Cryo-EM structures and functional characterization of murine Slc26a9 reveal mechanism of uncoupled chloride transport
- University of Zurich, Switzerland
Abstract
The epithelial anion transporter SLC26A9 contributes to airway surface hydration and gastric acid production. Colocalizing with CFTR, SLC26A9 has been proposed as a target for the treatment of cystic fibrosis. To provide molecular details of its transport mechanism, we present cryo-EM structures and a functional characterization of murine Slc26a9. These structures define the general architecture of eukaryotic SLC26 family members and reveal an unusual mode of oligomerization which relies predominantly on the cytosolic STAS domain. Our data illustrates conformational transitions of Slc26a9, supporting a rapid alternate-access mechanism which mediates uncoupled chloride transport with negligible bicarbonate or sulfate permeability. The characterization of structure-guided mutants illuminates the properties of the ion transport path, including a selective anion binding site located in the center of a mobile module within the transmembrane domain. This study thus provides a structural foundation for the understanding of the entire SLC26 family and potentially facilitates their therapeutic exploitation.
https://doi.org/10.7554/eLife.46986.001eLife digest
Many processes in the human body are regulated by chloride and other charged particles (known as ions) moving in and out of cells. Each cell is surrounded by a membrane barrier, which prevents ions from entering or exiting. Therefore, to control the levels of ions inside the cell, specific proteins in the membrane act as channels or transporters to provide routes for the ions to pass through the membrane.
Channel proteins form pores that, when open, allow a steady stream of ions to pass through the membrane. Transporter proteins, on the other hand, generally contain a pocket that is only accessible from one side of the membrane. When individual ions enter this pocket the transporter changes shape. This causes the entrance of the pocket to close and then re-open on the other side of the membrane.
Inside the lung, an ion channel known as CFTR provides a route for chloride ions to move out of cells, which helps clear harmful material from the airways. Mutations affecting this protein cause the mucus lining the airways to become very sticky, leading to a severe disease known as cystic fibrosis. CFTR works together with another protein that is also found in the membrane, called SLC26A9. Previous studies have suggested that SLC26A9 also allows chloride ions to pass through the membrane. It was not clear, however, if SLC26A9 operates as an ion channel or a transporter protein, or how the protein is arranged in the membrane.
Now, Walter, Sawicka and Dutzler combined two techniques known as cryo-electron microscopy and patch-clamp electrophysiology to reveal the detailed three-dimensional structure of the mouse version of SLC26A9, which is highly similar to the human form. The experiments found that mouse SLC26A9 proteins form pairs in the membrane referred to as homodimers, which arranged themselves in an unexpected way. Further investigation into the structure of these homodimers suggests that despite having many channel-like properties, SLC26A9 operates as a fast transporter, rather than a true channel.
These findings help us understand the role of SLC26A9 and other similar proteins in the lung and other parts of the body. In the future it may be possible to develop drugs that target SLC26A9 to treat cystic fibrosis and other severe lung diseases.
https://doi.org/10.7554/eLife.46986.002Introduction
Solute Carrier 26 family member A9 (SLC26A9) is a membrane-transport protein that in recent years has gained considerable attention due to its intriguing functional properties and promising therapeutic potential (Balázs and Mall, 2018). It is found abundantly in the epithelia of lung and stomach (Chang et al., 2009b; Lohi et al., 2002; Xu et al., 2005), where it mediates luminal electrogenic chloride transport and contributes to airway clearance and the production of gastric acid. Slc26a9–/– knockout mice develop airway and gastrointestinal pathologies (Anagnostopoulou et al., 2012; Liu et al., 2015; Xu et al., 2008), and human genetic polymorphisms in SLC26A9 are correlated with altered severity of airway clearance disorders (Anagnostopoulou et al., 2012; Miller et al., 2015; Sun et al., 2012). Owing to these associations, as well as its co-localization and proposed regulatory interactions with the cystic fibrosis transmembrane conductance regulator (CFTR) (Avella et al., 2011; Bertrand et al., 2017; Bertrand et al., 2009; Chang et al., 2009a; Strug et al., 2016), SLC26A9 has been proposed as a therapeutic target for treating complications associated with cystic fibrosis and other diseases of the airways and gastrointestinal tract (Balázs and Mall, 2018; Mall and Galietta, 2015). The protein is a member of the SLC26 family of anion transporters, which play indispensable roles in a wide variety of physiological processes such as hearing, cartilage and bone formation, and epithelial secretion and absorption (Alper and Sharma, 2013; Dorwart et al., 2008b). The ten functional SLC26 paralogs expressed in mammals are targeted to specific tissues and show a distinct transport mechanism and substrate preference (Alper and Sharma, 2013), with the human ortholog SLC26A5 (Prestin) having insignificant transport function and instead operating as voltage-dependent motor protein which is responsible for electromotility in cochlear outer hair cells (Dallos and Fakler, 2002). The transport mode of SLC26A9 is controversial, with some reports providing evidence of chloride/bicarbonate exchange (Chang et al., 2009b; Xu et al., 2005), yet others claiming that SLC26A9 has negligible bicarbonate permeability and instead confers an uncoupled, constitutively-active, channel-like Cl– conductance (Bertrand et al., 2009; Dorwart et al., 2007; Loriol et al., 2008). Although less characterized, uncoupled Cl– transport has also been reported for SLC26A7 (Kim et al., 2005) and SLC26A11 (Rahmati et al., 2013) whereas evidence for mixed uncoupled and coupled exchange properties was shown for SLC26A3 and SLC26A6 (Ohana et al., 2011; Shcheynikov et al., 2006).
No high-resolution structures have yet been reported for SLC26A9 or any other eukaryotic SLC26 family member, and detailed structural information about a full-length SLC26 protein is limited to a single crystal structure of a bacterial fumarate transporter (SLC26Dg) (Geertsma et al., 2015). The monomeric SLC26Dg structure provided valuable information about the basic elements of the SLC26 architecture consisting of a transport domain of 14 membrane-inserted α-helices, and a C-terminal cytosolic STAS domain of unknown function. However, SLC26 transporters assemble into functional oligomers (Compton et al., 2014; Detro-Dassen et al., 2008; Zheng et al., 2006), and the quaternary arrangement of an oligomeric eukaryotic SLC26 transporter has remained unknown. Moreover, structural knowledge of mammalian paralogs is indispensable for the comprehension of the molecular basis for the mechanistic diversity of the SLC26 family.
Here, we present cryo-electron microscopy (cryo-EM) structures of dimeric murine Slc26a9 accompanied by its detailed functional characterization. The structures of an inward-facing conformation as well as a potentially intermediate state of the transport cycle reveal a protein architecture that is likely representative for all mammalian SLC26 paralogs, including the unusual mode of dimerization in which nearly all the interactions between the two subunits are mediated by the STAS domains. In combination with electrophysiological experiments, our work demonstrates how Slc26a9 mediates the uncoupled transport of Cl– via a rapid alternate-access mechanism.
Results
Functional characterization of Slc26a9
Upon transfection of HEK293 cells with DNA encoding full-length murine Slc26a9, high expression is observed, but the protein is poorly detergent-extractable and exhibits a tendency to aggregate. Therefore, we designed a minimal construct in which two predicted intrinsically disordered regions (IDRs) were removed from the C-terminal part of the protein (Figure 1—figure supplement 1), a strategy which allowed crystallization of the isolated STAS domain of Prestin (Lolli et al., 2016; Pasqualetto et al., 2010). One excised region consists of residues P558-V660 and encompasses a hypervariable IDR which was previously termed the STAS intervening sequence (IVS) (Dorwart et al., 2008a; Pasqualetto et al., 2010). The second deleted region comprises the final 44 residues of the C-terminus (CT, residues P745-L790) subsequent to the predicted STAS domain fold and contains a terminal PDZ motif which commonly interacts with multi-domain scaffolding proteins (Kim and Sheng, 2004). The resulting dual ∆IVS/∆CT truncation construct has been designated Slc26a9T.
A striking observation from HEK293 cells expressing fluorescently-tagged Slc26a9T is that the truncations have apparently resulted in greater surface expression (Figure 1—figure supplement 2A). This is also observed for a construct only lacking the IVS (SLC26A9ΔIVS), but not for a construct solely lacking the CT (SLC26A9ΔCT), thus suggesting that the IVS might be responsible for the retention of the protein in intracellular membranes (Figure 1—figure supplement 2A). Relative to the full-length protein, Slc26a9T exhibits several additional favorable qualities, including more efficient detergent extraction and the capacity to be purified to homogeneity in a dimeric state (Figure 1—figure supplement 2B–C). To assess whether Slc26a9T retains transport activity, the purified protein was reconstituted into lipid vesicles and tested for electrogenic Cl– transport using a fluorometric assay (Kane Dickson et al., 2014). In these experiments, we observed robust transport activity that can be diminished in a dose-dependent manner by a known inhibitor of Slc26a9, implying that neither the removal of the IDRs from the STAS domain nor the purification procedure has compromised the function of Slc26a9T (Figure 1A, Figure 1—figure supplement 2D). Observation of Cl– transport by reconstituted Slc26a9T also demonstrates that the protein does not require the presence of endogenous cellular factors for its activity.
Figure 1 with 3 supplements see all
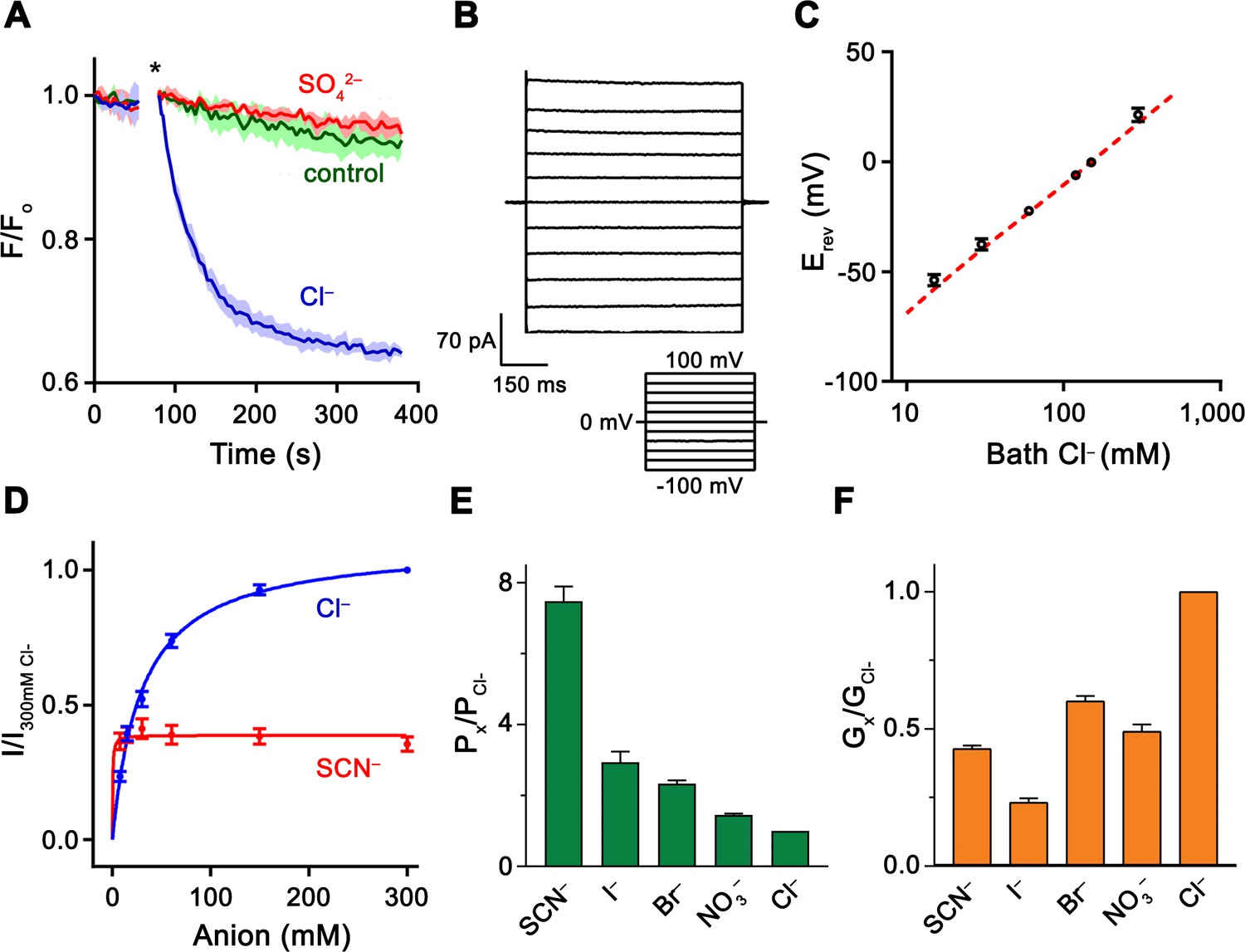
Functional properties of Slc26a9.
(A) Cl– transport of Slc26a9T reconstituted into proteoliposomes, monitored by the fluorescence change of the pH gradient-sensitive fluorophore ACMA. (*) Indicates addition of the H+ ionophore CCCP, which allows counterion movement and electrogenic Cl– transport to proceed. Green trace (control) corresponds to liposomes not containing any protein in the presence of an inward Cl– gradient, red trace (SO42–) to proteoliposomes exposed to external SO42– and blue trace (Cl–) to proteoliposomes exposed to external Cl–. Traces show mean of three technical replicates. (B) Representative current trace recorded from excised patches from HEK293T cells expressing Slc26a9T at symmetrical 150 mM Cl–. Inset shows voltage protocol. (C) Reversal potentials at 150 mM extracellular NaCl and varying intracellular NaCl concentrations recorded from inside-out patches (PNa+/PCl–=0.02). Red line corresponds to the Cl– Nernst potential. Presented data is the mean of 5 biological replicates, except for 300 mM, for which n = 3. (D) Conductance–concentration relationships of anion transport across Slc26a9T. Data were recorded at −100 mV from inside-out patches with a pipette solution containing 7.5 mM Cl– and varying intracellular Cl– (blue) or SCN– (red) concentrations. Km (Cl–)=29.5 mM; Km (SCN–)=0.5 mM; n = 5, errors are s.e.m.. (E) Permeabilities and (F) conductance ratios (recorded at −100 mV) were obtained from bi-ionic substitution experiments. Biological replicates for Px/PCl and Gx/GCl: Cl–, n = 10; Br–, n = 10; NO3–, n = 9; SCN–, n = 8; I–, n = 6. C-E, Errors are s.e.m.
For a thorough functional characterization of the transport activity, we recorded currents from HEK293T cells transfected with DNA encoding either full-length Slc26a9 or Slc26a9T. In the whole-cell configuration, both constructs yield constitutively active, anion-selective currents with linear current-voltage (I-V) relationships, as reported previously for human SLC26A9 (Bertrand et al., 2009; Chang et al., 2009b; Dorwart et al., 2007; Loriol et al., 2008) (Figure 1—figure supplement 2E–H). As expected from the higher surface expression, Slc26a9T-transfected cells show raw currents which are, on average, at least four-fold larger than currents associated with full-length Slc26a9 (Figure 1—figure supplement 2E–H). Moreover, whereas we could not reliably measure macroscopic currents in inside-out patches from full-length Slc26a9-transfected cells, robust activity was observed for Slc26a9T (Figure 1B). As for whole-cell experiments, these currents are constitutive and exhibit a linear I-V relationship. Contrary to a previous study (Chang et al., 2009b), we did not observe any discrete single-channel transitions in any of our recordings. Since excised patches allow better control of intra- and extracellular conditions, we used this method to characterize functional properties for the remainder of our investigations.
To assess the relative anion/cation selectivity, we measured I-V relationships for inside-out patches of Slc26a9T-expressing cells that were subjected to different NaCl gradients. In all cases, the currents reverse very close to the Nernst potential of Cl–, implying that Cl– is the only transported charge in the system and excluding obligate coupling to other ions (Figure 1C; Figure 1—figure supplement 3A). Next, we evaluated the relationship between substrate concentration and ion flux and found a saturation of currents at high Cl– concentrations (with a Km of 30 mM, Figure 1D), consistent with a weak interaction of the protein with its transported ion. Although the high Cl– selectivity, fast kinetics, and weak substrate interaction would all be consistent with a channel-like mechanism, they do not exclude transport by an alternate-exchange mechanism, since a similar high Km for Cl– (i.e. ~46 mM) was described for the rapid Cl–/HCO3– exchanger SLC4A1/Band 3 (Liu et al., 1996).
To characterize the ability of Slc26a9T to discriminate between different anions, we pursued bi-ionic substitution experiments. The observed permeability sequence (SCN–>I–>Br– > NO3–>Cl–) follows a lyotropic series (Wright and Diamond, 1977), which favors more easily dehydrated anions (Figure 1E; Figure 1—figure supplement 3B,C). In contrast, we measured negligible permeabilities for the anions F–, HCO3–, and SO42– (Figure 1—figure supplement 3D–F). The permeability sequence agrees with previous measurements of human SLC26A9 derived from oocyte voltage-clamp experiments (Dorwart et al., 2007; Loriol et al., 2008). However, the absence of HCO3– permeability is in contrast to some reports showing evidence of Cl–/HCO3– exchange by Slc26a9 which were based on measurements of intracellular pH changes in response to removal of external Cl– in the presence of bath HCO3– (Chang et al., 2009b; Xu et al., 2005). While we cannot fully account for this discrepancy we note that all reports of electrogenic transport indicate negligible movement of HCO3–. To assess relative transport rates, macroscopic conductance ratios (Gx/GCl) of permeable anions revealed a conductivity sequence of Cl–>Br– > NO3–~SCN–>I– (Figure 1F; Figure 1—figure supplement 3B,C). Notably, the conductivity pattern deviates from the permeability sequence, with Cl– being considerably more conductive than any other tested anion. The high relative permeability of lyotropic anions shows that they face a lower energy barrier for dehydration and entry into the transport pathway compared with Cl–, while their lower relative conductance implies that they also encounter a deeper energy well, which in turn would decrease their overall turnover rate. These properties are also reflected in the lower Km and Vmax of SCN– compared to Cl– (Figure 1D) and suggest that Slc26a9 has evolved as an efficient uncoupled Cl– transporter.
Slc26a9 structure
To reveal how the functional features of Slc26a9 as a Cl– transport protein with channel-like properties relate to the protein architecture, we determined its structure by cryo-EM (Figure 2—figure supplements 1–3). A 3.96 Å dataset of Slc26a9T purified in the synthetic digitonin analogue glyco-diosgenin (GDN) was of high quality and allowed the unambiguous interpretation of the cryo-EM density by an atomic model (Figure 2; Figure 2—figure supplements 1 and 2; Table 1; Video 1). To exclude a possible bias from a detergent environment, we have in parallel reconstituted the purified protein into lipid nanodiscs and determined its structure at low (7.77 Å) resolution (Figure 2—figure supplements 3 and 4; Video 2). Both datasets display similar general features of the protein with respect to the organization of subunits and their oligomeric arrangement, although the nanodisc structure reveals localized rigid-body movements indicative of a different functional conformation.
Figure 2 with 4 supplements see all

Slc26a9T structure.
(A) Cryo-EM density of Slc26a9T in the detergent GDN at 3.96 Å contoured at 5σ. Density corresponding to distinct subunits in the dimeric protein is colored in blue and red respectively. Residual density outside the protein region probably corresponding to detergent or lipids is colored in gray. The view is from within the membrane; the boundary of the bilayer is indicated. (B) Ribbon representation of Slc26a9T with subunits colored in blue and red. Protein dimensions are indicated. View is as in (A).
Table 1
Cryo-EM data collection, refinement and validation statistics.
https://doi.org/10.7554/eLife.46986.012| Dataset 1 Slc26a9T in detergent (EMD-4997) (PDB 6RTC) | Dataset 2 Slc26a9T in nanodiscs (EMD-4998) (PDB 6RTF) | |
|---|---|---|
| Data collection and processing | ||
| Microscope | FEI Titan Krios | FEI Tecnai G2 Polara |
| Camera | Gatan K2 Summit + GIF | Gatan K2 Summit + GIF |
| Magnification | 6,511 | 37,313 |
| Voltage (kV) | 300 | 300 |
| Electron exposure (e–/Å2) | 70 | 60 |
| Defocus range (μm) | −0.5 to −3.0 | −0.5 to −3.0 |
| Pixel size (Å)* | 1.075 (0.5375) | 1.34 |
| Symmetry imposed | C2 | C2 |
| Initial particle images (no.) | 416,164 | 711,032 |
| Final particle images (no.) | 112,930 | 17,442 |
| Map resolution (Å) FSC threshold 0.143 | 3.96 | 7.77 |
| Map resolution range (Å) | 3.0–4.2 | 6.0–10.0 |
| Refinement | ||
| Model resolution (Å) FSC threshold 0.5 | 4.0 | 8.0 |
| Model resolution range (Å) | 118–3.96 | |
| Map sharpening b-factor (Å2) | −205 | −512 |
| Model composition Non-hydrogen atoms Protein residues | 9570 1242 | 9570 1242 |
| B factors (Å2) Protein | 73 | 73 |
| Refmac FSCavg/Rfactor | 0.851/0.351 | |
| R.m.s. deviations Bond lengths (Å) Bond angles (°) | 0.005 0.888 | 0.006 1.195 |
| Validation MolProbity score Clashscore Poor rotamers (%) | 1.10 1.19 0 | 1.07 0.78 0 |
| Ramachandran plot Favored (%) Allowed (%) Disallowed (%) | 96.10 3.90 0 | 95.28 4.72 0 |
-
*Values in parentheses indicate the pixel size in super-resolution.
Video 1
Cryo-EM density map of the dimeric Slc26a9T.
Shown is the cryo-EM map of the protein in detergent with the refined model of the inward-facing state superimposed.
Video 2
Cryo-EM density map of the dimeric Slc26a9T in nanodiscs.
Shown is the cryo-EM map of the protein in nanodiscs with the modeled structure of the intermediate state superimposed.
The structure of Slc26a9T is displayed in Figure 2B. The protein is a dimer of elongated shape that is approximately 100 Å long and 50 Å wide (Video 3). Next to the membrane-inserted transport domains, two prominent structural features, which are striking in the cryo-EM density, extend beyond the lipid bilayer and flank the membrane-spanning regions. The interacting domain-swapped STAS domains, appearing as a knob centered on one side of Slc26a9T particles, protrude 38 Å into the cytosol, while a spiky loop on each subunit projects towards the extracellular solution (Figure 2; Figure 2—figure supplements 1–4). The interactions between both subunits of the dimeric protein are highly unusual since, unlike other proteins with a similar fold, they only exhibit minimal contacts between the membrane–inserted domains at the intracellular end of the last transmembrane helix (Figure 2). Of the 8’114 Å2 of the combined molecular surface buried in the dimer, only 11% can be attributed to interactions directly between the transmembrane domains (TMDs). Conversely, 16% of the buried surface is contributed by mutual contacts between STAS domains whereas the remaining interactions are mutually formed between the swapped STAS domain and the transmembrane part of the opposing subunit. Another remarkable feature of the dimer interface concerns the binding of the extended N-terminus to the STAS domain, which appears to contribute to dimer stability (Figure 3—figure supplement 1).
Video 3
The structure of Slc26a9T.
Shown are unique features of a mammalian SLC26 transporter. The oligomerization interface is minimal between the transmembrane domains and is predominantly mediated by the swapped STAS domains. The view of the transmembrane domain only shows two independent modules: core and gate.
The STAS domain as interaction platform
In the Slc26a9T dimer, the cytoplasmic STAS domains act as a platform for interactions between subunits, thus explaining the deleterious transport phenotype observed upon mutations or its removal in different family members (Babu et al., 2010; Dorwart et al., 2008a; Geertsma et al., 2015; Sharma et al., 2011) (Figure 3, Video 3). This cytoplasmic unit closely resembles known STAS domain structures from eukaryotic SLC26A5 (Prestin) (Lolli et al., 2016; Pasqualetto et al., 2010) and the bacterial homolog SLC26Dg (Geertsma et al., 2015) (Figure 3—figure supplement 2A,B). It consists of a six-stranded mixed β-sheet core, which is decorated with four interspersed α-helices (Figure 1—figure supplement 1 and Figure 3—figure supplement 2A). Several of the loop regions connecting β-strands with their adjoining α-helices (i.e. the β3-Cα1, β4-Cα2, β5-Cα3 and the Cα3-β6 loops) make contacts between the STAS domains and the TMD of the opposing subunit at parts that are close to the membrane (at the α4-α5, α8-α9, α12- α13 loops and the terminal helix α14) (Figure 3). The strong conservation of residues at the contact region with the TMD points towards the preservation of this interface among family members (Figure 1—figure supplement 1A and Figure 3—figure supplement 2C). At the bottom part of the STAS dimer, interactions with the N-terminus are mediated by the β1-β2, β2-β3 and β6-Cα4 loops (Figure 3—figure supplement 2D). Between both regions, on the side of the STAS dimer, a negatively charged groove at the dimer interface could potentially interact with cellular factors (Figure 3—figure supplement 2E,F). Finally, in the observed dimeric architecture of the STAS domain, the two unstructured regions removed in the Slc26a9T construct would be located opposite to the dimer-interface and thus be available for interactions with accessory proteins (Figure 3—figure supplement 2G).
Figure 3 with 2 supplements see all
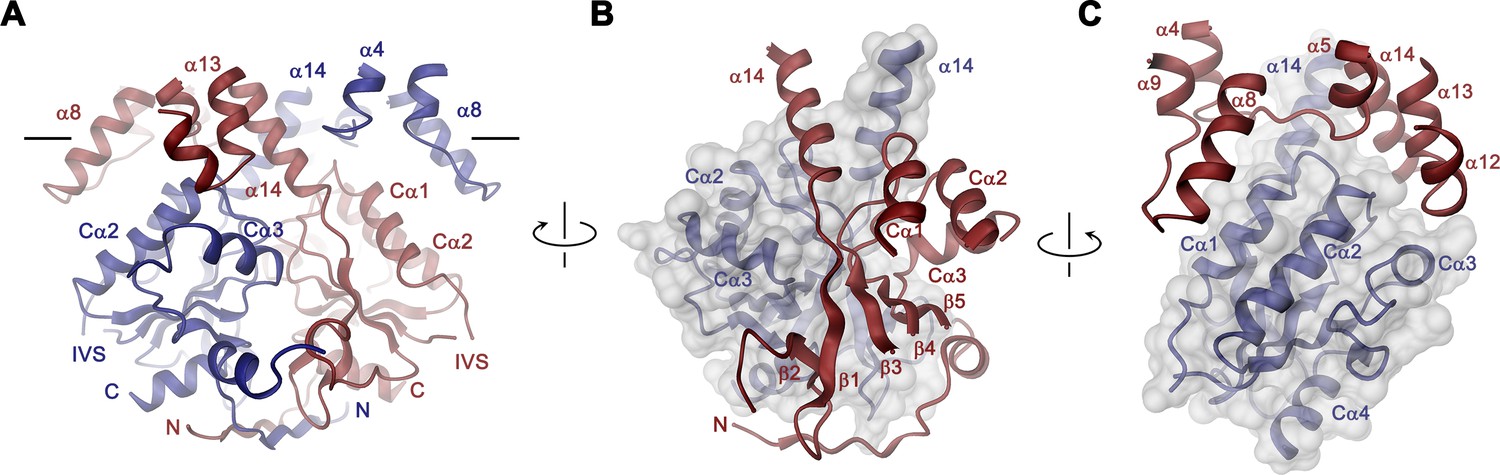
STAS domain and dimer interface.
(A) Ribbon representation of the STAS domain dimer and interacting parts of the N-terminus and the TMD. The view is as in Figure 2B. (B) Elements of mutual STAS domain interactions and contacts with the N-terminus. (C) STAS domain-TMD interactions. B and C, Molecular surface of the STAS domain and the C-terminal part of α14 is shown superimposed on one subunit (blue), segments of the interacting subunit are shown in red. The relationship between views is indicated. A-C, The intracellular membrane boundary is indicated.
The transmembrane domain
The overall architecture of the TMD conforms to the 7 + 7 inverted repeat topology (Figure 1—figure supplement 1 and Figure 4—figure supplement 1), which was previously observed in the structure of the prokaryotic homologue SLC26Dg (Geertsma et al., 2015) and in members of the distantly related SLC4 (Arakawa et al., 2015; Huynh et al., 2018) and SLC23 (Alguel et al., 2016; Lu et al., 2011) families. Consistent with the known features of this arrangement, the Slc26a9T TMD is composed of 14 membrane-inserted α-helices, with α8–14 representing a similar yet inverted organization to α1–7 (Figure 4—figure supplement 1). The TMD is segregated into two subdomains, a convex core module (consisting of α1–4, and 9–11) carrying a putative ion-binding site, and a concave gate module (consisting of α5–7 and 12–14) (Figure 4, Video 3). Both modules constitute independent structural units that interact via an extended interface (Figure 4A). Although displaying channel-like functional properties, the structure of the TMD of Slc26a9T shows hallmarks of a transporter. The striking degree of similarity revealed from a superposition with its equivalent unit of SLC26Dg (RMSD of 2.0 Å) emphasizes the high conservation within the family and suggests that both structures display equivalent conformations (Figure 5A–C). The large cavity between gate and core modules is accessible from the intracellular side but does not extend into a contiguous pore region towards the extracellular side, suggesting that the Slc26a9T structure represents an inward-open state of the protein that facilitates transport by an alternate-access mechanism. This assumption is further supported by the comparison of TMDs of Slc26a9T from cryo-EM maps acquired either in detergent micelles or lipid nanodiscs (Figure 2—figure supplement 4C–E). The nanodisc TMD structure differs in the conformation of the core module, which has rotated as rigid body by approximately 14° and moved by 4 Å towards the extracellular side, whereas the remainder of the protein appears largely unaltered. This conformation thus likely shows an intermediate state of the protein on its transition towards an outward-facing state (Figure 5D; Video 4).
Figure 4 with 1 supplement see all
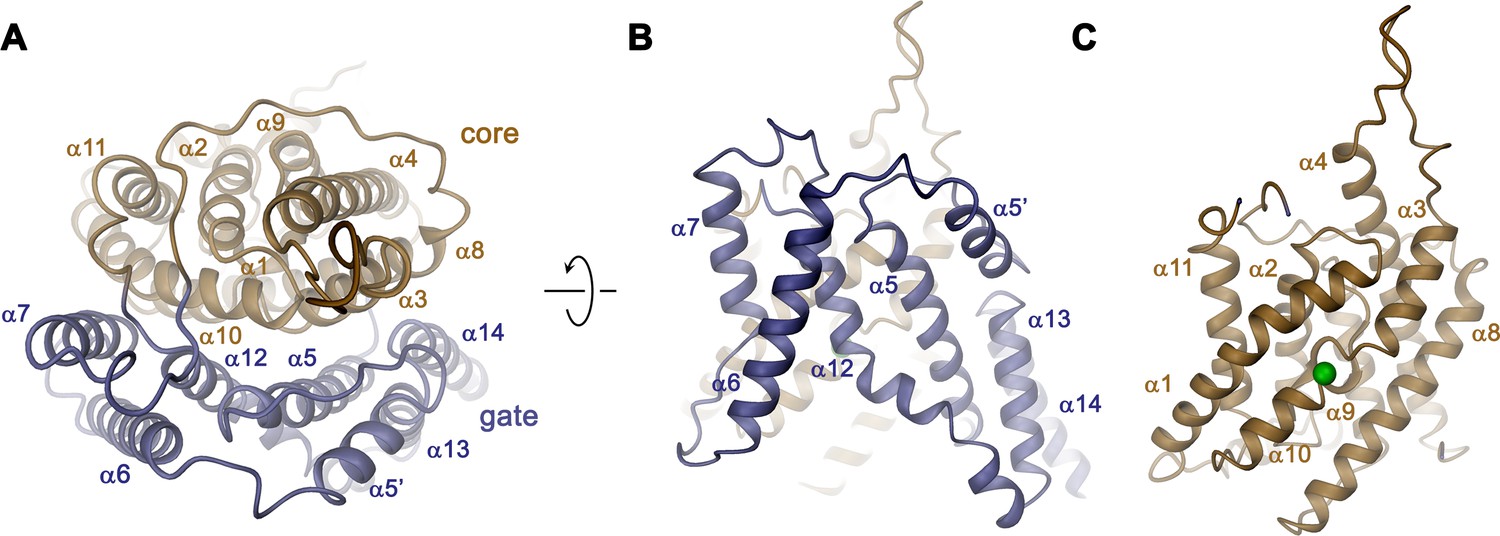
Transmembrane domain.
(A) Ribbon representation of the TMD of a single subunit of Slc26a9T viewed from the extracellular side. (B) View of the gate and (C) the core module from within the membrane. Green sphere indicates predicted position of bound Cl–.
Figure 5
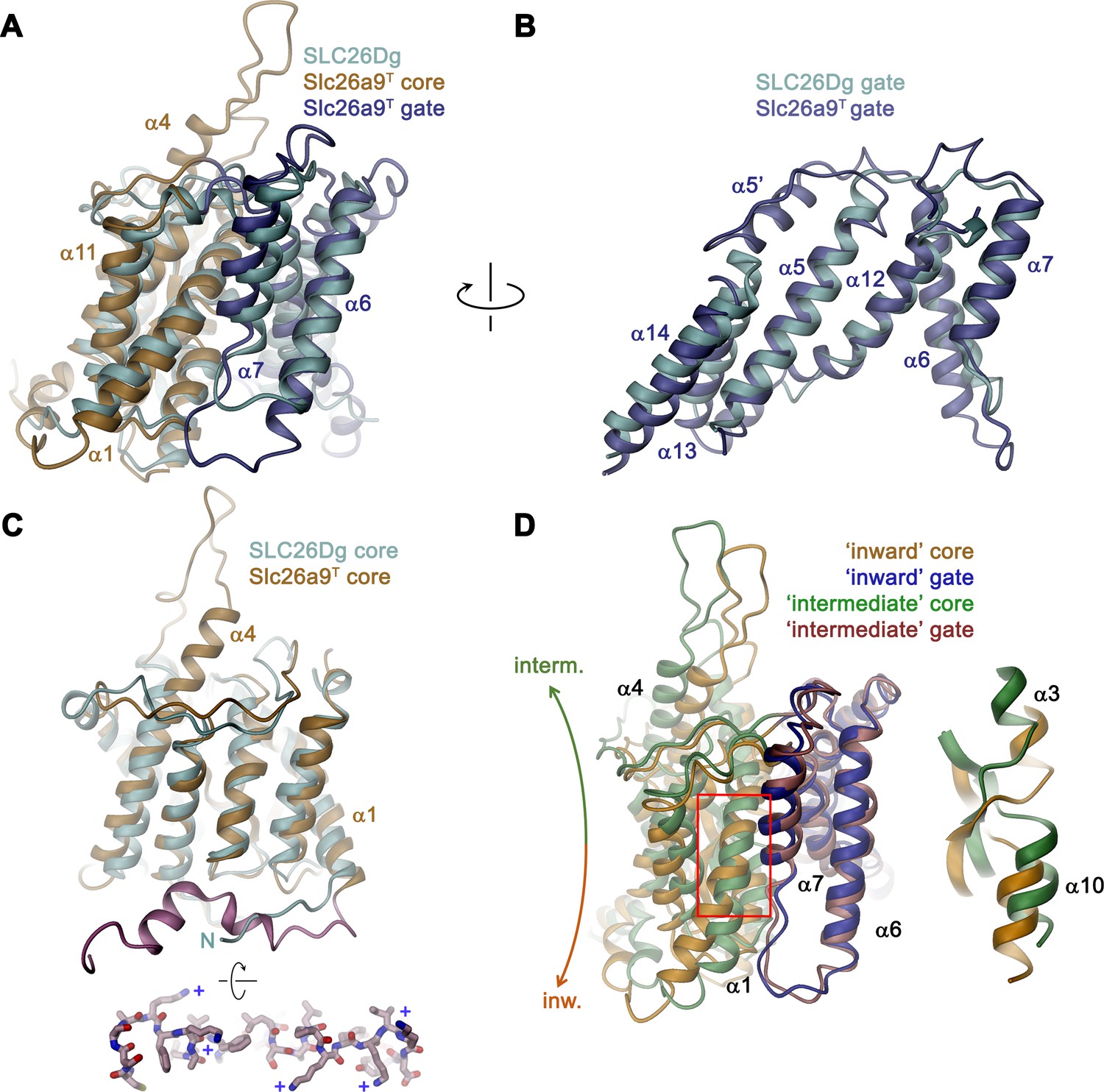
Comparison of the TMD.
(A) Structural superposition of the TMDs of Slc26a9T (beige and blue) and SLC26Dg (PDBID: 5DA0, cyan). (B) Superposition of the gate domains and (C) core domains of Slc26a9T and SLC26Dg. N-terminal region of Slc26a9T preceding α1 is colored in pink and displayed as sticks from an intracellular perspective below. The location of basic residues is indicated (+). (D) Superposition of the TMD of detergent-solubilized Slc26a9T, showing an inward-facing state onto the TMD of lipid nanodisc-reconstituted Slc26a9T, indicating an intermediate state on its transition towards the outside. The movement of the core domains is indicated. Inset (right) shows blow-up of the region around the Cl–-binding site (indicated by red box). The core and gate modules of the intermediate conformation of Slc26a9T determined in nanodiscs are colored in green and pink, respectively.
Video 4
Conformational transition from the inward-facing to the intermediate state.
Ribbon representation of the transmembrane domain as a morph between the inward-facing and intermediate states. The structure is viewed from the peripheral. Residues 276 to 312 are removed for clarity. The putative trajectory of the chloride ion is also shown.
Despite the general similarity between the TMDs of Slc26a9T and SLC26Dg, there are several structural features that distinguish the eukaryotic transporter form its prokaryotic homologue. First, a loop-helix structure connecting α5-α6 (α5’) fills a depression in the structure created by the short α13–α14 turn, which would not otherwise reach the boundary of the outer leaflet of an undistorted bilayer (Figures 4B and 5B). A second distinguishing feature of Slc26a9T concerns the loop region connecting α3 and the extended α4, which protrudes into the extracellular space and which contains two predicted glycosylation sites (Li et al., 2014) near the tip of the spiky extension (Figures 4C and 5C). Finally, the extended N-terminus of Slc26a9T, preceding α1, is significantly longer than the equivalent region in SLC26Dg (Figure 5C). In the mammalian transporter, the N-terminus is partly mobile and contains extended regions with interspersed secondary structure elements. Whereas the initial part of the N-terminus engages in the earlier described interaction with the STAS domains, a mobile linker bridges this region to a structured part preceding α1, which forms scattered helical segments peripherally contacting the inner membrane leaflet and containing several basic residues (Figure 5C, bottom inset). Due to the high sequence conservation between paralogs (Figure 1—figure supplement 1A), all three structural features are likely preserved in mammalian SLC26 transporters.
Structural and functional properties of the ion transport pathway
In the observed inward-facing conformation of the TMD, transported anions enter the protein to access their binding site via a wide aqueous vestibule, presumably attracted by the positive electrostatic potential that is conferred by an excess of basic amino acids at its intracellular entrance (Figure 6A,B). The importance of these basic residues for transport is illustrated in the comparison of conductance properties of Slc26a9T and constructs wherein selected positively charged residues at the intracellular entrance were mutated to glutamate. For wild-type (WT) Slc26a9T, the linear I-V relationship is compatible with symmetric rate-limiting barriers on the transport cycle in the absence of applied voltage (Lam and Dutzler, 2018; Paulino et al., 2017). This linear relation is minimally perturbed upon introducing the mutations R205E or K358E, which are positioned at the outermost regions of the intracellular vestibule (Figure 6A; Figure 6—figure supplement 1A). In contrast, mutations K270E and K443E, located deeper in the intracellular cavity and close to the presumed anion binding site, both altered the electrostatics and displayed considerable outward rectification (Figure 6A–C). This behavior indicates a relative increase of the inward barrier by either impeding the access of anions from the intracellular side or alternatively by hampering conformational transitions of the protein. Complimentary to the intracellular charge-reversal mutations, we find a reciprocal effect leading to inward rectification upon mutation of extracellular basic residues K221 and K431 that are predicted to line the aqueous cavity to the binding site in an outward-facing conformation (Figure 6—figure supplement 1B,C).
Figure 6 with 1 supplement see all
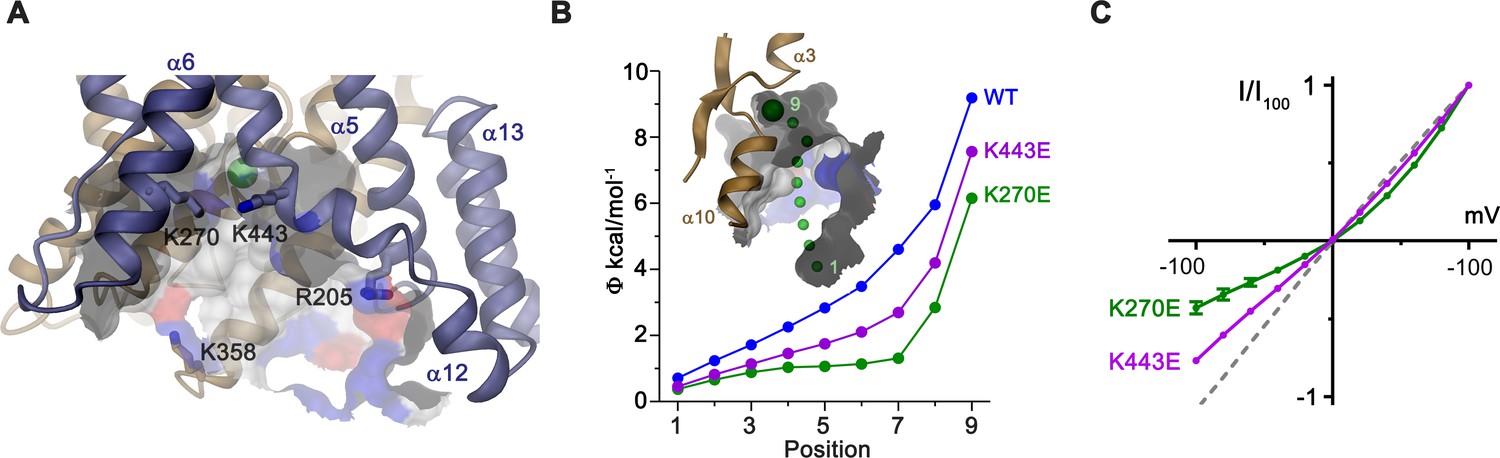
Electrostatic properties of the intracellular cavity.
(A) View of the intracellular cavity leading to the Cl–-binding site. Section of the molecular surface is shown with contact region of acidic residues colored in red and basic residues in blue. (B) Electrostatic potential within the aqueous cavity leading to the Cl–-binding site of WT and charge reversing mutants of basic residues facing the cavity. Inset shows aqueous access path of ions. Positions sampling the electrostatic potential are shown as green spheres. A, B, Bound Cl– is shown as a large green sphere. (C) I-V relationships of charge-reversing mutants K270E and K443E facing the aqueous cavity (K270E, n = 3; K443E, n = 4). Data was recorded from excised patches in symmetric 150 mM Cl–.
Although in our cryo-EM reconstructions we did not observe any density attributable to bound Cl–, its absence is concordant with its measured low apparent affinity. We infer an anion binding site to be located in a pocket of the core domain between the N-termini of the membrane-inserted partial helices α3 and α10 (Figures 4C and 7A; Video 5). The equivalent location was proposed as a substrate-binding site for the fumarate-proton symporter SLC26Dg (Chang et al., 2019a) and the chloride-bicarbonate transporter SLC4A1/Band 3 (Arakawa et al., 2015) and confirmed for the SLC23 transporters UraA (Lu et al., 2011) and UapA (Alguel et al., 2016). This site would be ideally positioned at the apex of the inward-open cavity, with its dual pseudo-symmetrically related helix dipoles arranged in a manner frequently observed for binding sites of anion transporters (Dutzler et al., 2002; Screpanti and Hunte, 2007) (Figures 4C and 7A). Compared to SLC26Dg, the pocket of Slc26a9T is considerably smaller, as a consequence of the conformation of F128 projecting its side-chain into the pocket, thus creating a site of appropriate size for a Cl– ion (Figure 7A). In this site, several residues could plausibly interact with the transported substrates. Besides F128, these include the nonpolar residues F92 and L391, all contributing to the partly hydrophobic character frequently found in anion binding sites which likely account for the observed lyotropic permeability sequence of anions (Figure 1E) (Dani et al., 1983; Dutzler et al., 2002). Dehydrated anions could be stabilized by side-chain interactions with Q88, S392, and possibly T127, as well as main-chain interactions at the N-terminus of α10, which together form a cradle to coordinate the desolvated ion (Figure 7A). To probe the functional importance of potential binding-site residues for anion interactions, we individually mutated them to alanine and used inside-out patch recordings to assess any corresponding changes in transport behavior (Figure 7—figure supplement 1A–G). Several mutants altered the linear I-V relationship, with the resulting inward rectification of the mutant S392A indicating a relative increase of the outer barrier and conversely, the outward rectification in the mutants F128A and Q88A, a relative increase of the inner barrier during transport (Figure 7B, Figure 7—figure supplement 1A,D,F). In contrast, only minor perturbations of the linear I–V relationship are found for the binding site residues F92A and L391A and no change is observed for the mutant N441A residing on the gate module, which, although proximal to the site, is not predicted to participate in ion coordination (Figure 7—figure supplement 1B,E,G). The change in the conductance properties upon mutation of Gln 88 is even more pronounced in the mutant Q88E, which replaces the amino acid conserved in mammalian paralogs with a negatively charged residue found in several prokaryotic transporters (Geertsma et al., 2015). Despite the strong apparent surface expression and high stability of the mutant, there are no currents observed in excised patches and severely decreased currents in whole-cell measurements are strongly outward-rectifying, consistent with a compromised transport activity due to the interference of the negatively charged residue with the transported anions (Figure 7—figure supplement 1H,I). A striking property of Slc26a9T is the inverted relationship between permeability and conductivity of transported ions, with larger anions being more permeable but less conductive than Cl– (Figure 1D and E). Although affected in all binding-site mutants (Figure 7—figure supplements 2 and 3A–E; Table 2), changes are most pronounced for S392A and F128A, where chloride becomes less conductive than other permeant anions, a behavior that is also reflected in the kinetic parameters obtained from the concentration dependence of transport of Cl– and SCN– (Figure 7C–H; Figure 7—figure supplement 3; Table 2). A pronounced difference is also observed for the mutant L391A where the Km of SCN– increases by nearly two orders of magnitude (Figure 7—figure supplement 3I; Table 2). Whereas the strong effect of different binding-site mutants on conduction underlines their importance for anion transport, in no case did we detect major increases in permeation by either HCO3– or SO42–, nor did we observe any evidence of coupled transport (Figure 7—figure supplements 1A–G and 2; Table 2).
Figure 7 with 3 supplements see all
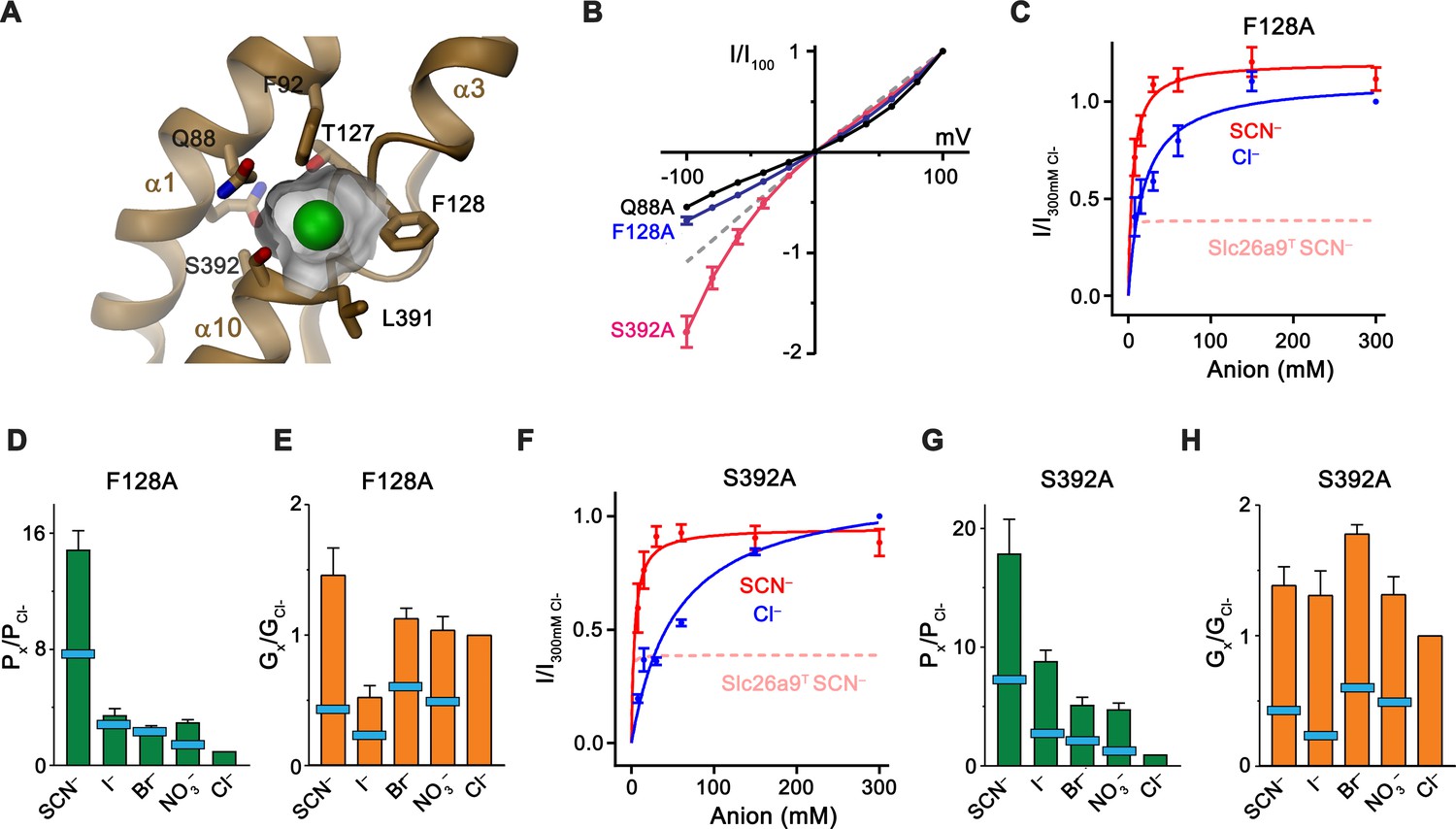
Substrate binding site.
(A) Structure of the Cl– binding site. The molecular surface of the binding pocket is shown. Selected residues are displayed as sticks. Bound Cl– is shown as a green sphere. (B) I-V relationships of selected Cl– binding site mutants (Q88A, n = 5; F128A, n = 8; S392A, n = 5). Data for WT Slc26a9T is shown as dashed line for comparison. (C) Conductance-concentration relationships of anion transport across the Slc26a9T mutant F128A (n = 3). For comparison, SCN– conductance–concentration relationship for non-mutated Slc26a9T is shown as a pink dotted line. (D) Permeability (Px/PCl) and (E) conductance ratios (Gx/GCl) of the Slc26a9T mutant F128A obtained from bi-ionic substitution experiments (Cl–, Br–, NO3–, SCN–, n = 8; I–, n = 3). The light-blue bars indicate corresponding values for WT Slc26a9T. (F) Conductance-concentration relationships of anion transport across the Slc26a9T mutant S392A (n = 5). For comparison, SCN– conductance–concentration relationship for non-mutated Slc26a9T is shown as a pink dotted line. (G) Permeability (Px/PCl) and (H) conductance ratios (Gx/GCl) of the Slc26a9T mutant S392A obtained from bi-ionic substitution experiments (Cl–, Br–, n = 5; NO3–, SCN–, I–, n = 4). The light-blue bars indicate corresponding values for WT Slc26a9T. Data was recorded from excised patches either in symmetric 150 mM Cl– (B), the indicated intracellular anion-concentrations with 7.5 mM extracellular Cl– (C and F), or at equimolar bi-ionic conditions containing 150 mM extracellular Cl– and 150 mM of the indicated intracellular anion (D, E, G, H). Data show mean values of the indicated number of biological replicates, errors are s.e.m.. C-H, Data were recorded at −100 mV.
Table 2
Transport properties of WT and mutated Slc26a9T.
https://doi.org/10.7554/eLife.46986.029| WT* | Q88A | F92A | T127A | F128A | L391A | S392A | N441A | ||
|---|---|---|---|---|---|---|---|---|---|
| SCN– | Erev (mV)† Px/PCl Gx/GCl | 50.5 7.5 0.43 | 58.4 10.6 0.55 | 37.8 4.5 0.44 | 59.3 10.7 0.80 | 68.7 14.9 1.46 | 61.5 12.0 1.10 | 71.5 18.0 1.39 | 52.9 8.2 0.57 |
| I– | Erev (mV) Px/PCl Gx/GCl | 26.6 2.9 0.23 | 16.3 1.9 0.14 | 14.1 1.8 0.25 | 9.5 1.5 0.17 | 31.0 3.5 0.53 | 23.5 2.6 0.54 | 53.2 8.4 1.31 | 29.1 3.2 0.35 |
| Br– | Erev (mV) Px/PCl Gx/GCl | 21.3 2.4 0.60 | 3.7 1.2 0.32 | 5.1 1.2 0.72 | 7.6 1.4 0.40 | 24.6 2.7 1.13 | 16.1 1.9 1.11 | 40.7 5.2 1.78 | 20.0 2.2 0.70 |
| NO3– | Erev (mV) Px/PCl Gx/GCl | 9.4 1.5 0.49 | 32.6 3.7 1.20 | 11.2 1.6 0.52 | 29.0 3.2 1.46 | 28.5 3.1 1.04 | 20.7 2.3 0.70 | 39.1 4.8 1.31 | 16.9 2.0 0.77 |
| Cl– | Erev (mV) Px/PCl Gx/GCl | −0.1 1 1 | −1.4 1 1 | −1.5 1 1 | −0.5 1 1 | 0 1 1 | −0.7 1 1 | 0.3 1 1 | −0.3 1 1 |
| F– | Erev (mV) Px/PCl Gx/GCl | −81.1 .05 nd¶ | −64.2 .08 nd | −81.0 .04 nd | −74.5 .07 nd | −67.6 .07 nd | −81.0 .05 nd | −58.8 0.11 nd | −86.7 0.03 nd |
| HCO3– | Erev (mV) Px/PCl Gx/GCl | −81.7 0.05 nd | −54.4 0.12 nd | −69.5 0.07 nd | −84.7 0.04 nd | −47.3 0.16 nd | −93.1 0.03 nd | −57.4 0.10 nd | −76.0 0.05 nd |
| SO42– | Erev (mV) Px/PCl Gx/GCl | −96.9 0.01 nd | −102.2 0.01 nd | −78.0 0.01 nd | −104.5 0 nd | −70.5 0.02 nd | −93.6 0.01 nd | −71.2 0.02 nd | −82.4 0.01 nd |
| E30mM Cl‡ | −37.7 | −39.3 | −38.5 | −39.8 | −42.1 | −37.9 | −39.0 | −38.3 | |
| Km,Cl (mM) § | 29.5 | 2.3 | 19.2 | 4.7 | 18.6 | 44.0 | 54.1 | 31.3 | |
| Km,SCN (mM) § | 0.5 | 0.2 | 0.8 | 1.6 | 5.2 | 29.4 | 3.8 | 0.7 | |
| VSCN/VCl § | 0.44 | 0.63 | 0.44 | 0.74 | 1.08 | 1.08 | 0.83 | 0.68 |
-
*WT refers to non-mutated Slc26a9T.
†Erev values, Px/PCl, and Gx/GCl were derived from bi-ionic data in Figure 1—figure supplement 3B–F, and Figure 7—figure supplement 2.
-
‡Measured reversal potential, in mV, under a 5-fold NaCl gradient (ECl = –40.2 mV). Extracted from data in Figure 1C and Figure 7—figure supplement 1A–H.
§Km and VSCN/VCl values are derived from conductance–concentration data presented in Figure 1D, Figure 7, and Figure 7—figure supplement 3F–J.
-
¶nd, not determined.
Video 5
Structure of the presumed ion binding site.
Ribbon representation of the anion binding site with side-chains of interacting residues displayed as sticks. Surface of the binding pocket around a modeled chloride (green sphere) is shown. View is as in Figure 7A.
Discussion
Our study has revealed the architecture of the murine Cl– transport protein Slc26a9 and it has provided the first direct glimpse at the relationship between structure and function in a mammalian SLC26 family member. Owing to the conservation between paralogs, the observed structural features are likely general for the family. The dimeric organization of Slc26a9 differs markedly from structurally related transport proteins of the SLC4 and SLC23 families, which interact via an extended interface between gate modules (Alguel et al., 2016; Arakawa et al., 2015; Chang and Geertsma, 2017; Huynh et al., 2018; Yu et al., 2017) (Figure 8—figure supplement 1). In contrast, in Slc26a9 the interface between the TMDs is minimal whereas the bulk of subunit interactions are mediated by the cytoplasmic STAS domains involving contacts with the N-terminus and intracellular loop regions of the gate module (Figure 2). A similar relationship between transmembrane domains was recently proposed to underlie dimerization of the prokaryotic transporter SLC26Dg in a membrane environment, based on EPR-derived distance constraints (Chang et al., 2019b). Although such minimal contacts in the membrane-inserted domain are unusual for oligomeric membrane proteins, they have been recently observed for the mechanosensitive channel OSCA (Jojoa-Cruz et al., 2018; Zhang et al., 2018), where subunit interactions are mediated exclusively between residues of the cytoplasmic domains.
As previously shown based on whole-cell experiments (Bertrand et al., 2009; Chang et al., 2009b; Dorwart et al., 2007; Loriol et al., 2008) and confirmed here with excised patches, Slc26a9 shows large uncoupled macroscopic Cl– currents that saturate only at high mM concentration. For this reason, the protein has been generally referred to as ion channel. However, such properties would be entirely consistent with a fast bidirectional uniport mechanism as evidenced by other structural and functional data shown in our study. The high Km of Cl– matches its intracellular concentration and would foster high turnover leading to the observed conduction phenotype. Moreover, transport of the hydrophobic ion SCN– saturates at lower concentration consistent with a mechanism that is mediated by an anion binding site, which would be expected in a transporter. Additionally, our structural data clearly suggest that Slc26a9 works by an alternate-access mechanism akin to other family members. This is evidenced by a comparison between the inward-facing structure of Slc26a9T determined in detergent, where the ion binding site is accessible to the cytoplasm, and an intermediate conformation of the lipid nanodisc-reconstituted transporter, where an upward movement of the core module has caused the binding site to be buried between core and gate modules (Figure 8A,B). In combination, both structures provide insight into conformational changes in which the relatively unconstrained core module, providing the substrate binding site, makes ascending and descending rigid-body movements relative to the immobilized gate module-STAS domain complex, in a process that resembles the elevator transport mechanism (Drew and Boudker, 2016; Ficici et al., 2017; Reyes et al., 2009) (Figure 8B). Such mechanism would allow for fast transport kinetics as observed here and as previously described for the Cl–/HCO3– transporter SLC4A1/Band 3, which shares a similar architecture (Arakawa et al., 2015; Brahm, 1977). The plausible homology to the known outward-facing structure of SLC4A1/Band 3 (Arakawa et al., 2015) also permits the anticipation of the possible extent of conformational changes, where basic residues on the rim of the resulting extracellular vestibule likely create a positive electrostatic environment which is similar to that observed in the vestibule of the inward-facing state. The role of electrostatics in this hypothesis is supported by functional experiments and model-based calculations (Figure 6A–C; Figure 6—figure supplement 1). During transport, the negatively charged Cl– is dragged across the transmembrane electric field, thus likely facilitating conformational changes. A similar mechanism may underlie the substrate-conferred electromotility observed in the homologous motor protein Prestin (Oliver et al., 2001).
Figure 8 with 3 supplements see all
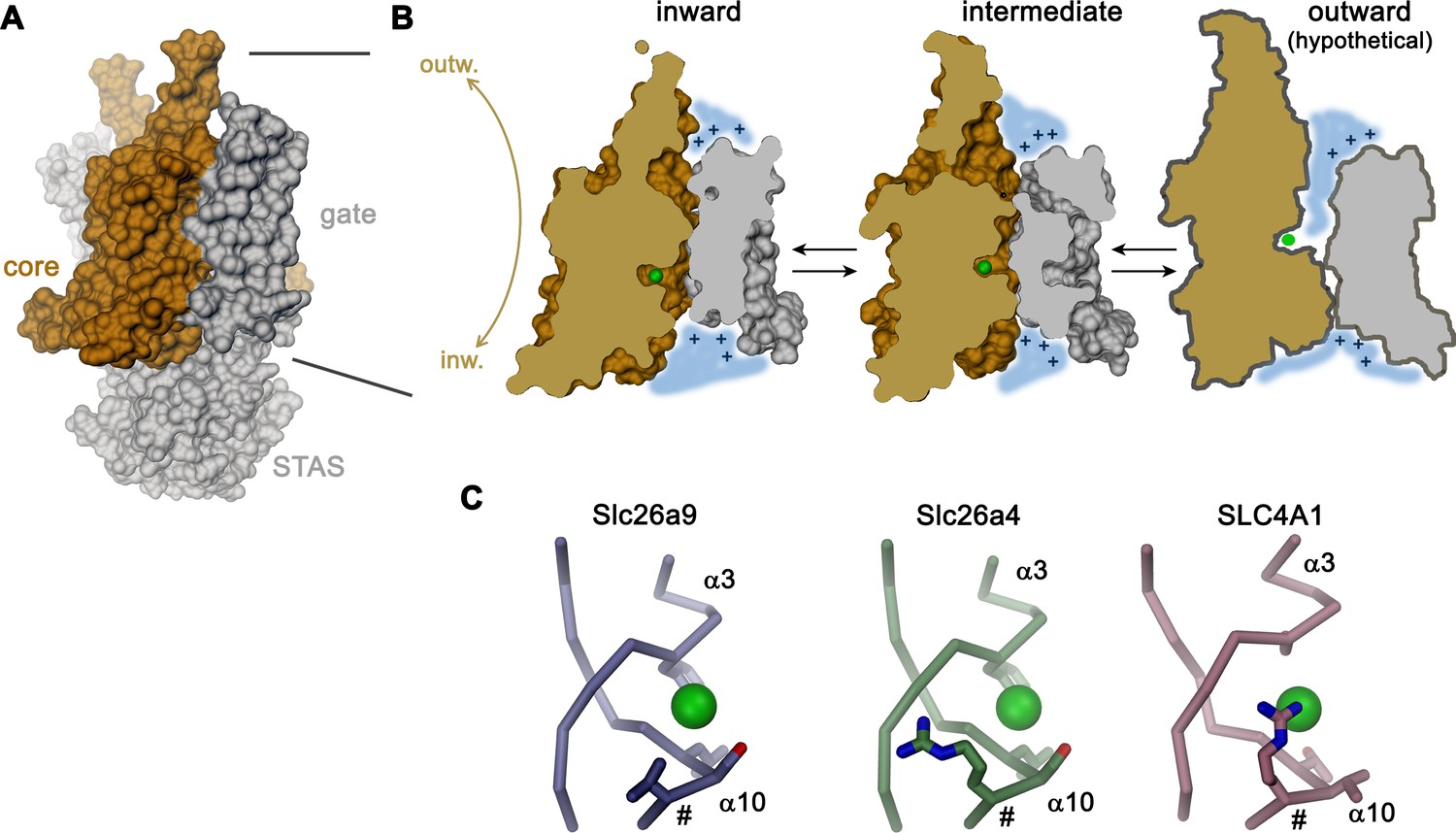
Transport mechanism.
(A) Molecular surface of Slc26a9T viewed towards the long dimension of the molecule. (B) Sections of the TMD in the inward-facing conformation defined by the Slc26a9T detergent structure (left), an intermediate conformation defined by the Slc26a9T nanodisc structure (center) and a schematic drawing of a potential outward-facing state (right). (C) Putative Cl–-binding site of Slc26a9 (left), a homology model of Slc26a4 (Pendrin, center) and SLC4A1/Band 3 (PDBID: 4YZF). A position that contains a basic residue in SLC26A4 and all mammalian SLC26 paralogs except for Slc26a9 is labeled (#). A and B, STAS domain and gate modules are colored in gray, the core module in beige. A-C, The putative position of a bound Cl– is indicated as green sphere.
A characteristic feature of Slc26a9 compared to many of its paralogs of the SLC26 family is its distinct anion selectivity and transport mechanism. Unlike several other family members, which are proposed to work as coupled exchangers, transport in Slc26a9 is not coupled to another ion (Figure 1C). Moreover, whereas other family members are capable of transporting the divalent anion SO42– or the ubiquitous monovalent HCO3–, neither of these anions is a substrate for electrogenic transport by Slc26a9. The lyotropic permeability sequence of Slc26a9 suggests that ions interact with a low field-strength binding site. The inability to transport SO42– is thus probably related to the fact that interactions within the binding site, which does not contain positive countercharges to balance the negatively-charged substrate, may not be sufficiently strong for the coordination of divalent anions. Remarkably, all other mammalian family members harbor very similar predicted binding sites to that of Slc26a9, with the notable exception that they additionally contain a positively charged amino acid at the equivalent position of V393, which is located on the N-terminal end of helix α10 (Figure 8C; Figure 8—figure supplement 2). Homology models predict this side-chain to reside very close to the substrate binding site (Figure 8—figure supplement 2), and an arginine at the equivalent location, which likely contributes to anion interactions, is also found in the Cl–/HCO3– transporter SLC4A1/Band 3 (Arakawa et al., 2015) (Figure 8C). The inability of Slc26a9 to transport HCO3– is particularly remarkable, since the nearly isosteric molecular anion NO3– is a transported substrate (Figure 1E). Although the pKa for the deprotonation of HCO3– is high (i.e. 10.3) and consequently the concentration of the divalent anion CO32– is negligible under physiological conditions, the equilibrium could be perturbed at the binding site, with the large positive electrostatic potential within the aqueous cavity stabilizing the divalent state of the ion, for which interactions with the anion binding site would not be sufficiently strong to make it a transported substrate.
The high homology between mammalian SLC26 paralogs suggests a common general architecture and also implies that other family members may undergo similar conformational transitions to those shown here for Slc26a9T. Therefore, the diversity of functional phenotypes ranging from uncoupled bidirectional uniport as observed in Slc26a9 (and possibly SLC26A7 and/or SLC26A11), to coupled exchangers such as SLC26A2/DTDST and SLC26A4/Pendrin, or even to the motor activity of SLC26A5/Prestin (Alper and Sharma, 2013), could rely on comparatively small localized structural differences which in turn impose variable energy barriers for conformational changes (Figure 8—figure supplement 3). Uncoupled uniporters like Slc26a9 presumably possess small conformational energy barriers between inward and outward states in the presence or absence of substrate which would permit transition of an unloaded transporter. In contrast, for coupled transport the presumed increase in ion binding affinity paired with an enlarged barrier prohibiting conformational transitions of the unloaded transport domain would itself lead to stoichiometric exchange. In this view, the electromotile motor function of SLC26A5/Prestin could be a consequence of an insurmountable energy barrier preventing the last phase of the transition to an outward-facing state, in line with prior suggestions of SLC26A5/Prestin exhibiting an ‘incomplete’ transport cycle (Schaechinger and Oliver, 2007). While it is currently not possible to pinpoint exact structural determinants of different SLC26 mechanisms, the architectural elements identified here provide a baseline for further studies of SLC26 structure-function relationships.
Since our electrophysiology experiments show instantaneous activity of the transporter that is not obviously dependent on activating ligands (Figure 1B), the question concerning the regulation of Slc26a9 activity under physiological conditions is pertinent. This question is particularly important in light of a potential role of human SLC26A9 to compensate for the loss of CFTR mediated Cl– transport in patients suffering from cystic fibrosis (Balázs and Mall, 2018; Mall and Galietta, 2015). A potential activation mechanism could be the mobilization of SLC26A9 from intracellular stores, which might be regulated by phosphorylation, as suggested in a previous study (Dorwart et al., 2007) and which is also consistent with the predominantly intracellular localization of full-length Slc26a9 observed in our study (Figure 1—figure supplement 2A). The fact that removal of the IVS results in significantly greater surface expression suggests that this region may harbor modifiable motif elements which can be involved in surface-trafficking regulation, but this hypothesis must be further evaluated in more physiologically relevant contexts. Another, still unresolved question concerns the precise role of the IVS and the CT in the interaction with accessory proteins including the chloride channel CFTR. The resolution of these questions will be subject of future studies, for which our current work provides a structural and functional foundation.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifier | Additional information |
|---|---|---|---|---|
| Chemical compound, drug | Pro293S-CDM medium | Lonza | Cat#BE02-025Q | |
| Chemical compound, drug | HyClone HyCell TransFx-H medium | GE Healthcare | Cat#SH30939.02 | |
| Chemical compound, drug | L-glutamine | Millipore Sigma | Cat#G7513 | |
| Chemical compound, drug | Penicillin-streptomycin | Millipore Sigma | Cat#P0781 | |
| Chemical compound, drug | Fetal bovine serum | Millipore Sigma | Cat#F7524 | |
| Chemical compound, drug | Pluronic F-68 | ThermoFisher Scientific | Cat#24040032 | |
| Chemical compound, drug | Polyethylenimine 25 K MW, linear | Polysciences | Cat#23966–1 | |
| Chemical compound, drug | Dulbecco’s Modified Eagle’s Medium (DMEM) | Millipore Sigma | Cat#D5546 | |
| Chemical compound, drug | Valproic acid | Millipore Sigma | Cat#P4543 | |
| Chemical compound, drug | cOmplete, EDTA-free Protease Inhibitor Cocktail | Roche | Cat#5056489001 | |
| Chemical compound, drug | Digitonin | AppliChem | Cat#A1905 | |
| Chemical compound, drug | Glyco-diosgenin | Anatrace | Cat#GDN101 | |
| Chemical compound, drug | D-desthiobiotin | Millipore Sigma | Cat#D1411 | |
| Chemical compound, drug | n-dodecyl-β-D-maltoside (DDM) | Anatrace | Cat#D310 | |
| Chemical compound, drug | Cholesteryl hemisuccinate, tris salt (CHS) | Anatrace | Cat#CH210 | |
| Chemical compound, drug | 1-palmitoyl-2-oleoyl-sn-glycero-3-phosphoethanolamine (POPE) | Avanti Polar Lipids, Inc | Cat#850757 | |
| Chemical compound, drug | 1-palmitoyl-2-oleoyl-sn-glycero-3-phospho-(1'-rac-glycerol) (POPG) | Avanti Polar Lipids, Inc | Cat#840457 | |
| Chemical compound, drug | Diethyl ether | Millipore Sigma | Cat#296082 | |
| Chemical compound, drug | Triton X-100 | Millipore Sigma | Cat#T9284 | |
| Chemical compound, drug | Biotin | Millipore Sigma | Cat#B4501 | |
| Chemical compound, drug | 9-amino-6-chloro-2-methoxyacridine (ACMA) | Thermo Fisher Scientific | Cat#A1324 | |
| Chemical compound, drug | carbonyl cyanide 3-chloropheny lhydrazone (CCCP) | Merck Millipore | Cat#C2759 | |
| Chemical compound, drug | 4,4’-Diisothiocyanatostilbene-2,2’-disulfonic acid (DIDS) | Millipore Sigma | Cat#D3514 | |
| Commercial assay or kit | QuikChange site-directed mutagenesis kit | Agilent | Cat#200523 | |
| Commercial assay or kit | NucleoBond Xtra Maxi kit | Macherey-Nagel | Cat#740416 | |
| Commercial assay or kit | StrepTactin Superflow affinity resin slurry | IBA Lifesciences | Cat#2-1206-002 | |
| Commercial assay or kit | Superose 6 10/300 GL | GE Healthcare | Cat#17-5172-01 | |
| Commercial assay or kit | Zorbax GF-450 | Agilent | Cat#884973–902 | |
| Commercial assay or kit | Superose 6 5/150 | GE Healthcare | Cat#29091597 | |
| Commercial assay or kit | Pierce Streptavidin Plus UltraLink Resin | Thermo Fisher Scientific | Cat#53117 | |
| Commercial assay or kit | Bio-Beads SM-2 | Bio-Rad | Cat# 1523920 | |
| Commercial assay or kit | Avestin LiposoFast Liposome Factory Basic | Millipore Sigma | Cat#Z373400 | |
| Commercial assay or kit | 400 nm polycarbonate filters for LiposoFast | Millipore Sigma | Cat#Z373435 | |
| Commercial assay or kit | 96-well black-walled microplate | Thermo Fisher Scientific | Cat#M33089 | |
| Commercial assay or kit | 200 mesh Au 1.2/1.3 cryo-EM grids | Quantifoil | Cat#N1-C14nAu20-01 | |
| Commercial assay or kit | Amicon 100 kDa MWCO centrifugal filter | EMD Millipore | Cat#UFC910008 | |
| Commercial assay or kit | 0.22 µm Ultrafree-MC Centrifugal Filter | EMD Millipore | Cat#UFC30GV | |
| Commercial assay or kit | Borosilicate glass capillary with filament | Sutter Instrument | Cat#BF150-86-10HP | |
| Cell line (human) | HEK293S GnTI- | ATCC | CRL-3022 | |
| Cell line (human) | HEK-293T | ATCC | CRL-1573 | |
| Recombinant DNA | Mus musculus Slc26a9 ORF shuttle clone | Source BioScience | ORFeome# OCACo5052B0115D; GenBank BC160193 | |
| Recombinant DNA | pcDNA 3.1 (+)vector, Invitrogen | Thermo Fisher Scientific | Cat# V79020 | |
| Recombinant DNA | Modified pcDNA 3.1 vector with C-terminal 3C protease cleavage site, Venus and Myc tags and streptavidin binding peptide | Raimund Dutzler laboratory | N/A | |
| Recombinant DNA | Modified pcDNA 3.1 vector with C-terminal 3C protease cleavage site, Myc tag and streptavidin binding peptide | Raimund Dutzler laboratory | N/A | |
| Recombinant DNA | Expression vector encoding membrane scaffold protein (MSP) E3D1, pMSP1E3D1 | Denisov et al., 2007 | Addgene, Cat#20066 | |
| Software,algorithm | SerialEM 3.5.0 | Mastronarde, 2005 | http://bio3d.colorado.edu/SerialEM/ | |
| Software, algorithm | RELION-3.0 | Scheres, 2012 | https://www2.mrc-lmb.cam.ac.uk/relion/ | |
| Software, algorithm | CTFFIND4.1 | Rohou and Grigorieff, 2015 | http://grigoriefflab.jan elia.org/ctf | |
| Software, algorithm | Bsoft 1.9.5 | Heymann and Belnap, 2007 | https://lsbr.niams.nih.gov/bsoft/ | |
| Software, algorithm | Coot 0.8.8 | Emsley and Cowtan, 2004 | https://www2.mrc-lmb.cam.ac.uk/person al/pemsley/coot/ | |
| Software, algorithm | PHENIX 1.14 | Adams et al., 2002 | http://phenix-online.org/ | |
| Software, algorithm | REFMAC5 | Murshudov et al., 2011 | http://www.ccpem.ac.uk/ | |
| Software, algorithm | MSMS | Sanner et al., 1996 | http://mgltools.scripps.edu/packages/MSMS/ | |
| Software, algorithm | DINO 0.9.4 | http://www.dino3d.org | http://www.dino3d.org | |
| Software, algorithm | PyMOL 2.3.0 | DeLano, 2002 | https://pymol.org/2/ | |
| Software, algorithm | Chimera 1.13.1 | Pettersen et al., 2004 | http://www.cgl.ucsf.edu/chimera/ | |
| Software, algorithm | ChimeraX 0.7 | Goddard et al., 2018 | https://www.cgl.ucsf.edu/chimerax/ | |
| Software, algorithm | CHARMM | Brooks et al., 1983 | https://www.charmm.org/charmm/ | |
| Software, algorithm | SWISS-MODEL | Biasini et al., 2014 | https://swissmodel.expasy.org/ | |
| Software, algorithm | Axon Clampex 10.6 | Molecular Devices | N/A | |
| Software, algorithm | Axon Clampfit 10.6 | Molecular Devices | N/A | |
| Software, algorithm | Prism 7 | GraphPad | https://www.graphpad.com/ |
Cell lines
Request a detailed protocolGnTI- cells used for protein expression and purification were obtaiend from ATTC (ATCC CRL-3022). Adherent HEK293T cells used for electrophysiology were obtained from ATTC (ATCC CRL-1573). Both cell-lines were tested negative for mycoplasma contamination. Suspension-adapted HEK293S GnTI- cells expressing murine Slc26a9 were grown at 37°C and 5% CO2 in either Pro293S-CDM or HyClone TransFx-H media, supplemented with 2 mM L-glutamine, 100 U ml–1 penicillin/streptomycin, 1% FBS, and 1% Pluronic F-68. Adherent HEK293T cells were grown in DMEM media supplemented with 1 mM L-glutamine, 100 U ml–1 penicillin/streptomycin, 10% FBS and 1 mM sodium pyruvate.
Construct generation
Request a detailed protocolDNA encoding the open reading frame (ORF) for mouse Slc26a9 (GenBank accession: BC160193) was PCR-amplified from a cDNA clone (Source BioScience) and shuttled into a pcDNA 3.1 vector (Invitrogen) which was modified to be compatible with FX cloning technology (Geertsma and Dutzler, 2011). Unless stated otherwise, all expression constructs also encoded a C-terminal Rhinovirus 3C protease cleavage site followed by venus YFP (vYFP), a myc epitope tag and a streptavidin binding peptide (SBP), giving the general construct scheme ORF-3C-vYFP-myc-SBP. Assembly of the Slc26a9T dual-truncation construct entailed removal of the STAS IVS region, via replacement of residues Pro558–Val660 with a Gly-Ser linker, and deletion of C-terminal residues Pro745–Leu790, akin to a described procedure for the isolated STAS domain from rat Prestin (Pasqualetto et al., 2010). Further deletion of N-terminal residues Met1–Ala30 resulted in the construct Slc26a9(Δ1-30)T. Mutations Q88A, Q88E, F92A, T127A, F128A, L391A, S392A, R205E, K221E, K270E, K431E, and K441E were introduced into Slc26a9T using the QuikChange site-directed mutagenesis method (Agilent).
Protein expression and purification
Request a detailed protocolSuspension-adapted HEK293S GnTI- cells (ATCC CRL-3022) were grown in either Pro293S-CDM (Lonza) or HyClone TransFx-H (GE Healthcare) media, supplemented with 2 mM L-glutamine (Sigma), 100 U ml–1 penicillin/streptomycin (Sigma), 1% FBS, and 1% Pluronic F-68. Cultures were maintained in TubeSpin Bioreactor 600 vessels (TPP), shaken at 185 rpm with an orbital radius of 50 mm, and incubated at 37°C and 5% CO2. For protein expression, a transient transfection protocol relying on 25 kDa linear polyethylenimine (PEI, Polysciences) was employed. One day prior to transfection, cells at high density (3–5 × 106 ml–1) were diluted into fresh media to a density of 0.6–0.8 × 106 ml–1. Transfection-grade plasmid DNA was purified from MC1061 E. coli culture using the NucleoBond Xtra Maxi kit (Macherey-Nagel), and a ratio of 1.3 μg DNA per 106 of HEK293S-GnTI- cells was used for transfection. Plasmid DNA was diluted into non-supplemented DMEM media (Sigma) at a concentration of 0.015 μg μl−1, and PEI was added to a concentration of 0.038 μg μl–1 from a 1 mg ml–1, pH 7 stock solution. After 10–15 min, the DNA-PEI mixtures were diluted 10-fold directly into the cultured cells, and valproic acid (Sigma) was added to a final concentration of 3 mM. After 40–48 hr, cells were harvested by centrifugation at 500 g for 10 min, washed with PBS, and then either directly used for protein extraction and purification or flash frozen in liquid nitrogen and stored at –80°C.
All subsequent protein extraction and purification procedures were carried out at 4°C. Cell pellets from 4–liter expression batches were resuspended in 60 ml of resuspension buffer (25 mM HEPES, pH 7.4, 200 mM NaCl, 5% glycerol, 2 mM CaCl2, and 2 mM MgCl2, 10 μg ml–1 DNase, and protease inhibitors (cOmplete EDTA-free, Roche). For cryo-EM analysis of detergent-solubilized Slc26a9T, the protein was extracted in digitonin (AppliChem), and subsequently purified in the presence of the synthetic digitonin substitute glyco-diosgenin (GDN, Anatrace). To extract membrane proteins, 2% (w/v) digitonin powder was directly dissolved in the cell resuspension, and the mixture was incubated for 1.5 hr under gentle agitation. Insoluble material was removed via ultracentrifugation for 40 min at 150,000 g and the supernatant was passed through a 5 μm syringe filter (Sartorius). The clarified extract was applied to 12 ml of StrepTactin Superflow affinity resin slurry (IBA Lifesciences), which was pre-equilibrated in wash buffer composed of 25 mM HEPES, pH 7.4, 200 mM NaCl, 5% glycerol, and 0.02% GDN. The affinity resin was washed with 20 CV of wash buffer, before elution with 15 ml of wash buffer supplemented with 10 mM D-desthiobiotin (Sigma). The protein was concentrated using a 100 kDa molecular weight cut-off (MWCO) centrifugal filter (Amicon) to 500 μl, typically resulting in a concentration of 1–2 mg ml–1 fusion protein, and 3C protease was added at a protein:protease mass ratio of 1:2. After a 1 hr incubation, the sample was centrifuged at 10,000 g for 3 min to pellet aggregated material, and the supernatant was passed through a 0.22 μm centrifugal filter (Millipore), before being injected onto a Superose 6 10/300 column (GE Healthcare) equilibrated in SEC buffer, 10 mM HEPES, pH 7.4, 200 mM NaCl, 0.02% GDN. Protein from peak fractions containing cleaved Slc26a9T protein was pooled, concentrated to 3 mg ml–1, and immediately used for cryo-EM grid preparation.
For the preparation of Slc26a9T protein to be reconstituted into either liposome or lipid nanodiscs, a similar extraction and purification procedure was performed, with the following modifications. Resuspended cells were extracted with a mixture of 1.5% n-dodecyl-β-D-maltoside (DDM, Anatrace) and 0.15% cholesteryl hemisuccinate (CHS, Anatrace), and protein was purified in the presence of 0.03% DDM and 0.003% CHS. For protein which was designated for liposome reconstitution, all other purification procedures were equivalent to the protocol used for cryo-EM sample preparation. However, for nanodisc preparations, an uncleaved variant of Slc26a9T, lacking a fluorescent fusion protein and therefore consisting of Slc26a9T-3C-myc-SBP (henceforth abbreviated as Slc26a9T-SBP), was purified and used for nanodisc assembly.
The behavior of all vYFP-tagged Slc26a9 constructs in detergent extracts was assessed with an HPLC system equipped for fluorescence-coupled size exclusion chromatography (FSEC, Agilent), using a Zorbax GF-450 column (Agilent). The same system was also employed to monitor the quality of purified proteins, using UV detection with a Superose 6 5/150 column (GE Healthcare). All protein samples were purified to ≥95% homogeneity, as assessed by SDS-PAGE.
Liposome and nanodisc reconstitution
Request a detailed protocolFor reconstitution of Slc26a9T into liposomes, a procedure relying on detergent-destabilization of preformed liposomes was employed, as previously described (Geertsma et al., 2008). Synthetic POPE and POPG lipids (Avanti Polar Lipids) at a mass ratio of 3:1 POPE:POPG were washed in diethyl ether, dried under N2 in a glass round-bottom flask, and hydrated via gentle sonication in liposome buffer, 10 mM HEPES, pH 7.4, 100 mM KCl, to a concentration of 20 mg ml–1. The lipid mixture was subjected to three freeze-thaw cycles, followed by extrusion through two 400 nm polycarbonate filters (LiposoFast Basic, Avestin) to form large unilamellar vesicles (LUVs). LUVs were diluted in liposome buffer to 4 mg ml–1, and 10% (w/v) Triton X-100 was added dropwise until the solution absorbance at 540 nm value reached a maximum, indicating suitable destabilization of liposomes for incorporation of membrane protein. Purified Slc26a9T (in DDM-CHS) was added to the destabilized LUVs at a protein:lipid ratio of 1:80 (w/w). After 20 min of incubation at RT, the sample was cooled to 4°C and detergent was removed via sequential additions of 250 mg SM-2 Bio-Beads (Bio-Rad) per 5 ml, every 24 hr for three days. Bio-Beads were removed via gravity filtration, and Slc26a9T proteoliposomes were pelleted via ultracentrifugation at 150,000 g for 30 min, resuspended in liposome buffer to 20 mg ml–1, frozen in liquid N2, and stored at –80°C. Identical procedures were used to produce mock liposomes lacking any reconstituted membrane protein.
Slc26a9T-SBP was reconstituted into lipid nanodiscs as described (Ritchie et al., 2009), with subtle modifications. We utilized the engineered membrane scaffold protein (MSP) MSP1-E3D1 because the estimated resultant nanodisc dimeter of 12 nm is ideal for the incorporation of Slc26a9T, which our preliminary cryo-EM analysis had suggested to possess a length of 10–11 nm when solubilized in detergent. MSP1-E3D1 was purified as described (Ritchie et al., 2009) and a 3:1 (w/w) mixture of the synthetic lipids POPC:POPG was dried and hydrated in 10 mM HEPES, pH 7.4, 100 mM KCl, as described above for proteoliposome preparations, except the final lipid concentration was 10 mM, and DDM was added to the stock lipid mixture to a final concentration of 27.5 mM. To assemble nanodiscs, purified Slc26a9T-SBP was diluted into 10 mM HEPES, 200 mM NaCl, 0.5 mM DDM, to a final protein concentration of 6 μM. Lipids were added to a concentration of 1.9 mM, and the mixture was incubated on ice for 30 min. Next, MSP1-E3D1 was added to a concentration of 20 μM, giving a final protein:lipid:MSP molar ratio of 1:450:20, at a final volume of 750 μl. After an additional 30 min of incubation on ice, 200 mg of SM2 Bio-Beads was added to remove detergent, and the reaction was allowed to incubate overnight under slow rotation at 4°C. Bio-beads were removed via gravity filtration, and Slc26a9T-SBP-reconstituted nanodiscs were isolated from empty nanodiscs via secondary purification with 3 ml of Streptavidin Plus UltraLink affinity resin (ThermoFisher) using detergent-free nanodisc buffer, 10 mM HEPES, pH 7.4, 150 mM NaCl, and elution with 5 mM biotin. Finally, nanodiscs were injected onto a Superose 6 5/300 column equilibrated in nanodisc buffer, and a peak containing Slc26a9T-SBP nanodiscs was concentrated to 1 mg ml–1 and immediately used for cryo-EM grid preparation.
Liposome anion transport assay
Request a detailed protocolTo measure electrogenic anion transport in Slc26a9T proteoliposomes, we used a method based on a previously described fluorometric assay (Kane Dickson et al., 2014) in which the internal liposome buffer ideally contains no permeant ions. Since Slc26a9 has low permeability to cations and sulfate, we exchanged the internal buffer of Slc26a9T proteoliposomes and mock liposomes to 10 mM HEPES, 50 mM Na2SO4 by pelleting the proteoliposomes (150,000 g, 30 min), resuspending in internal sulfate buffer to a concentration of 1 mg ml–1, and subjecting the sample to three freeze-thaw cycles. Finally, liposomes were again pelleted and resuspended to 20 mg ml–1 in internal sulfate buffer. All subsequent procedures were carried out at RT to restrict formation of multilamellar vesicles, and mock liposomes were always assayed in parallel as a negative control. To form small unilamellar vesicles, 20 μl aliquots of liposomes in 0.2 ml conical tubes were placed in a bath sonicator for 5–10 s, until the opaque solution became translucent. Liposomes were diluted 100-fold into flux buffer, consisting of 10 mM HEPES, pH 7.4, 75 mM NaCl, and 2 μM of the fluorophore 9-amino-6-chloro-2-methoxyacridine (ACMA, ThermoFisher), and 100 μl aliquots were transferred to a 96-well black-walled microplate (ThermoFisher). ACMA fluorescence was monitored with an Infinite M1000 spectrofluorometer (Tecan) in 5 s intervals, using excitation and emission wavelengths of 412 nm and 482 nm, respectively. After recording baseline fluorescence for 60 s, data collection was paused, 300 nM of the proton ionophore carbonyl cyanide 3-chlorophenylhydrazone (CCCP, Sigma) was added, and fluorescence measurements were immediately continued. Fluorescence intensity for each experimental condition was normalized to the initial value directly following addition of CCCP. For inhibition experiments, diluted proteoliposomes (0.2 mg ml–1) were pre-incubated for 5 min with 0–250 µM 4,4’−2,2’-disulfonic acid (DIDS, Sigma) and dimethyl sulfoxide (DMSO) was added to all samples to a final concentration of 1% to maintain solubility of DIDS.
Cryo-EM sample preparation and data collection
Request a detailed protocolFor structure determination of Slc26a9T by cryo-EM, 2.5 μl samples of GDN-purified protein at a concentration of 3 mg ml–1 were applied to glow-discharged holey carbon grids (Quantifoil R1.2/1.3 Au 200 mesh). For the structural characterization of the protein in a membrane-like environment, 2.5 μl samples of Slc26a9T reconstituted in E3D1 lipid nanodiscs at a concentration of 0.5–1 mg ml–1 were applied in a similar manner. Excess liquid was removed in a controlled environment (4°C and 100% relative humidity) by blotting grids for 3–6 s. Grids were subsequently flash frozen in liquid propane-ethane mix using a Vitrobot Mark IV (Thermo Fisher Scientific). Slc26a9T in detergent (dataset 1) was imaged in a 300 kV Titan Krios (Thermo Fisher Scientific) with a 100 μm objective aperture. Slc26a9T in nanodiscs (dataset 2) was recorded on a 300 kV Tecnai G2 Polara (FEI) with a 100 μm objective aperture. All data were collected using a post-column quantum energy filter (Gatan) with a 20 eV slit and a K2 Summit direct detector (Gatan) operating in super-resolution (for dataset 1) and counting (for dataset 2) modes. Dose-fractionated micrographs were recorded in an automated manner using SerialEM (Mastronarde, 2005) with a defocus range of –0.5 to –3.0 μm. Dataset one was recorded at a nominal magnification of 46,511 corresponding to a pixel size of 1.075 Å/pixel (0.5375 Å/pixel in super-resolution) with a total exposure time of 13 s (65 individual frames) and a dose of approximately 1.1 e–/Å2/frame. Dataset two was recorded at a nominal magnification of 37,313 corresponding to a pixel size of 1.34 Å/pixel with a total exposure time of 12.5 s (50 individual frames) and a dose of approximately 1.2 e–/Å2/frame. The total electron dose on the specimen level for dataset 1 and 2 was approximately 70 e–/Å2 and 60 e–/Å2, respectively.
Cryo-EM image processing
Request a detailed protocolThe recorded super-resolution images of Slc26a9T in detergent (dataset 1) were down-sampled twice by Fourier cropping and all individual frames were used for correction of beam-induced movement using a dose-weighting scheme in RELION’s own implementation of MotionCor2 algorithm available in version 3.0 (Zivanov et al., 2018a). The CTF parameters were estimated on summed movie frames using CTFFIND4.1 (Rohou and Grigorieff, 2015). All individual frames of Slc26a9T reconstituted in nanodiscs (dataset 2) were pre-processed in the same manner. Low-quality micrographs showing a significant drift or poor CTF estimates were discarded resulting in datasets of 2838 images of Slc26a9T in detergent and 3134 images of Slc26a9T in nanodiscs, which were subjected to further data processing in RELION (Scheres, 2012). From dataset one 416,164 particles were picked automatically using low-pass filtered 2D templates generated from an initial reference-free 2D classification. The particles were extracted with a box size of 232 pixels, down-scaled twice and subjected to a couple of rounds of 2D classification. Having discarded false positives and particles of poor quality, the dataset was reduced to 241,438 particles. The initial 3D reconstruction, which was generated from 3600 randomly chosen cleaned particles, was low-pass filtered to 60 Å and used as a template in a subsequent 3D classification. Multiple independent rounds of non-symmetrized 3D classification with a different number of classes were performed in order to isolate the most homogeneous subset of particles. One out of five classes, contained almost two-thirds of all the particles and showed clear two-fold symmetry. 157,644 particles belonging to this class were subjected to auto-refinement with imposed C2 symmetry. The particles were then unbinned to a pixel size of 1.075 Å/pixel and refined to 4.3 Å using C2 symmetry and a soft mask around the protein-detergent micelle density. The reconstruction was further improved to 4.09 Å by performing per-particle defocus and beam tilt corrections followed by Bayesian polishing (Zivanov et al., 2018a; Zivanov et al., 2018b). To enhance features of the peripheral helices α6 and α7, which were of lower local resolution than the core of the transmembrane domain, the density representing the detergent belt was subtracted from individual particles. In silico modified particles were used as an input in a subsequent masked 3D classification (Scheres, 2016). To separate any remaining heterogeneity particle angles were kept the same as the orientations from the consensus model. The final class containing 112,930 homogeneous particles was further auto-refined with C2 symmetry and in the presence of a soft mask around the protein density. The final map at 3.96 Å was sharpened using an isotropic b-factor of –205 Å2. The initial dataset of Slc26a9T in E3D1 lipid nanodiscs contained 711,032 particles, which were subjected to 2D classification in order to discard particles of poor quality, small aggregates and any remaining empty nanodiscs. After a stringent selection 308,872 particles were used in a subsequent non-symmetrized 3D classification. Similarly to the dataset of Slc26a9T in detergent, multiple independent runs were performed to optimally separate the heterogeneity in the dataset. Here, 3D classes showed more apparent heterogeneity compared to the detergent dataset. One out of seven classes containing 64,670 particles displayed well-resolved two-fold symmetric STAS domains although the extracellular loops located between the helices α3 and α4 of each subunit showed poorer density indicating increased flexibility of this region. As at this level of 3D classification the quality of the reconstruction did not allow to observe whether this flexibility is further extended to the TM domain or whether is affecting one or two monomers, further refinement was continued, side-by-side, without imposing any symmetry as well as with imposing C2 symmetry. Within the auto-refined non-symmetrized class one monomer was better resolved comparing to the second monomer and it also showed similar arrangement as either monomer in the structure of the protein in detergent. On the other hand, the second monomer showed increased flexibility only in the TM domain and the extracellular region as its STAS domain was identical as in the first monomer and as the STAS domain in the detergent structure. These results have suggested that despite eliminating particles with large heterogeneous differences during previous rounds of 2D and 3D classification, small conformational changes accounted for the remaining flexibility. In order to separate these changes into discrete classes a focused 3D classification was performed (Scheres, 2016). In this approach the mask around the flexible monomer was applied and the angles were kept fixed at the orientations from the auto-refined model. Given the classification focused only on a small region of the protein, the regularization parameter T was increased to 40. Approximately one-fourth of the remaining particles from both non-symmetrized and C2-symmetrized consensus models contributed to a class that recovered the density representing the extracellular loop. After identification of these homogeneous subsets that represent the stable monomer, final 3D auto-refinement of the whole dimeric particles was performed either with C1 or C2 symmetry imposed, yielding in both cases two-fold symmetric reconstructions of 8.4 Å and 7.77 Å, respectively. In all cases resolution was estimated in the presence of a soft solvent mask and based on the gold standard Fourier Shell Correlation (FSC) 0.143 criterion (Chen et al., 2013; Rosenthal and Henderson, 2003; Scheres, 2012; Scheres and Chen, 2012). High-resolution noise substitution was applied to correct FSC curves for the effect of soft masking in real space (Chen et al., 2013). The local resolution was estimated using BlocRes from the Bsoft package (Cardone et al., 2013; Heymann and Belnap, 2007).
Model building and refinement
Request a detailed protocolThe model of Slc26a9Tin the inward-facing state was built in Coot (Emsley and Cowtan, 2004) using the transmembrane domain structure of SLC26Dg (PDBID: 5DA0) and the chicken Prestin STAS domain (PDBID: 5EZB) as templates. The cryo-EM density of Slc26a9T in detergent was of sufficiently high resolution to unambiguously assign residues 5–27, 42–559 and 661–740. The model was improved iteratively by cycles of real-space refinement in PHENIX (Adams et al., 2002) with secondary structure and 2-fold NCS constraints applied, reciprocal-space refinement in REFMAC5 (Brown et al., 2015; Murshudov et al., 2011) and manual corrections in Coot. Validation of the refinement of the model was performed in REFMAC5, distributed as part of the CCP-EM suite (Burnley et al., 2017), and represented as Fourier Shell Correlation (FSCsum) between the refined model and the corresponding final cryo-EM density map. For cross-validation the detergent cryo-EM dataset was split into two subsets which were used to calculate two independent maps (half map one and half map 2). To detect possible overfitting and hence over-estimation of the resolution, random shifts, up to 0.5 Å, were applied to the coordinates of the model of the inward-facing state, which was then refined against the unfiltered half map 1. The cross-validation was done by comparing the FSCwork (estimated for the shaken-refined model and half map 1) and the FSCfree (estimated for the shaken-refined model and half map 2, which was not used in the refinement). The model of the intermediate state of Slc26a9T in nanodiscs was assembled by splitting the model of the inward-facing state into six independent entities, the STAS, gate and core domains, and refining them in PHENIX as rigid-bodies. In the next stage, the STAS, gate and core domain were linked into a single polypeptide chain and refined as described above with global minimization applied. The secondary structure and NCS constrains were maintained at all times. As the protein side-chains were not defined in our 7.77 Å cryo-EM reconstruction of the intermediate state, we have truncated all the side-chains to alanine. The validation of the refinement was performed as described above and represented as FSCsum. Owing to the low-resolution of the 3D reconstruction from the dataset in nanodiscs, FSCwork and FSCfree were not calculated. Surfaces were calculated with MSMS (Sanner et al., 1996). Figures and videos containing molecular structures and densities were prepared with DINO (http://www.dino3d.org), PyMOL (DeLano, 2002), Coot (Emsley and Cowtan, 2004),Chimera (Pettersen et al., 2004) and ChimeraX (Goddard et al., 2018).
Modeling and Poisson-Boltzmann calculations
Request a detailed protocolThe electrostatic potential in the intracellular vestibule leading to the Cl–-binding site was calculated by solving the linearized Poisson–Boltzmann equation in CHARMM (Brooks et al., 1983; Im et al., 1998) on a 150 Å ×170 Å × 200 Å grid (1 Å grid spacing) followed by focusing on a 120 Å x 130 Å x 160 Å grid (0.5 Å grid spacing). Partial protein charges were derived from the CHARMM36 all-hydrogen atom force field. Hydrogen positions were generated in CHARMM. Histidines were protonated. The protein was assigned a dielectric constant (ϵ) of 2. Its transmembrane region was embedded in a 30 Å-thick slab (ϵ = 2) representing the hydrophobic core of the membrane and two adjacent 15 Å-thick regions (ϵ = 30) representing the headgroups. The membrane region contained a 22 Å-high and 30 Å-wide aqueous cylinder (ϵ = 80) covering the intracellular vestibule of the protein and was surrounded by an aqueous environment (ϵ = 80). Calculations were carried out in either 150 mM of monovalent mobile ions in the aqueous regions (except for the membrane-inserted cylinder). The electrostatic surface potential shown in Figure 3—figure supplement 2F was calculated and displayed with COOT (Emsley and Cowtan, 2004). Homology models of murine SLC26 paralogs were prepared with the SWISS-MODEL homology modelling server (Biasini et al., 2014).
Electrophysiology
Request a detailed protocolAdherent HEK293T cells (ATCC CRL-1573) were grown in DMEM media supplemented with 1 mM L-glutamine (Sigma), 100 U ml–1 penicillin/streptomycin (Sigma), 1 mM sodium pyruvate (Sigma), and 10% FBS, at 37°C and 5% CO2, in 6 cm culture dishes. For transfection, 2.5–5 μg plasmid DNA encoding the construct of interest was mixed with 25 kDa linear polyethylenimine (Polysciences) in a ratio of 1:2.5 (w/w) in 0.3 ml PBS, incubated for 10 min at room temperature, and added dropwise to cells. Transfected cells were used within 24 hr for whole-cell recordings, or within 40 hr for excised patch recordings. Borosilicate glass capillaries (OD = 1.5 mm, ID = 0.86 mm, Sutter) were pulled and fire-polished, giving patch pipettes with resistances of 3–8 MΩ when backfilled with 150 mM NaCl pipette solution. All voltage-clamp signals were recorded using an Axopatch 200B amplifier (Molecular Devices) digitized with a Digidata 1440A A/D converter (Molecular Devices), filtered at 5 kHz, sampled at 20 kHz, and collected with Clampex 10.6 (Molecular Devices). For full-length Slc26a9, membrane seal resistance was typically 2–10 GΩ. However, for Slc26a9T, membrane seal resistance was frequently <1 GΩ, owing to its considerably higher constitutive Cl- conductance. Membrane seals for Slc26a9T-expressing cells showing resistance <0.2 GΩ were rejected. In whole-cell measurements of full-length Slc26a9, typical cell capacitance upon break-in was 15–30 pF, and series resistance was <10 MΩ after 60–80% compensation. Capacitive transients and series resistance could not be measured for Slc26a9T upon break-in, presumably due to its high constitutive conductance. Therefore, Slc26a9T whole-cell currents could not be expressed as normalized current densities (pA pF–1) and instead average raw currents were used for comparisons. For all experiments, the typical data acquisition protocol consisted of a holding potential of 0 mV for 0.2 s, followed by 0.2–0.4 s voltage steps from –100 mV to +100 mV in 20 mV increments, before returning to 0 mV. Calculated liquid junction potentials (JPCalcW, Molecular Devices) were corrected if they were greater than 2 mV, but never exceeded 5 mV. For all experiments, cells or excised patches expressing Slc26a9 constructs were only accepted for analysis if they displayed negligible conductance of NMDG-methylsulfonate.
Excised inside-out patches were employed for all ion selectivity experiments, in which the standard pipette solution contained 146 mM NaCl, 2 mM MgCl2, and 5 mM EGTA 10 mM HEPES, pH 7.4. For all solutions, pH was adjusted using NMDG or methanesulfonic acid. For assessment of anion/cation selectivity, patches were intracellularly perfused with solutions containing the same components as the pipette solution, except the NaCl concentration was varied between 15–150 and balanced to the ion concentrations of the pipette solution with NMDG-methylsulfonate. When employing perfusate concentrations above 150 mM NaCl, KCl was substituted for in order to minimize the liquid junction potential. For bi-ionic anion substitution experiments, the intracellular solution contained 150 mM sodium salts of substitute anions Br–, I–, F–, SCN–, NO3–, and SO42–, as well as 2 mM Mg(OAc)2, 5 mM EGTA, and 10 mM HEPES, pH 7.4. For the measurement of bicarbonate selectivity, a perfusate concentration of 165 mM NaHCO3 was prepared, because [HCO3–] / [HCO3–] + [H2CO3 + CO2]=0.9 at pH 7.4, due to the pKa of 6.4 for protonation of HCO3–. Fresh NaHCO3 solution was prepared every 1–3 hr, and pH monitoring of HCO3– solutions indicated that the decrease in [HCO3-] due to production of CO2 gas never exceeded 20% during recordings. In all selectivity experiments, Px/PCl values were calculated according to the GHK equation, and I-V plots were normalized to the current at +100 mV in symmetrical 150 mM NaCl. To obtain Gx/GCl conductance ratios, macroscopic chord conductance was calculated according to the equation , using the efflux current at –100 mV. For the determination of Km and relative Vmax values for Cl– and SCN–, the pipette solution contained 7.5 mM NaCl, 142.5 mM NMDG-methylsulfonate, 2 mM Mg(OAc)2, 5 mM EGTA, and 10 mM HEPES, pH 7.4, and the perfusion solutions contained 7.5–300 mM chloride or thiocyanate salts (sodium salts were used for all solutions except for 300 mM KCl). The current amplitude at 300 mM intracellular Cl– and –100 mV was used to normalize datasets. To calculate macroscopic SCN– conductances, Erev values for each perfusion concentration of SCN- were calculated according to the GHK equation, using PSCN/PCl values which were earlier determined in bi-ionic 150 mM SCNin/Clout selectivity experiments. Fitting the conductance vs. anion concentration data with the Michaelis-Menten equation allowed determination of Vmax and Km values. All electrophysiology data were analyzed with Clampfit 10.6 (Molecular Devices), Excel (Microsoft), and Prism 7 (GraphPad).
Statistics and reproducibility
Request a detailed protocolElectrophysiology data were repeated multiple times from different transfections with very similar results. Conclusions of experiments were not changed upon inclusion of further data. In all cases, leaky patches were discarded.
Accession codes
Request a detailed protocolThe cryo-EM density maps of Slc26a9T in detergent and lipid nanodiscs have been deposited in the Electron Microscopy Data Bank under ID codes EMD-4997 and EMD-4997, respectively. The coordinates for the atomic model of Slc26a9T refined against the 3.96 Å cryo-EM density in detergent and the coordinates of Slc26a9T with side-chains truncated to their β-positions refined against the 7.77 Å cryo-EM density in lipid nanodiscs have been deposited in the Protein Data Bank under ID codes 6RTC and 6RTF, respectively.
Data availability
The cryo-EM density maps of Slc26a9T in detergent and lipid nanodiscs have been deposited in the Electron Microscopy Data Bank under ID codes EMD-4997 and EMD-4997, respectively. The coordinates for the atomic model of Slc26a9T refined against the 3.96 Å cryo-EM density in detergent and the coordinates of Slc26a9T with sidechains-truncated to their β-positions refined against the 7.77 Å cryo-EM density in lipid nanodiscs have been deposited in the Protein Data Bank under ID codes 6RTC and 6RTF.
-
Electron Microscopy Data BankID EMD-4997. Cryo-EM density of murine Solute Carrier 26 family member A9 (Slc26a9) anion transporter in the inward-facing state.
-
Electron Microscopy Data BankID EMD-4998. Cryo-EM density of murine Solute Carrier 26 family member A9 (Slc26a9) anion transporter in an intermediate state.
-
Protein Data BankID 6RTC. Structure of murine Solute Carrier 26 family member A9 (Slc26a9) anion transporter in the inward-facing state.
-
Protein Data BankID 6RTF. Structure of murine Solute Carrier 26 family member A9 (Slc26a9) anion transporter in an intermediate state.
-
Protein Data BankID 5DA0. Structure of the the SLC26 transporter SLC26Dg in complex with a nanobody.
-
Protein Data BankID 4YZF. Crystal structure of the anion exchanger domain of human erythrocyte Band 3.
-
Protein Data BankID 5I6C. The structure of the eukaryotic purine/H+ symporter, UapA, in complex with Xanthine.
References
-
PHENIX: building new software for automated crystallographic structure determinationActa Crystallographica Section D Biological Crystallography 58:1948–1954.https://doi.org/10.1107/S0907444902016657
-
The SLC26 gene family of anion transporters and channelsMolecular Aspects of Medicine 34:494–515.https://doi.org/10.1016/j.mam.2012.07.009
-
SLC26A9-mediated chloride secretion prevents mucus obstruction in airway inflammationJournal of Clinical Investigation 122:3629–3634.https://doi.org/10.1172/JCI60429
-
SLC26A9 stimulates CFTR expression and function in human bronchial cell linesJournal of Cellular Physiology 226:212–223.https://doi.org/10.1002/jcp.22328
-
SLC26A9 is a constitutively active, CFTR-regulated anion conductance in human bronchial epitheliaThe Journal of General Physiology 133:421–438.https://doi.org/10.1085/jgp.200810097
-
The CFTR trafficking mutation F508del inhibits the constitutive activity of SLC26A9American Journal of Physiology-Lung Cellular and Molecular Physiology 312:L912–L925.https://doi.org/10.1152/ajplung.00178.2016
-
SWISS-MODEL: modelling protein tertiary and quaternary structure using evolutionary informationNucleic Acids Research 42:W252–W258.https://doi.org/10.1093/nar/gku340
-
Temperature-dependent changes of chloride transport kinetics in human red cellsThe Journal of General Physiology 70:283–306.https://doi.org/10.1085/jgp.70.3.283
-
CHARMM: a program for macromolecular energy, minimization, and dynamics calculationsJournal of Computational Chemistry 4:187–217.https://doi.org/10.1002/jcc.540040211
-
Tools for macromolecular model building and refinement into electron cryo-microscopy reconstructionsActa Crystallographica Section D Biological Crystallography 71:136–153.https://doi.org/10.1107/S1399004714021683
-
Recent developments in the CCP-EM software suiteActa Crystallographica Section D Structural Biology 73:469–477.https://doi.org/10.1107/S2059798317007859
-
One number does not fit all: mapping local variations in resolution in cryo-EM reconstructionsJournal of Structural Biology 184:226–236.https://doi.org/10.1016/j.jsb.2013.08.002
-
Slc26a9 is inhibited by the R-region of the cystic fibrosis transmembrane conductance regulator via the STAS domainJournal of Biological Chemistry 284:28306–28318.https://doi.org/10.1074/jbc.M109.001669
-
Slc26a9--anion exchanger, channel and na+ transporterJournal of Membrane Biology 228:125–140.https://doi.org/10.1007/s00232-009-9165-5
-
The novel class of seven transmembrane segment inverted repeat carriersBiological Chemistry 398:165–174.https://doi.org/10.1515/hsz-2016-0254
-
Conserved structure and domain organization among bacterial Slc26 transportersBiochemical Journal 463:297–307.https://doi.org/10.1042/BJ20130619
-
Prestin, a new type of motor proteinNature Reviews Molecular Cell Biology 3:104–111.https://doi.org/10.1038/nrm730
-
Lyotropic anions. Na channel gating and Ca electrode responseThe Journal of General Physiology 81:255–281.https://doi.org/10.1085/jgp.81.2.255
-
Pymol: An open-source molecular graphics toolCCP4 Newsletter On Protein Crystallography 40:82–92.
-
Cooperativity in cytochrome P450 3a4: linkages in substrate binding, spin state, uncoupling, and product formationThe Journal of Biological Chemistry 282:7066–7076.https://doi.org/10.1074/jbc.M609589200
-
Conserved dimeric subunit stoichiometry of SLC26 multifunctional anion exchangersJournal of Biological Chemistry 283:4177–4188.https://doi.org/10.1074/jbc.M704924200
-
SLC26A9 is a cl(-) channel regulated by the WNK kinasesThe Journal of Physiology 584:333–345.https://doi.org/10.1113/jphysiol.2007.135855
-
Congenital chloride-losing diarrhea causing mutations in the STAS domain result in misfolding and mistrafficking of SLC26A3Journal of Biological Chemistry 283:8711–8722.https://doi.org/10.1074/jbc.M704328200
-
Shared molecular mechanisms of membrane transportersAnnual Review of Biochemistry 85:543–572.https://doi.org/10.1146/annurev-biochem-060815-014520
-
Coot: model-building tools for molecular graphicsActa Crystallographica. Section D, Biological Crystallography 60:2126–2132.https://doi.org/10.1107/S0907444904019158
-
Asymmetry of inverted-topology repeats in the AE1 anion exchanger suggests an elevator-like mechanismThe Journal of General Physiology 149:1149–1164.https://doi.org/10.1085/jgp.201711836
-
Structure of a prokaryotic fumarate transporter reveals the architecture of the SLC26 familyNature Structural & Molecular Biology 22:803–808.https://doi.org/10.1038/nsmb.3091
-
Bsoft: image processing and molecular modeling for electron microscopyJournal of Structural Biology 157:3–18.https://doi.org/10.1016/j.jsb.2006.06.006
-
Continuum solvation model: computation of electrostatic forces from numerical solutions to the Poisson-Boltzmann equationComputer Physics Communications 111:59–75.https://doi.org/10.1016/S0010-4655(98)00016-2
-
SLC26A7 is a Cl- channel regulated by intracellular pHJournal of Biological Chemistry 280:6463–6470.https://doi.org/10.1074/jbc.M409162200
-
PDZ domain proteins of synapsesNature Reviews Neuroscience 5:771–781.https://doi.org/10.1038/nrn1517
-
N-glycosylation and topology of the human SLC26 family of anion transport membrane proteinsAmerican Journal of Physiology-Cell Physiology 306:C943–C960.https://doi.org/10.1152/ajpcell.00030.2014
-
Loss of Slc26a9 anion transporter alters intestinal electrolyte and HCO3 - transport and reduces survival in CFTR-deficient micePflügers Archiv - European Journal of Physiology 467:1261–1275.https://doi.org/10.1007/s00424-014-1543-x
-
Functional characterization of three novel tissue-specific anion exchangers SLC26A7, -A8, and -A9Journal of Biological Chemistry 277:14246–14254.https://doi.org/10.1074/jbc.M111802200
-
The STAS domain of mammalian SLC26A5 prestin harbours an anion-binding siteBiochemical Journal 473:365–370.https://doi.org/10.1042/BJ20151089
-
Characterization of SLC26A9, facilitation of cl(-) transport by bicarbonateCellular Physiology and Biochemistry : International Journal of Experimental Cellular Physiology, Biochemistry, and Pharmacology 22:15–30.https://doi.org/10.1159/000149780
-
Targeting ion channels in cystic fibrosisJournal of Cystic Fibrosis 14:561–570.https://doi.org/10.1016/j.jcf.2015.06.002
-
Automated electron microscope tomography using robust prediction of specimen movementsJournal of Structural Biology 152:36–51.https://doi.org/10.1016/j.jsb.2005.07.007
-
Variants in solute carrier SLC26A9 modify prenatal exocrine pancreatic damage in cystic fibrosisThe Journal of Pediatrics 166:1152–1157.https://doi.org/10.1016/j.jpeds.2015.01.044
-
REFMAC5 for the refinement of macromolecular crystal structuresActa Crystallographica Section D Biological Crystallography 67:355–367.https://doi.org/10.1107/S0907444911001314
-
Determinants of coupled transport and uncoupled current by the electrogenic SLC26 transportersThe Journal of General Physiology 137:239–251.https://doi.org/10.1085/jgp.201010531
-
UCSF chimera--a visualization system for exploratory research and analysisJournal of Computational Chemistry 25:1605–1612.https://doi.org/10.1002/jcc.20084
-
Slc26a11 is prominently expressed in the brain and functions as a chloride channel: expression in purkinje cells and stimulation of V H+-ATPasePflügers Archiv - European Journal of Physiology 465:1583–1597.https://doi.org/10.1007/s00424-013-1300-6
-
Chapter 11 - Reconstitution of membrane proteins in phospholipid bilayer nanodiscsMethods in Enzymology 464:211–231.https://doi.org/10.1016/S0076-6879(09)64011-8
-
CTFFIND4: fast and accurate defocus estimation from electron micrographsJournal of Structural Biology 192:216–221.https://doi.org/10.1016/j.jsb.2015.08.008
-
RELION: implementation of a bayesian approach to cryo-EM structure determinationJournal of Structural Biology 180:519–530.https://doi.org/10.1016/j.jsb.2012.09.006
-
Processing of structurally heterogeneous Cryo-EM data in RELIONMethods in Enzymology 579:125–157.https://doi.org/10.1016/bs.mie.2016.04.012
-
Prevention of overfitting in cryo-EM structure determinationNature Methods 9:853–854.https://doi.org/10.1038/nmeth.2115
-
Discontinuous membrane helices in transport proteins and their correlation with functionJournal of Structural Biology 159:261–267.https://doi.org/10.1016/j.jsb.2007.01.011
-
STAS domain structure and functionCellular Physiology and Biochemistry 28:407–422.https://doi.org/10.1159/000335104
-
Coupling modes and stoichiometry of cl-/HCO3- exchange by slc26a3 and slc26a6The Journal of General Physiology 127:511–524.https://doi.org/10.1085/jgp.200509392
-
Cystic fibrosis gene modifier SLC26A9 modulates airway response to CFTR-directed therapeuticsHuman Molecular Genetics 25:4590–4600.https://doi.org/10.1093/hmg/ddw290
-
Anion selectivity in biological systemsPhysiological Reviews 57:109–156.https://doi.org/10.1152/physrev.1977.57.1.109
-
SLC26A9 is expressed in gastric surface epithelial cells, mediates cl-/HCO3- exchange, and is inhibited by NH4+American Journal of Physiology. Cell Physiology 289:C493–C505.https://doi.org/10.1152/ajpcell.00030.2005
-
Structure of the mechanosensitive OSCA channelsNature Structural & Molecular Biology 25:850–858.https://doi.org/10.1038/s41594-018-0117-6
-
Analysis of the oligomeric structure of the motor protein prestinJournal of Biological Chemistry 281:19916–19924.https://doi.org/10.1074/jbc.M513854200
Article and author information
Author details
Funding
Schweizerischer Nationalfonds zur Förderung der Wissenschaftlichen Forschung (31003A_163421)
- Raimund Dutzler
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank O Medalia and M Eibauer, the Center for Microscopy and Image Analysis (ZMB) of the University of Zurich, and the Mäxi foundation for the support and access to the electron microscopes, S Klauser and S Rast for their help in establishing the computer infrastructure. All members of the Dutzler lab are acknowledged for their help at various stages of the project. This research was supported by a grant from the Swiss National Science Foundation (No. 31003A_163421).
Copyright
© 2019, Walter et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 7,354
- views
-
- 851
- downloads
-
- 126
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 126
- citations for umbrella DOI https://doi.org/10.7554/eLife.46986
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Cryo-EM structures and functional characterization of murine Slc26a9 reveal mechanism of uncoupled chloride transport
eLife 8:e46986.
https://doi.org/10.7554/eLife.46986




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}