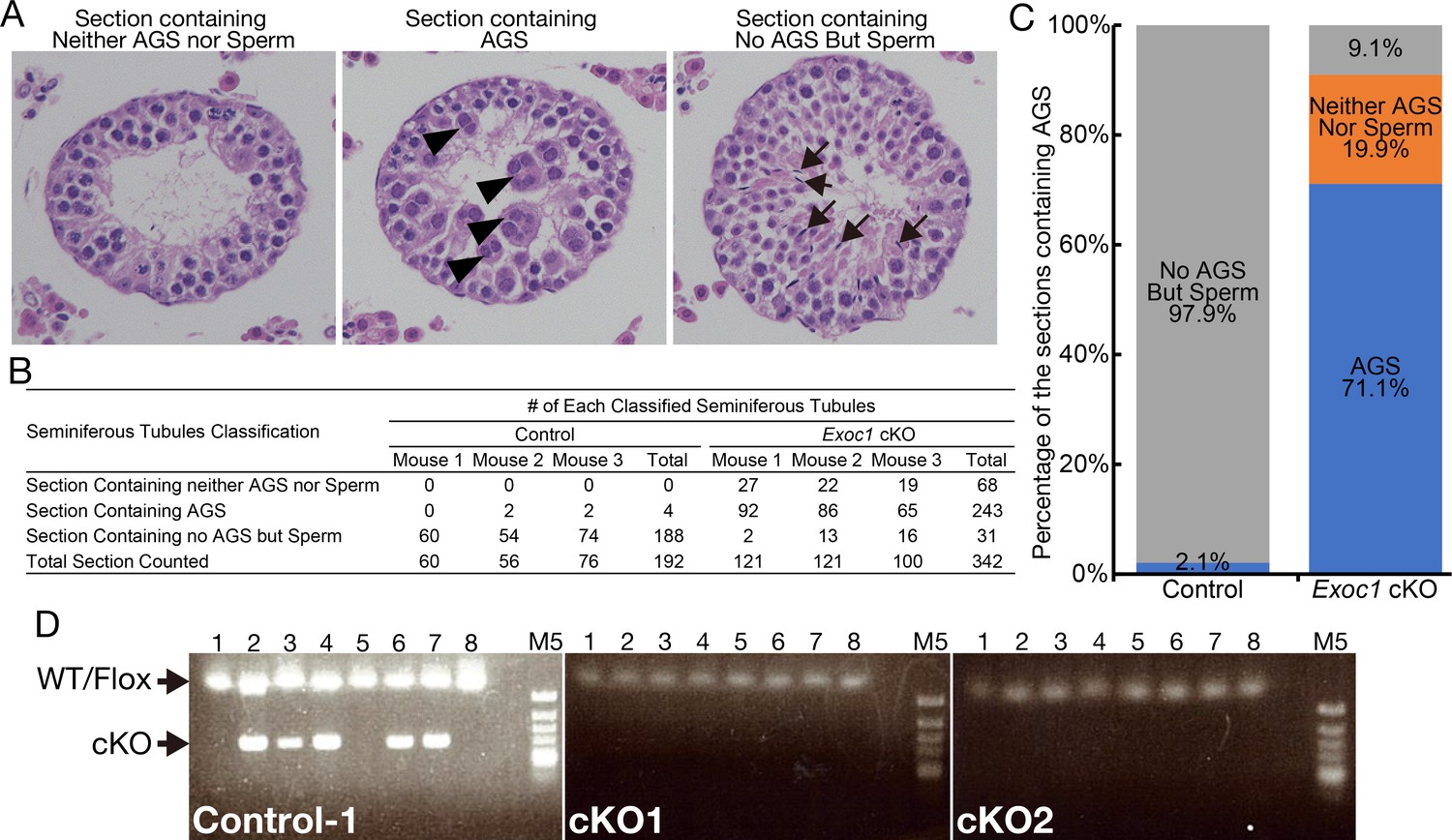
The occurrence frequency of aggregates of syncytia (AGS) in the Exoc1 cKO.
(A) Classification reference image of cross-sections of seminiferous tubules to evaluate the incidence of AGS. Cross-sections with AGS indicated by arrowheads were classified as ‘Section containing AGS’. Cross-sections with nuclear condensed sperm (including spermatid and spermatozoa) indicated by arrow were classified as ‘Section containing No AGS But Sperm’. Cross-sections with no AGS or Sperm and empty space in the lumen were classified as ‘Section containing neither AGS nor Sperm’. (B) Cross-sections of the seminiferous tubules of Exoc1 cKO and control (Exoc1flox/wt:: Nanos3+/Cre) adult mice (n = 3 in each genotype) were classified into three categories based on the criteria in (A). (C) Graphical representation of the results in (B). (D) Genotypic analysis of blastocysts obtained by in vitro fertilization of sperm from Exoc1 cKO mice with wild-type oocytes revealed no cKO alleles that underwent Cre-LoxP recombination (0/16, n = 2). Control: Exoc1flox/cKO. M5: Marker 5 (Nippon Genetics).
Figure 2—figure supplement 2
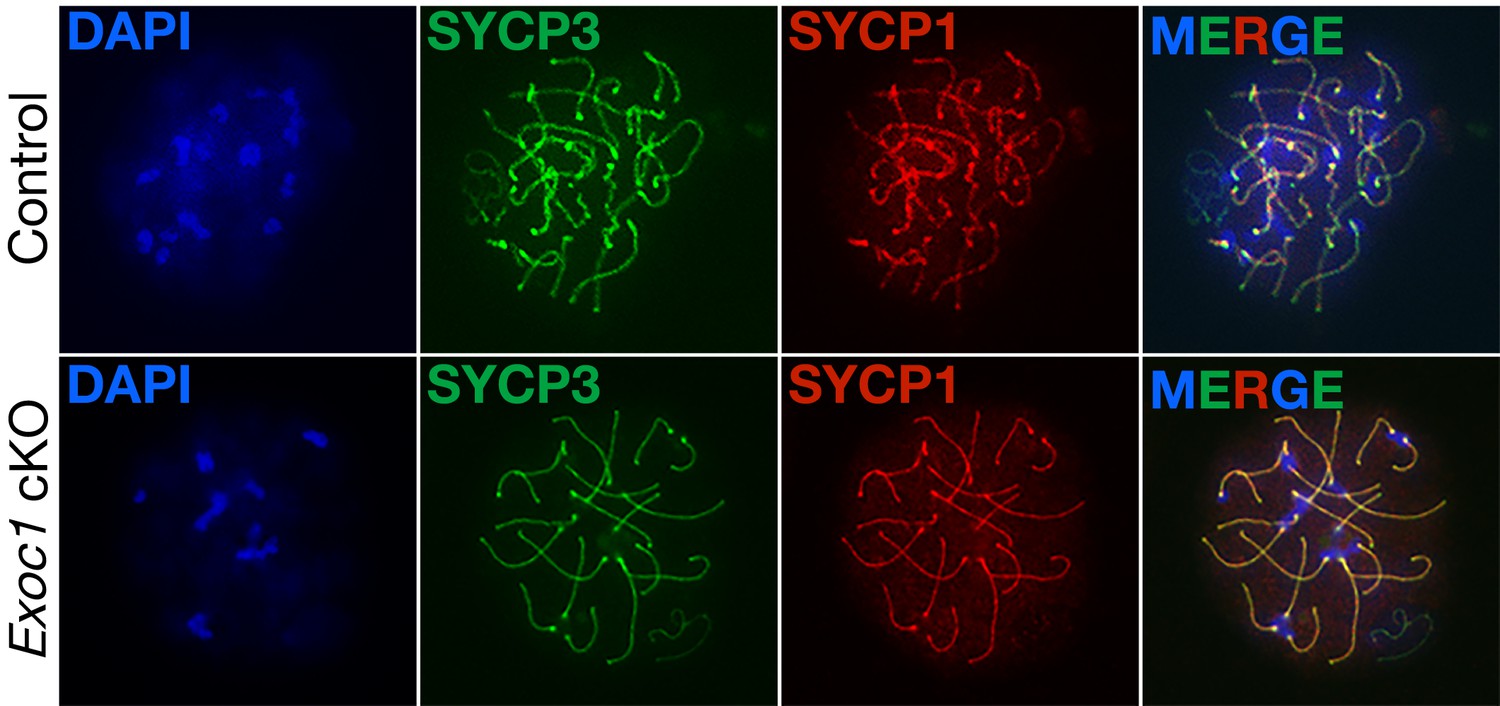
Normal meiotic chromosome synapsis in Exoc1 cKO mice.
Meiotic chromosome synapsis was confirmed by immunofluorescence. In pachytene stage, both the homologous pairing (marked by Sycp3) and synaptonemal complex (marked by Sycp1) were normal in Exoc1 cKO and control (wild-type) autosomal chromosomes (n = 2 in each genotype, 8–17 cells in each mouse).
Figure 3 with 2 supplements
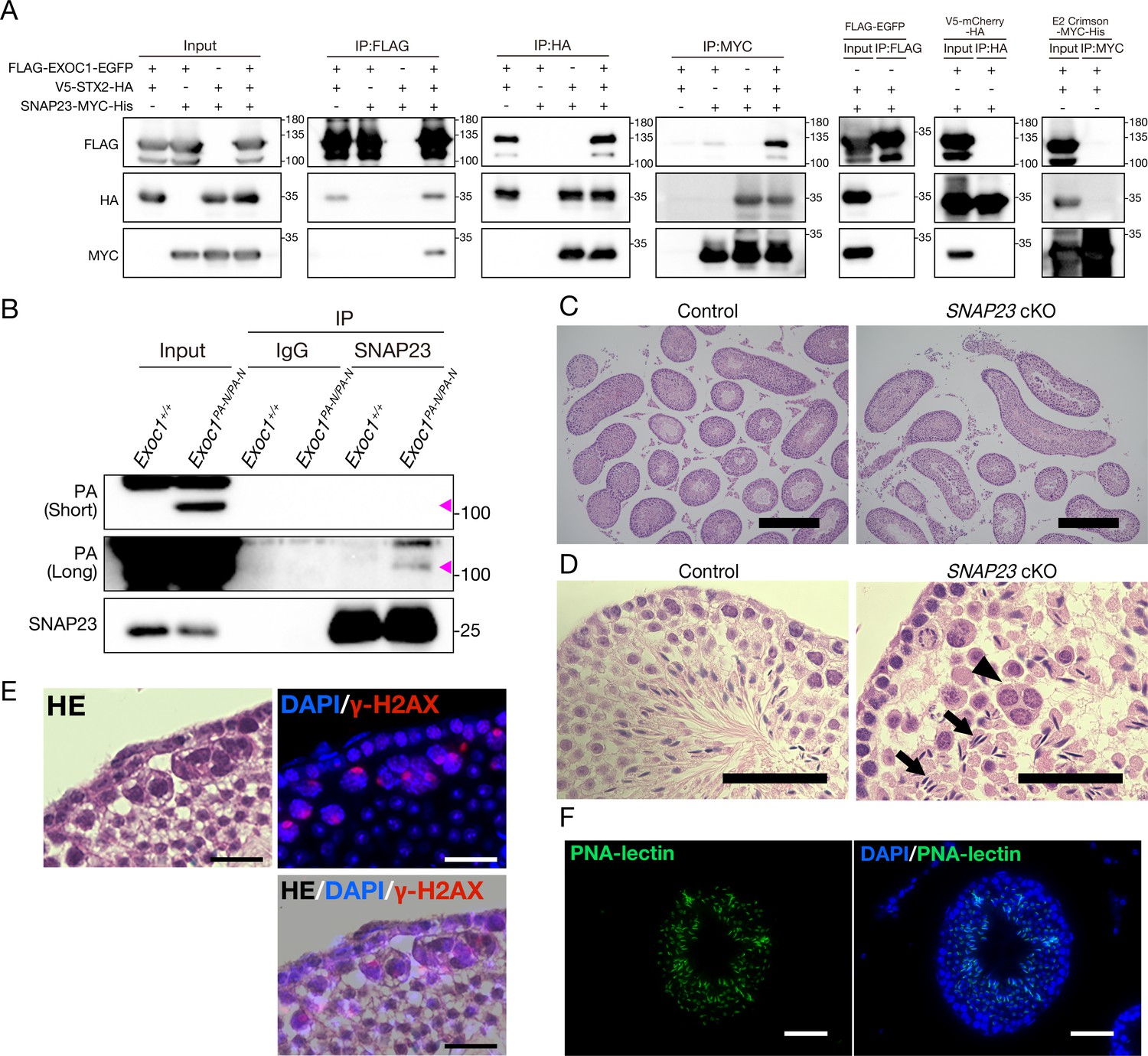
EXOC1 regulates ICB formation in cooperation with STX2 and SNAP23.
(A) Co-immunoprecipitation of EXOC1-STX2-SNAP23 complex in vitro. FLAG-tagged mouse EXOC1, HA-tagged mouse STX2, and Myc-tagged mouse SNAP23 were co-overexpressed in HEK293T cells. The binding of the three factors was confirmed in all combinations of Co-IP experiments. FLAG-EGFP, V5-mCherry-HA, and E2 crimson-MYC-His were used as negative controls. (B) Interaction of EXOC1 with SNAP23 in vivo. PA-tagged EXOC1 was co-immunoprecipitated with endogenous SNAP23 in the adult Exoc1PA-N testis. The upper and middle panels show short and long period exposure images, respectively. Arrowhead indicates PA-EXOC1. (C) The macroscopic images for H and E staining. Sperms were found in frequent seminiferous tubules in adult Snap23 cKO mice. Scale bars: 300 μm. Control: Snap23flox/wt:: Nanos3+/Cre mice. (D) The mesoscopic images for H and E staining of Snap23 cKO adult testis. Large and circular cells containing multiple nuclei (arrowheads), appearing to be aggregates of syncytia (AGS) were observed. In contrast with Exoc1 cKO, every seminiferous tubule had sperms with elongated nuclei (arrows). Scale bars: 50 μm. Control: Snap23flox/wt:: Nanos3+/Cre mice. (E) The serial section overlay image of Snap23 cKO testis. Immunofluorescence signals of γ-H2AX were found in AGS. Scale bars: 50 μm. (F) PNA-lectin staining of Snap23 cKO testis. PNA-lectin that was used to detect the acrosome is observed in the lumen of the seminiferous tubule of Snap23 cKO testis. Scale bars: 50 μm.
-
Figure 3—source data 1
Raw data of the in vitro immunoprecipitation.
- https://cdn.elifesciences.org/articles/59759/elife-59759-fig3-data1-v1.pptx
-
Figure 3—source data 2
Raw data of the in vivo immunoprecipitation.
- https://cdn.elifesciences.org/articles/59759/elife-59759-fig3-data2-v1.pptx
-
Figure 3—source data 3
Occurrence rate of AGS per area in the extracted Section containing AGS.
- https://cdn.elifesciences.org/articles/59759/elife-59759-fig3-data3-v1.xlsx
Figure 3—figure supplement 1
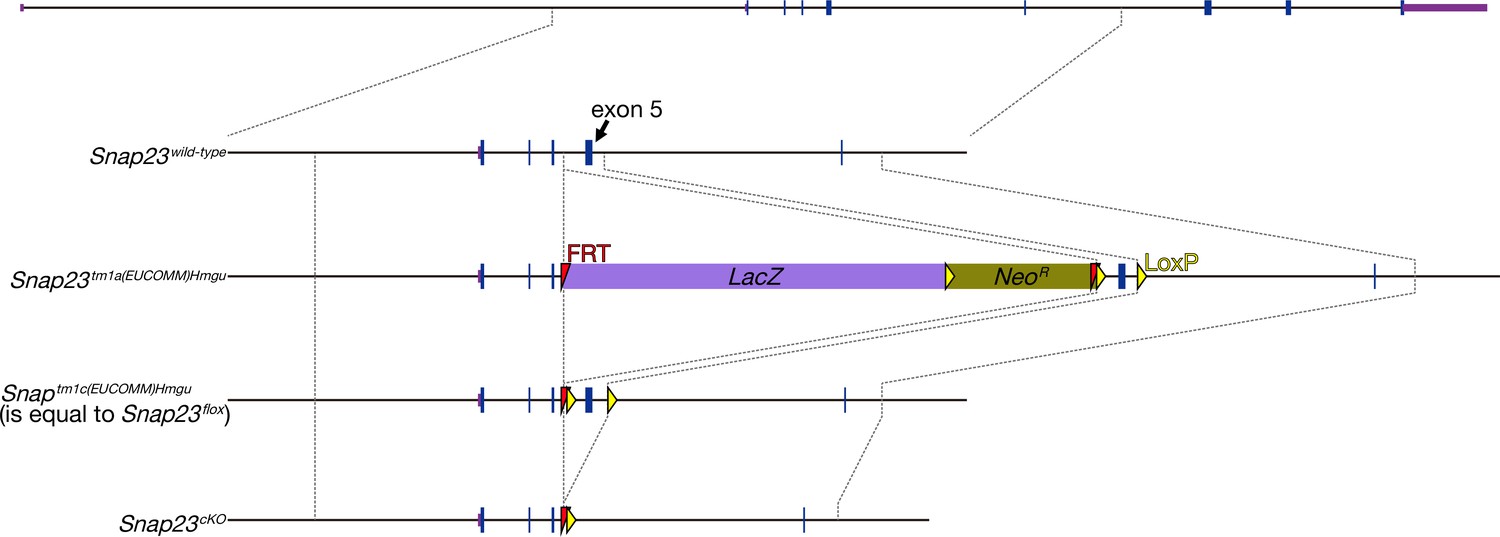
The production of Snap23 flox mice.
Snap23 flox mice were from the Snap23tm1a(EUCOMM)Wtsi frozen sperms. Snap23tm1a(EUCOMM)Wtsi frozen sperms were thawed and fertilized to wild-type oocyte in vitro. Flpe mRNA was electroplated into the fertilized embryos. Snap23tm1a(EUCOMM)Wtsi was changed to Snap23tm1c(EUCOMM)Wtsi (equal to Snap23 flox) allele in every newborn. Exon 5 of Snap23 was floxed in this allele. Arrows indicate the primers for the detection of flox allele.
Figure 3—figure supplement 2
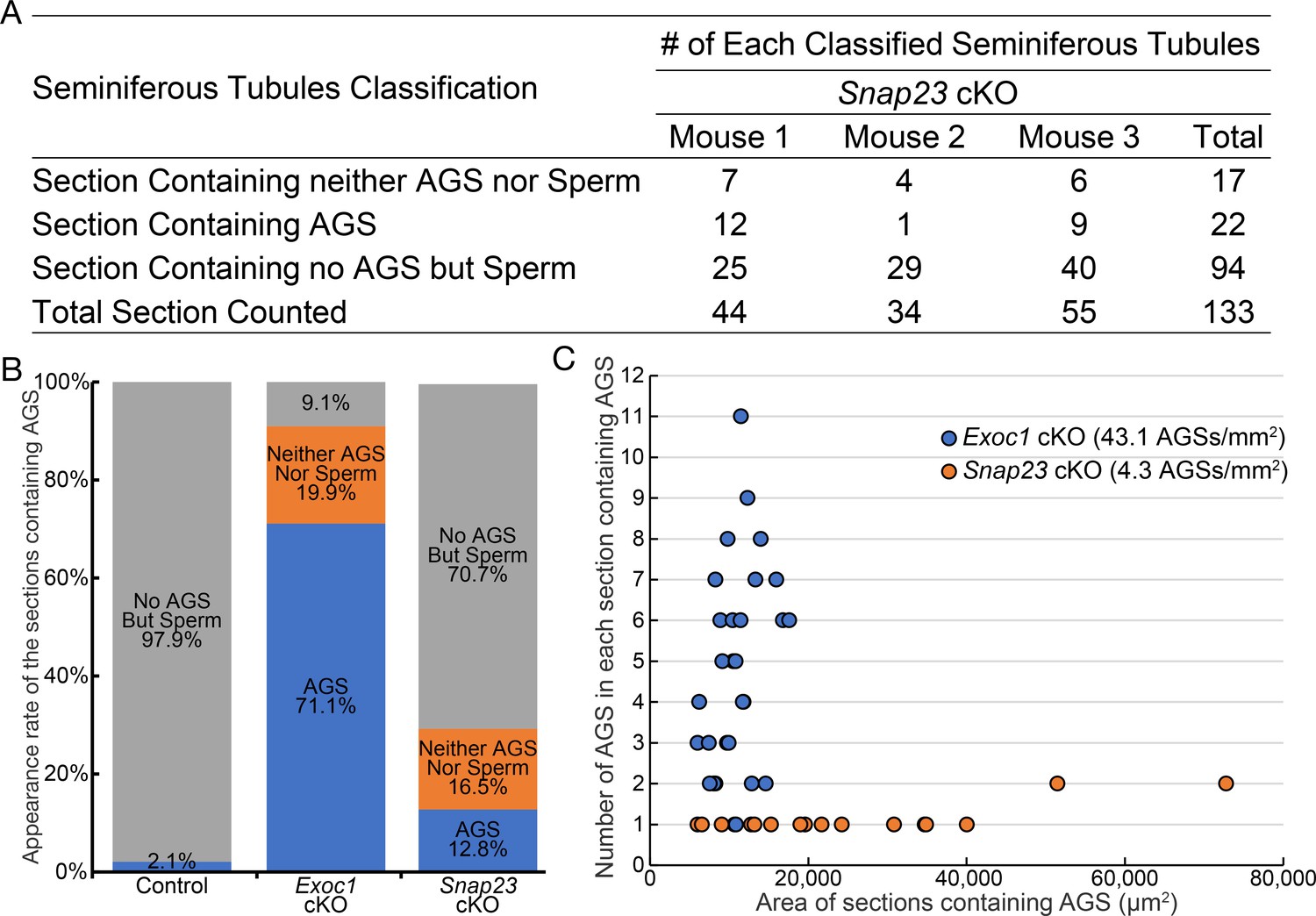
The occurrence frequency of aggregates of syncytia (AGS) in the Snap23 cKO.
(A) Results of classifying the cross-section of the seminiferous tubules of adult Snap23 cKO (n = 3) using the same criteria as in Figure 3—figure supplement 1A. (B) Graph is based on Figure 3—figure supplement 1C, with Snap23 cKO data (A) added to it. (C) ‘Section containing AGS’ classified images were extracted and plotted with the area of each cross-section on the X-axis and the number of AGS in each cross-section on the Y-axis.
Figure 4 with 1 supplement
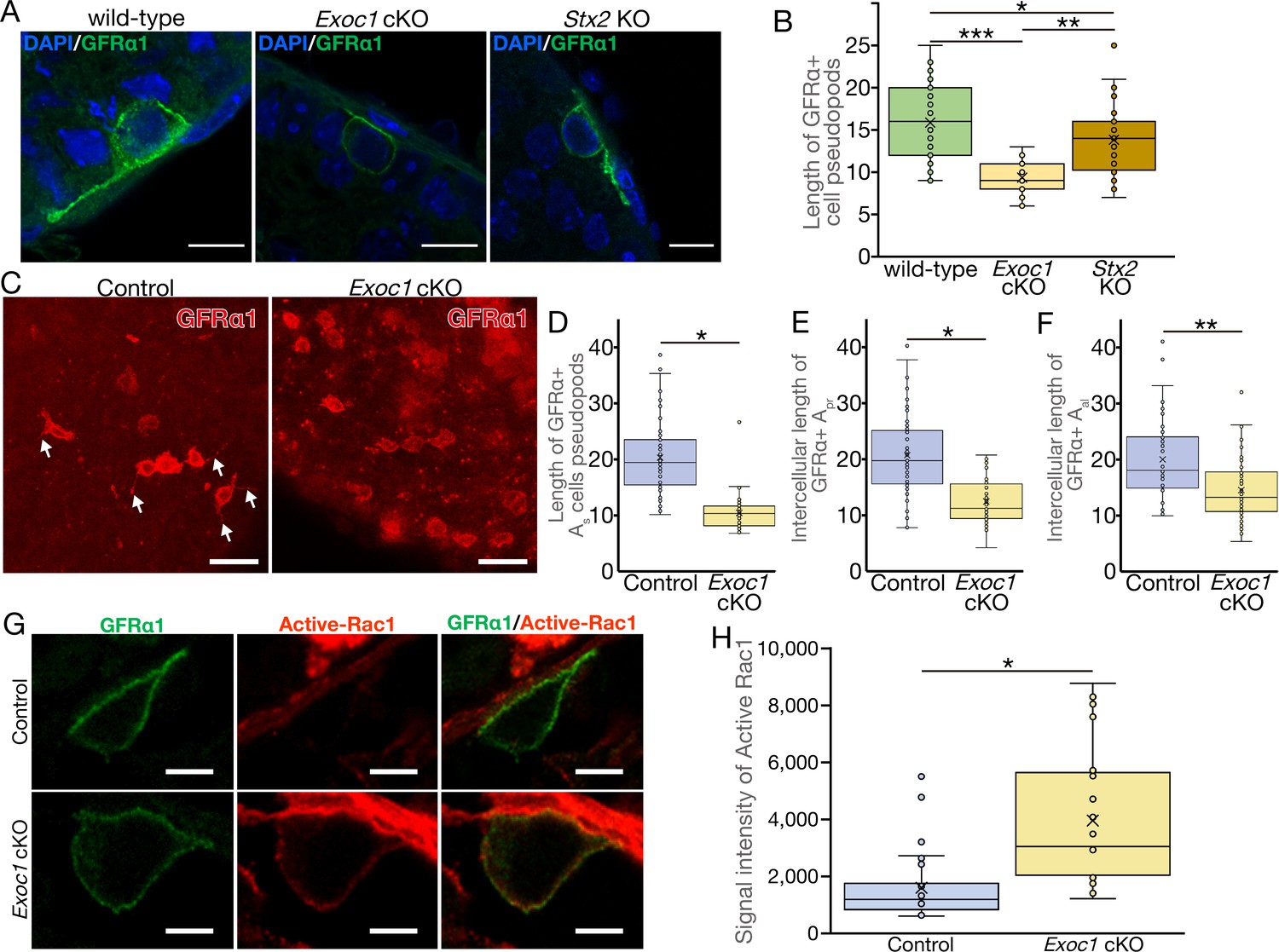
EXOC1 regulates pseudopod elongation via Rac1 inactivation.
(A) A representative image of GFRα1+ undifferentiated spermatogonia in Exoc1 cKO and Stx2 KO. Pseudopod elongation was impaired in Exoc1 cKO, but not in Stx2 KO. Scale bars: 10 μm. (B) Pseudopod length quantification using sections. Average length of GFRα1+ spermatogonia pseudopods in Exoc1 cKO was shorter than that of Stx2 KO and wild type (n = 3 in each genotype, 25–36 cells in each mouse). *p=0.052, **p=1.8 × 10−6, ***p=9.5 × 10−9. one-way ANOVA. (C) Pseudopod length quantification through whole-mount immunofluorescence staining of adult testes. GFRα1+ spermatogonia with elongated pseudopod (white arrows) were frequently observed in control (Exoc1flox/cKO) mice, whereas they were rarely observed in Exoc1 cKO mice. Scale bars: 30 μm. (D) Measurement of the length of pseudopod of Asingle GFRα1+ cells based on whole-mount immunofluorescence images (n = 3 in each genotype, 20 cells in each mouse). *p=7.2 × 10−17, Student’s t-test. Control: Exoc1flox/cKO. (E, F) Measurement of intercellular length in connected Apair (n = 3 in each genotype, 20 intercellular distances in each mouse) or Aaligned (n = 3 in each genotype, 14–19 intercellular distances in each mouse) based on whole-mount immunofluorescence images. *p=3.6 × 10−12, **p=0.00014, Student’s t-test. Control: Exoc1flox/cKO. (G) A representative image of active-Rac1 in GFRα1+ spermatogonia of Exoc1 cKO adult testis. In control mice (Exoc1flox/wt:: Nanos3+/Cre), active-Rac1 signal was lower than the detection limit. Non-polar active-Rac1 signal was detected in Exoc1 cKO adult testis. Scale bars: 5 μm. (H) Quantification of signal intensity of active-Rac1 in GFRα1+ spermatogonia based on immunostaining images. The average intensity in each cell is higher in the Exoc1 cKO group than that in the control group (n = 3 in genotype, 8–10 cells in each mouse). *p=0.000036, Student’s t-test. Control: Exoc1flox/flox.
-
Figure 4—source data 1
Measurement of the length of the pseudopodia of GFRα1+ cells in section observation.
- https://cdn.elifesciences.org/articles/59759/elife-59759-fig4-data1-v1.xlsx
-
Figure 4—source data 2
Measurement of the length of the pseudopodia of GFRα1+ cells in whole-mount observation.
- https://cdn.elifesciences.org/articles/59759/elife-59759-fig4-data2-v1.xlsx
-
Figure 4—source data 3
Intensity of active Rac1 signal in each cell.
- https://cdn.elifesciences.org/articles/59759/elife-59759-fig4-data3-v1.xlsx
Figure 4—figure supplement 1
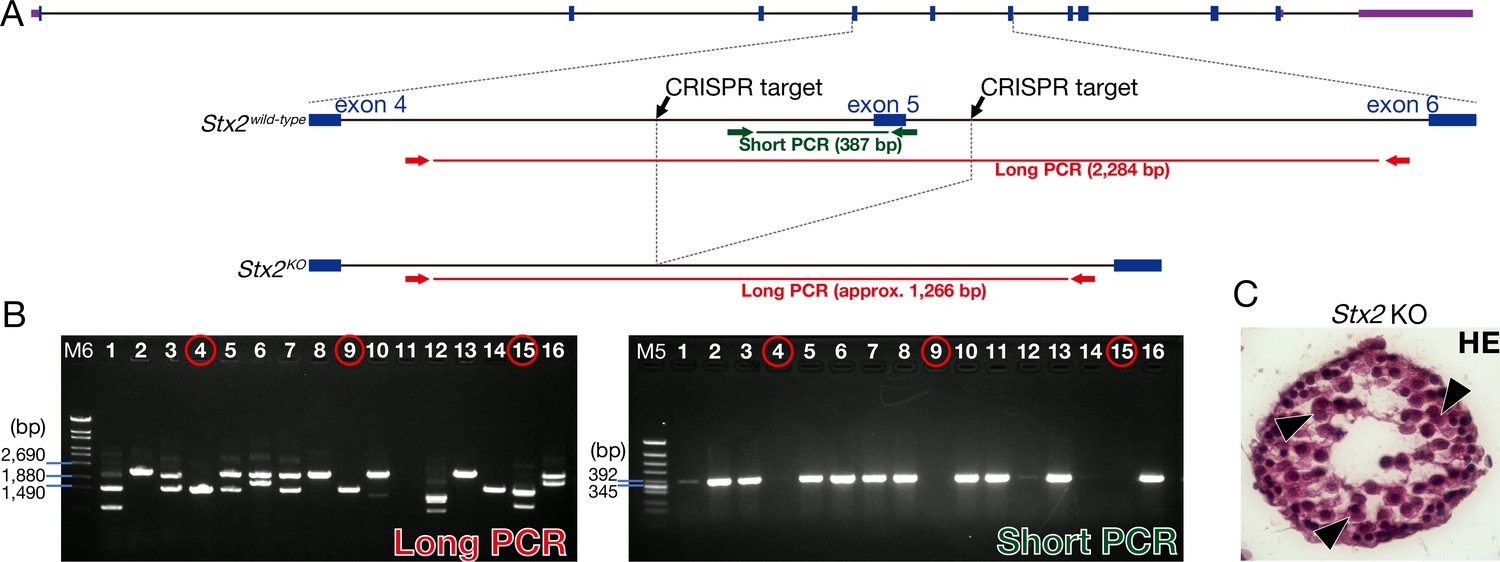
The production of the Stx2 KO.
(A) Stx2 KO mice generated by the CRISPR-Cas9 genome editing in mouse zygotes. Two CRISPR targets located 482 bp upstream and 148 bp downstream of exon 5 of Stx2. Arrows indicate long and short PCR primers for detecting the deletion of exon 5 of Stx2. (B) Electrophoresis data of PCR genotyping. We used #4, 9, and 15 as the Stx2 KO mice. M5: Marker 5. M6: Marker 6. (C) H and E staining of Stx2 KO testis. Consistent with reported Stx2 null phenotype (Fujiwara et al., 2013), our Stx2 KO also showed aggregates of syncytia (AGS) (arrowheads).
Figure 5 with 1 supplement
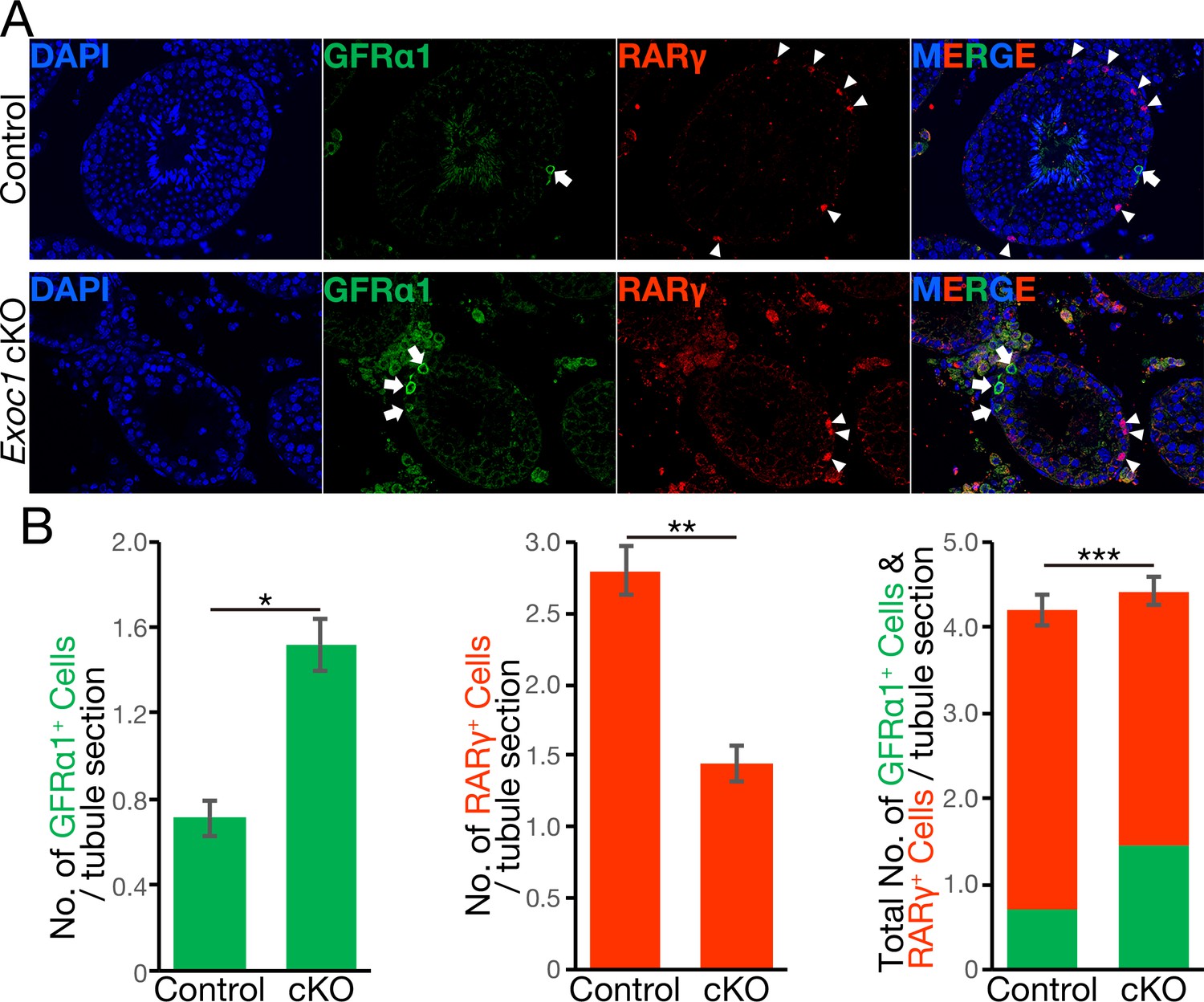
The balance of spermatogonial differentiation is perturbed in Exoc1 cKO.
(A) Representative immunofluorescence image of adult Exoc1 cKO seminiferous tubule. GFRα1+ spermatogonia (arrow) density in the cKO was higher than that in control mice. RARγ+ spermatogonia (arrowhead) density decreased in the cKO. Control: Exoc1flox/flox mice. (B) The number of spermatogonia in a cross-section of seminiferous tubule (n = 3 in each genotype, 46–87 sections in each mouse). The number of GFRα1+ cell per section was significantly increased in the cKO. The number of RARγ+ cell in the cKO was significantly smaller than that in control (Exoc1flox/flox) mice. *p=1.9 × 10−7, **p=3.6 × 10−9, ***p=0.035, Student’s t-test.
-
Figure 5—source data 1
The number of GFRα1+ and RARγ+ cells.
- https://cdn.elifesciences.org/articles/59759/elife-59759-fig5-data1-v1.xlsx
-
Figure 5—source data 2
The number of c-Kit+ cells.
- https://cdn.elifesciences.org/articles/59759/elife-59759-fig5-data2-v1.xlsx
Figure 5—figure supplement 1
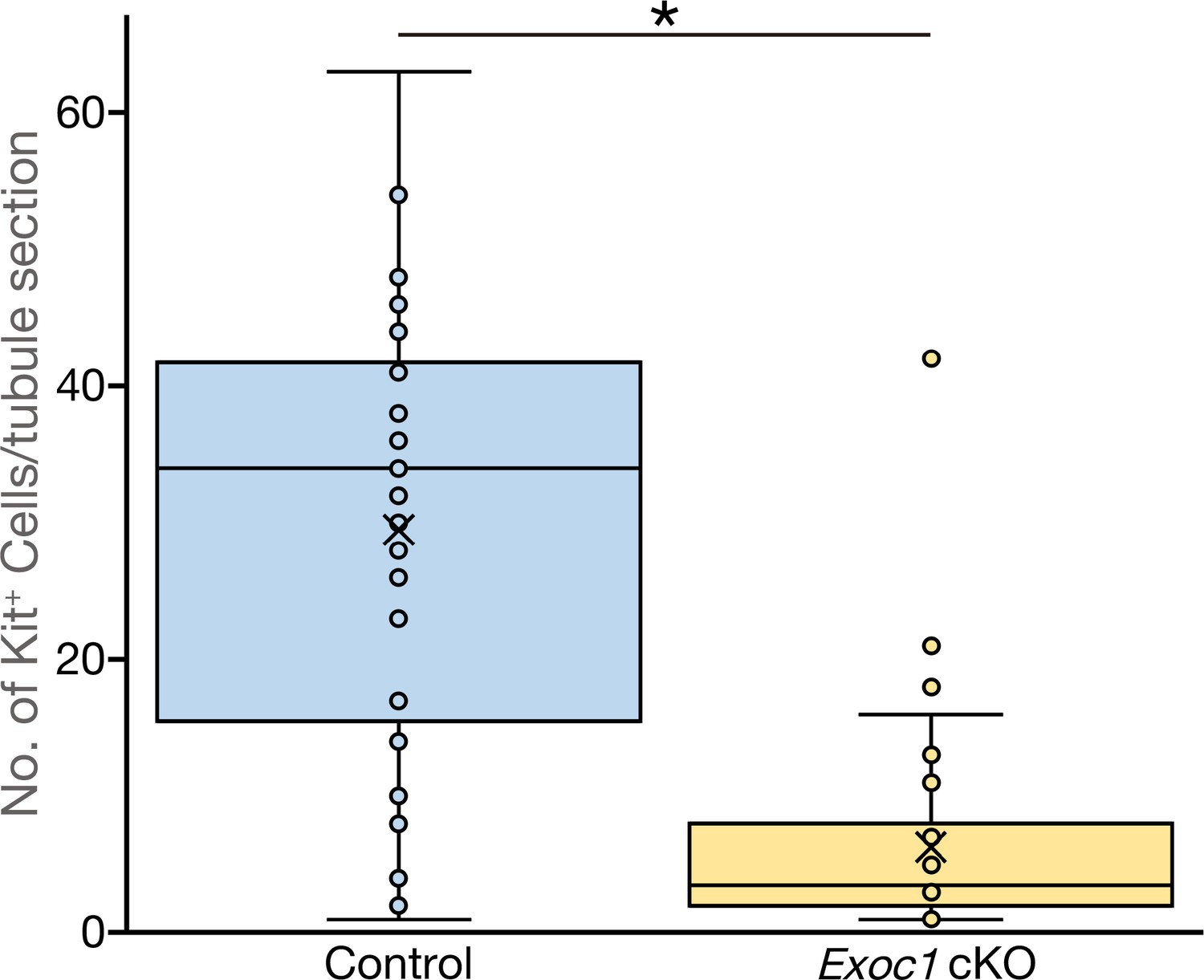
The number of Kit+ differentiating spermatogonia are reduced in Exoc1 cKO.
The number of Kit+ spermatogonia in each one cross-section of seminiferous tubule in adult mice (n = 3 in each genotype, 20 sections in each mouse). *p=0.000012, Student’s t-test. Control: Exoc1flox/flox.
Author response image 1
Author response image 2
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (M. musculus) | C57BL/6J | Charles River Laboratories Japan | Stock No: 000664 | |
| Strain, strain background (M. musculus) | Crl:CD1(ICR) | Charles River Laboratories Japan | ||
| Genetic reagent (M. musculus) | Exoc1 flox | This paper | This is from the Exoc1tm1a(EUCOMM)Hmgu. IKMC Project #78575 | |
| Genetic reagent (M. musculus) | Exoc1PA-C | This paper | ||
| Genetic reagent (M. musculus) | Exoc1PA-N | This paper | ||
| Genetic reagent (M. musculus) | Snap23 flox | This paper | This is from the Snap23tm1a(EUCOMM)Wts. Colony Name BLA3054 | |
| Genetic reagent (M. musculus) | Stx2 KO | This paper | ||
| Genetic reagent (M. musculus) | Nanos3tm2 (cre)Ysa | Suzuki et al., 2008 | Prof. Saga, RIKEN BRC (RDB13130) | RBRC02568 |
| Genetic reagent (M. musculus) | B6;SJL-Tg(ACTFLPe)9205Dym/J | The Jackson Laboratory | Stock No: 003800 | |
| Cell line (Human) | HEK293T cell | ATCC | ATCC Sales Order: SO0623448 | FTA Barcode: STRB4056 |
| Antibody | Monoclonal anti-PA-tag, Biotin conjugated (Rat) | FUJIFILM Wako Chemicals | Cat#017–27731 | WB (1:500) |
| Antibody | Polyclonal anti-b-actin (Rabbit) | MEDICAL and BIOLOGICAL LABORATORIES | Cat#PM053 | WB (1:3000) |
| Antibody | Monoclonal anti-DYKDDDDK tag (Mouse) | FUJIFILM Wako Chemicals | Cat# 018–22381 | WB (1:2000) |
| Antibody | Monoclonal anti-DYKDDDDK tag (Rat) | FUJIFILM Wako Chemicals | Cat# 018–23621 | WB (1:2000) |
| Antibody | Monoclonal anti-HA-tag (Rabbit) | Cell Signaling Technology | Cat#3724 | WB (1:2000) |
| Antibody | Monoclonal anti-HA-tag (Mouse) | BioLegend | Cat#901513 | WB (1:1000) |
| Antibody | Monoclonal anti-Myc (Mouse) | MEDICAL and BIOLOGICAL LABORATORIES | Cat#M192-3 | WB (1:1000) |
| Antibody | Monoclonal anti-SNAP23 (Mouse) | Santa Cruz Biotechnology | Cat#sc-166244 | WB (1:1000) |
| Antibody | Anti-Rat IgG, HRP-linked (Goat) | GE Healthcare | Cat#NA935V | WB (1:30000) |
| Antibody | Anti-Rabbit IgG, HRP-linked (Donkey) | GE Healthcare | Cat#NA934V | WB (1:30000) |
| Antibody | Anti-Mouse IgG, HRP-linked (Sheep) | GE Healthcare | Cat#NA931V | WB (1:30000) |
| Antibody | Normal IgG (Rabbit) | FUJIFILM Wako Chemicals | Cat#148–09551 | Co-IP |
| Antibody | Polyclonal anti-SNAP23 (Rabbit) | Abcam | Cat#ab3340 | Co-IP |
| Antibody | Monoclonal anti-PA-tag (Rat) | FUJIFILM Wako Chemicals | Cat#016–25861 | IF (1:1000) |
| Antibody | Monoclonal anti-γH2AX (Mouse) | Merck-Millipore | Cat#05–636 | IF (1:100) |
| Antibody | Polyclonal anti-SYCP1 (Rabbit) | Novus Biological | Cat#NB300-299 | IF (1:50) |
| Antibody | Monoclonal anti-SYCP3 (Mouse) | Santa Cruz Biotechnology | Cat#sc-74569 | IF (1:50) |
| Antibody | Polyclonal anti-GFRα1 (Goat) | R and D systems | Cat#AF560 | IF (1:400 for section) IF (1:1000 for whole mount) |
| Antibody | Monoclonal anti-RARγ1 (Rabbit) | Cell Signaling Technology | Cat#8965S | IF (1:200) |
| Antibody | Monoclonal anti-active rac1 (Mouse) | NewEast Biosciences | Cat#26903 | IF (1:1000) |
| Antibody | Polyclonal anti-Exoc1 (Rabbit) | Proteintech | Cat#11690–1-AP | IF (1:50) |
| Antibody | Polyclonal anti-Exoc1 (Rabbit) | Atlas Antibodies | Cat#HPA037706 | IF (1:50) |
| Antibody | Anti-Goat IgG, Alexa Fluor 488 (Chicken) | Thermo Fisher Scientific | Cat#A21467 | IF (1:200 for section) |
| Antibody | Anti-Goat IgG, Alexa Fluor 594 (Chicken) | Thermo Fisher Scientific | Cat#A21468 | IF (1:400 for whole mount) |
| Antibody | Anti-Rat IgG, Alexa Fluor 555 (Donkey) | Abcam | Cat#ab150154 | IF (1:1000) |
| Antibody | Anti-Mouse IgG, Alexa Fluor 555 (Goat) | Thermo Fisher Scientific | Cat#A28180 | IF (1:200) |
| Antibody | Anti-Mouse IgG, Alexa Fluor 555 (Donkey) | Thermo Fisher Scientific | Cat#A31570 | IF (1:200) |
| Antibody | Anti-Rabbit IgG, Alexa Fluor 647 (Goat) | Thermo Fisher Scientific | Cat#A27040 | IF (1:200) |
| Recombinant DNA reagent | pcDNA3.1 (+) Mammalian Expression Vector | Invitrogen | V79020 | |
| Recombinant DNA reagent | T7-NLS hCas9-pA plasmid | Yoshimi et al., 2016 | RIKEN BRC (RDB13130) | |
| Recombinant DNA reagent | pCAG-Flpe | Matsuda and Cepko, 2007 | Addgene (Plasmid #13787) | |
| Recombinant DNA reagent | pT7-Flpe-pA | This paper | RIKEN BRC (RDB16011) | |
| Sequence-based reagent | All primers in Supplementary file 1b | Thermo Fisher Scientific | ||
| Sequence-based reagent | All primers in Supplementary file 1b | Thermo Fisher Scientific | ||
| Sequence-based reagent | Exoc1 PA-C ssODN | Integrated DNA Technologies | GAATTCACTATTCAGGACATTCTGGATTATTGCTCCAGCATCGCACAGTCCCACGGCTCAACCAGCGGATCTGGTAAGCCAGGTAGTGGAGAAGGCAGCACCAAGCCTGGCGGCGTCGCCATGCCTGGAGCCGAGGATGATGTCGTGTAAGCCCTAGGAAAGAGGAGAAAGAAGTGAGCATGCATTCTCAGTCCAGCAAA | |
| Sequence-based reagent | Exoc1 PA-N ssODN | Integrated DNA Technologies | GGAGGGCAGTGGTTTTGAGAATTATTCTAAATGTTTTTCAGCTGAGAAAAGATGGGCGTCGCCATGCCTGGAGCCGAGGATGATGTCGTGGGCTCAACCAGCGGATCTGGTAAGCCAGGTAGTGGAGAAGGCAGCACCAAGCCTGGCACAGCAATCAAGCATGCGCTGCAGAGAGATATCTTCACACCAAATGATGAACG | |
| Software, algorithm | The R Foundation | https://www.r-project.org/foundation/ | ||
| Other | Streptavidin-HRP | Nichirei Biosciences | Cat#426061 | WB (1:1000 in 2% BSA/TBS-T) |
| Other | Lectin from Arachis hypogaea, FITC | Sigma-Adrich | Cat#L7381 | Lectin staining (1:100) |
Additional files
-
Supplementary file 1
Data of differentiating spermatogonia aggregation and primers for genotyping.
(a) Differentiating spermatogonia are not aggregated. Examination of aggregated Kit+ differentiating spermatogonia in adult Exoc1 cKO mice. Aggregation was determined by observation of immunofluorescence images with anti-Kit antibody. (b) Primers for genotyping The following is a list of primers used for genotyping or vector construction.
- https://cdn.elifesciences.org/articles/59759/elife-59759-supp1-v1.xlsx
-
Transparent reporting form
- https://cdn.elifesciences.org/articles/59759/elife-59759-transrepform-v1.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
EXOC1 plays an integral role in spermatogonia pseudopod elongation and spermatocyte stable syncytium formation in mice
eLife 10:e59759.
https://doi.org/10.7554/eLife.59759




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}