Peer review process
Revised: This Reviewed Preprint has been revised by the authors in response to the previous round of peer review; the eLife assessment and the public reviews have been updated where necessary by the editors and peer reviewers.
Read more about eLife’s peer review process.Editors
- Reviewing EditorWeibin ZhouIcahn School of Medicine at Mount Sinai, New York, United States of America
- Senior EditorDidier StainierMax Planck Institute for Heart and Lung Research, Bad Nauheim, Germany
Reviewer #1 (Public Review):
Induction of beta cell regeneration is a promising approach for the treatment of diabetes. In this study, Massoz et.al., identified calcineurin (CaN) as a new potential modulator of beta cell regeneration by using zebrafish as model. They also showed that calcineurin (CaN) works together with Notch signaling to promote the beta cell regeneration. Overall, the paper is well organized, and technically sound. However, some evidences seem weak to get the conclusion.
Reviewer #2 (Public Review):
This work started with transcriptomic profiling of ductal cells to identify the upregulation of calcineurin in the zebrafish after beta-cell ablation. By suppressing calcineurin with its chemical inhibitor cyclosporin A and expressing a constitutively active form of calcineurin ubiquitously or specifically in ductal cells, the authors found that inhibited calcineurin activity promoted beta-cell regeneration transiently while ectopic calcineurin activity hindered beta-cell regeneration in the pancreatic tail. They also showed similar effects in the basal state but only when it was within a particular permissive window of Notch activity. To further investigate the roles of calcineurin in the ductal cells, the authors demonstrated that calcineurin inhibition additionally induced the proliferation of the ductal cells in the regenerative context or under a limited level of Notch activity. Interestingly, the enhanced proliferation was followed by a depletion of ductal cells, suggesting that calcineurin inhibition would exhaust the ductal cells. Based on the data, the authors proposed a very attractive and intriguing model of the role of calcineurin in maintaining the balance of the progenitor proliferation and the endocrine differentiation. However, the conclusions of this paper are only partially supported by the data as some evidence of the lineage between ductal cells and beta cells remains suggestive.
Author Response
The following is the authors’ response to the original reviews.
eLife assessment
This work presents some valuable information regarding the molecular mechanisms controlling the regeneration of pancreatic beta cells following induced cell ablation. However, the study lacks the critical lineage tracing result to support the conclusion about the origin of the regenerated beta cells. The results of the pharmacological manipulation of CaN signaling are also incomplete. In particular, these manipulation are not cell-specific, making it difficult to interpret and thus genetic approach is recommended.
Public Reviews:
Reviewer #1 (Public Review):
Induction of beta cell regeneration is a promising approach for the treatment of diabetes. In this study, Massoz et.al., identified calcineurin (CaN) as a new potential modulator of beta cell regeneration by using zebrafish as model. They also showed that calcineurin (CaN) works together with Notch signaling calcineurin (CaN) to promote the beta cell regeneration. Overall, the paper is well organized, and technically sound. However, some evidence seems weak to get the conclusion.
Reviewer #2 (Public Review):
This work started with transcriptomic profiling of ductal cells to identify the upregulation of calcineurin in the zebrafish after beta-cell ablation. By suppressing calcineurin with its chemical inhibitor cyclosporin A and expressing a constitutively active form of calcineurin ubiquitously or specifically in ductal cells, the authors found that inhibited calcineurin activity promoted beta-cell regeneration transiently while ectopic calcineurin activity hindered beta-cell regeneration in the pancreatic tail. They also showed similar effects in the basal state but only when it was within a particular permissive window of Notch activity. To further investigate the roles of calcineurin in the ductal cells, the authors demonstrated that calcineurin inhibition additionally induced the proliferation of the ductal cells in the regenerative context or under a limited level of Notch activity. Interestingly, the enhanced proliferation was followed by a depletion of ductal cells, suggesting that calcineurin inhibition would exhaust the ductal cells. Based on the data, the authors proposed a very attractive and intriguing model of the role of calcineurin in maintaining the balance of the progenitor proliferation and the endocrine differentiation. However, the conclusions of this paper are only partially supported by the data as some evidence from the data remains suggestive.
(1) In the transcriptomic profiling, genes differentially regulated in the ablated adults could be solely due to the chemical effects of metronidazole instead of the beta-cell ablation. A control group without ins:NTR-mCherry but treated with metronidazole is necessary to exclude the side effects of metronidazole.
We believe that it is unlikely that the differential regulation observed is due to metronidazole rather than the beta cell loss. This experimental strategy as proven successful in well-published studies to identify regulators of beta cell regeneration in the zebrafish larvae. Importantly, the candidates identified in these studies were subsequently functionally validated in mammalian models (Lu et al. 2016, Karampelias 2021). Moreover, in our study, we also used another chemical compound, the nifurpirinol (Bergemann et al., 2018), to ablate the beta cells. Regardless of whether we employed metronidazole or nifurpirinol for beta cell ablation, our results consistently indicate a notable involvement of calcineurin. Of note, the nifurpirinol molecule is commonly used in fishkeeping without toxicity reported on the global health of the fish.
(2) Although it has been shown that the pancreatic duct is a major source of the secondary islets in the pancreatic tail in previous studies, there is no direct evidence showing the cyclosporin A-induced cells share the source in this manuscript. Without any proper lineage tracing work, the origin of those cyclosporin A-induced cells cannot be concluded.
Our experimental setting is similar to the one described in Ninov et al. 2013, where lineage tracing experiments demonstrate an increase of beta cell formation in the pancreatic tail that originate from the pancreatic ducts. In our study, we performed the same experiment with the addition of CsA and showed more ductal cell proliferation (Figure 5G) followed by a 19% increase of beta cell regeneration compared to nonregenerative conditions (Figure 2B). It is unlikely that the additional 19% of regenerated beta cells under CaN inhibition come from another source than the 68% first.
On the other hand, the acinar cells cannot be consider as another source of regenerated beta cell as they are not able to form beta cells unless they are artificially reprogrammed (Maddison et al., 2012). Therefore the only other potential source of regenerated beta cell is the endocrine compartment. However at the stage where we performed beta cell ablation, there are no endocrine cell in the pancreatic tail. Moreover, there are no evidence that secondary islets could come from the principal islet, they are tightly associated with the ducts and differentiate form ductal cell (Mi et al., 2023).
Importantly, we demonstrated that overexpression of CaN specifically in the pancreatic ducts prevents beta cell regeneration. CaN effect is therefore intrinsic to the ducts. Moreover, we showed that CsA increase beta cells formation when Notch signalling is repressed. Given that Notch signalling is known to act on the ductal cell population, this strongly suggests again that CsA exacerbate beta cells formation from the ducts.
All of these compelling evidences strongly support the notion that the cyclosporininduced beta cells originate from the ductal cells.
(3) It is interesting to see an increase of beta cells in the primary islet after cyclosporin A treatment (Supplemental Fig 2B). However, it remains unclear if their formation shares the same mechanism with the newly formed beta cells in the pancreatic tail.
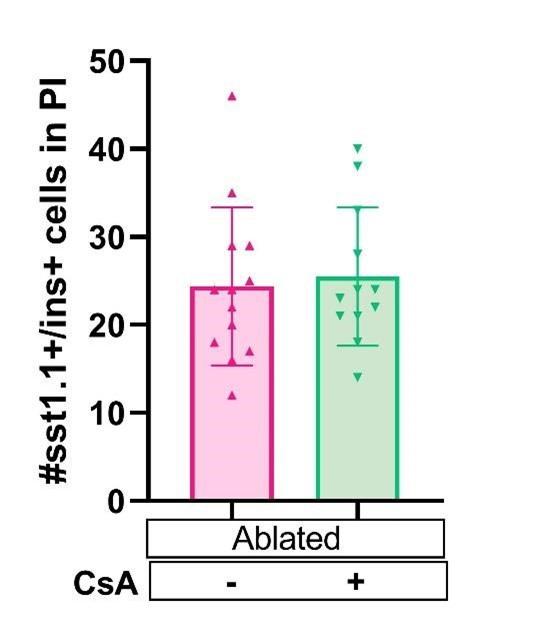
There are indeed several source of beta cell regeneration in the primary islet. However, a recent study showed that the contribution of alpha cell to regeneration is minor and the main contributors are ductal and sst1.1 cells (Mi et al., 2023). In our previous publication, we indeed showed that a major source of beta cell in the principal islet is the delta 1.1 cell population. Those sst1.1 cells begin to express insulin and therefore are named ‘bihormonal’ (Carril et al., 2022). We tested if this population is impacted by CsA treatment and we showed below that CsA does not affect bi-hormonal cell formation (Figure 2D supplemental). These new results suggest that the CsA mediated increase of beta cells in the principal islet arise from the ductal cells as observed in the tail. These results were added in the manuscript as Figure 2D supplemental.
Author response image 1.
Tg (sst1.1:GFP); Tg (ins:NTR*-mCherry) larvae were treated at 3dpf with NFP 4µM to induce beta cell ablation. Then larvae were treated with CsA 1µM from 4 to 6 dpf (or ctl with DMSO); prior fixation and analysis of bi-hormonal cells in the principal islet at 6dpf.
(4) The conclusion of the effect of cyclosporin A on the endocrine progenitors (Line 175) is not convincing because the data cannot distinguish the endocrine progenitors from the insulin-expressing cells. Indeed, Figure 2E shows that neurod1+ cells are fewer than ins+ cells (Figure 2D) in the pancreatic tail at 10 dpt, suggesting that all or at least the majority of neurod1+ cells are already ins+.
The neurod1+ cells population indeed included both endocrine progenitor cells and differentiated endocrine cells. However, we would like to point out that the timing of the analysis is essential to reach our conclusion. When we treat with CsA, we show an increase of neurod1+ cells already at 4dpt. At this time point, no hormone- producing cell can yet be detected (Figure 2E). Those additional neurod1+ cell are therefore endocrine progenitors and not beta cells. This result shows that CaN inhibition induces pro-endocrine cell formation in regenerative conditions.
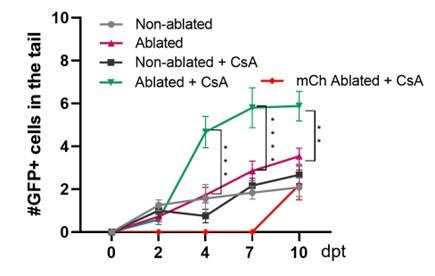
At 10dpt, the neurod1+ cells population includes beta cells as well as endocrine progenitor cell. We agree that the way the data are presented in figure 2D and 2E can be confusing. Those 2 figures come form 2 separated experiments, the number of beta cell in figure 2D can therefore not be compared to the number of Neurod1+ cell in figure 2E. Indeed, from one experiment to another the efficiency and rate of regeneration can vary, independently of calcineurin. To clarify, we added the number of beta cells regenerated in the experiment of figure 2E (see Author response image 2 in red). As you can see in this experiment, regeneration was a bit slower than usual.
Author response image 2.
Tg (neurod1:GFP); Tg (ins:NTR*-mCherry) larvae were treated at 3dpf with NFP 4µM to induce beta cell ablation. Then larvae were treated with CsA 1µM from 4 to 6 dpf (or ctl with DMSO); prior fixation and analysis of GFP+ cells (in grey, pink, dark grey and green), and mCherry+ cells for the condition ablated + CsA in red from 2 to 10 dpf.
(5) Figure 5D shows a significant loss of nkx6.1+ cells in the combined treatment group but there is no direct evidence showing this was a result of differentiation as the authors suggested. This cell loss also outnumbered the increase in ins+ cells (Figure 4D). The cell fates of these lost cells are still undetermined, and the authors did not demonstrate if apoptosis could be a reason of the cell loss.
Firstly, as you can notice on the graphs, we encountered a very high variability between individuals within the same condition. We decided to show this variability by presenting the raw data. This high variability could partially explain the differences that you underline. Moreover, we would like to point out that independently of CaN inhibition the progenitor loss (nkx6.1+ cell) outnumber the gain of beta cells. Indeed, in average there is a loss of 29% (41 GFP+) of the nkx6.1+ cells and a gain of only 6 beta cells after Notch inhibitory treatment. The other progenitors cells being differentiated into other endocrine cell types (pro-endocrine, alpha, delta). In the combined treatment (Notch and CaN inhibitors), we decreased the number of progenitors cell by 50%, i.e 21% (20 cells) more than without CaN inhibitor. However, we increased the number of regenerated beta cells by two fold (6 cell to 12 cells). In brief, the important progenitors cell loss could be explained by precocious differentiation in the pro-endocrine and endocrine cells type. It is therefore normal than the number of beta cells regenerated do not match the progenitors cell number loss and this in presence or absence of CaN inhibition.
Recommendations for the authors:
Reviewer #1 (Recommendations For The Authors):
Major concerns:
(1) The evidence to indicate the proliferating ductal cell differentiate into beta cell is weak. They should use linkage tracing, or other marker genes immunostaining to confirm that.
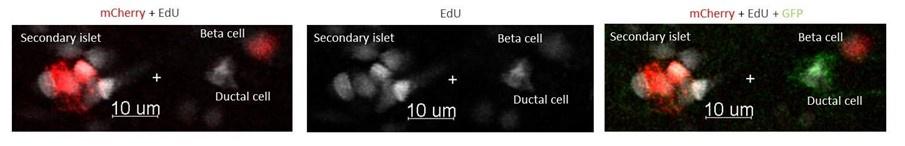
The experiment from the Figure 5 A-D is a short term tracing experiment and should have been presented as such in the manuscript. After LY411575 (Notch inhibitor) and CsA treatments at 3dpf, we exposed the larvae to EdU at 4dpf during 8 hours (Figure 5A). We showed that EdU is incorporated in dividing ductal cells at 4dpf (Figure 5C) ant that 2 days later there are newly form beta cells that are EdU+.(see Author response image 3) To reinforce our conclusion, the image below will be added to the manuscript.
Author response image 3.
Tg (nkx6.1:GFP); Tg (ins:NTR*-mCherry) larvae were treated at 3dpf with both CsA 1µM and LY411575 5µM. At 4dpf, the larvae were exposed to EdU 4mM during 8 hours, before analysis at 6 dpf.
(2) To inhibition of CaN and Notch pathway, they just used the pharmacological approaches, genetical approaches should be used to get stronger evidence.
We employed two distinct inhibitors specifically targeting calcineurin (CsA and FK506) for CaN inhibition. While these inhibitors have distinct chemical structures and potential non-specific effects, they both yield the same result of increased beta cell formation under Notch repression (see Figure 4D and Figure 4B in the supplementary data). This convergence of outcomes strongly suggests that the observed effect is primarily attributable to the specific inhibition of calcineurin.
Furthermore, we complemented our inhibitor-based approach with a genetic strategy involving CaN overexpression (see Figure 3). Notably, the overactivation of CaN resulted in a reduction of beta cell regeneration. Given that this genetic approach generated an effect contrary to that achieved with the inhibitors, it provides robust support for our model, which postulates that calcineurin plays a critical role in the regulation of beta cell regeneration (see Figure 3, panels C-E).
As for Notch inhibition, previous published data from our laboratory compared the effects of Notch inhibitor (LY411575) and genetic approaches (mib mutant and transgenic line) on pro-endocrine cell (ascl1b+) and ductal cell (nkx6.1+) formation. This study showed that both Notch inhibitor (LY411575) and Notch repression using genetic approaches recapitulate the same effect: an induction of pro-endocrine cells formation. The specificity of this inhibitor being validated (Ghaye et al., 2015), we did not consider the need of a genetic approach.
(3) The most enriched pathways among the up-regulated genes were DNA replication and cell cycle, which suggested that these genes are more important for the duct cell proliferation, how is Calcineurin related to these pathways, such as regulating the genes important for proliferation?
The transcriptomic data presented in this manuscript suggest that the ductal cells undergo a strong proliferative response after beta cell ablation. This is in accordance with our experimental data showing activation of ductal proliferation after beta cell ablation (Ghaye at al., 2015) and data from this manuscript (Figure 1 I-J).
Calcineurin is a well-known regulator of the cell cycle, and can either promote or repress the cell cycle depending on the cell type. For example, stressing the cell provokes an entry of calcium and subsequently a CaN activation which result in cell cycle arrest (Leech et al. 2020). Nevertheless, depending the cell type, CaN can be either necessary or deleterious to cell proliferation (Goshima et al. 2019; Masaki and Shimada 2022). The intriguing dual role of CaN in cell cycle is well illustrated in β cell regeneration. While CaN should be repressed to enable ductal progenitor amplification and subsequent endocrine differentiation, CaN is then necessary for β cell function and for their replication (Dai et al. 2017; Heit et al. 2006). Moreover, CaN is related to cellular senescence and CaN function is important for proper fin regeneration in zebrafish.
(4) It is hard to understand why they pick up the pathway of cellular senescence signature for the duct cell progenitor neogenesis? Moreover, among these senescence genes, many genes are cell cycle regulators.
In response to beta cell ablation, the ductal cells undergo a strong proliferative response, as shown in our previous data (Ghaye 2015). It was therefore not surprising that many differentially expressed genes are cell cycle regulators. On the other hand, the cellular senescence signature was surprising. Indeed, senescence is usually associated with cell cycle arrest and aging. However, recent studies showed that cellular senescence is required for proper development and regeneration. We therefore wanted to investigate this pathway and more particularly the function of calcineurin, which can either promote or repress the cell cycle in different cell types (see comment above).
(5) The RNA-seq data obtained from adult fish, while the authors use larvae to explore the CaN functions, it may have different conclusion using adult fish. Moreover, it is unclear whether the CaN increased when the beta cell ablated in young larvae.
We decided to first perform functional experiment in the larvae as this model unable the quantification of beta cell regeneration from the ducts in the pancreatic tail. However, to validate our results in non-developmental stages, we perform experiments in juveniles (2 months old) and adults. CsA treatments in juveniles zebrafish recapitulated the same results that in larvae (Figure 2B and Figure 6A-C). Moreover, we showed that CaN overactivation delayed glycemia recovery after ablation adults (Figure 6D-E), which is in accordance with an impaired regeneration. Altogether, these results strongly suggest that CaN act as regulator of beta cell regeneration both in the juvenile/adult and larval stages.
Concerning the expression of CaN in the zebrafish larvae, we tried to detect the level of CaN in the different experimental conditions by in situ hybridization. However, we were not able to detect it using this technique. We also tried immunostaining with antiphospho-nfact3 ser165 polyclonal antibody (Invitrogen) but this antibody does not seem to work in zebrafish. Finally, we tried to sort ductal cell at larval stage to perform a transcriptomic analysis but we were unable to collect enough ductal cells to proceed further. Indeed our staining experiment showed that there are only around 150 ductal cells (nkx6.1+, Figure 5D) at this stage.
(6) The beta cell regeneration in the young larvae usually recovers within ~ 5 days in principle islet. Please also show the beta cell number (PI) during the beta cell recovery after ablation.
We did show beta cell regeneration in the principal islet in Figure 2A-B supplemental. While new beta cells appears quickly in this islet (Carril, Massoz, Dupont et al., 2023), the principal islet has not yet fully recover at 5dpt.
(7) Since the studies did not show the CaN level in Fig.3, it is hard to know that the CaN is exactly expressed.
In the figure 3B, using Tg(hsp70:GFP-CaNCA), it is indeed not possible to see CaN expression at 10 dpt as the heat shocks induce only transiently CaNCA overexpression. However, the transient expression was detected in live shortly after the heat shocks. On the other hand, with the transgenic line Tg(UAS:GFP-CaNCA); Tg(cftr:Gal4), in which GFPCaNCA is continuously expressed allowing us to show CaNCA expression in the pancreatic ducts (Figure 3).
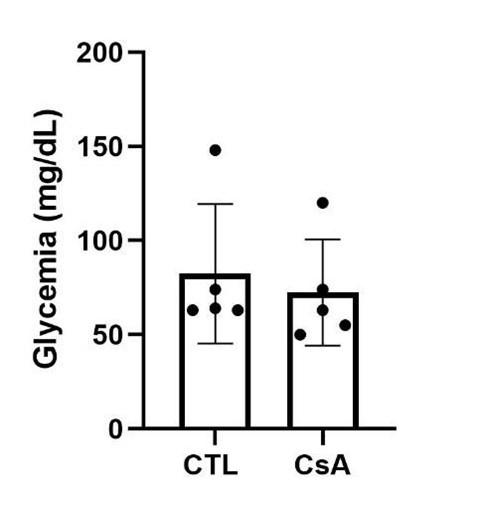
(8) In Fig.6 D and 6E, did these drug treatments change the glucose level in nonablated fish?
As you can see below, the CaN inhibitor, CsA does not affect the glycemia of the fish in non-regenerative conditions.
Author response image 4.
Glycemia of non-ablated fish, 3 days after drug treatment.
(9) The logic of writing in Results is very hard to understand.
We proofed read the paper in an effort to clarify it.
Minor concerns,
(1) Make a scheme for ablation and RNA-seq, and indicate the age of the fish used in Fig. 1.
We added the scheme in Figure 1 supplemental.
(2) In Fig. 1G, two arrows indicated mCherry+ cells is hard to see in the non-ablated fish.
One arrow was indeed mislocated, we moved the arrow and try to improve the intensity of red. However, the only cells are indeed small and can be difficult to see.
(3) In Fig.6, it is hard to know that the arrows indicated islets are small islets (up to 5 cells), how they compared with big islets and defined as small islet. Moreover, some of these islets are almost invisible.
We now show a close up of a portion of the pancreatic tail and show the beta cells with arrows only in this picture, to enhance clarity.
Reviewer #2 (Recommendations For The Authors):
(1) This manuscript needs more proofreading and polishing to increase its readability.
We proofread the manuscript and change some paragraph for more clarity.
(2) The extensive use of words like "modulate" or "regulate" sometimes makes the text ambiguous as the effect is not stated directly and clearly.
We re-wrote some parts of the text and try to avoid using “regulate” as often.
However, as we used both repression and over-activation of CaN, we still use words as regulate to stipulate general conclusions on the function of CaN.
(3) The list of individual differentially regulated genes after the beta-cell ablation in the RNAseq seems missing. This list could be interesting and helpful for other researchers. We added it.
(4) In Figure 1D, "modulated" genes are shown but were they all upregulated like those in Figure 1A? The modulation should be indicated more clearly (e.g. up- or down-regulated) in the figure. The authors can use different colours to illustrate that.
Done.
(5) Is Figure 2D showing the same data extracted from Figure 2B? Does Figure 2D add any information to the data?
No, it does not add data. We actually add the Figure 2D for a better visualisation of the increase at 10dpt.
(6) In the y-axis of Figure 3E, it should be "mCherry".
It already is. We did check all the axis again to be sure it is correct.
(7) Line 219, "Figure 4E supplemental" instead of "Figure 4D supplemental"
Done.
(8) Line 266, "ablated juveniles" instead of "ablated larvae"
Done. Thank you for noticing these mistakes.
(9) In Figure 6A, many mCherry+ cells are hardly visible and there are some greyish white signals in the images that are supposed to show the mCherry channel only. What are those grey signals?
There is no channel showing grey on the picture, I improved the overall quality of this pictures and show close up to improve the figure.
(10) In Figure 6D and 6E, CaNCA overexpression had a significant effect on the glycemia. But did the overexpression affect the beta cell formation or regeneration? We showed that CaNCA overexpression did not affect beta cell formation in absence of regeneration in the larvae (Figure 3E). Moreover, it does not affect the glycemia of the fish in non-regenerative conditions (Author response image 5). As for regenerative conditions, CaN overexpression decreased the regeneration in the larvae (Figure 3E).
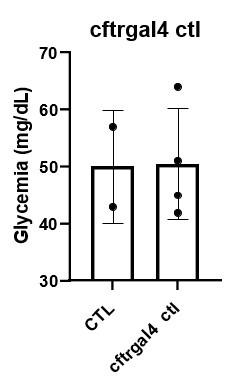
Author response image 5.
Glycemia of Tg(UAS:GFP-CaNCA); Tg(cftr:Gal4) fish, overexpressing CaNCA, compared to controls fish, in non-regenerative conditions.
(11) The role of calcineurin seems transient (e.g. Figure 2B and 4E) and does not play a significant role in long term. It would be interesting to see if long-term/repeated treatments of calcineurin inhibitors and overexpression/knockout of important members of calcineurin signaling would affect the pool of progenitors in long term.
We were also interested in the consequences of CaN overexpression on the long term. Our overexpression tool Tg(UAS:CaNCA) allow to address this question, as CaN is overexpress permanently. We assessed the structure of the ducts and the number of beta cells in transgenic larvae and did not see any defects of the ducts whether in regenerative context or not. On the other hand, we showed in this manuscript that CaN effect is specific to regenerative conditions. As a consequence, it is not likely that repeated treatments long after the ablation would continue to affect beta cell formation and the progenitors pool.



