Reactive oxygen species regulate activity-dependent neuronal plasticity in Drosophila
- University of Cambridge, United Kingdom
- HHMI Janelia Research Campus, United States
- University of Manchester, United Kingdom
- University of York, United Kingdom
Figures
Figure 1
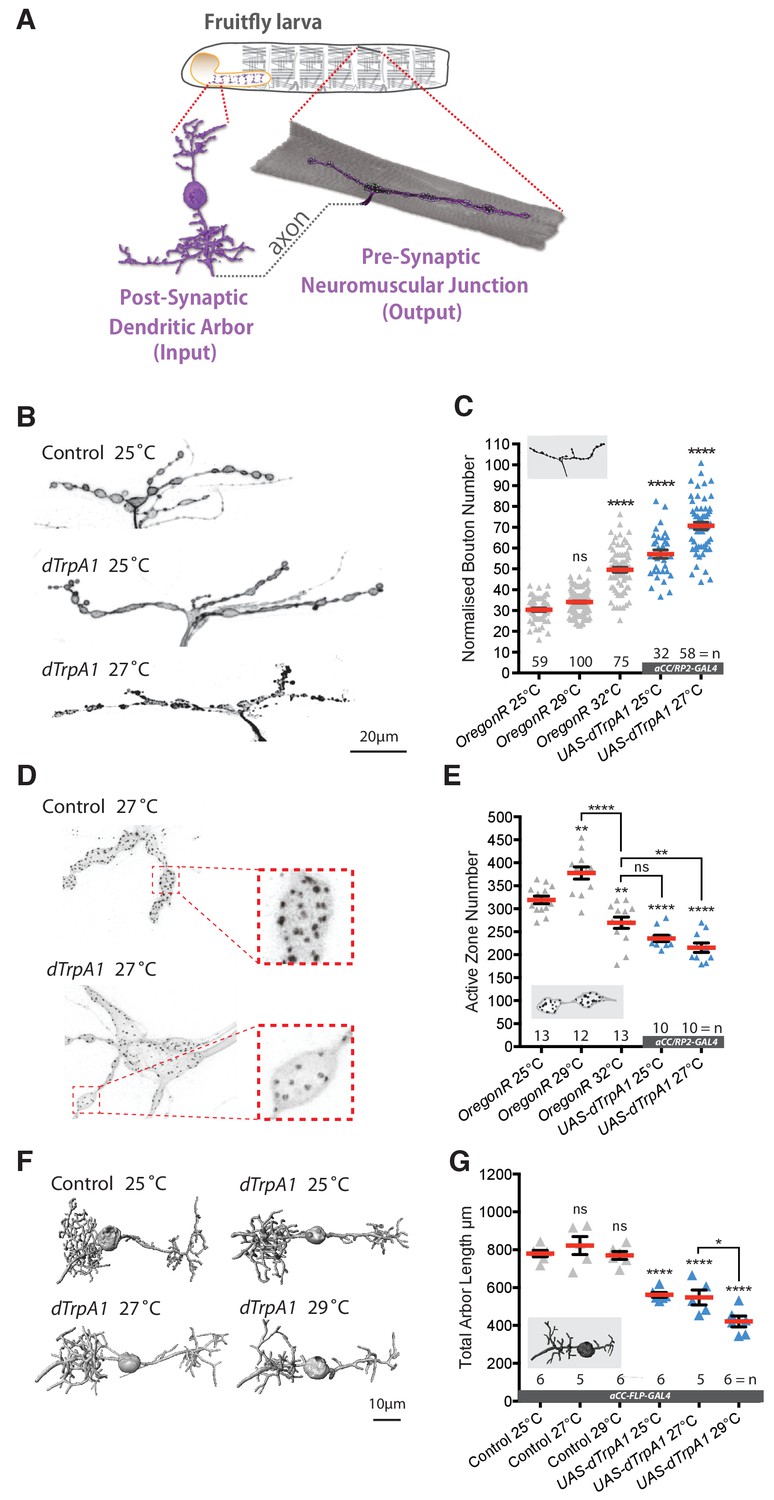
Adaptive structural synaptic plasticity at motoneuron input and output terminals in response to increased neuronal activity.
(A) Graphical illustration of a stereotypical larval motoneuron (MN) (adapted from Kohsaka et al., 2012). Pre-motor interneurons make synaptic connections with the MN dendritic arbor (input) in the larval ventral nerve cord (equivalent of mammalian spinal cord). The MN extends an axonal projection into the periphery where it connects with a target muscle via an NMJ, characterized by varicose swellings (boutons) each containing multiple individual neurotransmitter release sites (active zones). (B and C) Representative images of muscle DA1 [muscle one according to (Crossley, 1978)] NMJs from third instar larvae (100 hr ALH). Dot-plot quantification shows NMJ bouton number increases in response to systemic and cell-specific activity increases. (D and E) Active zone number increases following low-level overactivation (29°C), but progressively reduces upon stronger overactivation. (F and G) Digital reconstructions and dot plots show that overactivation leads to reduced total dendritic arbor length of aCC motoneurons (24 hr ALH). ‘aCC/RP2-GAL4’ expresses GAL4 in all, ‘aCC-FLP-GAL4’ in single aCC and RP2 motoneurons (see Methods for details); ‘Control’ in (B–G) is heterozygous aCC/RP2-GAL4 or aCC-FLP-GAL4, achieved by crossing the respective GAL4 line to Oregon-R wild type. Mean ± SEM, ANOVA, ns = not significant, *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, n = replicate number. Comparisons with control are directly above data points.
-
Figure 1—source data 1
Source data for Figure 1.
- https://doi.org/10.7554/eLife.39393.003
Figure 2
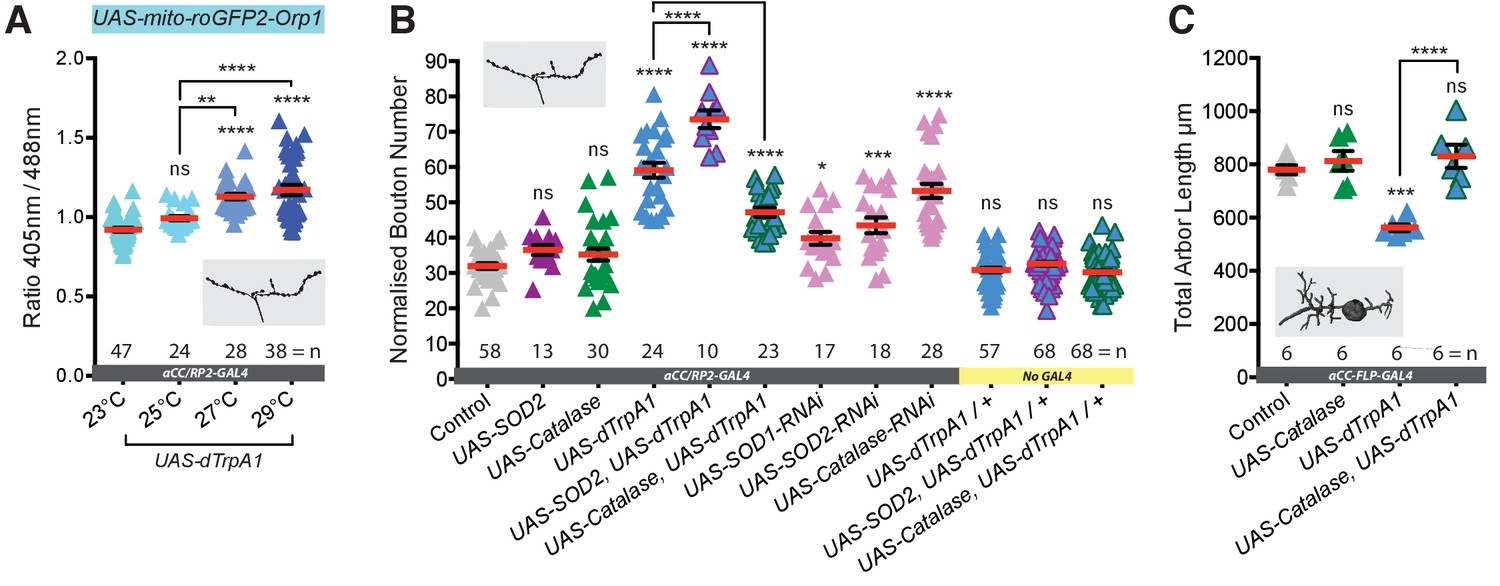
Neuronal activation leads to generation of synaptic ROS that regulate structural plasticity at input and output terminals.
(A) Elevated cell-specific activity increases mitochondrial ROS production at NMJs. Dot plots of mitochondrion-targeted ratiometric H2O2 sensor (UAS-mito-roGFP2-Orp1 [Gutscher et al., 2009]) in wandering third instar larval NMJs (100 hr ALH) at 23˚C (control, dTrpA1 inactive), 25˚C (moderate), 27˚C (strong) and 29˚C (very strong) dTrpA1-mediated overactivation. (B) Bouton number at the NMJ is increased by UAS-dTrpA1-mediated overactivation. This is exacerbated by co-expression of UAS-SOD2 (converts O2- to H2O2) and rescued by expression of the H2O2 scavenger UAS-Catalase. Cell-specific ROS elevation by scavenger knockdown is sufficient to induce NMJ elaboration. aCC/RP2-GAL4, ‘Control’ is aCC/RP2-GAL4 alone. (C) Total dendritic arbor length is reduced by single cell overactivation, but rescued by co-expression of the H2O2 scavenger UAS-Catalase (aCC motoneurons, 24 hr ALH). aCC-FLP-GAL4, ‘Control’ is aCC-FLP-GAL4 alone, heterozygous, achieved by crossing the GAL4 driver to Oregon-R wild type flies. Larvae were reared at 25°C unless indicated otherwise. Mean ± SEM, ANOVA, ns = not significant, *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001, n = replicate number..
-
Figure 2—source data 1
Source data for Figure 2.
- https://doi.org/10.7554/eLife.39393.005
Figure 3 with 3 supplements
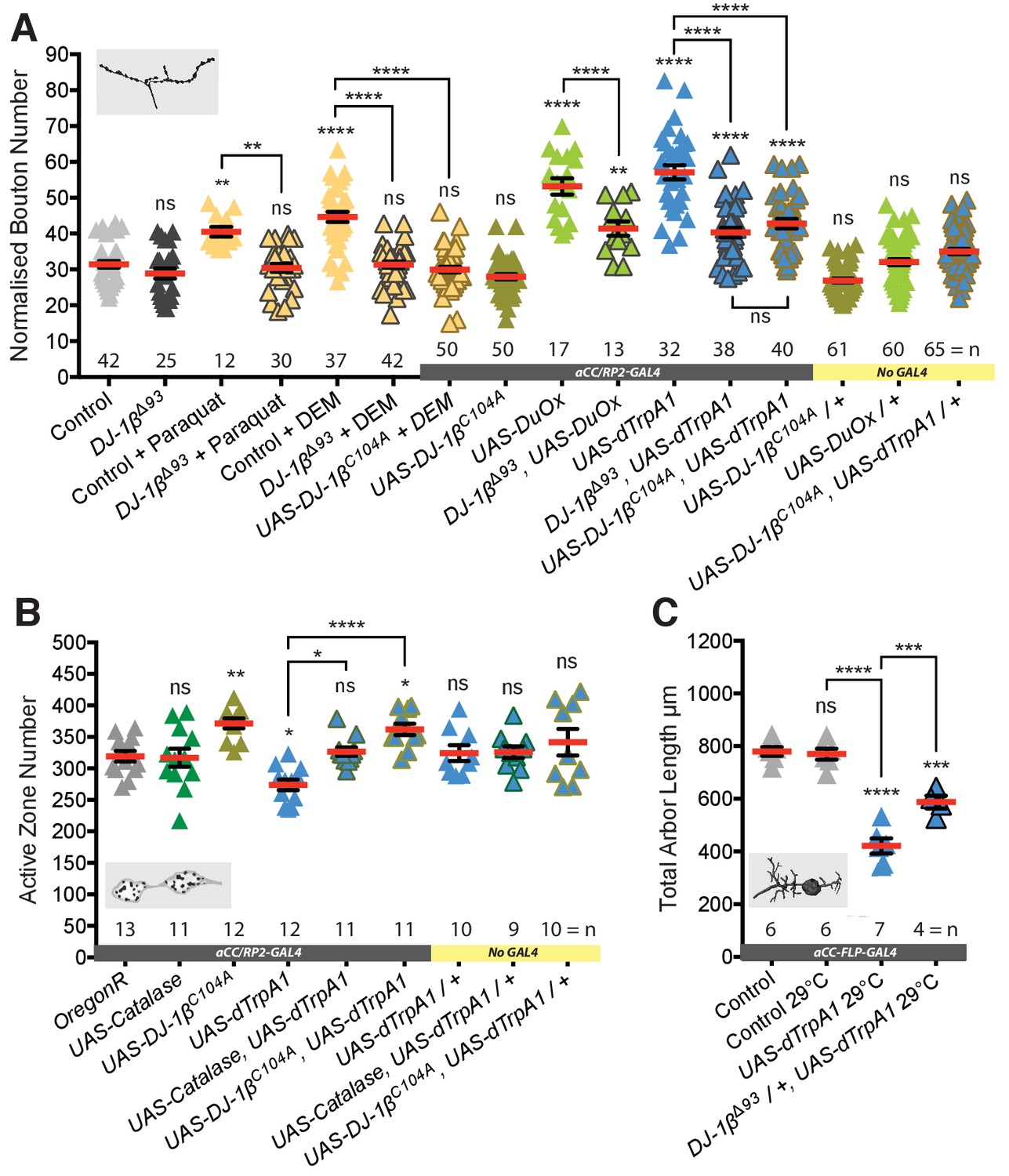
DJ-1β senses ROS and regulates activity-induced neural plasticity.
(A) DJ-1β is required for ROS and neuronal-activity-induced NMJ elaboration (100 hr ALH). Larvae reared at 25°C. (B) Cell-specific expression of DJ-1βC104A, non-oxidizable on conserved cysteine C104, prevents activity-induced reduction of active zone number. ‘Control’ is aCC/RP2-GAL4 alone. Larvae were reared at 25°C. (C) Activity-generated ROS sensing is dose sensitive. Removal of one copy of DJ-1β (in DJ-1βΔ93 /+ heterozygotes) is sufficient to significantly rescue activity-induced reduction of total dendritic arbor length of motoneurons in 24 hr ALH larvae. ‘Control’ is aCC-FLP-GAL4 alone, heterozygous, achieved by crossing the GAL4 driver to Oregon-R wild type flies.
-
Figure 3—source data 1
Source data for Figure 3.
- https://doi.org/10.7554/eLife.39393.010
Figure 3—figure supplement 1
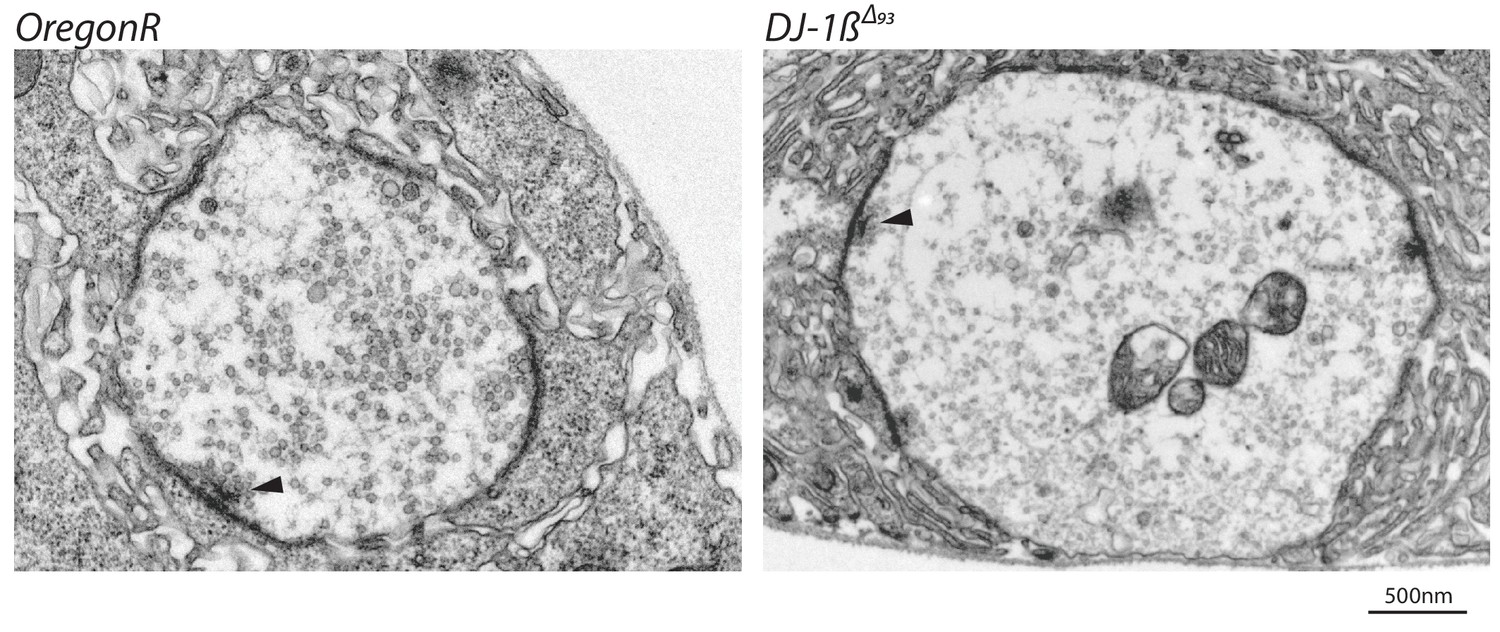
DJ-1βΔ93 mutant third instar larval NMJs are phenotypically normal.
Representative TEM bouton cross-sectional images showing that presynaptic architecture is intact including active-zones with associated clustered synaptic vesicles (arrowed).
Figure 3—figure supplement 2
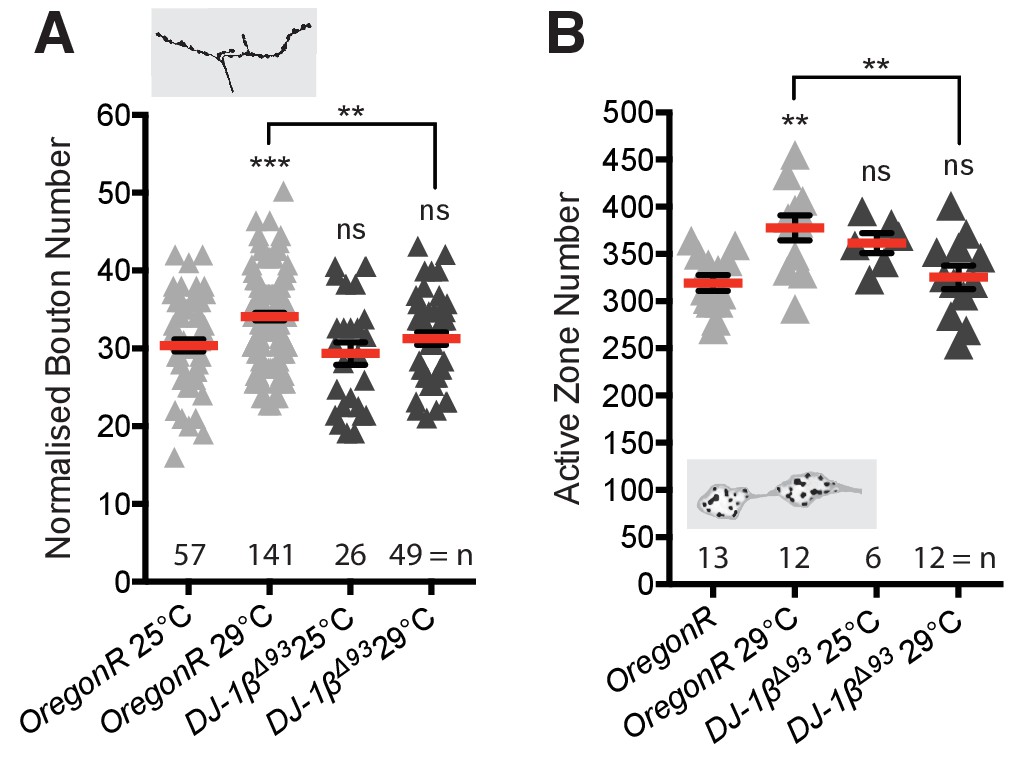
DJ-1βΔ93 mutant third instar larval NMJs fail to elaborate bouton number or active zone number upon rearing at 29ºC.
(A) DJ-1β null mutant (DJ-1βΔ93) larvae do not show systemic activity-dependent NMJ elaboration. Normalised bouton number dot plot showing significantly increased NMJ elaboration in OregonR, but not DJ-1βΔ93, larvae reared at 29°C vs 25°C. (B) DJ-1βΔ93 larvae also do not show systemic activity-dependent increase in active zone number. OregonR, but not DJ-1βΔ93, show significantly increased NMJ active zone number when reared at 29°C vs 25°C. Mean ± SEM, ANOVA **p<0.01, ***p<0.001, n = replicate number.
Figure 3—figure supplement 3
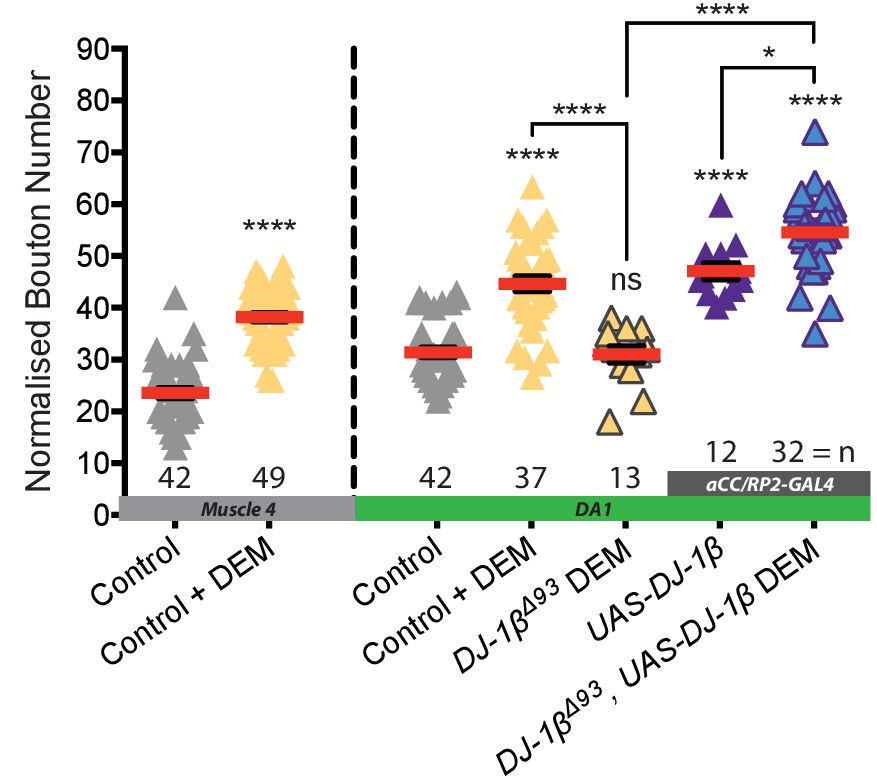
10 mM DEM feeding induces increased bouton number at both muscle four and DA1.
This response is absent in DJ-1β null mutant larvae and is re-established via neuronal (aCC and RP2) specific miss-expression of UAS-DJ-1β. UAS-DJ-1β expression alone elevates bouton number, which is exacerbated when combined with DEM feeding. Normalised bouton number dot plot, Control is aCC-FLP-GAL4 alone in heterozygous condition. Mean ± SEM, ANOVA *p<0.05, ****p<0.0001, n = replicate number.
Figure 4
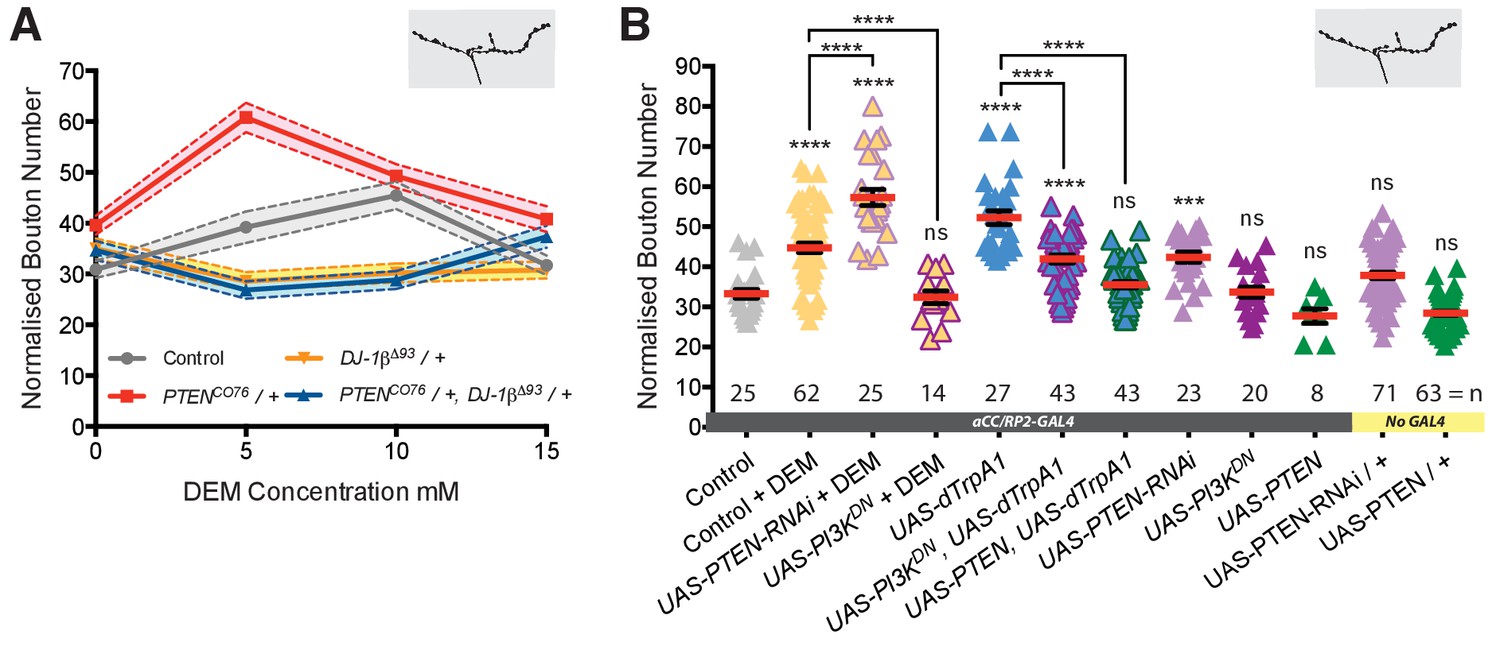
DJ-1β signals via PTEN and PI3Kinase to regulate ROS and activity-induced NMJ elaboration.
(A) DJ-1β and PTEN genetically interact to regulate systemic ROS-induced NMJ elaboration. NMJ bouton number varies with ROS (DEM) levels (grey data). Removal of one copy of PTEN sensitizes (red) while heterozygosity for DJ-1β desensitizes NMJs to ROS levels (yellow), partially restored in double heterozygotes (blue). Dashed boundaries indicate 95% confidence intervals (n ≥ 38). (B) Systemic ROS and activity-induced NMJ structural adjustments require PTEN and PI3Kinase signaling. Over-expression of the PI3Kinase antagonist PTEN or a dominant negative PI3Kinase form abrogates activity-induced NMJ elaboration. ‘Control’ is aCC/RP2-GAL4 alone, heterozygous, achieved by crossing the GAL4 driver to Oregon-R wild type flies. Mean ± SEM, ANOVA, ns = not significant, **p<0.01, ***p<0.001, ****p<0.0001, n = replicate number.
-
Figure 4—source data 1
Source data for Figure 4.
- https://doi.org/10.7554/eLife.39393.012
Figure 5
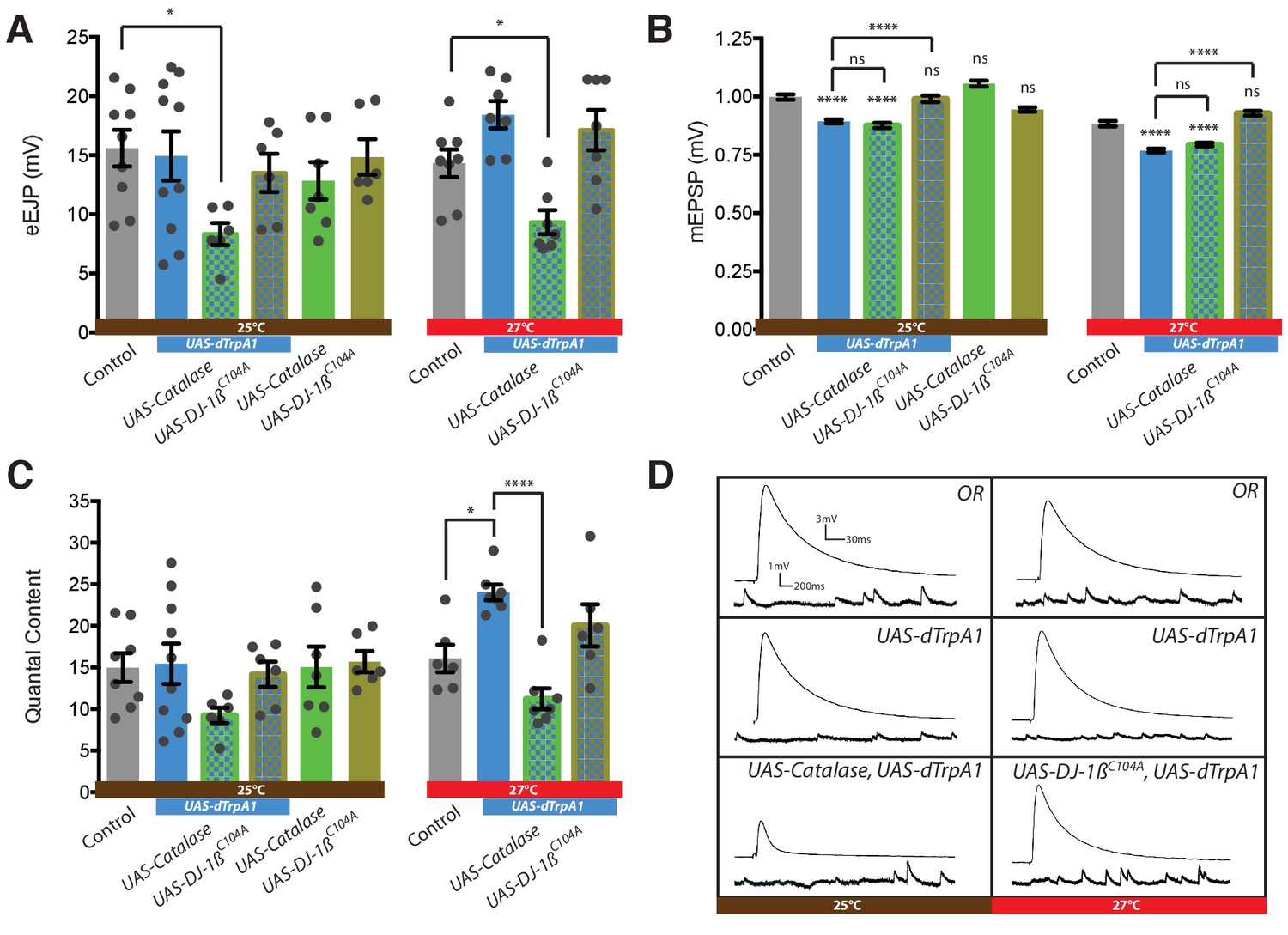
Presynaptic ROS regulate the maintenance of eEJP amplitude and DJ-1β function is required for the reduction of mEJP amplitude during overactivation.
(A) Overactivated motoneurons (expressing dTrpA1 at 25°C, 27°C) show similar eEJP amplitude to control, despite reduced mEJP amplitude (see B). Catalase co-expression prevents this adaptation, significantly reducing eEJP amplitude, whereas DJ-1β appears not to be required. (B) Increased neuronal activity, mediated by dTrpA1 expression at 25°C or by elevating ambient temperature to 27°C, reduces mEJP amplitude. Co-expression of DJ-1βC104A rescues this effect. (C) Quantal content (eEJP amplitude/mEJP amplitude) shows no significant difference between groups due to high variance within some conditions. (D) Representative eEJP and mEJP traces. ANOVA, ns = not significant, *p<0.05, ****p<0.0001.
-
Figure 5—source data 1
Source data for Figure 5.
- https://doi.org/10.7554/eLife.39393.014
Figure 6

Adaptive behavioral plasticity in response to chromic locomotor overactivation.
(A) Larval motor network activity, assayed by crawling speed 72 hr after larval hatching (AHL), increases in response to acute temperature upshifts (blue) in wild type larvae. In contrast, with prolonged exposure (grey) to elevated temperatures (29°C or 32°C) the motor network adapts homeostatically generating the same crawling speed as 25°C reared controls. This adaptation is further revealed by acute temperature downshifts (green). Each data point represents crawling speed from an individual uninterrupted continuous forward crawl, n = specimen replicate number, up to three crawls assayed for each larva. Genotype = OregonR. (B) Prolonged overactivation targeted to motoneurons (dVGlut-GAL4; UAS-dTrpA1) also leads to adaptation with reduced crawling speed (dTRPA1 channels open at 27°C, closed at 22°C). Mean ±SEM, control is dVGlut-GAL4 / +. ****p<0.0001 students t test, n = replicate number. (C) Larvae with expression of UAS-DJ-1βC104A targeted to motoneurons (dVGlut-GAL4) are unable to adapt motor network output (crawling speed) to elevated rearing temperatures. Control is dVGlut-GAL4 alone, in heterozygous condition. Each data point represents crawling speed from an individual uninterrupted continuous forward crawl, n = specimen replicate number, up to three crawls assayed for each larva. Mean ± SEM, ANOVA, ns = not significant, *p<0.05, **p<0.01, ***p<0.001, ****p<0.0001.
-
Figure 6—source data 1
Source data for Figure 6.
- https://doi.org/10.7554/eLife.39393.016
Figure 7
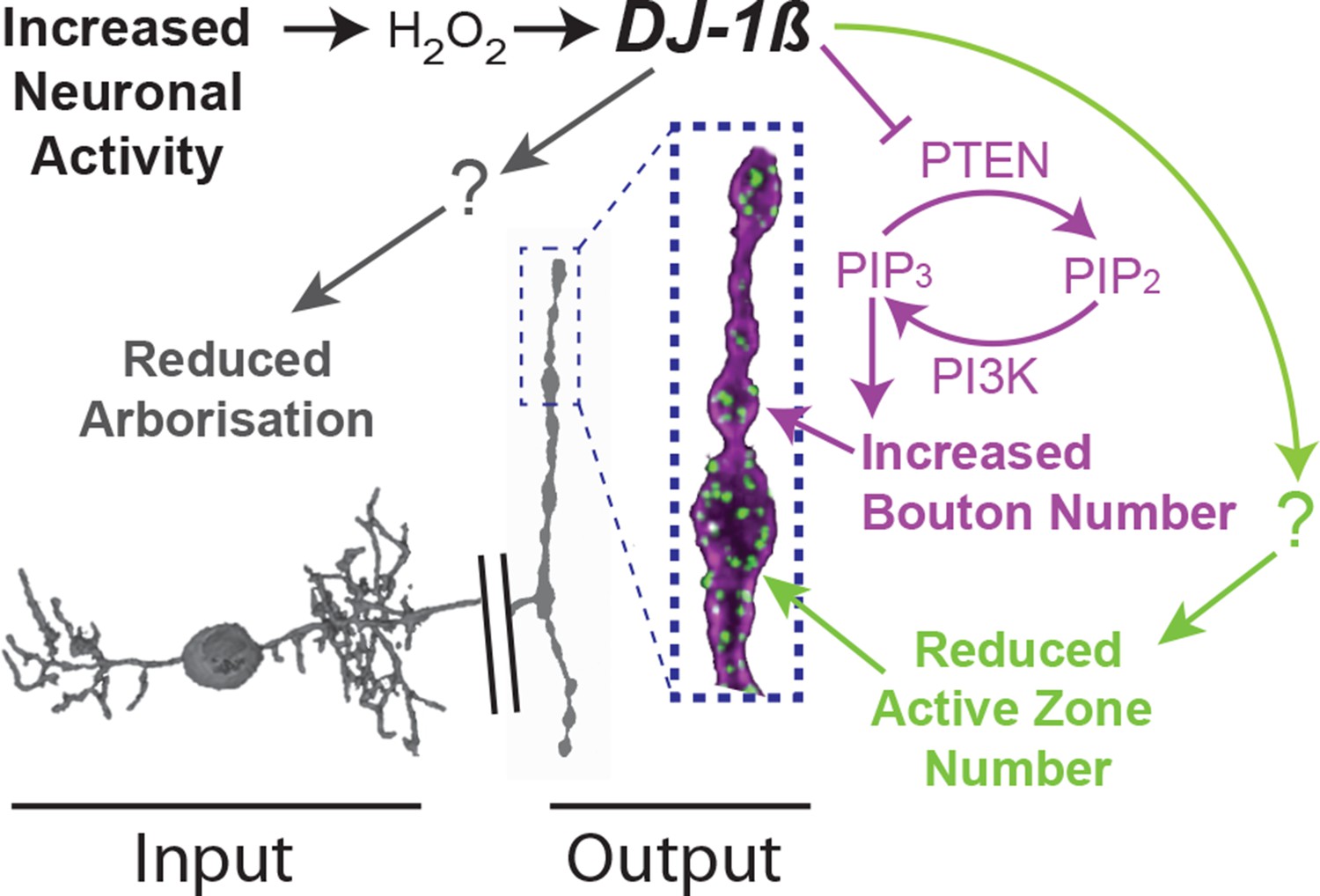
Model summary.
DJ-1β is a redox signaling hub that coordinates structural synaptic plasticity at motoneuron synaptic input and output terminals. Activity-induced ROS oxidize DJ-1β, leading to PTEN inhibition and thus to a gain in PI3Kinase signaling, which regulates activity-induced NMJ elaboration of boutons and active zones. At higher activity/ROS thresholds additional, yet to be defined, pathways downstream of DJ-1β are activated, implementing adaptive reductions of active zones at the NMJ and dendritic arbor length in the CNS.
Additional files
-
Transparent reporting form
- https://doi.org/10.7554/eLife.39393.018
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Reactive oxygen species regulate activity-dependent neuronal plasticity in Drosophila
eLife 7:e39393.
https://doi.org/10.7554/eLife.39393




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}