A TORC1-histone axis regulates chromatin organisation and non-canonical induction of autophagy to ameliorate ageing
- Max Planck Institute for Biology of Ageing, Germany
- Institute of Healthy Ageing, Department of Genetics, Evolution and Environment, University College London, United Kingdom
- Department of Pathology, University of Michigan, United States
Figures
Figure 1 with 3 supplements
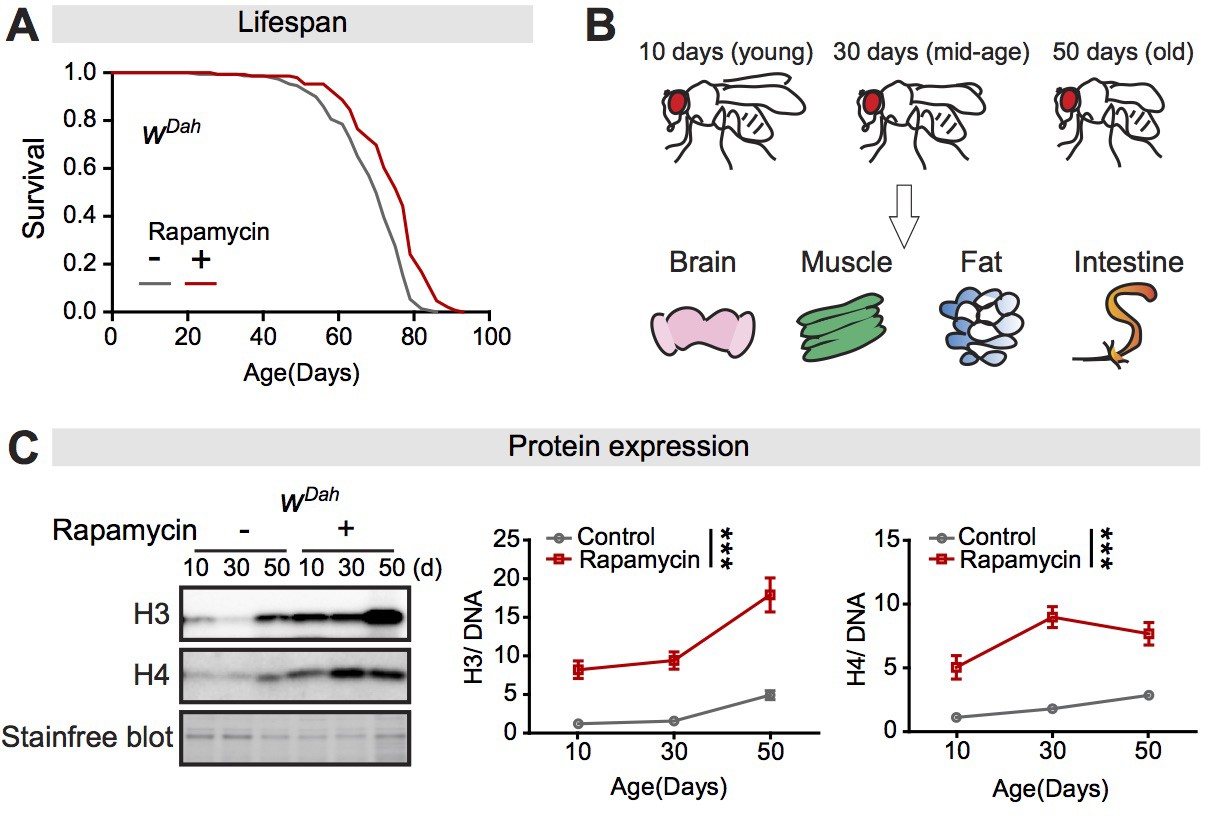
Expression of core histones in the fly intestine increases with age and in response to rapamycin treatment.
(A) Adult-onset rapamycin treatment extended the lifespan of wDah females (log-rank test, p=7.4E-08). See also Supplementary file 1. (B) Four tissues were dissected: brain, muscle, fat, and intestine, at 10 days, 30 days, and 50 days of adult age. (C) Expression of H3 and H4 in dissected intestines of wDah controls significantly increased with age. Rapamycin substantially increased the expression of H3 and H4 in intestine (n = 4 biological replicates of 10 intestines per replicate, two-way ANOVA, H3 and H4, age p<0.05, treatment p<0.001, interaction p>0.05). The amount of protein was normalised to DNA, shown by stain-free blot.
-
Figure 1—source data 1
Source data pertaining to Figure 1.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig1-data1-v2.xlsx
Figure 1—figure supplement 1
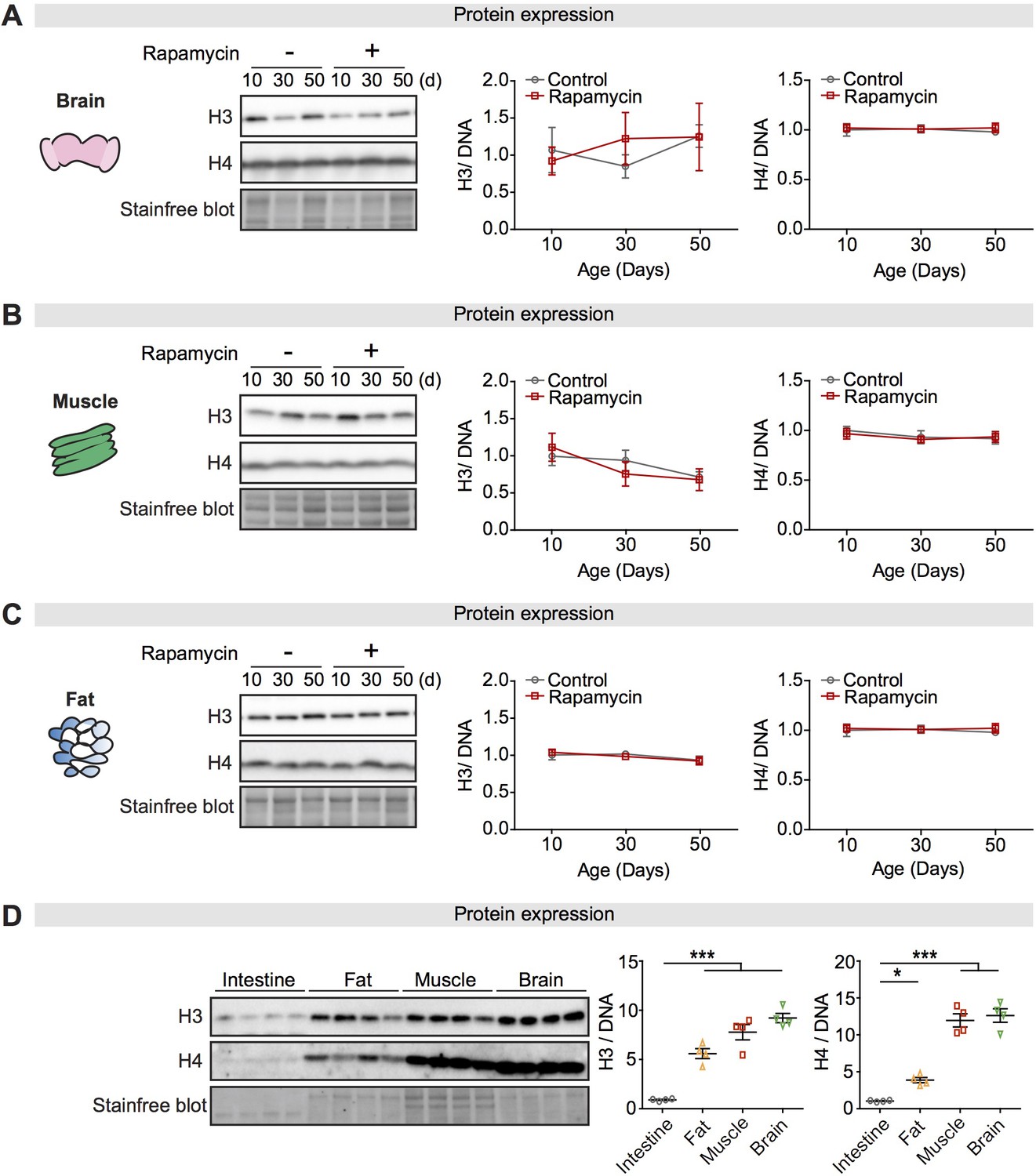
Expression of core histones in different tissues and with rapamycin treatment.
(A–C) Expression of H3 and H4 in dissected brain, muscle, or fat of wDah control flies did not change significantly with age or rapamycin treatment (n = 4 biological replicates of 10 tissues per replicate, two-way ANOVA, H3 and H4, age p>0.05, treatment p>0.05, interaction p>0.05). (D) Expression of H3 and H4 in intestine was markedly lower than in fat, muscle, or brain of wDah control flies at 5 days of age (n = 4 biological replicates of 10 tissues per replicate, one-way ANOVA; post-hoc test, *p<0.05, ***p<0.001). The amount of protein was normalised to DNA, shown by stain-free blot.
-
Figure 1—figure supplement 1—source data 1
Source data pertaining to Figure 1—figure supplement 1.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig1-figsupp1-data1-v2.xlsx
Figure 1—figure supplement 2
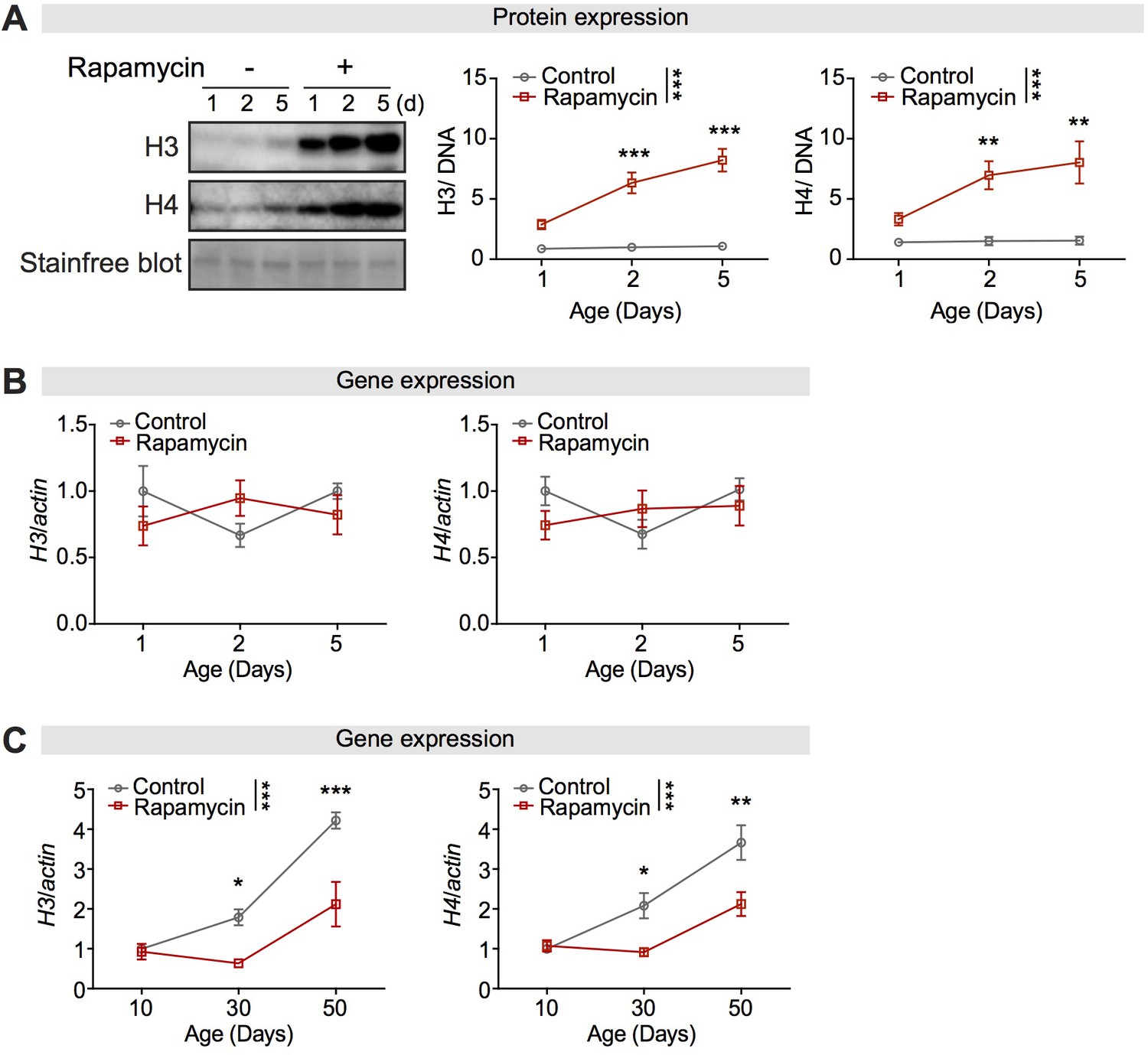
Rapamycin increased expression of histone proteins without affecting their transcripts in the fly intestine.
(A) Expression of H3 and H4 in dissected intestines was unchanged over 5 days in wDah controls, but had substantially increased after 2 days in rapamycin-treated flies (n = 4 biological replicates of 10 intestines per replicate, two-way ANOVA, H3 and H4, age p<0.05, treatment p<0.001, interaction p<0.05; post-hoc test, NS p>0.05, **p<0.01, ***p<0.001). The amount of protein was normalised to DNA, shown by stain-free blot. (B) Rapamycin treatment did not affect the expression of H3 and H4 transcripts in the intestine of young flies at 1, 2, and 5 days of age (n = 3–4 biological replicates of 15 intestines per replicate, two-way ANOVA, age p>0.05, treatment p>0.05, interaction p>0.05). (C) Expression of H3 and H4 transcripts in dissected intestines of control flies significantly increased with age. Rapamycin treatment attenuated the age-associated increase in H3 and H4 in the intestine of flies at 30 days and 50 days of age (n = 3–4 biological replicates of 15 intestines per replicate, two-way ANOVA; H3 and H4, age p<0.001, treatment p<0.001, interaction, p<0.01).
-
Figure 1—figure supplement 2—source data 1
Source data pertaining to Figure 1—figure supplement 2.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig1-figsupp2-data1-v2.xlsx
Figure 1—figure supplement 3
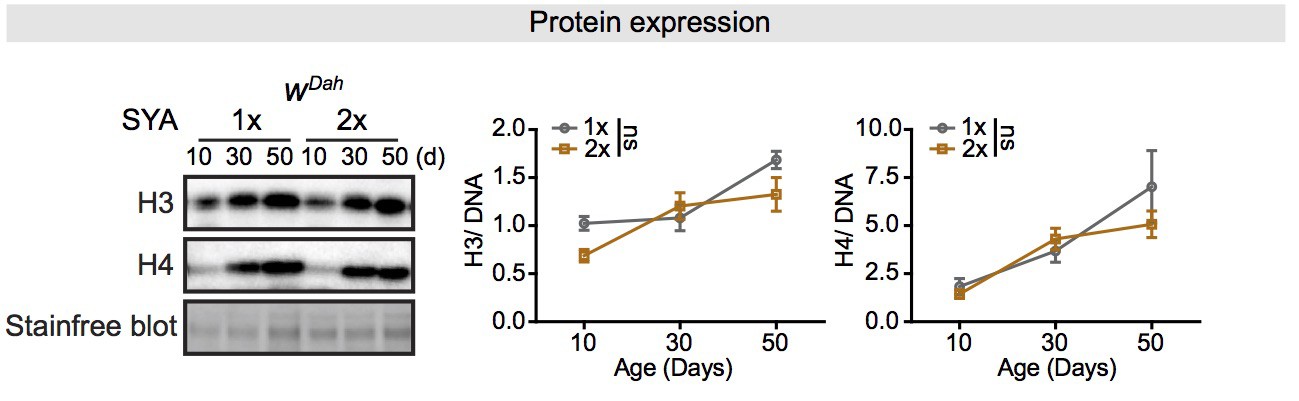
Expression of core histones in the fly intestine increases with age but does not affect by dietary restriction (DR).
Expression of H3 and H4 in dissected intestines of wDah controls significantly increased with age. DR (1× vs. 2× SYA food) did not influence the expression of H3 and H4 in intestine (n = 4 biological replicates of 10 intestines per replicate, two-way ANOVA, H3 and H4, age p<0.001, treatment p>0.05, interaction p>0.05). The amount of protein was normalised to DNA, shown by stain-free blot.
-
Figure 1—figure supplement 3—source data 1
Source data pertaining to Figure 1—figure supplement 3.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig1-figsupp3-data1-v2.xlsx
Figure 2 with 2 supplements
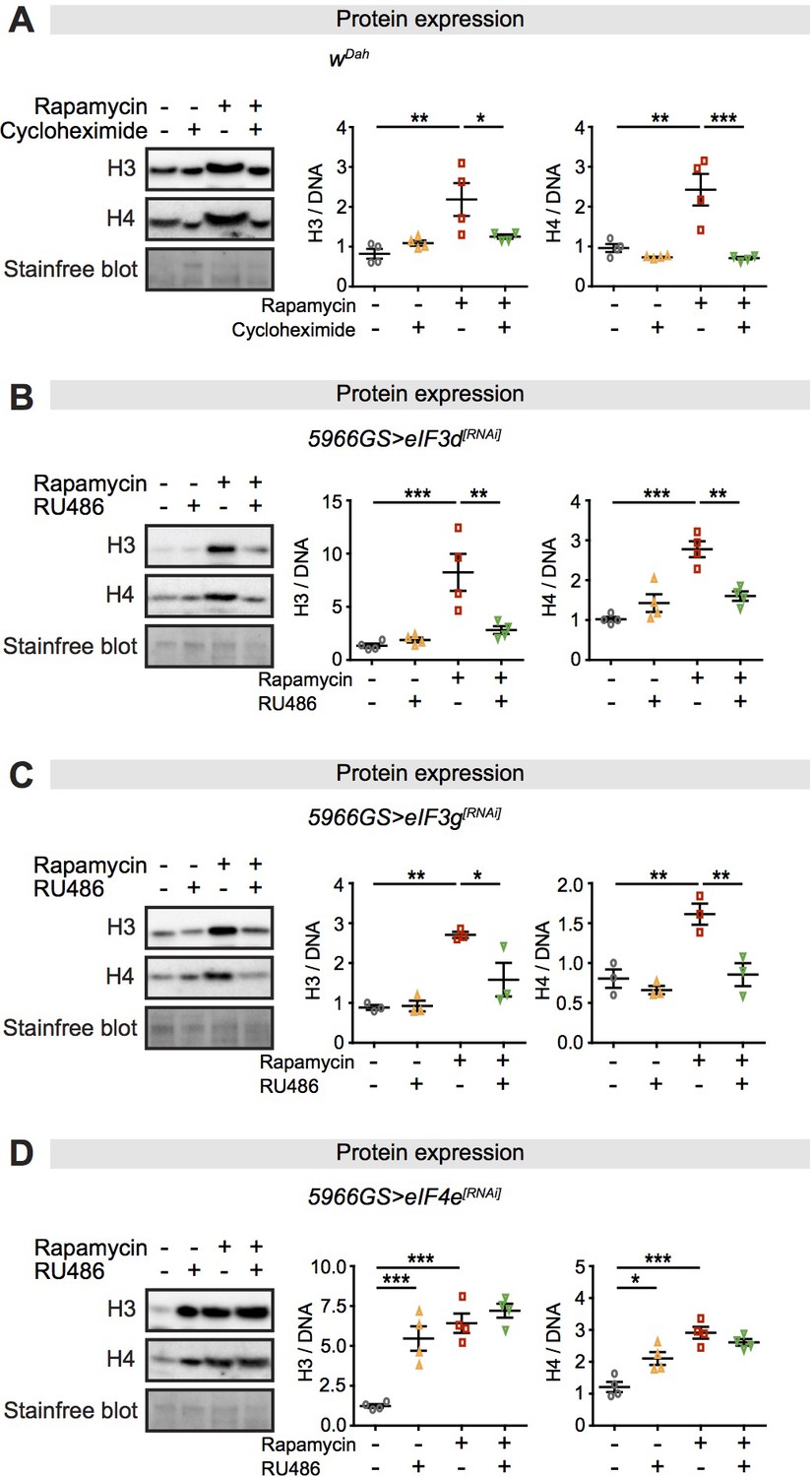
Expression of core histones in the fly intestine in response to rapamycin treatment and inhibition of translation or translation factors eukaryotic initiation factor (eIF)3 and eIF4.
(A) Adult-onset cycloheximide treatment (1 mM) alone had no effect on histone expression but blocked increased expression of histones H3 and H4 in response to rapamycin treatment in intestines of flies at 2 days of age (n = 4 biological replicates of 10 intestines per replicate, two-way ANOVA, interaction, H3 p<0.05, H4 p<0.01; post-hoc test, *p<0.05, **p<0.01, ***p<0.001). (B, C) Adult-onset, enterocyte (EC)-specific knock-down of eIF3d or eIF3g by RNAi alone had no effect on histone expression but blocked increased expression of H3 and H4 in response to rapamycin treatment in intestine of flies at 20 days of age (n = 4 biological replicates of 10 intestines per replicate, two-way ANOVA, interaction, eIF3d RNAi H3 p<0.01, H4 p<0.001; eIF3g RNAi H3 p<0.05, H4 p<0.05; post-hoc test, *p<0.05, **p<0.01, ***p<0.001). (D) Adult-onset, EC-specific knock-down of eIF4e by RNAi alone increased expression of H3 and H4 to the same extent as did rapamycin treatment, with no additional effect of their combination in intestine of flies at 20 days of age (n = 4 biological replicates of 10 intestines per replicate, two-way ANOVA, interaction, H3 and H4 p<0.01; post-hoc test, *p<0.05, **p<0.01, ***p<0.001). The amount of protein was normalised to DNA, shown by stain-free blot.
-
Figure 2—source data 1
Source data pertaining to Figure 2.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig2-data1-v2.xlsx
Figure 2—figure supplement 1
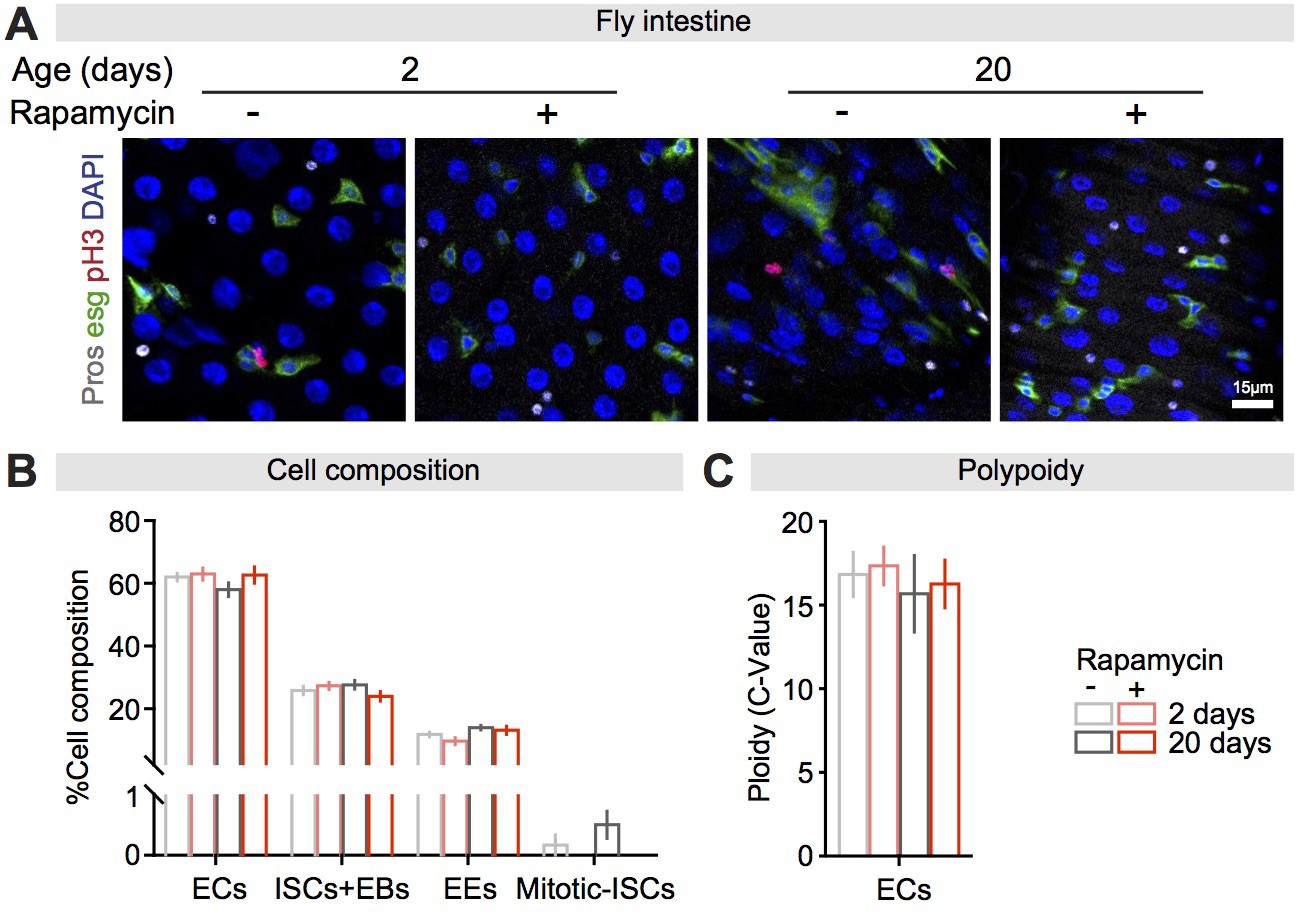
Rapamycin did not affect cell composition or enterocyte (EC) polyploidisation in the fly intestine.
(A) The fly intestine consists of ECs (labelled by DAPI only), intestinal stem cells (ISCs) + enteroblasts (EBs) (esg-GFP positive), enteroendocrine cells (EEs) (Pros positive), and mitotic-ISCs (pH3 positive). (B) Rapamycin treatment did not affect the proportion of the four cell types (EC, ISCs + EB, EE), but reduced ISC mitosis (n = 6 intestines per condition; two-way ANOVA; rapamycin ECs, ISCs + EBs, EEs p>0.05, Mitosis-ISCs, p<0.05). (C) Rapamycin treatment did not affect EC polyploidy (n = 5 intestines per condition; n = 12–15 ECs were observed per intestine, data points represent the average value per intestine; two-way ANOVA; rapamycin, p>0.05).
-
Figure 2—figure supplement 1—source data 1
Source data pertaining to Figure 2—figure supplement 1.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig2-figsupp1-data1-v2.xlsx
Figure 2—figure supplement 2

Perturbation of autophagy or proteosome activity has no effect on expression of core histones in the fly intestine in response to rapamycin treatment.
(A) Adult-onset, knock-down of Atg5 ubiquitously by RNAi had no effect on increased histone expression in response to rapamycin treatment in intestines of flies at 20 days of age (n = 4 biological replicates of 10 intestines per replicate, two-way ANOVA, interaction, H3 and H4 p>0.05; post-hoc test, NS p>0.05, **p<0.01, ***p<0.001). (B) Adult-onset bortezomib treatment (2 µM) had no effect on increased expression of H3 and H4 in response to rapamycin treatment in intestine of flies at 20 days of age (n = 4 biological replicates of 10 intestines per replicate, two-way ANOVA, interaction, H3 and H4 p>0.05; post-hoc test, NS p>0.05, ***p<0.001). The amount of protein was normalised to DNA, shown by stain-free blot.
-
Figure 2—figure supplement 2—source data 1
Source data pertaining to Figure 2—figure supplement 2.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig2-figsupp2-data1-v2.xlsx
Figure 3 with 1 supplement
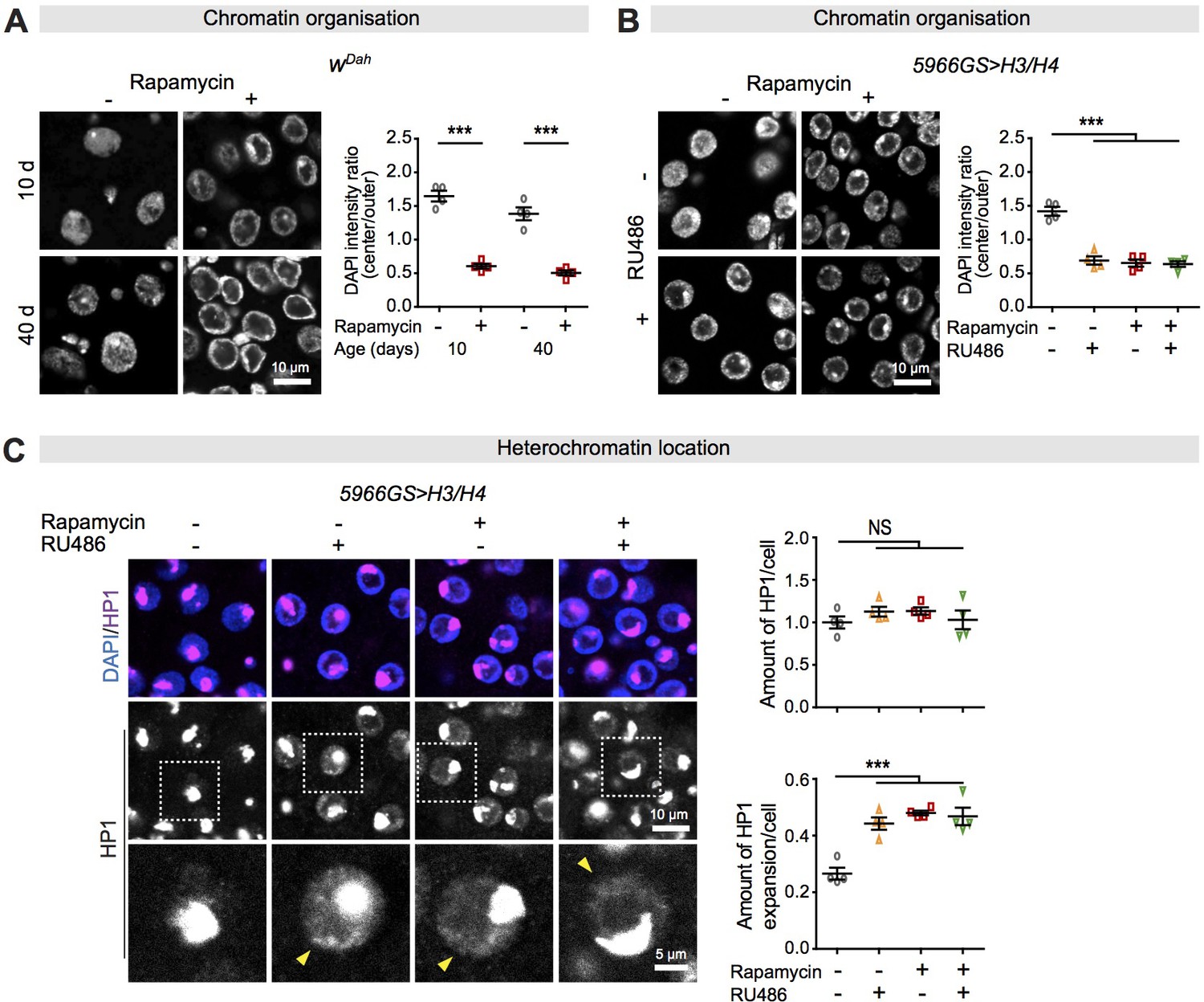
Increased histone expression in response to rapamycin treatment causes chromatin rearrangement and heterochromatin expansion across the nucleus in intestinal enterocytes (ECs).
(A) Rapamycin induced a substantial accumulation of chromatin at the nuclear envelope in ECs (linear mixed model, interaction, p>0.05; post-hoc test, ***p<0.001). (B) Adult-onset, EC-specific expression of H3/H4 by the 5966GS driver recapitulated the effect of rapamycin on the accumulation of chromatin at the nuclear envelope in intestine of flies at 20 days of age (linear mixed model, interaction, p<0.001; post-hoc test, ***p<0.001). (C) Adult-onset, EC-specific expression of H3/H4 by the 5966GS driver had no effect on the total amount of HP1 in the presence or absence of rapamycin (linear mixed model, interaction, p>0.05; post-hoc test, NS p>0.05), but it altered the distribution of HP1 in the nucleus in the intestine of flies at 20 days of age (linear mixed model, interaction, p<0.001; post-hoc test, ***p<0.001). The yellow arrow indicates the expansion of HP1 to the whole nucleus. Each data point (n = 4 intestines) represents an average value from 3 to 5 ECs per intestine.
-
Figure 3—source data 1
Source data pertaining to Figure 3.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig3-data1-v2.xlsx
Figure 3—figure supplement 1
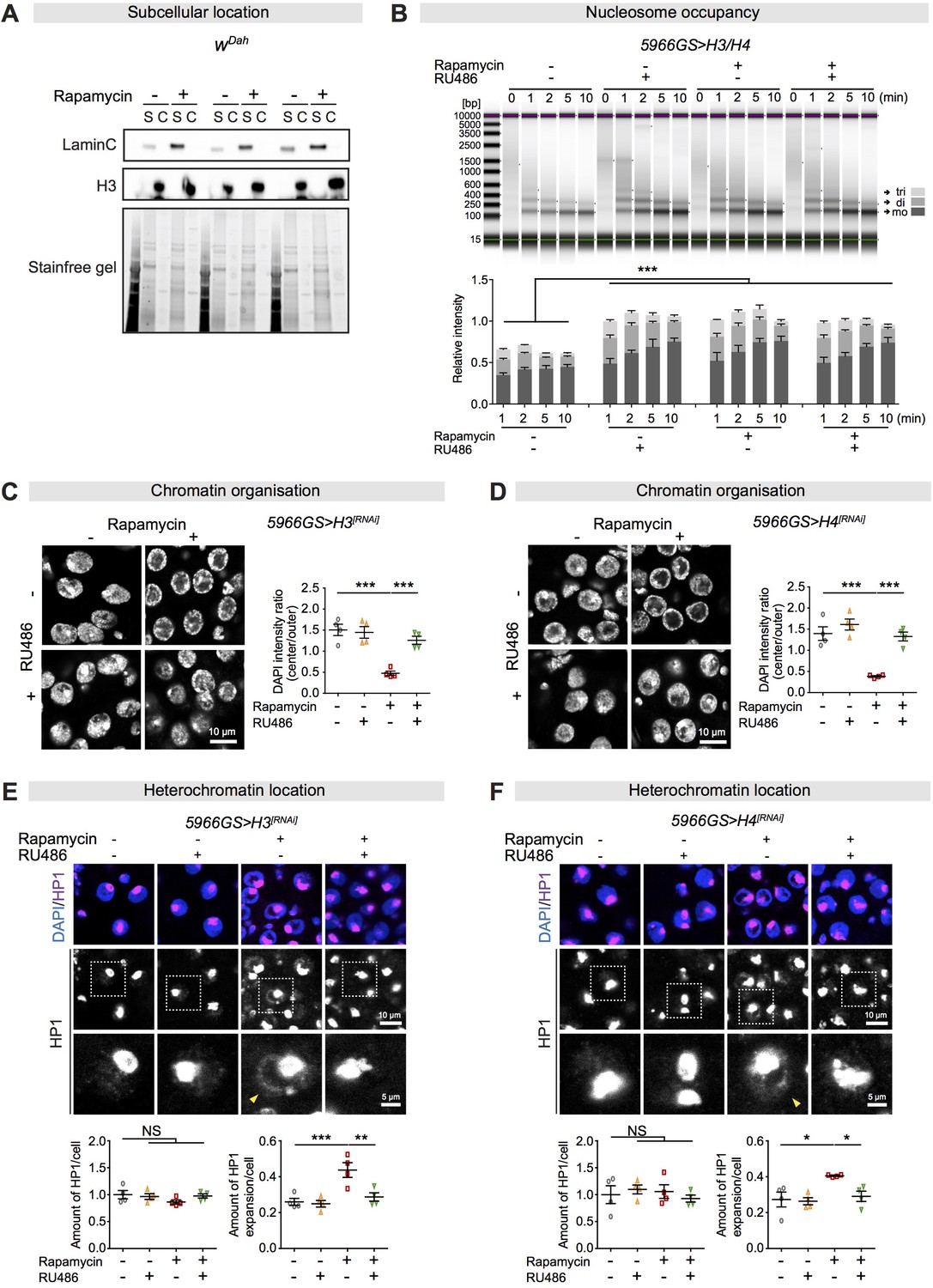
Increased histones are required for rapamycin-induced chromatin rearrangement and heterochromatin expansion in the fly intestine.
(A) Rapamycin treatment did not alter the subcellular location of H3, which was located in the chromatin pellet (C) but not in the soluble cellular components (S) in fly intestine at 10 days of age, in all three biological replicates of 10 intestines per replicate. (B) The number of nucleosomes in the intestine of flies at 20 days of age had increased markedly in response to increased expression of histone in enterocytes (ECs) or upon rapamycin treatment. Gel electrophoresis of time course (0–10 min micrococcal nuclease [MNase] digestions showed the majority of nucleosomes after digestion were trinucleosomes [tri], dinucleosomes [di], and mononucleosomes [mo]). The number of nucleosomes was normalised to input (0 min) (n = 3 biological replicates of 20 intestines per replicate, two-way ANOVA, compared to controls, ***p<0.001). (C, D) Knock-down of H3 or H4 in adult ECs blocked the chromatin rearrangement from rapamycin treatment in fly ECs at 20 days of age (n = 4 intestines per condition; n = 3–5 ECs were observed per intestine, data points represent the average value per intestine; linear mixed model, interaction, p<0.001; post-hoc test, ***p<0.001). (E, F) Knock-down of H3 or H4 in adult ECs had no effect on total amount of HP1 (n = 4 intestines per condition; n = 3–5 ECs were observed per intestine, data points represent the average value per intestine; two-way ANOVA, interaction, p>0.05; post-hoc test, NS p>0.05), but abolished rapamycin-induced HP1 expansion across the nucleus in ECs in fly intestines at 20 days of age (linear mixed model, interaction, p<0.05; post-hoc test, NS p>0.05, *p<0.05, **p<0.01, ***p<0.001). The yellow arrow indicates the expansion of HP1 to the whole nucleus.
-
Figure 3—figure supplement 1—source data 1
Source data pertaining to Figure 3—figure supplement 1.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig3-figsupp1-data1-v2.xlsx
Figure 4

Increased histone expression in adult enterocytes (ECs) mediates lifespan extension and intestinal homeostasis from rapamycin treatment.
(A, B) Rapamycin extended lifespan of control flies (log-rank test, H3RNAi p=3.80E-08; H4RNAi p=2.61E-12), but not of flies with knock-down of H3 or H4 by RNAi in adult ECs (H3RNAi p=0.74; H4RNAi p=0.06). See also Supplementary file 2. (C) Adult-onset expression of H3/H4 in adult ECs extended lifespan (log-rank test, p=0.001) and had no additional effect on lifespan in the presence of rapamycin (Rapamycin vs. Rapamycin + RU, p=0.48). See also Supplementary file 3. (D, E) Knock-down of H3 or H4 in adult ECs by RNAi counteracted the effects of rapamycin on intestinal stem cell (ISC) proliferation in flies at 20 days of age (n = 23–25 intestines, two-way ANOVA, interaction, p>0.05; post-hoc test, *p<0.05, **p<0.01, ***p<0.001). (F) Expression of H3/H4 in adult ECs reduced ISC proliferation in intestine of flies at 20 days of age (n = 23–24 intestines, two-way ANOVA, interaction, p<0.001; post-hoc test, ***p<0.001). (G) Knock-down of H3 in adult ECs by RNAi partially blocked the effects of rapamycin on intestinal dysplasia in flies at 50 days of age (n = 10–12 intestines, two-way ANOVA, interaction, p>0.05; post-hoc test, NS p>0.05, ***p<0.001). (H) Expression of H3/H4 in adult ECs reduced intestinal dysplasia in 50-day-old flies (n = 9–12 intestines, two-way ANOVA, interaction, p<0.01; post-hoc test, ***p<0.001).
-
Figure 4—source data 1
Source data pertaining to Figure 4.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig4-data1-v2.xlsx
Figure 5 with 3 supplements
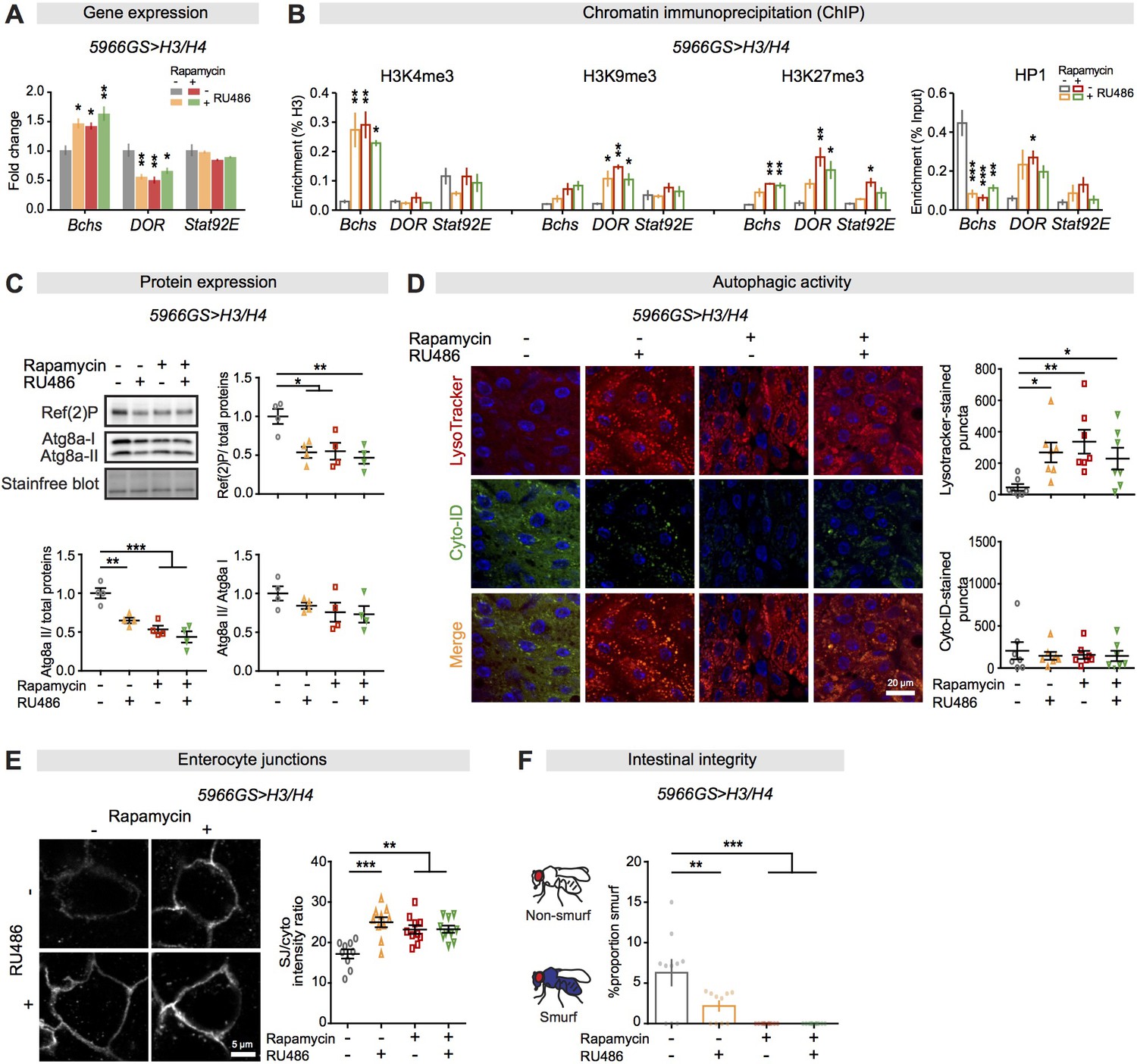
Increased histone expression in enterocytes from rapamycin treatment activates autophagy by altered histone marks and maintains gut barrier function.
(A) Expression of H3/H4 in adult enterocytes (ECs) regulated expression of Bchs and DOR in the intestine of flies at 20 days of age (n = 4 biological replicates of 15 intestines per replicate, two-way ANOVA; post-hoc test, compared to controls, *p<0.05, **p<0.01). (B) Expression of H3/H4 in adult ECs mediated enrichment of H3K4me3, H3K9me3, H3K27me3, and HP1 on Bchs and DOR transcriptional start sites in the intestine of flies at 20 days of age (n = 3 biological replicates of 25 intestines per replicate, two-way ANOVA;post-hoc test, compared to controls, *p<0.05, **p<0.01, ***p<0.001). (C) Expression of H3/H4 in adult ECs decreased the amount of Atg8a-II and Ref(2)P (n = 4 biological replicates of 10 intestines per replicate, two-way ANOVA, interaction, p<0.05; post-hoc test, *p<0.05, **p<0.01, ***p<0.001). (D) Expression of H3/H4 in adult ECs substantially increased the number of LysoTracker-stained puncta and had no effect on the number of Cyto-ID-stained puncta in the intestine (n ≥ 6 intestines per condition; n = 2–3 pictures per intestine, data points represent the average value per intestine; linear mixed model, interaction, LysoTracker-stained puncta p<0.01; post-hoc test, *p<0.05, **p<0.01). (E) Expression of H3/H4 in adult ECs improved maintenance of coracle at septate junctions between ECs in the intestine of flies at 50 days of age. The ratio of septate junction (SJ)/cytoplasm fluorescence for coracle was high in the intestine of flies fed RU486 and/or rapamycin (n ≥ 9 intestines per condition; n = 3–5 ECs were observed per intestine, linear mixed model, interaction, p<0.01; post-hoc test, **p<0.01, ***p<0.001). (F) The number of Smurfs was significantly reduced in response to increased expression of H3/H4 in ECs and/or rapamycin at 60 days of age. Bar charts with n = 10 biological replicates of 15–20 flies per replicate (two-way ANOVA, interaction, p<0.05; post-hoc test, **p<0.01, ***p<0.001).
-
Figure 5—source data 1
Source data pertaining to Figure 5.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig5-data1-v2.xlsx
Figure 5—figure supplement 1
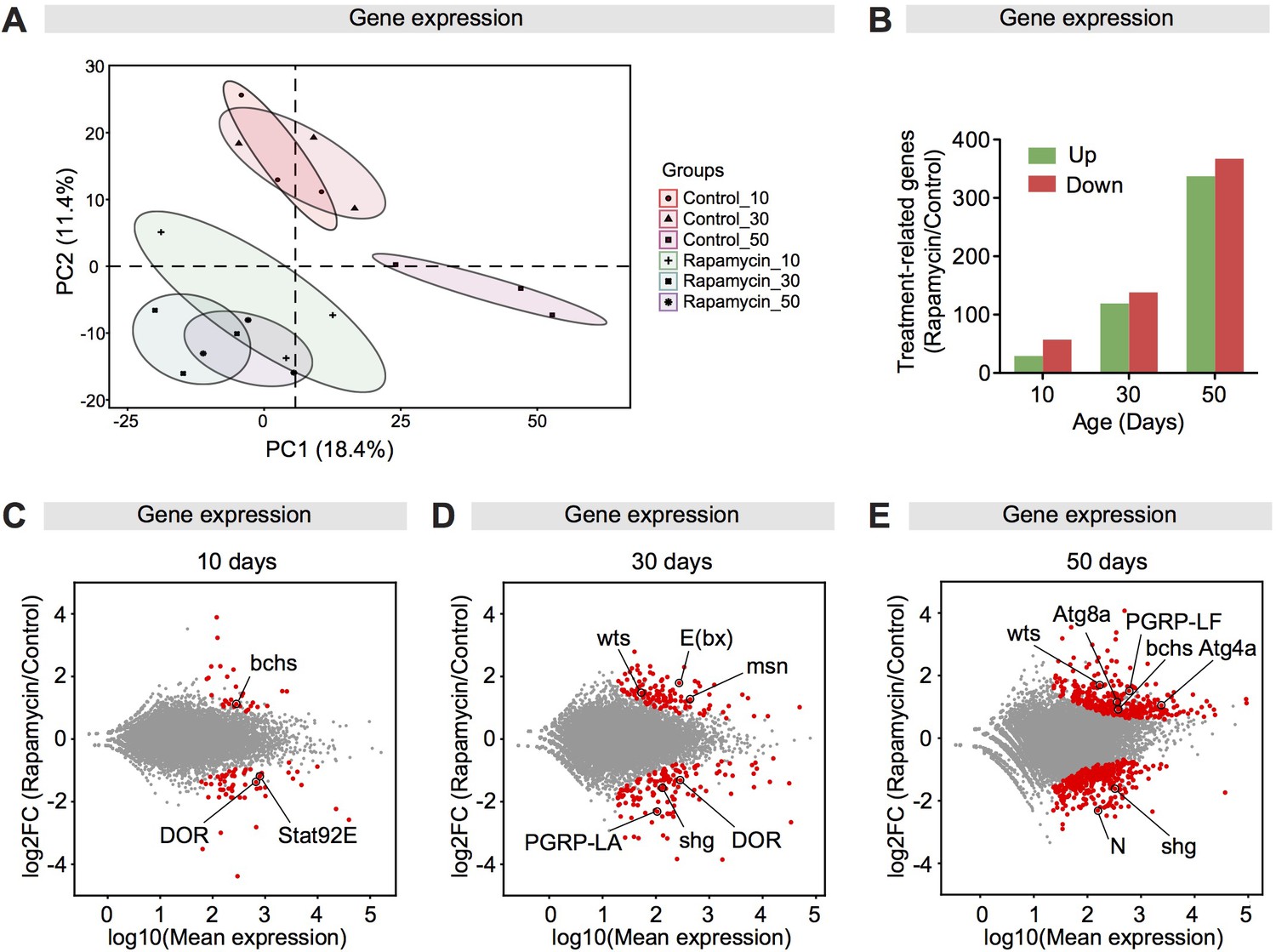
Rapamycin meditates a transcriptional response of autophagy-related genes in the fly intestine.
(A) Rapamycin induced a global transcriptional change. (B) The number of significantly up-regulated and down-regulated genes in intestines of rapamycin-fed flies compared to controls at 10, 30, and 50 days of age. (C–E) MA-Plots showed the log2 fold change of gene expression levels in intestines of rapamycin-fed flies compared to controls at 10, 30, and 50 days of age. Genes showing significantly differential expression between rapamycin groups and control groups are marked in red. Autophagy-related genes are highlighted.
-
Figure 5—figure supplement 1—source data 1
Source data pertaining to Figure 5—figure supplement 1.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig5-figsupp1-data1-v2.xlsx
Figure 5—figure supplement 2
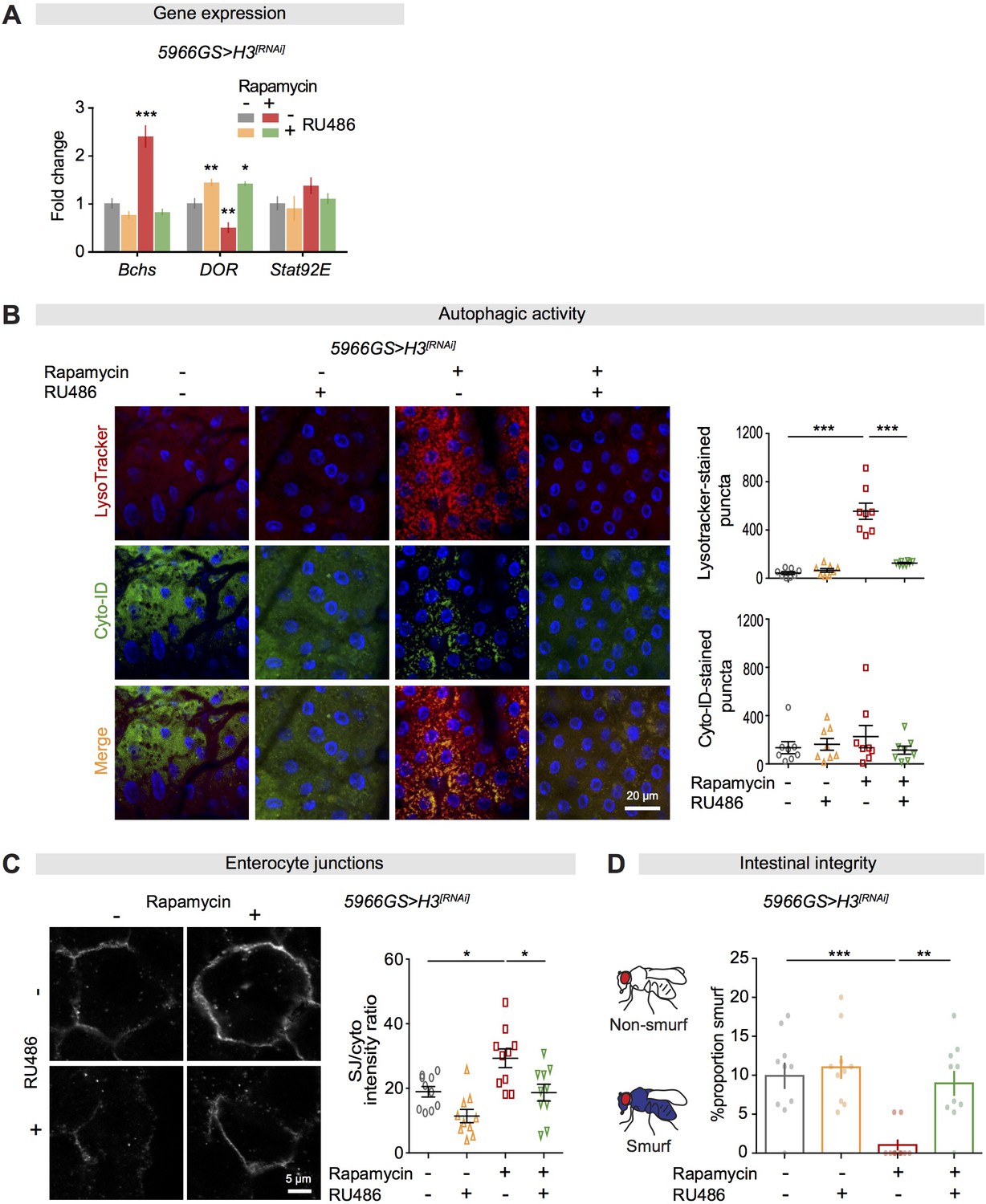
Increased histone expression in enterocytes is required for activation of autophagy and maintenance of gut barrier function from rapamycin treatment.
(A) Knock-down of H3 in adult enterocytes (ECs) abolished the effect of rapamycin on expression of Bchs and DOR in the intestine of flies at 20 days of age (n = 4 biological replicates of 15 intestines per replicate, two-way ANOVA; post-hoc test, compared to controls, *p<0.05, **p<0.01, ***p<0.001). (B) Knock-down of H3 in adult ECs abolished the effect of rapamycin on the number of LysoTracker-stained puncta, but had no effect on the number of Cyto-ID-stained puncta (n = 8 intestines per condition; n = 2–3 pictures per intestine, data points represent the average value per intestine; linear mixed model, interaction, LysoTracker-stained puncta p<0.001; post-hoc test, ***p<0.001). (C) Knock-down of H3 in adult ECs abolished the effect of rapamycin on septate junctions between ECs in the intestine of flies at 50 days of age. The ratio of SJ/cytoplasm fluorescence for coracle was high in the intestine of flies fed rapamycin compared to other treatments (n = 10 intestines per condition; n = 3–5 ECs were observed per intestine, linear mixed model, interaction, p>0.05; post-hoc test, *p<0.05). (D) Knock-down of H3 in adult ECs abolished the effect of rapamycin on proportion of Smurfs at 60 days of age. Bar charts with n = 10 biological replicates of 15–20 flies per replicate (two-way ANOVA, interaction, p<0.05; post-hoc test, **p<0.01, ***p<0.001).
-
Figure 5—figure supplement 2—source data 1
Source data pertaining to Figure 5—figure supplement 2.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig5-figsupp2-data1-v2.xlsx
Figure 5—figure supplement 3
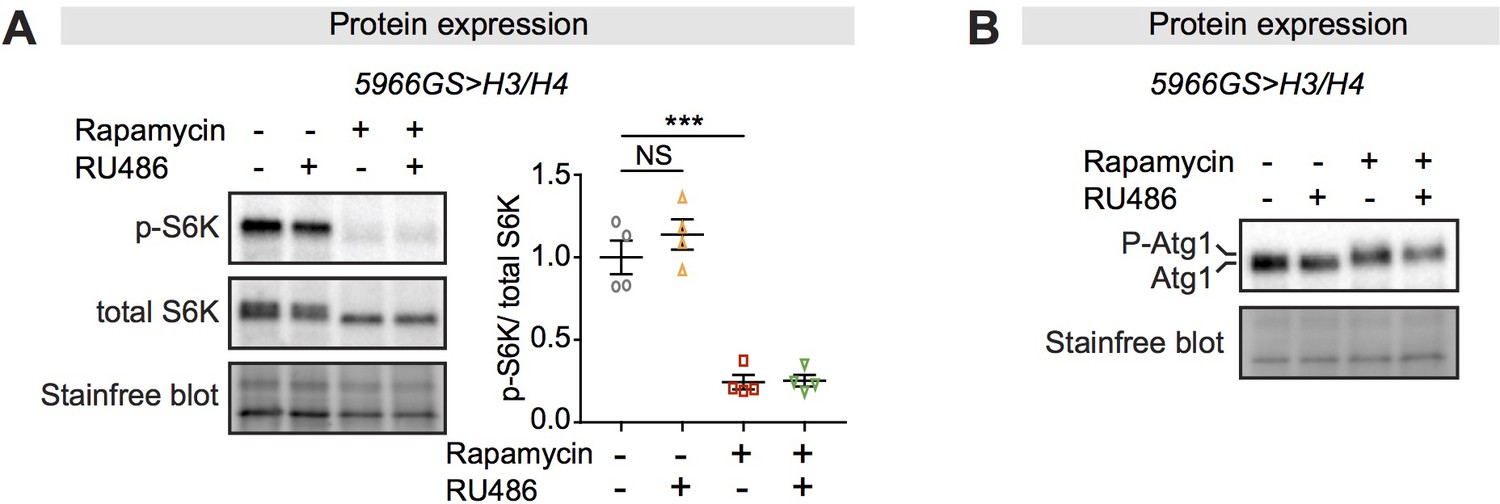
Increased histone expression does not affect mTORC1 activity.
(A) The level of phospho-S6K in intestines was unaffected by increased expression of H3/H4 in adult enterocytes (ECs), while it was substantially reduced by rapamycin treatment (n = 4 biological replicates of 10 intestines per replicate, two-way ANOVA, interaction, p>0.05; post-hoc test, NS p>0.05, ***p<0.001). (B) A slower-migrating form of Atg1 appeared only in the presence of rapamycin (n = 4 biological replicates of 10 intestines per replicate).
-
Figure 5—figure supplement 3—source data 1
Source data pertaining to Figure 5—figure supplement 3.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig5-figsupp3-data1-v2.xlsx
Figure 6 with 1 supplement
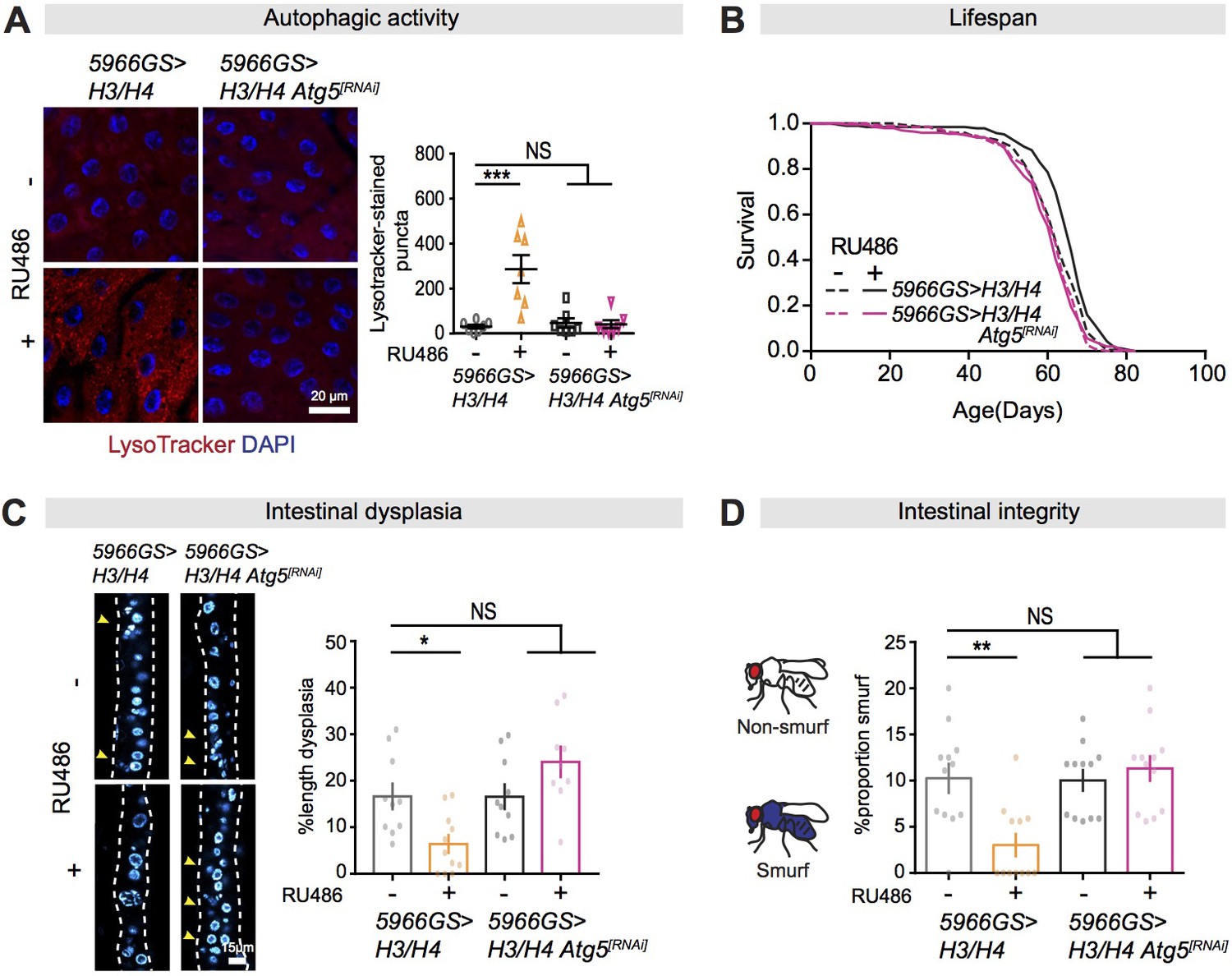
Autophagy activation is necessary for mTORC1-histone axis on survival and intestinal homeostasis.
(A) Knock-down of Atg5 abolished the effect of expression of H3/H4 in enterocytes (ECs) on induction of LysoTracker-stained puncta in the intestine of flies at 20 days of age (n ≥ 6 intestines per condition; n = 2–3 images per intestine, data points represent the average value per intestine; linear mixed model, interaction, p<0.001; post-hoc test, ***p<0.001). (B) Knock-down of Atg5 abolished the increase in lifespan in response to expression of H3/H4 in adult ECs. 5966GS>H3/H4 females showed increased lifespan in the presence of RU486 (log-rank test, p=0.0001), but 5966GS>H3/H4 Atg5[RNAi] females did not (p=0.49). See also Supplementary file 4. (C) Knock-down of Atg5 blocked the effect of expression of H3/H4 in adult ECs on intestinal dysplasia at 50 days of age (n = 9–12 intestines, two-way ANOVA, interaction, p<0.01; post-hoc test, NS p>0.05, ***p<0.001). (D) Knock-down of Atg5 abolished the effects of expression of H3/H4 in adult ECs on gut integrity at 60 days of age. Bar charts with n = 12 biological replicates of 15–20 flies per replicate (two-way ANOVA, interaction, p<0.01; post-hoc test, **p<0.01, ***p<0.001).
-
Figure 6—source data 1
Source data pertaining to Figure 6.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig6-data1-v2.xlsx
Figure 6—figure supplement 1
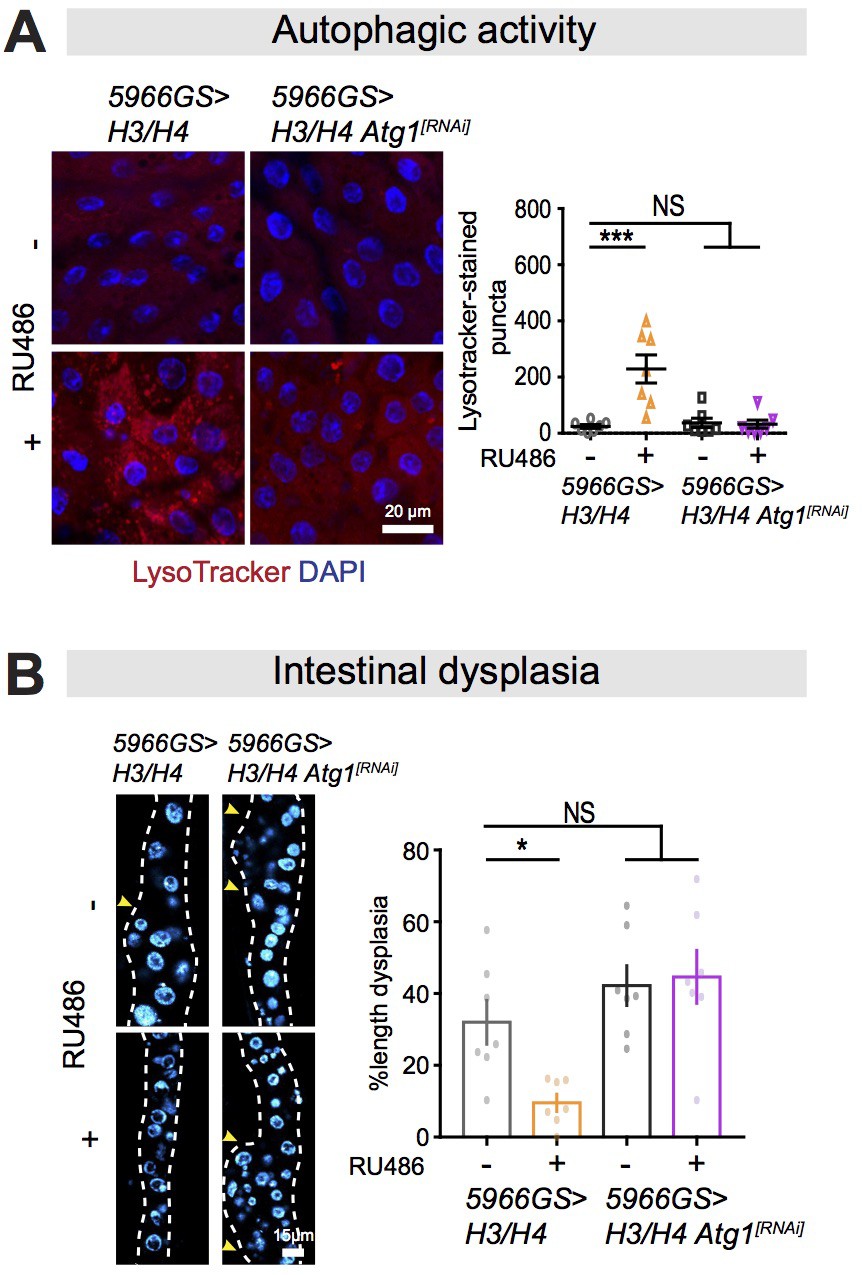
Autophagy activation is required for mTORC1-histone axis on survival and intestinal homeostasis.
(A) Knock-down of Atg1 abolished the effect of expression of H3/H4 in enterocytes (ECs) on induction of LysoTracker-stained puncta in the intestine of flies at 20 days of age (n = 7 intestines per condition; n = 2–3 images per intestine, data points represent the average value per intestine; linear mixed model, interaction, p<0.01; post-hoc test, NS p>0.05, ***p<0.001). (B) Knock-down of Atg1 blocked the effect of expression of H3/H4 in adult ECs on intestinal dysplasia at 50 days of age (n = 7 intestines, two-way ANOVA, interaction, p<0.05; post-hoc test, NS p>0.05, *p<0.05).
-
Figure 6—figure supplement 1—source data 1
Source data pertaining to Figure 6—figure supplement 1.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig6-figsupp1-data1-v2.xlsx
Figure 7 with 2 supplements
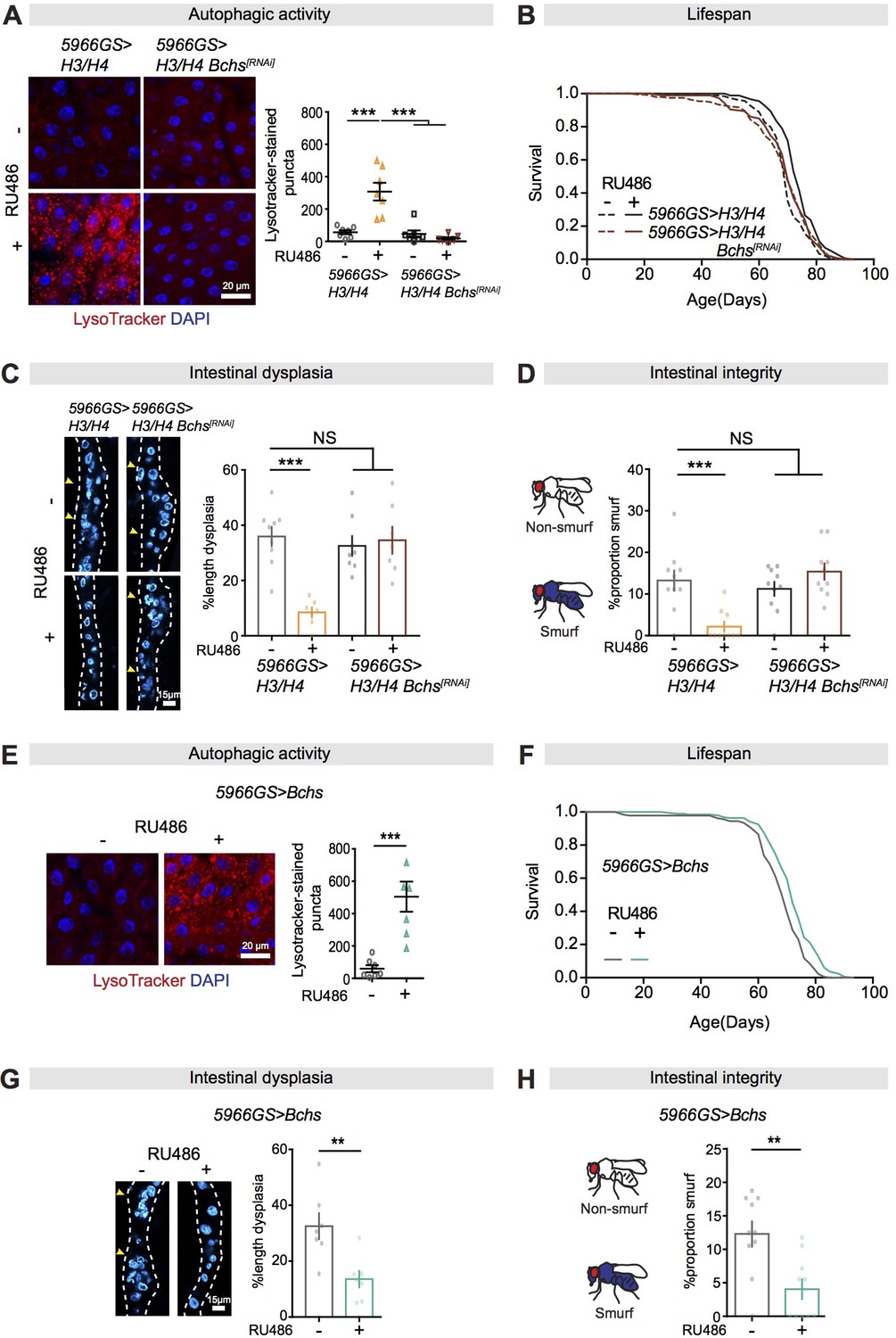
Bchs is a required target for autophagy activation, lifespan extension, and intestinal homeostasis from the mTORC1-histone axis.
(A) Knock-down of Bchs abolished the effect of expression of H3/H4 in enterocytes (ECs) on induction of LysoTracker-stained puncta in the intestine of flies at 20 days of age (n = 7 intestines per condition; n = 3 images per intestine, data points represent the average value per intestine; linear mixed model, interaction, p<0.001; post-hoc test, ***p<0.001). (B) Knock-down of Bchs abolished the effects of expression of H3/H4 in adult ECs on lifespan. 5966GS>H3/H4 females showed increased lifespan in the presence of RU486 (log-rank test, p=7.55E-08), but 5966GS>H3/H4 Bchs[RNAi] females did not (p=0.90). See also Supplementary file 5. (C) Knock-down of Bchs blocked the effect of expression of H3/H4 in adult ECs on intestinal dysplasia at 50 days of age (n = 7–9 intestines, two-way ANOVA, interaction, p<0.001; post-hoc test, ***p<0.001). (D) Knock-down of Bchs abolished the beneficial effects of expression of H3/H4 in adult ECs on gut integrity at 60 days of age. Bar charts with n = 10 biological replicates of 15–20 flies per replicate (two-way ANOVA, interaction, p<0.001; post-hoc test, **p<0.01, ***p<0.001). (E, F) Expression of Bchs in adult ECs substantially increased the number of LysoTracker-stained puncta in the intestine (n = 7 intestines per condition; n = 3 images per intestine, data points represent the average value per intestine; linear mixed model, ***p<0.001) and extended lifespan (log-rank test, p=4.92E-06). See also Supplementary file 7. (G) Expression of Bchs in adult ECs reduced intestinal dysplasia in 50-day-old flies (n = 7 intestines, Student's t test, **p<0.01). (H) The proportion of Smurfs at 60 days of age was significantly reduced in response to increased expression of Bchs in ECs and/or rapamycin treatment. Bar charts with n = 10 biological replicates of 15–20 flies per replicate (Student's t test, **p<0.01).
-
Figure 7—source data 1
Source data pertaining to Figure 7.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig7-data1-v2.xlsx
Figure 7—figure supplement 1
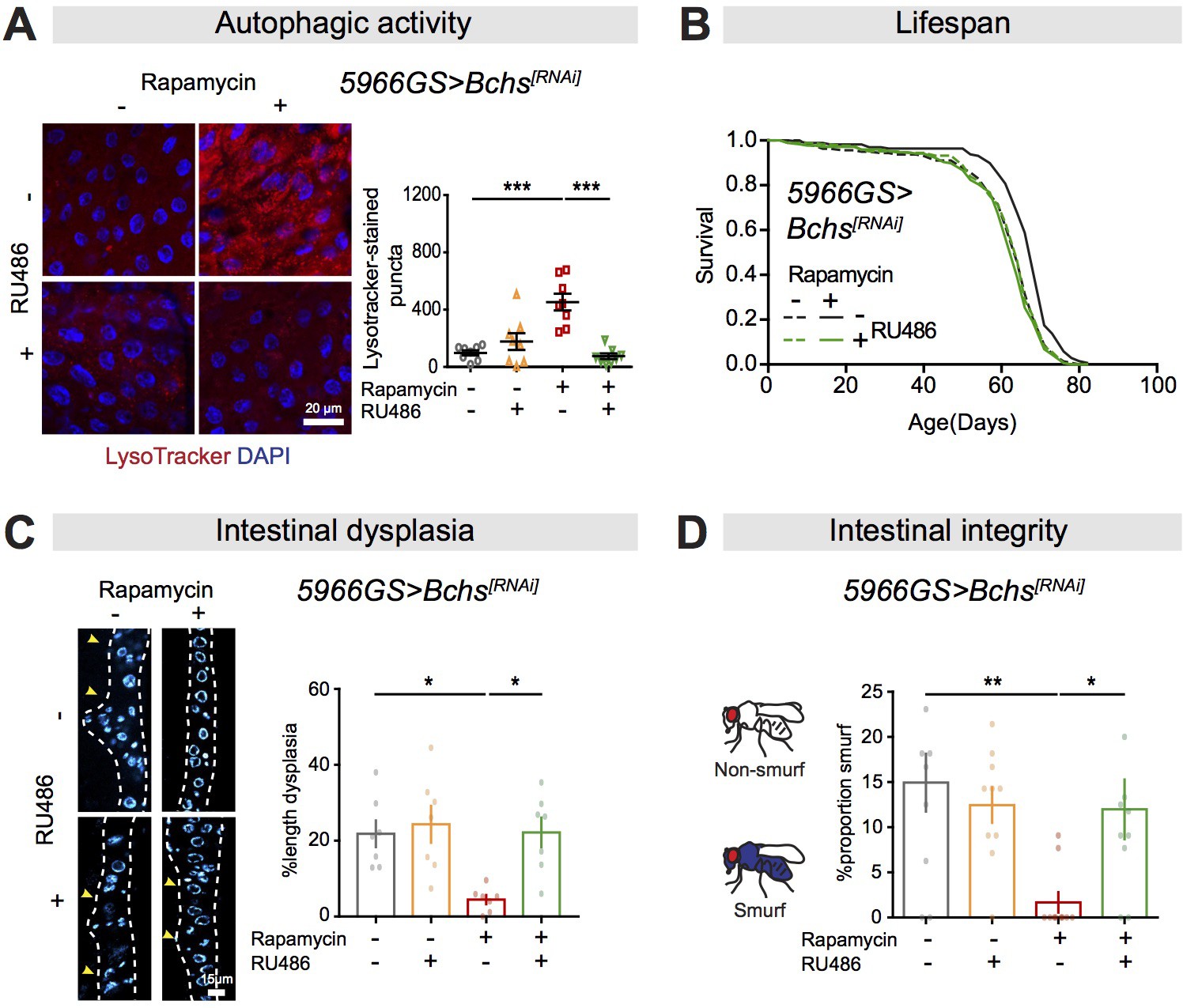
Bchs is a required autophagic target for rapamycin-induced lifespan extension and intestinal homeostasis.
(A) Knock-down of Bchs abolished the effect of rapamycin on induction of LysoTracker-stained puncta in the intestine of flies at 20 days of age (n = 8 intestines per condition; n = 3 images per intestine, data points represent the average value per intestine; linear mixed model, interaction, p<0.001; post-hoc test, ***p<0.001). (B) Rapamycin extended lifespan of control flies (log-rank test, p=7.67E-07), but not of flies with knock-down of Bchs in adult enterocytes (ECs) (log-rank test, p=0.34). See also Supplementary file 6. (C) Knock-down of Bchs in adult ECs abolished the effects of rapamycin on intestinal dysplasia in flies at 50 days of age (n = 7 intestines, two-way ANOVA, interaction, p<0.05; post-hoc test, *p<0.05). (D) Knock-down of Bchs in adult ECs abolished the effect of rapamycin on proportion of Smurfs at 60 days of age. Bar charts with n = 10 biological replicates of 15–20 flies per replicate (two-way ANOVA, interaction, p<0.05; post-hoc test, *p<0.05, **p<0.01).
-
Figure 7—figure supplement 1—source data 1
Source data pertaining to Figure 7—figure supplement 1.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig7-figsupp1-data1-v2.xlsx
Figure 7—figure supplement 2

Manipulated Bchs expression does not influence mTORC1-dependent phosphorylation of Atg1.
(A) Knock-down of Bchs did not abolish the effect of rapamycin on the induction of a slower-migrating form of Atg1 (n = 4 biological replicates of 10 intestines per replicate). (B) Over-expression of Bchs did not lead to a slower-migrating form of Atg1 (n = 4 biological replicates of 10 intestines per replicate).
Figure 8
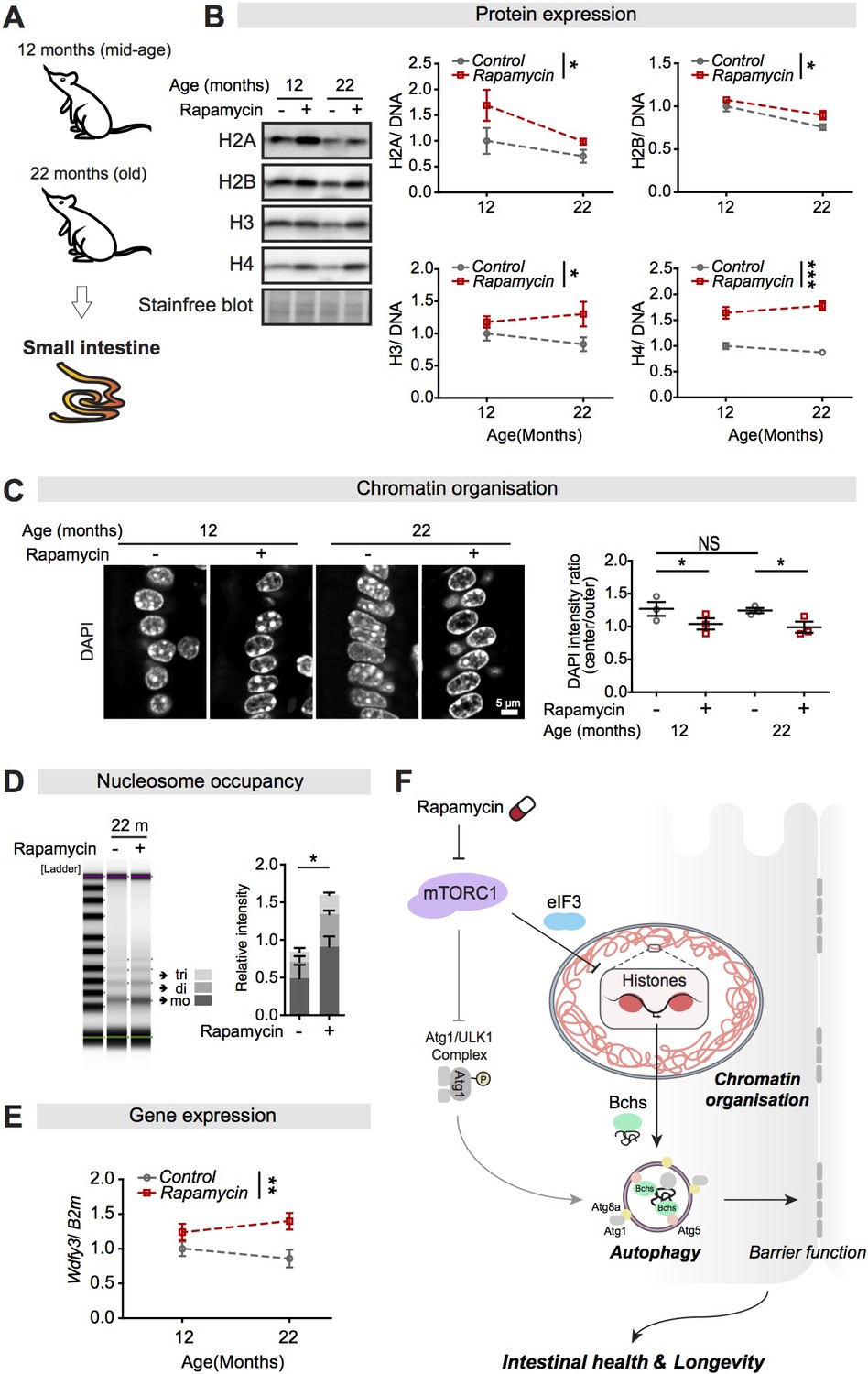
Rapamycin treatment upregulates expression of histones and Wdfy3 transcript, alters chromatin structure, and increases the number of nucleosomes in the small intestine of mice.
(A) Female mice were sacrificed at 12 months and 22 months of age, and the jejunum of the small intestine was dissected. (B) Rapamycin substantially increased expression of H2A, H2B, H3, and H4 compared to controls in the small intestine of mice (n = 3 jejunums, two-way ANOVA; treatment *p<0.05; ***p<0.001). The amount of protein was normalised to DNA, shown by stain-free blot. (C) Rapamycin induced a substantial accumulation of chromatin at the nuclear envelope in cells in villi of the small intestine of mice at 12 months and 22 months of age (n = 3 jejunums per condition; n = 40–45 cells were observed per intestine, linear mixed model; post-hoc test, NS p>0.05, *p<0.05). (D) The number of nucleosomes in the intestine increased markedly in response to rapamycin treatment in mice at 22 months of age. Gel electrophoresis of 5 min micrococcal nuclease (MNase) digestions showed that the majority of nucleosomes after digestion were trinucleosomes (tri), dinucleosomes (di), and mononucleosomes (mo). The number of nucleosomes was normalised to input (0 min) (n = 3 jejunums, two-way ANOVA, post-hoc test, *p<0.05). (E) Rapamycin substantially increased Wdfy3 in the small intestine of mice compared to controls at 22 months of age (n = 3 jejunums, mean ± SEM, two-way ANOVA; treatment **p<0.01). (F) Model of the relationship linking mTORC1, histones, autophagy, and longevity.
-
Figure 8—source data 1
Source data pertaining to Figure 8.
- https://cdn.elifesciences.org/articles/62233/elife-62233-fig8-data1-v2.xlsx
Author response image 1
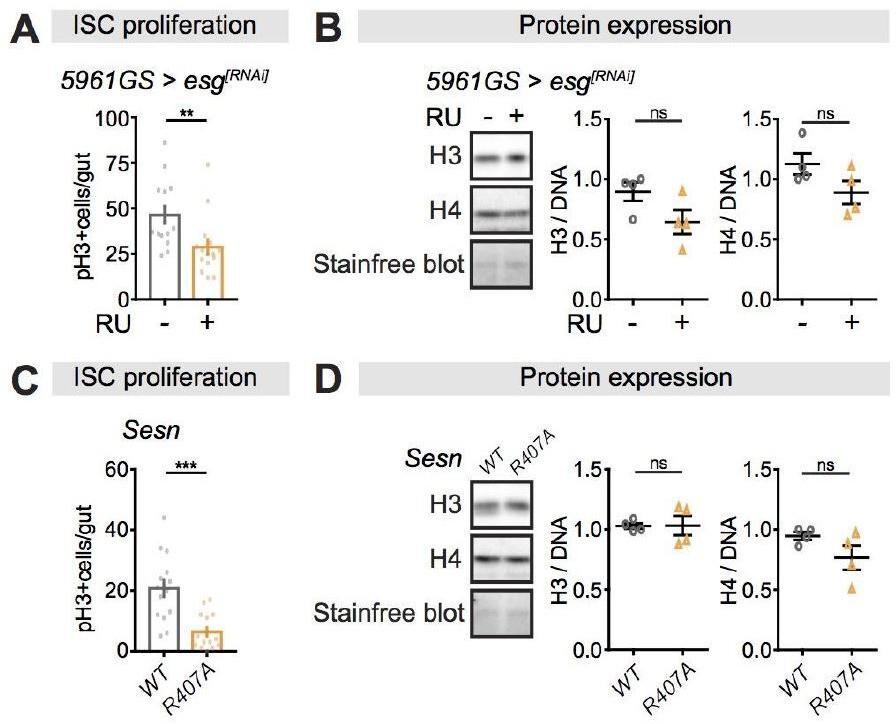
Perturbations of ISC proliferation via Escargot or Sestrin activity have no effect on histone protein levels in the fly intestine.
(A-B) Adult-onset, ISC-specific knock-down of escargot by RNAi reduced ISC proliferation in intestine of flies (n = 15 intestines, Students t test, **p<0.01), but it had not effect on the expression of H3 and H4 at 20 days of age. (n = 4 biological replicates of 10 intestines per replicate, Students t test, ns p>0.05). The amount of proteins was normalized to DNA, shown by stain-free blot. (C-D) Sestrin mutant flies (SesnR407A) showed lower ISC proliferation in the intestine compared to wild type flies (n = 15 intestines, Students t test, ***p<0.001), but it had not effect on the expression of H3 and H4 at 20 days of age. (n = 4 biological replicates of 10 intestines per replicate, Students t test, ns p>0.05). The amount of proteins was normalized to DNA, shown by stain-free blot.
Author response image 2
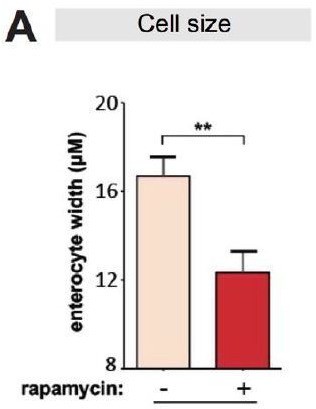
Rapamycin reduces cell size of enterocytes.
(A) Cell size of enterocytes in wDah females significantly reduced upon rapamycin treatment at 28 days of age. (n = 10 intestines, n = 10-20 ECs were observed per intestine, Student t-test, **p<0.01).
Tables
Appendix 1—key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Antibody | Histone H2A antibody (rabbit polyclonal) | Active Motif | Cat# 39209 RRID:AB_2793184 | WB (1:1000) |
| Antibody | Anti-histone H2B antibody – ChIP Grade (mouse monoclonal) | Abcam | Cat# ab52484 RRID:AB_1139809 | WB (1:10,000) |
| Antibody | Anti-histone H3 antibody – ChIP Grade (rabbit polyclonal) | Abcam | Cat# ab1791 RRID:AB_302613 | WB (1:10,000) ChIP (2 µg) |
| Antibody | Histone H4 antibody (pAb) (rabbit polyclonal) | Active Motif | Cat# 39269 RRID:AB_2636967 | WB (1:3000) |
| Antibody | Lamin C (mouse monoclonal) | Developmental Studies Hybridoma Bank | Cat# LC28.26 RRID:AB_528339 | WB (1:3000) |
| Antibody | Atg1 (rabbit polyclonal) | From Jun Hee Lee’s lab, USA | WB (1:2000) | |
| Antibody | Atg8a (rabbit polyclonal) | From Péter Nagy’s lab, Hungary | WB (1:3000) | |
| Antibody | Phospho-Drosophila p70 S6 Kinase (Thr398) (rabbit polyclonal) | Cell Signaling | Cat#9209 RRID:AB_2269804 | WB (1:1000) |
| Antibody | Total S6K (rabbit polyclonal) | Self-made | WB (1:1000) | |
| Antibody | Anti-histone H3 (trimethyl K4) antibody – ChIP Grade (rabbit polyclonal) | Abcam | Cat# ab8580 RRID:AB_306649 | ChIP (2 µg) |
| Antibody | Anti-histone H3 (trimethyl K9) antibody – ChIP Grade (rabbit polyclonal) | Abcam | Cat# ab8898 RRID:AB_306848 | ChIP (2 µg) |
| Antibody | Anti-trimethyl-histone H3 (Lys27) Antibody (rabbit polyclonal) | Merck Millipore | Cat# 07-449 RRID:AB_310624 | ChIP (2 µg) |
| Antibody | HP1 (mouse monoclonal) | Developmental Studies Hybridoma Bank | Cat# C1A9 RRID:AB_528276 | IF (1:500) |
| Antibody | Mouse (G3A1) mAb IgG1 Isotype Control (mouse monoclonal) | Cell Signaling | Cat# 5415 RRID:AB_10829607 | ChIP (2 µg) |
| Antibody | Rabbit IgG, polyclonal - Isotype Control (rabbit polyclonal) | Abcam | Cat# ab171870 RRID:AB_2687657 | ChIP (2 µg) |
| Antibody | Coracle (mouse monoclonal) | Developmental Studies Hybridoma Bank | Cat# C615.16 RRID:AB_1161644 | IF (1:100) |
| Antibody | Phospho-Histone H3 (Ser10) (rabbit polyclonal) | Cell Signaling | Cat# 9701 RRID:AB_331535 | IF (1:500) |
| Antibody | Lamin A/C (rabbit polyclonal) | Cell Signaling | Cat# 2032 RRID:AB_2136278 | IF (1:500) |
| Antibody | Prospero (mouse monoclonal) | DSHB | Cat# MR1A RRID:AB_528440 | IF (1:200) |
| Antibody | Goat anti-Mouse IgG (H+L) Cross-Adsorbed Secondary Antibody, Alexa Fluor 488 | Thermo Fisher Scientific | Cat# A11001 RRID:AB_2534069 | IF (1:1000) |
| Antibody | Goat anti-Rabbit IgG (H+L) Cross-Adsorbed Secondary Antibody, Alexa Fluor 594 | Thermo Fisher Scientific | Cat# A11012 RRID:AB_2534079 | IF (1:1000) |
| Chemical compound, drug | Rapamycin | LC Laboratories | Cat# R-5000 | |
| Chemical compound, drug | RU486 (Mifepristone) | Sigma | Cat# M8046 | |
| Chemical compound, drug | Cycloheximide | Sigma | Cat# 239763 | |
| Chemical compound, drug | Bortezomib | Sigma | Cat# 5043140001 | |
| Chemical compound, drug | Brilliant Blue FCF | Sigma | Cat# 80717 | |
| Commercial assay or kit | TRIzol Reagent | Invitrogen | Cat#15596018 | |
| Commercial assay or kit | SuperScript III First Strand Master Mix | Invitrogen | Cat#15596018 | |
| Commercial assay or kit | High Sensitivity D5000 ScreenTape | Agilent Technologies | Cat# 5067-5593 | |
| Commercial assay or kit | VECTASHIELD Antifade Mounting Medium | Vector Laboratories | Cat# H-1000 RRID:AB_2336789 | |
| Commercial assay or kit | RIPA Lysis and Extraction Buffer | Thermo Scientific | Cat# 89901 | |
| Commercial assay or kit | 4–20% Criterion TGX Stain-Free Protein Gel, 26 well, 15 µl | Bio-Rad | Cat# 5678095 | |
| Commercial assay or kit | Amersham Hybond P Western blotting membranes, PVDF | GE Healthcare Amersham | Cat# GE10600023 | |
| Commercial assay or kit | PhosSTOP | Roche | Cat# 04906837001 | |
| Commercial assay or kit | cOmplete, Mini, EDTA-free Protease Inhibitor Cocktail | Roche | Cat# 11836170001 | |
| Commercial assay or kit | Qubit dsDNA HS Assay Kit | Invitrogen | Cat# Q32854 | |
| Commercial assay or kit | Qubit dsRNA HS Assay Kit | Invitrogen | Cat# Q32852 | |
| Commercial assay or kit | Pierce BCA Protein Assay Kit | Thermo Scientific | Cat# 23227 | |
| Commercial assay or kit | Power SYBR Green PCR Master Mix | Applied Biosystems | Cat# 4367659 | |
| Commercial assay or kit | TaqMan Gene Expression Master Mix | Applied Biosystems | Cat# 4369016 | |
| Commercial assay or kit | EZ Nucleosomal DNA Prep Kit | Zymo Research | Cat# D5220 | |
| Commercial assay or kit | LysoTracker Red DND-99 | Invitrogen | Cat# L7528 | |
| Commercial assay or kit | Cyto-ID Autophagy detection kit 2.0 | Enzo Life Sciences | Cat# ENZ-KIT175 | |
| Strain, strain background (Drosophila melanogaster) | wDah | This lab | ||
| Genetic reagent (D. melanogaster) | DaGS | Bloomington Drosophila Stock Center | 8641 | |
| Genetic reagent (D. melanogaster) | 5966GS | Guo et al., 2014 | ||
| Genetic reagent (D. melanogaster) | esgGal4 | Kyoto Stock Center | 104863 | |
| Genetic reagent (D. melanogaster) | UAS-H3[RNAi] | Vienna Drosophila Resource Center | KK109374 | |
| Genetic reagent (D. melanogaster) | UAS-H4[RNAi] | Vienna Drosophila Resource Center | KK109059 | |
| Genetic reagent (D. melanogaster) | UAS-Bchs[RNAi] | Vienna Drosophila Resource Center | KK110785 | |
| Genetic reagent (D. melanogaster) | UAS-Bchs.HA | Bloomington Drosophila Stock Center | 51636 | |
| Genetic reagent (D. melanogaster) | UAS-Atg5[RNAi] | Scott et al., 2004; Ren et al., 2009 | ||
| Genetic reagent (D. melanogaster) | UAS-Atg1[RNAi] | Bloomington Drosophila Stock Center | 26731 | |
| Genetic reagent (D. melanogaster) | UAS-eIF3d[RNAi] | Vienna Drosophila Resource Center | 330545 | |
| Genetic reagent (D. melanogaster) | UAS-eIF3g[RNAi] | Vienna Drosophila Resource Center | GD28937 | |
| Genetic reagent (D. melanogaster) | UAS-eIF4e[RNAi] | Vienna Drosophila Resource Center | GD7800 | |
| Genetic reagent (D. melanogaster) | UAS-H3/H4 | This lab | ||
| Genetic reagent (D. melanogaster) | UAS-mCD8::GFP.L | Bloomington Drosophila Stock Center | 5137 | |
| Strain, strain background (mouse) | UM-HET3 stock (CByB6F1 × C3D2F1) | This lab | ||
| Sequence-based reagent | Primer for Q-RT-PCR Act5C_F1: AGGCCAACCGTGAGAAGATG | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR Act5C_R1: GGGGAAGGGCATAACCCTC | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR His3_F1: CCACGCAAACAACTGGCTAC | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR His3_R1: TGCGGATTAGAAGCTCGGTG | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR His4_F1: CGGATAGCAGGCTTCGTGAT | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR His4_R1: GGTCGTGGTAAAGGAGGCAA | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR Bchs_F1: AGCCTCACCACGCTAAAGAAG | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR Bchs_R1: CTCATGTCGTTTGACGGACAG | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR DOR_F1: CTTGATCTCGGGGTGTCGAC | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR DOR_R1: CTTCAACTGTACGGCCGCAT | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR Stat92_F1: AAGCTGCTTGCCCAAAACTAC | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR Stat92_R1: GACGCATTGTGAGTACGATGG | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR B2m_Mouse_F1: TTCTGGTGCTTGTCTCACTGA | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR B2m_Mouse_R1: CAGTATGTTCGGCTTCCCATTC | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR Wdfy3_Mouse_F1: GAGGCTCTGGAGTGTGATTACG | This paper | ||
| Sequence-based reagent | Primer for Q-RT-PCR Wdfy3_Mouse_R1: GTGGCCGTCTCCTTCAGTG | This paper | ||
| Sequence-based reagent | Primer for ChIP-Q-PCR Bchs_ChIP_F1: AGAACAGCTGTCTCGCACAA | This paper | ||
| Sequence-based reagent | Primer for ChIP-Q-PCR Bchs_ChIP_R1: CCTACATAGCGAGCAAGCGA | This paper | ||
| Sequence-based reagent | Primer for ChIP-Q-PCR DOR_ChIP_F1: ATTCGCTCGTCAGTCGTTGT | This paper | ||
| Sequence-based reagent | Primer for ChIP-Q-PCR DOR_ChIP_R1: TAACTGACGGGGGTGAGAGT | This paper | ||
| Sequence-based reagent | Primer for ChIP-Q-PCR Stat92E_ChIP_F1: AAGCGATCCACATGCGATACT | This paper | ||
| Sequence-based reagent | Primer for ChIP-Q-PCR Stat92E_ChIP_R1: TCCTATCTTCCCGGTTTGGC | This paper | ||
| Recombinant DNA reagent | Plasmid: pUAST attb H3 | This paper | ||
| Software, algorithm | Microsoft Excel | Microsoft | https://www.microsoft.com/en-gb/ | |
| Software, algorithm | GraphPad Prism | GraphPad | https://www.graphpad.com/scientific-software/prism/ | |
| Software, algorithm | Adobe Illustrator | Adobe | https://www.adobe.com/uk/products/illustrator.html | |
| Software, algorithm | ImageJ | ImageJ | https://imagej.nih.gov/ij/ | |
| Software, algorithm | Leica LAS X-3D | Leica | https://www.leica-microsystems.com/de/produkte/mikroskop-software/details/product/leica-las-x-ls/ | |
| Software, algorithm | Imaris | Bitplane | http://www.bitplane.com/imaris | |
| Software, algorithm | Flexbar | Dodt et al., 2012 | ||
| Software, algorithm | HiSat | Kim et al., 2015 | ||
| Software, algorithm | SAMtools | Li et al., 2009 | http://samtools.sourceforge.net/ | |
| Software, algorithm | StringTie | Pertea et al., 2015 | ||
| Software, algorithm | R statistics package | R Core Team | https://www.r-project.org/ | |
| Other | Rapamycin gut RNA-seq analysed data | This paper | GEO: GSE148002 |
Additional files
-
Supplementary file 1
Inhibition of mTORC1 activity by rapamycin treatment extends lifespan in females.
Related to Figure 1.
- https://cdn.elifesciences.org/articles/62233/elife-62233-supp1-v2.xlsx
-
Supplementary file 2
Knock-down of histone H3 or H4 in adult enterocytes blocks rapamycin-induced lifespan extension.
Related to Figure 4.
- https://cdn.elifesciences.org/articles/62233/elife-62233-supp2-v2.xlsx
-
Supplementary file 3
Over-expression of H3/H4 in enterocytes recapitulates rapamycin-induced lifespan extension.
Related to Figure 4.
- https://cdn.elifesciences.org/articles/62233/elife-62233-supp3-v2.xlsx
-
Supplementary file 4
Knock-down of Atg5 associated with the expression of H3/H4 in enterocytes abolished the benefits of increased histones on lifespan extension.
Related to Figure 6.
- https://cdn.elifesciences.org/articles/62233/elife-62233-supp4-v2.xlsx
-
Supplementary file 5
Knock-down of Bchs associated with the expression of H3/H4 in enterocytes abolished the benefits of increased histones on lifespan extension.
Related to Figure 7.
- https://cdn.elifesciences.org/articles/62233/elife-62233-supp5-v2.xlsx
-
Supplementary file 6
Knock-down of Bchs in enterocytes abolished rapamycin-induced lifespan extension.
Related to Figure 7—figure supplement 1.
- https://cdn.elifesciences.org/articles/62233/elife-62233-supp6-v2.xlsx
-
Supplementary file 7
Over-expression of Bchs in enterocytes extends lifespan in females.
Related to Figure 7.
- https://cdn.elifesciences.org/articles/62233/elife-62233-supp7-v2.xlsx
-
Transparent reporting form
- https://cdn.elifesciences.org/articles/62233/elife-62233-transrepform-v2.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
A TORC1-histone axis regulates chromatin organisation and non-canonical induction of autophagy to ameliorate ageing
eLife 10:e62233.
https://doi.org/10.7554/eLife.62233




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}