Fruitless mutant male mosquitoes gain attraction to human odor
- Laboratory of Neurogenetics and Behavior, The Rockefeller University, United States
- Kavli Neural Systems Institute, United States
- Howard Hughes Medical Institute, United States
Abstract
The Aedes aegypti mosquito shows extreme sexual dimorphism in feeding. Only females are attracted to and obtain a blood-meal from humans, which they use to stimulate egg production. The fruitless gene is sex-specifically spliced and encodes a BTB zinc-finger transcription factor proposed to be a master regulator of male courtship and mating behavior across insects. We generated fruitless mutant mosquitoes and showed that males failed to mate, confirming the ancestral function of this gene in male sexual behavior. Remarkably, fruitless males also gain strong attraction to a live human host, a behavior that wild-type males never display, suggesting that male mosquitoes possess the central or peripheral neural circuits required to host-seek and that removing fruitless reveals this latent behavior in males. Our results highlight an unexpected repurposing of a master regulator of male-specific sexual behavior to control one module of female-specific blood-feeding behavior in a deadly vector of infectious diseases.
eLife digest
Sexual dimorphism is a phenomenon among animals, insects and plants where the two sexes of a species show differences in body size, physical features or colors. The bushy mane of a male lion, for example, is nowhere to be seen on a female lioness, and only male peacocks have extravagant tails. Most examples of sexual dimorphism, such as elaborate visual displays or courtship behaviors, are linked to mating.
However, there are a few species where behavioral differences between the sexes are not connected to mating. Mosquitoes are an example: while female mosquitoes feed on humans, and are attracted to a person’s body heat and odor, male mosquitoes have little interest in biting humans for their blood. Therefore, female mosquitoes are the ones responsible for transmitting the viruses that cause certain blood-borne diseases such as dengue fever or Zika. Determining which genes are linked to feeding behaviors in mosquitoes could allow researchers to genetically engineer females so they no longer bite people, thus stopping the spread of these diseases. Unfortunately, the genes that control mosquito feeding behaviors have not been well studied.
In other insects, some of the genes that control mating behaviors that depend on sex have been identified. For example, a gene called fruitless controls courtship behaviors in male flies and silkworms, and is thought to be the ‘master regulator’ of male sexual behavior across insects. Yet it remains to be seen whether the fruitless gene has any effect in mosquitoes, where sex differences relate to feeding habits.
To investigate this, Basrur et al. removed the fruitless gene from Aedes aegypti mosquitoes. The genetically altered male mosquitoes became unable to mate successfully, but – similar to unmodified males – still preferred sugar water over blood when feeding. Unlike unmodified males, however, the male mosquitoes lacking fruitless were attracted to the body odor of a person’s arm (like females).
These results reveal that fruitless, a gene that controls sex-specific mating behaviors in other insects, controls a sex-specific feeding behavior in mosquitoes. The fruitless gene, Basrur et al. speculate, likely gained this role controlling mosquito feeding behavior in the course of evolution. More research is required to fully understand the effects of the fruitless gene in male and female mosquitoes.
Introduction
Across animals, males and females of the same species show striking differences in behavior. Male Paradisaeidae birds-of-paradise perform an elaborate courtship dance to seduce prospective female partners, contorting their bodies in forms resembling flowers, ballerinas, and smiling faces (Scholes, 2008). Female Serromyia femorata midges pierce and suck conspecific males dry during mating, breaking off his genitalia inside her, thereby supplying the female with both nutrition and sperm (Edwards, 1920). Although an astonishing diversity of sexually dimorphic behaviors exists across species, most insights into the genetic and neural basis of sex-specific behaviors have come from a limited set of model organisms (Matthews and Vosshall, 2020). Which genes control sexual dimorphism in specialist species that have evolved novel behaviors? Do conserved genes control sexual dimorphism in species-specific behaviors, or do novel genes evolve to control new behaviors?
Many advances in understanding the genetics of sexually dimorphic behaviors have come from the study of Drosophila melanogaster fly courtship, where a male fly orients toward, taps, and follows a female fly, extending a wing to produce a courtship song before tasting, mounting, and copulating with her (Hall, 1994). Courtship comprises behavioral modules, which are simple discrete behaviors that must be combined to perform a complex behavior and are elicited by different sensory modalities and subsets of fruitless-expressing neurons (Clowney et al., 2015; Clyne and Miesenböck, 2008; Kohl et al., 2013; Ruta et al., 2010; von Philipsborn et al., 2011). Courtship modules include orienting, which is driven by visual information (Ribeiro et al., 2018) and persistent following and singing, which are triggered by chemical cues on a female fly (Clowney et al., 2015) and guided by vision (Ribeiro et al., 2018; Sten et al., 2020). The fruitless gene is sex-specifically spliced in the brain of multiple insect species including mosquitoes (Bertossa et al., 2009; Gailey et al., 2006; Salvemini et al., 2013) and has been proposed to be a master regulator of male courtship and mating behavior across insects (Clowney et al., 2015; Demir and Dickson, 2005; Hall, 1994; Ryner et al., 1996; Seeholzer et al., 2018; Tanaka et al., 2017). Sex-specific splicing of the fruitless gene controls several aspects of courtship behavior. Male flies mutant for fruitless promiscuously court other males and cannot successfully mate with females (Ito et al., 1996; Ryner et al., 1996). Forcing male fruitless splicing in females triggers orientation and singing behaviors normally only performed by males (Demir and Dickson, 2005). In addition, fruitless is required for sex-specific aggressive behaviors (Vrontou et al., 2006). Fruitless encodes a BTB zinc-finger transcription factor that is thought to control cell identity and connectivity during development (Ito et al., 2016; Neville et al., 2014), as well as the functional properties of neurons in adulthood (Sethi et al., 2019). Fruitless modulates the expression of a number of potential downstream target genes (Neville et al., 2014; Sato and Yamamoto, 2020; Vernes, 2015) in a cell-type-specific manner (Brovkina et al., 2020). Moreover, fruitless has a conserved role controlling courtship in multiple Drosophila species (Seeholzer et al., 2018; Tanaka et al., 2017), and sex-specific fruitless splicing is conserved across wasps (Bertossa et al., 2009) and mosquitoes (Gailey et al., 2006; Salvemini et al., 2013), suggesting that fruitless may act as a master regulator of sexually dimorphic mating behaviors across insects.
Mosquitoes display striking sexually dimorphic mating and feeding behaviors. Only male mosquitoes initiate mating, and only females drink blood, which they require to develop their eggs (Bowen, 1991; Galun et al., 1963; Jové et al., 2020; Klowden, 1995). Sexual dimorphism in blood-feeding is one of the only instances of a completely sexually dimorphic feeding behavior because male mosquitoes never pierce skin or engorge on blood. While part of this dimorphism is enforced by sex-specific genitalia (Spielman, 1964) or feeding appendages (Jones and Pilitt, 1973), there is also a dramatic difference in the drive to hunt hosts between males and female mosquitoes (Bowen, 1991; Roth, 1948). To blood-feed, females combine multiple behavioral modules (Bowen, 1991). Female Aedes aegypti mosquitoes take flight when exposed to carbon dioxide (Bowen, 1991; McMeniman et al., 2014) and are attracted to human olfactory (DeGennaro et al., 2013; Dekker et al., 2005; Zwiebel and Takken, 2004), thermal, and visual cues (Liu and Vosshall, 2019; McMeniman et al., 2014; van Breugel et al., 2015), and integrate at least two of these cues to orient toward and land on human skin. Engorging on blood is triggered by specific sensory cues tasted by the female (Galun et al., 1963; Jové et al., 2020). It is not known which genes have evolved to control this unique sexually dimorphic and mosquito-specific feeding behavior. Here, we generate fruitless mutant Aedes aegypti mosquitoes and show that consistent with observations in Drosophila, fruitless is required for male mating behavior. Unexpectedly, fruitless mutant male mosquitoes gain the ability to host-seek, specifically driven by an attraction to human odor. Our results demonstrate that sexual dimorphism in a single module of a mosquito-specific behavior is controlled by a conserved gene that we speculate has gained a new function in the course of evolution.
Results
Fruitless is sex-specifically spliced in the mosquito nervous system
We used an arm-next-to-cage assay (Figure 1A) to monitor attraction of male and female Aedes aegypti mosquitoes to a live human arm. Consistent with their sexually dimorphic blood-feeding behavior, only females were strongly attracted to the arm (Figure 1B–C). What is the genetic basis for this extreme sexual dimorphism? We reasoned that fruitless, which is alternatively spliced in a sex-specific manner and promotes male courtship and copulation in D. melanogaster flies (Ito et al., 1996; Ryner et al., 1996), may play similar roles in controlling sexually dimorphic behaviors in Aedes aegypti. Fruitless is a complex gene with multiple promoters and multiple alternatively spliced exons. Downstream promoters drive broadly expressed non-sex-specific fruitless transcripts and proteins (Lee et al., 2000). A previous study showed (Salvemini et al., 2013) and we confirmed that transcripts from the upstream neuron-specific (P1) promoter in the Aedes aegypti fruitless gene are sex-specifically spliced (Figure 1D–H). Both male and female transcripts include a short male ‘m’ exon, and female transcripts additionally include a longer female ‘f’ exon with an early stop codon, predicted to yield a truncated Fruitless protein in the female. However, it is unlikely that any sex-specific Fruitless protein or peptides are stably expressed in adult females. In Drosophila, sex-specific female fruitless peptides are not detected (Lee et al., 2000), and transformer is thought to inhibit translation by binding to female fruitless P1 transcripts (Usui-Aoki et al., 2000). P1 transcripts of both sexes splice to the first common ‘c1’ exon, but only male transcripts are predicted to encode full-length Fruitless protein with BTB and zinc-finger domains (Figure 1E). By analyzing previously published tissue-specific RNA-seq data (Matthews et al., 2016), we verified that of all the fruitless exons, only the f exon was sex-specific in Aedes aegypti brains (Figure 1G). Moreover, while the c1 exon was broadly expressed through downstream promoters, P1 transcripts were specifically expressed in the brain and the antenna, the major olfactory organ of the mosquito (Figure 1H), consistent with fruitless expression in Drosophila (Stockinger et al., 2005).
Figure 1
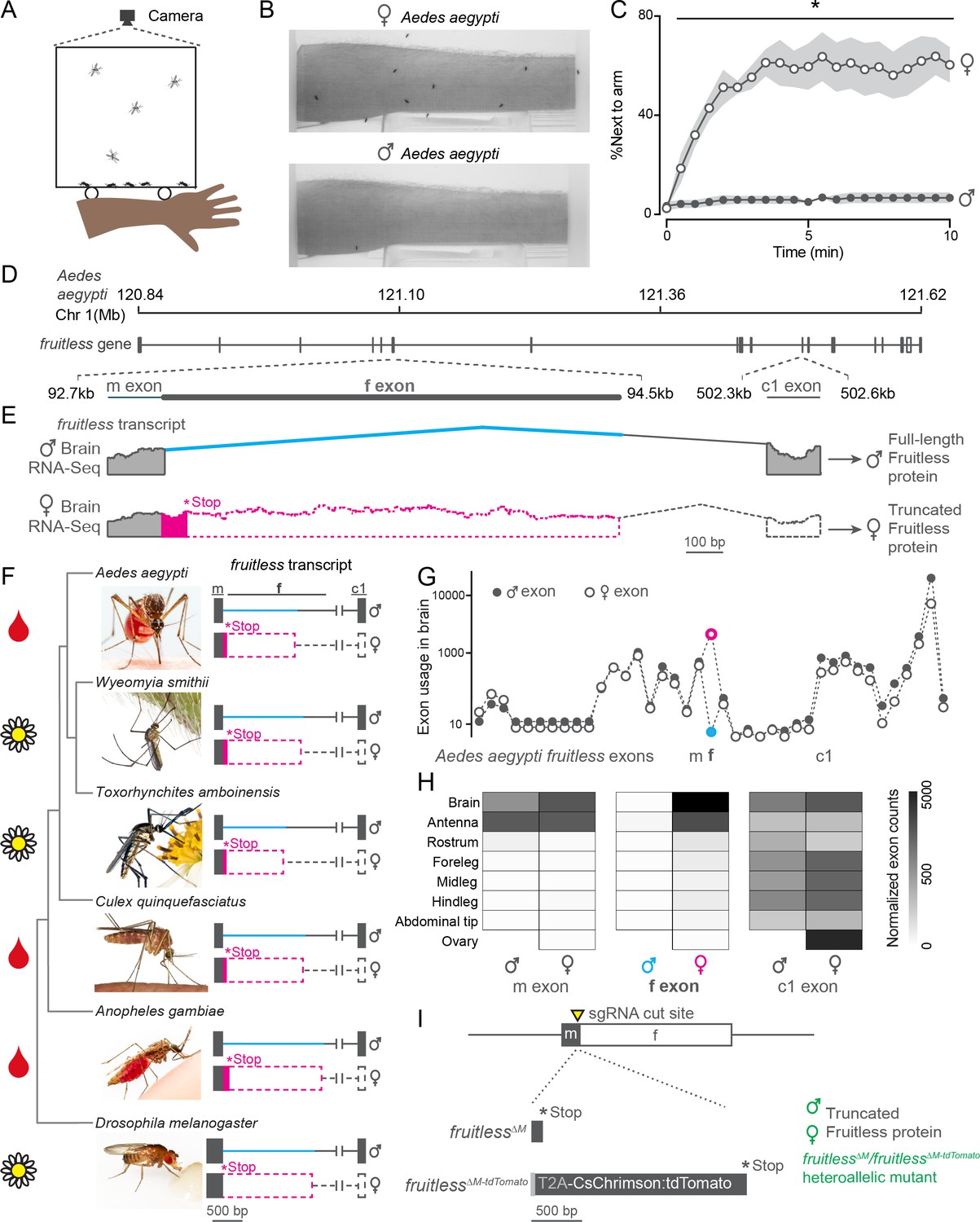
Sex-specific mosquito attraction to humans and fruitless splicing.
(A, B) Arm-next-to-cage assay schematic A and image B with female (top) and male (bottom) Aedes aegypti mosquitoes. (C) Percent mosquitoes next to arm measured every 30 s. Data are mean ± s.e.m., n = 6 trials, n = 20 mosquitoes/trial; *p<0.05, Mann-Whitney test for each time point. (D, E) Schematic of Aedes aegypti fruitless genomic locus D and sex-specific splicing region with RNA-seq read evidence E. (F) Phylogeny of mosquito species with outgroup Drosophila melanogaster, with conserved fruitless exon structure inferred from de novo transcriptome assembly. In E, F, coding and non-coding exons are represented by filled and open dashed bars, respectively. Toxorhynchites rutilus and Culex salinarius images were used to represent Toxorhynchites amboinensis or Culex quinquefasciatus, respectively. See acknowledgments for photo credits. Cartoons indicate blood-feeding (blood drop) and non-blood-feeding (flower) species. (G, H) Aedes aegypti fruitless exon usage based on male and female RNA-seq data (normalized counts) from the indicated tissue plotted for each exon G and m, f, and c1 exons H n = 3–4 independent RNA-seq replicates (Matthews et al., 2016). (I) Schematic of generation of fruitless∆M and fruitless∆M-tdTomato mutants.
-
Figure 1—source data 1
Source data for Figure 1.
- https://cdn.elifesciences.org/articles/63982/elife-63982-fig1-data1-v3.xlsx
To ask if fruitless splicing was conserved across mosquitoes, we sequenced RNA from male and female brains of five different species and assembled de novo transcriptomes for each sex. Three of these species are important arboviral disease vectors because their females blood feed on humans, whereas the two other species only feed on plants (Bradshaw et al., 2018; Zhou et al., 2014; Figure 1F). We identified orthologues of fruitless in each species and found that all had conserved ‘m’ and ‘c1’ exons and distinct ‘f’ exons. Fruitless was sex-specifically spliced in each of these species with a female-specific ‘f’ exon and early stop codon, predicted to produce a full-length fruitless protein only in males.
We used CRISPR-Cas9 genome editing (Kistler et al., 2015) to disrupt P1 neural-specific fruitless transcripts in Aedes aegypti to investigate a possible role of fruitless in sexually dimorphic mosquito behaviors. We generated two alleles, fruitless∆M, which introduces a frameshift that is predicted to produce a truncated protein in males, and fruitless∆M-tdTomato, in which the fruitless gene is disrupted by a knocked-in CsChrimson:tdTomato fusion protein (Figure 1I). In both alleles, the protein is truncated before the downstream BTB and zinc-finger domains. The fruitless∆M-tdTomato line allowed us to visualize cells that express the fluorescent tdTomato reporter under the control of the endogenous fruitless regulatory elements. To control for independent background mutations, we used the heteroallelic fruitless∆M/fruitless∆M-tdTomato mutant strain in all subsequent behavior assays (Figure 1I). In this heteroallelic mutant, fruitless P1 transcripts are disrupted in both males and females. Since full-length fruitless protein is male-specific, we expected that only fruitless∆M/fruitless∆M-tdTomato male mosquitoes would display altered behavioral phenotypes.
Sexually dimorphic expression of fruitless in the mosquito brain and antenna
In D. melanogaster, P1 fruitless transcripts are expressed in several thousand cells comprising about ~2% of the neurons in the adult brain (Stockinger et al., 2005). To examine the distribution of cells expressing fruitless in male and female Aedes aegypti mosquitoes, we carried out whole mount brain staining to reveal the tdTomato marker expressed from the fruitless locus. Fruitless >tdTomato is expressed in a large number of cells in both male and female brains (Figure 2A–F), as well as in the ventral nerve cord (Figure 2—figure supplement 1A–B). Fruitless >tdTomato expressing cells innervate multiple regions of the mosquito brain, including the suboesophageal zone, the lateral protocerebral complex, and the lateral horn. These areas have been implicated in feeding (Jové et al., 2020), mating (Seeholzer et al., 2018), and innate olfactory behaviors (Datta et al., 2008) respectively, and also receive projections from fruitless-expressing neurons in Drosophila (Seeholzer et al., 2018; Stockinger et al., 2005). The projections of fruitless >tdTomato neurons are dramatically sexually dimorphic, with denser innervation in the female suboesophageal zone and the male lateral protocerebral complex (Figure 2A–F). We did not detect any gross anatomical differences between heterozygous and heteroallelic fruitless mutant male brains or the pattern of fruitless >tdTomato expression (Figure 2B,C,E,F). We cannot exclude the possibility that there are subtle differences that can only be observed with sparse reporter expression in subsets of cells.
Figure 2 with 1 supplement see all
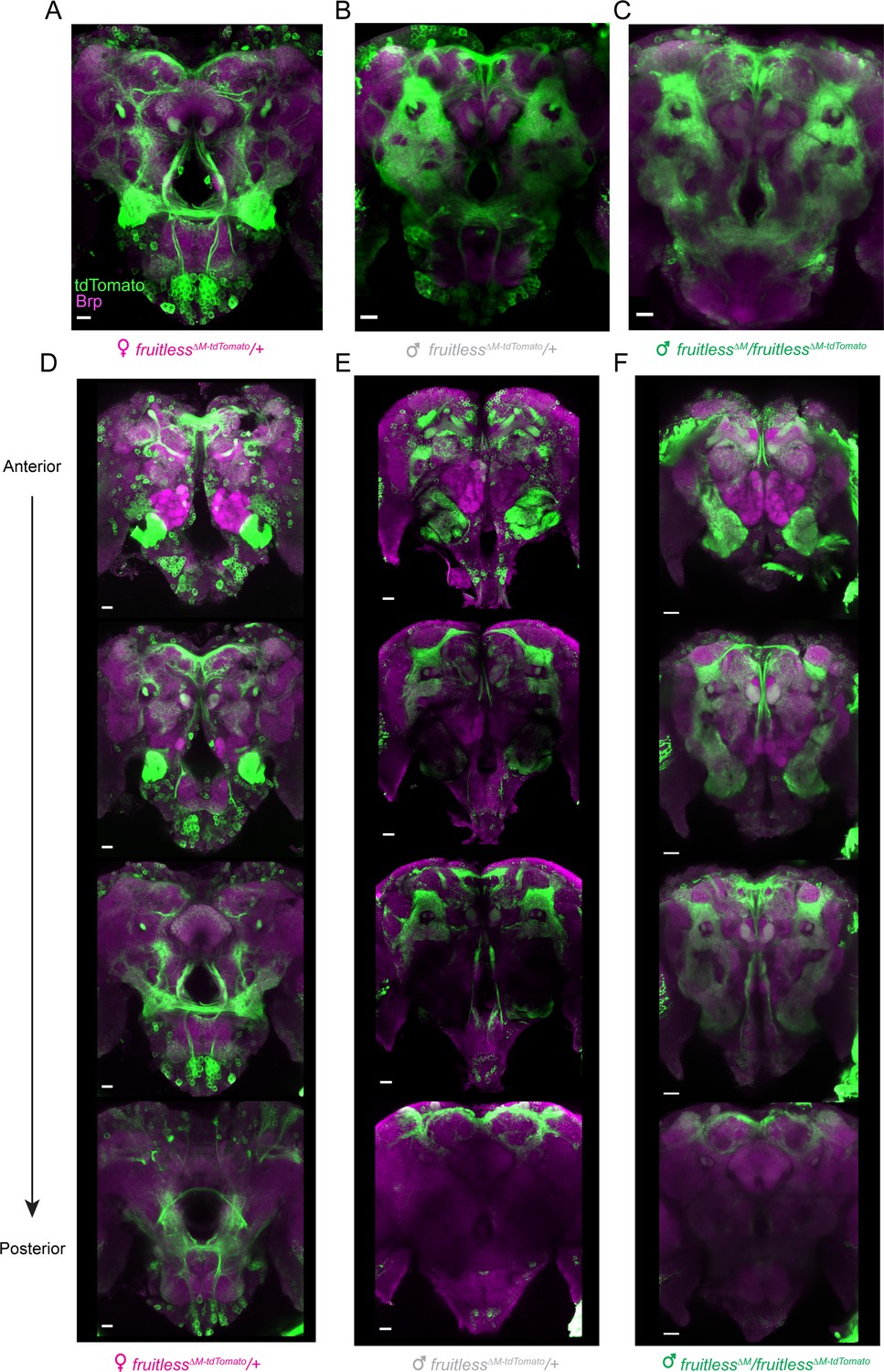
Expression of fruitless in the mosquito brain.
(A–F) Confocal images of brains of the indicated genotypes showing fruitless >tdTomato (green) and Brp (magenta) expression. Scale bars, 20 µm. (D–F) Top-to bottom images are optical sections of the same brain, arranged from anterior to posterior.
We also examined fruitless expression in the periphery. Odors are sensed by olfactory sensory neurons in the mosquito antenna, and each type of neuron projects to a single glomerulus in the antennal lobe of the mosquito brain (Figure 3A). We found that, as is the case in Drosophila (Stockinger et al., 2005), fruitless >tdTomato is expressed in olfactory sensory neurons in the antenna of both male and female mosquitoes, and that some of these neurons co-express the olfactory receptor co-receptor Orco (Figure 3B–E). Fruitless >tdTomato labels a subset of glomeruli in the antennal lobe, with females having about twice as many positive glomeruli compared to males of either genotype (Figure 3F–L). There was no difference in the number of fruitless >tdTomato labeled glomeruli between wild-type and fruitless mutant males (Figure 3F), suggesting that fruitless does not control sexual dimorphism in the number of glomeruli labeled by fruitless >tdTomato.
Figure 3
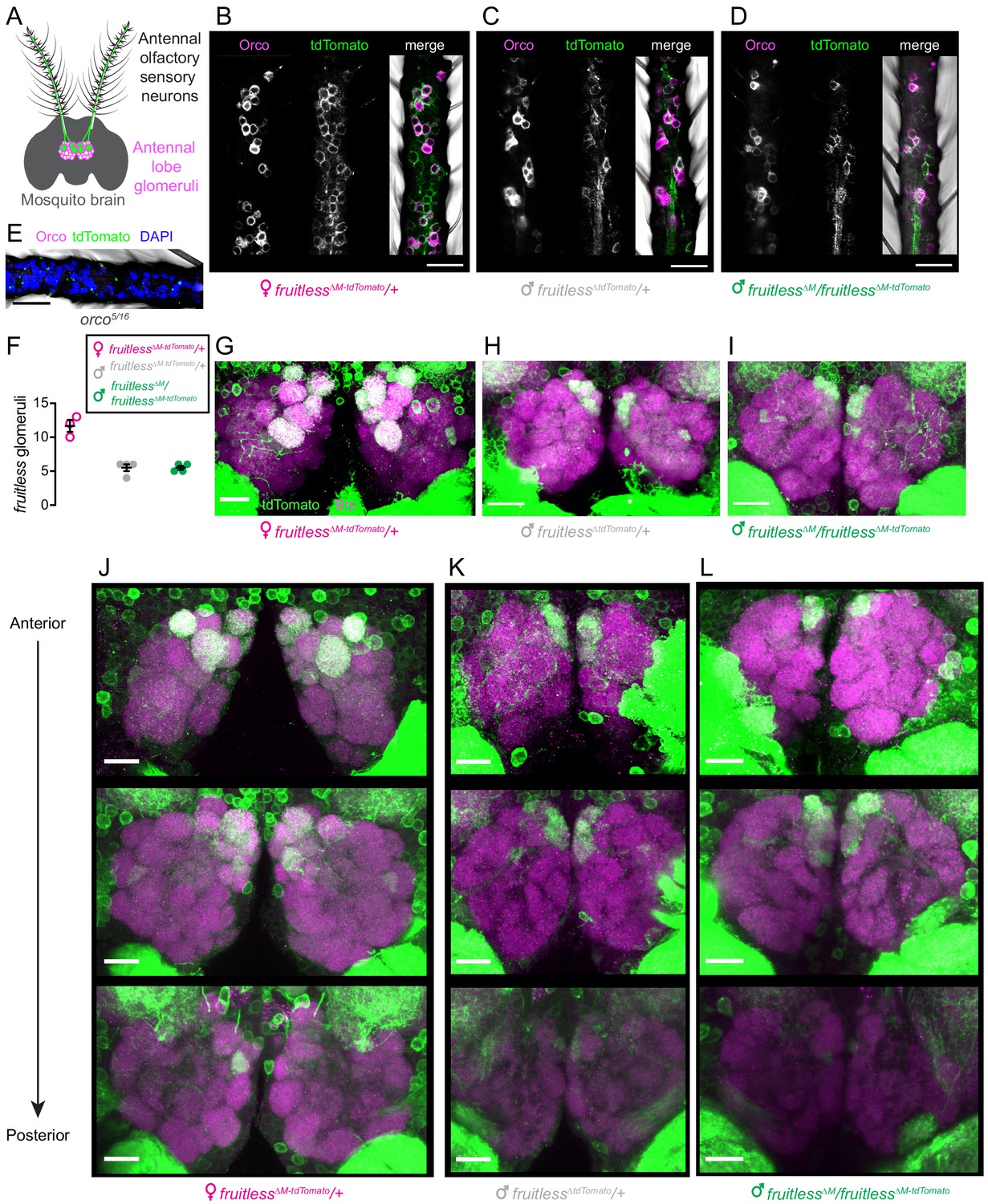
Expression of fruitless in the mosquito olfactory system.
(A) Schematic of antennal olfactory sensory neurons and their projections to the antennal lobe of the mosquito brain. (B–D) Confocal images of antennae of the indicated genotypes with fruitless >tdTomato (green) and Orco (magenta) expression. (E) Confocal image of orco mutant antenna, as negative control for Orco and tdTomato antibodies, with DAPI (blue). (F) Number of antennal lobe glomeruli labeled by fruitless >tdTomato in the indicated genotypes. (G–I) Confocal images of antennal lobes of the indicated genotypes with fruitless >tdTomato (green) and Brp (magenta) expression. (J–L) Confocal images of antennal lobes of the indicated genotypes showing fruitless >tdTomato (green) and Brp (magenta) expression. Top-to bottom images are optical sections of the same lobes, arranged from anterior to posterior. All scale bars, 20 µm.
-
Figure 3—source data 1
Source data for Figure 3.
- https://cdn.elifesciences.org/articles/63982/elife-63982-fig3-data1-v3.xlsx
Fruitless mutant males gain attraction to live human hosts
Given the broad neural expression and sexual dimorphism in fruitless circuits, we asked if fruitless mutant males showed any defects in sexually dimorphic feeding and mating behaviors. Since only female mosquitoes have the anatomical capacity to pierce skin and artificial membranes (Jové et al., 2020; Klowden, 1995), we developed a feeding assay in which both females and males are able to feed from warmed liquids through a net without having to pierce a membrane to access the meal (Figure 4A). Both wild-type males and females reliably fed on sucrose and did not feed on water. Only wild-type females fed on blood. Even when warm blood was offered and available to males for ready feeding, they still did not find it appetizing (Figure 4B). Fruitless mutant males fed similarly to their wild-type male counterparts on all meals, suggesting that this behavioral preference is not under the control of fruitless in males (Figure 4B).
Figure 4
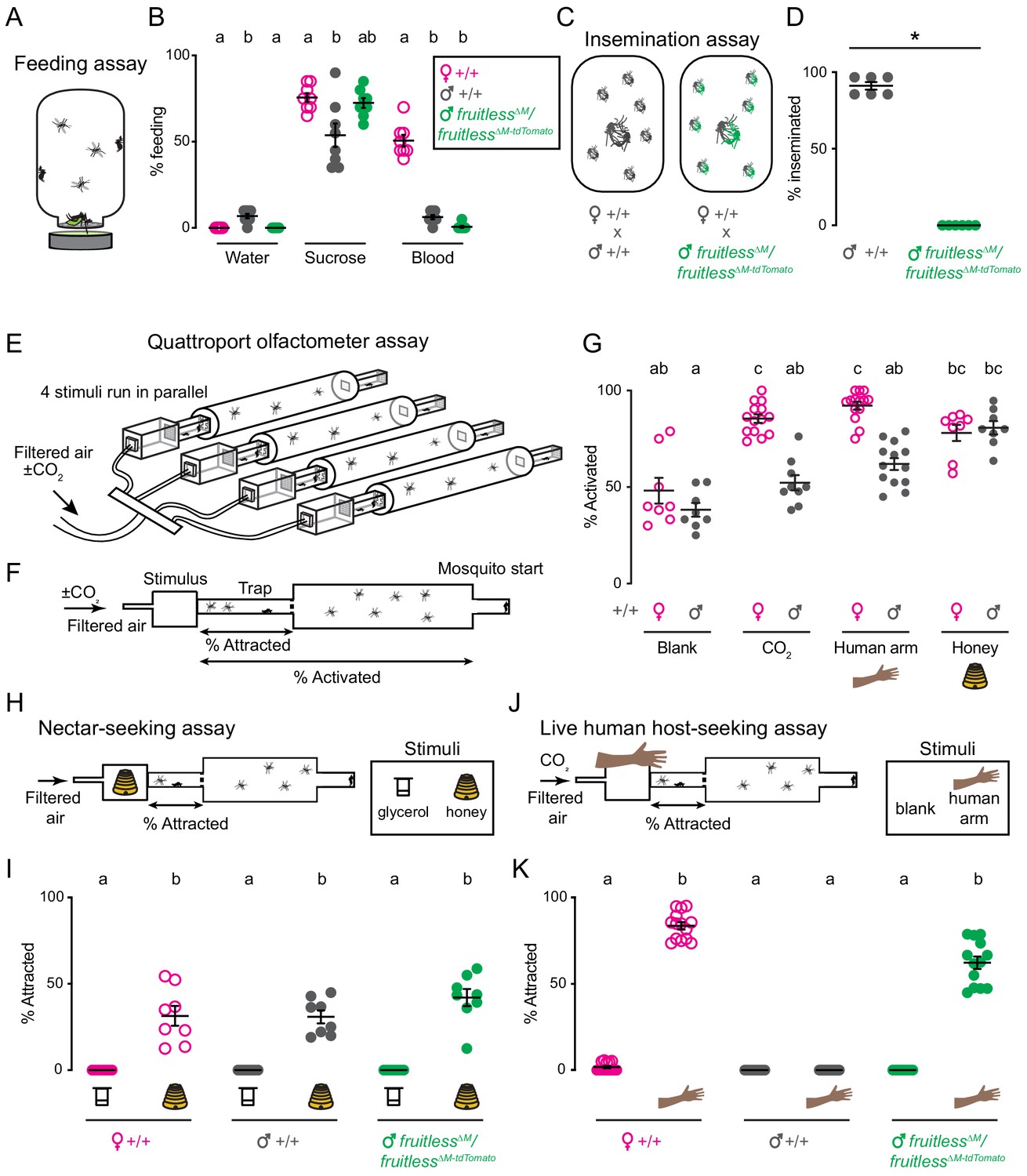
Male fruitless mutant mosquitoes gain attraction to a live human host.
(A) Feeding assay schematic. (B) Feeding on indicated meal (n = 8 trials/meal; n = 20 mosquitoes/trial). (C) Insemination assay schematic. (D) Insemination of wild-type females by males of the indicated genotype (n = 6 trials/male genotype, n = 20 females/trial; *p=0.0022, Mann-Whitney test). (E) Schematic of Quattroport assay, highlighting ability to run multiple stimuli and genotypes simultaneously. (F) Side-view schematic of Quattroport, highlighting close-range (attraction) and long-range (activation) metrics (G) Percent activated animals, n = 8–14 trials/group, n = 17–28 mosquitoes/trial. (H, J) Quattroport assay schematic for nectar-seeking H and live human host seeking J. 1% CO2 is added to the airstream in the live human host seeking assay J. (I, K) Percent of attracted animals (n = 8–14 trials per group, n = 17–28 mosquitoes/trial). Data in B, D, G, I, K are mean ± s.e.m. In B, I, G, K, data labeled with different letters are significantly different from each other (Kruskal-Wallis test with Dunn’s multiple comparisons, p<0.05). In B, comparisons are made between genotypes for each meal. In I, K, comparisons are made between all genotypes and stimuli.
-
Figure 4—source data 1
Source data for Figure 4.
- https://cdn.elifesciences.org/articles/63982/elife-63982-fig4-data1-v3.xlsx
Because fruitless plays a key role in male courtship and mating in Drosophila, we asked if it is similarly required in Aedes aegypti. Since mosquitoes show extremely rapid in-flight mating behavior that is completed in less than 30 s, it is difficult to directly observe or quantify (Hartberg, 1971). We used previously developed insemination assays (Degner and Harrington, 2016; Duvall et al., 2017) to quantify the ability of males to successfully mate (Figure 4C). We found that fruitless mutant males appeared to contact females but were unable to successfully inseminate wild-type females (Figure 4D). This mating failure is consistent with the established role of fruitless in Drosophila male sexual behavior (Demir and Dickson, 2005; Ryner et al., 1996).
We then turned to innate olfactory behaviors that govern the search for nectar, which is used as a source for metabolic energy by both males and females, and blood, which is required only by females for egg production. Consistent with the use of these meals, nectar-seeking behavior is not sexually dimorphic, but human host-seeking behavior is sexually dimorphic.
To measure these behaviors, we adapted the Uniport olfactometer (Liesch et al., 2013), which is only able to test one stimulus at a time, and developed the Quattroport, an olfactometer that tests attraction to four separate stimuli in parallel (Figure 4E). The Quattroport measures both the activation, the participation of the animals in the assay, and attraction, short range attraction to the stimulus (Figure 4F). In control experiments, we examined activation responses of wild-type male and females offered a blank, CO2, a human arm, or the floral odor of honey. While males and females showed equivalent activation with a blank and honey, females were more strongly activated to the host-related cues of CO2 and the human arm (Figure 4G). To model nectar-seeking behavior, we used honey as a floral odor and glycerol as a control odor as previously described (Figure 4H; DeGennaro et al., 2013). There was no difference in nectar-seeking as defined by attraction in the Quattroport between wild-type females, males, and fruitless mutant males (Figure 4I).
We next used the Quattroport with a live human host as a stimulus (Figure 4J). As expected, wild-type females robustly and reliably entered traps in response to a live human forearm. In contrast, zero wild-type males entered the trap, consistent with our observations in the arm-next-to-cage assay (Figure 1A–C). If fruitless function in Aedes aegypti were limited to mating and aggression as it is in Drosophila, we would expect fruitless mutant males to show no interest in a live human host. Unexpectedly, fruitless mutant males were as attracted to a live human host as wild-type females (Figure 4K). This indicates that fruitless males have gained the ability to host-seek, displaying the signature sexually dimorphic behavior of the female mosquito.
Olfactory and not heat cues attract fruitless mutant males to hosts
A live human arm gives off multiple sensory cues that are known to attract female mosquitoes, the most salient of which are body odor and heat. Fruitless mutant males might be attracted by heat alone or only the human odor, or to the simultaneous presentation of both cues. To disentangle the contribution of these complex sensory cues to the phenotype we observed, we tested the response of fruitless mutant males to each cue in isolation. We first used a heat-seeking assay (Corfas and Vosshall, 2015; McMeniman et al., 2014) to present heat to mosquitoes in the absence of human odor (Figure 5A). Neither fruitless mutant nor wild-type males were attracted to the heat cue at any temperature (Figure 5B). In contrast, wild-type females showed typical heat-seeking behavior that peaked near human skin temperature (Figure 5B).
Figure 5 with 2 supplements see all
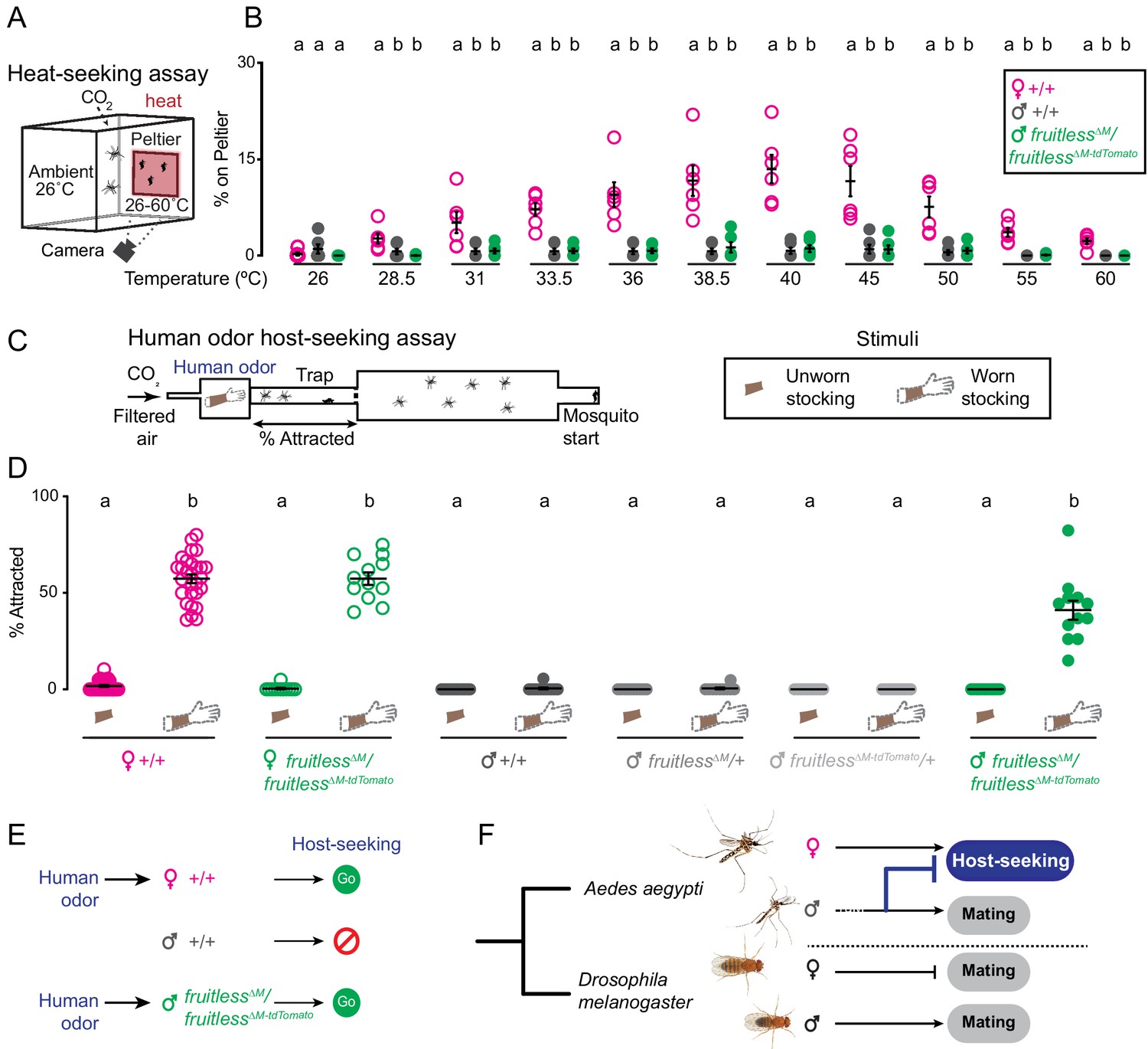
Olfactory cues selectively drive male fruitless mutant attraction to humans.
(A) Heat-seeking assay schematic. A 20 s pulse of 10% CO2 is added to the assay. (B) Percent of animals on Peltier. Data are mean ± s.e.m., n = 6 trials/temperature, n = 50 mosquitoes/trial. Data labeled with different letters are significantly different from each other, within each temperature. (C) Schematic of human odor host-seeking assay (left) and stimuli (right). (D) Percent of attracted animals. Data are mean ± s.e.m., n = 8–14 trials/group, n = 17–28 mosquitoes/trial. (E–F) Summary of results and model of gain of fruitless function in Aedes aegypti. Photo credit: Aedes aegypti (Alex Wild); D. melanogaster (Nicolas Gompel). In B, D, data labeled with different letters are significantly different from each other (Kruskal-Wallis test with Dunn’s multiple comparisons, p<0.05). In B, comparisons are made between genotypes at each temperature. In D, comparisons are made across all genotypes and stimuli.
-
Figure 5—source data 1
Source data for Figure 5.
- https://cdn.elifesciences.org/articles/63982/elife-63982-fig5-data1-v3.xlsx
To ask if fruitless mutant males are attracted to human host odor alone, we collected human scent on nylon stockings and presented this stimulus in the Quattroport (Figure 5C) to both male and female mosquitoes. Although wild-type males and heterozygous fruitless mutant males showed no response to human odor, wild-type females and fruitless∆M/fruitless∆M-tdTomato females were strongly attracted to human odor (Figure 5D). Normal host-seeking in fruitless mutant females is expected since full-length fruitless protein is translated only in males. These females also showed normal blood-feeding, egg-laying, and mating behaviors (Figure 5—figure supplement 1A–C), confirming our prediction that fruitless acts specifically in the male. We attempted to test the effect of forcing male fruitless splicing on female host-seeking (Figure 5—figure supplement 2A–C), but found that these animals were inviable due to blood-feeding and egg-laying defects (Figure 5—figure supplement 2D–I, Supplementary file 1). We then returned to the fruitless mutant males. Remarkably, heteroallelic fruitless mutant males were strongly attracted to human scent, at levels comparable to wild-type females (Figure 5D). These results demonstrate that fruitless mutant males have gained a specific attraction to human odor, which drives them to host-seek.
Discussion
Only female Aedes aegypti mosquitoes host-seek, and we have shown that mutating fruitless reveals an attraction to human odor in the male mosquito (Figure 5E). Previously, fruitless was shown to be required for male mating behavior in both Drosophila (Demir and Dickson, 2005) and Bombyx silkmoths (Xu et al., 2020). Our work demonstrates that in Aedes aegypti mosquitoes, fruitless has acquired a novel role in inhibiting female host-seeking behavior in the male (Figure 5F). Interestingly, fruitless also acts to suppress female-specific aggressive behaviors in male Drosophila in addition to its role in promoting male-specific courtship and aggression (Vrontou et al., 2006), suggesting a common theme where this gene can repress specific aspects of female-specific behavior. We cannot exclude the possibility that fruitless had a broader ancestral role in repressing male-specific host-seeking or feeding but consider this extremely unlikely given the rarity of sexually dimorphic feeding behaviors relative to sexually dimorphic mating behaviors.
Our results suggest that the neural circuits that promote female attraction to human scent are latent in males and suppressed by expression of fruitless either during development, or during adulthood. This is in contrast to a model where the ability to host-seek develops exclusively in females. Since males are able to host-seek in the absence of fruitless, other components of the sex-determination pathway do not intrinsically regulate the development and function of brain circuits controlling host-seeking behavior, even though this behavior is normally sex-specific. The concept that a latent sex-specific behavior can be revealed by knocking out a single gene was elegantly demonstrated in the mouse (Mus musculus). Only male mice court and initiate sexual contact with females and yet knocking out the Trpc2 gene causes female mice to display these male-specific behaviors (Kimchi et al., 2007).
There are field reports of Aedes aegypti males being collected near human hosts (Hartberg, 1971), which the experimenters interpreted as male Aedes aegypti attraction to humans. We note, however, that these field experiments did not control for the presence of females, suggesting that the collected males may have been attracted to the female mosquitoes that attempt to bite humans. In our well-controlled laboratory assays, we were unable to find any evidence of strong attraction to humans in mosquito males at close-range (Figure 1C, Figure 4K) or at long distances (Figure 4G). In these same assays, wild-type male mosquitoes showed strong attraction to floral cues (Figure 4I). We cannot exclude that male mosquitoes in the field show some attraction to a human host, but suggest that any attraction in males would be weaker than in wild-type females, or the attraction we demonstrate in fruitless mutant males here.
Host-seeking is the first step in a complex sequence of behaviors that lead to blood-feeding. After detecting and flying toward a human host, the female mosquito must land on the human, pierce the skin, and ultimately engorge on blood (Bowen, 1991). We have shown that fruitless has evolved to control sexual dimorphism in one module of this specialized behavior, the ability to host-seek. Females integrate multiple sensory cues to identify and approach human hosts, and we show that fruitless controls the response to just one of those cues, human odor. Sexual dimorphism in thermosensation, or in subsequent feeding behaviors does not appear to be controlled by fruitless in the male mosquito, since neither wild-type nor mutant fruitless males will drink warm blood. The modular genetic organization of mosquito behavior sparks intriguing parallels to other complex sexually dimorphic behaviors like mouse parenting (Kohl et al., 2018). To be effective parents, female mice must build nests, and then retrieve, groom, and nurse their pups. In the deer mouse Peromyscus, the conserved peptide vasopressin has evolved to control nest building (Bendesky et al., 2017). In both Peromyscus and Aedes, a conserved gene has gained control over a single aspect of a complex behavior. It has been hypothesized that certain classes of genes like neuromodulators and transcription factors are more likely to underlie phenotypic differences between species (Bendesky and Bargmann, 2011; Martin and Orgogozo, 2013; Tosches, 2017), and our study demonstrates that this is true even for an entirely novel behavior.
Where in the nervous system is fruitless required to suppress host seeking in male mosquitoes? fruitless might function in the antenna to modulate the detection of human odor in male mosquitoes, perhaps by tuning the functional or anatomical properties of olfactory sensory neurons. Such a role has been recently demonstrated in the aging-dependent sensitization of the male Drosophila antennal response to pheromones (Sethi et al., 2019; Zhang and Su, 2020; Zhao et al., 2020). Alternatively, both wild-type males and females might detect human odor, and fruitless could function in the central brain to reroute these signals to drive different motor outputs, as has been demonstrated with the sexually dimorphic response to Drosophila pheromones (Datta et al., 2008; Kohl et al., 2013; Ruta et al., 2010). To distinguish between these two models, we would require significant advances in technology and mosquito genetics, including a fruitless driver line to image neural responses to human odor. Despite significant effort, we were unable to generate a viable fruitless driver line both because of tight genetic linkage of fruitless to the sex-determining M locus (Hall et al., 2015) and because gene-targeted females failed to blood-feed and were therefore sterile (Supplementary file 1). To explore central brain fruitless+ circuits, we would need to be able to subset expression to label and drive reporters or rescue fruitless expression in sparse populations of neurons, a technology that is still out of reach. Advances in mosquito genetic tools, such as the successful implementation of orthogonal transcriptional activator reagents, combined with sparse labeling approaches will be required to gain mechanistic insight into fruitless function within mosquito host-seeking circuits. We note that these advances were not trivial in D. melanogaster, requiring efforts from multiple laboratories over the past decade (Datta et al., 2008; Kohl et al., 2013; Ruta et al., 2010), and expect that the generation of these tools and the subsequent characterization of the circuit will be significantly more challenging in the mosquito, a non-model organism.
Our work suggests that fruitless has evolved the novel function of enforcing female-specific host-seeking while maintaining its presumably ancestral male mating function. How might fruitless have evolved to control host-seeking? One possibility is that non-sex-specific host-seeking neural circuits first emerged in the ancestral mosquito, and then secondarily began to express fruitless to suppress the development or adult function of host-seeking circuits specifically in males. Another possibility is that mosquitoes duplicated and co-opted the ancestral fruitless-expressing mating neural circuits and retuned the inputs and outputs into this circuit to drive host-seeking. Circuit duplication is one of the mechanisms by which neural circuits are proposed to evolve (Tosches, 2017) and has been demonstrated in the case of vocal learning (Chakraborty and Jarvis, 2015) and in the evolution of cerebellar nuclei (Kebschull et al., 2020). In these duplicated mosquito circuits, fruitless function would have switched from promoting mating to inhibiting host-seeking in males. We speculate that both possibilities allow for fruitless to control both sex-specific host-seeking and mating behaviors, and identification and molecular profiling of the fruitless cells controlling host-seeking and mating will help distinguish between these models. Our work highlights fruitless as a potential means to investigate the circuit basis of Aedes aegypti host seeking, a behavior that is responsible for infecting millions of people with life-threatening pathogens.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene (Aedes aegypti) | fruitless | RefSeq | LOC5567734 | |
| Gene (Anopheles gambiae) | fruitless | VectorBase | AGAP000080 | |
| Gene (Culex quinquefasciatus) | fruitless | RefSeq | CPIJ010853 | |
| Gene (Wyeomyia smithii) | fruitless | This paper | Figure 1—source data 1 | |
| Gene (Toxorhynchites amboinensis) | fruitless | This paper | Figure 1—source data 1 | |
| Strain, strain background (Aedes aegypti, both sexes) | LVP-IB12 | BEI resources | MRA-735 | |
| Strain, strain background (Anopheles gambiae, both sexes) | G3 | Flaminia Catteruccia (Harvard University) | ||
| Strain, strain background (Culex quinquefasciatus, both sexes) | JHB | BEI resources | NR-43025 | |
| Strain, strain background (Wyeomyia smithii, both sexes) | PB | William Bradshaw and Christina Holzapfel (University of Oregon) | ||
| Strain, strain background (Toxorhynchites amboinensis, both sexes) | Laurence Zwiebel (Vanderbilt University) | |||
| Genetic reagent (Aedes aegypti, both sexes) | fruitless∆M | This paper | fruitless mutant strain; eggs available on request | |
| Genetic reagent (Aedes aegypti, both sexes) | fruitless∆M-tdTomato | This paper | fruitless mutant strain; eggs available on request | |
| Antibody | Mouse monoclonal anti-Apocrypta bakeri Orco antibody | Vanessa Ruta (The Rockefeller University) (PMID:30111839) | 15B2 | IF(1:50) |
| Antibody | Rabbit polyclonal anti-RFP antibody | Rockland | Cat# 600-401-379, RRID:AB_2209751 | IF(1:200) |
| Antibody | Mouse monoclonal anti-Brp antibody | DSHB | Cat# nc82, RRID:AB_2314866 | IF(1:5000) |
| Antibody | Goat polyclonal anti-mouse Alexa Fluor 488 antibody | Thermo Fisher | Cat# A-11001, RRID:AB_2534069 | IF(1:500) |
| Antibody | Goat polyclonal anti-mouse Alexa Fluor 555 Plus antibody | Thermo Fisher | Cat# A32732, RRID:AB_2633281 | IF(1:500) |
| Antibody | Goat polyclonal anti-mouse Alexa Fluor 647 polyclonal antibody | Thermo Fisher | Cat# A-21235, RRID:AB_2535804 | IF(1:500) |
| Recombinant DNA reagent | Fruitless∆-T2A-QF2 (plasmid) | This paper | Plasmid used to generatefruitless∆M strain | |
| Recombinant DNA reagent | Fruitless∆-T2A-CsChrimson-TdTomato (plasmid) | This paper | Plasmid used to generatefruitless∆M-tdTomato strain | |
| Commercial assay or kit | NEBuilder HiFi DNA Assembly | NEB | NEB:E5520S | |
| Commercial assay or kit | HiScribe Quick T7 kit | NEB | NEB:E2050S | |
| Peptide, recombinant protein | Cas9 | PNABio | PNABio:CP01-200 | |
| Sequence-based reagent | sgRNA_for | This paper | PCR primer for sgRNA | GAAATTAATACGACTCACTATAGCACCGAAGGTATGTTGAGGTTTTAGAGCTAGAAATAGC |
| Sequence-based reagent | sgRNA_rev | This paper | PCR primer for sgRNA | AAAAGCACCGACTCGGTGCCACTTTTTCAAGTTGATAACGGACTAGCCTTATTTTAACTTGCTATTTCTAGCTCTAAAAC |
| Software, algorithm | STAR v 2.5.2a | RNAseq alignment | ||
| Software, algorithm | Trinity v 2013-03-25 | De novo assembly | ||
| Software, algorithm | Prism v8 | GraphPad | Prism 8 | statistics |
| Software, algorithm | MacVector v15.0.3 | MacVector | MacVector | Plasmid cloning |
| Other | DAPI stain | Sigma | D9542 | 1:10,000 |
| Other | Quattroport | This paper | https://github.com/VosshallLab/Basrur_Vosshall2020; Basrur, 2021; (copy archived atswh:1:rev:529b0ba393bac09207d298e73e425b452338e876) |
Mosquito rearing and maintenance
Request a detailed protocolAedes aegypti wild-type laboratory strains (Liverpool-IB12) were maintained and reared at 25–28°C, 70–80% relative humidity with a photoperiod of 14 hr light: 10 hr dark (lights on at 7 a.m.) as previously described (DeGennaro et al., 2013). All behavioral assays were performed at these conditions of temperature and humidity. Adult females were blood-fed on mice for stock maintenance and on human subjects for initial stages of mutant generation. Anopheles gambiae (G3 strain), Wyeomyia smithii (PB strain), Toxorhynchites amboinensis, and Culex quinquefasciatus (JHB strain) were reared in similar conditions, following previously described protocols for each species (Bradshaw et al., 2018; Werling et al., 2019; Zhou et al., 2014). Adult mosquitoes of each species were provided constant access to 10% sucrose.
RNA-sequencing
Request a detailed protocolSeven- to 14-day-old mosquitoes of each species were cold-anesthetized and kept on ice for up to 1 hr or until dissections were complete. Brains were dissected in ice-cold RNase-free phosphate-buffered saline (PBS) (Invitrogen AM9625) on ice, moved into a microfuge tube with forceps, and immediately snap frozen in a cold block (Simport S700-14) chilled to −80°C on dry ice. Each sample group was dissected in parallel to avoid artefacts and batch effects, and five brains were used per sample. Dissected tissue was stored at −80°C until RNA extraction. RNA extraction was performed using the PicoPure Kit (ThermoFisher #KIT0204) following the manufacturer’s instructions, including DNase treatment. Samples were run on a Bioanalyzer RNA Pico Chip (Agilent #5067–1513) to determine RNA quantity and quality. Libraries were prepared using the Illumina TruSeq Stranded mRNA kit #20020594, following manufacturer’s instructions. Library quantity and quality were evaluated using High Sensitivity DNA ScreenTape Analysis (Agilent #5067–5585) prior to pooling. Barcoded samples from all non-Aedes tissues were pooled in an equal ratio before distributing the pool across two sequencing lanes. Sequencing was performed at The Rockefeller University Genomics Resource Center on a NextSeq 500 sequencer (Illumina). All reads were 2 × 150 bp. Data were de-multiplexed and delivered as fastq files for each library. Sequencing reads have been deposited at the NCBI Sequence Read Archive (SRA) under BioProject PRJNA612100.
Fruitless splicing analysis
Request a detailed protocolReads from individual Aedes libraries were mapped to the AaegL5 genome (Matthews et al., 2018) using STAR version 2.5.2a with default settings (Dobin et al., 2013). Raw counts were used for differential splicing analysis in Aedes aegypti using DEXSeq version 1.32.0 (Anders et al., 2012) as per author instructions. For the other mosquito species without genomes or incomplete genome annotations, we assembled sex-specific de novo transcriptomes using Trinity version 2013-03-25 with default settings (Grabherr et al., 2011). We then searched for fruitless orthologues in each species using BLAST 2.6.0 (Altschul et al., 1990), aligned hits to Aedes aegypti fruitless P1 transcripts using MacVector version 15.0.3, and picked the best match for each exon, species, and sex. Fruitless exon sequences are found in Figure 1—source data 1.
Fruitless∆M and fruitless∆M-tdTomato strain generation
Request a detailed protocolThe fruitless gene was targeted using CRISPR-Cas9 methods as previously described (Kistler et al., 2015). Gene-targeting reagents were injected into wild-type Liverpool-IB12 embryos at the Insect Transformation Facility at the University of Maryland Institute for Bioscience and Biotechnology Research. For each line, either 2000 or 1000 embryos were injected with 600 ng/µL plasmid, 300 ng/µL Cas9 protein (PNABio CP01-200), and 40 ng/µL sgRNA. Proper integration was confirmed in each strain using polymerase chain reaction (PCR) and sequencing. Animals were then back-crossed to wild-type Liverpool-IB12 for at least four generations.
All homology arms for homology-directed integration were isolated by PCR using Liverpool-IB12 genomic DNA. sgRNA DNA template was prepared by annealing oligonucleotides as previously described (Kistler et al., 2015). In vitro transcription of sgRNA template was performed using HiScribe Quick T7 kit (New England Biolabs #E2050S) following the manufacturer’s directions and incubating for 4 hr at 37°C. Following transcription and DNAse treatment for 15 min at 37°C, sgRNA was purified using Ampure RNAse-free SPRI beads (Beckman-Coulter #A63987) and eluted in Ultrapure water (Invitrogen #10977–015). For all plasmids, fragments were generated by PCR from the indicated template with the indicated primers (Supplementary file 2) and assembled using NEBuilder HiFi DNA Assembly (NEB E5520S). Plasmids were transformed into NEB competent cells (NEB C2987I), purified with the NucleoBond Xtra Midi Endotoxin Free kit (Clontech 740420.50), and sequence verified.
The fruitless∆M mutant was generated in the course of attempting to generate a fruitless QF2 knock-in mutant (Supplementary file 1) (see below). One of the families had viable 3xP3-dsRed-positive offspring and an out-of-frame QF2 insertion, which was predicted to produce a truncated fruitless protein in males. This was the fruitless∆M mutant allele we used in the study.
The fruitless∆M-tdTomato knock-in/knock-out strain was generated by inserting a cassette containing T2A followed by CsChrimson fused to the fluorescent protein tdTomato and the 3xP3-EYFP strain marker. We obtained two independent viable lines and selected one for use in this study. We used the CsChrimson:tdTomato protein expressed from the fruitless locus in fruitless∆M-tdTomato animals as a marker for fruitless expression in these studies.
CsChrimson is a red-light-activated cation channel, and we originally generated this animal with the intention of optogenetically manipulating behavior. However, CsChrimson:tdTomato intrinsic fluorescence was not visible under a confocal microscope, even at high laser intensities, and required immunofluorescent amplification in all our images. When animals were fed with retinal, the necessary co-factor which was absent in all other experiments, and we attempted to substitute human odor and CO2 with optogenetic activation of fruitless+ neurons in a blood-feeding assay, we did not observe increased feeding or any other behaviors (preliminary data not shown). Although we were unable to see evidence of CsChrimson activity in these optogenetic experiments, potential background levels of CsChrimson-driven activation of fruitless-expressing neurons is an important concern to rule out when considering the behavioral data in this paper. We note that animals were not fed trans-retinal, the necessary cofactor for CsChrimson activity. Fruitless∆M-CsChrimson-tdTomato/+ males and females were able to mate normally, and fruitless∆M-CsChrimson-tdTomato/+ females show normal blood-feeding and egg-laying behavior (Figure 5—figure supplement 1A–C). We have not examined fruitless∆M /∆M animals due to the difficulty of obtaining these animals without molecular genotyping of each individual. However, given the weak expression of CsChrimson and the robust behavior of heterozygote animals, we consider it unlikely that this allele is significantly affecting mosquito behavior.
Attempted generation of fruitless QF2/QF2w knock-in mutants
Request a detailed protocolWe attempted to generate fruitless P1-specific driver lines by knocking in a cassette containing the ribosomal-skipping peptide T2A followed by the transcriptional activator QF2 (Riabinina et al., 2015), with 3xP3-dsRed as an insertion marker as previously described (Matthews et al., 2019). In this knock-in/knock-out strain, we aimed to disrupt the fruitless gene as well as generate a driver line that would allow us to label and manipulate fruitless-expressing neurons. We recovered seven independent 3xP3-dsRed positive G1 families. However, all females with one copy of the correct integration did not blood-feed after many attempts using multiple different human hosts. Males with one copy of this insertion did not mate with wild-type females. Since blood-meals are required for Aedes aegypti egg-development, this line could not be maintained. We next tried to knock-in the weaker QF2w transcriptional activator, and recovered six independent families, all of which showed the same blood-feeding and mating defects (Supplementary file 1). We speculate that toxicity of QF2 or QF2w may affect the function or viability of fruitless-expressing neurons, leading to the behavioral defects we observed. The cause of Q-system toxicity, even attenuated from Q to QF2 to QF2w is unknown (Riabinina et al., 2015). We speculate that this toxicity is unrelated to the fruitless locus, because fruitless∆M/+ animals had no phenotype as heterozygotes, unlike fruitless∆QF2/+ and fruitless∆QF2w/+ animals.
Attempted generation of sex-switched fruitless mutants
Request a detailed protocolTo ask if fruitless protein was sufficient to inhibit host-seeking behavior in females, we attempted to force females to express male fruitless protein by deleting the female exon of fruitless P1 transcripts and forcing male fruitless splicing in female brains (Supplementary file 1). For the fruitless∆F line, embryos were injected with 300 ng/µL Cas9 protein, 125 ng/µL oligonucleotide with template repairing the splice site, 40 ng/µL each of two sgRNAs targeting the beginning and end of the female-specific exon. We recovered multiple G1 animals with the correct integration, as verified by PCR and sequencing. Male fruitless splicing in fruitless∆ females was verified with reverse-transcription PCR (data not shown). G2 fruitless∆F females did not fully blood-feed or lay eggs even though they were successfully inseminated by wild-type males (Figure 5—figure supplement 2D–I). It was therefore impossible to maintain these lines. We do not know if the blood-feeding defect was due to a failure to respond to the host or some other behavioral or anatomical defect. Since fruitless is tightly linked to the male-determining locus, it was not an option to maintain this targeted allele in males. Integrations on the male chromosome would yield ~1/500 females with the recombinant allele, and integrations on the female chromosome yield inviable females and rare recombinant males. In either scenario, the fruitless∆F insertion is unmarked and would need to be followed by PCR genotyping. We also attempted to generate a line where we both deleted the female fruitless exon and knocked-in an intronic 3xP3 fluorescent marker, which would allow us to maintain this line in males and use the marker to select rare recombinants for behavioral analysis. However, females with this integration did not have any behavioral phenotypes, suggesting that the intronic 3xP3 marker interfered with regular fruitless splicing in both males and females. These difficulties precluded any further investigation of the phenotype of expressing full-length fruitless protein in females.
Antibody staining – brain whole mounts
Request a detailed protocolDissection of adult brains and immunostaining was carried out as previously described (Jové et al., 2020; Matthews et al., 2019). Six- to 14-day-old mosquitoes were anesthetized on ice and decapitated. Heads were fixed in 4% paraformaldehyde (Electron Microscopy Sciences 15713 s), 1X Ca+2, Mg+2 free PBS (Thermo 14190144), 0.25% Triton X-100 (Sigma 93443), and nutated for 3 hr at 4°C. Brains were then dissected and placed in cell-strainer caps (Falcon #352235) in a 24-well plate. All subsequent steps were performed on a low-speed orbital shaker. Brains were washed for 15 min at room temperature in 1x PBS with 0.25% Triton X-100 (0.25% PBT) at least six times. Brains were permeabilized with 4% Triton X-100 with 2% normal goat serum (Jackson ImmunoResearch #005-000-121) in PBS at 4°C for 2 days. Brains were washed for 15 min at least sixtimes with 0.25%PBT at room temperature. Brains were incubated in 0.25% PBT plus 2% normal goat serum with primary antibodies at the following dilutions: rabbit anti-RFP (Rockland 600-401-379) 1:200 and mouse anti-Drosophila melanogaster Brp (nc82) 1:5000. The nc82 hybridoma developed by Erich Buchner of Universitätsklinikum Würzburg was obtained from the Developmental Studies Hybridoma Bank, created by the NICHD of the NIH and maintained at The University of Iowa, Department of Biology, Iowa City, IA 52242. Primary antibodies were incubated for 2 nights at 4°C and then washed at least six times for 15 min with 0.25% PBT at room temperature. Brains were incubated with secondary antibody for 2 nights at 4°C with secondary antibodies at 1:500% and 2% normal goat serum. Secondary antibodies used were goat anti-rabbit Alexa Fluor 555 (Thermo A32732) and goat anti-mouse Alexa Fluor 647 (Thermo A-21235). Brains were then washed for 15 min at least six times with 0.25% PBT at room temperature and mounted in Slowfade Diamond (Thermo S36972) using #1.5 coverslips as spacers before confocal imaging.
Antibody staining – antennal whole mounts
Request a detailed protocolThis protocol was adapted from a Drosophila embryo staining protocol (Manning and Doe, 2017). Six- to 10-day-old mosquitoes were anesthetized, decapitated, and placed in 1.5 mL 5 U/mL chitinase (Sigma C6137) and 100 U/mL chymotrypsin (Sigma CHY5S) in 119 mM NaCl, 48 mM KCl, 2 mM CaCl2, 2 mM MgCl2, 25 mM HEPES buffer on ice. Male heads were incubated for 5 min on a ThermoMixer (Eppendorf 5382000023), and 25 min in a rotating hybridization oven, and female heads were incubated for 10 min on the ThermoMixer and 50 min in rotating oven, all at 37°C. Heads were then rinsed once and fixed in 4% paraformaldehyde, 1X Ca+2, Mg+2 free PBS, and 0.25% Triton X-100 for 24 hr at room temperature on a rotator. All subsequent 4°C steps used a nutator, and room temperature steps used a rotator. Heads were washed for 30 min at room temperature at least three times in 1X PBS with 0.03% Triton X-100 (0.03% PBT). Antennae were then dissected into 0.5-mL microfuge tubes and dehydrated in 80% methanol/20% DMSO for 1 hr at room temperature. Antennae were washed in 0.03% PBT for 30 min at room temperature, and blocked/permeabilized in 1X PBS, 1% DMSO (Sigma 472301), 5% normal goat serum, 4% Triton X-100 for 24 hr at 4°C. Antennae were washed for 30 min at least five times with 0.03% PBT, 1% DMSO at room temperature, and then moved to primary antibody in 1X PBS, 1% DMSO, 5% normal goat serum, 0.03% Triton X-100 for 72 hr at 4°C. Primary antibodies used were mouse anti-Apocrypta bakeri Orco monoclonal antibody #15B2 (1:50 dilution, gift of Joel Butterwick and Vanessa Ruta), and rabbit anti-RFP (1:100, Rockland 600-401-379). Orco monoclonal antibody specificity was verified in Aedes aegypti by staining orco mutant antennae, which showed no staining (data not shown). Antennae were washed for 30 min at least five times with 0.03% PBT, 1% DMSO at room temperature, and then washed overnight in the same solution. Antennae were then moved to secondary antibody (1:200) and DAPI (1:10000, Sigma D9542) in 1X PBS, 1% DMSO, 5% normal goat serum, 0.03% Triton X-100 for 72 hr at 4°C. Secondary antibodies used were goat anti-mouse Alexa Fluor 488 (Thermo A-11001) and goat anti-rabbit Alexa Fluor 555 Plus (Thermo A32732). Antennae were washed for 30 min at least five times with 0.03% PBT, 1% DMSO at room temperature, and then washed overnight in the same solution. Antennae were rinsed in 1X PBS, rinsed three times in Slowfade Diamond (Thermo S36972), and mounted in Slowfade Diamond.
Confocal image acquisition
Request a detailed protocolImages were acquired with a Zeiss Axio Observer Z1 Inverted LSM 880 NLO laser scanning confocal microscope (Zeiss) with either 25x/0.8 NA (whole brains) or 40x/1.4 NA (antennal lobes, antennae) immersion-corrected objective at a resolution of 1024 × 1024 or 2048 × 2048 (brains) or 3024 × 1024 (antennae) pixels. Confocal images were processed in ImageJ (NIH).
Arm-next-to-cage assay
Request a detailed protocolThis assay was performed as described previously (DeGennaro et al., 2013). Briefly, for each trial, 20 adult mosquitoes were sorted under cold anesthesia (4°C) and placed in a cage and allowed to acclimate for 30 min. A human arm was placed 2.5 cm from one side of a standard 28 × 28×28 cm cage. Mosquitoes could not directly contact the human arm. A Logitech C920s HD Pro Webcam was positioned to take images of mosquitoes responding to the human arm. Trials ran for 10 min and images were acquired at a rate of 1 frame/s. To quantify mosquito responses, we manually counted the number of mosquitoes resting on the lower portion of the screen closest to the human arm.
Feeding assay
Request a detailed protocolMosquitoes were cold-anesthetized, and 20 mosquitoes were sorted into 250 mL bottles covered with a taut net secured with rubber bands. They were allowed to acclimate for 24 hr with access to water through cotton balls. The following meals were presented: water, 10% sucrose, or sheep’s blood (Hemostat DSB100) supplemented with 1 mM ATP (Sigma A6419). Meals were warmed to 45°C before being used in the assay. 10 mL of a given meal was pipetted into the bottle caps, animals were activated with a 30 s pulse of 4% CO2, and bottles were inverted on top of the caps. Mosquitoes were allowed to feed on each meal through the net for 10 min and were then anesthetized at 4°C and scored as fed if any level of feeding was observed, as assessed by visual inspection of the abdomen of the animal.
Insemination assay
Request a detailed protocolMosquitoes were separated by sex at the pupal stage and sex was confirmed within 24 hr of eclosion. For each trial, 10 female Liverpool-IB12 virgin mosquitoes were crossed to 11 virgin male mosquitoes of either Liverpool-IB12 or fruitless∆M/fruitless∆M-tdTomato genotype in a bucket cage for 24 hr, with access to 10% sucrose. Mosquitoes were then anesthetized at 4°C, females separated from males, and female spermathecae were dissected to score for insemination as a sign of successful mating (Degner and Harrington, 2016). Control virgin females were dissected in parallel to verify absence of insemination.
Quattroport olfactometer
Request a detailed protocolDetails of Quattroport fabrication and operation are available at https://github.com/VosshallLab/Basrur_Vosshall2020. Briefly, the Quattroport consists of four tubes, each with its own stimulus box, trap, and mosquito start chamber. There are adjustable gates between each chamber. The stimulus was placed upstream of a trap, and mosquitoes are prevented from contacting the stimulus by a mesh barrier. In each trial, four stimuli were run in parallel, with the positions of stimuli randomized and rotated between each trial. Air was filtered and pumped into each box, either at a final concentration of 1% CO2 (for host-seeking assays) or at ambient CO2 (honey-seeking assays). For all assays, ~20 mosquitoes were sorted and placed into canisters the day of behavior. Mosquitoes were allowed to acclimate in the assay for 10 min, then exposed to the stimulus for 30 s, after which gates were opened and animals allowed to fly for 5 min. After this time, gates were closed and mosquitoes were counted to quantify the percent of mosquitoes in the trap.
For honey assays, 3- to 7-day-old mosquitoes were fasted for 24 hr before the experiment by replacing 10% sucrose with a water source. CO2 was not added for honey assays. Either 1 g of leatherwood honey (Tasmanian Honey Company, Tasmania, Australia) or glycerol (Sigma G5516) was applied to a 55 mm diameter Whatman filter paper circle (GE Healthcare, Buckinghamshire, UK) and placed in a Petri dish.
For host-seeking assays, mosquitoes were allowed access to sucrose before the experiment. A final concentration of 1% CO2 was supplied in the airstream for the duration of the 5 min 30 s assay in all host-seeking assays (for both human forearm/odor stimuli and blank/unworn nylon controls). For live human host-seeking assays, a human subject placed their forearm on an acrylic box, exposing a 2.5 × 5 cm rectangle of skin to the airstream.
For human odor host-seeking assays, the same human subject wore a tan nylon sleeve (L'eggs Women's Comfortable Everyday Knee Highs Reinforced Toe Panty Hose, modified with scissors to remove the toe area) on their forearm. A second black nylon sleeve was worn on top of the tan nylon odor sampling sleeve to protect it from external odors. After 6 hr of continuous wear, the black nylon sleeve was discarded, and the tan nylon sleeve was frozen at −20°C. Nylons were used within 1 week of being worn. On the day of the assay, nylons were thawed for at least 1 hr at room temperature. A 10 × 14 cm piece of the sleeve was presented with the skin-contacting surface facing upward in the stimulus box along with CO2. Unworn nylons were similarly frozen, thawed, and cut to serve as negative controls.
Heat-seeking assay
Request a detailed protocolExperiments were performed as previously described (Corfas and Vosshall, 2015; McMeniman et al., 2014). Briefly, 45–50 mosquitoes were fasted for 3 hr before the experiment by replacing 10% sucrose with a water source and were then transferred into a custom-made Plexiglass box (30 × 30×30 cm), with carbon-filtered air pumped continuously into the box via a diffusion pad installed on the ceiling of the enclosure. All stimulus periods lasted 3 min and were presented on a single Peltier element (6 × 9 cm, Tellurex) covered with a piece of standard white letter-size printer paper (NMP1120, Navigator) cut to 15 × 17 cm and held taut by a magnetic frame. CO2 pulses (20 s, to >1000 ppm above background levels) were added to the air stream and accompanied all stimulus period onsets. Mosquito landings on the Peltier were monitored by fixed cameras (FFMV-03M2M-CS, Point Grey Research) with images acquired at 1 Hz. Images were analyzed using custom MATLAB scripts to count mosquito landings within a fixed target region. Mosquito occupancy on the Peltier was quantified during seconds 90–180 of each stimulus period.
Quantification, statistical analysis, and reproducibility
Request a detailed protocolAll statistical analyses were performed using GraphPad Prism Version 8. Data collected as percent of total are shown as mean ± s.e.m. Details of statistical methods are reported in the figure legends. Preliminary experiments were used to assess variance and determine sample sizes before carrying out experiments. Typically, sample sizes were n = 6–14 groups of 10-20 mosquitoes in behavioral assays. We used similar sample sizes for all experiments where the same variable was being compared. No data were excluded from this study. Since mosquito behavior is variable, all olfactometer experiments with a human arm were carried out repeatedly to assess the effect of external environmental conditions on behavior. No experiments were performed on days when < 40% of wild-type females responded to a live human arm. No data met these exclusion criteria. All attempts at replication over multiple days were successful. We carried out all experiments with different groups of animals hatched up to 4 weeks apart, and over multiple days. Several experiments were carried out repeatedly over the course of this study, namely the wild-type female response to a live human arm in Quattroport olfactometer assays. These results were robust and reliable over the course of the many years it took to complete this study. For all experiments, mosquitoes from a cage were randomly selected and sorted by sex into groups for behavioral assays. All stimuli and genotypes were interspersed, and positions were randomized when possible. Every experiment involved replicates collected over multiple days, to ensure that there was no effect of daily environmental or experimental conditions. We also collected a similar sample size for each variable every time the experiment was run, to ensure no effect of external conditions. Blinding to genotype was performed in the heat-seeking assays. The experimenter was not blinded to genotype in the host-seeking assays because the mutant phenotype we describe is so robust it was impossible to be blinded in these assays.
Data and software availability
Request a detailed protocolAll raw data are provided in the accompanying Source Data files. Plasmids are available at Addgene (#141099, #141100). RNA-seq data are available in the Short Read Archive at Genbank (Bioproject: PRJNA612100). Details of Quattroport fabrication and operation are available at Github: https://github.com/VosshallLab/Basrur_Vosshall2020.
Data availability
All raw data are provided in the accompanying source data files. Plasmids are available at Addgene (#141099, #141100). RNA-seq data are available in the Short Read Archive at Genbank (Bioproject: PRJNA612100). Details of Quattroport fabrication and operation are available at Github: https://github.com/VosshallLab/Basrur_Vosshall2020 (copy archived at https://archive.softwareheritage.org/swh:1:rev:529b0ba393bac09207d298e73e425b452338e876).
-
NCBI BioProjectID PRJNA612100. Sex-specific mosquito brain transcriptomes.
References
-
Basic local alignment search toolJournal of Molecular Biology 215:403–410.https://doi.org/10.1016/S0022-2836(05)80360-2
-
Detecting differential usage of exons from RNA-seq dataGenome Research 22:2008–2017.https://doi.org/10.1101/gr.133744.111
-
SoftwareBasrur_Vosshall2020, version swh:1:rev:529b0ba393bac09207d298e73e425b452338e876Software Heritage.
-
Genetic contributions to behavioural diversity at the gene-environment interfaceNature Reviews Genetics 12:809–820.https://doi.org/10.1038/nrg3065
-
The fruitless gene in Nasonia displays complex sex-specific splicing and contains new zinc finger domainsMolecular Biology and Evolution 26:1557–1569.https://doi.org/10.1093/molbev/msp067
-
The sensory physiology of host-seeking behavior in mosquitoesAnnual Review of Entomology 36:139–158.https://doi.org/10.1146/annurev.en.36.010191.001035
-
Brain evolution by brain pathway duplicationPhilosophical Transactions of the Royal Society B: Biological Sciences 370:20150056.https://doi.org/10.1098/rstb.2015.0056
-
A mosquito sperm's journey from male ejaculate to egg: Mechanisms, molecules, and methods for explorationMolecular Reproduction and Development 83:897–911.https://doi.org/10.1002/mrd.22653
-
Carbon dioxide instantly sensitizes female yellow fever mosquitoes to human skin odoursJournal of Experimental Biology 208:2963–2972.https://doi.org/10.1242/jeb.01736
-
STAR: ultrafast universal RNA-seq alignerBioinformatics 29:15–21.https://doi.org/10.1093/bioinformatics/bts635
-
Some records of predaceous ceratopogoninae (Diptera)Entomologist's Monthly Magazine 56:203–205.
-
Functional conservation of the fruitless male sex-determination gene across 250 myr of insect evolutionMolecular Biology and Evolution 23:633–643.https://doi.org/10.1093/molbev/msj070
-
Full-length transcriptome assembly from RNA-Seq data without a reference genomeNature Biotechnology 29:644–652.https://doi.org/10.1038/nbt.1883
-
Observations on the mating behaviour of Aedes aegypti in natureBulletin of the World Health Organization 45:847–850.
-
Blood-feeding behavior of adult Aedes aegypti mosquitoesThe Biological Bulletin 145:127–139.https://doi.org/10.2307/1540353
-
Functional and genetic characterization of neuropeptide Y-like receptors in Aedes aegyptiPLOS Neglected Tropical Diseases 7:e2486.https://doi.org/10.1371/journal.pntd.0002486
-
How to turn an organism into a model organism in 10 ‘easy’ stepsThe Journal of Experimental Biology 223: jeb218198.https://doi.org/10.1242/jeb.218198
-
Improved and expanded Q-system reagents for genetic manipulationsNature Methods 12:219–222.https://doi.org/10.1038/nmeth.3250
-
A study of mosquito behavior an experimental laboratory study of the sexual behavior of Aedes aegypti (Linnaeus)American Midland Naturalist 40:265–352.https://doi.org/10.2307/2421604
-
The mode of action of fruitless: is it an easy matter to switch the sex?Genes, Brain and Behavior 19:e12606.https://doi.org/10.1111/gbb.12606
-
The mechanics of copulation in Aedes aegyptiThe Biological Bulletin 127:324–344.https://doi.org/10.2307/1539230
-
Optogenetic activation of the fruitless-labeled circuitry in Drosophila subobscura males induces mating motor actsThe Journal of Neuroscience 37:11662–11674.https://doi.org/10.1523/JNEUROSCI.1943-17.2017
-
Developmental and genetic mechanisms of neural circuit evolutionDevelopmental Biology 431:16–25.https://doi.org/10.1016/j.ydbio.2017.06.016
-
Mosquitoes use vision to associate odor plumes with thermal targetsCurrent Biology 25:2123–2129.https://doi.org/10.1016/j.cub.2015.06.046
-
Fruitless regulates aggression and dominance in DrosophilaNature Neuroscience 9:1469–1471.https://doi.org/10.1038/nn1809
-
Olfactory regulation of mosquito-host interactionsInsect Biochemistry and Molecular Biology 34:645–652.https://doi.org/10.1016/j.ibmb.2004.03.017
Article and author information
Author details
Maria Elena De Obaldia

Ricarda K von Heynitz
Funding
Howard Hughes Medical Institute (Vosshall-Investigator)
- Leslie B Vosshall
National Center for Advancing Translational Sciences (UL1 TR000043)
- Leslie B Vosshall
Harvey L Karp Discovery Award (Postdoctoral fellowship)
- Maria Elena De Obaldia
- Takeshi Morita
Japan Society for the Promotion of Science (JSPS Overseas Research Fellowship)
- Takeshi Morita
Helen Hay Whitney Foundation (HHW Fellowship)
- Maria Elena De Obaldia
National Center for Advancing Translational Sciences (UL1 TR001866)
- Maria Elena De Obaldia
National Institute on Deafness and Other Communication Disorders (F30DC017658)
- Margaret Herre
National Institute of General Medical Sciences (T32GM007739)
- Margaret Herre
Quadrivium Foundation (Vosshall/Herre-Investigator)
- Margaret Herre
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank Richard Benton, Josie Clowney, Lindy McBride, Vanessa Ruta, David Stern, Nilay Yapici, and members of the Vosshall Lab for comments on the manuscript; Gloria Gordon and Libby Mejia for expert mosquito rearing; Joel Butterwick and Vanessa Ruta for sharing unpublished anti-Apocrypta bakeri Orco monoclonal antibodies; the colleagues who shared mosquito species with us [William Bradshaw and Christina Holzapfel (Wyeomyia smithii); Larry Zwiebel and Jason Pitts (Toxorhynchites amboinensis); Flaminia Catteruccia (Anopheles gambiae)] and those who shared insect photos with us [Alex Wild (Aedes aegypti, Anopheles gambiae), Lawrence Reeves of Florida Medical Entomology Laboratory (Wyeomyia smithii, Toxorhynchites rutilus, Culex salinarius), Francisco Romero, Veronica Corrales-Carvajal, Carlos Ribeiro, and Nicolas Gompel (Drosophila melanogaster)]; Erich Jarvis, Vanessa Ruta, and Li Zhao for advice and discussion; Ben Matthews and Zach Gilbert for assistance in designing and testing of sgRNAs and advice on the CRISPR protocol; Ben Matthews for advice on bioinformatics; Rob A Harrell II at the Insect Transgenesis Facility at the University of Maryland for CRISPR-Cas9 injections; Jim Petrillo and Kunal Shah at the Rockefeller Precision Instrument Technology center for help in development and construction of the Quattroport assay; Meg Younger for advice on brain dissections and antibody staining; Ella Jacobs for training in spermathecae dissections; Christina Pyrgaki, Carlos Rico, and Alison North at the Rockefeller Bio-Imaging Resource Center for assistance with confocal imaging; Connie Zhao at the Rockefeller Genomics Resource Center for advice on RNA-seq library preparation and sequencing; Román Corfas for assistance with the heat-seeking assay.
Ethics
Human subjects: Blood-feeding procedures and behavioral experiments with human volunteers were approved and monitored by The Rockefeller University Institutional Review Board (IRB protocol LV-0652). Human subjects gave their written informed consent to participate.
Animal experimentation: Blood-feeding procedures with live mice were approved and monitored by The Rockefeller University Institutional Animal Care and Use Committee (IACUC protocol 17018).
Copyright
© 2020, Basrur et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 7,290
- views
-
- 820
- downloads
-
- 58
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 58
- citations for umbrella DOI https://doi.org/10.7554/eLife.63982
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Fruitless mutant male mosquitoes gain attraction to human odor
eLife 9:e63982.
https://doi.org/10.7554/eLife.63982




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}