The DEG/ENaC cation channel protein UNC-8 drives activity-dependent synapse removal in remodeling GABAergic neurons
- Vanderbilt University, United States
- University of Illinois at Chicago, United States
- University of Miami, United States
- Tokyo Women's Medical University, Japan
Figures
Figure 1 with 1 supplement
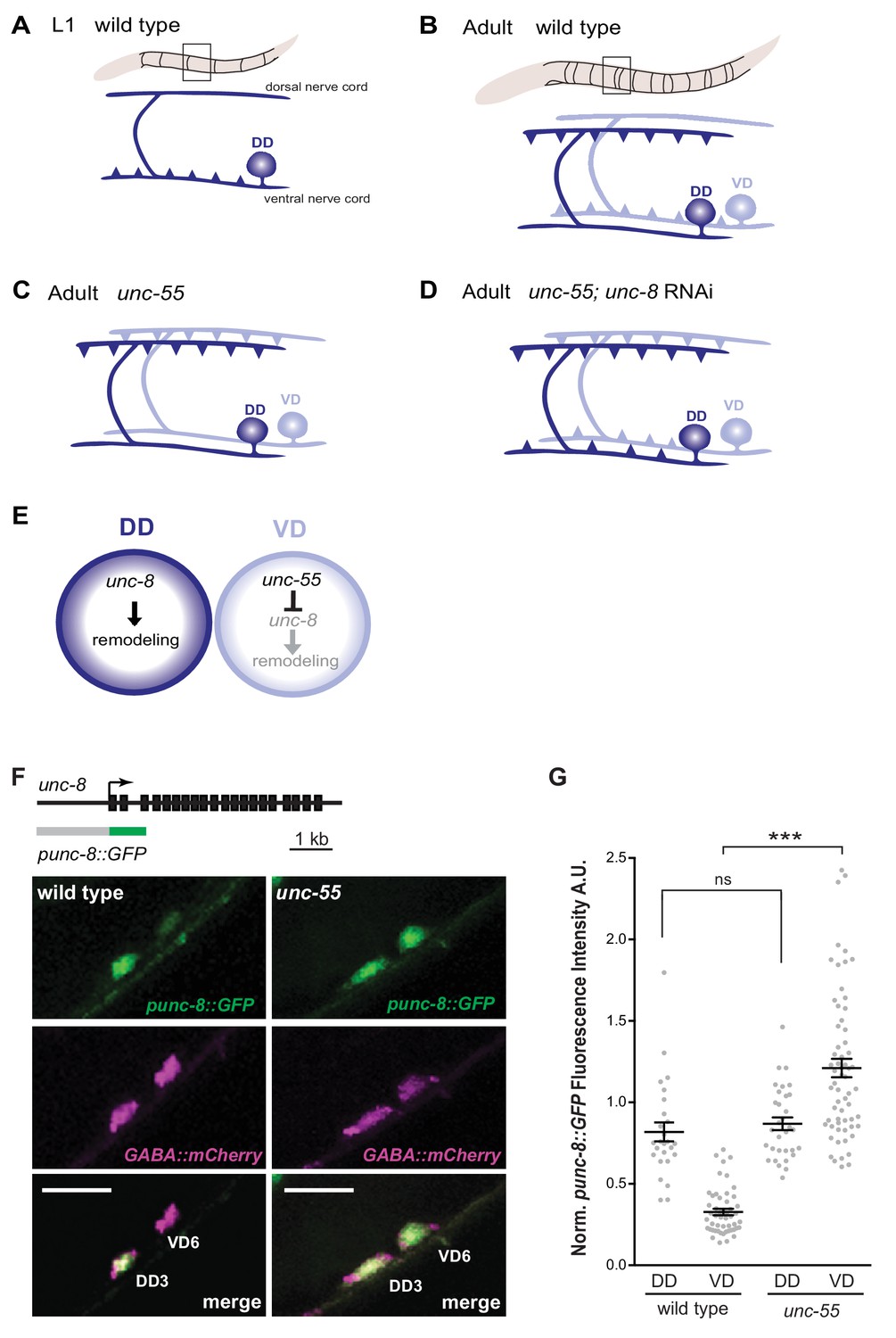
GABAergic neuron synaptic remodeling is transcriptionally controlled and depends on UNC-8.
(A) Dorsal D (DD) GABAergic motor neurons (dark blue) synapse with ventral muscles during embryonic development. (B) DD synapses are relocated to dorsal muscles at the end of the first larval stage (L1). Ventral D (VD) GABAergic motor neurons (light blue) are generated in the late L1 and innervate ventral muscles. (C) The COUP/TF transcription factor UNC-55 is expressed in VD neurons and blocks remodeling; VD neurons relocate synapses to the dorsal side in loss-of-function unc-55 mutants. (D/E) UNC-8/DEG/ENaC expression is negatively regulated by UNC-55 and RNAi knockdown of unc-8 suppresses ectopic VD remodeling in unc-55 mutants. These results suggest that UNC-8 may also promote DD remodeling. Schematics modified from Petersen et al. (2011). (F) unc-8 expression in remodeling neurons is visualized with a punc-8::GFP reporter gene. Strong punc-8::GFP (green) expression was observed in DD motor neurons in wild type, but was also detected in VD motor neurons in unc-55 animals. GABAergic motor neurons are labeled with pttr-39::mCherry (magenta). Scale bar is 10 μm. (G) Normalized fluorescence intensity is plotted on the Y-axis in arbitrary units (A.U.). punc-8::GFP expression is enhanced in VDs, but not DDs, in unc-55 mutants (***p<0.001, ns is not significant, One-Way ANOVA with Bonferroni correction). n ≥ 26 DDs and n ≥ 51 VDs per genotype, data are mean ± SEM.
Figure 1—figure supplement 1

UNC-8 is expressed in remodeling GABA neurons.
(A) A GFP C-terminal fusion with the intact UNC-8 protein was generated by recombineering (see Materials and methods). (B) UNC-8::GFP shows mosaic expression in the ventral nerve cord (VNC) of early L2 animals in GABAergic DD neurons (asterisks) and in cholinergic DA and DB neurons (upper left). Schematic denotes VNC neurons that express UNC-8::GFP (upper right). The six DD neurons are labeled with punc-25::mCherry. mCherry-labeled cell bodies between the DD neurons are the newly born VD neurons (lower left). Merged image of UNC-8::GFP fosmid and punc-25::mCherry-labeled GABA neurons (lower right). Asterisks denote an expression of UNC-8::GFP in DD3 and DD4. Scale bar is 20 μm. (C) UNC-8::GFP is expressed in DD and VD neurons in unc-55 mutants. GABA neurons are labeled with wdIs90 [punc-25::mCherry::RAB-3]. UNC-8::GFP-expressing GABA neurons are outlined. Scale bar is 10 μm. UNC-8::GFP rescues the unc-55 remodeling phenotype in unc-55; unc-8 animals (***p<0.001, n ≥ 20 animals, One-Way ANOVA with Bonferroni correction, data are mean ± SD).
Figure 2 with 1 supplement
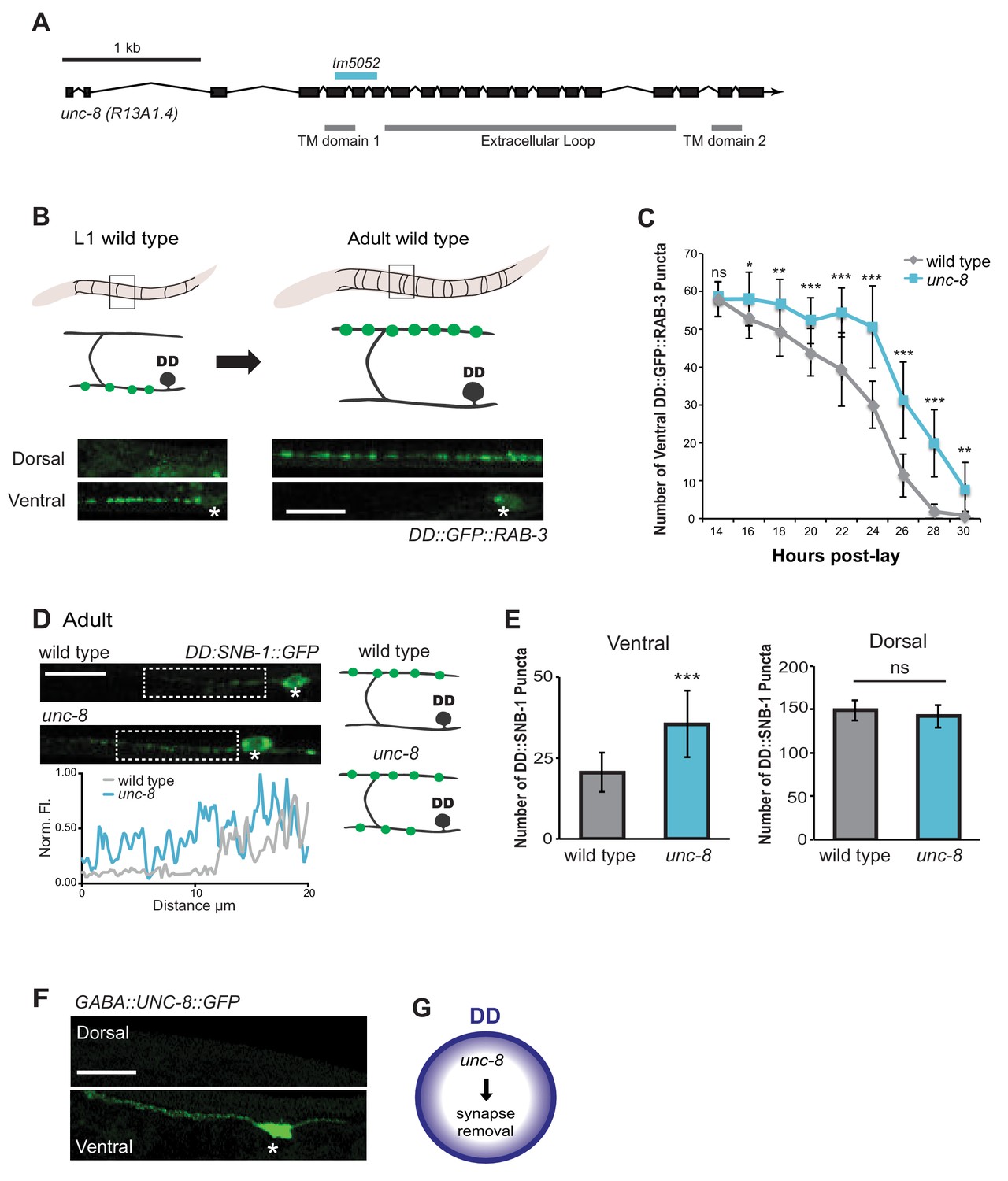
The DEG/ENaC subunit UNC-8 promotes removal of ventral DD synapses.
(A) Schematic of the unc-8 gene and predicted UNC-8 protein. DEG/ENaC channel subunits contain two transmembrane domains (TM domains) and a large extracellular loop (gray bars). The unc-8 deletion allele tm5052 is indicated (blue bar). (B) DD GABA neuron synapses (green) with ventral muscles are relocated to the dorsal side during development. DD-specific GFP-tagged RAB-3 (pflp-13::GFP::RAB-3) labels synapses in the ventral nerve cord of early L1 larvae and the dorsal nerve cord of adults. Asterisk denotes DD4 soma. Scale bar is 10 μm. (C) DD remodeling was quantified by counting GFP::RAB-3 puncta during larval stages. Removal of ventral DD synapses is significantly delayed in unc-8 animals (*p<0.05, **p<0.01, ***p<0.001, ns is not significant, Student’s t-test, data are mean ± SD). (D) Representative images of wild-type and unc-8 adult ventral nerve cords (asterisk denotes DD5 soma). Ventral DD synapses labeled with GFP-tagged synaptobrevin (pflp-13::SNB-1::GFP) are retained in unc-8 mutant adult animals. Scale bar is 10 μm. Inset shows pixel intensity over a 20 μm region (indicated by dashed boxes) of the ventral nerve cord in wild-type and unc-8 animals. (E) Removal of DD::SNB-1::GFP puncta is defective in unc-8 mutant adults; however, dorsal DD synaptic assembly is not affected (***p<0.001, n ≥ 20, ns is not significant, Student’s t-test, data are mean ± SD). (F) Ventral localization of GFP-tagged UNC-8 in GABA neurons. Asterisk denotes DD5 soma. Scale bar is 10 μm. (G) UNC-8 promotes removal of ventral presynaptic components in DD neurons, but is not required for dorsal synapse formation.
Figure 2—figure supplement 1
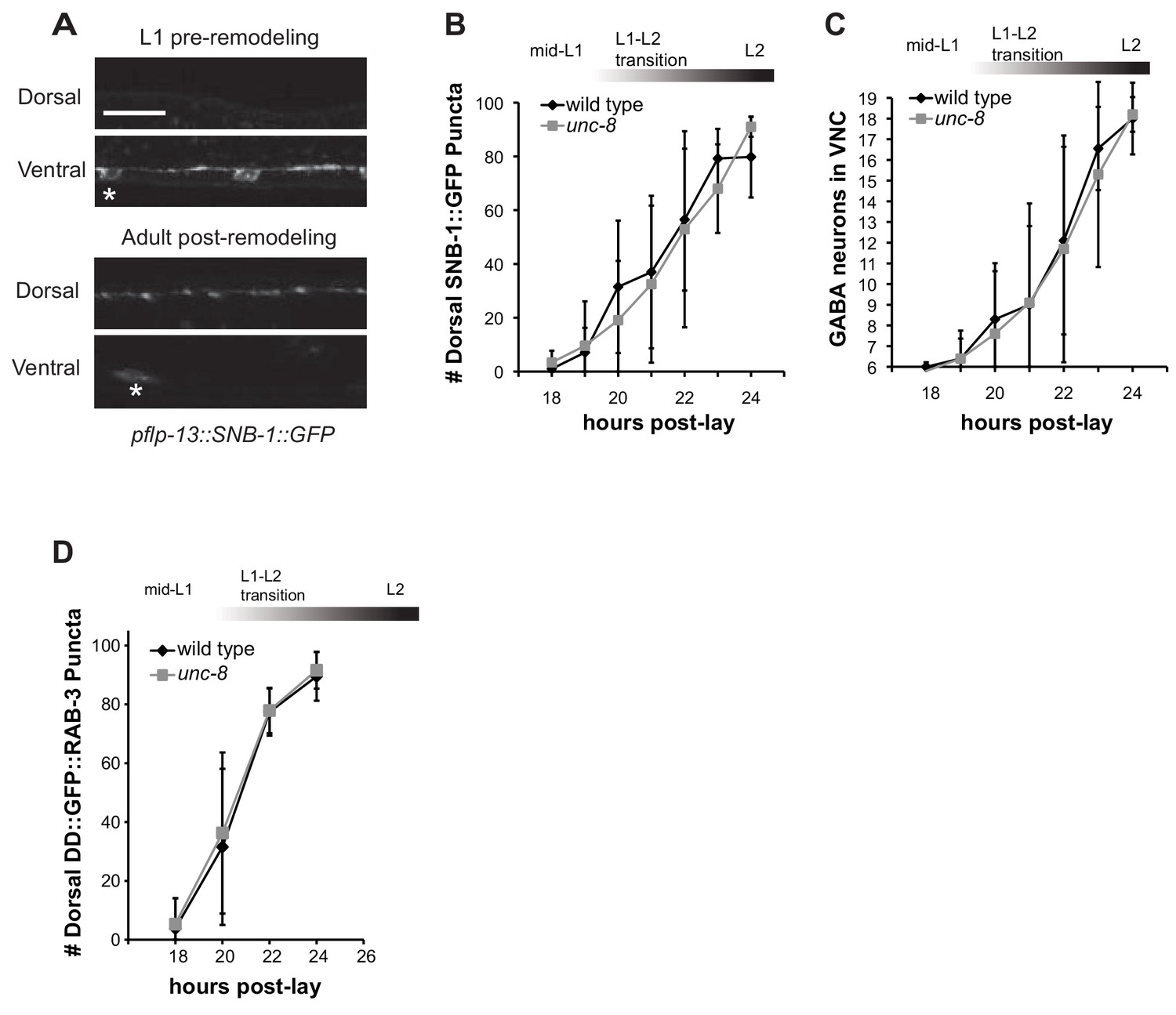
UNC-8 removes ventral synapses, but is not required for assembly of dorsal synapses.
(A) DD-specific expression of SNB-1::GFP (pflp-13::SNB-1::GFP) labels DD synapses. In the wild type, all SNB-1::GFP puncta are ventrally localized in early L1 larvae, but are strictly dorsal in adults after DD remodeling. Scale bar is 10 μm. Asterisk denotes DD4 cell soma. (B) Loss of unc-8 function does not delay dorsal DD synapse formation. Dorsal SNB-1::GFP puncta were counted in developing wild-type and unc-8 mutant L1/L2 larvae. (C) GABA neuron development is not delayed in unc-8. GABA neurons (DD + VD) in the ventral cord, marked with SNB-1::GFP, were counted in developing wild-type and unc-8 mutant L1/L2 larvae. (D) DD remodeling was quantified by counting GFP::RAB-3 puncta during the L1-L2 larval stages. Assembly of dorsal DD synapses is not significantly different between wild-type and unc-8 animals (Student’s t-test, data are mean ± SD). Results for (B–D) were pooled from 3 independent experiments.
Figure 3 with 2 supplements
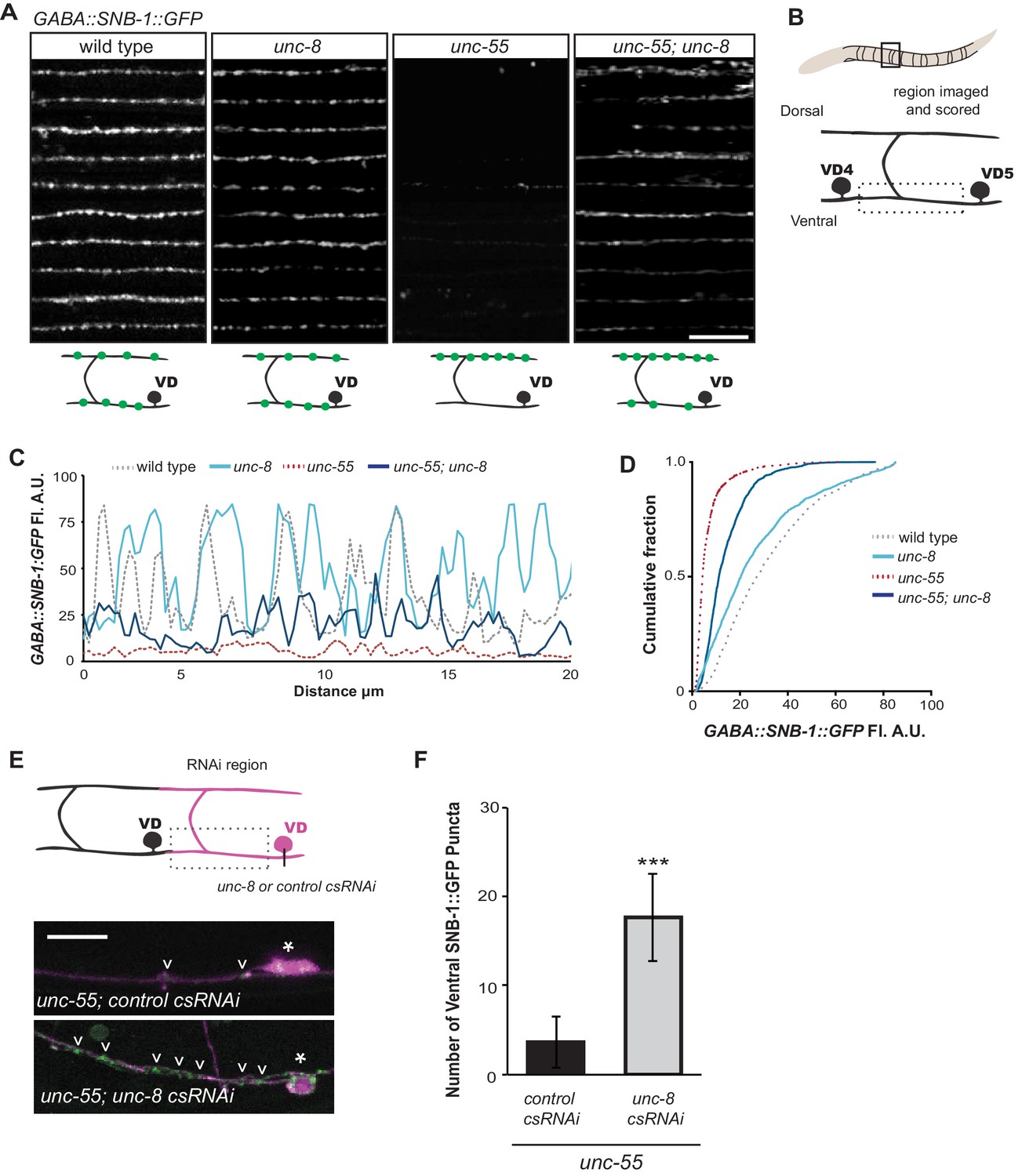
UNC-8 drives removal of ventral GABAergic synapses.
(A) Ventral GABA synapses labeled with GFP-tagged synaptobrevin (punc-25::SNB-1::GFP) for 10 adult animals. Wild-type and unc-8(tm5052) show similar distributions of SNB-1::GFP puncta. Ventral SNB-1::GFP is depleted from unc-55 due to VD remodeling, but partially restored in unc-55; unc-8 animals. (B) Data for Figure panels A, C, D were collected from the ventral nerve cord between VD4 and VD5. (C) SNB-1::GFP fluorescent intensity measurements from each genotype. Each line represents the pixel intensity over a 20 μm region of the VNC from a single representative animal. (D) Cumulative frequency curves for SNB-1::GFP fluorescence intensity for each genotype (n > 10 animals). unc-55 animals show a significant loss of ventral SNB-1::GFP fluorescence (p< 0.0001 vs wild type). SNB-1::GFP fluorescence is partially restored in unc-55; unc-8 animals, demonstrating the role of UNC-8 for synapse removal (p<0.0001 vs unc-55). p values calculated with Kruskal-Wallis and Dunn’s post test. (E) Knockdown of unc-8 by GABA-neuron-specific RNAi (unc-8 csRNAi) restored SNB-1::GFP puncta to the VNC of unc-55; juIs1 animals vs control animals expressing the empty vector RNAi (control csRNAi). GABAergic neurons are labeled with punc-25::mCherry (magenta). Asterisks denote GABA neuron cell bodies and arrowheads point to SNB-1::GFP-labeled ventral synapses. (F) Quantification of ventral synapses in the region anterior to each cell body expressing the RNAi construct (n > 60 animals). RNAi knockdown of unc-8 in unc-55 mutant GABA neurons significantly suppresses synapse removal (***p<0.001, Student’s t-test. Data are mean ± SD). Scale bars are 10 μm.
Figure 3—figure supplement 1
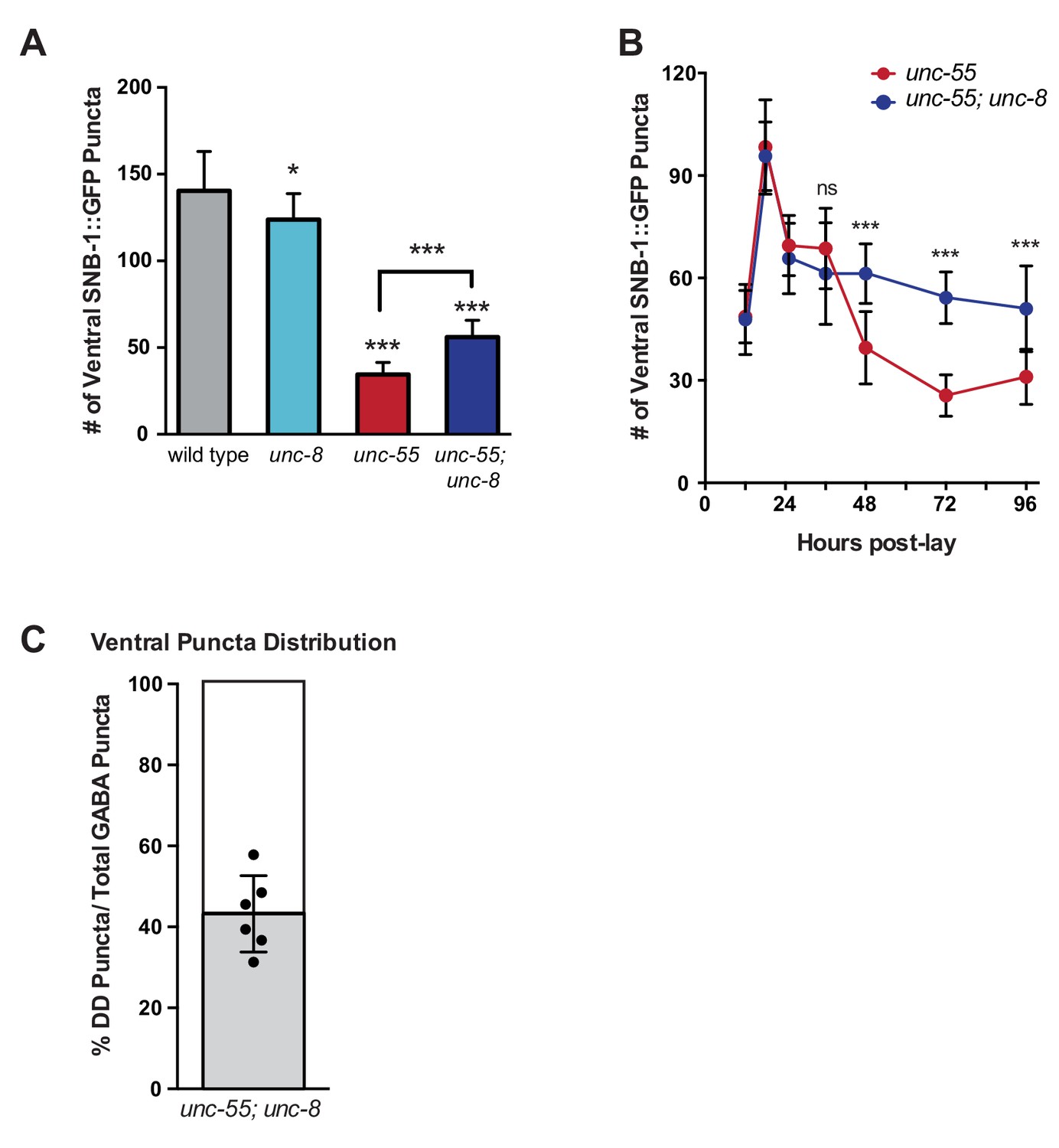
UNC-8 promotes synapse disassembly in remodeling GABAergic neurons.
Fluorescent puncta for the presynaptic protein SNB-1::GFP were counted in the ventral nerve cord from VD3 to VD11. (A) SNB-1::GFP puncta are removed from ventral synapses in unc-55 and this effect partially depends on unc-8 (***p<0.001, *p<0.05, n ≥ 10). One-Way ANOVA with Bonferroni correction, data are mean ± SD. (B) Time course experiments with unc-55 and unc-55; unc-8 animals show the removal of ventral GABAergic synapses (punc-25::SNB-1::GFP) over time. Ventral VD synapses are initially established at 24-hr post lay in both unc-55 and unc-55; unc-8 and these synapses are largely removed in unc-55 animals. Loss of unc-8 function disrupts ventral synapse removal in unc-55 animals and results in the retention of significantly more ventral synapses. (***p<0.001, ns is not significant, Student’s t-test). n ≥ 78 animals per genotype, data are mean ± SD. (C) Ventral GABA synapses in unc-55; unc-8 animals were co-labeled with punc-25::SNB-1::GFP, which marks all synapses (DD and VD) and pflp-13::mCherry::RAB-3, which labels DD synapses only. The proportion of ventral DD synapses (labeled with pflp-13::mcherry::RAB-3) in unc-55; unc-8 adults are shown in gray as a percentage of total ventral synapses.
Figure 3—figure supplement 2

UNC-8 functions cell autonomously and is sufficient to promote ventral synapse elimination in GABA neurons.
(A) UNC-8 function is cell-autonomous. Ventral synapses were counted in unc-55; unc-8 animals injected with GABA::unc-8cDNA and the GABA marker punc-25::mCherry (left). Asterisk denotes cell expressing unc-8cDNA and punc-25::mCherry. Scale bar is 10 μm, anterior to left. GABA neurons expressing unc-8cDNA showed fewer ventral puncta than neighboring unc-55; unc-8 neurons that do not express the unc-8cDNA transgene, indicating that UNC-8 functions in GABA neurons to promote synapse removal (***p<0.001, n ≥ 15 animals, Student’s t-test, data are mean ± SD). (B) Forced expression of unc-8cDNA in GABA neurons induces disassembly of of ventral synapses. VD neurons expressing unc-8cDNA (co-labeled with GABA::mCherry, VD8 denoted with arrow) show significantly fewer ventral SNB-1::GFP (punc-25::SNB-1::GFP) puncta than neighboring VD neurons that do not express unc-8cDNA which are unaffected (e.g., VD9 denoted with arrowhead). Anterior to left. Ectopic expression of unc-8cDNA in VD neurons has no effect on dorsal synapses (***p<0.001, ns is not significant, Student’s t-test, n ≥ 51 VDs (ventral) and n = 7 VDs (dorsal), data are mean ± SEM).
Figure 4 with 3 supplements
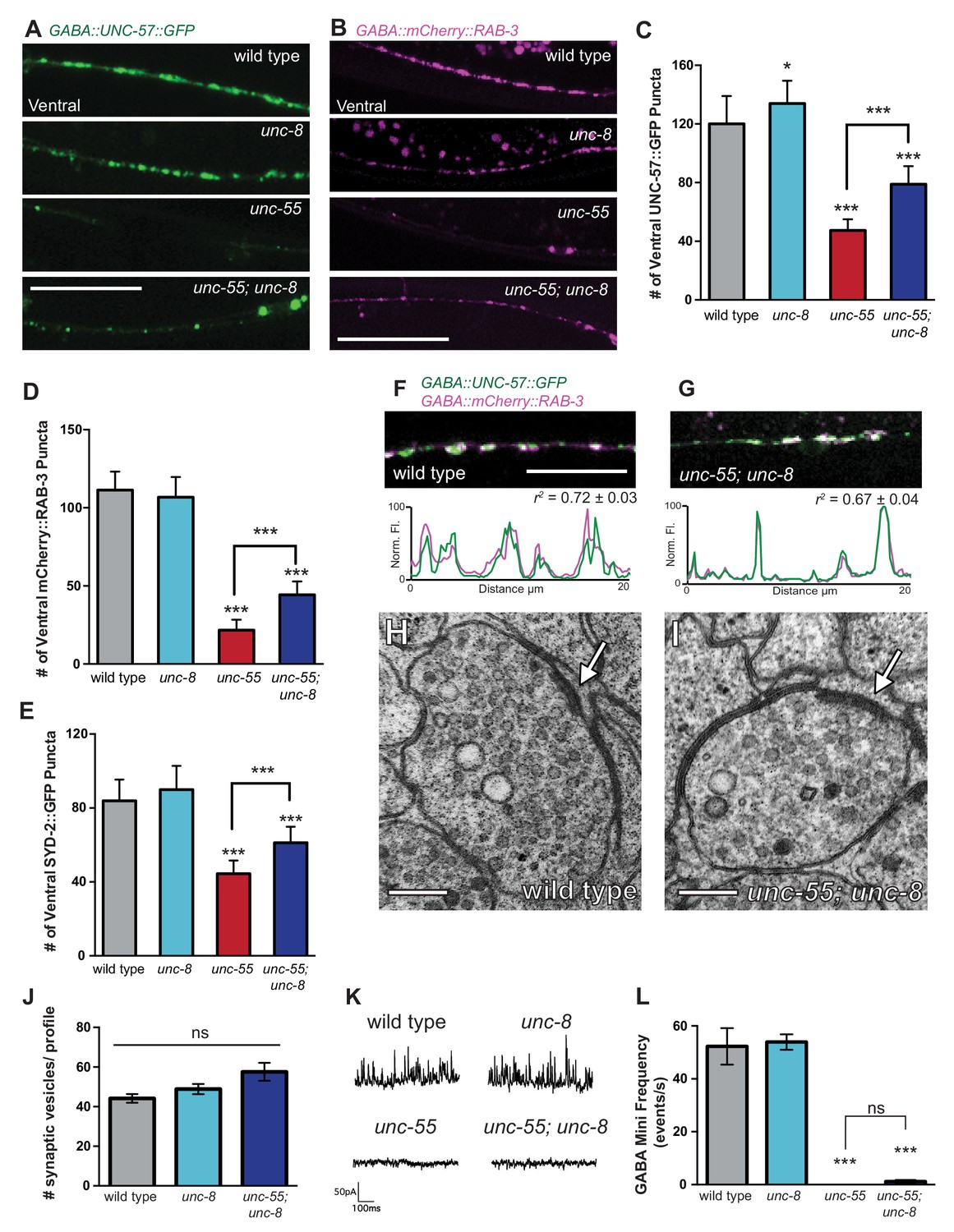
UNC-8 promotes disassembly of the presynaptic apparatus in GABAergic motor neurons.
Fluorescent puncta for presynaptic proteins (UNC-57::GFP, mCherry::RAB-3, and SYD-2::GFP) were counted in the ventral nerve cord from VD3 to VD11. (A/C) Representative images (A) and quantification (C) of endophilin/UNC-57 indicate that unc-8 promotes removal of UNC-57::GFP from ventral synapses in remodeling neurons (*p<0.05, ***p<0.001, n ≥ 25). (B/D) Representative images (B) and quantification (D) show reduced removal of the presynaptic G protein RAB-3 in unc-55; unc-8 animals (***p<0.001, n ≥ 21). Scale bars are 10 μM. (E) Efficient removal of the presynaptic density protein α-liprin/SYD-2 from ventral synapses in unc-55 requires unc-8 (***p<0.001, n ≥ 21, One-Way ANOVA with Bonferroni correction, data are mean ± SD). (F/G) GFP-tagged endophilin (punc-25::UNC-57::GFP) and mCherry::RAB-3 (punc-25::mCherry::RAB-3) are co-localized in GABA neurons of wild-type and unc-55;unc-8 animals. Representative images and normalized fluorescence intensity plots for a 20 μm region of the ventral nerve cord are shown. Scale bar is 10 μm. r2 is Pearson’s correlation coefficient (n > 10, mean ± SEM). Presynaptic components are co-localized in wild type (r2 = 0.72 ± 0.03) and unc-55;unc-8 (r2 = 0.67 ± 0.04). Average r2 value for unc-55;unc-8 is not statistically different from the average r2 value for wild type (p<0.001, Mann-Whitney test, see Figure 4—figure supplement 1). (H/I) Electron micrographs of GABA synapses with ventral muscles in (H) wild type and (I) unc-55;unc-8. No ventral GABA presynaptic densities were detected in unc-55. Arrows point to presynaptic density, scale bars are 200 nm. (J) Synaptic vesicles were quantified in ventral GABAergic synapses. Synapses in wild-type, unc-8 and unc-55;unc-8 animals contain comparable numbers of synaptic vesicles (N > 5 for each genotype, ns is not significant). (K) Representative traces of ventral mini-iPSCs from each genotype. (L) The high frequency of ventral mini-iPSCs in wild-type and in unc-8 animals were not observed in unc-55 or unc-55;unc-8 (***p<0.001, ns is not significant, n ≥ 5, data are mean ± SEM, One-Way ANOVA with Bonferroni correction).
Figure 4—figure supplement 1
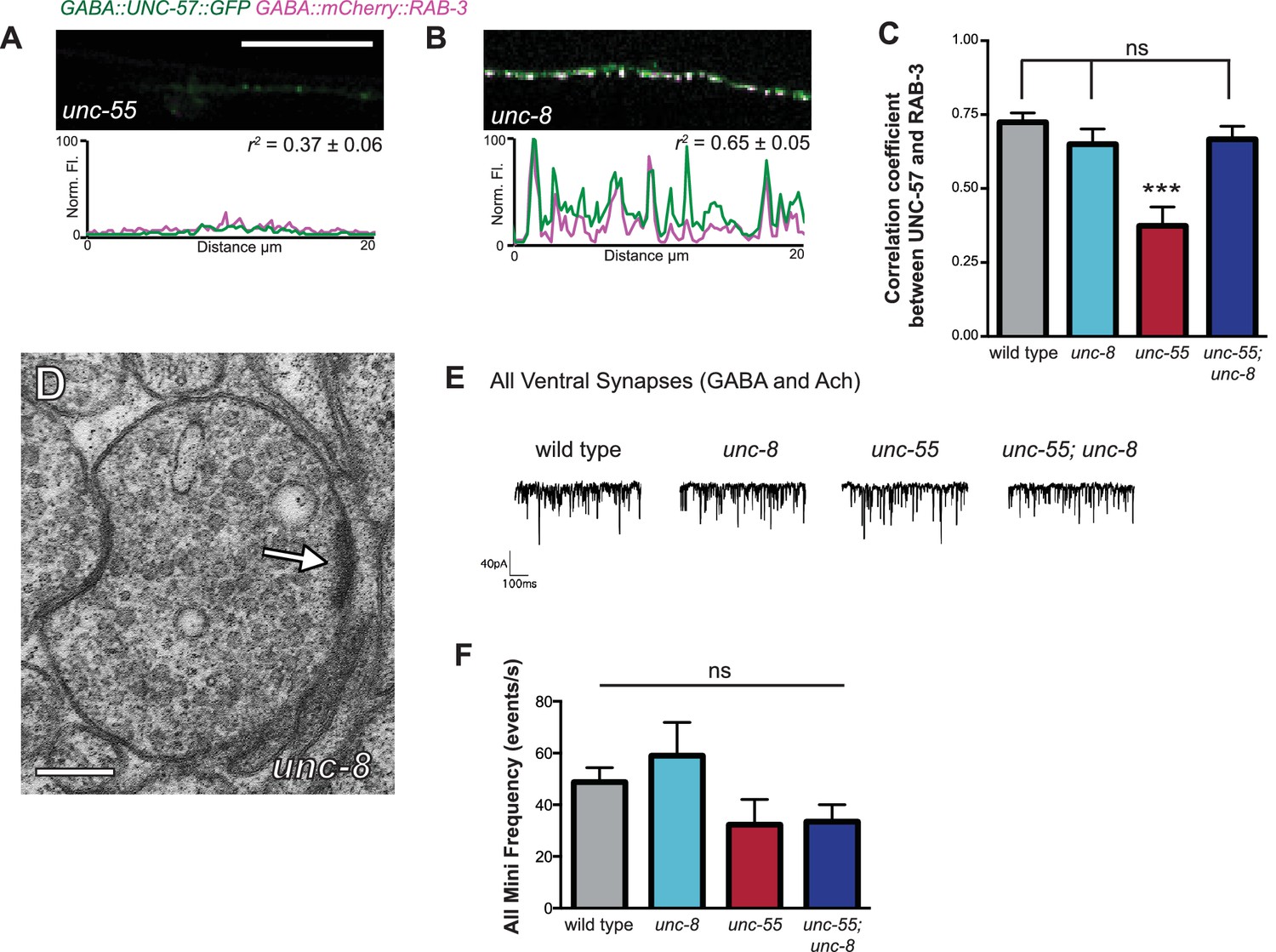
Ventral synapses in unc-55;unc-8 mutants are well-organized.
(A–B) unc-55 and unc-8 animals expressing GFP-tagged endophilin (punc-25::UNC-57::GFP) and mCherry::RAB-3 (punc-25::mCherry::RAB-3) were examined for co-localization. Representative images and normalized fluorescence intensity plots for a 20 μm ventral cord region of unc-55 and unc-8 animals are shown. Scale bar is 10 μm. r2 is the coefficient of determination (n > 10, mean ± SEM). UNC-57::GFP and mCherry::RAB-3 puncta are co-localized in unc-8 (r2 = 0.65 ± 0.05); but not in unc-55 mutants which show few residual ventral VD synapses (r2 = 0.37 ± 0.06). (C) Average coefficient of determination for UNC-57::GFP and mCherry::RAB-3. Wild-type, unc-8, and unc-55; unc-8 animals show no significant differences; whereas, unc-55 animals show much lower r2 values (n ≥ 11, data are mean ± SEM, Mann-Whitney test, ***p<0.001, ns is not significant). (D) Representative electron micrograph of a ventral GABAergic synapse in an unc-8 animal. Arrow points to presynaptic density, scale bar is 200 nm. (E) Representative traces of spontaneous activity (GABAergic and cholinergic) in ventral muscles. (F) Frequency of ventral spontaneous events (GABAergic and cholinergic) demonstrate that the loss of GABA activity in unc-55 and unc-55; unc-8 animals is not due to defects in neurotransmission to ventral muscles (n ≥ 5 animals, ns is not significant, mean ± SEM, One-Way ANOVA with Bonferroni correction).
Figure 4—figure supplement 2
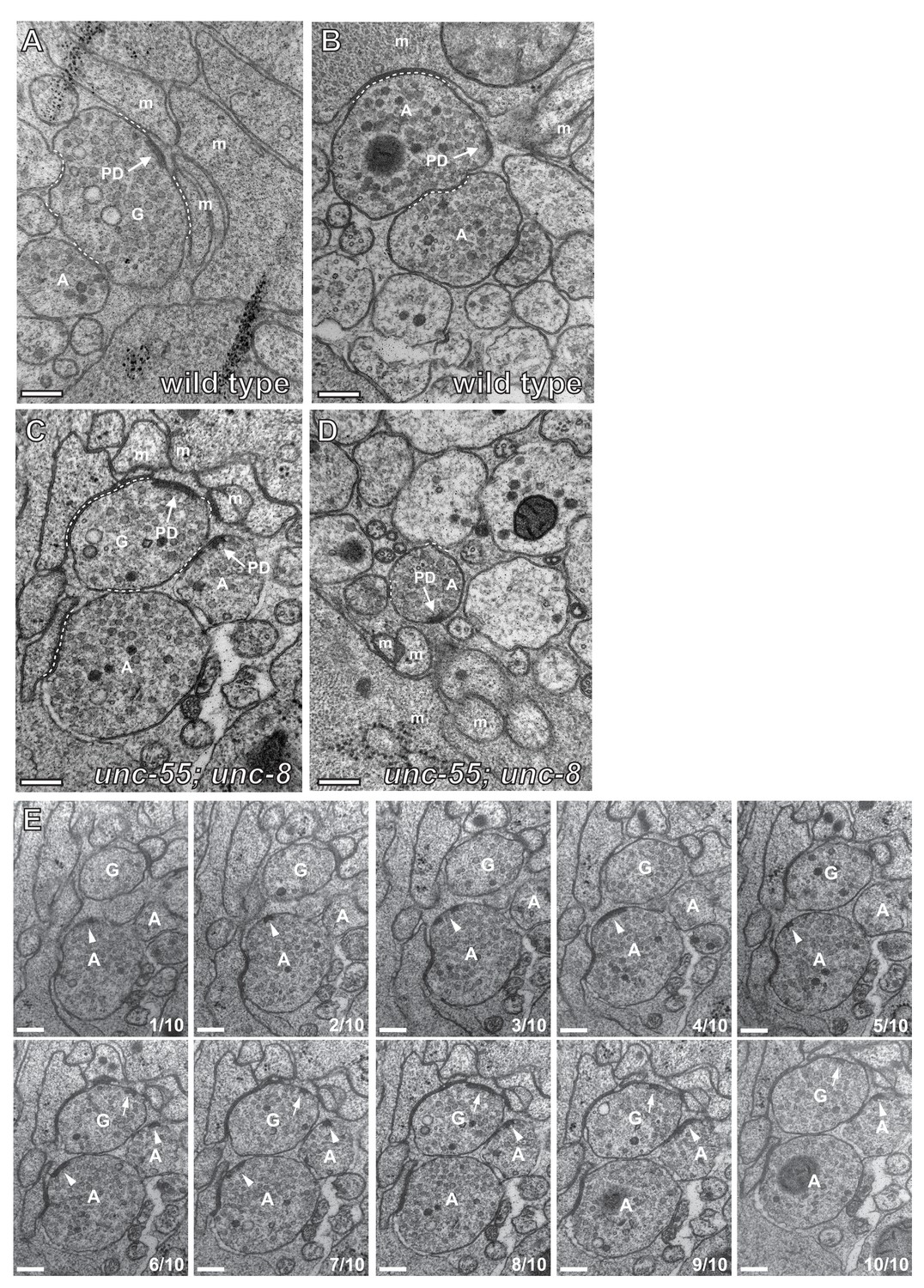
GABAergic and cholinergic synapses are detectable in electron micrographs of wild-type and unc-55; unc-8 animals.
(A/B) Ventral GABAergic (A) and cholinergic (B) presynaptic densities are visible in electron micrographs from wild-type animals. (C/D) Ventral GABAergic (C) and cholinergic (C/D) presynaptic densities were also detected in unc-55; unc-8 animals, suggesting that wild-type unc-8 function is required for the complete removal of ventral GABA synapses in unc-55 mutants. Arrows denote presynaptic densities, PD (presynaptic density), G (GABAergic neuron), A (cholinergic neuron), m (muscle arm), dotted lines are adherens or gap junctions. (E) Serial slices from the ventral nerve cord of the unc-55; unc-8 animals. 10 serial slices span a region with GABAergic neuron terminal labeled 'G' and cholinergic neuron terminal labeled 'A'. Each micrograph slice number is listed in lower right corner. Cholinergic (arrowhead) and GABAergic (arrow) presynaptic densities are labeled. GABAergic and cholinergic neuron synapses were distinguished by previously established criteria (see Materials and methods). Scale bars are 200 nm.
Figure 4—figure supplement 3

The postsynaptic UNC-49 GABAA receptor co-localizes with the presynaptic domains of remodeling GABAergic neurons.
Presynaptic regions (GABAergic neurons) are labeled with mCherry::RAB-3 (magenta) and the postsynaptic compartment (body muscle) is marked with UNC-49::GFP (green); arrows denote regions of co-localization. Arrowheads point to GABA neuron cell soma. Asterisks mark intestinal autofluorescent granules. (A). Pre-remodeling DD synapses with ventral muscles (early L1 larva, 2 hr post hatch). (B) Pre-remodeling VD synapses with ventral muscles (L2 larva, 18 hr post hatch). Note co-localization of mcherry::RAB-3 and UNC-49::GFP in both unc-55 and unc-55; unc-8 animals before remodeling. Scale bars are 10 μm. (C) Ventral GABAergic synapses in young adults. Few ventral mCherry::RAB-3-marked presynaptic domains are detected in the unc-55 mutant background due to ectopic remodeling but are abundant in both unc-8 and unc-55; unc-8. Scale bars are 25 μm. (D) Average correlation coefficient for ventral UNC-49::GFP and mCherry::RAB-3 puncta before remodeling. unc-8 is not significantly different from wild type (p>0.05, n ≥ 3 L1 larvae, ns is not significant) and unc-55; unc-8 is not significantly different from unc-55 (p>0.05, n ≥ 5 L2 larvae, ns is not significant, data are mean ± SD, Mann-Whitney test). (E) Average coefficients of determination for ventral UNC-49::GFP and mCherry::RAB-3 puncta after remodeling are not significantly different among wild-type, unc-8 and unc-55; unc-8 animals. unc-55 shows significantly lower values likely due to the relative depletion of mCherry::RAB-3 vs UNC-49::GFP (**p<0.01, ns is not significant, n ≥ 5 young adults, data are mean ± SD, Kruskal-Wallis with Dunn’s multiple comparisons test). Mutant strains were unc-55(e1170) and unc-8(tm5052).
Figure 5 with 1 supplement
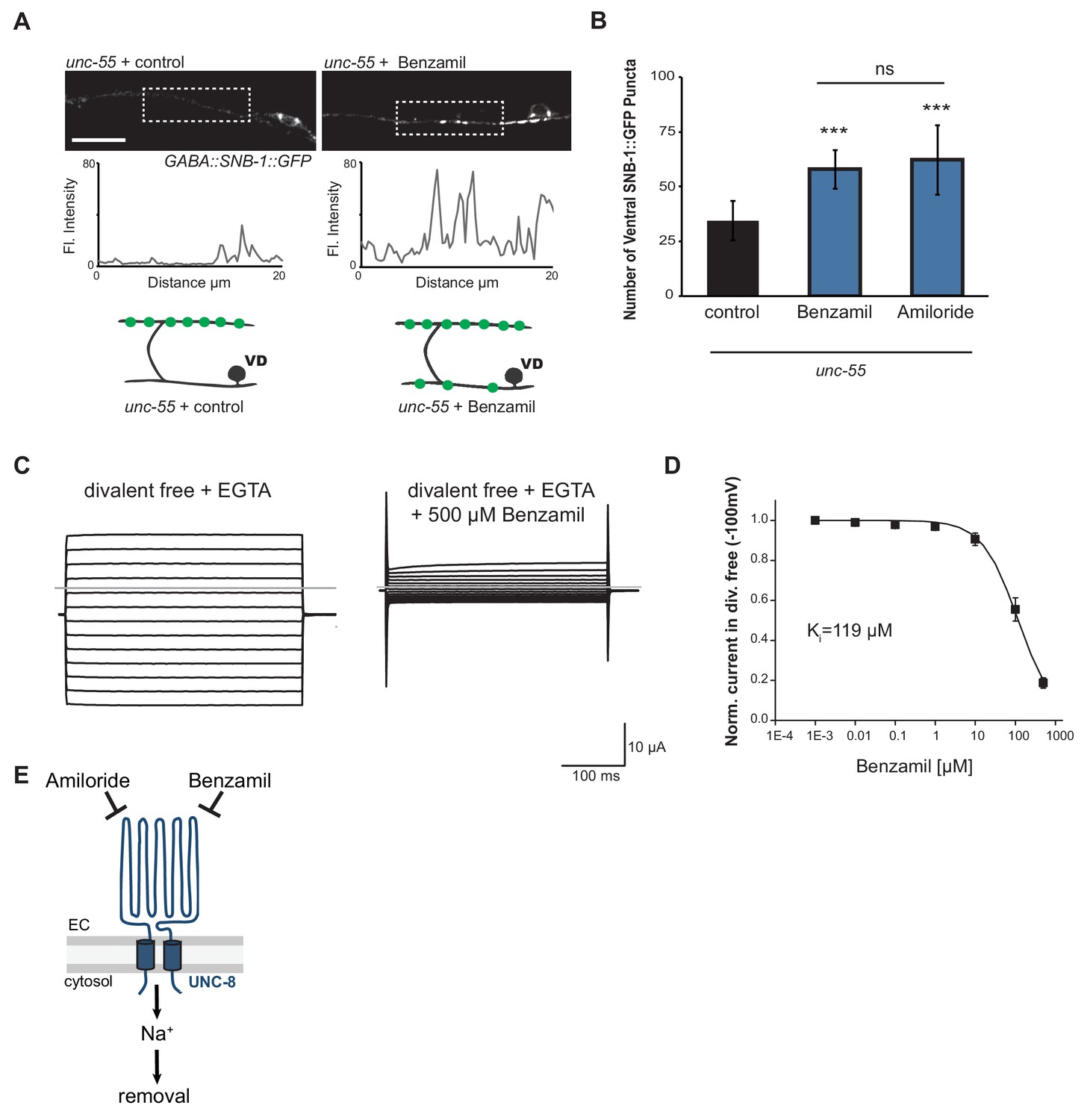
UNC-8 cation channel activity promotes the removal of ventral synapses in remodeling GABA neurons.
(A) Representative images and fluorescence intensity plots (generated from 20 μm dashed region) of SNB-1::GFP-marked ventral GABA neuron synapses in unc-55 animals treated with either 3 mM Benzamil or water (control). Scale bar is 10 μm. (B) Benzamil and Amiloride antagonize the removal of ventral GABA synapses in unc-55 mutant animals. Ventral GABA neuron synapses were quantified by counting SNB-1::GFP puncta (***p<0.001, ns is not significant, n ≥ 25 animals, data are mean ± SD, One-Way ANOVA with Bonferroni correction). (C) Benzamil blocks UNC-8(G387E) current in Xenopus oocytes. Representative currents from oocyte expressing UNC-8(G387E) in a bath of divalent cation-free solution plus EGTA. Currents elicited by 20mV voltage steps from -160mV to +100mV. The holding potential was -30mV. The gray line represents the zero current level (left). The same oocyte exposed to 500µM Benzamil (right). (D) Benzamil dose-response curve in divalent cation-free bath solution. Currents recorded with Benzamil were normalized against recordings in divalent cation-free bath solution plus EGTA at -100mV. Data were fitted to the Boltzmann's equation to derive Ki = 119 µM (n = 10 oocytes). Data are mean ± SEM. (E) Pharmacological inhibition of UNC-8 channel activity with either Benzamil or Amiloride blocks removal of ventral synapses in remodeling GABA neurons synaptic remodeling (EC is extracellular).
Figure 5—figure supplement 1
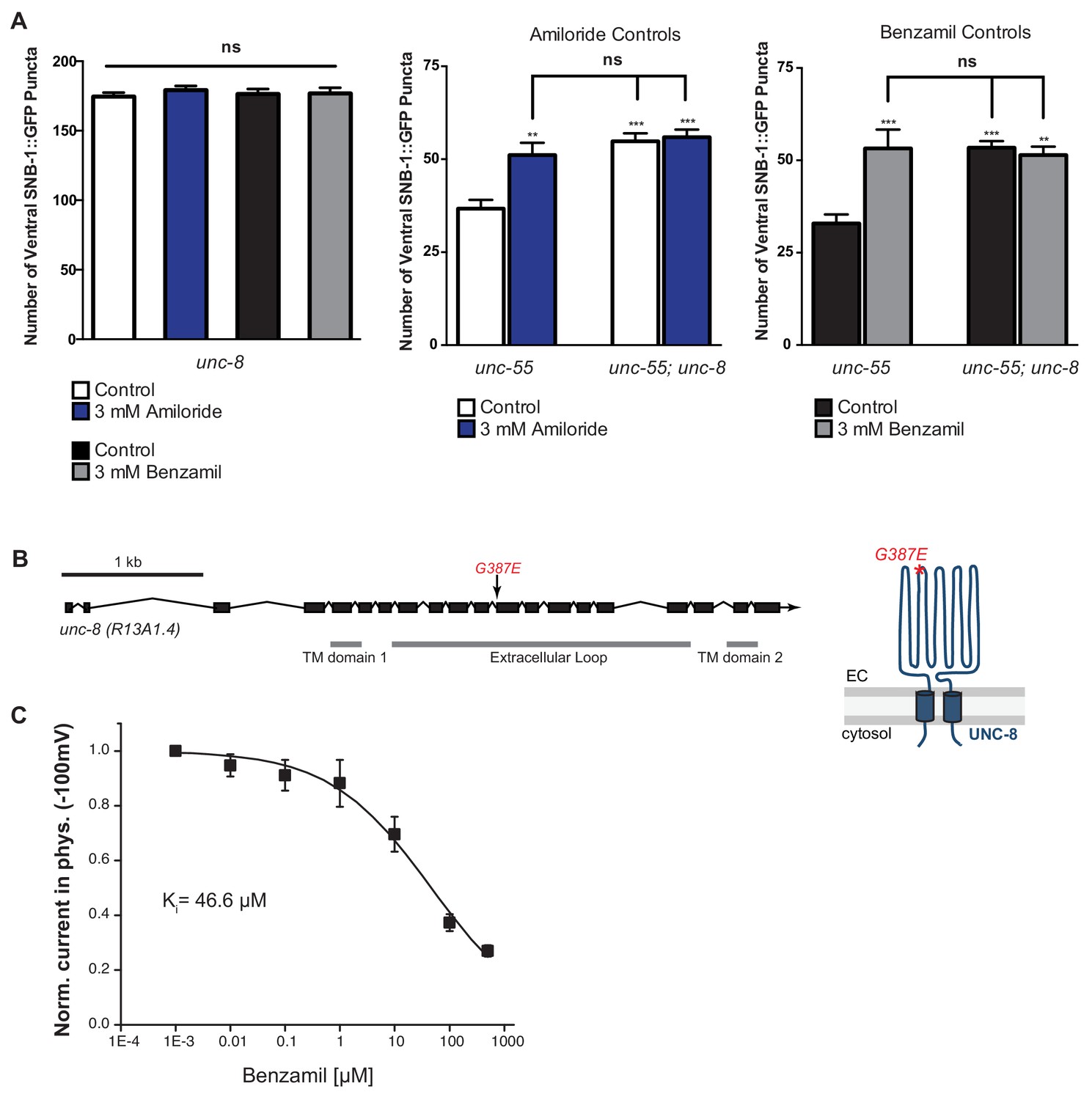
UNC-8 is required for the inhibitory effect of Benzamil and Amiloride on synaptic removal.
(A) Treatment of unc-8 and unc-55; unc-8 animals with either 3 mM Amiloride or 3 mM Benzamil does not limit removal of ventral synapses. Vehicle control for Amiloride and Benzamil treatment is water (n ≥ 10, **p<0.01, ***p<0.001, ns is not significant, mean ± SEM, One-Way ANOVA with Bonferroni correction). (B) The point mutant G387E renders the UNC-8 channel constitutively active and was used for in vitro oocyte experiments. Schematic shows the unc-8 genomic region and denotes the locations of transmembrane domains TM1 and TM2 and the predicted extracellular loop region. (C) UNC-8(G387E) currents recorded in physiological solution plus the DEG/ENaC inhibitor Benzamil at -100mV were normalized against currents recorded in physiological solution at -100mV. Data were fitted with the Boltzmann's equation for Ki = 46.6 µM (n = 10 oocytes). Data are mean ± SEM.
Figure 6
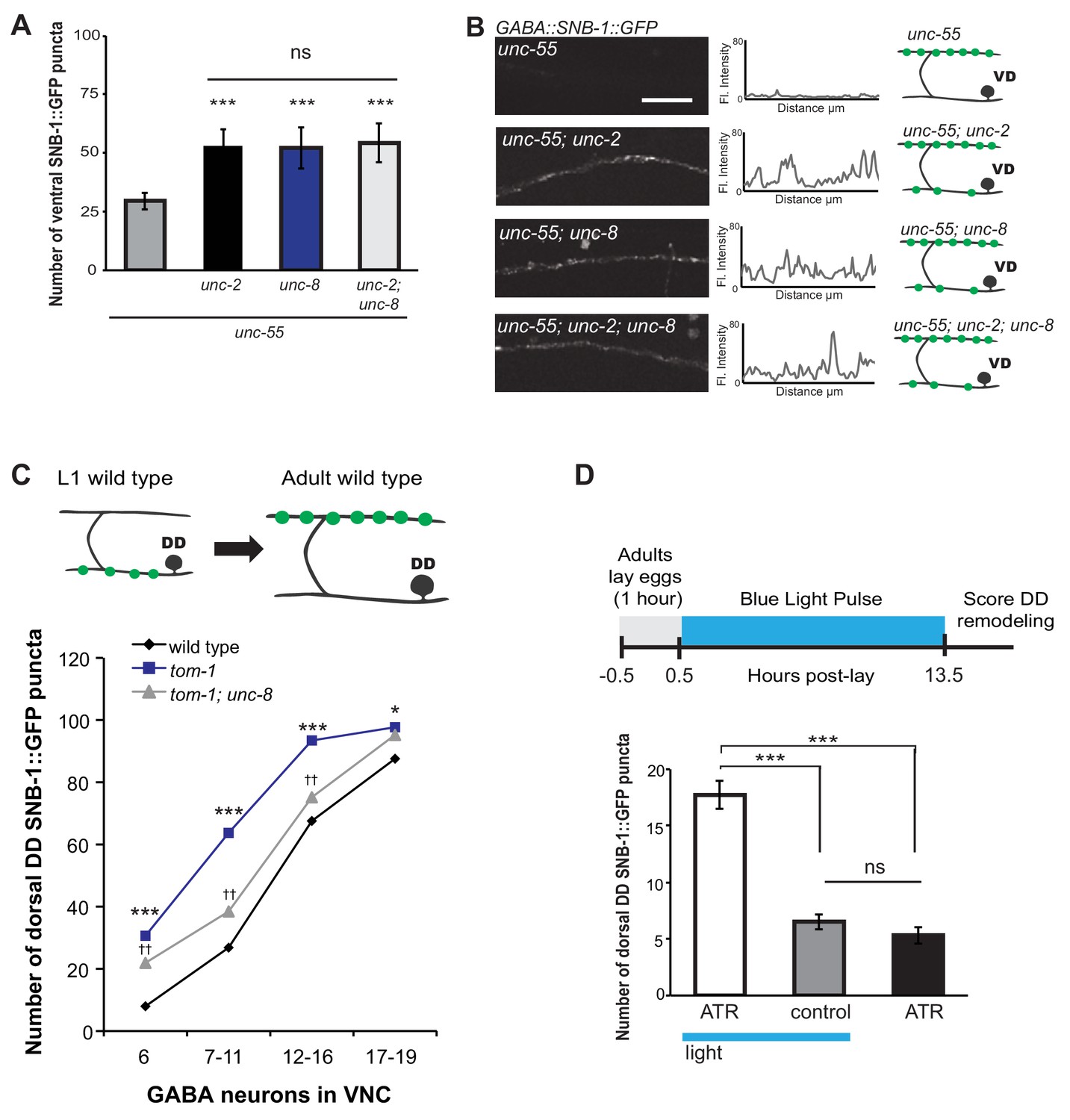
UNC-8 drives synaptic remodeling in an activity-dependent pathway that requires neurotransmitter release.
(A) Loss-of-function mutations in either unc-2 (VGCC) or unc-8 impairs removal of ventral SNB-1::GFP in unc-55 animals. The unc-8 mutation does not enhance the unc-55;unc-2 remodeling defect, demonstrating that UNC-2/VGCC and UNC-8/DEG/ENaC function in a common pathway to promote GABA synapse removal (***p<0.001 vs unc-55, One-Way ANOVA with Bonferroni correction, data are mean ± SD, n ≥ 17). (B) Representative images and insets show fluorescence intensity plots over a 20 μm region of the ventral nerve cord for each genotype. Scale bar is 5 μm. (C) DD synapses are precociously remodeled in tom-1 mutants. This effect is suppressed in unc-8; tom-1 animals (*p<0.05, ***p<0.001 vs wild type, ††p<0.01 vs tom-1 Student’s t-test, results pooled from > 3 independent experiments per genotype) Plots are normalized for the total number of GABA neurons labeled with SNB-1::GFP in the ventral cord to account for developmental delay in tom-1 mutants. (D) Optogenetic stimulation of channelrhodopsin (ChR2)-induced activity in GABA neurons for 13 hr (0.5 Hz) results in precocious appearance of SNB-1::GFP marked DD synapses in the dorsal nerve cord (n ≥ 18 animals, ***p<0.001, data are mean ± SEM, One-Way ANOVA with Bonferroni correction), ATR is all-trans retinal. Mutant alleles were unc-2(e55) and tom-1(ok2437).
Figure 7
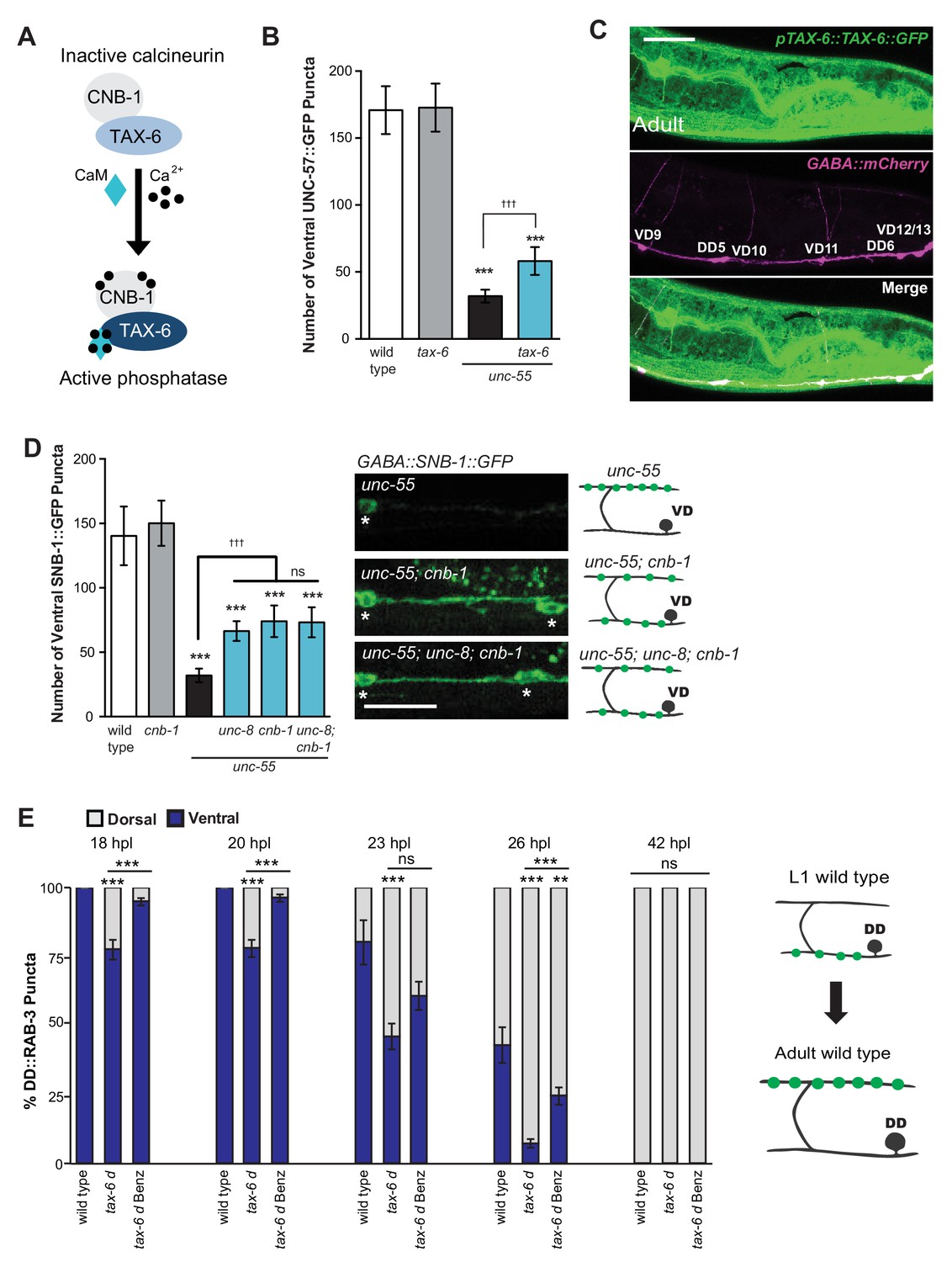
The calcium/calmodulin-dependent phosphatase calcineurin promotes GABA synapse removal in the UNC-8 pathway.
(A) Calcineurin A and B subunits (TAX-6 and CNB-1, respectively) require calcium and calmodulin (CaM) to activate phosphatase activity. (B) Loss of tax-6 partially suppresses the unc-55 remodeling phenotype in GABA neurons (***p<0.001, ns is not significant, One-Way ANOVA with Bonferroni correction, data are mean ± SD, n ≥ 25). (C) GFP-tagged TAX-6 under the control of the tax-6 promoter region (ptax-6::TAX-6::GFP) is expressed in GABA neurons (punc-47::mCherry). Scale bar is 20 μm. (D) CNB-1 is required for remodeling in unc-55 animals. Loss of cnb-1 function does not enhance the unc-55; unc-8 remodeling defect, suggesting that calcineurin and UNC-8 promote synapse removal in a common genetic pathway (left, ***p<0.001 compared to wild type, tttp<0.001 compared to unc-55, ns is not significant, One-Way ANOVA with Bonferroni correction, data are mean ± SD, wild type n = 10, mutants n = 20). Representative images of ventral nerve cords in unc-55, unc-55; cnb-1 and unc-55; unc-8; cnb-1 animals. Asterisks denote GABA neuron soma, scale bar is 20 μm (right). (E) Gain-of-function tax-6 (tax-6d) mutants remodel precociously and this effect is suppressed by Benzamil. Percentage of ventral (blue) vs dorsal (gray) DD synapses (pflp-13::GFP::RAB-3, **p<0.01, ***P<0.001, ns is not significant, One-Way ANOVA with Bonferroni correction, n ≥ 8 animals per timepoint, data are mean ± SEM). Results for tax-6d and for the tax-6d control for Benzamil treatment (see Materials and methods) are combined because they were not significantly different. Benz denotes 3mM DEG/ENaC inhibitor Benzamil. Mutant alleles were tax-6(p675), tax-6d(jh107), cnb-1(ok276).
Figure 8
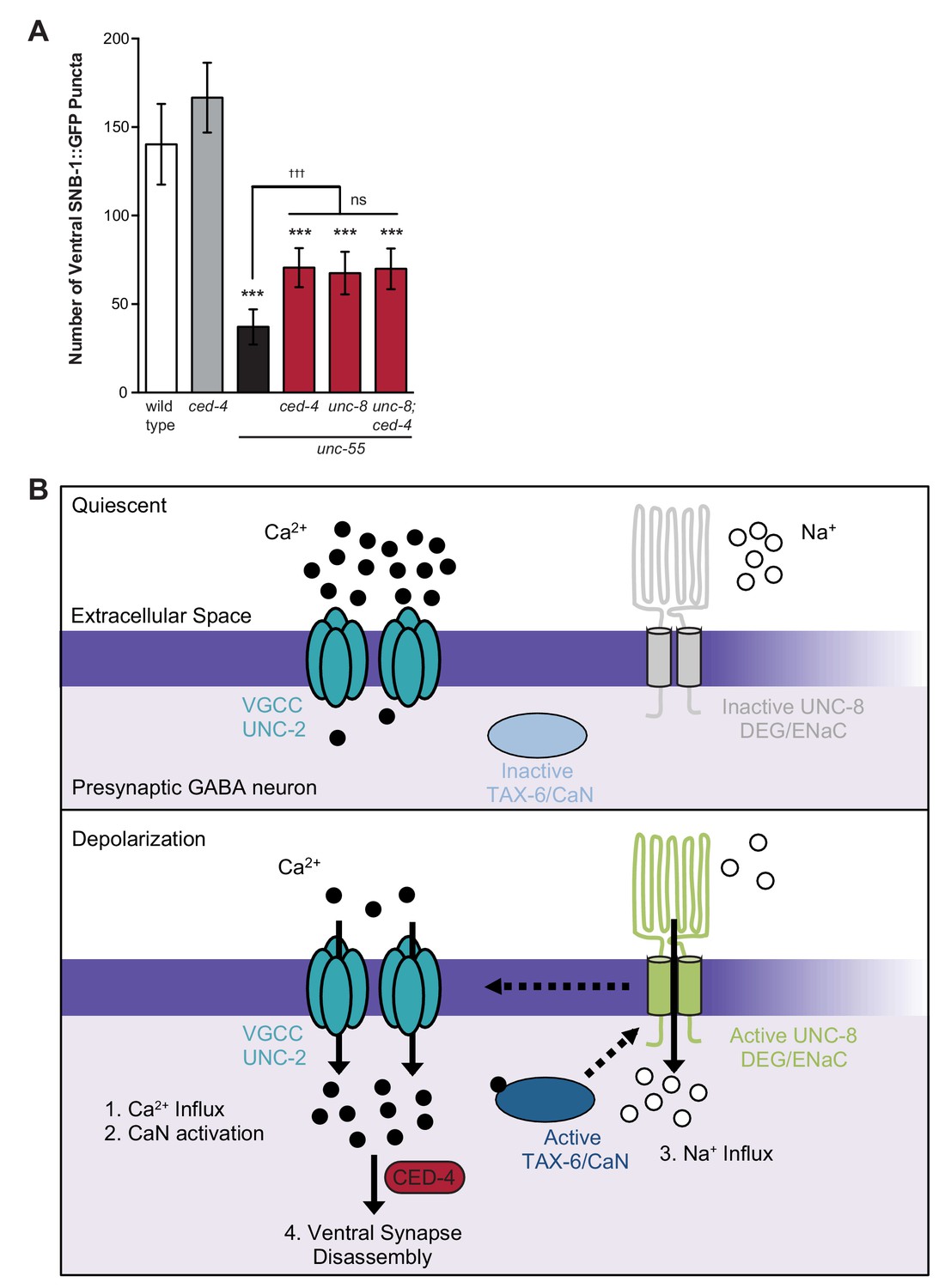
Model of UNC-8-driven synapse disassembly in the GABA neuron remodeling program.
(A) A loss-of-function mutation in the pro-apoptotic gene ced-4 partially suppresses GABA neuron remodeling in unc-55 animals. Genetic ablation of unc-8 in the triple mutant unc-55; ced-4; unc-8 mutants does not enhance the remodeling defect and thus suggests that UNC-8 and CED-4 promote synapse elimination in a common pathway (***p<0.001 compared to wild type, tttp<0.001 compared to unc-55, ns is not significant, One-Way ANOVA with Bonferroni correction, data are mean ± SD, wild type n = 10, mutants n ≥ 20) Mutant allele is ced-4(n1162). (B) Model of a calcium-dependent mechanism for removal of the presynaptic apparatus. A DEG/ENaC channel containing UNC-8 is not active (gray) in quiescent GABA motor neurons (top panel). (1) GABA neuron depolarization activates the voltage-gated calcium channel (VGCC), UNC-2, to allow calcium entry (black circles bottom panel). (2) Intracellular calcium activates the calcium/calmodulin-dependent phosphatase, calcineurin (CaN/TAX-6). (3) CaN phosphatase may activate UNC-8, which results in the movement of sodium ions (white circles) into the presynaptic GABA neuron, further depolarizing the presynaptic membrane and activating VGCCs. This positive feedback loop is predicted to further elevate intracellular calcium. (4) Our results show that UNC-8 drives the removal of presynaptic components and functions in a common genetic pathway with calcium signaling and with the apoptotic protein CED-4. Therefore, we hypothesize that selective expression of UNC-8 in remodeling GABA neurons effectively boosts the level of intracellular calcium to activate a CED-4-dependent pathway for removal of the presynaptic apparatus.
Additional files
-
Supplemental file 1
C. elegans strains used in this study.
- https://doi.org/10.7554/eLife.14599.019
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
The DEG/ENaC cation channel protein UNC-8 drives activity-dependent synapse removal in remodeling GABAergic neurons
eLife 5:e14599.
https://doi.org/10.7554/eLife.14599




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}