Cis-regulatory modes of Ultrabithorax inactivation in butterfly forewings
- Department of Biological Sciences, The George Washington University, United States
- Smithsonian Tropical Research Institute, Panama
- Department of Integrative Biology, University of Texas – Austin, United States
Figures
Figure 1

Annotation of the Ubx genomic interval in four butterflies of the Nymphalinae sub-family.
(A) Genomic intervals spanning Antp, Ubx, and abd-A, featuring published transcript annotations from NCBI Reference Genomes of V. cardui and A. io, and manual re-annotations of the J. coenia and K. inachus genomes using published RNAseq dataset (see Methods). Exons are shown with coding (thick) and non-coding (thin) sections. No lincRNA:Ubx-AS5’ transcripts were detected in K. inachus. (B) Expression profiling of transcripts of the Ubx region in K. inachus, based on a reanalysis of published wing RNA-seq transcriptomes (Wang et al., 2022). Expression levels are plotted as DESeq2 normalized counts plots. Pairwise Wald tests adjusted for multiple test correction each assess differential expression between forewings and hindwings. ns: non-significant; *: p<0.05; **: p<0.01; ***: p<0.001.
Figure 2 with 1 supplement

A region of hindwing-specific chromatin-opening is bordered by a TAD BE in the last intron of Ubx.
(A) Hi-C contact heatmap in fifth instar forewings of J. coenia and TAD separation scores around Ubx. A TAD boundary element (Antp-Ubx_BE) is inferred in the last intron of Ubx (vertical dotted line). (B) Differential ATAC-seq profiles, re-analyzed from a previous dataset (Mazo-Vargas et al., 2022). Top: open-chromatin profiles of forewings (FW, magenta), and hindwings (HW, green), respectively subtracted from larval head signal (purple, negative when wing signals at background-level). Bottom: subtractive ATAC-seq profile (HW-FW) revealing hindwing-enriched chromatin in the Ubx locus. Antp-Ubx_BE is in the vicinity of an isolated region of forewing-enriched opening (blue arrowhead). (C) PhastCons genomic alignment scores, with overall alignability suggesting minimal structural variation across this interval in Lepidoptera and Trichoptera.
Figure 2—figure supplement 1
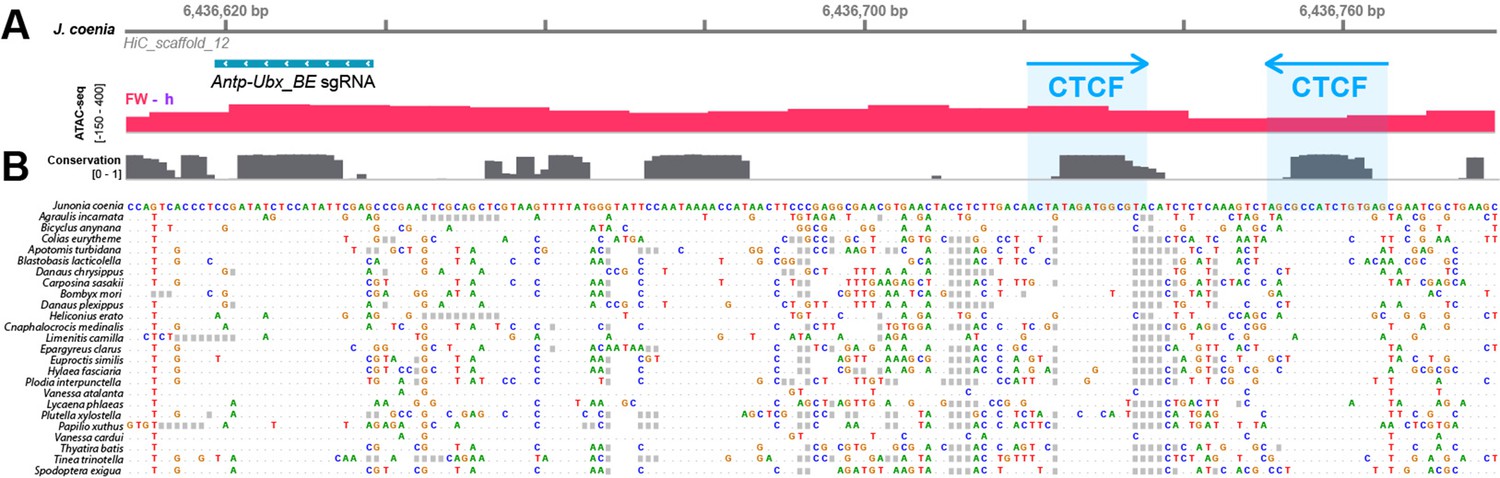
Prediction of two conserved CTCF binding sites at Antp-Ubx_BE.
(A) Sequence-level view of a 180 bp genomic interval including the Antp-Ubx_BE sgRNA (turquoise) in J. coenia, overlapping with an ATAC-seq peak of forewing-enriched chromatin opening (red). The CRISPR target is about 100 bp away from two predicted binding sites for the Drosophila CTCF insulator protein. (B) High-level of nucleotide conservation at thesgRNA site and CTCF motifs across Lepidoptera and Trichoptera representative genomes, indicative of functional constraints on these sequences. CTCGAATATGGAGATATCGGAGG.
Figure 3

Hindwing-enriched chromatin-opening around Ubx, and the Antp-Ubx_BE boundary, are both maintained in mid-pupal hindwings.
(A) Hi-C heatmap in J. coenia fifth instar larval forewings, and subtractive ATAC-seq profiles at this stage (hindwing-forewing), as expanded from Figure 2 across the Hox cluster. (B) Hi-C heatmap in J. coenia mid-pupal hindwings, and subtractive ATAC-seq profiles at this stage (forewing-hindwing). Inferred TAD boundaries are shown as vertical dotted lines. Blue arrowhead: position of the Antp-Ubx_BE sgRNA.
Figure 4 with 4 supplements

CRISPR perturbation of Antp-Ubx_BE results in FW➞HW homeoses.
(A) Antp-Ubx_BE sgRNA targeting (cyan triangle) of a FW-enriched ATAC-peak (magenta) within the Ubx last intron. (B–C) Two examples of J. coenia Antp-Ubx_BE crispants showing mosaic FW➞HW homeoses, shown in dorsal views. CL-WT: contralateral, horizontally flipped images of forewings from the same individuals. WT HW: wild type hindwings from the same individual and mutant forewing side. Both individuals show disruption of their Radial veins (R1-R5 area). The specimen shown in C displays a partial, ectopic eyespot (asterisk). (D–E) Immunofluorescent detection of the UbdA epitope (green) in fifth instar wings disks of Antp-Ubx_BE crispants, revealing ectopic antigenicity in forewings. WT forewings of similar stage, and HW from the same crispant individuals, are shown for comparison as insets. Green autofluorescence was observed in tracheal tissues. Scale bars: B-C = 500 μm; D-E = 100 μm.
Figure 4—figure supplement 1

CRISPR perturbation of the Antp-Ubx boundary element results in FW-to-HW homeosis.
(A) Example of an Antp-Ubx_BE crispant with a unilateral phenotype on the right forewing. (B) Additional examples of forewing homeoses in Antp-Ubx_BE crispant. Wing sets (forewing mKO mutants and corresponding contralateral WT) are shown with one of the wings horizontally flipped to show the mutant wings in left-to-right orientation. Arrows: small mutant clones. Asterisks: large mutant clones.
Figure 4—figure supplement 2
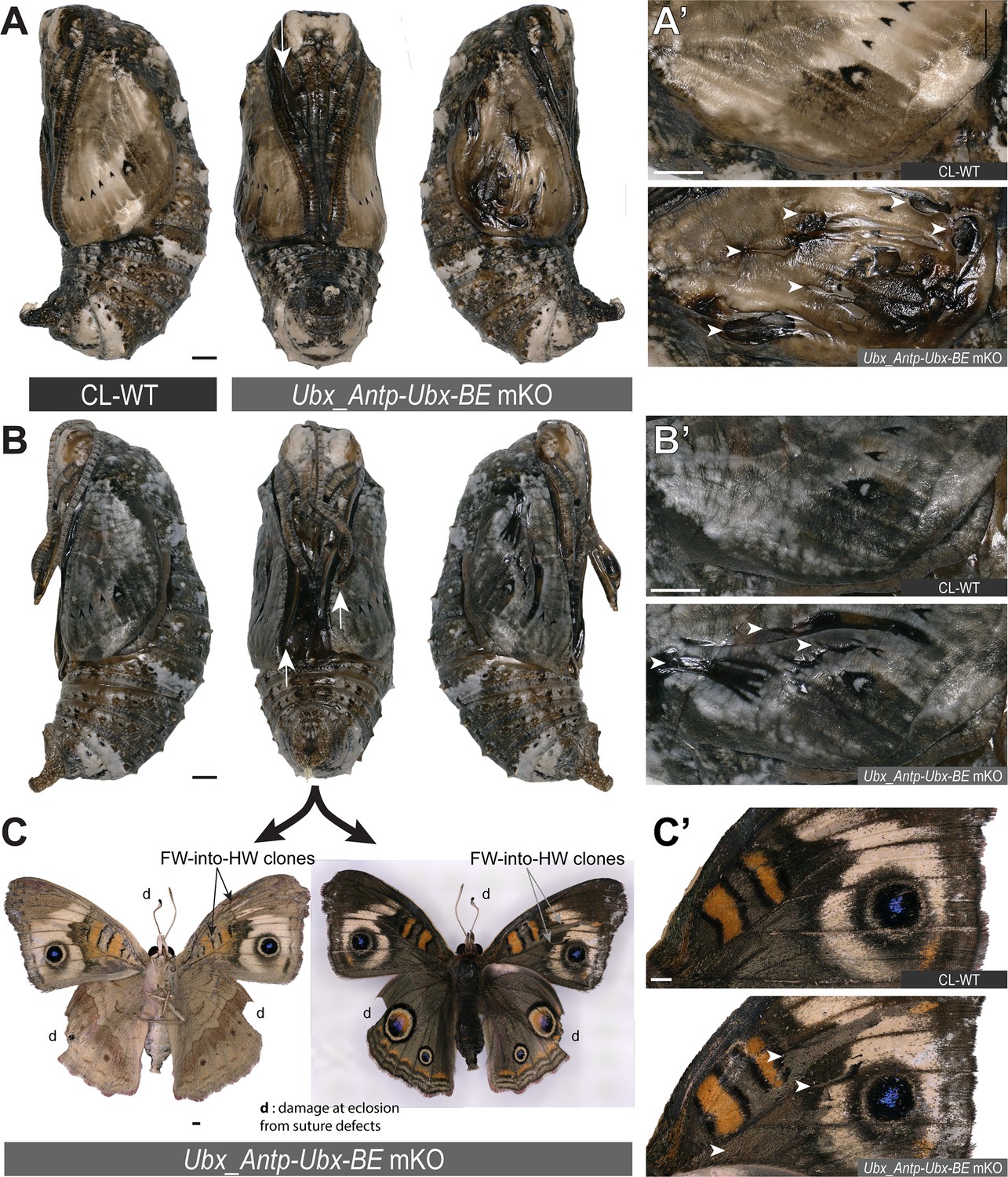
Pupal defects following FW➞HW homeosis in Antp-Ubx_BE crispants.
(A–B) Contralateral (CL) and, forewing mosaic knockout (mKO) mutants following CRISPR targeting of Antp-Ubx_BE in J. coenia. The two pupae show suture defects in the midline appendages (arrows). (A’-B’) Magnified views of the crispant forewings, showing defective cuticle (arrowheads). (C-C’) Crispant adult butterfly emerged from the pupa in panel B. White arrowheads in C’ highlight the match between dorsal forewing clones and the pupal forewing cuticle defects shown in B’. Scale bars: 1 mm.
Figure 4—figure supplement 3
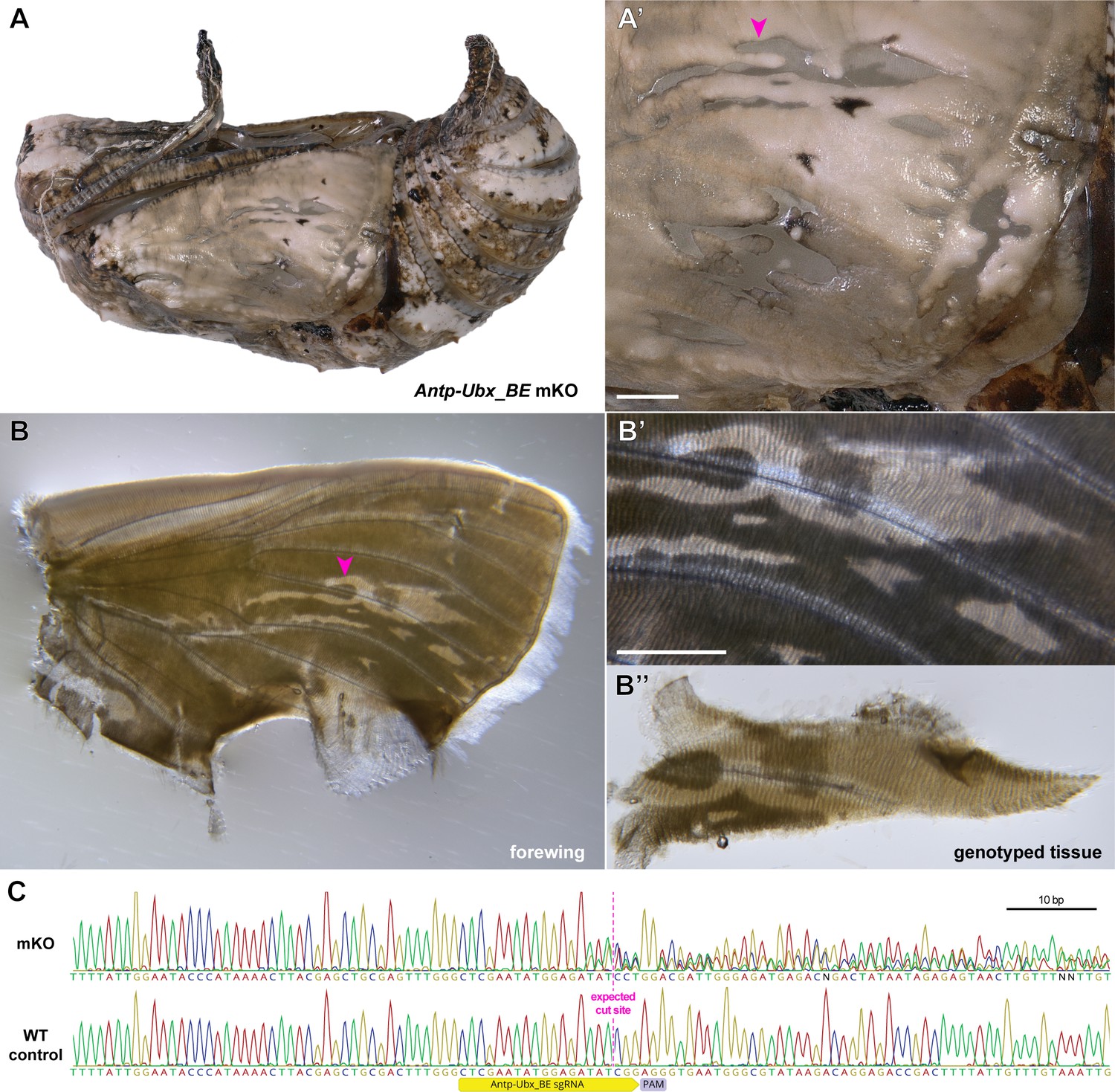
Validation of CRISPR-induced DNA lesions in an Antp-Ubx_BE crispant pupal forewing.
(A-A’) Pupal forewing cuticle phenotype of an Antp-Ubx_BE J. coenia crispant, as in Figure 4—figure supplement 2. (B-B’’) Aspect of the same forewing under trans-illumination following dissection out of the pupal case. Regions from mutant clones have a more transparent appearance. (C). Sanger sequencing of an amplicon targeting the Antp-Ubx_BE region in the mutant tissue shown in panel B”, compared to a control wing tissue, showing mixed chromatogram around the expected CRISPR cutting site due to indel mutations from non-homologous end-joining. Scale bars: 500 μm.
Figure 4—figure supplement 4
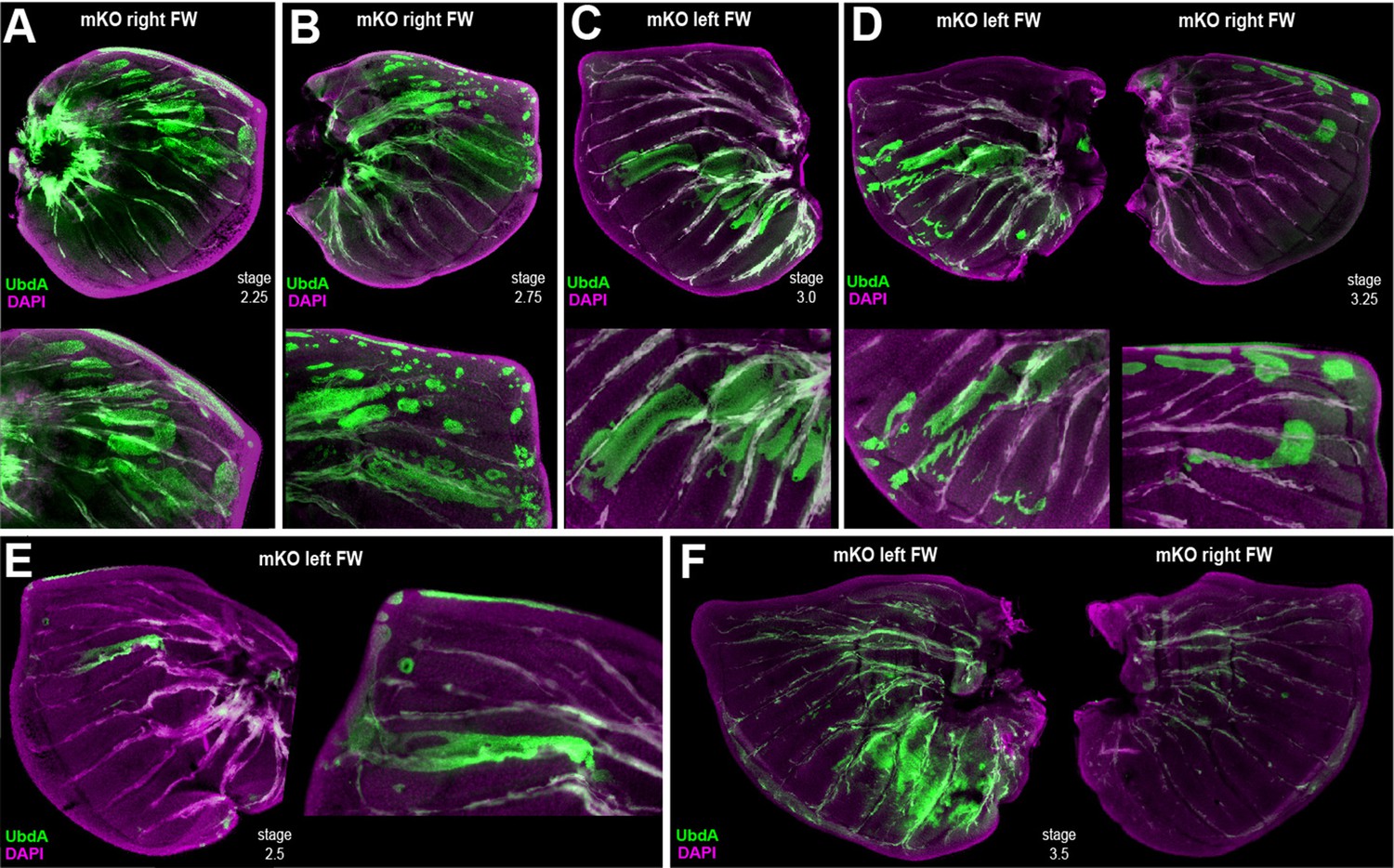
Additional examples of ectopic UbdA and FW➞HW homeosis in Antp-Ubx_BE crispant larval forewings.
(A–F) Each panel shows forewings with ectopic detection of UbdA (FP6.87 monoclonal antibody, green), dissected at the fifth instar stage. Panels D and F are wing sets from individual crispants. Panels E and C are mutant contralateral wings of the mutant forewings shown in Figure 4D and E, respectively.
Figure 5 with 1 supplement

Rare, dual homeoses obtained from CRISPR mutagenesis of the lncRNA_Ubx-IT1 5’ region.
(A) Genomic context of the sgRNA targets (here shown in J. coenia), in the promoter and first exon of the non-coding Ubx-IT1 transcript. (B–C) Dorsal and ventral views of a J. coenia crispant displaying dual homeoses, that is with both FW➞HW (presumably due to Ubx gain-of-expression), and HW➞FW clones (akin to Ubx null mutations). Insets on the right describe forewing mutant clones (IT1 mKO), in apposition to CL-WT (contralateral forewings from the same individual), and WT HW (wild type hindwings from the same individual and mutant forewing side). (D) Examples of dual homeoses obtained when targeting orthologous sites in V. cardui. Scale bars: 1 mm.
Figure 5—figure supplement 1

Additional mutant phenotypes from CRISPR-mediated interrogation of lncRNA_Ubx-IT1 5’ region in J. coenia (top) and V. cardui (bottom).
Cyan arrows: mutant clones.
Figure 6 with 1 supplement
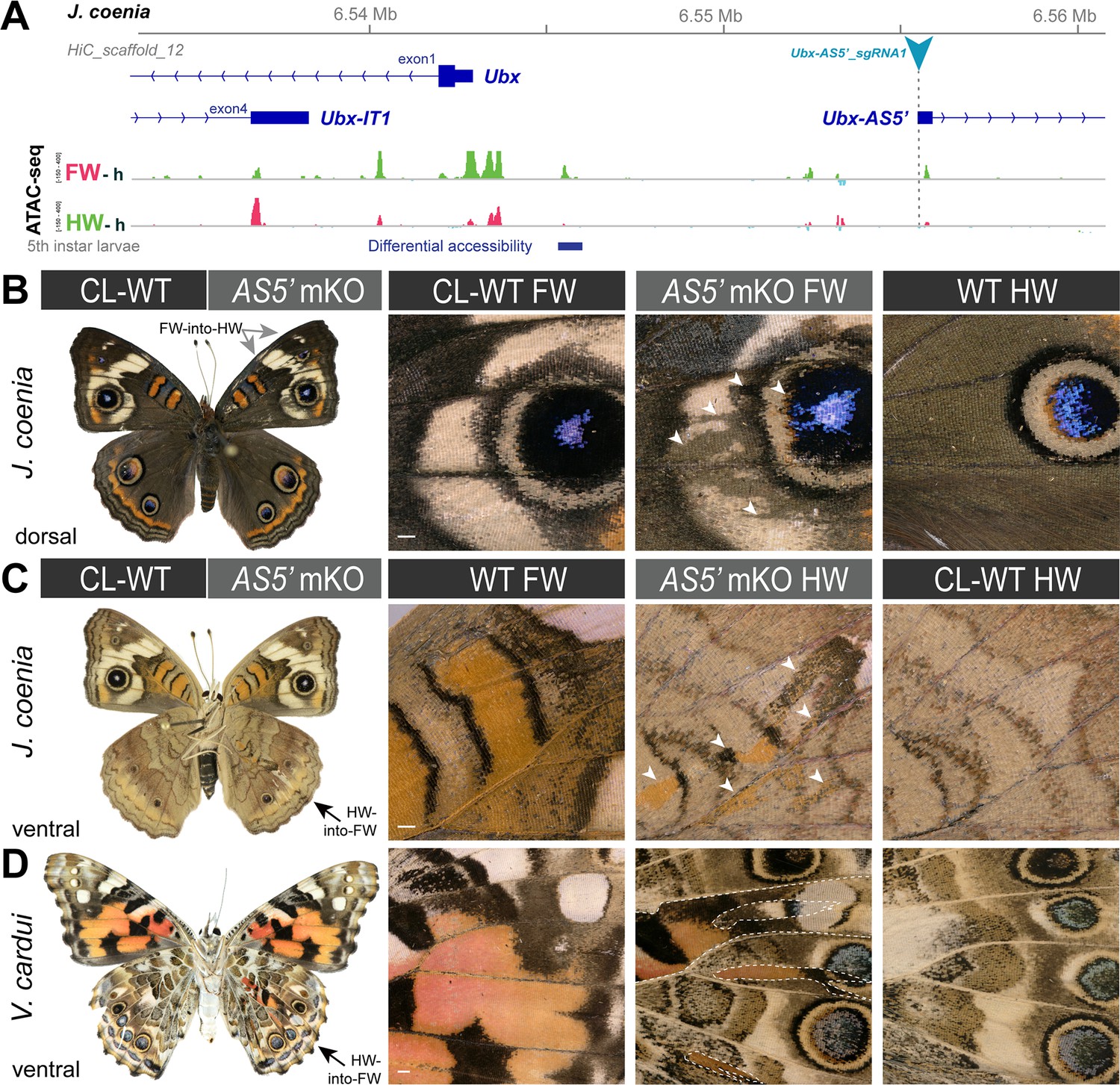
Homeoses obtained from CRISPR mutagenesis of the lncRNA Ubx-AS5’ first exon.
(A) CRISPR sgRNA targets (here shown in J. coenia), in the first exon of the non-coding Ubx-AS5’ transcript. (B) A single J. coenia crispant showed a FW➞HW transformation. Insets on the right describe forewing mutant clones (AS5’ mKO), in apposition to CL-WT (contralateral forewings from the same individual), and WT HW (wild-type hindwings from the same individual and mutant forewing side). (C–D) Examples of HW➞FW homeoses obtained in J. coenia or when targeting orthologous sites in V. cardui. Scale bars: 500 μm.
Figure 6—figure supplement 1

Additional mutant phenotypes from CRISPR-mediated interrogation of the lncRNA_Ubx-AS5’ region in J. coenia and V. cardui.
Arrows: mutant clones. Arrowheads: white eyespot foci.
Figure 7 with 2 supplements

CRISPR perturbation of Ubx CRM11 generates occasional dual homeotic phenotypes.
(A) Overview of ATAC-seq differential chromatin accessibility profiles (hindwing - head tissues, green; forewing - head tissue, magenta) across the Ubx first exon. Several regions show differential opening between wings, one of which (CRM11), was targeted here for CRISPR perturbation (sites a2 and c5 indicate sgRNA targets). (B) Dual homeosis phenotypes obtained in V. cardui following dual-targeting of UbxCRE11a2c5, including homeoses in color patterns and scale morphology. (D) Additional example of a V. cardui UbxCRE11a2c5 crispant with a forewing phenotype (gain of hindwing hair patches, arrowheads). (E) Example of mild hindwing homeoses showing a white eyespot focus on the dorsal and ventral sides. These effects were previously shown to occur in coding Ubx CRISPR knock-out experiments (Tendolkar et al., 2021). Contralateral (CL) WT wings are shown for comparison with mutant wings (B–E). Colored dashed lines: wing veins. Scale bars: 500 μm.
Figure 7—figure supplement 1

Additional mutant phenotypes from CRISPR-mediated interrogation of CRM11 in J. coenia and V. cardui show bidirectional homeoses and non-homeotic eyespot changes.
Arrows: mutant clones. Arrowheads: white eyespot foci.
Figure 7—figure supplement 2

CRISPR perturbation of the conserved Ubx_CRE11b results in HW➞FW homeoses.
(A–B) The UbxCRE11b9 sgRNA targets a hindwing-enriched ATAC peak with strong conservation across genomes from 23 Lepidoptera and 2 Trichoptera species (gray: PhastCons scores). Colored bars denote variation from the J. coenia reference. (C) Jc_UbxCRE_11b9 crispant butterflies exclusively showed HW➞FW transformed clones (blue arrows in both J. coenia and V. cardui).
Figure 8 with 2 supplements

Mosaic forewing homeoses in Heliconius butterfly spontaneous mutants.
Wild-type and mutant sides from the same individuals are shown in each panel, with one side digitally flipped to match left-to-right orientation. (A) Heliconius melpomene rosina, ventral view. Wild-caught in the Osa Peninsula (Costa Rica), October 1989. (B) Heliconius cydno galanthus, ventral view (magnified inset in B’). Stock culture from Organisation for Tropical Studies station, La Selva (Costa Rica), June 1990 (C) Heliconius himera, dorsal view (magnified inset in C’). Stock Culture in the butterfly farm Heliconius Butterfly Works in Mindo (Ecuador), March 2008.
Figure 8—figure supplement 1

Hindwing homeoses in Heliconius butterfly spontaneous mutants from pure stocks, hybrid cultures and wild-caught individuals from the L.
E. Gilbert collection (UT Austin). White arrowheads: homeotic clones including the acquisition of ventral forewing coupling scales. Asterisks: local deformation of hindwings relative to wild-type. All hindwing homeoses are ventral except in panel L. (A) Heliconius cydno galanthus x H. melpomene rosina (Costa Rica), cross J31, August 1987. (B) Heliconius cydno gustavi, captive stock from Saladito (Colombia), September 1991. (C) Heliconius melpomene madeira (Brazil) x Heliconius melpomene plesseni (Ecuador), September 2012. (D) H. m. rosina (Costa Rica) x Heliconius melpomene madeira (Brazil) x H. cydno galanthus (Costa Rica) mixed population, December 2015. (E) H. m. rosina, captive stock from Osa Peninsula (Costa Rica), September 1991. (F) Heliconius hewitsoni, captive stock from Osa Peninsula (Costa Rica), July 2005.
Figure 8—figure supplement 2

Hindwing homeoses in Heliconius butterfly spontaneous mutants from pure stocks, hybrid cultures and wild-caught individuals from the L.
E. Gilbert collection (UT Austin). White arrowheads: homeotic clones including the acquisition of ventral forewing coupling scales. Asterisks: local deformation of hindwings relative to wild-type. All hindwing homeoses are ventral except in panel L. (A) Heliconius cydno cydnides, captive stock from natural hybrid zone in Dagua Pass (Colombia), May 1989. (B) H. m. rosina (Costa Rica) x H. m. madeira (Brazil) x H. c. galanthus (Costa Rica) mixed population, June 2016. (C) H. c. galanthus x H. m. rosina crossed three times, and back to H. c. galanthus, August 2014. (D) Heliconius melpomene malleti (Ecuador) x H. m. plesseni (Ecuador) hybrid stock, 2010. (E) H. m. rosina captive stock, Costa Rica. (F) H. m. rosina captive stock, Osa Peninsula (Costa Rica), March 1987, in dorsal view.
Figure 9
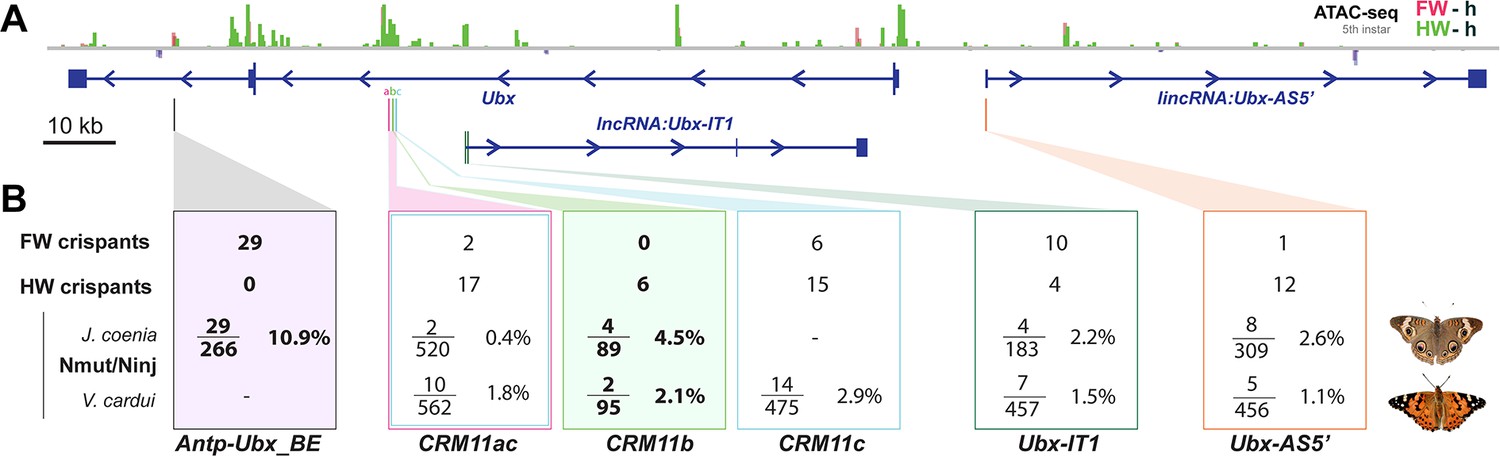
Summary of wing homeosis phenotypes obtained from mutational interrogation.
(A) CRISPR targets at non-coding regions across the Ubx region, here visualized in J. coenia. (B) Summary of injection and adult phenotype data obtained across CRISPR experiments. FW/HW crispants: total number of individuals with forewing or hindwing homeotic clones, regardless of the injected species. Individuals with dual homeosis are counted in both categories. Nmut/Ninj: number of crispants obtained (Nmut), over the number of injected embryos for each species. Bold: experiments with consistent effects in only one segment. See Table 1 for details.
Author response image 1

Validation of CRISPR-induced DNA Lesions in an Antp-Ubx_BE crispant pupat forewing.
(A-A') Pupal forewing cuticle phenotype of an Antp-Ubx_BE J. coenia crispant, as in Fig. S3. (B-B") Aspect of the same forewing under trans-illumination following dissection out of the pupal case. Regions from mutant clones have a more transparent appearance. (C). Sanger sequencing of an amplicon targeting the Antp-Ubx_BE region in the mutant tissue shown in panel B", compared to a control wing tissue, showing mixed chromatogram around the expected CRISPR cutting site due to indel mutations from non-homologous end-joining.
Tables
Table 1
CRISPR mutational interrogation experiments at putative Ubx regulatory regions.
| Species | sgRNA(s) | Inj. EmbryosNinj | L1 larvaeNhat | Pupae orL5 larvae | AdultsNadu | CrispantsNmut | Inj. timeh AEL | Cas9:sgRNA ng/µL | Hatching RateNhat/Ninj | Crispant RateNmut/Ninj |
|---|---|---|---|---|---|---|---|---|---|---|
| J. coenia | Antp-Ubx_BE | 59 | 50 | 50 | 44 | 6 | 2.5–3.5 | 500 : 250 | 84.7% | 10.2% |
| 118 | 40 | 40 | 31 | 6 | 1.75–2.75 | 250 : 125 | 33.9% | 5.1% | ||
| 89 | 44 | 44 | 39 * | 17 | 2.25–3.5 | 500 : 250 | 49.4% | 19.1% | ||
| Total | 266 | 90 | 134 | 115 | 29 | 33.8% | 10.9% | |||
| V. cardui | IT1_sgRNA1 | 204 | 67 | 50 | 50 | 2 | 1–3 | 250 : 125 | 32.8% | 1.0% |
| 108 | 49 | 31 | 31 | 3 | 2–3 | 125 : 62.5 | 45.4% | 2.8% | ||
| 145 | 60 | 39 | 39 | 2 | 2.25–3.5 | 500 : 250 | 41.4% | 1.4% | ||
| Total | 457 | 176 | 120 | 120 | 7 | 38.5% | 1.5% | |||
| J. coenia | IT1_sgRNA2 | 59 | 40 | 7 | 6 | 0 | 0.5–2.5 | 500 : 250 | 67.8% | 0.0% |
| 124 | 112 | 112 | 110 | 4 | 2.25–4.75 | 500 : 250 | 90.3% | 3.2% | ||
| Total | 183 | 152 | 119 | 116 | 4 | 83.1% | 2.2% | |||
| V. cardui | AS5_sgRNA1 | 334 | 183 | 57 | 52 | 5 | 2–3 | 250 : 125 | 54.8% | 1.5% |
| 122 | 87 | 2 | 2 | 0 | 2–4 | 500 : 250 | 71.3% | 0.0% | ||
| Total | 456 | 270 | 59 | 54 | 5 | 59.2% | 1.1% | |||
| J. coenia | AS5_sgRNA1 | 309 | 181 | 181 | 181 | 8 | 2–4.5 | 500 : 250 | 58.6% | 2.6% |
| J. coenia | Ubx11a2+3 +c5+6 | 317 | 18 | - | - | 2 | 1–3 | 500 : 75 ea. | 5.7% | 0.6% |
| 203 | 35 | 0 | 0 | 0 | 1.5–3.5 | 500 : 75 ea. | 17.2% | 0.0% | ||
| Total | 520 | 53 | - | - | 2 | 10.2% | 0.4% | |||
| V. cardui | Ubx11a2+c5 | 50 | 5 | 3 | 3 | 2 | 4–4.5 | 500 : 500 | 10.0% | 4.0% |
| 151 | 29 | 6 | 5 | 2 | 2–2.75 | 500:125:125 | 19.2% | 1.3% | ||
| 361 | 18 | 13 | 16 | 6 | 0.5–2 | 500:125:125 | 5.0% | 1.7% | ||
| Total | 562 | 52 | 22 | 24 | 10 | 9.3% | 1.8% | |||
| V. cardui | Ubx11c5 | 168 | 99 | 27 | 26 | 3 | 3.75–4.75 | 250 : 125 | 58.9% | 1.8% |
| 62 | 22 | 9 | 9 | 2 | 0.5–0.75 | 500 : 250 | 35.5% | 3.2% | ||
| 131 | 93 | 8 | 8 | 3 | 1.5–3 | 500 : 250 | 71.0% | 2.3% | ||
| 114 | 63 | 20 | 20 | 6 | 3.5–4.5 | 500 : 250 | 55.3% | 5.3% | ||
| Total | 475 | 277 | 64 | 63 | 14 | 58.3% | 2.9% | |||
| V. cardui | Ubx11b9 | 32 | 18 | 6 | 5 | 1 | 1.25–2.25 | 500 : 250 | 56.3% | 3.1% |
| 63 | 49 | 9 | 6 | 1 | 3.5–4.5 | 500 : 250 | 77.8% | 1.6% | ||
| Total | 95 | 67 | 15 | 11 | 2 | 70.5% | 2.1% | |||
| J. coenia | Ubx11b9 | 41 | 13 | 13 | 13 | 3 | 2.5–4 | 125 : 62.5 | 31.7% | 7.3% |
| 48 | 21 | 14 | 14 | 1 | 2–3 | 250 : 125 | 43.8% | 2.1% | ||
| Total | 89 | 34 | 27 | 27 | 4 | 38.2% | 4.5% | |||
| *: upper estimate, includes 16 fifth instars larvae that were dissected for immunostainings, of which 7 were mutants (as evidenced by ectopic UbdA in forewings), and 3 dissected mutant pupae | ||||||||||
Additional files
-
Supplementary file 1
List of sgRNAs used in CRISPR experiments.
- https://cdn.elifesciences.org/articles/90846/elife-90846-supp1-v1.xlsx
-
MDAR checklist
- https://cdn.elifesciences.org/articles/90846/elife-90846-mdarchecklist1-v1.pdf
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Cis-regulatory modes of Ultrabithorax inactivation in butterfly forewings
eLife 12:RP90846.
https://doi.org/10.7554/eLife.90846.3




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}