A switch in surface polymer biogenesis triggers growth-phase-dependent and antibiotic-induced bacteriolysis
- Harvard Medical School, United States
- University of Sheffield, United Kingdom
- Howard Hughes Medical Institute, United States
Figures
Figure 1 with 1 supplement
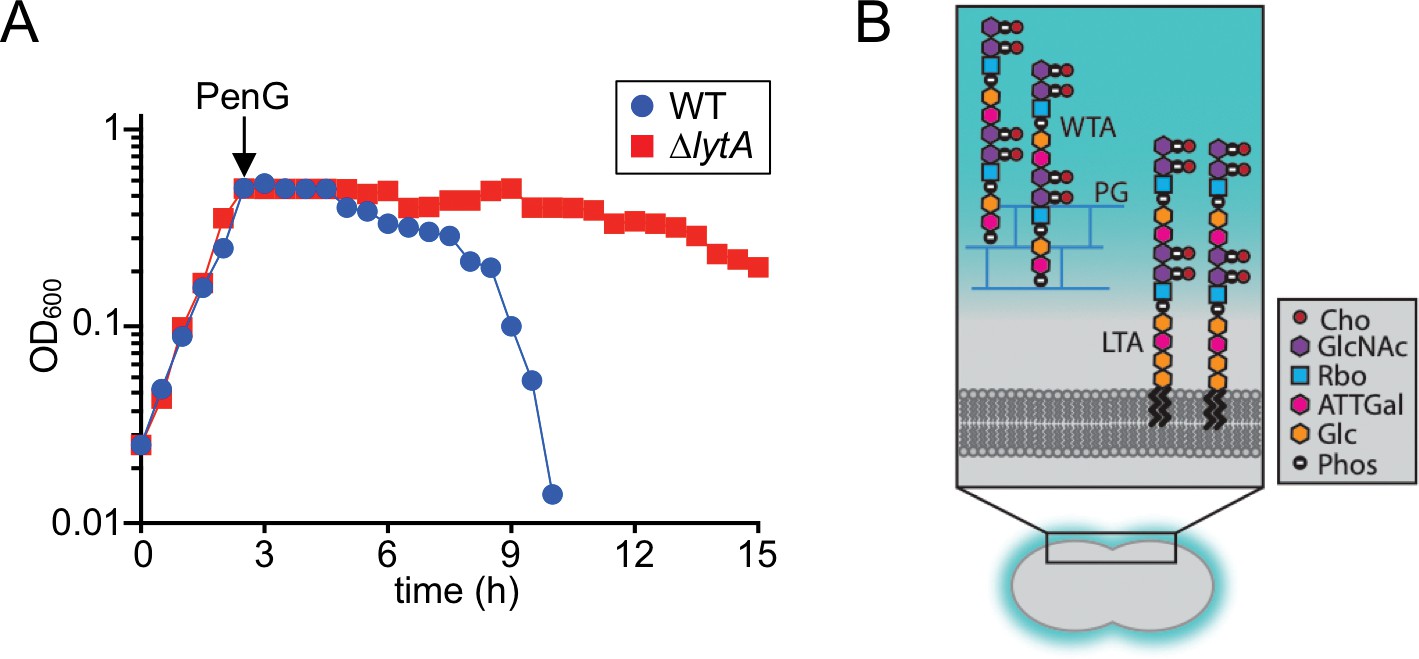
Beta-lactam induced lysis of Sp and overview of its cell envelope.
(A) The indicated strains were grown in THY at 37 ˚C in 5% CO2. At an OD600 of ~0.5, they were challenged with penicillin G (PenG) (0.5 μg/ml final). Growth was monitored every 30 min for 15 hr. (B) Schematic diagram of the cell envelope of Sp. The cell wall peptidoglycan (PG) (blue) contains Wall Teichoic Acid (WTA) polymers and the lipid bilayer contains Lipoteichoic Acid (LTA). The constituents of the repeating unit in LTAs and WTAs are indicated; Cho, choline; GlcNac, N-acetylglucosamine; Rbo, ribitol; ATTGal, 2-acetamido-4-amino-2,4,6-trideoxygalactose; Glc, glucose; Phos, phosphate. The following figure supplement is available for Figure 1.
Figure 1—figure supplement 1
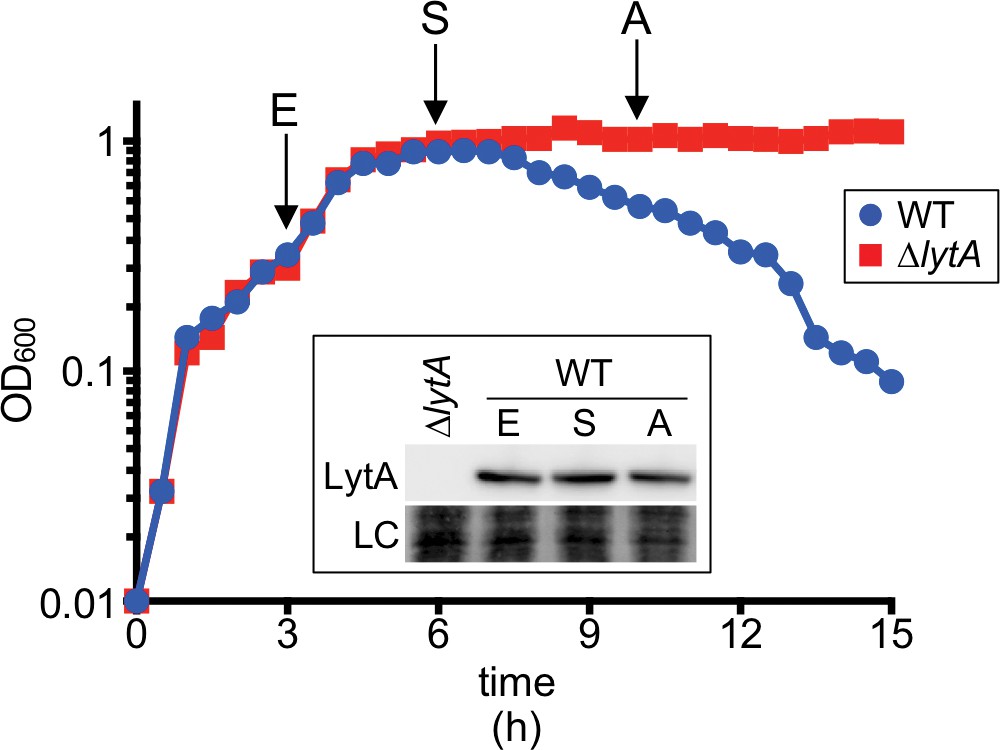
LytA levels remain constant before and at the onset of growth-phase-dependent autolysis.
Growth curves of the indicated strains at 37°C in 5% CO2. Measurements of culture OD600 were recorded approximately every 30 min for 15 hr. Samples were collected at the indicates time points (E, exponential; S, stationary; A, autolysis), normalized to an OD600 of 0.5 and analyzed by SDS-PAGE and immunoblotting using an anti-LytA antibody (LytA). LC (loading control) indicates a Ponceau S stain of the nitrocellulose membrane used for immunoblot analysis.
Figure 2
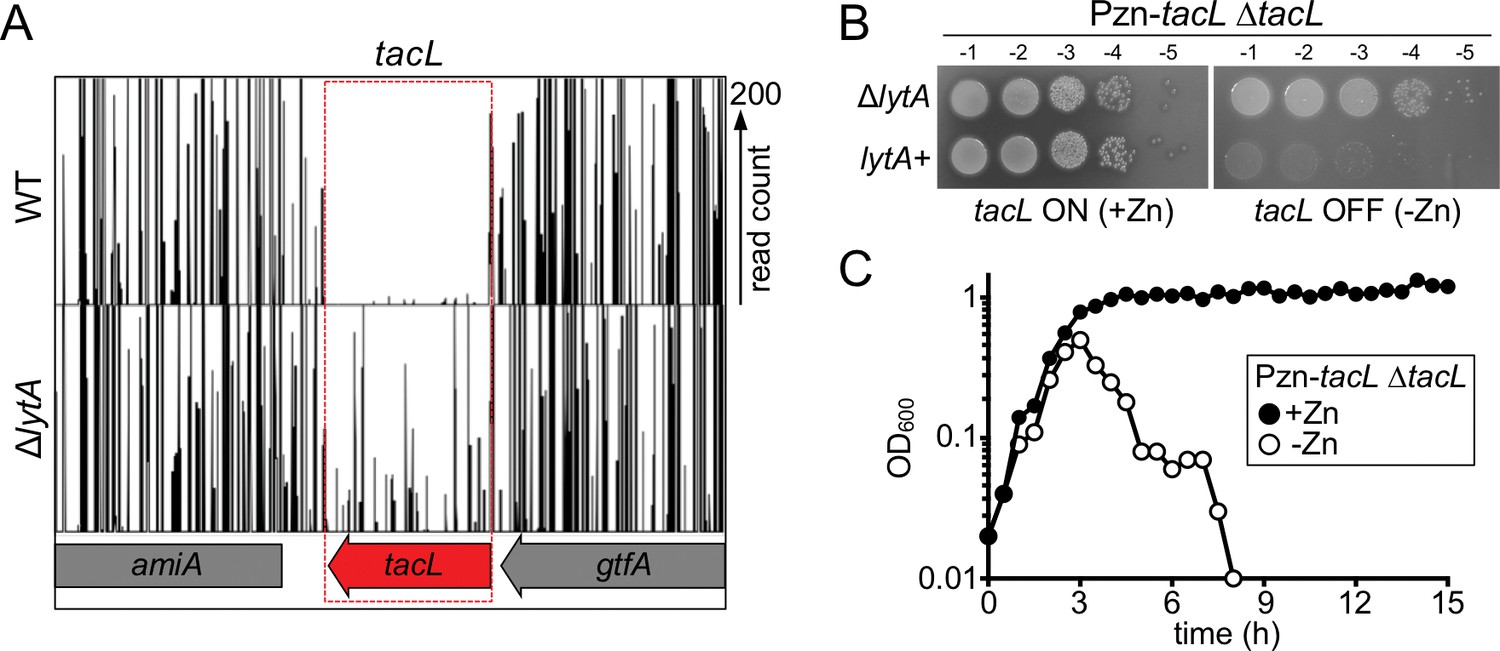
The essential gene tacL can be inactivated in cells lacking lytA.
(A) Transposon insertion profiles from the Tn-Seq screen. Mariner transposon libraries were generated in wild-type (WT) and ΔlytA mutant strains and insertion sites were mapped to the Sp genome using Illumina sequencing. The height of each line reflects the number of sequencing reads at each position. Note that transposon insertions in tacL were much more readily isolated in cells lacking lytA. (B) Spot dilutions of the indicated strains in the presence and absence of inducer (Zn). The indicated strains were grown to exponential phase, normalized for OD600 and serially diluted. Aliquots (5 μl) of each dilution were spotted onto TSAII 5% SB plates in the presence or absence of 100 µM ZnCl2. Plates were incubated at 37 ˚C in 5% CO2 and imaged. (C) Depletion of tacL results in growth arrest and lysis in exponential phase while its overexpression results in protection against growth-phase-dependent autolysis. Strains containing a zinc-inducible tacL allele (Pzn-tacL) were grown in THY to mid-exponential phase. Cultures were diluted into fresh THY to an OD600 of 0.025 in the presence or absence of 100 µM ZnCl2 and grown at 37 ˚C in 5% CO2. Growth was monitored by taking OD600 measurements approximately every 30 min for 15 hr.
Figure 3
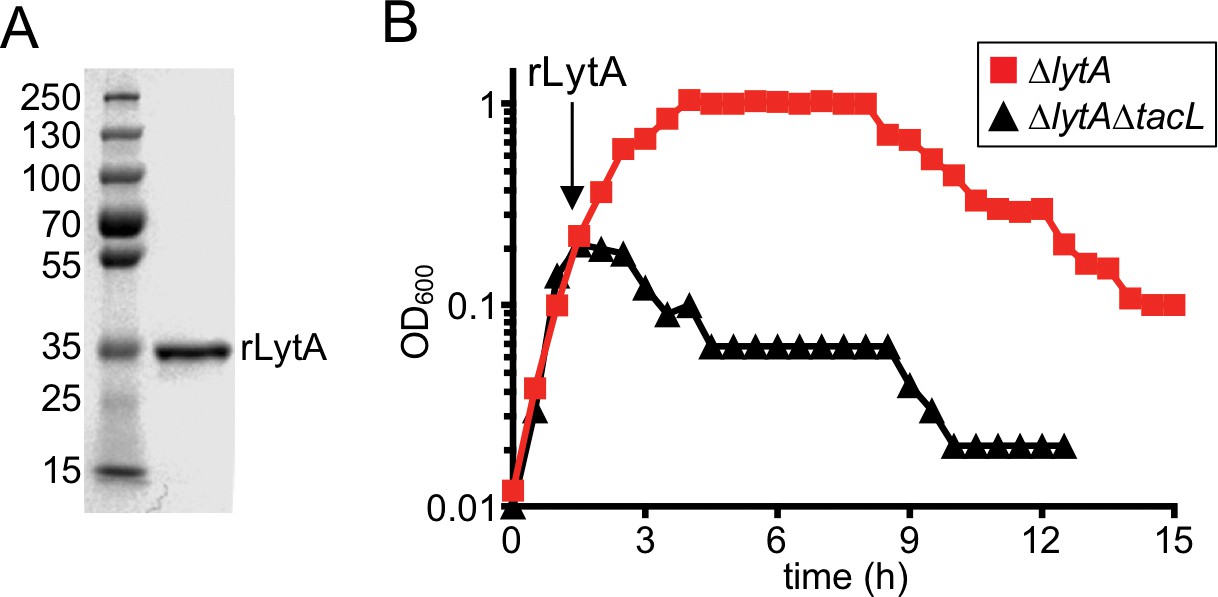
Cells lacking TacL are hypersensitive to exogenous LytA.
(A) Coomassie-stained gel of recombinant LytA (rLytA) purified from E. coli. Molecular weight markers (in kDa) are shown. (B) Growth curves of the indicated strains before and after the addition of 1 μg/ml rLytA. Cells lacking lytA (ΔlytA) or tacL and lytA (ΔlytA ΔtacL) were challenged with rLytA at an OD600 of ~0.2. In the absence of TacL, cells rapidly lyse after rLytA addition. By contrast and as reported previously, the ΔlytA (TacL+) strain only lyses in stationary phase in a manner similar to LytA+ cells (Fernebro et al., 2004; Mellroth et al., 2012).
Figure 4
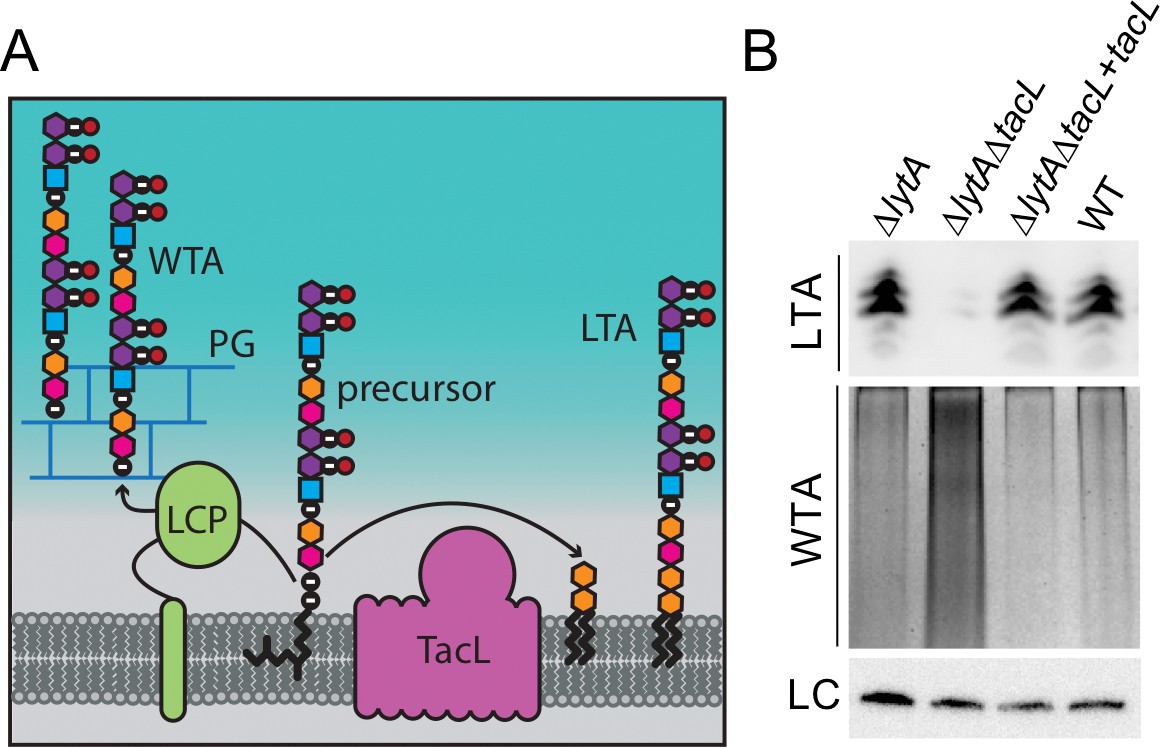
Cells lacking TacL contain altered levels of teichoic acids.
(A) Schematic model depicting the final steps in the synthesis of WTA and LTA in Sp. WTAs and LTAs are thought to be synthesized from a common precursor polymer that is linked to an undecaprenyl phosphate lipid carrier (Denapaite et al., 2012). The polymer is transferred to the PG by LCP proteins to form WTA (Denapaite et al., 2012). TacL is hypothesized to transfer the precursor to a glycolipid anchor to generate LTA (Heß et al., 2017). (B) Analysis of LTA and WTA levels during exponential growth in wild-type (WT) or cells lacking LytA (∆lytA), LytA and TacL (∆lytA∆tacL) or the double mutant harboring a Zn-inducible tacL allele (+tacL) grown in the presence of 100 µM ZnCl2. Top: Immunoblot analysis of membrane-associated LTAs separated by 16% Tris-tricine SDS-PAGE and probed with a monoclonal antibody specific for phosphocholine. Middle: Analysis of WTAs released from purified cell wall sacculi and separated by SDS-PAGE followed by alcian blue-silver staining. Bottom: LC, loading control.
Figure 5
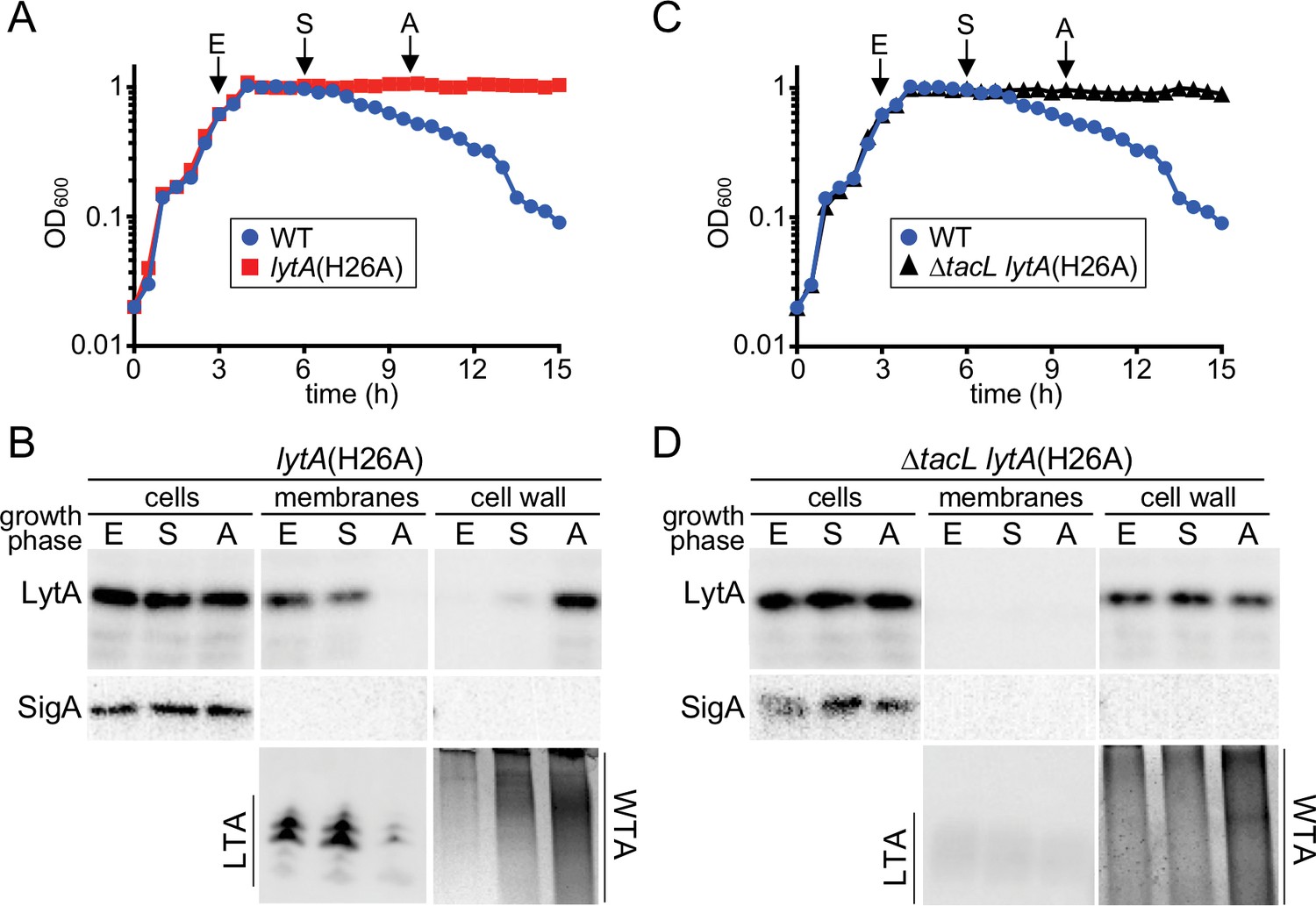
A switch from LTA to WTA synthesis and LytA localization occurs at the onset of autolysis.
(A and C) Growth curves of the indicated strains cultured at 37 ˚C in 5% CO2. At the indicated time points (E, exponential; S, stationary; A, autolysis), samples from the lytA(H26A) mutants were collected, normalized to an OD600 of 0.5 and processed as described in Materials and methods. The WT growth curve was used as a reference for the timing of autolysis. (B and D) Samples from (A) and (C) were analyzed to detect LytA(H26A) in whole cell lysates (cells), associated with protoplast membranes, or cell wall sacculi. The cytoplasmic protein SigA was used as a control for protoplast integrity. The immunoblots were from the same membrane and exposure but were cropped to re-order the lanes for clarity. LTAs in the membrane fraction and WTA in the cell wall fraction were monitored by immunoblot and alcian blue-silver staining, respectively.
Figure 6
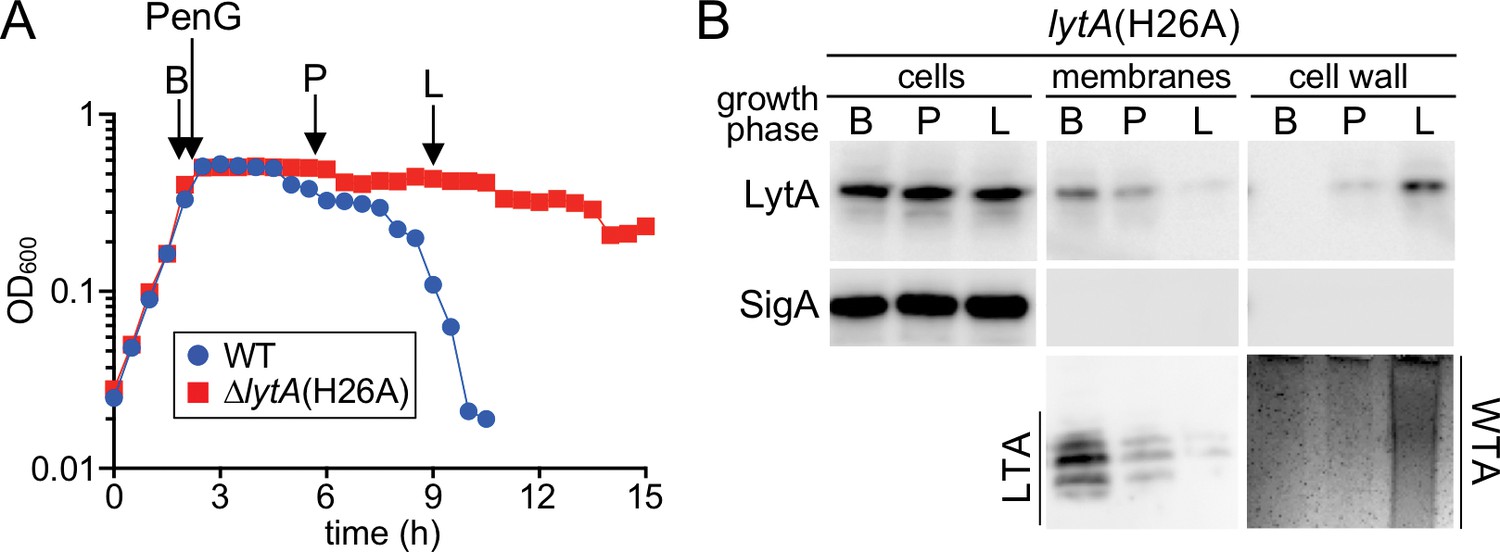
A switch from LTA to WTA synthesis and LytA localization occurs after exposure to penicillin.
(A) Growth curves of the indicated strains before and after challenge with penicillin G (PenG) (0.5 μg/ml final). At the indicates time points (B, before PenG addition; A, after addition; L, lysis), samples from the lytA(H26A) strain were collected, normalized to an OD600 of 0.5 and processed as described in Materials and methods. The growth curve of WT treated with PenG was used as reference for the timing of lysis (B) Samples from (A) were analyzed to detect LytA(H26A) in whole cell lysates (cells), associated with protoplast membranes, or with cell wall sacculi. The cytoplasmic protein SigA was used as a control for protoplast integrity. The immunoblots were from the same membrane and exposure but were cropped to re-order the lanes for clarity. LTAs in the membrane fraction and WTA in the cell wall fraction were monitored by immunoblot and alcian blue-silver staining, respectively.
Figure 7
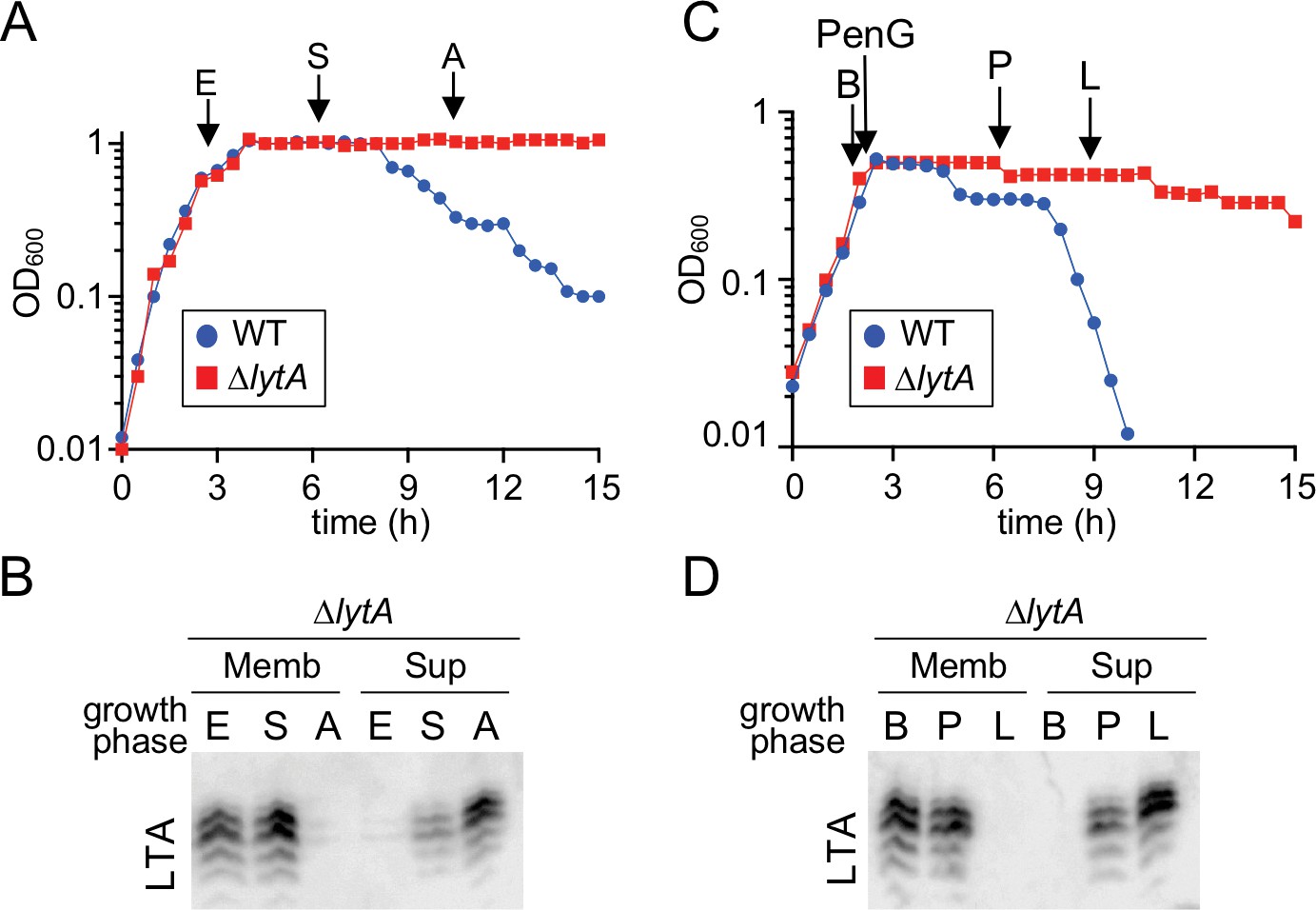
Teichoic acids are released into the culture medium during autolysis.
(A) Growth curve of the indicated strains cultured at 37 ˚C in 5% CO2. At the indicated time points (E, exponential; S, stationary; A, autolysis), samples from the ΔlytA mutant were collected, normalized to an OD600 of 0.5 and processed as described in Materials and methods. The WT growth curve was used as a reference for the timing of autolysis. (B) Samples from (A) were analyzed to detect LTA in membranes (Memb) or culture medium (Sup, supernatant) by immonoblot. (C) Growth curve of the indicated strains cultured at 37 ˚C in 5% CO2. At an OD600 of ~0.5 the cultures were challenged with penicillin G (PenG) (0.5 µg/ml final). At the indicated time points (B, before PenG addition; A, after addition; L, lysis), samples from the ∆lytA mutant were collected and normalized to an OD600 of 0.5. (D) Samples from (C) were analyzed to detect LTA in membranes (Memb) or supernatant (Sup) by immunoblot.
Figure 8 with 2 supplements
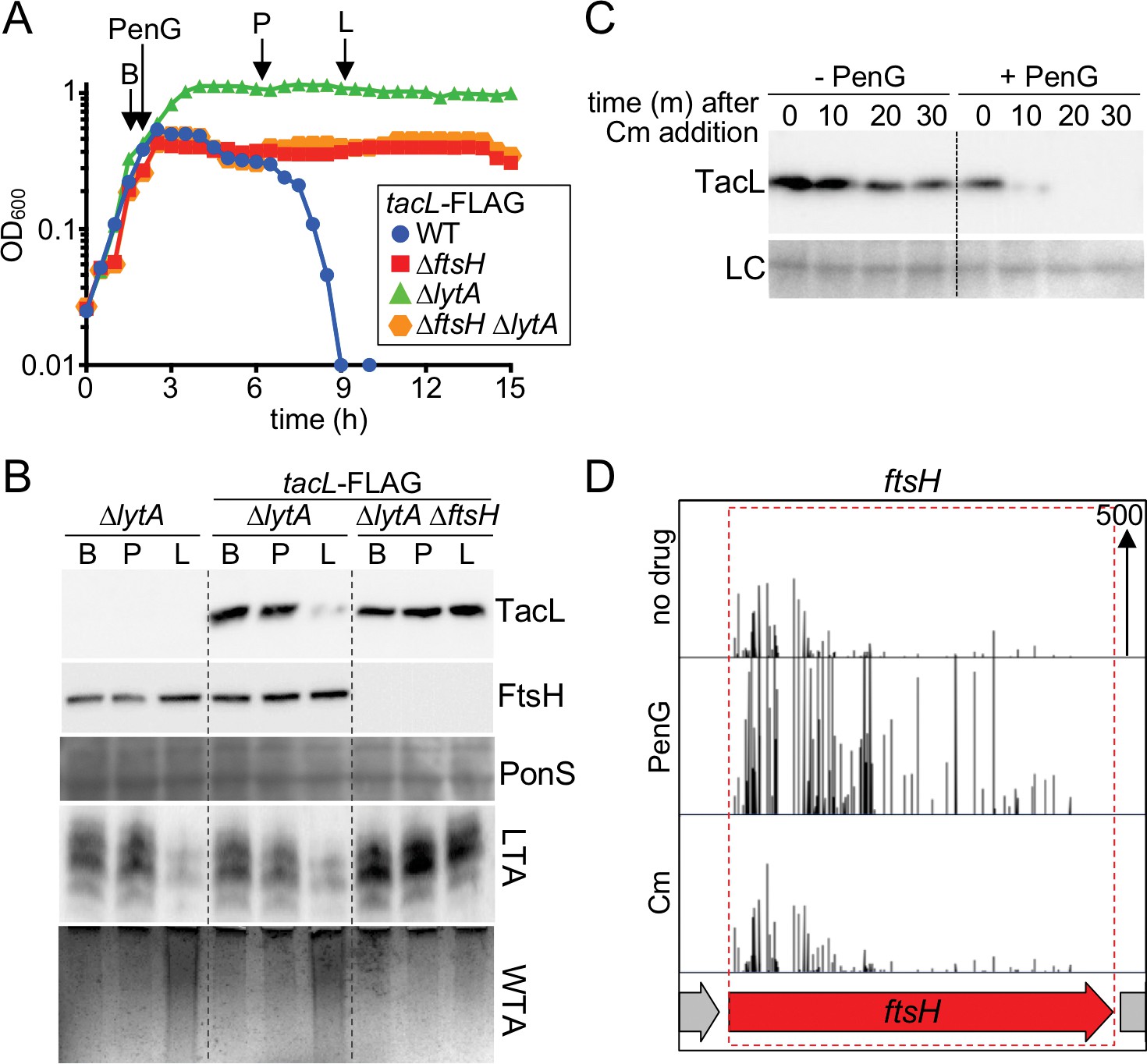
Penicillin treatment causes a reduction in TacL protein levels that depends on FtsH.
(A) Growth curves of the indicated strains harboring a functional TacL-FLAG fusion as the sole source of TacL. At an OD600 of ~0.5 the cultures were challenged with penicillin G (PenG) (0.5 µg/ml final). At the indicated time points (B, before penG addition; A, after addition; L, lysis), samples from the ∆lytA and ∆ftsH∆lytA strains were collected and normalized to an OD600 of 0.5. Samples from a TacL+ ∆lytA strain were collected at the same time points. The growth curve of this strain was omitted from the figure for clarity. The growth curve of the TacL-FLAG LytA+ strain (WT) treated with PenG was used as reference for the timing of lysis. (B) Samples from (A) were processed as described in Materials and methods to detect TacL-FLAG and FtsH from whole cell lysates, LTA from membrane preparations, and WTAs released from purified cell wall sacculi. A region of the nitrocellulose membrane used for immunoblot analysis was stained with Ponceau S (PonS) to control for loading. (C) TacL-FLAG stability before and after PenG exposure. Wild-type Sp harboring TacL-FLAG was grown as in (A). Prior to PenG exposure (-PenG) or 2 hr after addition (+PenG), cultures were treated with chloramphenicol (Cm) (50 μg/ml final) to block translation. Samples were collected immediately before and 10, 20, 30 min after Cm addition and analyzed by SDS-PAGE and anti-FLAG immunoblotting to detect TacL. A region of the nitrocellulose membrane used for immunoblot analysis was stained with Ponceau S (PonS) to control for loading. (D) Transposon insertion profiles from a mariner transposon library generated in wild-type Sp and grown in the presence or absence of sub-inhibitory doses of penicillin G (PenG) or chloramphenicol (Cm). Transposon insertions in ftsH were significantly enriched (p<0.0001) in the presence of PenG compared to no drug or the Cm-treated control. The following figure supplements are available for Figure 8.
Figure 8—figure supplement 1

tacL overexpression prevents growth-phase-dependent and antibiotic-induced autolysis.
(A) Growth curves of wild-type (WT) Sp and a strain harboring a zinc-inducible tacL allele (WT +tacL) Exponentially growing cultures were diluted into fresh THY at an OD600 of 0.025 in the presence of 100 µM ZnCl2 and incubated at 37°C in 5% CO2. (B) The same strains as in (A) were grown in THY in the presence of 100 µM ZnCl2 to an OD600 of 0.5 and then challenged with penicillin G (0.5 μg/ml final). Growth was monitored approximately every 30 min for 15 hr. Note that tacL overexpression blocks cell lysis in both conditions.
Figure 8—figure supplement 2
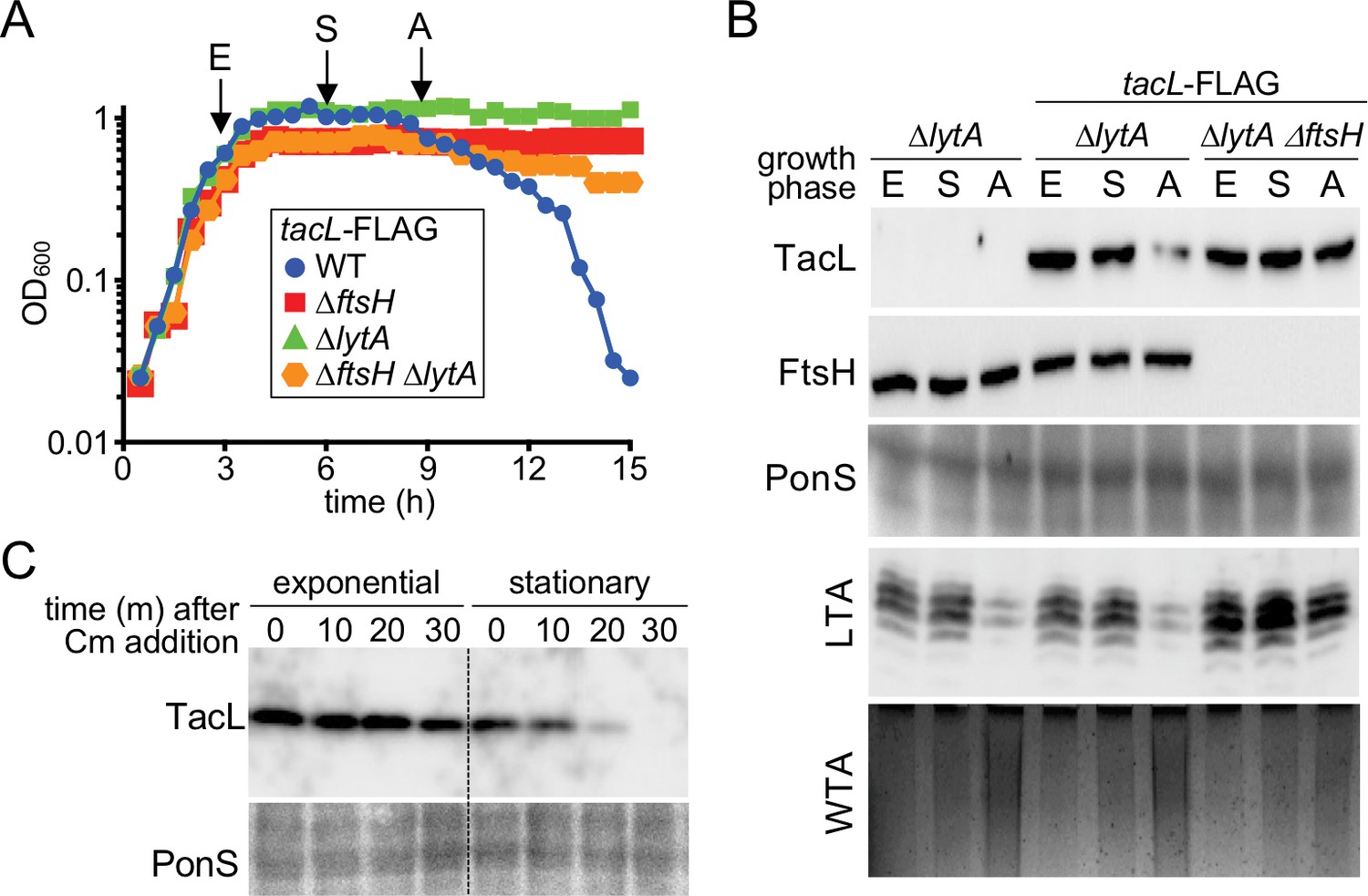
Reduction in TacL protein levels correlates with the growth phase-dependent switch from LTA to WTA synthesis and autolysis.
(A) Growth curves of the indicated strains harboring a functional TacL-FLAG fusion as the sole source of TacL. At the indicated time points (E, exponential phase; S, stationary phase; A, autolysis), samples from the ∆lytA and ∆ftsH ∆lytA strains were collected and normalized to an OD600 of 0.5. Samples from a TacL+ ∆lytA strain were collected at the same time points. The growth curve of this strain was omitted from the figure for clarity. The growth curve of the TacL-FLAG LytA+ strain (WT) was used as reference for the timing of autolysis. (B) Samples from (A) were processed as described in Materials and methods to detect TacL-FLAG and FtsH from whole cell lysates, LTA from membrane preparations, and WTAs released from purified cell wall sacculi. A region of the nitrocellulose membrane used for immunoblot analysis was stained with Ponceau S (PonS) to control for loading. (C) TacL-FLAG stability in exponential and stationary phases. Wild-type Sp harboring TacL-FLAG was grown as in (A). In exponential phase (E) or 2 hr after entry into stationary phase (S), cultures were treated with chloramphenicol (Cm) (50 μg/ml final) to block translation. Samples were collected immediately before and 10, 20, 30 min after Cm addition and analyzed by SDS-PAGE and anti-FLAG immunoblotting to detect TacL. A region of the nitrocellulose membrane used for immunoblot analysis was stained with Ponceau S (PonS) to control for loading.
Figure 9
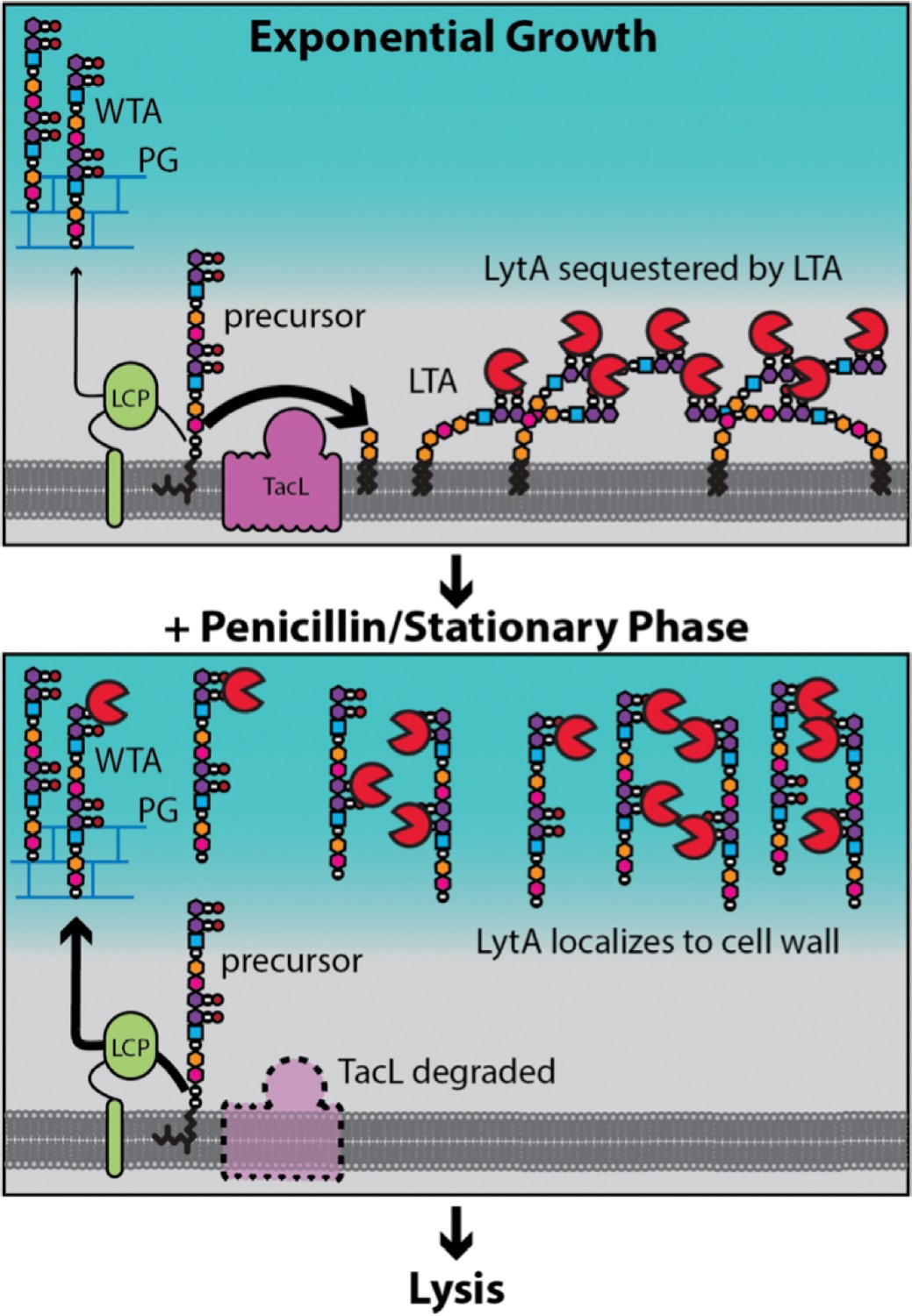
Model for LytA regulation.
During exponential growth, TacL-dependent LTA synthesis dominates over LCP-dependent production of WTA. LytA bound to the phosphocholine moeities (purple/black balls) on LTA is sequestered away from WTA preventing LytA from targeting the cell wall. Upon entry into stationary phase or exposure to cell wall targeting antibiotics (penicillin), TacL is degraded in an FtsH-dependent manner leading to a reduction in LTA synthesis and an increase in WTA levels. Association of LytA with newly synthesized WTA leads to cell wall cleavage and lysis.
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Strain (Streptococcus pneumoniae(Sp) D39 Δcps) | WT (Sp D39 Δcps) | Malcolm Winkler lab (Lanie et al., 2007) | Wildtype S. pneumoniae D39 Δcps | |
| Strain, strain background (Sp D39 Δcps) | AKF_Spn001 | Fenton et al., 2016 | ΔbgaA::kan | |
| Strain, strain background (Sp D39 Δcps) | AKF_Spn002 | Fenton et al., 2016 | ΔbgaA::add9(spec) | |
| Strain, strain background (Sp D39 Δcps) | AKF_Spn003 | Fenton et al., 2016 | ΔbgaA::tetM(tet) | |
| Strain, strain background (Sp D39 Δcps) | AKF_Spn004 | Fenton et al., 2016 | ΔbgaA::cat | |
| Strain, strain background (Sp D39 Δcps) | AKF_Spn005 | Fenton et al., 2016 | ΔbgaA::erm | |
| Strain, strain background (Sp D39 Δcps) | AKF_Spn351 | Fenton et al., 2016 | ΔlytA::cat | |
| Strain, strain background (Sp D39 Δcps) | AKF_Spn704 | This study | ΔlytA::erm | (see Materials and methods) |
| Strain, strain background (Sp D39 Δcps) | JFK_Spn001 | This study | ΔlytA::erm ΔtacL::cat | (see Materials and methods) |
| Strain, strain background (Sp D39 Δcps) | JFK_Spn002 | This study | ΔftsH::cat | (see Materials and methods) |
| Strain, strain background (Sp D39 Δcps) | JFK_Spn003 | This study | ΔlytA::erm ΔftsH::cat | (see Materials and methods) |
| Strain, strain background (Sp D39 Δcps) | JFK_Spn004 | This study | lytA(H26A), erm | (see Materials and methods) |
| Strain, strain background (Sp D39 Δcps) | JFK_Spn005 | This study | lytA(H26A), erm ΔtacL::cat | (see Materials and methods) |
| Strain, strain background (Sp D39 Δcps) | JFK_Spn006 | This study | tacL-FLAG, spec | (see Materials and methods) |
| Strain, strain background (Sp D39 Δcps) | JFK_Spn007 | This study | tacL-FLAG, spec ΔftsH::cat | (see Materials and methods) |
| Strain, strain background (Sp D39 Δcps) | JFK_Spn008 | This study | tacL-FLAG, spec ΔlytA::erm | (see Materials and methods) |
| Strain, strain background (Sp D39 Δcps) | JFK_Spn009 | This study | tacL-FLAG, spec ΔlytA::erm ΔftsH::cat | (see Materials and methods) |
| Strain, strain background (Sp D39 Δcps) | JFK_Spn010 | This study | ΔbgaA::(PZn-tacL, tet) | (see Materials and methods) |
| Strain, strain background (Sp D39 Δcps) | JFK_Spn011 | This study | ΔbgaA::(PZn-tacL, tet) ΔlytA::erm | (see Materials and methods) |
| Strain, strain background (Sp D39 Δcps) | JFK_Spn012 | This study | ΔbgaA::(PZn-tacL, tet) ΔtacL::cat | (see Materials and methods) |
| Strain, strain background (Sp D39 Δcps) | JFK_Spn013 | This study | ΔbgaA::(PZn-tacL, tet) ΔtacL::cat ΔlytA::erm | (see Materials and methods) |
| Strain, strain background (E. coli DH5a) | DH5a | Gibco BRL | F-hsdR17 Δ(argF-lacZ)U169 phoA glnV44 Φ80dlacZ Δ M15 gyrA96 recA1 endA1 thi-1 supE44 deoR | |
| Strain, strain background (E. coli BL21) | BL21 | New England Biolabs | E. coli B F– ompT gal dcm lon hsdSB(rB–mB–) [malB+]K-12(λS) | |
| Recombinant DNA reagent | pLEM023 | Fenton et al., 2016 | bgaA’::Pzn::MCS::tetM::bgaA’ bla | plasmid |
| Recombinant DNA reagent | pET24a | Novagen | PT7, lacIq; vector used for protein expression | plasmid |
| Recombinant DNA reagent | pMagellan6 | Fenton et al., 2016 | IRL(MmeI)::add9::IRR(MmeI) bla | plasmid |
| Recombinant DNA reagent | ‘pMalC9’ | Fenton et al., 2016 | MBP::Himar1 bla | plasmid |
| Recombinant DNA reagent | pJFK_001 | This study | tacL in pLEM023 | plasmid (see Materials and methods) |
| Recombinant DNA reagent | pJFK_002 | This study | lytA in pET24a | plasmid (see Materials and methods) |
| Antibody | LytA (rabbit polyclonal) | This study | 1:50000 (see Materials and methods) | |
| Antibody | FtsH (rabbit polyclonal) | Kotschwar et al., 2004 | 1:10000 | |
| Antibody | SigA (rabbit polyclonal) | Fujita, 2000 | 1:10000 | |
| Antibody | FLAG (TacL-FLAG; rabbit polyclonal) | Sigma | RRID:AB_796202 | 1:1000 |
| Antibody | LTA (mouse monoclonal) | Sigma | RRID:AB_1163630 | 1:1000; anti-Phosphocholine TEPC-15 |
| Antibody | anti-rabbit IgG-HRP | BioRad | RRID:AB_1102634 | 1:20000 |
Additional files
-
Supplementary file 1
Table of oligonucleotides used in this study
- https://doi.org/10.7554/eLife.44912.014
-
Transparent reporting form
- https://doi.org/10.7554/eLife.44912.015
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
A switch in surface polymer biogenesis triggers growth-phase-dependent and antibiotic-induced bacteriolysis
eLife 8:e44912.
https://doi.org/10.7554/eLife.44912




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}