An FGF-driven feed-forward circuit patterns the cardiopharyngeal mesoderm in space and time
- College of Arts and Science, New York University, United States
Abstract
In embryos, multipotent progenitors divide to produce distinct progeny and express their full potential. In vertebrates, multipotent cardiopharyngeal progenitors produce second-heart-field-derived cardiomyocytes, and branchiomeric skeletal head muscles. However, the mechanisms underlying these early fate choices remain largely elusive. The tunicate Ciona emerged as an attractive model to study early cardiopharyngeal development at high resolution: through two asymmetric and oriented divisions, defined cardiopharyngeal progenitors produce distinct first and second heart precursors, and pharyngeal muscle (aka atrial siphon muscle, ASM) precursors. Here, we demonstrate that differential FGF-MAPK signaling distinguishes between heart and ASM precursors. We characterize a feed-forward circuit that promotes the successive activations of essential ASM determinants, Hand-related, Tbx1/10 and Ebf. Finally, we show that coupling FGF-MAPK restriction and cardiopharyngeal network deployment with cell divisions defines the timing of gene expression and permits the emergence of diverse cell types from multipotent progenitors.
https://doi.org/10.7554/eLife.29656.001Introduction
Developmental genetics knowledge guided progress towards driving mammalian stem cells into forming pure cultures of selected cell types in vitro (e.g. [Kattman et al., 2011; Mazzoni et al., 2011; Peljto and Wichterle, 2011]). By contrast, in the embryo, pluripotent cells generate diverse cell types in defined proportions, as they divide before individual daughter cells adopt distinct fates.
Subsets of the heart and head/neck myocytes recently emerged as related derivatives of multipotent progenitors located in the cardiopharyngeal mesoderm (Diogo et al., 2015a; Tzahor, 2009; Tzahor and Evans, 2011). Early lineage tracing, transplantations and controlled explant culture experiments demonstrated that the anterior splanchnic/pharyngeal mesoderm of amniote embryos can produce either skeletal muscles or heart tissue, depending upon exposure to growth factors and signaling molecules (Nathan et al., 2008; Tirosh-Finkel et al., 2006; Tzahor et al., 2003; Tzahor and Lassar, 2001). Clonal analyses in the mouse further revealed the existence of common Mesp1-expressing progenitors for subsets of the second heart field-derived cardiomyocytes and branchiomeric facial, jaw, neck and even œsophageal muscles (Gopalakrishnan et al., 2015; Lescroart et al., 2014; Lescroart et al., 2015; Lescroart et al., 2010; Lescroart et al., 2012). In pluripotent stem cells, controlled Mesp1 expression can drive mesodermal progenitors towards cardiac and/or skeletal muscle fates (Bondue et al., 2008; Chan et al., 2016; Chan et al., 2013). Proper development of the pharyngeal apparatus and second heart field derivatives require shared inputs from Tbx1, Nkx2.5 and Islet1 transcription factors (e.g. [Cai et al., 2003; George et al., 2015; Jerome and Papaioannou, 2001; Kelly et al., 2004; Merscher et al., 2001; Mosimann et al., 2015; Nevis et al., 2013; Prall et al., 2007; Tzahor and Evans, 2011; Vitelli et al., 2002a; Watanabe et al., 2012; Witzel et al., 2017; Yagi et al., 2003; Zhang et al., 2006]). Taken together, this growing body of evidence points to the existence of a mesodermal field of multipotent progenitors capable of producing either SHF-derived cardiomyocytes or branchiomeric skeletal muscles in early vertebrate embryos (Diogo et al., 2015; Mandal et al., 2017). However, the mechanisms that distinguish fate-restricted heart and head muscle precursors remain largely elusive.
The tunicate Ciona, which is among the closest living relatives to the vertebrates (Delsuc et al., 2006; Putnam et al., 2008), has emerged as a simple chordate model to characterize multipotent cardiopharyngeal progenitors and the mechanisms that initiate heart vs. pharyngeal muscle fate choices (Kaplan et al., 2015; Razy-Krajka et al., 2014; Stolfi et al., 2010; Tolkin and Christiaen, 2016; Wang et al., 2013). Ciona tailbud embryos possess two multipotent cardiopharyngeal progenitors on either side. Like their vertebrate counterparts, these cells emerge from Mesp+ progenitors towards the end of gastrulation; they are induced by FGF-MAPK signaling and have been termed trunk ventral cells (aka TVCs; [Christiaen et al., 2008; Davidson and Levine, 2003; Davidson et al., 2006; Davidson et al., 2005; Satou et al., 2004; Stolfi et al., 2010]). TVCs activate conserved cardiac markers, including Hand, Gata4/5/6 and Nk4/Nkx2-5, and migrate as polarized pairs of cells, until the left and right pairs meet at the ventral midline and begin to divide asymmetrically along the mediolateral axis (Figure 1A; [Christiaen et al., 2008; Davidson et al., 2005; Satou et al., 2004; Stolfi et al., 2010]). The first oriented asymmetric divisions produce small median first heart precursors (FHPs), and large lateral second trunk ventral cells (STVCs), which specifically activate Tbx1/10 (Davidson et al., 2005; Stolfi et al., 2010; Wang et al., 2013). STVCs later divide again to produce small median second heart precursors (SHPs), and large lateral atrial siphon muscle founder cells (ASMFs), which activate Ebf (aka COE; [Razy-Krajka et al., 2014; Stolfi et al., 2010; Stolfi et al., 2015]). The transcription factors Hand-related (Hand-r)/Notrlc, which is expressed in the TVCs and maintained in the STVCs and ASMFs after each division, and Tbx1/10 are required for Ebf activation in the ASMFs, whereas Nk4/Nkx2.5 represses Tbx1/10 and Ebf expression in the second heart precursors (SHPs)(Razy-Krajka et al., 2014; Tolkin and Christiaen, 2016; Wang et al., 2013). Conversely, Tbx1/10 and Ebf inhibit cardiac markers, and likely determinants, such as Gata4/5/6 and Hand (Razy-Krajka et al., 2014; Stolfi et al., 2010, 2014a; Wang et al., 2013). These regulatory cross-antagonisms underlie the transition from transcriptionally primed multipotent progenitors to separate fate-restricted precursors, by limiting the deployment of the heart- and pharyngeal-muscle-specific programs to their corresponding specific precursors (Kaplan et al., 2015).
Figure 1 with 2 supplements see all
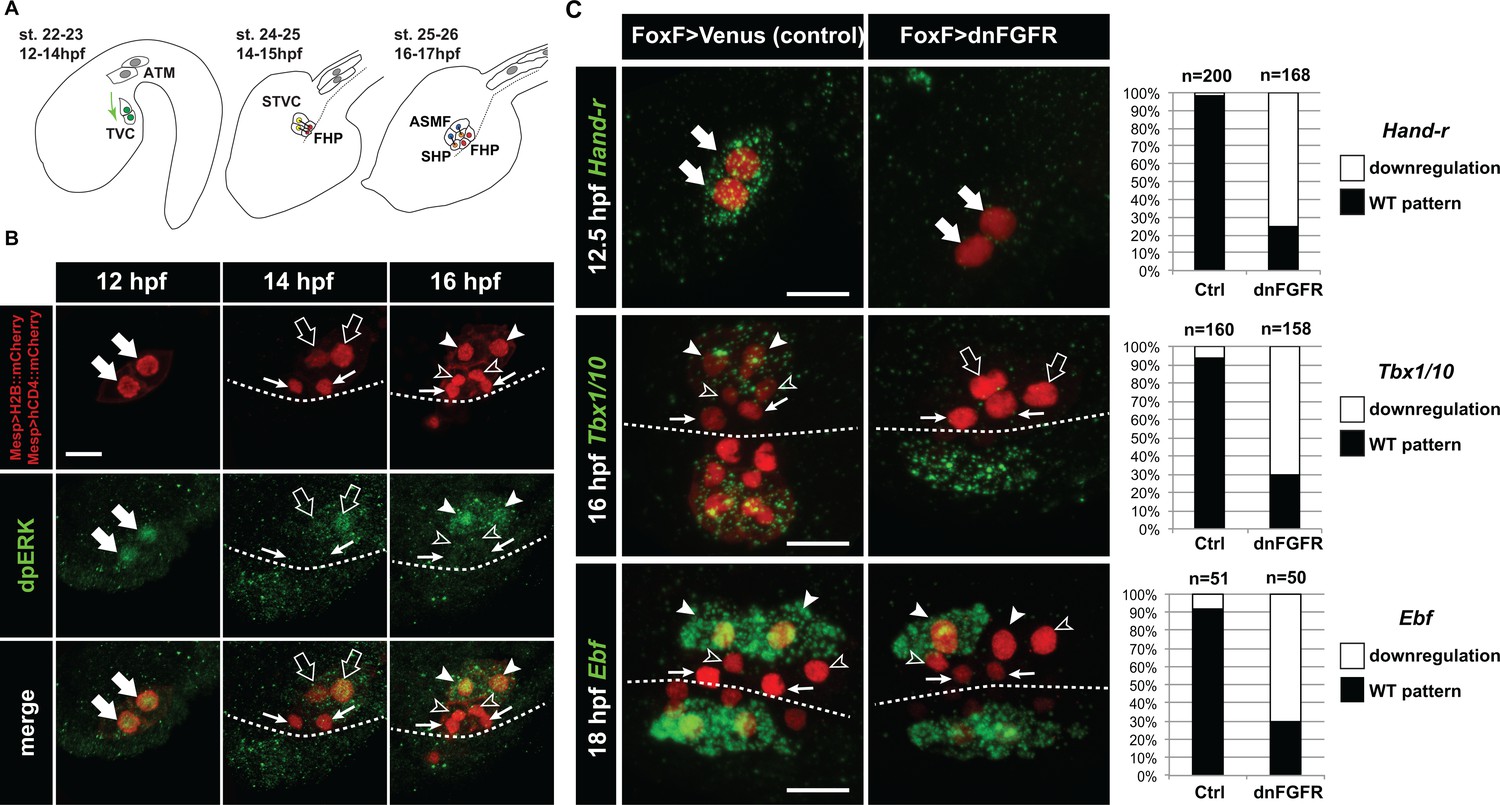
Spatio-temporal restriction of ERK activity reflects FGF requirement for the specification of cardiopharyngeal progenitors.
(A) Schematic of Ciona development showing asymmetric cell divisions and resulting cell fates of the cardiopharyngeal mesoderm (CPM). Embryonic and larval stages (St) according to (Hotta et al., 2007) with hours post fertilization (hpf) at 18°C. Anterior tail muscle (ATM, gray), trunk ventral cell (TVC, green), secondary TVC (STVC, green), first heart precursor (FHP, red), second heart precursor (SHP, orange), atrial siphon founder cell (ASMF, blue). Black bars link sister cells. Dashed lines: ventral midline. The first stage presents a quasi-lateral view while the second and third stages present quasi-ventral views. Anterior is to the left. Scale bar, 50 µm. (B) ERK activity visualized by anti-dpERK antibody (green). TVCs and their progeny are marked by mCherry driven by Mesp and revealed by anti-mCherry antibody (red). H2B::mCherry and hCD4::mCherry accumulate in the nuclei and at the cell membrane, respectively. Arrowheads indicate STVCs and ASMFs at 14 and 16 hpf, respectively. Arrows indicate FHPs and open arrowheads mark SHPs. Anterior to the left. Scale bar, 10 µm. See also Figure 1—figure supplement 1 for broader time series of dpERK immunostaining in the B7.5 lineage. (C, D) TVC-specific overexpression of dnFGFR induces loss of expression of key lateral CPM markers visualized by in situ hybridization. (C) Representative expression patterns of key CPM genes (Hand-related, Tbx1/10, Ebf) in control embryos (Ctrl, electroporated with Foxf(TVC):bpFOG-1>Venus) and TVC-specific dnFGFR expression (electroporated with Foxf(TVC):bpFOG-1>dnFGFR::mCherry) individuals. TVCs and progeny are marked with Mesp > NLS::lacZ (red). Loss of expression in half of the TVC progeny, as presented for Ebf, is assumed to be due to left-right mosaicism. Arrowheads mark the ASMFs. Anterior is to the left. Scale bar, 10 µm. (D) Corresponding histograms with the phenotype proportions. For simplicity, loss of gene expression in half or all of the TVCs and their progeny were combined in the same category. ‘n’ corresponds to the number of individual halves documented per condition.
Here, we identify regulatory mechanisms ensuring the emergence of diverse fate-restricted precursors from multipotent progenitors. We show that differential FGF-MAPK signaling, feed-forward regulatory circuits and coupling with the cell cycle control the spatially restricted activation of Tbx1/10 and Ebf, successively, thus permitting the emergence of both first and second heart precursors, and ASM/pharyngeal muscle precursors from common multipotent progenitors.
Results
MAPK signaling is active in the multipotent cardiopharyngeal progenitors and progressively restricted to the pharyngeal muscle precursors
During the earliest stages of cardiopharyngeal development in ascidians, multipotent progenitors display multilineage transcriptional priming, (Razy-Krajka et al., 2014; Stolfi et al., 2014b), and subsequent regulatory cross-antagonisms segregate distinct cardiopharyngeal programs to their corresponding fate-restricted progenitors (Stolfi et al., 2010; Wang et al., 2013); reviewed in [Kaplan et al., 2015]). For instance, the ASM-specific factor Ebf is necessary and sufficient to terminate the heart program and impose a pharyngeal muscle fate (Razy-Krajka et al., 2014; Stolfi et al., 2010). However, the symmetry-breaking events leading to cardiopharyngeal mesoderm patterning and ASM-specific expression of Ebf remain unknown. We surmised that differential signaling inputs determine the stereotyped spatio-temporal patterning of early cardiopharyngeal progenitors.
The Ciona homologs of specific FGF-MAPK pathway components, including FGF receptor substrate 2/3 (Frs2/3; [Gotoh et al., 2004]), Ets.b, and Fgf4/5/6, are preferentially expressed in the TVCs, in the STVCs and in the ASMFs as cells transition from multipotent progenitor to distinct heart vs. ASM fate-restricted states (Razy-Krajka et al., 2014). These patterned expressions of MAPK effector genes prompted us to evaluate a role for FGF-MAPK pathway in cardiopharyngeal fate decisions.
We first used an antibody specific to the dual phosphorylated form of Extracellular Regulated Kinase (dpERK) to monitor Mitogen Activated Protein Kinase (MAPK) activity in the cardiopharyngeal mesoderm. We detected dpERK staining in the newly born TVCs, marked by the B7.5-lineage-specific Mesp >H2B::mCherry transgene (Figure 1—figure supplement 1), as previously observed (Davidson et al., 2006). We also detected weaker but persistent dpERK staining in the TVCs during migration (Figure 1 and Figure 1—figure supplement 1). Following the first and second asymmetric divisions of the TVCs and STVCs, dpERK staining was successively restricted to the more lateral STVCs and ASMFs, respectively (Figure 1A,B; Figure 1—figure supplement 1).
The canonical FGF/Ras/MEK/ERK pathway is necessary and sufficient to promote pharyngeal muscle specification in the cardiopharyngeal lineage
This exclusion of MAPK activity from the medial first and second heart precursors opened the possibility that differential ERK activity is required for proper STVC and ASMF vs. heart precursors fate decisions. In Ciona, signaling through the sole FGF receptor (FGFR) governs ERK activity in several developmental processes, including neural induction (Bertrand et al., 2003; Hudson et al., 2003) and central nervous system patterning (Haupaix et al., 2014; Racioppi et al., 2014; Stolfi et al., 2011; Wagner et al., 2014), early endomesoderm and notochord fate specification (Imai et al., 2002; Picco et al., 2007; Shi and Levine, 2008; Shi et al., 2009; Yasuo and Hudson, 2007). Notably, FGF-MAPK signaling is active in the Mesp+ cardiogenic B7.5 blastomeres (Imai et al., 2006; Shi and Levine, 2008), where targeted misexpression of a dominant negative form of FGFR (dnFGFR) blocks TVC induction (Christiaen et al., 2008; Davidson et al., 2006). We used a TVC-specific Foxf enhancer (Foxf(TVC):bpFog-1>dnFGFR::mCherry, hereafter called Foxf>dnFGFR; [Beh et al., 2007]), to bypass early effects and achieve later misexpression of dnFGFR in the TVCs and their progeny. Importantly, although TVC fate specification and the onset of Foxf expression require FGF-MAPK signaling (Beh et al., 2007; Davidson et al., 2006), we confirmed that this perturbation altered neither initial TVC induction, nor the expression of the Foxf driver (Figure 1—figure supplement 2). Consistent with proper TVC induction, Foxf>dnFGFR prevented neither TVC migration nor asymmetric divisions, but it abolished the expression of both Tbx1/10 in the STVCs and Ebf in the ASMFs (Figure 1C). This data indicate that FGF-MAPK signaling is required in the cardiopharyngeal progenitors and/or their progeny for ASM fate specification, beyond the initial TVC induction.
Upon FGF-MAPK-dependent induction, the TVCs express Hand-related/Hand-r (renamed after Notrlc/Hand-like; [Christiaen et al., 2008; Davidson and Levine, 2003; Davidson et al., 2006; Satou et al., 2004; Stolfi et al., 2015; Woznica et al., 2012]), which encodes a basic helix-loop-helix (bHLH) transcription factor necessary for Ebf expression in the ASMFs (Razy-Krajka et al., 2014). Moreover, the Hand-r TVC enhancer contains putative Ets1/2 binding sites, which are necessary for reporter gene expression, and presumably mediate the transcriptional inputs of FGF-MAPK (Woznica et al., 2012). Since Hand-r and Foxf expressions start at approximately the same time in newborn TVCs, we used Foxf>dnFGFR, which did not alter the onset of Hand-r expression in the TVCs (Figure 1—figure supplement 2), to test whether the maintenance of Hand-r expression in migratory TVCs requires prolonged FGF-MAPK inputs. Foxf>dnFGFR inhibited Hand-r expression in late TVCs (Figure 1C), indicating that sustained Hand-r expression requires continuous FGF-MAPK signaling, as did TVC-expressed FGF-MAPK pathway components (Figure 1—figure supplement 2).
To test whether the spatial restriction of MAPK activity explains the patterned expressions of Hand-r, Tbx1/10 and Ebf following asymmetric cell divisions, we used gain-of-function perturbations to force FGF-MAPK activity throughout the cardiopharyngeal mesoderm and assayed gene expression (Figure 2). We focused on the canonical FGF-MAPK pathway where signal transduction involves Ras, Raf, MEK and ERK downstream of FGFR and upstream of transcriptional effectors (Lemmon and Schlessinger, 2010). We first used M-RasG22V, a defined constitutively active form of M-Ras, which mediates FGF signaling in Ciona, where other classical Ras genes are missing (Keduka et al., 2009), and focused on Htr7 and Tbx1/10 expression following the first asymmetric TVC division in 15 hours post-fertilization (hpf) embryos. Htr7 encodes a trans-membrane G-protein coupled receptor and, like Hand-r, its expression and maintenance in the TVCs require MAPK activity (Figure 1—figure supplement 2; [Razy-Krajka et al., 2014]), and become restricted to the lateral STVC following asymmetric division. However, Htr7 mRNAs are cleared more rapidly from the FHPs, making the patterned expression easier to analyze than that of Hand-r (Figures 2 and 3D; [Razy-Krajka et al., 2014]). Importantly, misexpression of M-RasG22V using the TVC-specific Foxf enhancer altered cell division asymmetry and/or orientation in under 50% of the embryos, still allowing us to identify large lateral and small median cells in a small majority of embryos (Figure 2—figure supplement 1). Compared to control embryos overexpressing wild-type M-Ras (M-RasWT), TVC-specific gain of M-Ras function caused both persistent Htr7 expression and ectopic activation of Tbx1/10 in the small median cells following asymmetric divisions (Figure 2B,C). Similarly, Foxf>M-RasG22V-expressing 18hpf larvae displayed ectopic Ebf activation throughout the cardiopharyngeal mesoderm (Figure 2B,C). This cannot be simply accounted for by general disruption of cell division patterns at this later stage (Figure 2—figure supplement 1), since similar disruptions can be caused by Foxf>dnFGFR, which inhibits Ebf expression (Figure 1C). These results indicated that forced M-Ras activity is sufficient to upregulate STVC and ASMF markers ectopically. This is consistent with the idea that spatially defined signaling upstream of M-Ras restricts MAPK activity, thus localizing STVC- and ASM-specific gene activities.
Figure 2 with 2 supplements see all
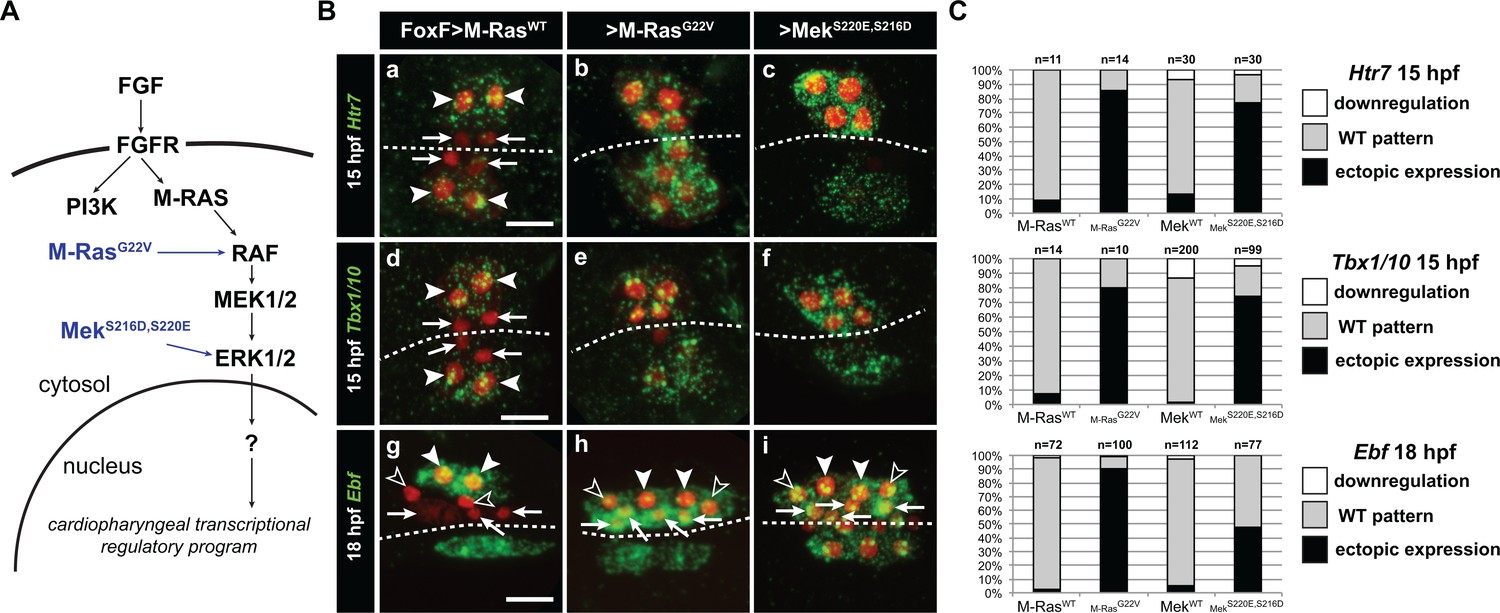
Constitutively active M-Ras and MEK are sufficient to impose a pharyngeal muscle fate in the cardiopharyngeal lineage.
(A) Diagram of the FGF-MAPK transduction pathway with constitutive activation by M-RasG22V and MEKS216D,S220E mutants. (B) Expression patterns of markers of the lateral TVC progeny, Htr7 (a, b, c,), Tbx1/10 (d, e, f) and Ebf (g, h, i), visualized by in situ hybridization following TVC-specific over-expression of M-RasWT (as control), M-RasG22V and MEKS216D,S220E. M-RasWT overexpression (a, d, g) does not alter the wild-type spatial expression patterns of Htr7, Tbx1/10 and Ebf in lateral TVC derivatives (STVC and ASMF) and excluded from the median heart precursors. TVC-specific over-expression of M-RasG22V (b, e, h) or MEKS216D,S220E (c, f, i) induces ectopic expression of STVC and/or ASMF markers (Htr7, Tbx1/10 and Ebf) in the more median cells, that normally form cardiac precursors. Arrowheads indicate STVCs and ASMFs at 15 and 18 hpf, respectively. Arrows indicate FHPs and open arrowheads mark SHPs. At 18 hpf, the FHPs start dividing or have divided into 4 cells. Anterior to the left. Scale bar, 10 µm. (C) Corresponding histograms: Larvae with TVC-specific over-expression of MEKWT retain the wild-type expression patterns. For simplicity, ectopic expressions in half to all of the cardiac precursors were combined in the same phenotype category. ‘n’ corresponds to the number of embryo halves documented per condition. See also Figure 1—figure supplement 2.
Figure 3
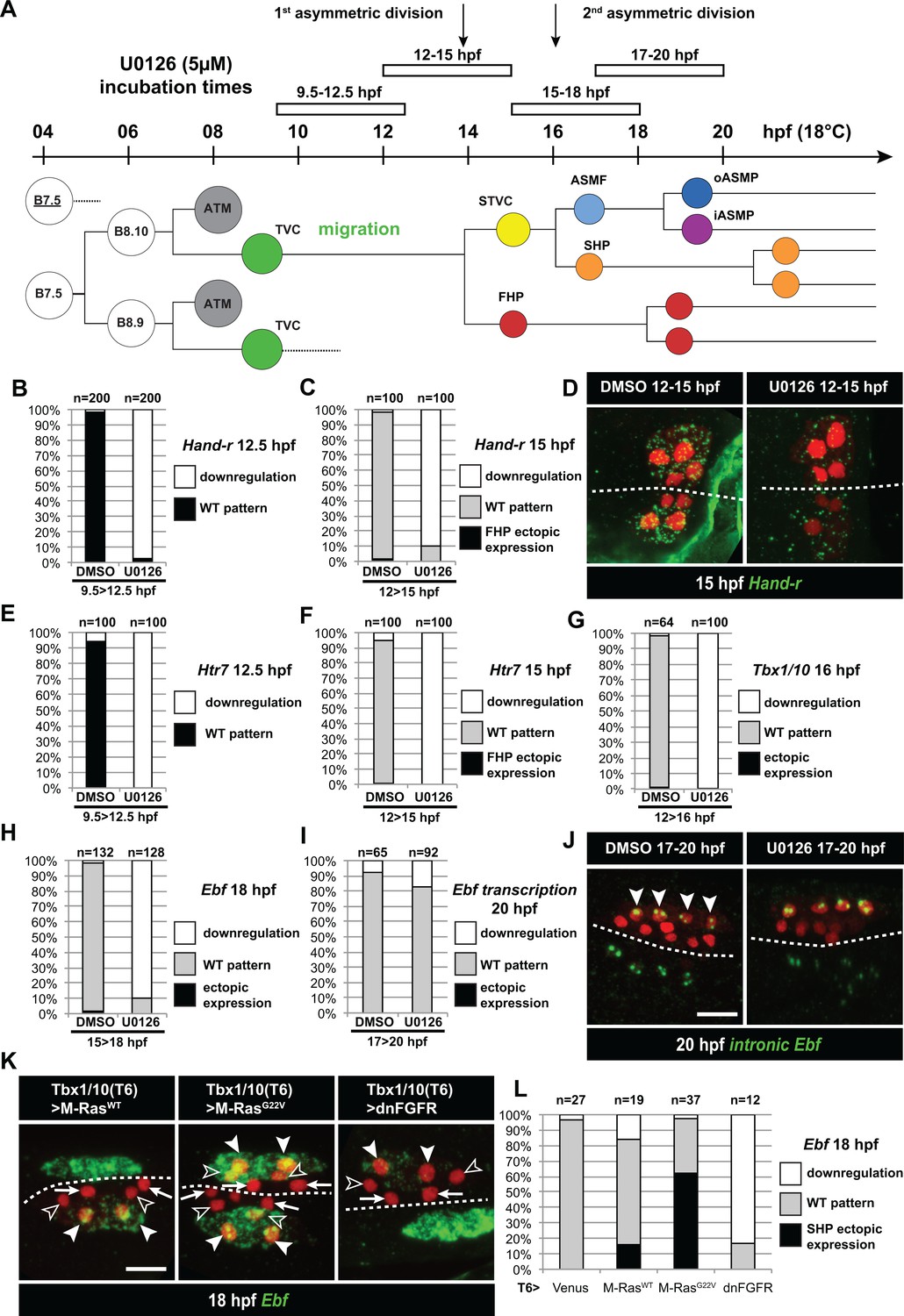
Temporal requirement for MAPK activity permits the progressive deployment of the cardiopharyngeal regulatory program.
(A) Summary of the CPM cell lineage showing the different U0126 treatments with regard to the timing of cell divisions. Abbreviations and color codes as in Figure 1. (B, C) Proportions of embryo halves with wild-type or downregulated expression of Hand-r at 12.5 hpf (B) and 15 hpf (C) following 3 hr incubations in U0126 (with DMSO as control treatment). (D) Hand-r expression visualized by in situ hybridization at 15 hpf in control (DMSO treated) and U0126 treated embryos. In control embryos, Hand-r remains expressed in the STVCs and downregulated in the FHPs. In U0126 (12–15 hpf) treated embryos, downregulation of Hand-r expression is observed throughout the TVC progeny (STVCs and FHPs), suggesting inhibition of transcription and inheritance of remnant transcripts following TVC divisions. (E, F) Proportions of embryo halves with wild-type or downregulated expression of Htr7 at 12.5 hpf (E) and 15 hpf (F) following 3 hr incubations in U0126 (with DMSO as control treatment). (G) Proportions of larvae with wild-type expression or downregulated expression of Tbx1/10 at 16 hpf following 4 hr incubation in U0126 (with DMSO as control). (H) Proportions of larvae with wild-type or downregulated expression of Ebf at 18 hpf following a three hour incubation in U0126 (with DMSO as control). (I) Proportions of larvae with wild-type or downregulated transcription of Ebf at 18 hpf following a three hour incubation in U0126 (DMSO as vehicle control). (J) Pattern of nascent Ebf transcripts visualized by in situ hybridization with intronic probes (green) at 20 hpf. The nuclear dots reveal the active transcription sites in the four ASMPs per side in larvae, both control/DMSO- and U0126-treated from 17 to 20 hpf. (K) Ebf expression (green) in 18hpf larvae expressing control M-RasWT, constitutively active M-RasG22V or dominant negative dnFGFR under the control of the T12 element, an STVC-specific Tbx1/10 enhancer. Arrows: first heart precursors (FHP); open arrowhead: second heart precursors (SHPs); closed arrowheads: ASM founder cells (ASMFs); dotted line: midline. (L) Proportions of larvae with wild-type or downregulated expression of Ebf at 18 hpf in larvae with Venus (control), M-RasWT, M-RasG22, or dnFGFR driven by Tbx1/10 cis-regulatory sequence and overexpressed in the STVCs. ‘n’: number of individual halves documented per condition.
To further probe the signal transduction pathway, we engineered a constitutively active version of the Ciona Mek1/2 protein by introducing phosphomimetic mutations of two conserved serine residues in the catalytic domain, as previously shown for the mammalian homolog (Cowley et al., 1994; Mansour et al., 1994). Early misexpression of MekS220E,S216D in the B7.5 lineage using the Mesp enhancer caused ectopic TVC induction, mimicking the effects of gain of Ets1/2 function (Figure 2—figure supplement 2; [Davidson et al., 2006]). As seen with M-RasG22V, TVC-specific misexpression of MekS220E,S216D using the Foxf enhancer also caused ectopic expression of Htr7 and Tbx1/10, and Ebf in 15 and 18hpf larvae, respectively (Figure 2B,C). Taken together, these results indicate that activity of the canonical FGF-Ras-MEK-ERK pathway is progressively restricted to the STVC and ASMF, and is both necessary and sufficient to promote STVC- and ASMF-specific gene expressions.
Continuous FGF-MAPK activity is required for the successive activations of Tbx1/10 and Ebf
FGF-MAPK signaling is sufficient and necessary to maintain Hand-r expression in late TVCs (Figure 1), and Hand-r is necessary for Ebf expression in the ASMF (Razy-Krajka et al., 2014). Therefore, it is possible that later FGF-MAPK signaling is dispensable for Tbx1/10 and Ebf activation and ASM specification, as long as STVC and ASMF cells inherit sustained levels of Hand-r mRNAs and/or proteins. To disentangle late from early requirements of FGF-MAPK signaling, we incubated embryos at different stages with the MEK/MAPKK inhibitor U0126, which abolishes dual ERK phosphorylation and the initial MAPK-dependent TVC induction in Ciona embryos (Figure 1—figure supplement 1; [Davidson et al., 2006; Hudson et al., 2003]). MEK inhibition during TVC migration (i.e. between 9.5 and 12.5 hpf, Figure 3A) blocked the expression of Hand-r and Htr7 in late TVCs (Figure 3B,E). U0126 treatments in late TVCs, and through the first asymmetric division (i.e. between 12 and 15 hpf, Figure 3A) did not alter TVC division patterns (Figure 2—figure supplement 1), but it blocked both the maintenance of Hand-r and Htr7, and the activation of Tbx1/10 in the STVCs (Figure 3C,D,F,G). Finally, MEK inhibition in late STVCs and through asymmetric divisions (i.e. between 15 and 18 hpf) also did not alter STVC divisions (Figure 2—figure supplement 1), but it blocked the ASMF-specific expression of Ebf (Figure 3H). These results indicate that continuous MEK activity is required throughout cardiopharyngeal development to successively activate TVC-, STVC-, and ASMF-expressed genes.
Since Ebf expression is maintained for several days in the ASMF derivatives as they differentiate into body wall and siphon muscles (Razy-Krajka et al., 2014), we tested whether continued MEK activity is also required for the maintenance of Ebf expression past its onset and cells' commitment to an ASM fate. Using both regular and intron-specific antisense probes, which specifically detect nascent transcripts (Wang et al., 2013), we showed that later MEK inhibition (i.e. U0126 incubation between 17 and 20 hpf) did not block the maintenance of Ebf transcription in the ASMPs (Figure 3I,J). This indicates that sustained MEK activity is required until the onset of Ebf expression, but not beyond, and the maintenance of Ebf expression during ASM development is independent of MAPK.
Since U0126 treatments affect the whole embryo, we sought to further confirm the later roles for FGF-MAPK signaling specifically in the cardiopharyngeal mesoderm. To this aim, we used an STVC-specific enhancer from the Tbx1/10 locus (termed T6; Figure 3K,L; Figure 4—figure supplement 1; (Tolkin and Christiaen, 2016; Racioppi et al., in preparation) to drive expression of either dnFGFR or the constitutively active M-RasG22V starting at ~14 hpf, and assayed Ebf expression at 18hpf (Figure 3K,L). These perturbations minimally affected the cell division patterns (Figure 2—figure supplement 1), such that cells corresponding to FHP, SHP and ASMF could be identified by their position relative to the midline in many embryos (Figure 3K). M-RasG22V misexpression caused conspicuous ectopic Ebf expression in the SHPs, whereas dnFGFR-mediated inhibition of MAPK activity blocked Ebf activation in the lateral ASMFs. These results support the notion that localized FGF-MAPK activity is necessary and sufficient for ASMF-specific expression of Ebf.
Coherent feed-forward circuits for cardiopharyngeal mesoderm patterning and ASM fate specification
The above results indicate that Hand-r, Tbx1/10 and Ebf require ongoing FGF-MAPK activity for their successive activations in the TVCs, STVCs and ASMFs, respectively. We previously showed that RNAi and/or CRISPR-mediated inhibition of either Hand-r or Tbx1/10 function blocks Ebf activation in the ASMFs, where both Hand-r and Tbx1/10 expressions are maintained (Razy-Krajka et al., 2014; Tolkin and Christiaen, 2016; Wang et al., 2013). We used epistasis assays to systematically test whether early regulators mediate the effects of FGF-MAPK on later gene expression and ASM fate specification, or whether FGF-MAPK signaling acts both upstream and in parallel to early regulators in a more complex regulatory circuit.
We first revisited the regulatory relationships between FGF-MAPK, Hand-r and Tbx1/10 in late TVCs and early STVCs. We validated single guide RNAs (sgRNAs) for CRISPR/Cas9-mediated mutagenesis of Hand-r (Supplementary file 1; [Gandhi et al., 2017]), and determined that Hand-r function is necessary for Tbx1/10 activation in the STVCs (Figure 4A). Co-expression of a CRISPR-resistant Hand-r cDNA (Hand-rPAMmis) rescued Tbx1/10 expression in the STVCs, indicating that Tbx1/10 down-regulation in this CRISPR ‘background’ is specifically due to Hand-r loss-of-function (Figure 4A). To further probe if Hand-r activity is necessary for FGF-MAPK-dependent Tbx1/10 expression, we used gain of M-Ras function in a Hand-r CRISPR ‘background’. Whereas, misexpression of the constitutively active M-RasG22V caused ectopic Tbx1/10 expression, concomitant loss of Hand-r function diminished both endogenous and ectopic Tbx1/10 expression in the STVC and FHP, respectively (Figure 4A). Although, remaining ectopic activation could still be observed, possibly because M-RasG22V could boost Hand-r expression in heterozygous cells where CRISPR/Cas9 disrupted only one copy of the gene. This data indicate that Hand-r function is necessary for FGF-MAPK-induced activation of Tbx1/10.
Figure 4 with 1 supplement see all
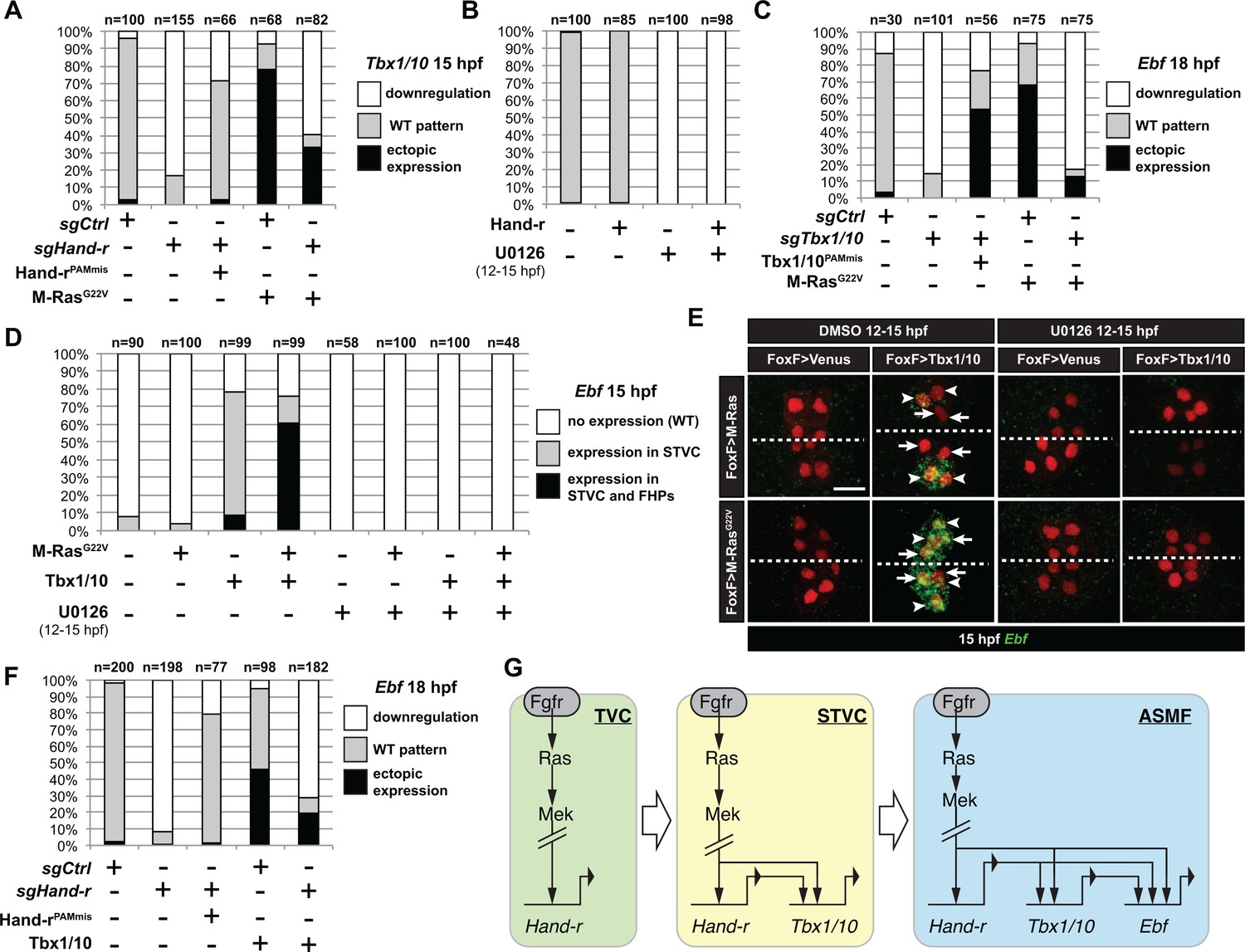
M-Ras/MAPK-driven feed-forward subcircuits control the successive activations of Hand-r, Tbx1/10 and Ebf.
(A) Proportions of embryo halves with indicated Tbx1/10 expression patterns following TVC-specific CRISPR/Cas9-mediated mutagenesis of Neurogenin/Neurog as a control (sgCtrl), and Hand-r (sgHand-r). TVC-specific overexpression of a CRISPR/Cas9-resistant form of Hand-r with mutation in the PAM sequence (Hand-rPAMmis) rescued Tbx1/10 expression in the sgHand-r ‘background’. TVC-specific overexpression of a constitutively active M-Ras mutant (M-RasG22) (control: M-RasWT) was sufficient to induce ectopic expression of Tbx1/10 in the FHPs in sgCtrl embryos but not in sgHand-r embryos indicating that Hand-r is necessary for M-Ras-dependent activation of Tbx1/10 transcription. (B) Proportions of embryo halves with indicated Tbx1/10 expression patterns following TVC-specific overexpression of Hand-r or a neutral reporter (Venus) and treated from 12 to 15hpf with the MEK inhibitor U0126 (+) or with DMSO (-) as control. Hand-r overexpression is not sufficient to rescue loss of Tbx1/10 expression due to MAPK inhibition indicating that M-Ras/MAPK activity is required in parallel of Hand-r expression to activate Tbx1/10 transcription in the TVC progeny. (C) Tbx1/10 is necessary downstream of M-Ras/MAPK activity to activate Ebf transcription in the TVC progeny. Shown are proportions of Ebf expression phenotypes following TVC-specific CRISPR/Cas9-mediated loss of Tbx1/10 function (sgTbx1/10), with Neurog-targeting sgRNA as control (sgCtrl). Specificity of Tbx1/10 loss of function was validated through rescue of Ebf expression with TVC-specific overexpression of a CRISPR/Cas9 resistant form of Tbx1/10 (Tbx1/10PAMmis). Ectopic Ebf expression in SHPs in Tbx1/10PAMmis larvae is explained by precocious misexpression of Tbx1/10 in the TVC as described in Wang et al. (2013). TVC-specific overexpression of M-RasG22 (M-RasG22), with wild type M-Ras (M-RasWT) as control, was sufficient to induce ectopic expression of Ebf in the cardiac precursors in sgCtrl embryos but not in sgTbx1/10 embryos indicating that Tbx1/10 is necessary for M-Ras-dependent activation of Ebf transcription. (D, E) Proportions (D) and examples (E) of 15hpf larvae halves showing indicated Ebf expression phenotypes in sgCtrl and sgHand-r CRISPR/Cas9 conditions combined with TVC-specific overexpression of a neutral reporter (Venus), Hand-rPAMmis, or Tbx1/10, and with MEK inhibition by U0126 (+) or not (DMSO control (-)). Arrowhead: STVCs, Arrows: FHPs, dotted line: ventral midline (F) Loss of Hand-r function impaired the ability of Tbx1/10 to induce ectopic Ebf expression. For simplicity, ectopic expressions in half to all of the cardiac precursors were combined in the same phenotype category. ‘n=": number of individual halves documented per condition. (G) Summary model of the temporal deployment of FGF/MAPK-driven feed-forward sub-circuits leading to the sequential activations of Tbx1/10 and Ebf in the STVCs and ASMFs, respectively.
To further probe the epistatic relationships between Hand-r and MAPK signaling upstream of Tbx1/10, we attempted to rescue Tbx1/10 expression in U0126-treated embryos, by over-expressing Hand-r with the TVC-specific Foxf enhancer. Neither did Hand-r over-expression cause ectopic Tbx1/10 activation (in the FHPs), nor was it sufficient to rescue Tbx1/10 expression in 15hpf STVCs (Figure 4B). Taken together, these data indicate that both Hand-r and MAPK activities are required to activate Tbx1/10 in the STVCs. These results also imply that MAPK signaling is restricted to the STVC independently of Hand-r activity, which suffice to explain the STVC-specific activation of Tbx1/10. Finally, we isolated a minimal STVC-specific enhancer from the Tbx1/10 locus and identified conserved putative Ets binding sites, which were necessary for reporter gene expression (Figure 4—figure supplement 1). This suggests that the FGF-MAPK-Ets pathway directly regulates Tbx1/10 expression in the STVCs.
Next, we investigated the epistatic relationship between FGF-MAPK, Hand-r, and Tbx1/10 upstream of Ebf in the ASMFs. We first used previously validated CRIPSR/Cas9 reagents targeting the Tbx1/10 coding region (Tolkin and Christiaen, 2016), to confirm that B7.5-lineage-specific loss of Tbx1/10 function inhibited Ebf activation, and verified that this effect could be rescued by over-expression of a CRISPR/Cas9-resistant Tbx1/10 cDNA, expressed with a minimal TVC-specific Foxf enhancer (Figure 4C; Tbx1/10PAMmis). In these rescue experiments, we observed ectopic Ebf activation in the SHP, as previously described when driving Tbx1/10 expression with a TVC-specific Foxf enhancer (Wang et al., 2013). As explained below, this ectopic activation could be attributed to a precocious expression of Ebf in the STVCs (Figure 4E). To test whether Tbx1/10 was also required for ectopic Ebf expression in response to MAPK activation (see Figure 2), we combined CRISPR/Cas9-mediated Tbx1/10 knockout with constitutive MAPK activation using the M-RasG22V mutant and observed a significant inhibition of both endogenous and ectopic Ebf expression in the 18hpf ASMF and SHP, respectively (Figure 4C). Taken together, these results show that Tbx1/10 function is necessary for FGF-MAPK-induced expression of Ebf in the ASMFs.
To further test whether Tbx1/10 acts in parallel and/or downstream of MAPK to activate Ebf, we combined gain of Tbx1/10 function with perturbations of FGF-MAPK signaling and assayed Ebf expression. We realized that Foxf-driven misexpression of Tbx1/10 caused precocious Ebf activation in 15hpf STVCs (Figure 4D,E). This precocious expression remained remarkably patterned, suggesting that STVC-restricted FGF-MAPK activity prevented Ebf expression in the dpERK-negative, small median FHPs (Figures 1B and 4E, Figure 1—figure supplement 1). Indeed, co-expression of Tbx1/10 and M-RasG22V caused both precocious and ectopic Ebf expression in the 15hpf medial and lateral TVC derivatives, which would be FHPs and STVCs in control embryos, respectively. This data confirms that Tbx1/10 misexpression does not suffice to cause ectopic Ebf expression in the FHPs, because the latter presumably lack FGF-MAPK activity, as is the case in control embryos.
U0126-mediated MEK inhibition from 12 to 15hpf, that is after the onset of Foxf>Tbx1/10 misexpression, further confirmed that MAPK activity is required in parallel to Tbx1/10 for precocious Ebf activation in 15hpf STVCs (Figure 4D,E). Taken together, these results indicate that Tbx1/10 and MAPK are both required to activate Ebf in the cell cycle following that of Tbx1/10 onset.
Since Hand-r expression is maintained in the ASMF, and CRISPR/Cas9- or RNAi-mediated Hand-r knockdown blocked both Tbx1/10 (Figure 4A) and Ebf expression (Razy-Krajka et al., 2014), we reasoned that Hand-r could also act both upstream and in parallel to Tbx1/10 for Ebf activation. To test this possibility, we assayed Ebf expression in 18hpf ASMF following defined perturbations of Hand-r and Tbx1/10. As expected, CRISPR/Cas9-mediated Hand-r mutagenesis strongly inhibited Ebf expression, and this effect could be rescued by a CRISPR-resistant Hand-r cDNA (Figure 4F). To test whether this effect was mediated by a loss of Tbx1/10 expression, we attempted to rescue the Hand-r loss-of-function by over-expressing Tbx1/10 using the Foxf enhancer. As explained above, Foxf-mediated Tbx1/10 misexpression caused precocious and ectopic Ebf expression in larvae co-electroporated with control sgRNAs (Figure 4D,E,F). By contrast, combining loss of Hand-r function with Tbx1/10 misexpression inhibited both the endogenous and ectopic Ebf expression (Figure 4F), indicating that Hand-r is also required in parallel to Tbx1/10 for Ebf activation in the ASMFs.
Taken together, these analyses suggest that coherent feed-forward circuits govern the sequential activation of Hand-r, Tbx1/10 and Ebf in response to continuous but progressively restricted FGF-MAPK inputs (Figure 4G), thus linking spatial patterning to the temporal deployment of the regulatory cascade leading to localized Ebf activation and pharyngeal muscle specification.
The cell cycle entrains the temporal deployment of the cardiopharyngeal gene regulatory network
In principle, the feed-forward circuit described above is sufficient to explain the successive activations of Hand-r, Tbx1/10 and Ebf. However, Tbx1/10 and Ebf do not turn on until after TVC and STVC divisions, respectively. Notably, even when we misexpressed Tbx1/10 in the TVCs, Ebf was activated only after cell division and in the lateral-most cells, where FGF-MAPK signaling is normally maintained (Figures 1B and 4E). This sequence of events -divisions followed by gene activation- is paramount as it permits the birth of first and second heart precursors, whose fates are antagonized by Tbx1/10 and Ebf (Razy-Krajka et al., 2014; Stolfi et al., 2010; Wang et al., 2013). Therefore, we sought to investigate the role(s) of the cell cycle in controlling the timing of Tbx1/10 and Ebf activation.
We first evaluated the effects of cytochalasin B, a classic inhibitor of cytokinesis widely used to study cell fate specification in ascidians (Figure 5A; [Whittaker, 1973]). Treatments starting before TVC divisions (12 hpf) did not block Tbx1/10 or Ebf expression in embryos fixed after their normal onset at either 16 or 19hpf, respectively (Figure 5B,C). Similarly, treatment starting between the first and second asymmetric divisions (15hpf) did not block localized Ebf expression at 19hpf (Figure 5C). This indicates that Tbx1/10 and Ebf activations occur by default in the absence of cytokinesis, most likely because FGF-MAPK signaling persists throughout the shared cytoplasm. This data thus illustrates how the spatial restriction of FGF-MAPK signaling, following cell divisions, leads to the localized activations of Tbx1/10 and Ebf.
Figure 5 with 1 supplement see all
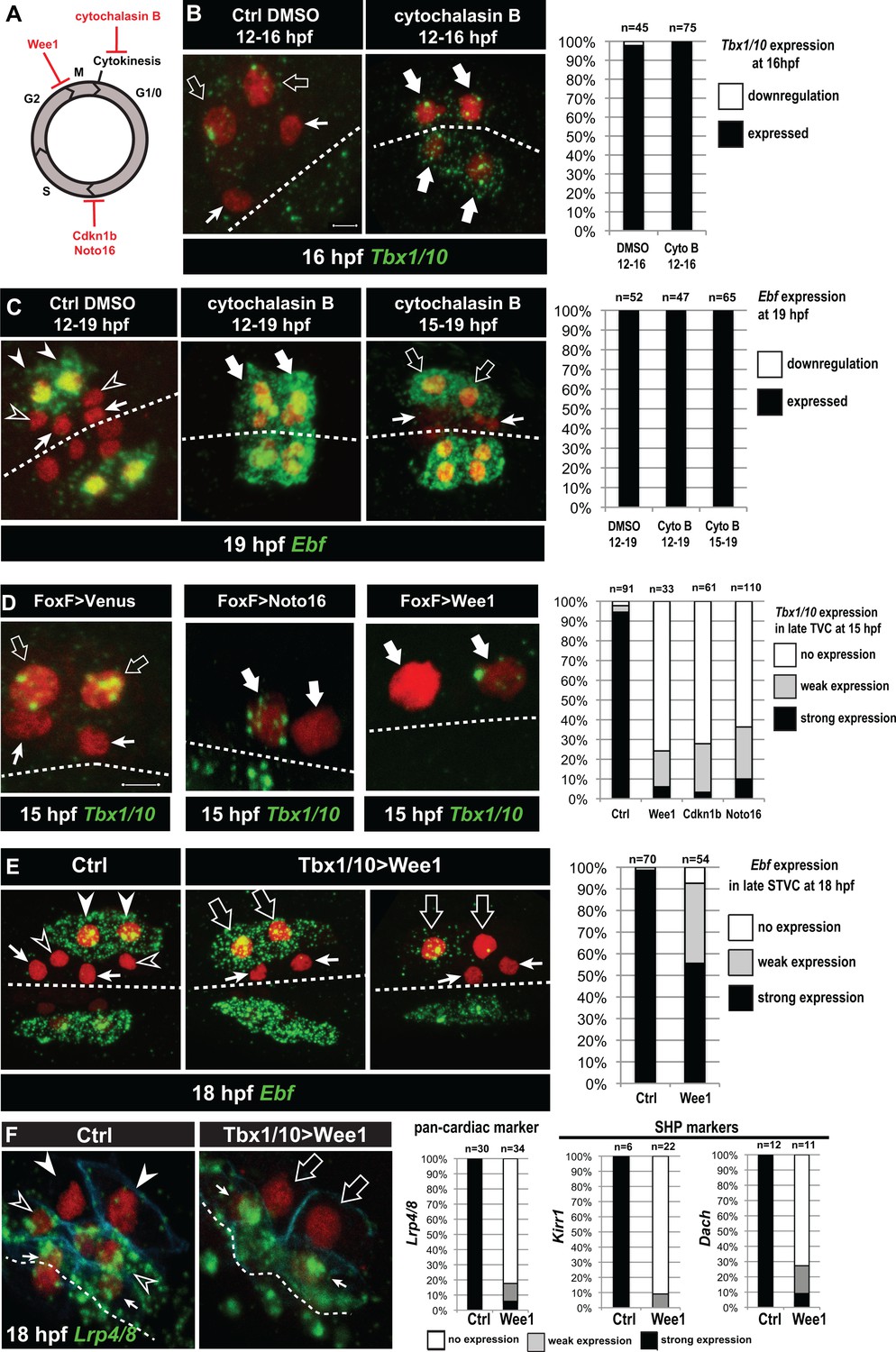
Temporal deployment of the cardiopharyngeal network is partially coupled with cell cycle progression.
(A) Schematic representation of the canonical eukaryotic cell cycle, and actions of the perturbations used in this study. (B,C) Tbx1/10 and Ebf expression at indicated time points, and following inhibition of cytokinesis by cytochalasin B treatment at indicated time points. Note that 15 to 19hpf treatment is applied AFTER the first division and birth the FHPs, which do not activate Ebf at 19hpf (right panel, arrows). (D) Inhibition of G1/S or G2/M blocks TVC division, and reduces Tbx1/10 expression. Pictures shows TVCs that have divided in controls but not in experimental cells, with one cell occasionnally turning on Tbx1/10, but not the other. Left: the proportions of embryos showing strong Tbx1/10 expression is substantially reduced compared to control embryos (e.g. Figure 1, and [Wang et al., 2013]). (E) Inhibition of G2/M in the STVCs by misexpression of Wee1 using the Tbx1/10 T6 enhancer inhibits STVC division, and has a mild impact on Ebf expression at 18hpf. Open arrows indicate STVCs that have not divided, but express high (middle) or low (right) levels of Ebf. Left: control larva showing high Ebf expression in the ASMF (closed arrowheads), but neither in the SHPs (open arrowheads) nor in the FHPs (Arrows). (F) Misexpression of Wee1 using the Tbx1/10 T6 enhancer (Tbx1 >Wee1) inhibits STVC division. Right: the proportions of embryos showing strong Lrp4/8, Kirr1 or Dach expression into late STVCs is reduced compared to SHPs in control embryos (T6 >NLS::LacZ) at 18 hpf. Notably, the pan-cardiac marker Lrp4/8 is still expressed in FHPs (arrows). Nuclei are marked in red with Mesp >NLS::lacZ, membranes in blue with Mesp >hCD4::mCherry, FHP labeled with arrows and SHP with arrowheads. 'n=', number of individual halves scored per condition. Scale bar, 5 µm. In all image panels, dotted line: ventral midline.
Cytochalasin treatments usually lead to the formation of polynucleated cells (e.g. Figure 5C, middle panel), because the cell cycle and nucleokinesis continue. To alter cell cycle progression more comprehensively, and specifically in the cardiopharyngeal lineage, we used genetically encoded inhibitors of cell cycle transitions: Cdkn1b.a and Cdkn1b.b (also known as Noto16), the ortholog of which is a potent inhibitor of the G1/S transition in the ascidian species Halocynthia roretzi (Kuwajima et al., 2014), and the G2/M inhibitor Wee1 (Dumollard et al., 2017). We used the TVC-specific Foxf enhancer to misexpress these negative regulators of cell cycle progression, monitored cell divisions and assayed Tbx1/10 expression at 15hpf, when control TVCs have divided and the lateral-most STVCs normally express Tbx1/10. Each perturbation efficiently inhibited TVC divisions, such that only two cells were visible on either side of the embryos (Figure 5D). In these delayed TVCs, Tbx1/10 expression was strongly reduced compared to control STVCs (Figure 5D). However, 20% to 40% of the delayed TVCs expressed Tbx1/10 to variable extents. This suggests that the cardiopharyngeal regulatory network can qualitatively unfold independently of cell cycle progression, but the latter is necessary for Tbx1/10 expression to its wild-type levels.
We next used the STVC-specific Tbx1/10 T6 enhancer (Figure 4—figure supplement 1), to misexpress Cdkn1b.a, Noto16 and Wee1, and assayed Ebf expression at later stages. Inhibitors of the G1/S transition failed to block STVC divisions (data not shown), most likely because Tbx1/10(T6)-driven products did not accumulate quickly enough to interfere with the G1/S transition in STVCs, since this cell cycle lasts only ~2 hr compared to ~6 hr for the TVC interphase. Therefore, we focused the analyses of Ebf response to cell cycle perturbations on misexpression of the G2/M inhibitor Wee1. Analyses of 18hpf larvae, fixed approximately 2 hr after the documented onset of Ebf expression in ASMFs (Razy-Krajka et al., 2014), indicated that Ebf can turn on in arrested STVCs that failed to divide upon Wee1 misexpression (Figure 5E).
Because ~30% of the embryos showed variable expression, as was the case for Tbx1/10 in the previous experiment, we reasoned that perturbations of the G2/M transition could alter the dynamics of Ebf upregulation. We investigated this possibility using embryos fixed every 30 min between 15.5hpf and 18hpf, when cells transition from a late Tbx1/10+; Ebf- STVC state to a committed Ebf+, Mrf+ ASMF state (Razy-Krajka et al., 2014; Wang et al., 2013). First, we scored the proportions of embryos with delayed STVCs or conspicuous ASMFs at each time point and showed that Wee1 misexpression strongly delays cell cycle progression, blocking cell divisions in a substantial fraction of embryos (Figure 5—figure supplement 1).
The proportion of Ebf+ ASMFs in control embryos progressively increased from ~20% at 15.5hpf to >90% by 18hpf, revealing an under-appreciated dynamic at the onset of Ebf expression, which appears to take at least one hour to be ‘strongly’ expressed in >75% of newborn ASMFs (Figure 5—figure supplement 1).
To evaluate the impact of Wee1-induced mitosis inhibition on Ebf accumulation, we focused on undivided STVCs at each time point (hence the lower numbers in Figure 5—figure supplement 1A compare to Figure 5—figure supplement 1B). By 17hpf, Wee1-expressing delayed STVCs showed ‘strong’ Ebf expression in proportions as high as for control ASMFs. However, these proportions were significantly lower at 16 and 16.5hpf (Chi-square tests, p=0.002 and p=0.0003, respectively), with ~1.5 and ~1.2 times less ‘strongly’ expressing cells than in the control distributions (hypergeometric tests, p=0.0005 and p=0.0001, respectively). These semi-quantitative analyses suggest that the cardiopharyngeal network can eventually unfold independently of cell divisions, leading to high levels of Ebf expression, albeit with a delay. This suggests that STVC division entrains Ebf upregulation in early ASMFs.
Finally, we reasoned that Wee1-expressed delayed STVC that activate Ebf would not turn on heart markers (Wang et al., 2017). Indeed, delayed STVCs failed to activate the pan-cardiac Lrp4/8, and the SHP-markers Kirr and Dach (Figure 5F; [Wang et al., 2017]). This indicated that coupling STVC division with localized FGF-MAPK activity and timed Ebf upregulation permits the localized activation of Ebf and the emergence of cardiac progenitors.
Transition from a MAPK-dependent to a MAPK-independent and autoregulative mode of Ebf expression in early ASMFs
We sought to further probe the mechanisms that regulate the initiation of Ebf expression in early ASMFs, and their biological significance for fate specification. Since we observed a progressive accumulation of Ebf mRNAs, and a transition from a MAPK-dependent onset to a MAPK-independent maintenance of Ebf transcription (Figure 3I,J), we reasoned that the window of MAPK-dependence might coincide with the accumulation of Ebf mRNAs between 16 and 17hpf. To test this possibility, we treated embryos with the MEK inhibitor U0126 at successive time points, assayed ongoing transcription using intronic probes and counted the numbers of Ebf transcribing cells (Figure 6A). This analysis revealed that Ebf transcription gradually lost its sensitivity to MAPK inhibition between 16 and 17hpf, that is during the first hour of the ASMF cycle when Ebf mRNAs normally accumulate (Figure 5—figure supplement 1A,B).
Because Ebf transcription becomes independent from MAPK by the time Ebf mRNA have accumulated to ‘high’ levels, and because Ebf expression lasts for several days in the progeny of the ASMFs, we reasoned that autoregulation might suffice to maintain high levels of Ebf mRNA past the MAPK-dependent onset. To test this possibility, we misexpressed the Ebf coding sequence using the STVC-specific T6 enhancer as described (Tolkin and Christiaen, 2016). Assaying endogenous Ebf transcription with intronic probes demonstrated that, in addition to its normal expression in the ASMFs, Ebf misexpression caused precocious and ectopic activation of the endogenous locus in the STVCs, and in the MAPK-negative SHPs, respectively (Figure 6C–F). This result suggests that Ebf transcription bypasses both requirements for cell-division coupling and MAPK inputs if high levels of Ebf gene products are present in the cell.
We reasoned that, if high levels of Ebf expression can promote its own transcription independently of MAPK signaling, then Ebf misexpression should be sufficient to rescue a chemical inhibition of MAPK at a critical stage. We tested this possibility by combining Ebf misexpression using the STVC-specific T6 enhancer and U0126 treatments starting at 16hpf, which normally block Ebf expression (Figure 6A,D–F). We observed that transcription of the endogenous Ebf locus became independent of early MAPK activity upon misexpression of an Ebf cDNA, further supporting the notion that high levels of Ebf expression suffice to maintain Ebf transcription independently of MAPK activity.
A potentially important implication of this transient MAPK requirement is to render Ebf expression initially reversible. For instance, Ebf occasionally turns on precociously in the STVCs of a small proportion of embryos (Figure 5—figure supplement 1C). Given the powerful anti-cardiogenic effects of Ebf (Razy-Krajka et al., 2014; Stolfi et al., 2010), persistent Ebf expression would have dramatic consequences for SHP development (Wang et al., 2013). However, because MAPK activity is excluded from the SHPs, and the early phase of Ebf expression depends upon continuous MAPK activity, we surmise that Ebf cannot be maintained in the SHPs. For instance, when embryos from the same electroporated batch were fixed at the time of early U0126 treatment (i.e. 15.75 and 16.25hpf) and ~4 hr later, at 20hpf, and assayed for Ebf transcription using intronic probes, initially wild-type patterns of Ebf transcription could not be maintained (Figure 6—figure supplement 1A). This suggests that, although Ebf can be activated precociously in a MAPK-dependent manner, its expression shuts off in the SHPs upon MAPK inhibition following STVC division.
We further addressed the interplay between cell division, MAPK signaling and Ebf expression. We reasoned that, if cell divisions entrain Ebf accumulation and the transition to a MAPK-independent autoregulative mode, then delaying STVC divisions should extend the period of MAPK-dependent Ebf transcription. We tested this possibility by expressing Wee1 under the control of the STVC-specific T6 enhancer, and treated embryos with U0126 at 17hpf, which inhibited the maintenance of Ebf transcription in only 15% to 20% of the control embryos (Figure 6A, Figure 6—figure supplement 1B). The proportion of embryos showing U0126-sensitive Ebf transcription increased to almost 50% upon Tbx1/10>Wee1 expression (Figure 6—figure supplement 1B), which is consistent with our hypothesis that inhibiting the G2/M transition delays the accumulation of Ebf gene products, thus postponing the transition from a low level/MAPK-dependent to an high level/MAPK-independent and self-activating mode of Ebf regulation.
Figure 6 with 1 supplement see all
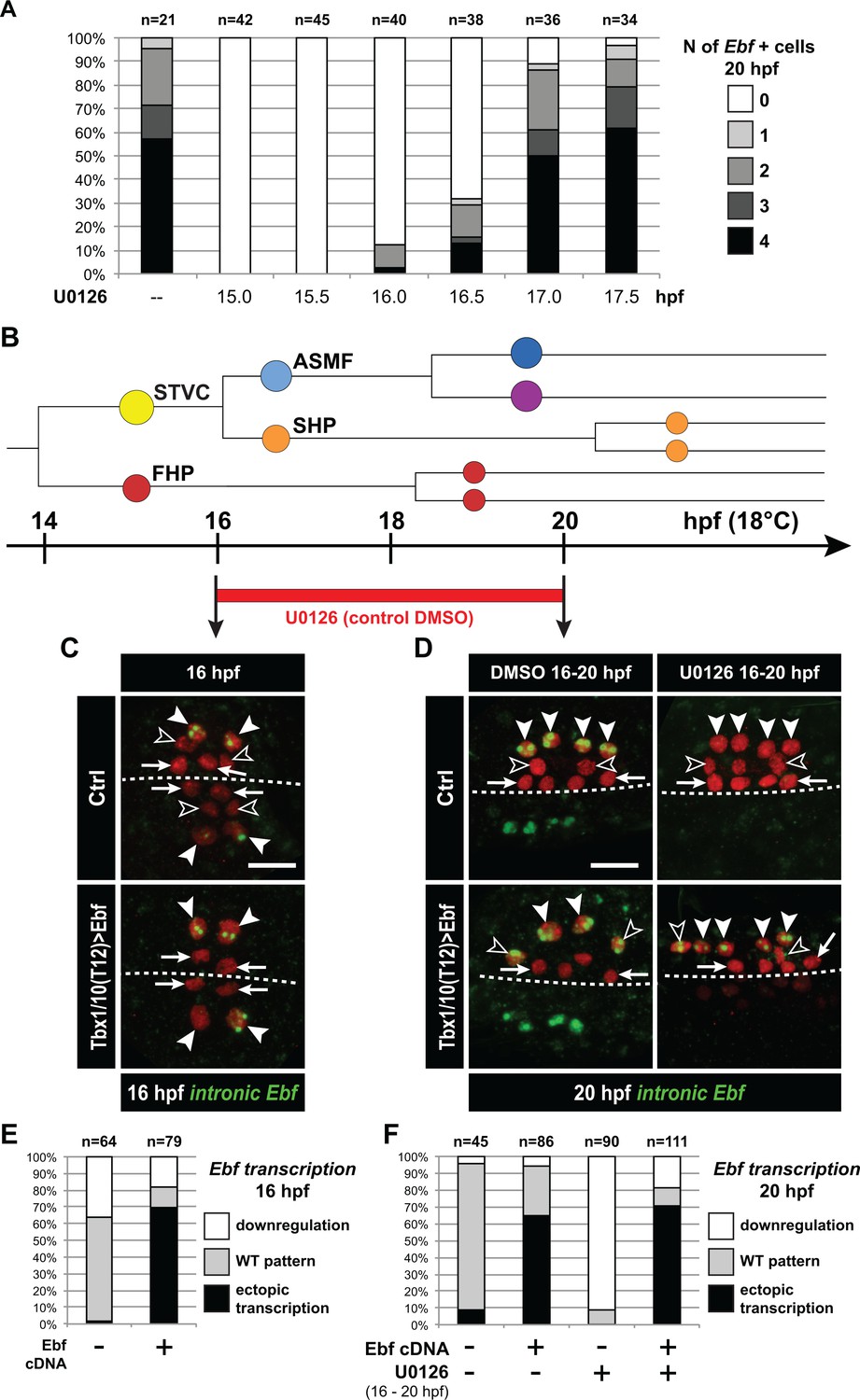
Ebf regulation transitions from MAPK-dependent to autoregulative during the early phase of ASMF cycle.
(A) Proportions of 20hpf larva halves showing the indicated number of Ebf-expressing cells following U0126 treatments started at the indicated time points. This indicates that, by 17hpf, Ebf expression, which started at ~16 hpf, has become largely insensitive to loss of MAPK activity. (B) Summary lineage diagram and time scale indicating the approximate stages for U0126 and DMSO (control) treatments for the results shown in (C, D). (C) Control (Ctrl) and Ebf-misexpressing embryos fixed at 16hpf, prior to chemical treatments, and stained for nascent transcripts with an intronic Ebf probe. In controls, the ASMFs (solid arrowhead), but neither the SHPs (open arrowheads) nor the FHPs (arrows), actively transcribe Ebf (green nuclear dots). In Larvae misexpressing the Ebf cDNA under the control of the STVC-specific Tbx1/10 enhancer, divisions are delayed and STVCs (solid arrowheads) activated transcription of endogenous Ebf loci (green nuclear dots). (D) After 4 hr, U0126 treated ASMFs no longer transcribe Ebf (top right image, solid arrowheads), whereas control DMSO-treated ASMFs do (top left, green nuclear dots). Upon misexpression of the Ebf cDNA in the STVCs and derivatives, ongoing Ebf transcription is detected at 20hpf in both DMSO and U0126-treated cells, and it persists in both ASMFs (solid arrowheads), and SHPs (open arrowheads). (E, F) Proportions of larvae halves showing the indicated Ebf transcription patterns, in indicated experimental conditions, as illustrated in C and D, respectively.
We propose a model for Ebf regulation whereby Hand-r, Tbx1/10, ongoing MAPK signaling and cell-cycle-regulated transcriptional input(s) govern the onset and initial accumulation of Ebf gene products during the first hour of the ASMF cycle, whereas the maintenance of Ebf expression relies primarily on MAPK-independent autoactivation, following initial accumulation (Figure 7).
Figure 7
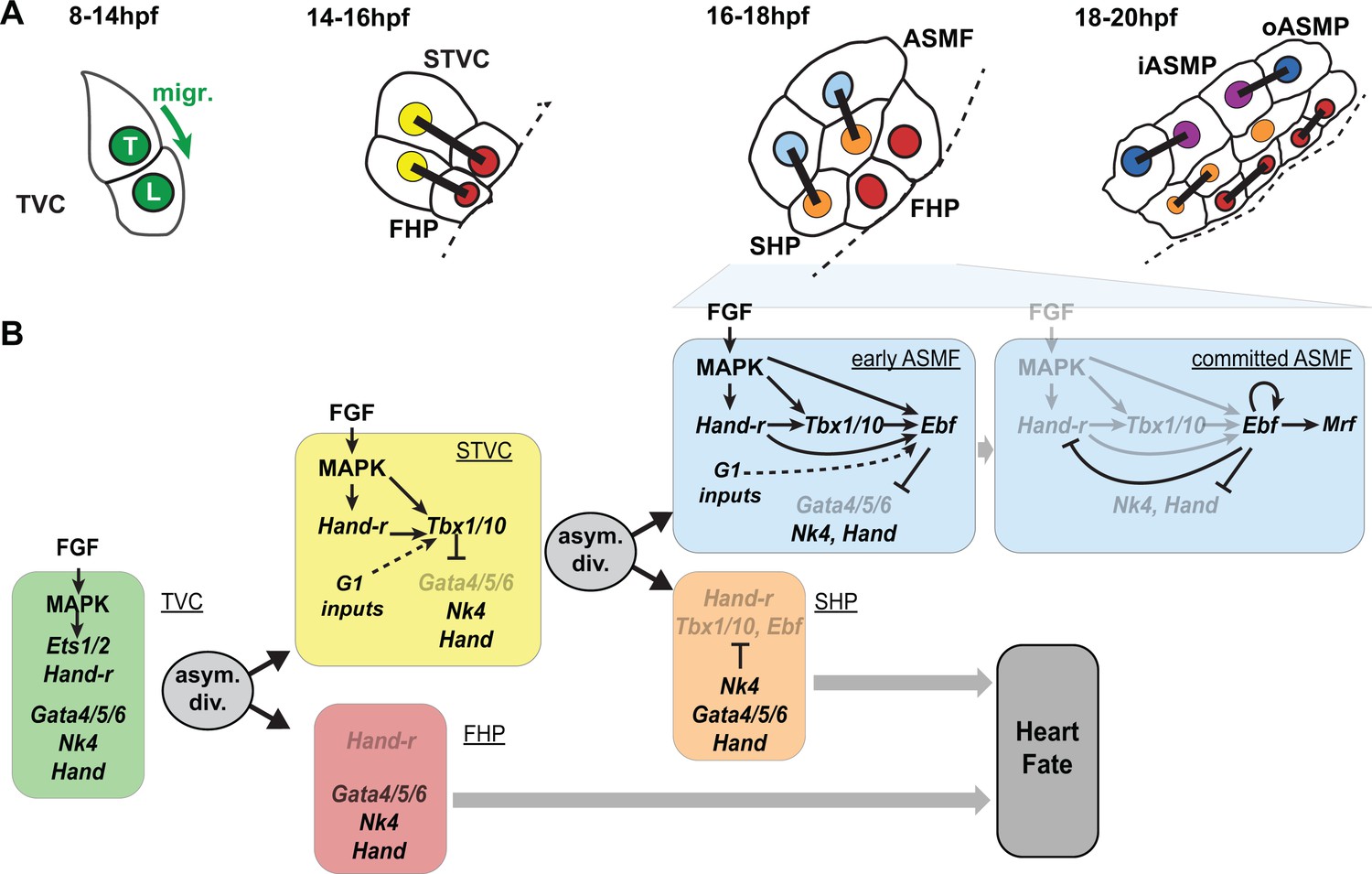
Summary model.
(A) Schematic representation of cardiopharyngeal lineage cells at successive time points representing the main fate transitions. hpf: hours post-fertilization; TVC: trunk ventral cells; L: Leader T: trailer; migr.: migration; STVC: second trunk ventral cells; FHP: first heart precursors; dotted line: midline; black bars link sister cells; ASMF: atrial siphon muscle founder cells; SHP: second heart precursors; iASMP: inner atrial siphon muscle precursors; oASMP: outer atrial siphon muscle precursor (these cells correspond to stem-cell-like Mrf-; Notch+ precursors and Mrf+; Notch- differentiating myoblasts, respectively; see (Razy-Krajka et al., 2014) for details). (B) Lineage diagram and documented regulatory relationships between indicated genes and pathways, as showing here and in (Razy-Krajka et al., 2014; Wang et al., 2013). In TVCs, primed heart and ASM markers are coexpressed, and maintenance of the STVC and ASM markers requires ongoing FGF/MAPK signaling. Following the first oriented and asymmetric cell division, FGF-MAPK is maintained only in the STVCs, which permits the continued expression of Hand-r and the activation of Tbx1/10. Cell division, presumably through G1-specific inputs, contributes to Tbx1/10 activation, and Tbx1/10 function antagonizes Gata4/5/6 expression (Wang et al., 2013). In the FHPs, termination of FGF-MAPK signaling inhibits Hand-r expression and prevents Tbx1/10 activation. Following oriented and asymmetric division of the STVCs, FGF/MAPK signaling persists only in the ASMFs, where it permits the transient maintenance of Hand-r and Tbx1/10, both of which act in parallel to FGF/MAPK to activate Ebf expression, together with contributions from presumed G1 inputs. Ebf activities further antagonize the cardiac program (marked by Gata4/5/6, Nk4/Nkx2.5 and Hand expression; [Razy-Krajka et al., 2014; Stolfi et al., 2010; Wang et al., 2013]). Once Ebf expression reaches ‘high levels’, its regulation becomes MAPK-independent and self-activating (this study). It also feeds back negatively on early activators such as Hand-r, and promotes the expression of the muscle determinant Mrf (Razy-Krajka et al., 2014; Tolkin and Christiaen, 2016). We propose that this transition represents commitment to an ASM fate. In the SHPs, termination of FGF/MAPK signaling prevents maintenance of Hand-r and Tbx1/10 expression, which, together with repressive inputs from Nk4/Nkx2.5, inhibits Ebf activation (Wang et al., 2013), and permits heart fate specification (Wang et al., 2017).
Discussion
Here, we demonstrated that the progressive restriction of FGF-MAPK signaling follows oriented and asymmetric cell divisions of multipotent progenitors and patterns the ascidian cardiopharyngeal mesoderm in space and time. Dynamic FGF-MAPK activity patterns lead to the localized expression of Hand-r, Tbx1/10 and Ebf in fate-restricted pharyngeal muscle precursors, and their concomitant exclusion for first and second heart precursors. We show that coherent feedforward circuits encode the successive activations of Hand-r, Tbx1/10 and Ebf, whereas cell divisions entrain the progression of this regulatory sequence and thus define the timing of gene expression. Finally, we provide evidence that the initiation of Ebf expression depends on MAPK activity in early ASMF, until Ebf accumulation permits MAPK-independent auto-activation. Given the potent anti-cardiogenic, and pro-pharyngeal muscle effects of Ebf (Razy-Krajka et al., 2014; Stolfi et al., 2010), we surmise that the latter switch corresponds to the transition from a cardiopharyngeal multipotent state to a committed pharyngeal muscle identity.
Spatial patterning by localized maintenance of FGF-MAPK signaling
Our results demonstrate that MAPK signaling is maintained only in the lateral-most daughter cells following each asymmetric division of multipotent cardiopharyngeal progenitors - the TVCs and STVCs. This asymmetric maintenance is necessary and sufficient for the progressive and localized deployment of the pharyngeal muscle network. Notably, the TVCs themselves are initially induced by similar polarized FGF-MAPK signaling coincidental to asymmetric cell divisions of their mother cells, the B8.9 and B8.10 founder cells (Davidson et al., 2006). At this stage, asymmetrical maintenance of sustained FGF-MAPK signaling involves intrinsic Cdc42-dependent polarity of the founder cells, which promotes polarized cell-matrix adhesion of the prospective TVC membrane to the ventral epidermis. The latter differential integrin-mediated adhesion promotes localized MAPK activation, leading to TVC induction (Cooley et al., 2011; Norton et al., 2013). It has been proposed that adhesion- and caveolin-dependent polarized FGFR recycling during mitosis accounts for the localized activation of MAPK in the prospective TVCs (Cota and Davidson, 2015). Whereas similar mechanisms could in principle account for asymmetric maintenance of FGF-MAPK signaling in STVCs and ASMFs, this has not been formally tested and there are notable differences opening the possibility that other mechanisms may be at work: during TVC induction, MAPK signaling is maintained in the smaller daughter cell that contacts the epidermis, whereas in the following divisions, MAPK activity persists in the larger daughter cells and all cells maintain contact with the epidermis (Kaplan et al., in preparation). Moreover, using fusion proteins as in previous studies, we could not observed a marked polarized distribution of FGFR molecules to the lateral-most cells (the STVCs and ASMFs; Kaplan et al., in preparation). However, the fact that constitutively active forms of M-Ras and Mek1/2 were sufficient to bypass the loss of MAPK activity, and impose pharyngeal muscle specification, indicates that differential FGF-MAPK activity is regulated upstream of M-Ras. Further work is needed to elucidate the cellular and molecular mechanisms governing the spatiotemporal patterns of FGF-MAPK signaling in the cardiopharyngeal mesoderm. In particular, it will be important to disentangle the relative impacts of extrinsic (i.e. tissues, contacts) vs. intrinsic (i.e. asymmetric cell division) effects onto FGF-MAPK signaling and the downstream transcriptional inputs.
Our preliminary analyses indicate that perturbations of the FGF-Ras-MEK pathway can alter cardiopharyngeal cell division patterns (Figure 2—figure supplement 1). While these effects did not account for the observed changes in gene expression, future studies will unravel FGF-MAPK-dependent and cardiopharyngeal-specific mechanisms governing the orientation and asymmetry of progenitor cell division (Kaplan et al., in preparation).
Transcriptional consequences of differential FGF-MAPK signaling
Differential FGF-MAPK signaling rapidly impacts cell-specific gene expression, we thus surmise that transcriptional effectors are dynamically regulated. Even though we have not formally identified the downstream DNA-binding transcription factor (see discussion below), it is conceivable that the phosphorylated forms of either transcriptional effector could persist through cell division upon maintenance of FGF-MAPK activity. However, we have shown that continuous MAPK activity is needed following each division. Therefore, we must invoke elusive phosphatase activities, such as dual-specificity phosphatases (DUSPs; [Patterson et al., 2009]), which would reset transcriptional effectors to a dephosphorylated state, thus rendering steady-state FGF-Ras-MAPK inputs necessary.
Systematic dephosphorylation of FGF-MAPK transcriptional effectors is particularly important for heart fate specification. As shown in our companion paper (Wang et al., 2017), whole genome analyses indicate that heart-specific de novo gene expression requires MAPK inhibition (Wang et al., 2017). The molecular mechanisms remain elusive, but one simple possibility is that, lest fate-restricted heart precursors inhibit MAPK activity, they will activate Tbx1/10 and Ebf, which will block the cardiac program (Razy-Krajka et al., 2014; Stolfi et al., 2010; Wang et al., 2013).
Finally, we previously proposed that repressor inputs from Nk4/Nkx2-5 are needed in the second heart precursors to avoid ectopic activation of Ebf (Wang et al., 2013). The observation that Nk4 transcripts are detected in all cardiopharyngeal cells opened the question as to how Ebf would escape repression by Nk4 in the ASMFs. Differential MAPK activity offers an intriguing possibility: for instance, Nk4/Nkx2.5-mediated repression in other species involves the co-repressor Groucho/TLE (Choi et al., 1999), which is strongly expressed in the cardiopharyngeal mesoderm (Razy-Krajka et al., 2014); and, in flies, MAPK-mediated phosphorylation of Groucho inhibits its repressor function (Cinnamon et al., 2008; Cinnamon and Paroush, 2008; Hasson et al., 2005). Therefore, it is possible that persistent MAPK signaling dampens Groucho/TLE-mediated repressive inputs on cell-specific regulatory genes like Ebf. Future studies will determine whether such mechanisms provide bistable switches underlying MAPK-dependent fate choices in the cardiopharyngeal mesoderm.
Temporal deployment of the pharyngeal muscle network
The localized and successive activation of Tbx1/10 and Ebf in STVCs, and ASMFs, respectively, are important features of the cardiopharyngeal network that permit the emergence of diverse cell fates: first and second heart precursors, and atrial siphon muscle precursors. Experimental misexpression of Ebf throughout the cardiopharyngeal mesoderm suffice to inhibit heart development (Razy-Krajka et al., 2014; Stolfi et al., 2010), illustrating the importance of Ebf restriction to the ASMF for the emergence of first and second heart precursors.
Our analyses indicate that the sequential activations of Hand-r, Tbx1/10 and Ebf is encoded in the feed-forward structure of this sub-circuit, whereas the continuous requirement for MAPK inputs and their progressive exclusion from heart progenitors restrict the competence to activate Tbx1/10 and Ebf to the most lateral cells, after each division. Our model implies that each gene may directly respond to transcriptional inputs from MAPK signaling. We have not formally identified the transcription factors(s) that mediate the transcriptional response to FGF-MAPK signaling. However, multipotent cardiopharyngeal progenitors express Ets1/2 and Elk, two common transcriptional effectors of FGF/MAPK signaling in Ciona (Bertrand et al., 2003; Christiaen et al., 2008; Davidson et al., 2006; Gainous et al., 2015), Ets1/2 has been implicated in the initial FGF-MAPK-dependent induction of multipotent TVCs (Christiaen et al., 2008; Davidson et al., 2006), and its expression is also progressively restricted to the lateral-most progenitors following each division (Razy-Krajka et al., 2014). Taken together, Ets1/2 and, to some extend, Elk are intriguing candidate transcriptional effectors of FGF/MAPK signaling in cardiopharyngeal development.
The binding preferences of Ets-family factors have been extensively studied in Ciona, and they do not depart markedly from conserved Ets-family binding sites with a GGAW core (Bertrand et al., 2003; Farley et al., 2015, 2016; Gueroult-Bellone et al., 2017; Khoueiry et al., 2010). Putative Ets-family binding sites in the TVC-specific Hand-r enhancer are conserved between Ciona intestinalis and its sibling species C. robusta and C. savignyi, and necessary for its activity in reporter assays (Woznica et al., 2012). Similarly, minimal STVC and ASM enhancers for Tbx1/10 and Ebf, respectively, contain conserved putative Ets-family binding sites, which contribute to proper reporter gene expression in transfection assays (Figure 4—figure supplement 1; (Razy-Krajka et al., 2014; Wang et al., 2013) and data not shown). Taken together, these observations suggest that the proposed feed-forward sub-circuit involves direct transcriptional inputs from FGF-MAPK-regulated Ets-family factors on the cardiopharyngeal enhancers of Hand-r, Tbx1/10 and Ebf.
Whereas the regulatory architecture of the MAPK; Hand-r; Tbx1/10; Ebf sub-circuit explains the sequence of activation events, it is also crucial for its correct deployment, and the generation of diverse cell identities, that genes are not fully activated before successive cell divisions. While divisions are not absolutely required for Ebf to eventually turn on, cell cycle progression appears to entrain the deployment of this network, especially for Tbx1/10 and Ebf activation in STVCs and ASMFs, respectively. These observations imply that the intrinsic dynamic of the network is slower than observed. This allows first and second heart precursors to be born prior to the onset of Tbx1/10 and Ebf, respectively. The latter sequence is essential for the heart progenitors to escape the anti-cardiogenic effects of Tbx1/10 (Wang et al., 2013), and Ebf (Razy-Krajka et al., 2014).
Initial Ebf expression in early ASMFs is also labile and MAPK-dependent for approximately one hour. This continued requirement for MAPK inputs ensures that, in the rare instances when Ebf expression starts in the multipotent STVC progenitors and/or expands to the nascent SHPs, inhibition of MAPK shuts off Ebf expression before it reaches the levels needed for commitment to an ASM fate. Indeed, our results indicate that, once Ebf mRNAs have accumulated to high levels, its expression becomes auto-regulative and MAPK-independent. We surmise that this transition coincides with a fundamental switch from a multipotent cardiopharyngeal state to a committed pharyngeal muscle identity.
From this standpoint, the observed entrainment of Ebf expression by the cell cycle can be seen as acceleration of the transition to commitment following asymmetric division of multipotent progenitors. Although the mechanisms remain elusive, it is likely that this requires the M/G1 transition, as the G1 phase has been shown to be particularly conducive to the expression of fate-specific regulators in mammalian pluripotent stem cells (Dalton, 2015; Pauklin et al., 2016; Pauklin and Vallier, 2013; Soufi and Dalton, 2016).
Conserved dual effects of FGF-MAPK signaling on heart development in chordates
Previous studies highlighted how FGF-MAPK signaling is necessary along side Mesp during early cardiac development in Ciona (Christiaen et al., 2008; Davidson, 2007; Davidson et al., 2006). This early requirement also exists in vertebrates (Abu-Issa et al., 2002; Alsan and Schultheiss, 2002; Barron et al., 2000; Brand, 2003; Reifers et al., 2000; Zaffran and Frasch, 2002). We now know that these early FGF-MAPK inputs induce and maintain multipotent cardiopharyngeal states in Ciona, including the Tbx1/10+ multipotent progenitors that eventually produce the second heart lineage ([Razy-Krajka et al., 2014; Stolfi et al., 2010; Wang et al., 2013], [Wang et al., 2017] and this study). Similarly, in vertebrates, regulatory interplay between Fgf8 and Fgf10 signaling and Tbx1 is required for development of both pharyngeal arch and second heart field derivatives, presumably in part by maintaining an undifferentiated and proliferative state (Abu-Issa et al., 2002; Aggarwal et al., 2006; Brown et al., 2004; Chen et al., 2009; Hu et al., 2004; Ilagan et al., 2006; Kelly and Papaioannou, 2007; Park et al., 2006; Park et al., 2008; Vitelli et al., 2002b; Watanabe et al., 2010; Watanabe et al., 2012). In fish, FGF signaling is necessary for cardiomyocyte addition at the arterial pole, in a manner reminiscent of its role in second heart field development (de Pater et al., 2009; Lazic and Scott, 2011). Notably, FGF signaling acts in successive phases, and its inhibition is necessary for final myocardial specification and differentiation (Hutson et al., 2010; Marques et al., 2008; Tirosh-Finkel et al., 2010; van Wijk et al., 2009). Conversely, continued FGF signaling beyond the multipotent mesodermal progenitor stages was shown to promote smooth muscle and epicardial differential in the heart (Hutson et al., 2010; van Wijk et al., 2009), and also myoblast specification and/or skeletal muscle differentiation in the head, with the expression of FGF ligands being maintained in the pharyngeal arches (Bothe et al., 2011; Buckingham and Vincent, 2009; Michailovici et al., 2015; Michailovici et al., 2014; von Scheven et al., 2006). Taken together, these and our data suggest that FGF-MAPK signaling plays evolutionary conserved roles during chordate cardiopharyngeal development, by promoting the specification of successive mesodermal and Tbx1+ multipotent states, and a fate-restricted non-cardiac muscle identity, while MAPK inhibition is required for myocardial specification and differentiation in the first and second heart field, successively.
Materials and methods
Animals, electroporations, and chemical treatments
Request a detailed protocolGravid wild Ciona intestinalis type A, now called Ciona robusta (Pennati et al., 2015), were obtained from M-REP (Carlsbad, CA, USA), and kept under constant light to avoid spawning. Gametes from several animals were collected separately for in vitro cross-fertilization followed by dechorionation and electroporation as previously described (Christiaen et al., 2009a, 2009b). Different quantities of plasmids were electroporated depending on the constructs. Typically, 50 µg of DNA was electroporated for NLS::lacZ or plain mCherry driving constructs but only 15 µg for Mesp-1 >H2B::mCherry. For perturbation constructs, 70 µg were usually electroporated, except for Mesp > NLS::Cas9::NLS (30 µg) and pairs of U6 >sgRNA plasmids (25 µg each). U0126 (Cell Signaling Technology, Danvers, MA) was used at 5 µM in artificial seawater from a stock solution of 20 mM in DMSO. Cytochalasin B (Sigma, Saint Louis, MO) was used at ~3 µg/mL from a 10 mg/mL stock solution in DMSO as previously performed (Jeffery et al., 2008). Control embryos were incubated in parallel with corresponding concentrations of DMSO alone.
In situ hybridization
Request a detailed protocolIn situ hybridizations were carried out essentially as described previously (Christiaen et al., 2009c; Razy-Krajka et al., 2014), using DIG labeled riboprobes, anti-DIG-POD Fab fragments (Roche, Indianapolis, IN), and Tyramide Amplification Signal coupled to Fluorescein (Perkin Elmer, MA). Reporters expressed in the lineage of interest were marked using anti-ß-galactosidase monoclonal mouse antibody (1:1000; Promega, Fitchburg, WI) or anti-mCherry rabbit polyclonal antibody (1:500; BioVision 5993–100), respectively targeted with anti-mouse or anti-rabbit secondary antibody coupled with Alexa 648 (1:500; Invitrogen, Carlsbad, CA). The different probes used in this study were described previously (Razy-Krajka et al., 2014; Stolfi et al., 2010; Wang et al., 2013).
dpERK/mcherry double fluorescent immunostaining
Request a detailed protocolSamples were fixed, as for in situ hybridizations, in MEM-PFA with Tween 20 (0.05%) but only for 30 min at room temperature, washed three times in PBSt (Tween 20 0.01%) for 10 min, gradually dehydrated every 10 min in Ethanol/PBS series (33%, 50%, 80%) and Methanol 100%. Samples were then gradually rehydrated every 10 min in Methanol/PBSt series, rinsed three times in PBSt, permeabilized with PBS Triton-100 (0.2%) for 30 min and incubated for 2 hr at room temperature with anti-dpERK mouse monoclonal antibody (1:200; Sigma, Saint Louis, MO) and anti-mCherry polyclonal antibody from rabbit (1:500; Biovision, Milpitas, CA) in PBS 0.01% Triton-100 (T-Pbs) supplemented with 2% normal goat serum. Samples were then washed three times in T-PBS and incubated in anti-mouse and anti-rabbit antibodies (1 :500 each), respectively coupled with Alexa 488 and Alexa 568 (Invitrogen, Carlsbad, CA), overnight at 4°C or for 2 hr at room temperature. Finally, samples were rinsed three times in T-PBS for 15 min and mounted in Prolong Gold (Molecular Probes, Eugene, OR).
Molecular cloning
Request a detailed protocolCoding sequences for wild-type M-Ras (KH.L172.2), Mek1/2 (KH.L147.22), Cdkn1b.a (Cdkn1b, KH.C14.564), and Cdkn1b.b (Noto16, KH.S643.6) were PCR-amplified from cDNA libraries prepared by reverse transcription of total RNA from mixed developmental stages. Insertion of the products into expressing vectors was performed using regular restriction/ligation or In-fusion (Clontech, Mountain View, CA) procedure. Oligonucleotide directed mutagenesis or two-step overlap PCRs were used to generate the point mutated forms M-RasG22V and MekS220E,S216D from the corresponding wild-type sequences. We also used oligonucleotide directed mutagenesis to generate mismatches in the PAM sequences adjacent to the sgRNA targets for Hand-r (153C > T 574C > T for Hand-rPAMmis) and Tbx1/10 (325G > A and 579G > A for Tbx1/10PAMmis). Due to the absence of a correct PAM sequence (NGG, (reverse complement CCN)), overexpressed Hand-rPAMmis and Tbx1/10PAMmis are resistant to the Cas9 nuclease activity. We also used oligonucleotide directed mutagenesis to generate the mutations in two putative Ets binding sites from the corresponding wild-type sequence of the Tbx1/10 enhancer element: −3646TC >CT −3638GA >AG upstream from the initiation codon (E1 and E2 in Figure 4—figure supplement 1, respectively). Primer sequences are listed in Supplementary file 1.
CRISPR/Cas9-mediated loss of Hand-r function
Request a detailed protocolThe pair of single guide RNA (sgRNA) targeting Tbx1/10 (sgTbx1/10) has been validated previously (Tolkin and Christiaen, 2016). Rescue of the Tbx1/10 loss-of-function was achieved by TVC-specific overexpression of Tbx1/10PAMmis driven by a Foxf enhancer (Foxf-1 > Tbx1/10PAMmis). For Hand-r loss of function, sgRNAs were first designed to avoid genomic off-targets and tested as described (Gandhi et al., 2017). In short, sgRNA expressing cassettes (U6 > sgRNA) were assembled by single step overlap PCR. Individual PCR products (~25 µg) were electroporated with EF1a > NLS::Cas9::NLS (30 µg), Myod905 > Venus (50 µg), driving ubiquitous expression of Cas9 and a widely expressed fluorescent reporter construct, respectively. Efficient electroporation was confirmed by observation of fluorescence before genomic DNA extraction around 16 hpf (18°C) using QIAamp DNA Micro kit (Qiagen, German Town, MD). Mutagenesis efficacy of individual sgRNAs, as a linear function of Cas9-induced indel frequency, was estimated from electrophoregrams following Singer sequencing of the targeted regions amplified from extracted genomic DNA by PCR. Result of the relative quantification of the indel frequency (‘corrected peakshift’ of 22% and 24%) was considered high enough for both sgRNAs targeting Hand-r, which were finally selected. The corresponding cassettes were cloned into plasmid for repeated electroporations to study the loss of function of Hand-r. Rescue of Hand-r loss-of-function was achieved by overexpression of Hand-rPAMmis driven by a Foxf TVC specific enhancer (Foxf-1 >Hand rPAMmis). In order to control the specificity of the CRISPR/Cas9 system, sgRNAs targeting Neurogenin, a gene not expressed in the TVC and their progeny, was electroporated in parallel. Sequences of the DNA targets and oligonucleotides used for the sgRNAs are listed in Supplementary file 1.
Tbx1/10 enhancer and cis-regulatory analysis
Request a detailed protocolTo isolate the minimal STVC-specific element of Tbx1/10, we used conserved non coding sequences between Ciona robusta and Ciona savignyi as a guide for molecular dissection (Figure 4—figure supplement 1A, http://genome.lbl.gov/vista/index.shtml; [Frazer et al., 2004]). We cloned a full-length cis-regulatory DNA construct (~7 kbp) that was analyzed by introducing large deletions to map the functional elements. We found a fragment of 1264 bp, that we called T6, located at −4682 bp upstream from the initiation codon that was sufficient for STVC expression as well as in the mesenchyme and endoderm of the reporter gene (Figure 4—figure supplement 1A,B). 5’ deletions of the T6 element show that the main cis-regulatory elements required exclusively for STVC expression map in a 575 bp element, which we called T12, at −4116 bp upstream from the initiation codon sufficient (Figure 4—figure supplement 1A). The sequence of this element, which we called T12, reveals the presence of putative Ets1/2 binding sites (Figure 4—figure supplement 1C) that were predicted using JASPAR (Khan et al., 2018) and CisBP (Weirauch et al., 2014) databases. The minimal Tbx1/10 STVC-specific enhancer was further analyzed using point mutations of the candidate Ets1/2 sites with highest predicted scores (Figure 4—figure supplement 1D).
Observation and imaging
Request a detailed protocolSamples were usually scored under a DM2500 epifluorescent microscope (Leica Microsystems, Wetzlar, Germany). Imaging was performed using a TCS SP8 X inverted confocal microscope equipped with a white light laser, AOBS and HyD detectors (Leica Microsystems).
References
-
Fgf8 is required for pharyngeal arch and cardiovascular development in the mouseDevelopment 129:4613–4625.
-
Dissection of Tbx1 and Fgf interactions in mouse models of 22q11DS suggests functional redundancyHuman Molecular Genetics 15:3219–3228.https://doi.org/10.1093/hmg/ddl399
-
Regulation of avian cardiogenesis by Fgf8 signalingDevelopment 129:1935–1943.
-
Dynamic control of head mesoderm patterningDevelopment 138:2807–2821.https://doi.org/10.1242/dev.062737
-
Distinct and dynamic myogenic populations in the vertebrate embryoCurrent Opinion in Genetics & Development 19:444–453.https://doi.org/10.1016/j.gde.2009.08.001
-
Tbx1 regulates proliferation and differentiation of multipotent heart progenitorsCirculation Research 105:842–851.https://doi.org/10.1161/CIRCRESAHA.109.200295
-
Electroporation of transgenic DNAs in the sea squirt CionaCold Spring Harbor Protocols 2009:pdb prot5345.https://doi.org/10.1101/pdb.prot5345
-
Isolation of sea squirt (Ciona) gametes, fertilization, dechorionation, and developmentCold Spring Harbor Protocols 2009:pdb prot5344.https://doi.org/10.1101/pdb.prot5344
-
Whole-mount in situ hybridization on sea squirt (Ciona intestinalis) embryosCold Spring Harbor Protocols 2009:pdb prot5348.https://doi.org/10.1101/pdb.prot5348
-
Context-dependent regulation of Groucho/TLE-mediated repressionCurrent Opinion in Genetics & Development 18:435–440.https://doi.org/10.1016/j.gde.2008.07.010
-
Cytoskeletal polarity mediates localized induction of the heart progenitor lineageNature Cell Biology 13:952–957.https://doi.org/10.1038/ncb2291
-
Linking the cell cycle to cell fate decisionsTrends in Cell Biology 25:592–600.https://doi.org/10.1016/j.tcb.2015.07.007
-
Ciona intestinalis as a model for cardiac developmentSeminars in Cell & Developmental Biology 18:16–26.https://doi.org/10.1016/j.semcdb.2006.12.007
-
VISTA: computational tools for comparative genomicsNucleic Acids Research 32:W273–W279.https://doi.org/10.1093/nar/gkh458
-
A cranial mesoderm origin for esophagus striated musclesDevelopmental Cell 34:694–704.https://doi.org/10.1016/j.devcel.2015.07.003
-
Fgf8 is required for anterior heart field developmentDevelopment 133:2435–2445.https://doi.org/10.1242/dev.02408
-
Regulatory blueprint for a chordate embryoScience 312:1183–1187.https://doi.org/10.1126/science.1123404
-
Early embryonic expression of FGF4/6/9 gene and its role in the induction of mesenchyme and notochord in Ciona savignyi embryosDevelopment 129:1729–1738.
-
DiGeorge syndrome phenotype in mice mutant for the T-box gene, Tbx1Nature Genetics 27:286–291.https://doi.org/10.1038/85845
-
Regulation and evolution of cardiopharyngeal cell identity and behavior: insights from simple chordatesCurrent Opinion in Genetics & Development 32:119–128.https://doi.org/10.1016/j.gde.2015.02.008
-
The del22q11.2 candidate gene Tbx1 regulates branchiomeric myogenesisHuman Molecular Genetics 13:2829–2840.https://doi.org/10.1093/hmg/ddh304
-
Wnt signaling balances specification of the cardiac and pharyngeal muscle fieldsMechanisms of Development 143:32–41.https://doi.org/10.1016/j.mod.2017.01.003
-
Craniofacial Muscle DevelopmentCurrent Topics in Developmental Biology 115:3–30.https://doi.org/10.1016/bs.ctdb.2015.07.022
-
Chamber identity programs drive early functional partitioning of the heartNature Communications 6:8146.https://doi.org/10.1038/ncomms9146
-
Tbx1 is required for second heart field proliferation in zebrafishDevelopmental Dynamics 242:550–559.https://doi.org/10.1002/dvdy.23928
-
Dual-specificity phosphatases: critical regulators with diverse cellular targetsBiochemical Journal 418:475–489.https://doi.org/10.1042/BJ20082234
-
Programming embryonic stem cells to neuronal subtypesCurrent Opinion in Neurobiology 21:43–51.https://doi.org/10.1016/j.conb.2010.09.012
-
Induction and differentiation of the zebrafish heart requires fibroblast growth factor 8 (fgf8/acerebellar)Development 127:225–235.
-
The ascidian Mesp gene specifies heart precursor cellsDevelopment 131:2533–2541.https://doi.org/10.1242/dev.01145
-
Tissue-specific genome editing in Ciona embryos by CRISPR/Cas9Development 141:4115–4120.https://doi.org/10.1242/dev.114488
-
Neural tube patterning by Ephrin, FGF and Notch signaling relaysDevelopment 138:5429–5439.https://doi.org/10.1242/dev.072108
-
Pharyngeal mesoderm development during embryogenesis: implications for both heart and head myogenesisCardiovascular Research 91:196–202.https://doi.org/10.1093/cvr/cvr116
-
Antagonists of Wnt and BMP signaling promote the formation of vertebrate head muscleGenes & Development 17:3087–3099.https://doi.org/10.1101/gad.1154103
-
Wnt signals from the neural tube block ectopic cardiogenesisGenes & Development 15:255–260.https://doi.org/10.1101/gad.871501
-
A genetic link between Tbx1 and fibroblast growth factor signalingDevelopment 129:4605–4611.
-
Islet is a key determinant of ascidian palp morphogenesisDevelopment 141:3084–3092.https://doi.org/10.1242/dev.110684
-
Isl2b regulates anterior second heart field development in zebrafishScientific Reports 7:41043.https://doi.org/10.1038/srep41043
-
Role of TBX1 in human del22q11.2 syndromeThe Lancet 362:1366–1373.https://doi.org/10.1016/S0140-6736(03)14632-6
-
Early signals in cardiac developmentCirculation Research 91:457–469.https://doi.org/10.1161/01.RES.0000034152.74523.A8
Article and author information
Author details
Funding
National Heart, Lung, and Blood Institute (HL108643)
- Lionel Christiaen
Fondation Leducq (15CVD01)
- Lionel Christiaen
AFM-Téléthon (20895)
- Basile Gravez
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank Robert Kelly (Université Aix-Marseille, CNRS, France) for feedbacks on the manuscript. We are grateful to Alberto Stolfi for collaborative inputs throughout the project. We thank Farhana Salek and Kristyn Millan for technical support. This project was funded by NIH/NHLBI R01 award HL108643, and trans-Atlantic network of excellence award 15CVD01 from the Leducq Foundation to LC.
Copyright
© 2018, Razy-Krajka et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 1,713
- views
-
- 281
- downloads
-
- 42
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 42
- citations for umbrella DOI https://doi.org/10.7554/eLife.29656
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
An FGF-driven feed-forward circuit patterns the cardiopharyngeal mesoderm in space and time
eLife 7:e29656.
https://doi.org/10.7554/eLife.29656




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}