Cohesin architecture and clustering in vivo
- Department of Molecular and Cell Biology, University of California, Berkeley, United States
Figures
Figure 1 with 1 supplement
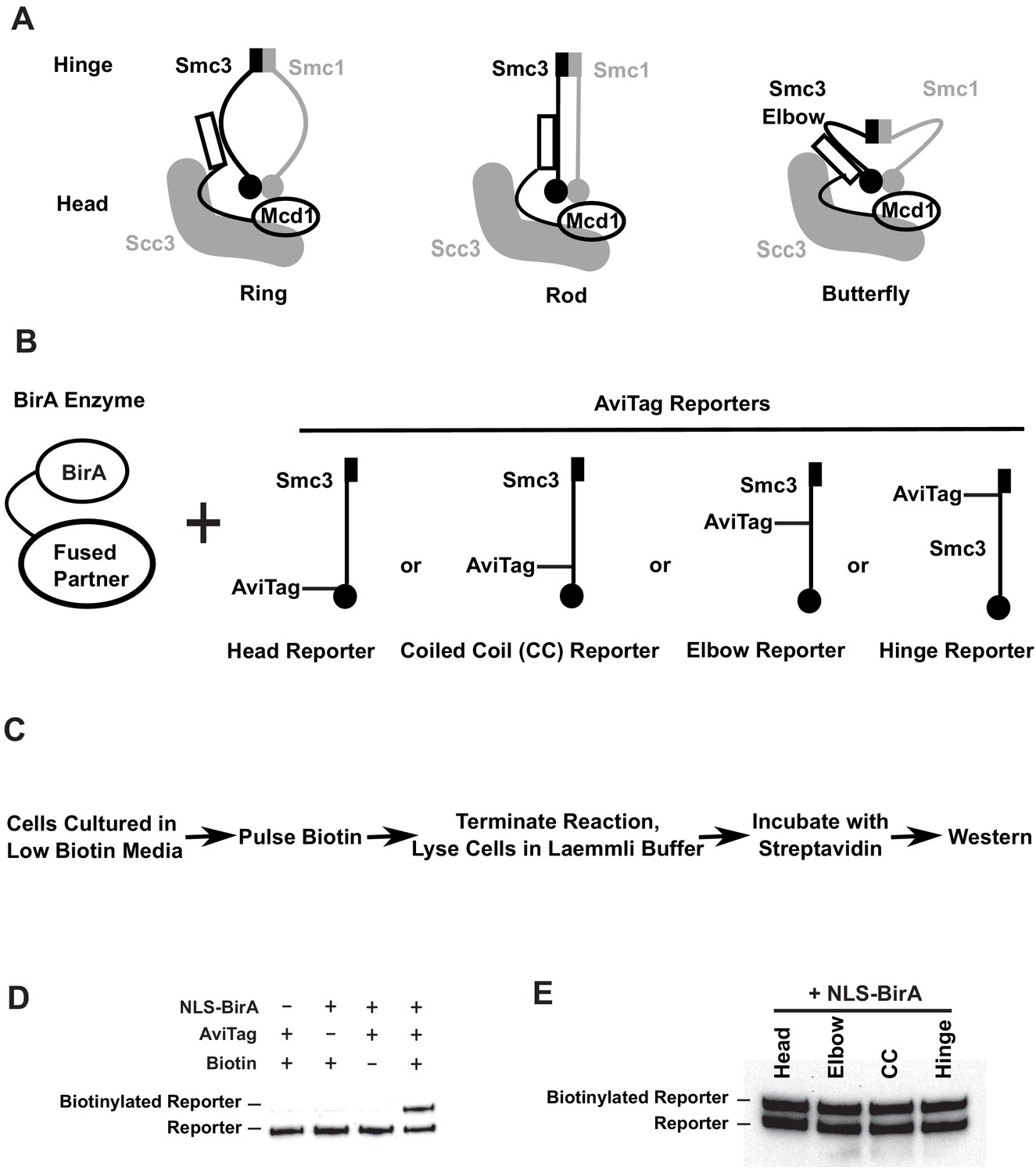
Smc3p reporters for proximity biotinylation experiments.
(A) Cartoons showing the proposed ring, rod, or butterfly conformation of budding yeast cohesin. Each cohesin complex consists of one single copy of each of the four subunits: Smc1p, Smc3p, Mcd1p, and Scc3p. (B) Cartoons depicting strains used in proximity biotinylation experiments. Each strain expresses one of the four AviTag reporters and a BirA enzyme fused to a partner. Each reporter had an AviTag fused to a flexible linker of 6xHA tag inserted into the head (head reporter), the joint of coiled coil (CC reporter), the elbow-proximal coiled coil (elbow reporter), or the hinge (hinge reporter). (C) Experimental regime of proximity biotinylation assays. Cells are cultured in synthetic medium with low biotin and pulsed with biotin briefly. The biotinylation reactions are terminated with the addition of trichloroacetic acid (TCA), and cell lysates are prepared in the Laemmli buffer. The samples are then diluted and incubated with streptavidin and subjected to western blot analysis using anti-HA antibodies to detect both biotinylated and non-biotinylated AviTag reporter. (D) Streptavidin gel shift assays (see 'Streptavidin gel shifts' in 'Materials and methods') to detect biotinylation of the head reporter. Cells were cultured in low biotin synthetic medium with galactose and arrested in mid-M phase by nocodazole. Cell lysates were prepared in sodium dodecyl sulfate (SDS) sample buffer, incubated with 1 mg/ml streptavidin for 15 min and biotinylation of the head reporter is seen as a slower migrating band (lane 4, SX80F). No reporter biotinylation was detected in cells missing BirA enzyme (lane 1, SX73), in cells with a dead AviTag (lane 2, SX222, no lysine in AviTag), or in cells treated with TCA before biotinylation (lane 3, SX80F). (E) Comparison of AviTag accessibility to BirA-mediated biotinylation. Cultures of strains with SMC3 reporter alleles (SX80B, SX80D, SX80E, and SX80F) were grown in low biotin synthetic media at 30°C overnight and treated with nocodazole for 2.5 hr to arrest cells in mid-M phase. Then NLS-BirA, a nuclear-localized version of BirA enzyme, was induced for 1 hr. About 10 nM biotin was added to initiate biotinylation. After a 7-min biotin pulse, the reactions were terminated by TCA. Efficiencies of reporter biotinylation were assessed by streptavidin gel shift.
Figure 1—figure supplement 1
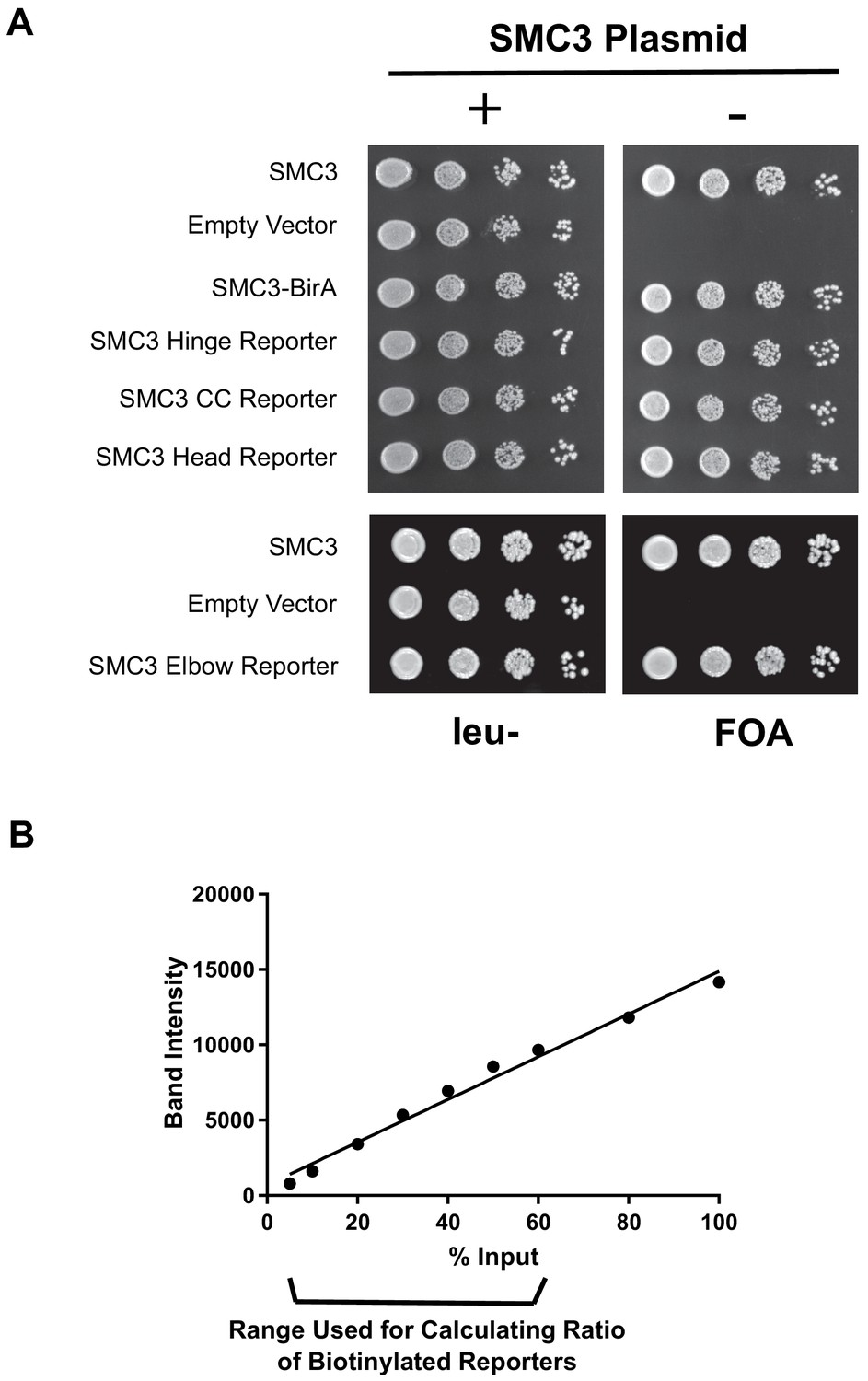
Tagged SMC3 alleles support cell viability.
Smc3p-AviTag yields western signals proportional to the protein levels. (A) Comparison of cells expressing wild-type or SMC3-AviTag as the sole source of SMC3. Shuffle strains carrying CEN URA3 SMC3 plasmid and plasmid expressing the tagged SMC3 alleles. Strains were grown overnight in (yeast extract peptone dextrose) YPD medium and plated at tenfold serial dilution on leucine- (leu-) or 5-fluoroorotic acid (FOA) plates. (B) Quantification of Smc3p-AviTag reporter Western bands blotted with HA antibody. The 5%, 10%, 20%, 30%, 40%, 50%, 60%, 80%, and 100% diluted cells lysates from Figure 1E (hinge reporter, SX80B), respectively, were loaded on the gel, blotted with HA antibody, and band intensities were measured using ImageJ. For gels measuring ratio of AviTag reporter proteins biotinylated, cell lysates were diluted 1.8× and loaded on gel (56% lysate, see 'Materials and methods').
Figure 2 with 1 supplement
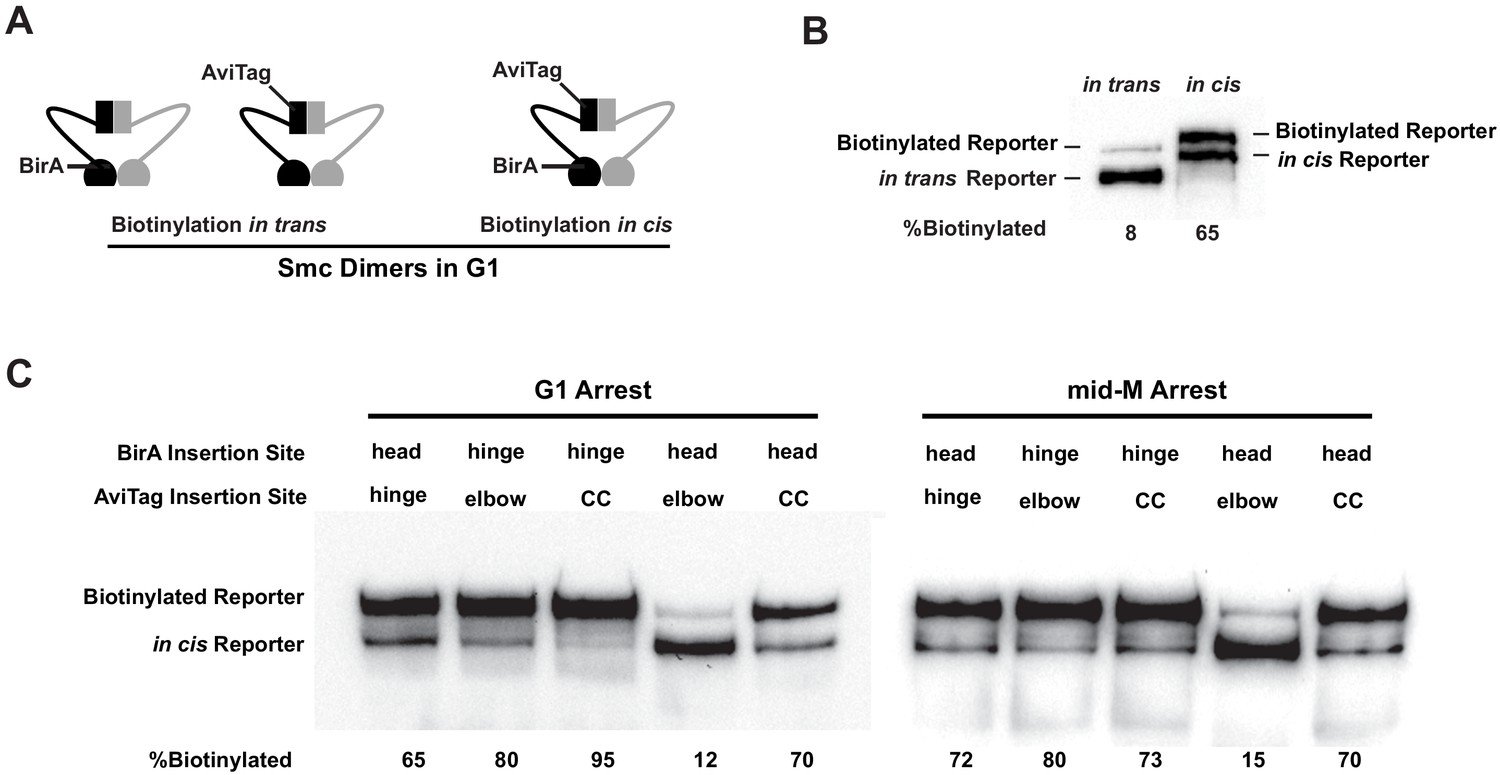
Cohesin adopts a dynamic butterfly conformation in vivo.
(A) Cartoons depicting inter- and intramolecular biotinylation of Smc3 hinge reporter. Both strains were cultured to early log phase and arrested in G1, where cohesin complexes are not fully assembled. In trans, biotinylation experiment was carried out in a strain carrying two SMC3 alleles; one allele was tagged with BirA enzyme in the head domain, while the second allele was tagged with AviTag in its hinge. In cis, biotinylation was carried out in the cells expressing double-tagged Smc3, with BirA in the head and AviTag inserted in the hinge domain of the same molecule. Mcd1p and Scc3p are omitted from the cartoon for clarity. (B) Comparison of biotinylation levels of Smc3 reporters in cis (SX246) and in trans (SX48B). Streptavidin gel shift assay as described in Figure 1D was used to compare Smc3p-AviTag biotinylation in trans and in cis. (C) Smc dimers in G1 and cohesin tetramers in mid-M arrested cells adopt the butterfly conformation. Each strain carries a cis-biotinylation reporter allele of SMC3 (SX246, SX247, SX248, SX249, and SX250). The strain was arrested in G1 with α-factor or mid-M using nocodazole. The cells were then treated with biotin pulse, and biotinylation efficiencies were assayed by streptavidin gel shift.
Figure 2—figure supplement 1
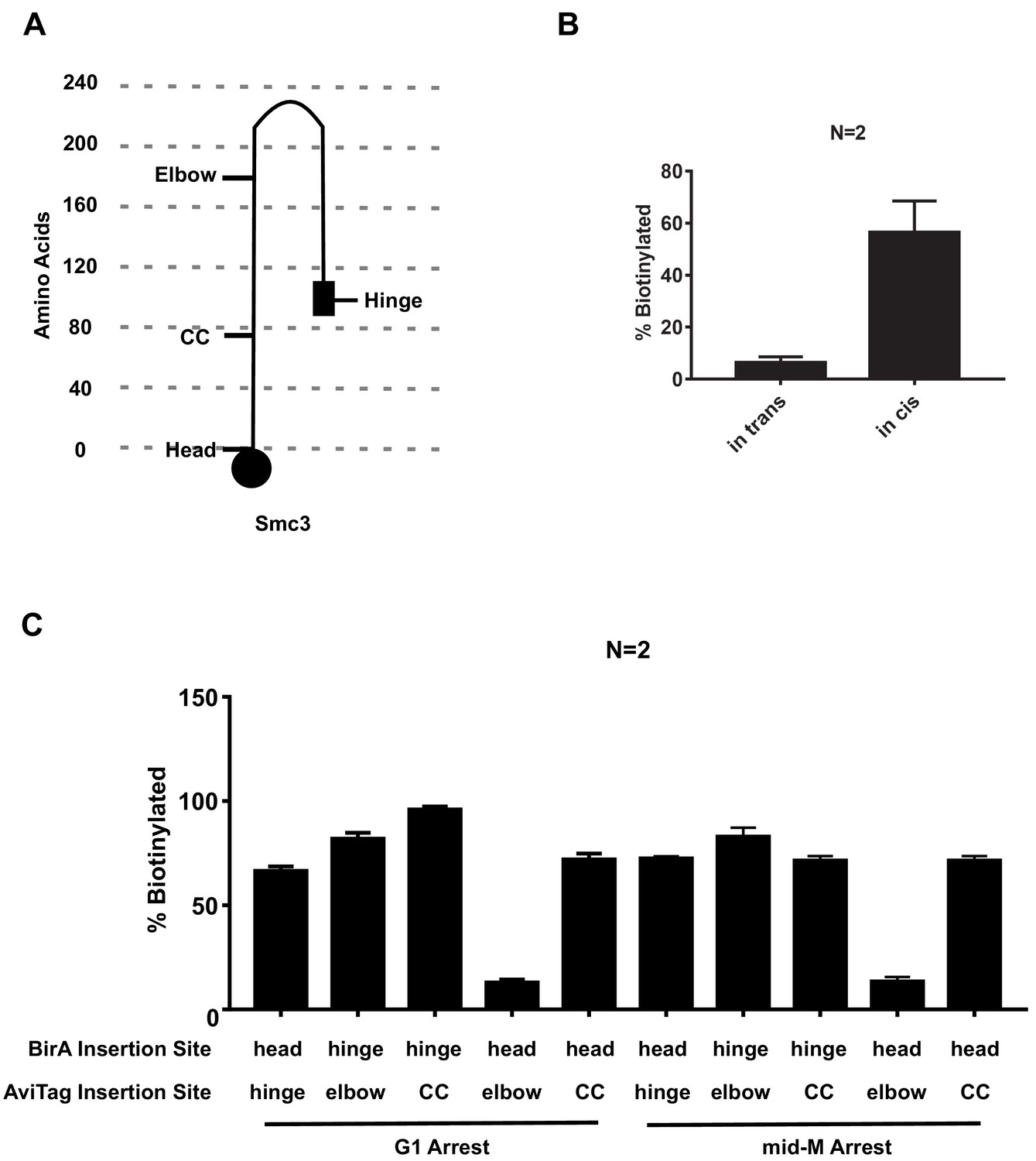
Cartoon of elbow folded Smc3p and quantitative analysis of Figure 2.
(A) Cartoon showing AviTag insertion sites on Smc3p in the elbow folded structure. (B) Quantitative analysis of Figure 2B. Ratios of Smc3p-AviTag reporter proteins biotinylated from two independent cultures were calculated based on band intensities (all error bars in this article are indicative of standard deviations). (C) Quantitative analysis of Figure 2C. Ratios of Smc3p-AviTag reporter proteins biotinylated from two independent cultures were calculated based on band intensities (all error bars in this article are indicative of standard deviations).
Figure 3 with 1 supplement
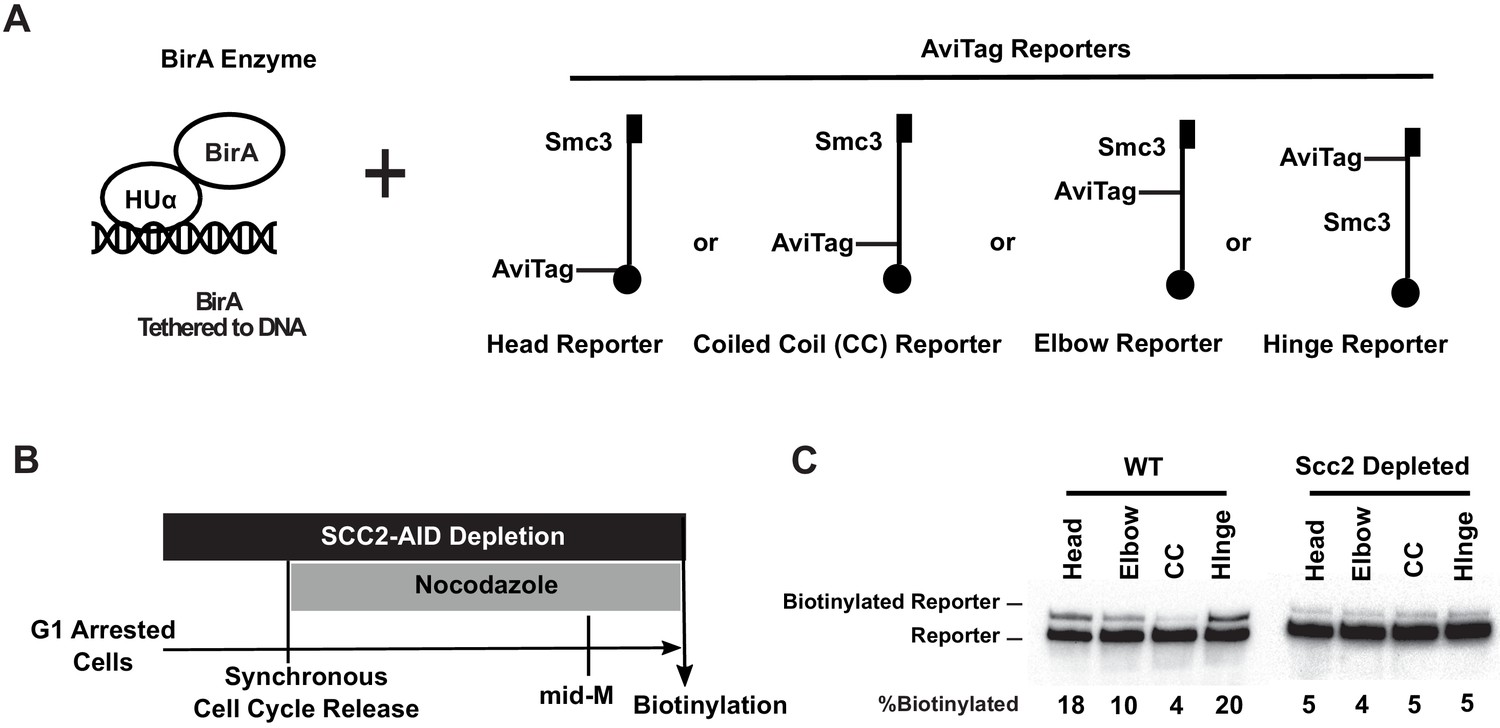
Cohesin binds DNA with both head and hinge domains on DNA.
(A) Cartoons depicting strains used in this figure. BirA enzyme was tethered to DNA by HUα, a bacterial non-specific DNA binding protein. This DNA-tethered BirA was expressed in cells carrying a head, elbow, coiled coil (CC), or hinge reporter Smc3, respectively. Mcd1p and Scc3p are omitted from the cartoon for clarity. (B) Experimental regimen used to test cohesin architecture on DNA. Cells were cultured in synthetic media with low biotin overnight to early log phase and arrested in G1 by α-factor. About 1 mM auxin was added to the arrested cells to deplete the cohesin loader in SCC2-AID cells. Cells were synchronously released from G1 and arrested in mid-M phase in low biotin media containing auxin and nocodazole, then the cultures were treated with biotin pulse. Half of the cells were analyzed by western blotting to assess biotinylation efficiencies of the Smc3 reporters. The other half of the cultures were fixed to assay cohesin on DNA by chromatin immunoprecipitation (Figure 3—figure supplement 1A). (C) Biotinylation of Smc3 reporters by DNA-tethered BirA. The gel on the left side shows biotinylation of Smc3 reporters in wild-type cells (SX136B, SX136D, SX136E, and SX136F). The gel on the right side shows biotinylation of Smc3 reporters in cells depleted of loader subunit Scc2p (SX221B, SX221D, SX221E, and SX221F).
Figure 3—figure supplement 1
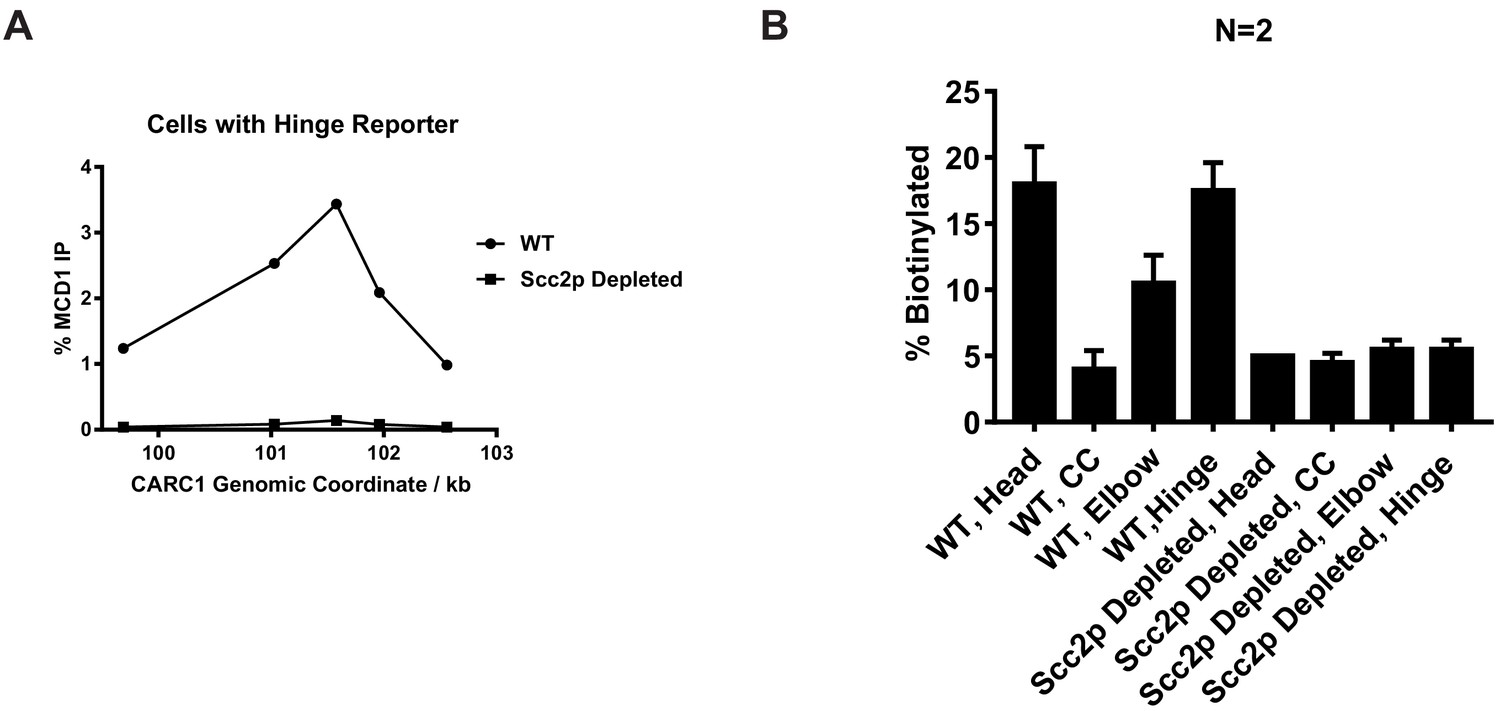
Confirmation of Scc2p-AID depletion and quantitative analysis of Figure 3.
(A) Cohesin failed to bind chromosome in Scc2p-AID-depleted cells. Cells from experiments described in Figure 3B (expressing hinge reporter) were fixed for chromatin immunoprecipitation experiments. Cohesin-bound chromosome was immunoprecipitated by anti-Mcd1 antibody and quantified by qPCR. Cohesin binding at a pericentric site (CARC1) in cells with or without Scc2p was plotted against genomic coordinates. (B) Quantitative analysis of Figure 3C. Ratios of Smc3p-AviTag reporter proteins biotinylated from two independent cultures were calculated based on band intensities.
Figure 4 with 1 supplement
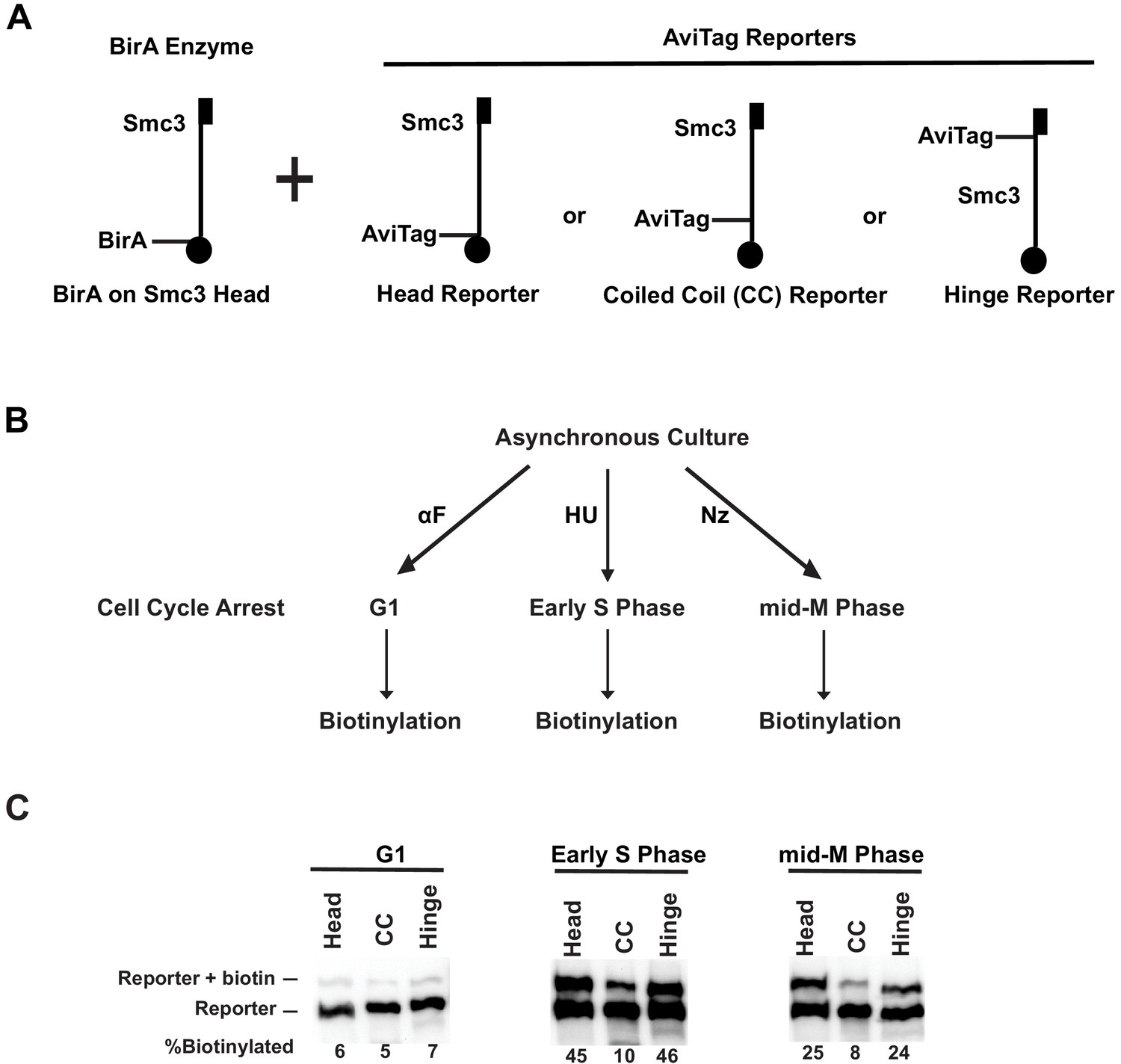
Cohesin forms clusters that are cell cycle-regulated.
(A) Cartoons depicting the strains used (SX48B, SX48E, and SX48F). Each strain carries two SMC3 alleles. One SMC3 allele is tagged with BirA enzyme in its head domain. The other SMC3 allele is the reporter, with AviTag fused to the head domain, the coiled coil (CC), or the hinge domain. Mcd1p and Scc3p are omitted from the cartoon for clarity. (B) Experimental regimen to assess cohesin clusters as a function of the cell cycle. Asynchronous cultures were treated with α-factor, hydroxyurea, or nocodazole to arrest cells in G1, early S phase, or mid-M, respectively. Cells were then treated with a biotin pulse to access levels of cohesin clusters. (C) Comparison of cluster formation in G1, S, and mid-M. Asynchronous cultures expressing Smc3p-BirA and one of the three Smc3p reporters as shown in Figure 3A were treated as described in Figure 3B to assess intermolecular biotinylation of the reporters. Biotinylation was assessed by streptavidin gel shift assay as described in Figure 1D.
Figure 4—figure supplement 1
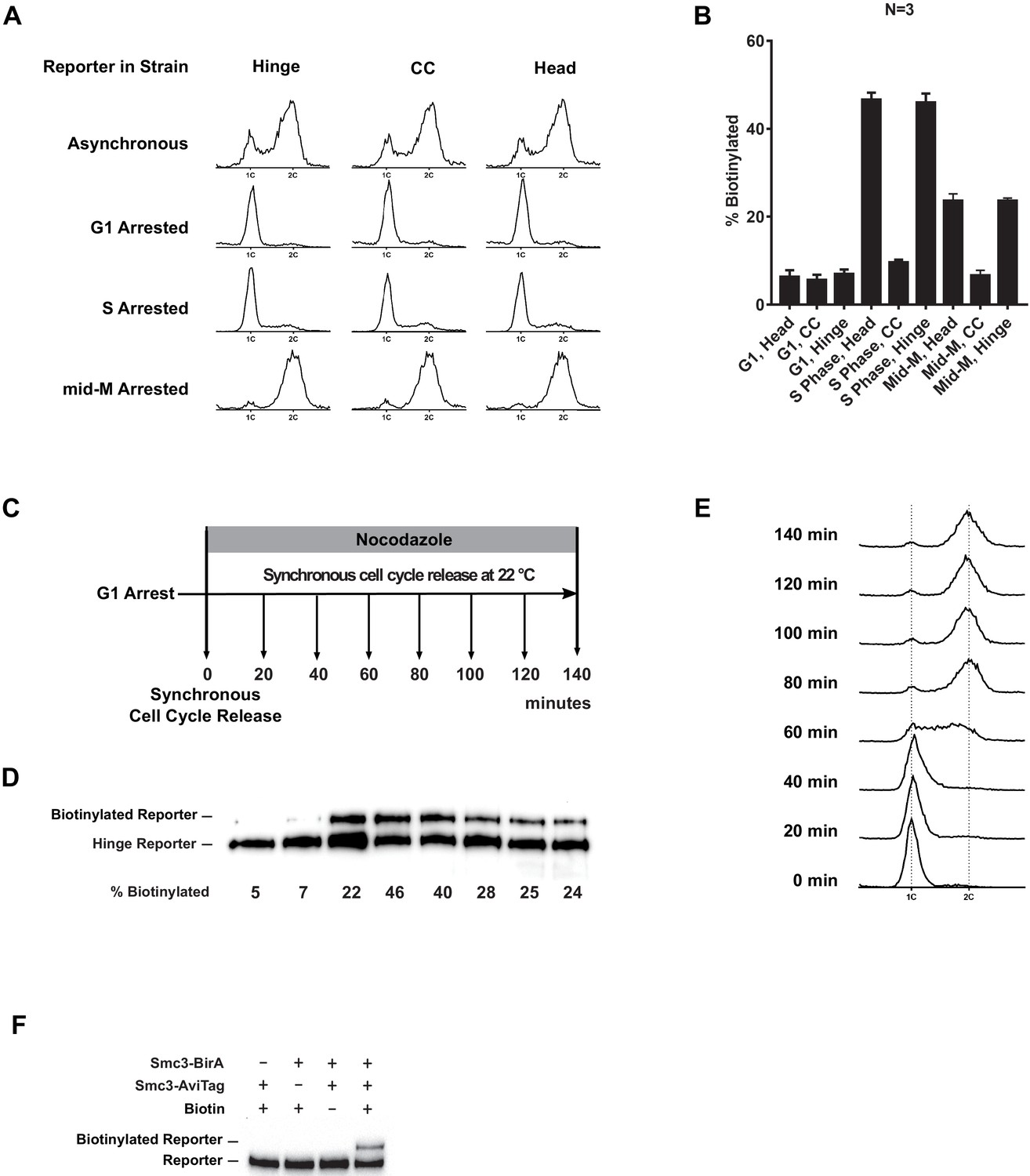
Cohesin clusters in cells synchronously released from G1 into mid-M phase.
(A) DNA content analysis in the G1, early S phase, and mid-M phase arrested cells (left side, comparing with DNA content in asynchronous culture, SX48B, SX48E, and SX48F). (B) Quantitative analysis of Figure 4C. Ratios of Smc3-AviTag reporter proteins biotinylated from three independent cultures were calculated based on band intensities. (C) Regime used to assay cohesin clusters in cells after synchronous G1 release. Cells carrying Smc3p hinge reporter (SX48B) were arrested in G1 and synchronously released into mid-M arrest. Aliquots of the culture were taken every 20 min, and cohesin clustering levels in the cells were assayed with proximity biotinylation. Cells were preserved in 20% trichloroacetic acid (TCA), and biotinylated reporters were assayed with streptavidin gel shift. (D) Cohesin clustering levels during cell cycle progression. A basal level of cohesin cluster was detected in G1 arrested cells. The ratio of cohesin in clusters increases during S phase and then drops to half of its peak level in mid-M arrested cells. (E) Cell cycle progression was shown with DNA content analysis using flow cytometry. (F) Trans-biotinylation of Smc3p-AviTag reporter requires AviTag, BirA, and biotin. Biotinylation of the head reporter is seen as a slower migrating band (lane 4, SX48F). No reporter biotinylation was detected in cells missing BirA enzyme (lane 1, SX73), in cells with a dead AviTag (lane 2, no lysine in AviTag, SX74), or in cells treated with TCA before biotinylation (lane 3, SX48F).
Figure 5 with 2 supplements
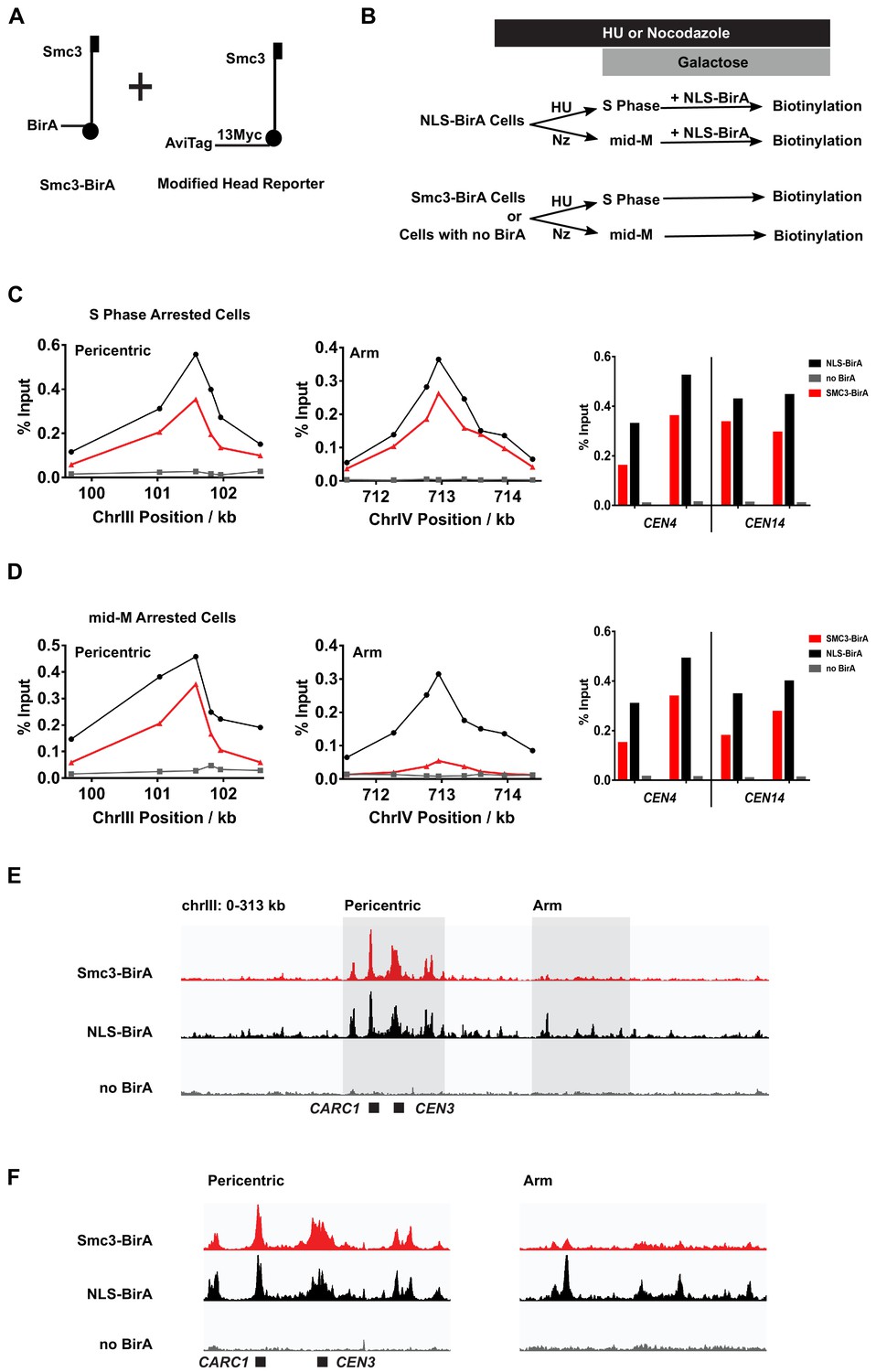
Cohesin clusters localize to centromeres and pericentric cohesin-associated regions (CARs) at high levels in both S and mid-M phases, but decrease at arm CARs in mid-M.
(A) The strain used to assay chromosomal localization of cohesin clusters. The strain carries two SMC3 alleles (SX155), one expresses Smc3 with BirA in the head domain (Smc3-BirA) while the other expresses a modified Smc3 head reporter with an AviTag fused to Smc3 C-terminus via a 13xMyc linker (Smc3-AviTag). Mcd1p and Scc3p are omitted from the cartoon for clarity. (B) Experimental regime used to study chromosomal localization of cohesin clusters. Cells were cultured in synthetic media, arrested in S phase (hydroxyurea) or mid-M (nocodazole), and NLS-BirA was induced in the NLS-BirA strain (SX173 expresses NLS-BirA and SX172 has no BirA). Reporter biotinylation was achieved by biotin pulse, and then cells were fixed and harvested for chromatin immunoprecipitation (ChIP) analysis as described in 'Materials and methods'. (C) Quantitative analysis of cohesin cluster localization by qPCR in S phase arrested cells. ChIP experiments were carried out as shown in (B) with cells arrested in S phase by hydroxyurea. Cohesin clusters in S phase arrested cells were detected on centromeres (right), pericentric CAR (left), and an arm CAR (middle). (D) Quantitative analysis of cohesin cluster localization by qPCR in mid-M arrested cells. ChIP experiments were carried out as shown in (B) with cells arrested in mid-M by nocodazole. In mid-M arrested cells, cohesin clusters were detected at high levels on centromeres (right) and pericentric CAR (left) but not on arm CAR (middle). (E) ChIP to assess cohesin binding to chromosomes. Next-generation sequencing experiments were carried out using samples in (D). Biotinylated cohesin clusters were shown in the red trace (SMC3-BirA). Sequencing results of the control strains were plotted at the same scale. The black trace shows the sequencing result from a positive control, where all Smc3 reporter proteins can be biotinylated (NLS-BirA). The gray trace shows sequencing results from a negative control with no BirA. Positions of the centromere and a pericentric CAR were labeled at the bottom of the traces. (F) Sequencing traces zoomed in the two highlighted regions in (E). The traces on the left show the pericentric region (chrIII: 90–142 kb), and the traces on the right show the chromosome arm (chrIII: 178–237 kb).
Figure 5—figure supplement 1
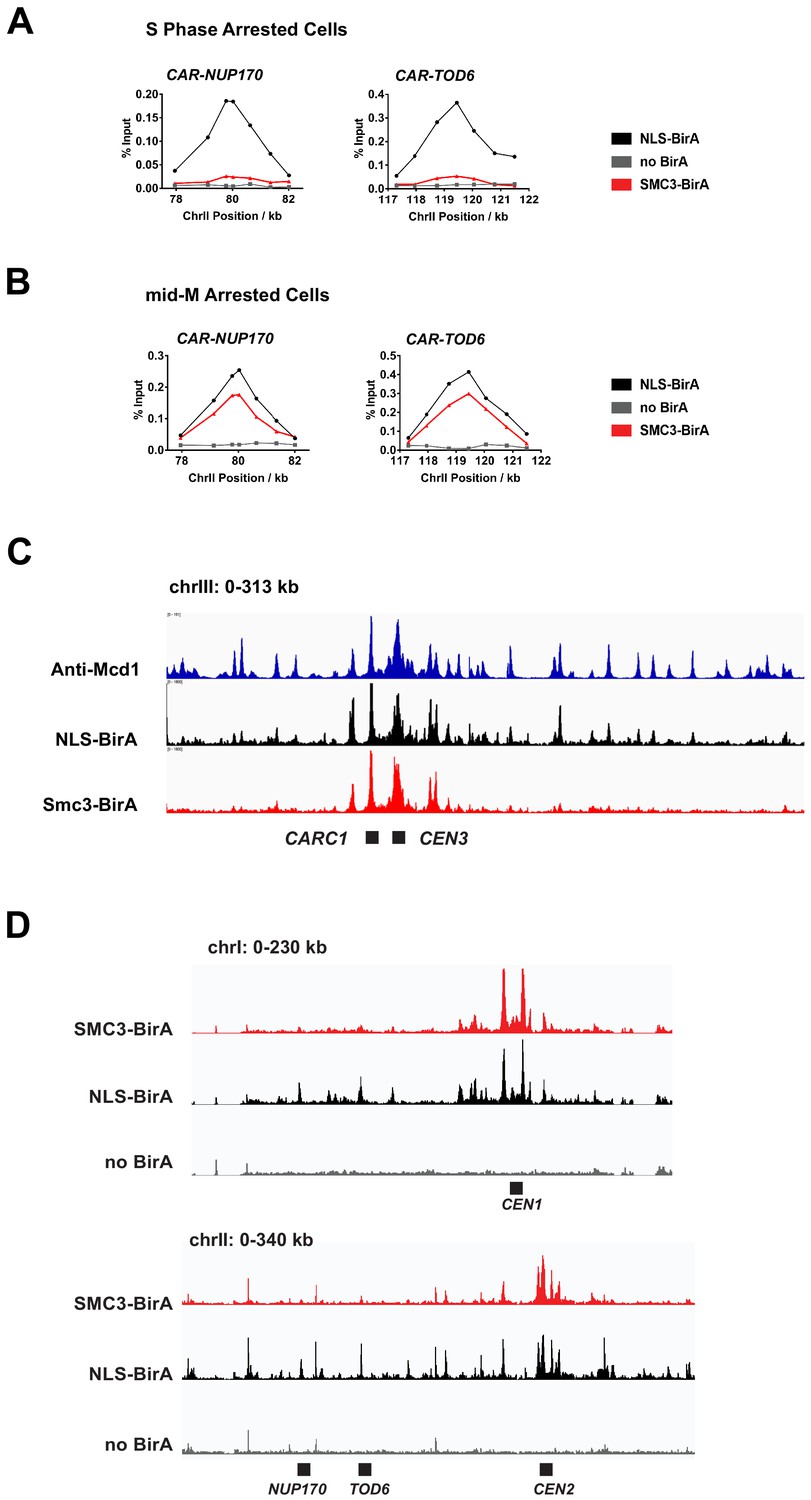
Chromatin immunoprecipitation (ChIP) experiments reveal the centromeric and pericentric localization of cohesin clusters.
(A) Cohesin clusters on the arm of chromosome II in S phase arrested cells. Samples in Figure 5C were analyzed by qPCR using primers targeting cohesin-associated region (CAR) sites at the 3′ end of NUP170 and TOD6. The two binding sites are both on the arm of chromosome II, as shown in (D). (B) Cohesin clusters on the arm of chromosome II in mid-M arrested cells. Samples in Figure 5D were analyzed by qPCR using primers targeting CAR sites at the 3′ end of NUP170 and TOD6. The two binding sites are both on the arm of chromosome II, as shown in (D). (C) Peaks of cohesin clusters correlate with previously identified CAR sites. Anti-Mcd1p ChIP-seq trace (Costantino et al., 2020) at chrIII was compared with ChIP-seq traces from Figure 5E. (D) ChIP-seq traces showing localization of cohesin clusters on chromosome I (top) and part of chromosome II (bottom). The figures were generated from the same sequencing data set used in Figure 5E. The trace with SMC3-BirA (red, SX155) shows the position of cohesin clusters; the trace with pGAL-BirA (black, SX173) shows cohesin binding sites; the no BirA trace (gray, SX172) is the negative control showing non-specific pull down from cells expressing no biotinylation enzyme.
Figure 5—figure supplement 2
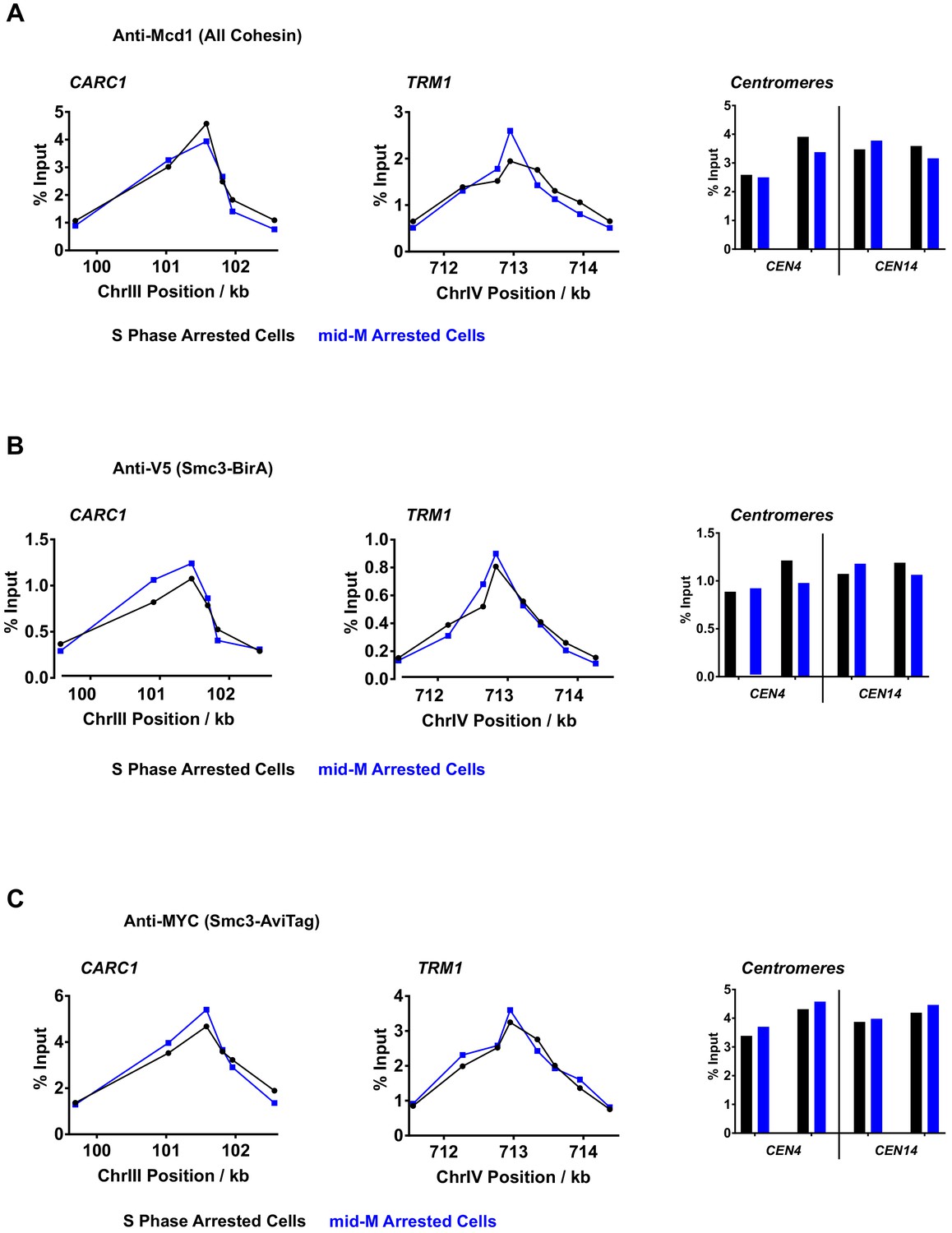
Chromatin immunoprecipitation (ChIP) experiments reveal the centromeric and pericentric localization of cohesin oligomers.
(A) Quantitative analysis of cohesin binding by qPCR in S phase (black) or mid-M (blue) arrested cells. ChIP experiments were carried out with formaldehyde fixed cells expressing both Smc3p-BirA and Smc3p-AviTag reporter using Mcd1 antibody (SX155). In both mid-M and S phase arrested cells, cohesins were detected at centromeres (right), pericentric CAR (left, CARC1), and on arm cohesin-associated region (CAR) (middle, TRM1). (B) Quantitative analysis of BirA-tagged cohesin binding by qPCR in S phase (black) or mid-M (blue) arrested cells. ChIP experiments were carried out with formaldehyde fixed cells using V5 antibody. In both mid-M and S phase arrested cells, BirA-tagged cohesins were detected at centromeres (right), pericentric CAR (left, CARC1), and on arm CAR (middle, TRM1). (C) Quantitative analysis of AviTag-tagged cohesin binding by qPCR in S phase (black) or mid-M (blue) arrested cells. ChIP experiments were carried out with formaldehyde fixed cells using Myc antibody. In both mid-M and S phase arrested cells, AviTag-tagged cohesins were detected at centromeres (right), pericentric CAR (left, CARC1), and on arm CAR (middle, TRM1).
Figure 6 with 1 supplement
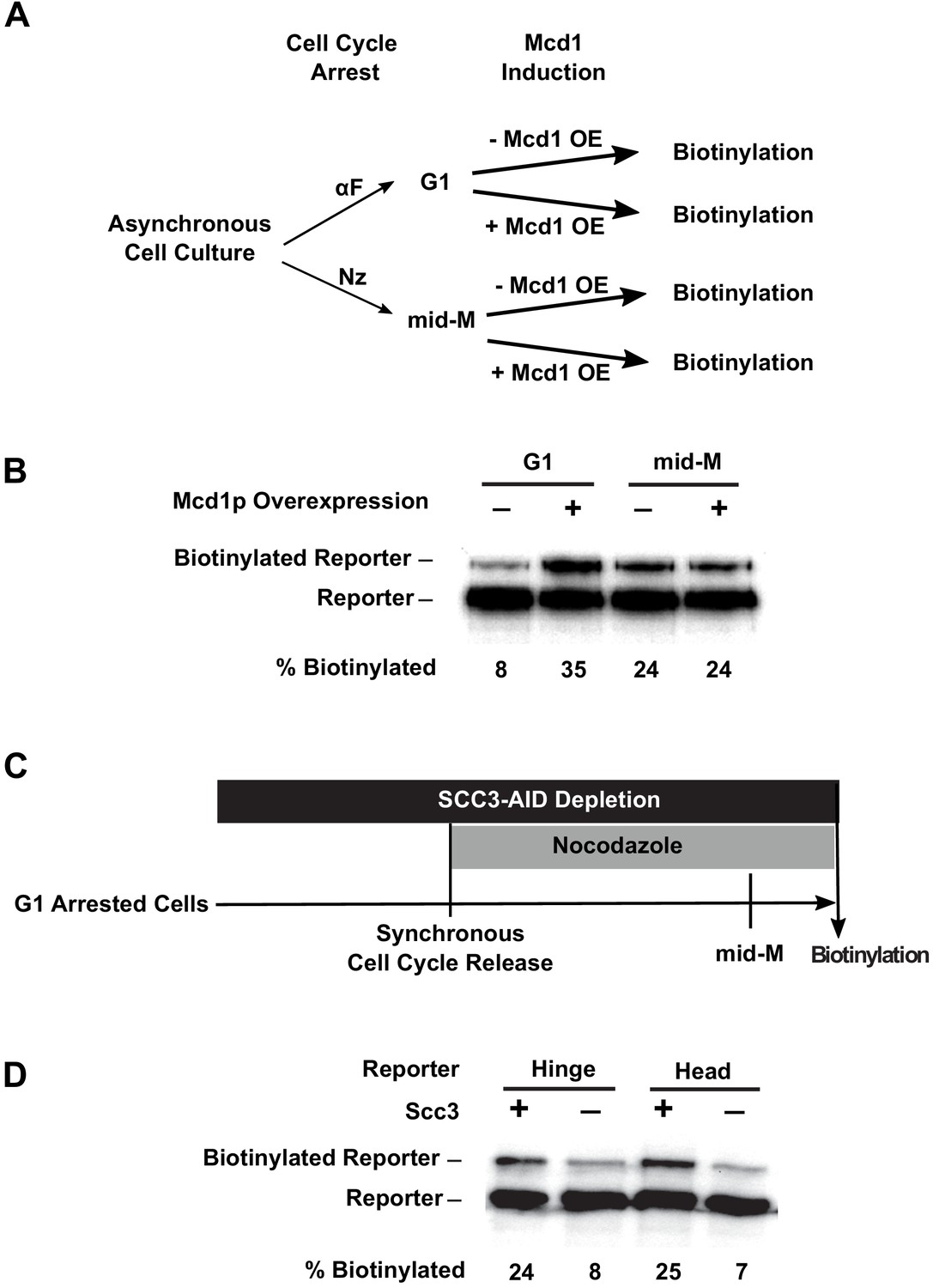
Assembly of the full cohesin tetramer is required for clustering.
(A) Experimental regime used to assess the requirement of Mcd1p in cohesin clustering. The strain carries two SMC3 alleles with one allele tagged with BirA in the head domain and the other allele tagged with AviTag in hinge domain as shown in Figure 4A (SX165). Cultures were arrested in G1 (when Mcd1p is absent) or mid-M (when Mcd1p is assembled in cohesin tetramer). Cultures were split into half, and galactose was added to one aliquot to induce Mcd1p overexpression in arrested cells followed by a short biotin pulse to allow proximity biotinylation of the reporters. (B) Effect of Mcd1p overexpression on cohesin clustering. Cells arrested in G1 or mid-M phase were grown with and without Mcd1p overexpression as described in (A). Levels of cohesin clusters were accessed using streptavidin gel shifts as described in Figure 1D. (C) Experimental regime used to assess the requirement of Scc3p in cohesin clustering. The strains carry two SMC3 alleles: one allele was tagged with BirA in the head domain and the other allele tagged with AviTag in the hinge or head domains as shown in Figure 4A (SX220B and SX220F). Cells were cultured in low biotin synthetic media, arrested in G1, and depletion of scc3p-AID was carried out by auxin addition. Synchronous cell cycle release was carried out in the presence of auxin, and cells were again arrested in mid-M phase by nocodazole. Proximity biotinylation experiments were carried out in the mid-M phase arrested cells. (D) Scc3p is required for cohesin clustering. Plus signs indicate wild-type cells expressing Scc3p, while minus signs indicate cells depleted of scc3p-AID.
Figure 6—figure supplement 1
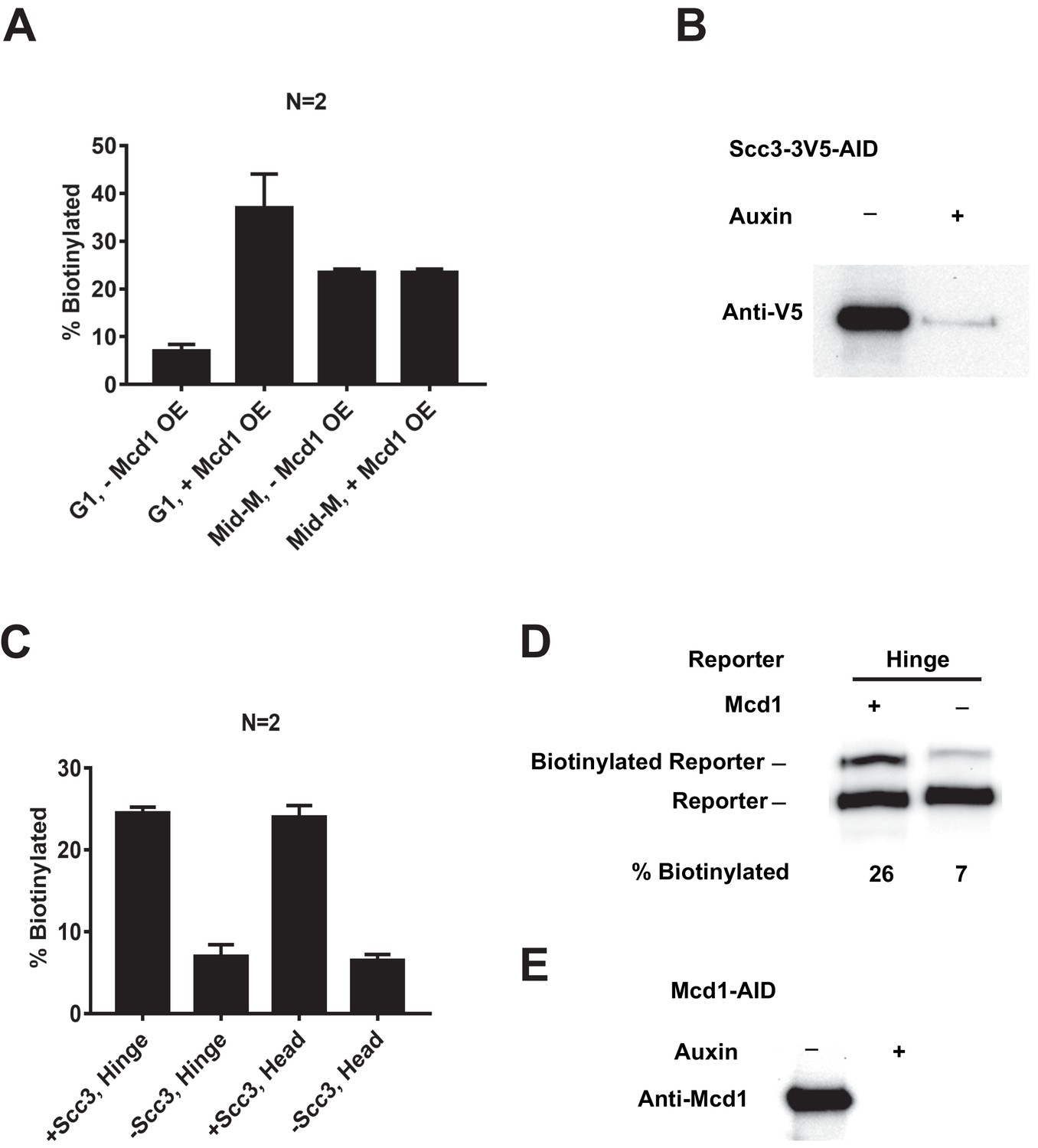
Depletion of SCC2-AID and MCD1-AID.
(A) Quantitative analysis of Figure 6B. Ratios of Smc3-AviTag reporter proteins biotinylated from independent cultures were calculated based on band intensities. (B) Western blot confirms depletion of SCC3-AID. (C) Quantitative analysis of Figure 6D. Ratios of Smc3-AviTag reporter proteins biotinylated from independent cultures were calculated based on band intensities. (D) MCD1 is required for cohesin clustering.Plus signs indicate wild-type cells expressing MCD1, while minus signs indicate cells depleted of MCD1-AID. Cells were cultured following the experiment regime in Figure 6C. Cells were cultured in low biotin synthetic media, arrested in G1, and depletion of MCD1-AID was carried out by auxin addition. Synchronous cell cycle release was carried out in the presence of auxin, and cells were again arrested in mid-M phase by nocodazole. Proximity biotinylation experiments were carried out in the mid-M phase arrested cells. (E) Confirmation of MCD1-AID depletion. Cells were cultured as (D) with (+) or without (−) adding auxin, and western blot was carried out with anti-Mcd1p antibody.
Figure 7 with 1 supplement
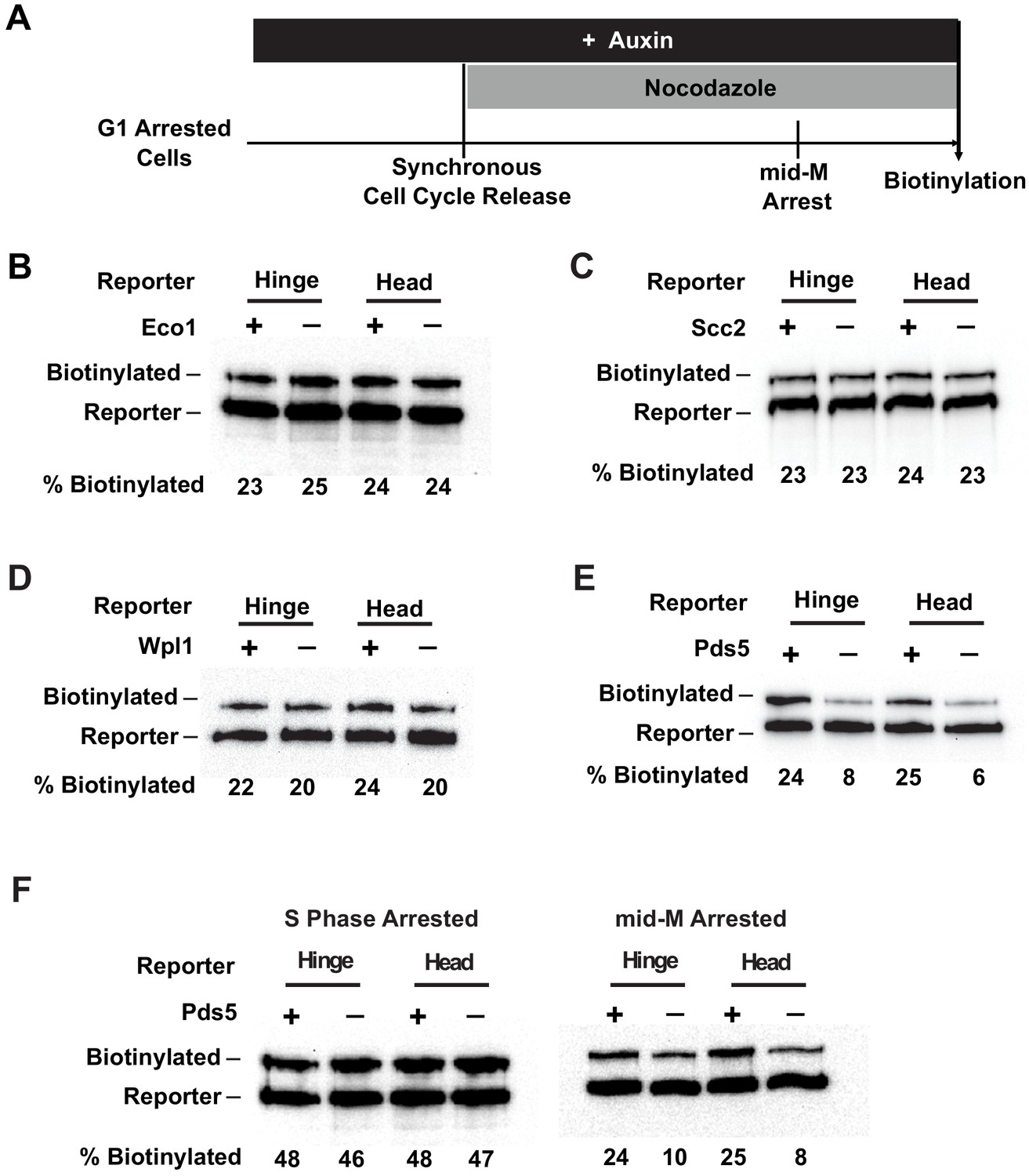
Cohesin clustering requires Pds5p, but not Eco1p, Wpl1p, or cohesin loader.
(A) Experimental regime used to assess the requirement of cohesin regulators for cohesin clusters. Cells were cultured in low biotin synthetic media, arrested in G1, and depletion of the regulator studied was carried out by addition of auxin (except for wpl1Δ strains). Synchronous cell cycle release was carried out in the presence of auxin, and cells were again arrested in mid-M by nocodazole. Proximity biotinylation experiments were carried out in the mid-M arrested cells. (B) Eco1p is not required for cohesin clustering (SX81B and SX81F). Plus signs indicate cells carrying wild-type Eco1p, while minus signs indicate strains depleted of eco1p-AID. eco1p-AID depletion was confirmed by western blotting (Figure 7—figure supplement 1A). (C) Scc2p is not required for cohesin clustering (SX82B and SX82F). Plus signs indicate cells carrying wild-type SCC2, while minus signs indicate strains depleted of scc2p-AID. scc2p-AID depletion was confirmed by western blotting (Figure 7—figure supplement 1C). Cohesin is not loaded onto DNA in the absence of Scc2p (Figure 7—figure supplement 1D). (D) Wpl1p is not required for cohesin clustering (SX66B and SX66F). Plus signs indicate strains carrying the wild-type WPL1 allele, while minus signs represent wpl1Δ strains. (E) Cohesin failed to cluster in pds5p-AID depleted cells (SX122B and SX122F). Plus signs indicate cells expressing wild-type PDS5, while minus signs indicate strains depleted of pds5p-AID. pds5p-AID depletion was confirmed by western blotting (Figure 7—figure supplement 1B). Hinge or head reporters were biotinylated to basal level in the absence of Pds5p. (F) Effects of Pds5p depletion on cohesin clustering in S phase or mid-M arrested cells (SX122B and SX122F). Cells were arrested in S phase or mid-M phase, respectively, and auxin was added to the cell cycle arrested cultures to deplete pds5p-AID. Cohesin clusters were assayed by biotin pulse after pds5p-AID depletion. Cohesin clustering does not require Pds5p in S phase arrested cells but requires Pds5p in mid-M arrested cells.
Figure 7—figure supplement 1
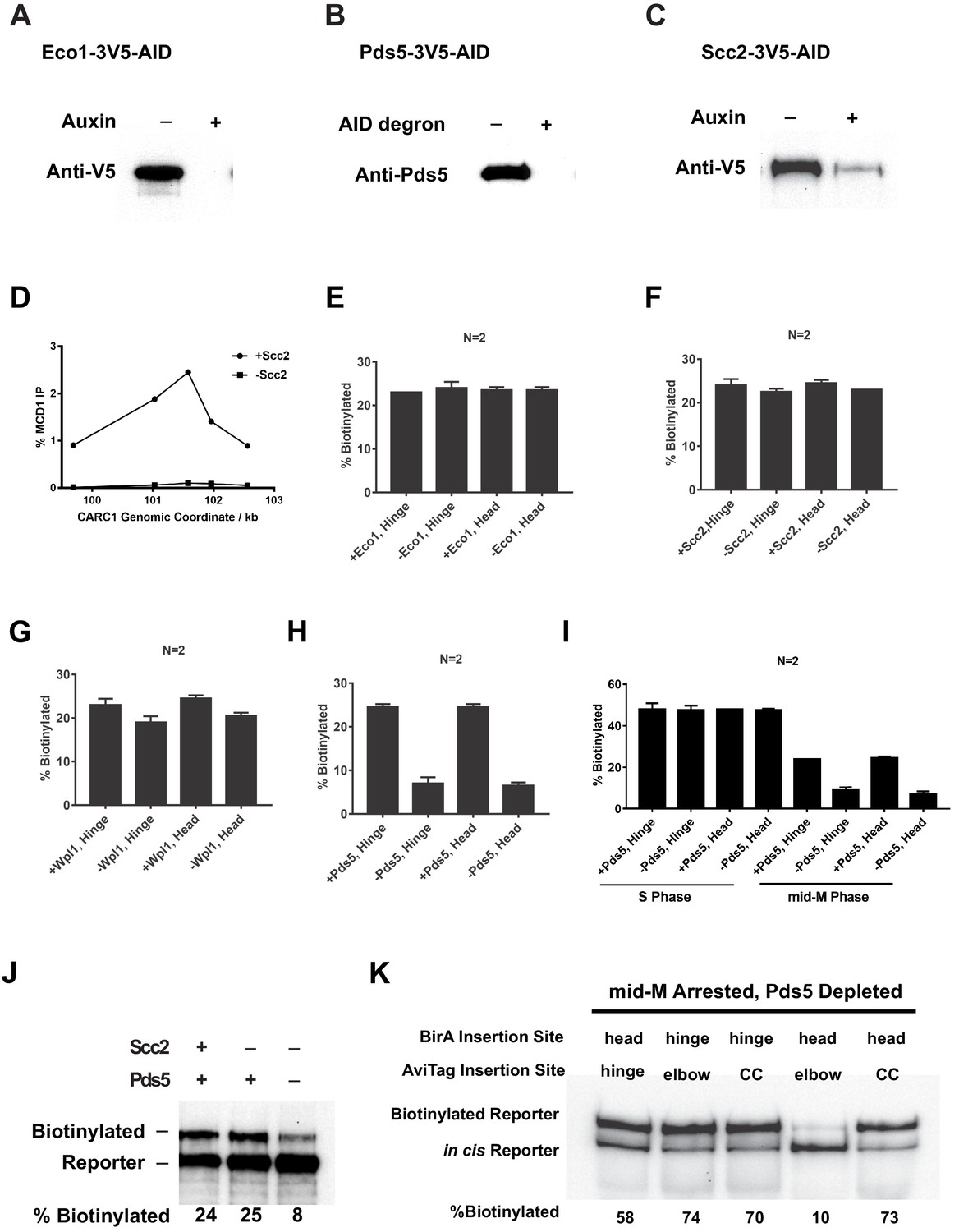
Depletion of Eco1, Pds5, Scc2, and Mcd1.
(A) Western blot showing Eco1p-AID depletion. Cells carrying hinge reporter (sample in Figure 7B) were arrested in G1. The −auxin sample was collected before auxin addition, while the +auxin sample was collected 1 hr after adding auxin. (B) Western blot showing Pds5p-AID depletion. The same samples used for the first two lanes of Figure 7E were analyzed by western blotting using anti-Pds5p antibody. The first lane shows Pds5 protein level in cells with wild-type PDS5. The second lane shows Pds5 protein level in cells with PDS5-AID. (C) Western blot showing Scc2p-AID depletion. Cells carrying hinge reporter (sample in Figure 7C) were arrested in G1. The −auxin sample was collected before auxin addition, while the +auxin sample was collected 1 hr after adding auxin. (D) Cohesin failed to bind chromosome in cells depleted of Scc2p-AID. Aliquots from cell cultures in the first two lanes of Figure 7C were fixed and subjected to chromatin immunoprecipitation (ChIP) against Mcd1. (E) Quantitative analysis of Figure 7B. Ratios of Smc3-AviTag reporter proteins biotinylated from two independent cultures were calculated based on band intensities. (F) Quantitative analysis of Figure 7C. Ratios of Smc3-AviTag reporter proteins biotinylated from two independent cultures were calculated based on band intensities. (G) Quantitative analysis of Figure 7D. Ratios of Smc3-AviTag reporter proteins biotinylated from two independent cultures were calculated based on band intensities. (H) Quantitative analysis of Figure 7E. Ratios of Smc3-AviTag reporter proteins biotinylated from two independent cultures were calculated based on band intensities. (I) Quantitative analysis of Figure 7F. Ratios of Smc3-AviTag reporter proteins biotinylated from two independent cultures were calculated based on band intensities. (J) Cohesin clusters formed in mid-M arrested cells both on and off DNA are Pds5p-dependent. Wild-type cells (first lane), cells depleted of Scc2p-AID (second lane), or cells depleted of both Scc2p-AID and Pds5p-AID (third lane) were synchronously released from G1 into mid-M phase and biotinylated. All three strains carry the Smc3p-BirA and the Smc3p hinge reporter. (K) Fully assembled cohesin tetramers on DNA adopt the butterfly conformation. The strain expresses an auxin-depletable Pds5p-AID and a cis-biotinylation reporter allele (as shown in Figure 2A, right. SX246, SX247, SX248, SX249, and SX250) of SMC3. The strain was arrested in mid-M using nocodazole, and auxin was added to the culture to deplete Pds5p-AID. The cells were then treated with biotin pulse. Trans-biotinylation was abolished in this condition (Figure 7E), and the biotinylation shown is the result of intramolecular reaction.
Figure 8 with 1 supplement

Cohesin mutants defective in cohesion maintenance failed to cluster.
(A) Experimental regime used to assay cohesin clusters in strains expressing wild-type or mutated Mcd1p. Each strain expresses both Smc3p-BirA and Smc3p reporter. Cells were cultured in low biotin synthetic media, arrested in G1, and depletion of wild-type mcd1p-AID was carried out by addition of auxin. Synchronous cell cycle release was carried out in the presence of auxin, and cells were again arrested in mid-M phase by nocodazole with Mcd1p (SX134), mcd1p-V137K (SX130), or mcd1p-q266 (SX129) as the sole Mcd1p protein in the cell. Proximity biotinylation experiments were carried out in the mid-M phase arrested cells. (B) Cohesin failed to form clusters in cells carrying either mcd1-Q266 or mcd1-V137K mutation. Each strain carries two MCD1 alleles: an auxin-depletable MCD1-AID and a test MCD1 mutant allele. Lane 1 has protein extracts from cells expressing wild-type Mcd1p (SX134); lane 2 has protein extracts from cells expressing mcd1p-V137K (SX130); and lane 3 has protein extracts from cells expressing mcd1p-Q266 (SX129). Cohesin clusters were assayed by biotinylation (left), and Mcd1p expression was confirmed by western blotting using Mcd1p antibody (right). (C) Cartoons showing SMC3 alleles used in (D). Each strain carries two SMC3 alleles. One allele expresses a wild-type, BirA-tagged version of SMC3. The other allele expresses a wild-type or D667-mutated version of Smc3p reporter (AviTag inserted in the head in SX49F, and in the hinge domain in SX49B). (D) smc3p-D667 reporters failed to cluster with wild-type Smc3p-BirA. Cells depicted in (C) were arrested in mid-M phase, and interactions between wild-type Smc3p and mutated smc3p-D667 reporters were assayed by proximity biotinylation.
Figure 8—figure supplement 1
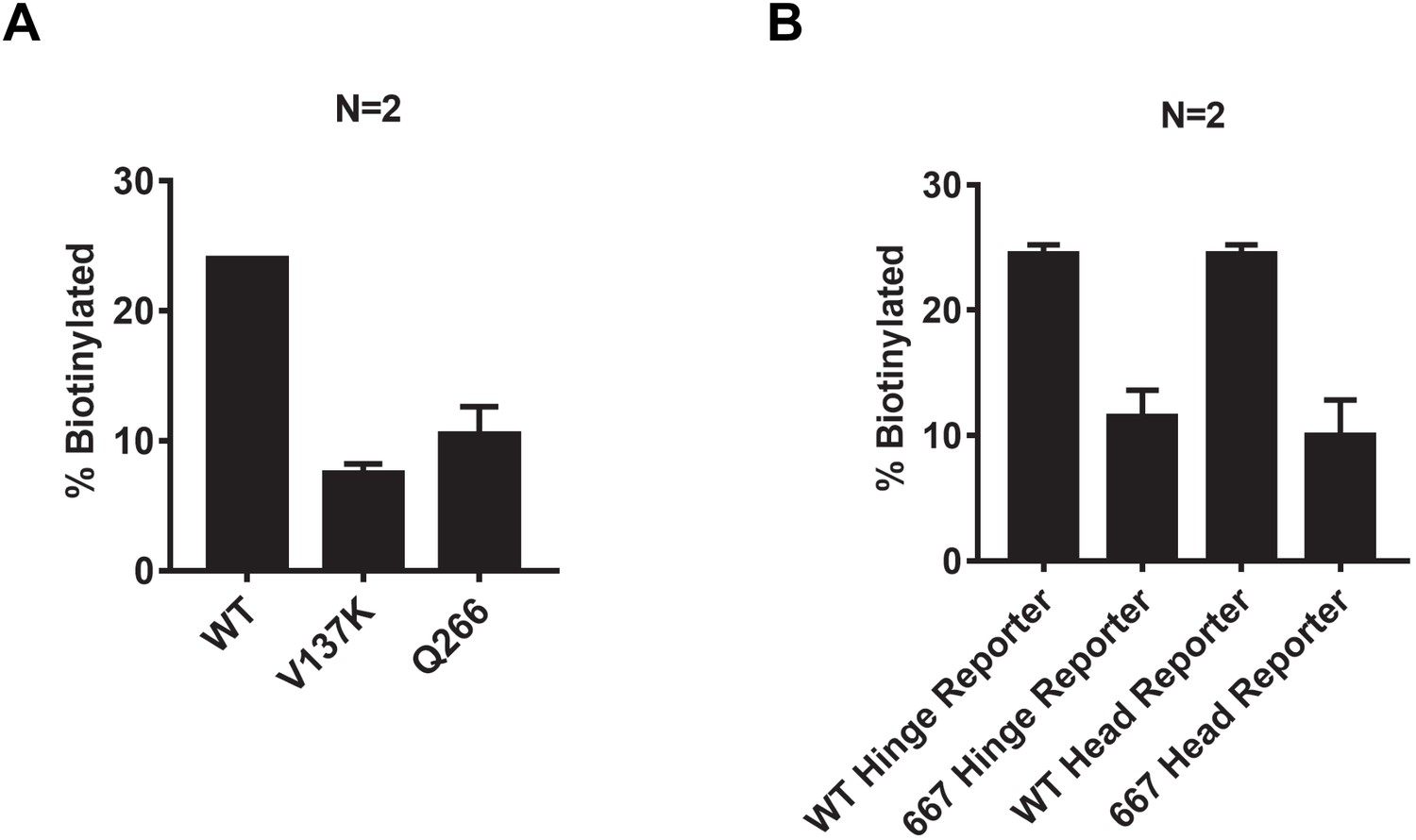
Quantification of Smc3 reporter biotinylation in cells expressing mcd1-V137K, mcd1-Q266, or smc3-D667.
(A) Quantitative analysis of Figure 8B. Ratios of Smc3p-AviTag reporter proteins biotinylated from two independent cultures were calculated based on band intensities. (B) Quantitative analysis of Figure 8D. Ratios of Smc3p-AviTag reporter proteins biotinylated from two independent cultures were calculated based on band intensities.
Figure 9

Model of cohesin cluster regulation during cell cycle.
(A) Cartoon showing the initial steps of asymmetric loop extrusion by cohesin. Mcd1p, Scc3p, and Pds5p are omitted from the cartoon for clarity. Cohesin monomer binds DNA with both the head and hinge domains (cohesin complex in extended conformation may adopt rod or open ring structures), then the coiled coil domain bends to form the butterfly conformation and extrude a DNA loop. In subsequent steps (not shown here), DNA is transferred from the hinge to a second DNA binding site on the head. The hinge binds a new region of DNA and repeats the cycle, increasing the size of the loop. Cohesin dimmers can extrude DNA symmetrically. This model has been proposed for the condensin complex (Ganji et al., 2018). Mcd1p and Scc3p are omitted from the model for simplicity. (B) Cartoon showing sister chromatid cohesion by cohesin. Cohesin binds sisters with its head and hinge domains, respectively, and tethers sister chromatids. This tethering state proceeds to the butterfly conformation where it is trapped. Clustering of cohesins in the butterfly conformation stabilizes tethering between sisters and maintains cohesion. (C) Cartoon showing regulation of cohesin clusters during cell cycle. Cohesin complexes form clusters at all previously identified binding sites at early S phase, including centromeres, pericentric cohesin-associated regions (CARs), and arm CAR sites. Most of the clusters at chromosome arms dissolve in mid-M phase. A small fraction of cohesin complexes remain in the clustering state and keep sister chromatids tethered. Pds5 protein participates in the stabilization of cohesin clusters through S phase into mid-M.
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain, strain background (Saccharomyces cerevisiae) | This paper | Supplementary file 1 | ||
| Sequence-based reagent | qPCR primers | IDT | Supplementary file 2 | |
| Antibody | Rabbit polyclonal anti-Mcd1 | V. Guacci via Covance | Anti-Mcd1 (555) | WB (1:10,000) ChIP (1:1000) |
| Antibody | Rabbit polyclonal anti-Pds5 | V. Guacci via Covance | Anti-Pds5 (556) | WB (1:20,000) |
| Antibody | Mouse monoclonal anti-HA (12CA5) | Roche | 11667203001 | WB (1:8000) |
| Antibody | Mouse monoclonal anti-MYC (9E10) | Roche | 11667203001 | WB (1:8000) |
| Antibody | Mouse monoclonal anti-V5 | Invitrogen | 46-0705 | WB (1:8000) |
| Antibody | Goat polyclonal HRP anti-rabbit | Biorad | 170-6515 | WB (1:8000) |
| Antibody | Goat polyclonal HRP anti-Mouse | Biorad | 170-6516 | WB (1:5000) |
| Other | Protein A Dynabeads | Invitrogen | 10002D | |
| Other | Dynabeads MyOne Streptavidin T1 | Invitrogen | 65601 | |
| Peptide, recombinant protein | Streptavidin | Invitrogen | S888 | |
| Chemical compound, drug | Auxin | Millipore-Sigma | C9911 | 1 mM final concentration, 0.5 M stock in dimethyl sulphoxide (DMSO) |
| Peptide, recombinant protein | Alpha factor | Millipore-Sigma | T6901 | 24 nM final concentration, 10 nM stock |
| Chemical compound, drug | Hydroxyurea | Millipore-Sigma | H8627 | 0.2 M final concentration |
| Chemical compound, drug | Nocodazole | Millipore-Sigma | M1404 | 0.012 mg/ml final concentration, 1.5 mg/ml stock in DMSO |
| Chemical compound, drug | Ethyl acetate ACS | Millipore-Sigma | 319902 | |
| Peptide, recombinant protein | Pronase protease | Millipore-Sigma | 537088 | 10 mg/ml stock in water |
| Other | YNB-biotin | Sunrise Science Products | 1523-100 | |
| Other | BSM powder | Sunrise Science Products | 1387-100 | |
| Chemical compound, drug | D-Biotin | Invitrogen | B20656 |
Additional files
-
Supplementary file 1
Yeast strains used in this study.
All strains listed were generated for this study and derived from the A364a background.
- https://cdn.elifesciences.org/articles/62243/elife-62243-supp1-v2.xlsx
-
Supplementary file 2
Oligonucleotides used for RT-qPCR.
- https://cdn.elifesciences.org/articles/62243/elife-62243-supp2-v2.docx
-
Transparent reporting form
- https://cdn.elifesciences.org/articles/62243/elife-62243-transrepform-v2.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Cohesin architecture and clustering in vivo
eLife 10:e62243.
https://doi.org/10.7554/eLife.62243




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}