Dzip1 and Fam92 form a ciliary transition zone complex with cell type specific roles in Drosophila
- Institut NeuroMyoGène, CNRS UMR 5310, INSERM U1217, Université Claude Bernard Lyon 1, France
- Institute of Human Genetics, Universitätsklinikum Düsseldorf, Heinrich-Heine-Universität Düsseldorf, Germany
- IRCM, INSERM, Université de Montpellier, ICM, France
Abstract
Cilia and flagella are conserved eukaryotic organelles essential for cellular signaling and motility. Cilia dysfunctions cause life-threatening ciliopathies, many of which are due to defects in the transition zone (TZ), a complex structure of the ciliary base. Therefore, understanding TZ assembly, which relies on ordered interactions of multiprotein modules, is of critical importance. Here, we show that Drosophila Dzip1 and Fam92 form a functional module which constrains the conserved core TZ protein, Cep290, to the ciliary base. We identify cell type specific roles of this functional module in two different tissues. While it is required for TZ assembly in all Drosophila ciliated cells, it also regulates basal-body growth and docking to the plasma membrane during spermatogenesis. We therefore demonstrate a novel regulatory role for Dzip1 and Fam92 in mediating membrane/basal-body interactions and show that these interactions exhibit cell type specific functions in basal-body maturation and TZ organization.
eLife digest
Many animal cells have hair-like structures called cilia on their surface, which help them to sense and interact with their surroundings. The cilia are supported by protein filaments and must assemble correctly because faulty cilia can lead to several life-threatening diseases. Problems in an area at the base of the cilia, known as the ‘transition zone’, account for the most severe forms of these diseases in humans.
The transition zone is responsible for selecting which proteins are allowed in and out of the cilia. The transition zone itself is made up of many proteins that work together to determine the cilia composition. But not all of these proteins are known, and it is unclear how those that are known affect cilia structure.
One protein found in transition zones of several animals, including fruit flies and mice, is called Cby. Lapart et al. set out to understand which other proteins interact with Cby in fruit flies to better understand what this protein does in the transition zone. A series of experiments showed that Cby interacts with two proteins called Dzip1 and Fam92 to regulate the assembly of transition zones. Together these three proteins constrain a core component of the transition zone, a fourth protein called Cep290, to the base of the cilia.
Fruit flies only have cilia on cells in their sensory organs and testes and, in both types of tissue, cilia could only form properly when Dzip1 and Fam92 were present. Lapart et al. also showed that, in the fruit fly testes, Dzip1 and Fam92 helped to anchor the newly forming cilia to the cell surface. This anchoring role was particularly important for the fruit flies’ sperm to grow their characteristic whip-like tails, which are a specialized type of cilia that allow sperm cells to move.
Overall, the findings show how some transition zone proteins work together and that they can have different effects in different tissues. Understanding the mechanisms behind healthy cilia assembly will likely be key to tackling cilia-related diseases.
Introduction
Cilia and flagella are highly conserved organelles present at the surface of eukaryotic cells. They play major physiological roles in animals such as cell or fluid mobility, signaling during development and cellular homeostasis. The importance of cilia and flagella is highlighted by the discovery of human diseases, classified as ciliopathies, that are associated with defects in cilia structure and/or function (Badano et al., 2006; Baker and Beales, 2009; Brown and Witman, 2014). Cilia and flagella are templated from the basal body, derived from the mother centriole, from which the microtubule based axoneme is assembled.
At the base of the cilium, a specific compartment, the transition zone (TZ) plays a critical role in cilium assembly and function. Many genes responsible for cilia associated diseases such as the Meckel syndrome (MKS), Joubert syndrome or nephronophthisis (NPHP) are caused by defects in proteins of the TZ (Czarnecki and Shah, 2012). The TZ functions as a ciliary gate by sorting selected components in and out of the cilium, thus controlling the specific composition of the ciliary compartment (Garcia and Reiter, 2016; Reiter et al., 2012; Gonçalves and Pelletier, 2017). TZ assembly starts during the first steps of cilia formation, when the mother centriole associates with cytoplasmic vesicles before docking to the plasma membrane. Assembly of TZ proteins is spatiotemporally controlled and requires, in most organisms, at least three different protein modules namely MKS, NPHP and CEP290. Extensive genetic, biochemical studies and super resolution microscopy analysis helped to establish a hierarchy of these components and a structural view of the TZ architecture (Williams et al., 2011; Garcia-Gonzalo et al., 2011; Sang et al., 2011; Chih et al., 2012; Gupta et al., 2015; Garcia and Reiter, 2016; Reiter et al., 2012; Gonçalves and Pelletier, 2017).
Although largely conserved from worms to mammals, all TZ proteins are not conserved in all ciliated species and variations exist between model organisms. For example, the NPHP module, present in mammals, worms and protozoa, is not conserved in flies, whereas CEP290 and several members of the MKS module are conserved in most organisms (Basiri et al., 2014; Pratt et al., 2016; Vieillard et al., 2016). In addition to the core TZ components, several others have been identified but their precise relationships with the core known TZ components are not understood. Among these other proteins, Chibby (Cby), a conserved TZ component in vertebrates and flies, is required for cilia function both in mammals and Drosophila (Burke et al., 2014; Voronina et al., 2009; Enjolras et al., 2012; Vieillard et al., 2016). In vertebrates, CBY has been shown to interact with several basal body (BB) associated proteins, such as ODF2 or CEP164 (Lee et al., 2014; Siller et al., 2017; Burke et al., 2014; Steere et al., 2012; Chang et al., 2013) and more recently DZIP1L, DZIP1 and FAM92a or b proteins (Wang et al., 2018; Li et al., 2016b; Breslow et al., 2017). However, the functional integration of CBY and these interactors in TZ assembly is unclear and some of those, such as Cep164 cannot likely sustain Cby function in Drosophila, as Cep164 does not seem to be expressed in Drosophila testes (Flybase).
We show here that the unique Drosophila orthologs Dzip1 (CG13617) and Fam92 (CG6405) of respectively, vertebrate DZIP1 or DZIP1L and FAM92a or b, interact and cooperate with Cby in flies. We demonstrate that all three proteins form a strictly ordered functional module, and cooperate in building the TZ in the two Drosophila ciliated tissues, with Dzip1 acting upstream of Fam92 and Cby. While our observations establish that Dzip1 and Fam92 localization at the TZ relies on Cep290, they reveal that Dzip1 and Fam92 exert a negative regulatory feedback loop by restraining Cep290 localization to the ciliary base.
Last, our work reveals remarkable differences in the role of Dzip1 and Fam92 in regulating basal bodies (BB) docking between the two Drosophila ciliated tissues. Whereas, loss of Dzip1 or Fam92 does not affect basal body docking in sensory cilia, it impairs BB-membrane growth and attachment in spermatocytes. As a consequence, we observed aberrant and premature elongation of the axoneme before completion of meiosis, highlighting a primary role of the BB-membrane associated compartment for regulating axonemal microtubule elongation in Drosophila spermatocytes. These aberrant elongations mostly affect the daughter centrioles, revealing functional differences of the mother and daughter centrioles in Drosophila spermatocytes.
Results
Dzip1/Fam92/Cby form a complex at the ciliary transition zone in Drosophila
We initially identified mouse CBY1 interactors by LAP-Tag affinity purification using IMCD3 cells (murine Inner Medullary Collecting Duct cells) stably expressing CBY1 fused to the EGFP-TEV-S peptide tag (see Materials and methods section). This EGFP-TEV-S tag in N-terminus allows to detect the protein in cells and its purification by a two-step affinity procedure (Torres et al., 2009). Analysis of LAP-CBY1 complex revealed the presence of DZIP1 and FAM92 among 22 identified proteins (see full list in Supplementary file 1), in agreement with recently published data (Li et al., 2016b; Breslow et al., 2018; Wang et al., 2018). DZIP1/1L and FAM92a/b each show a unique ortholog gene in Drosophila, CG13617 and CG6405 respectively (Figure 1—figure supplement 1A), but are absent, like CBY1, from the C. elegans genome. Hereafter, we name CG13617 and CG6405 as Dzip1 and Fam92, respectively. By co-immunoprecipitating Cby-GFP and HA-Dzip1 or HA-Fam92, we demonstrate that Drosophila Dzip1 and Fam92, each interact with Cby (Figure 1—figure supplement 1B–C). Dzip1 or Fam92 do not apparently interact with each other in these co-IP experiments (Figure 1—figure supplement 1D). However, when all three proteins are expressed together, immunoprecipitation of GFP-Dzip1 pulls down both Cby and Fam92, suggesting that all three proteins are present in one complex when co-expressed in mammalian cells (Figure 1—figure supplement 1D).
To identify the subcellular localization of Drosophila Dzip1 and Fam92, we generated transgenic flies expressing Dzip1-GFP or Fam92-GFP under the control of their respective promoters (Figure 1—figure supplement 2). We determined that both dzip1 and fam92 are exclusively expressed in the two kinds of Drosophila ciliated cell types, namely type I sensory neurons and male germ cells (Figures 1 and 2). Type I sensory neurons comprise chordotonal (Ch) and external sensory (ES) neurons, which harbor motile and immotile cilia respectively (Figure 1A) (Gogendeau and Basto, 2010; Jana et al., 2016). Each sensory neuron is enclosed in several support cells forming the sensory organ or scolopidia. Dzip1 and Fam92 decorate the base of the cilia at the tip of the sensory dendrites (labeled with 22C10) (Figure 1B, arrows). By performing, super-resolution 3D structured-illumination microscopy (3D-SIM), we confirmed that Dzip1 and Fam92 co-localize with Cby in sensory neurons (Figure 1C), demonstrating their restricted localization at the ciliary transition zone.
Figure 1 with 2 supplements see all
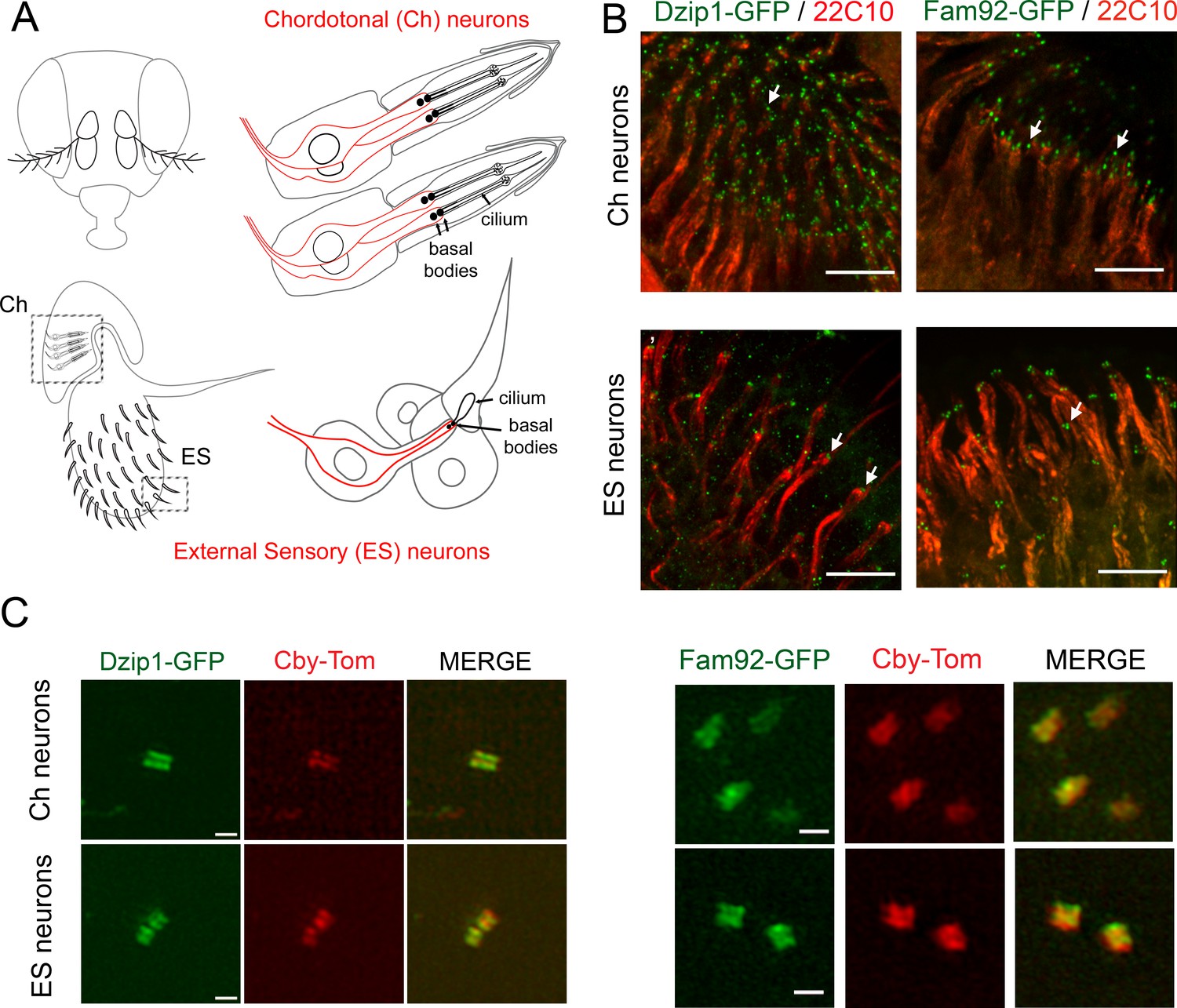
Drosophila Dzip1 and Fam92 are expressed in ciliated sensory neurons and locate at the ciliary transition zone.
(A) Scheme of the two types of sensory organs (or scolopidia) present in the antennae of the Drosophila head. Chordotonal scolopidia are proprioceptors present in the second antennal segment of the Drosophila head and respond to sound vibrations and gravity. Each chordotonal scolopidia is composed of several support cells (gray) and comprise two or three ciliated neurons (red). External sensory organs (or scolopidia) are present under each sensory sensilla of the third antennal segment and react to olfactory and chemical stimuli. In other body parts, ES organs also respond to mechanical stimuli. External sensory scolopidia comprise support cells (gray) and only one ciliated neuron (red). In ciliated sensory neurons, the two centrioles (or proximal and distal basal bodies) stand above each other at the tip of the dendrites. (B) Whole-mount staining of the second and third antennal segment showing neuronal cell bodies and dendrites (22C10), Dzip1-GFP and Fam92-GFP. Dzip1-GFP and Fam92-GFP are present at the tip of the dendrites in each type of neurons. Arrows point both to Dzip1-GFP and Fam92-GFP localization at the tip of the dendrites. (C) 3D-SIM imaging of Ch and ES neurons. Both Drosophila Dzip1 and Fam92 overlap with Cby at the transition zone. Bars = 10 μm for (B); = 0.5 μm for (C).
Figure 2

Drosophila Dzip1 and Fam92 are located at the transition zone during spermatogenesis.
(A) Scheme illustrating centriole/basal bodies (BB) behavior during spermatogenesis in male germ cells. In early spermatocytes, two pairs of centrioles (gray) are present in each cell. During spermatocyte maturation, centrioles convert to BB and dock to the plasma membrane while extending a primary like cilium which is entirely decorated with TZ proteins (green and red). In late spermatocytes, the TZ, also described as the primary like cilium, reaches approximatively 500 nm and protrudes at the cell surface. During meiosis, BB with primary like cilia/TZ are internalized, thus retaining the ciliary membrane (yellow) connected to the plasma membrane, hence creating a membrane cap in spermatids. In round spermatid, BB are apposed to the nuclear membrane and the ciliary cap (yellow) connected to the plasma membrane is extended. The ring centriole (green) marks the base of the ciliary cap which is decorated by TZ proteins. At the onset of axoneme elongation, axonemal microtubule nucleation inside the cap extends the ciliary cap/TZ, and as the axoneme grows (black), the ring centriole is pushed away from the basal body. (B–C) Confocal imaging of whole-mount testes showing Dzip1-GFP (B), Fam92-GFP (C), Cby-Tom and Asterless (centrioles). Dzip1 and Fam92 appear together with Cby in early spermatocytes at the tip of centrioles. In elongating spermatids, Drosophila Dzip1 and Fam92 mark the ring centriole (arrows) separating from the BB (asterless, gray). (D) 3D-SIM imaging of male germ cells. Drosophila Dzip1 and Fam92 overlap with Cby in spermatocytes. Plots of the intensity profile of the centrioles along the dotted lines illustrate the overlay between Cby-Tom and Dzip-GFP or Fam92-GFP. Bars = 1 μm.
In the male germline, Dzip1 and Fam92 appear first in spermatocytes (Figure 2) at the distal end of centrioles. In spermatocytes, centrioles have a specific behavior as both mother and daughter centrioles of each pair elongate and dock to the plasma membrane. All four basal bodies (BB) extend a TZ, also described as a primary cilium-like structure (Figure 2A) (Pasmans and Tates, 1971; Riparbelli et al., 2012; Gottardo et al., 2013; Vieillard et al., 2016). Subsequently, during meiosis, all four basal bodies are engulfed inside the cytoplasm, together with the primary like cilium which hence creates a ciliary cap that ensheaths each basal body distal end. After meiosis and at the onset of axoneme elongation, the ciliary cap is remodeled and a distinct domain, the ring centriole, appears at its base (Vieillard et al., 2016; Basiri et al., 2014; Fabian and Brill, 2012). The axoneme grows inside the ciliary cap, which concomitantly migrates, extruding the nascent axoneme out in the cytoplasm (Figure 2A) (Pasmans and Tates, 1971; Riparbelli et al., 2013; Gottardo et al., 2013). We observed that Dzip1, Fam92 and Cby all localize at the centriolar distal ends in early spermatocytes (Figure 2B–C). During centriolar/BB maturation, as the ciliary cap grows, Dzip1 and Fam92 localizations are extended and overlap with Cby, as revealed by 3D-SIM observations (Figure 2D). In spermatids, when the TZ migrates away from the basal body, we observed that Dzip1 and Fam92 strongly accumulate with Cby at the ring centriole (Figure 2B–C, arrows).
Together, these results strongly indicate that Dzip1/Fam92/Cby interact at the ciliary transition zone.
Dzip1 and Fam92 are required for cilia and flagella formation in Drosophila
To determine the roles of the Dzip1/Fam92 module in TZ assembly or function, we generated two deletion alleles of dzip1 or fam92 (Figure 1—figure supplement 2 and Materials and method section). In the dzip11 allele, the 65 N-ter codons (including start codon) were deleted and replaced by a 3 kb insertion including the mini-white gene (Figure 1—figure supplement 2). In fam921, exon two and part of exon three were removed (294 bp, Figure 1—figure supplement 2), leading to the deletion of 81 aa and a frameshift with several stop codons in the remaining downstream sequence, leaving only 47 aa of the WT protein and 39 amino acids of the −1 frameshifted open reading frame.
dzip11 and fam921 flies are viable but show typical behavioral defects associated with ciliary dysfunction in Drosophila. Fly geotaxis response (Figure 3A) was monitored by bang assay (Enjolras et al., 2012). Whereas most of the control flies reach and stay at the top of the tube 30 s after the bang, no dzip11 flies reached the top of the tube (Video 1). This phenotype was worsened in dzip11/Df as the flies not only fail to climb, but are completely immotile with held up wings (Video 2), indicating that dzip11 is likely not a complete null allele. Both dzip11 and dzip11/Df behavioral phenotypes were fully rescued by one copy of dzip1::GFP (not shown). In contrast, a few percentage (17%) of fam921 flies could reach the top of the tube (Figure 3A) and no differences could be observed when comparing fam921 flies and fam921 over its cognate deficiency (fam921/Df), indicating that fam921 is likely a null allele. The fam92::GFP rescue construct partially restores the climbing behavior of the fam921 mutant flies. These observations reveal a graded requirement for Fam92 and Dzip1 in Drosophila geotaxis response and indicate a regulatory role of this complex in sensory cilia assembly or function.
Figure 3
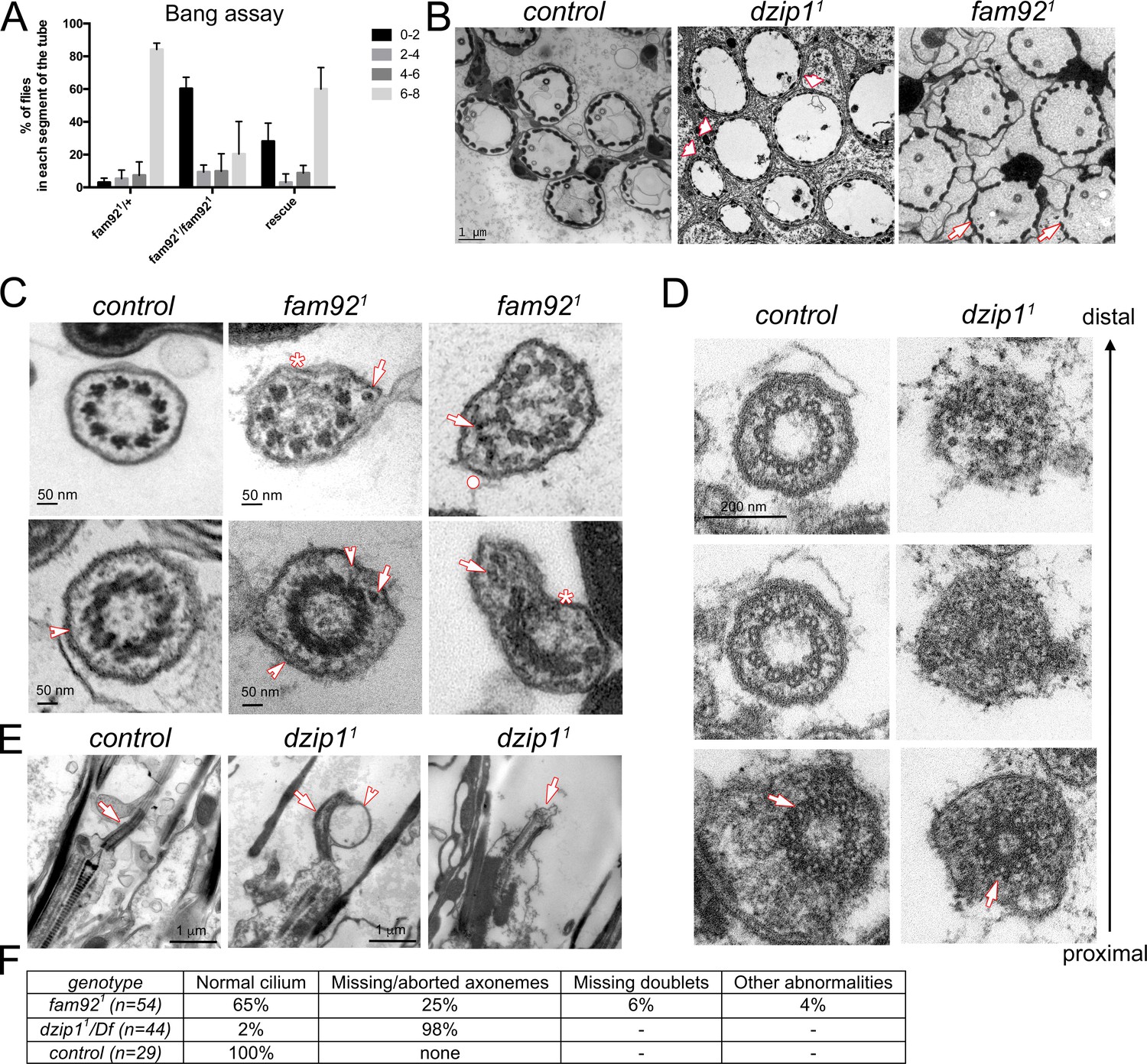
Dzip1 and Fam92 are required for ciliogenesis in Drosophila sensory cilia.
(A) Quantification by bang assay of geotaxis response of fam921 flies compared to control or rescued flies. The percentage of flies that reach a defined level in the tube (0–2, 2–4, 4–6, 6–8) is represented. fam921 flies are unable to climb along the tube compared to control flies. This defect is partially rescued by adding two copies of fam92::GFP transgene (fam921/+ n = 61, fam921/fam921 n = 63, rescue n = 68). (B) EM analysis of cross sections of antennal Chordotonal (Ch) neurons. Whereas two neurons/cilia can be observed in each control scolopidia, cilia are almost completely absent in dzip11 scolopidia (arrowheads). In fam921, reduced number of cilia are observed on several Ch neurons (arrows). (C) Ch neurons cilia ultrastructure of fam921 antennae, showing reduced number of microtubule doublets (asterisks) and/or accumulation of dense material (arrows) underneath the ciliary membrane and deformation of the membrane (dot). Similar defects are also observed on cross sections of the TZ (lower panels). Note that the linkers connecting the axoneme to the membrane are still present (arrowheads). (D) Serial sections of the basal body-transition zone region of Ch neurons in dzip11 compared to control, from the basal body (proximal, lower panels) to the TZ (distal, upper panels). Whereas doublet microtubules are present and symmetrically organized at the basal body level (arrows, lower panels), they fail to elongate along the transition zone which is incompletely assembled. (E) Longitudinal sections of dzip11 showing basal body with aberrant TZ compared to control (arrows). Membrane bulges (arrowhead) along the aberrant dzip11 TZ can also be detected. (F) Quantifications of the cilium defects observed in scolopidia.
Video 1
Real time imaging of dzip11adult flies.
Video 2
Real time imaging of dzip11/Df adult flies.
To determine the impact of Dzip1 or Fam92 loss on cilia assembly, we performed ultra-structural analysis by transmission electron microscopy (EM). Strikingly, we observed that cilia were essentially absent in dzip11 antennal chordotonal organs, but were still present in fam921 antennae (Figure 3B and F), in agreement with their milder geotaxis defect behavior. In fam921 flies, 25% of scolopidia (total observed n = 54) showed reduced cilia number (one or no cilia). In addition, defects in axonemal structure were occasionally observed, such as lack of microtubule doublets (Figure 3C, asterisks, 6%), excess accumulation of material beneath the ciliary membrane (arrows) and deformation of the membrane (Figure 3C, upper panel, dot, 4%). Most TZ sections showed normal ultrastructure, but a few (6%) showed incomplete radial symmetry and accumulating material as observed for cilia. Linkers connecting the axoneme to the membrane could still be observed (Figure 3C, arrowhead, lower panel). Transition zones were completely disorganized in dzip11 antennae as revealed by serial-cross and longitudinal sections (Figure 3D–E) and no more axoneme-to-membrane linkers could be observed. Basal bodies were normally present at the dendrite distal tip (Figure 3D, lower panel), but we observed a rapid disorganization of the axoneme, with its complete abrogation a few microns above the basal body (Figure 3D–E).
Hence, these observations demonstrate that loss of Dzip1 or Fam92 affects TZ assembly required for sensory cilia formation.
To address the function of Dzip1 and Fam92 in sperm flagellum assembly, we first investigated male fertility. We observed a strong reduction of the fertility of fam921 males compared to controls. This defect is restored by expressing Fam92-GFP (Figure 4A). Because dzip11 mutant flies are severely uncoordinated, their fertility could not be tested. However, dzip11 testes showed a marked dispersion of the nuclei along the cysts and, as a consequence, altered migration of sperm individualization complexes (Figure 4B and Figure 4—figure supplement 1A). Nuclear dispersion is also observed, to a lesser extent, in fam921 testes (Figure 4B and Figure 4—figure supplement 1A). One possible origin of nuclei dispersion is defective axonemal elongation (Vieillard et al., 2016; Soulavie et al., 2014) and EM analysis confirmed that axonemes were affected in fam921 cells (Figure 4D, 125/448 broken axonemes, 25/448 missing central pair, 52/448 missing axoneme). As well, in dzip11 or dzip11/Df mutant testes, marked axonemal defects could be observed. In dzip11 testes, we observed 6/126 broken axonemes, 1/126 missing central pair, 1/126 missing axonemes (Figure 4C). More severe defects were observed in very young dzip11/Df spermatid cysts, with up to 60% of spermatids with missing or broken axonemes (Figure 4—figure supplement 1B–C). However, in older cysts, the number of spermatids/cyst was much reduced compared to control (mean = 44 spermatids/cyst compared to control = 63.4), but almost all remaining elongated spermatids showed normal axonemes (Figure 4—figure supplement 1B). These observations suggest that young spermatids with severe axonemal defects fail to elongate and are not observed in cross sections of older spermatid cysts.
Figure 4 with 1 supplement see all
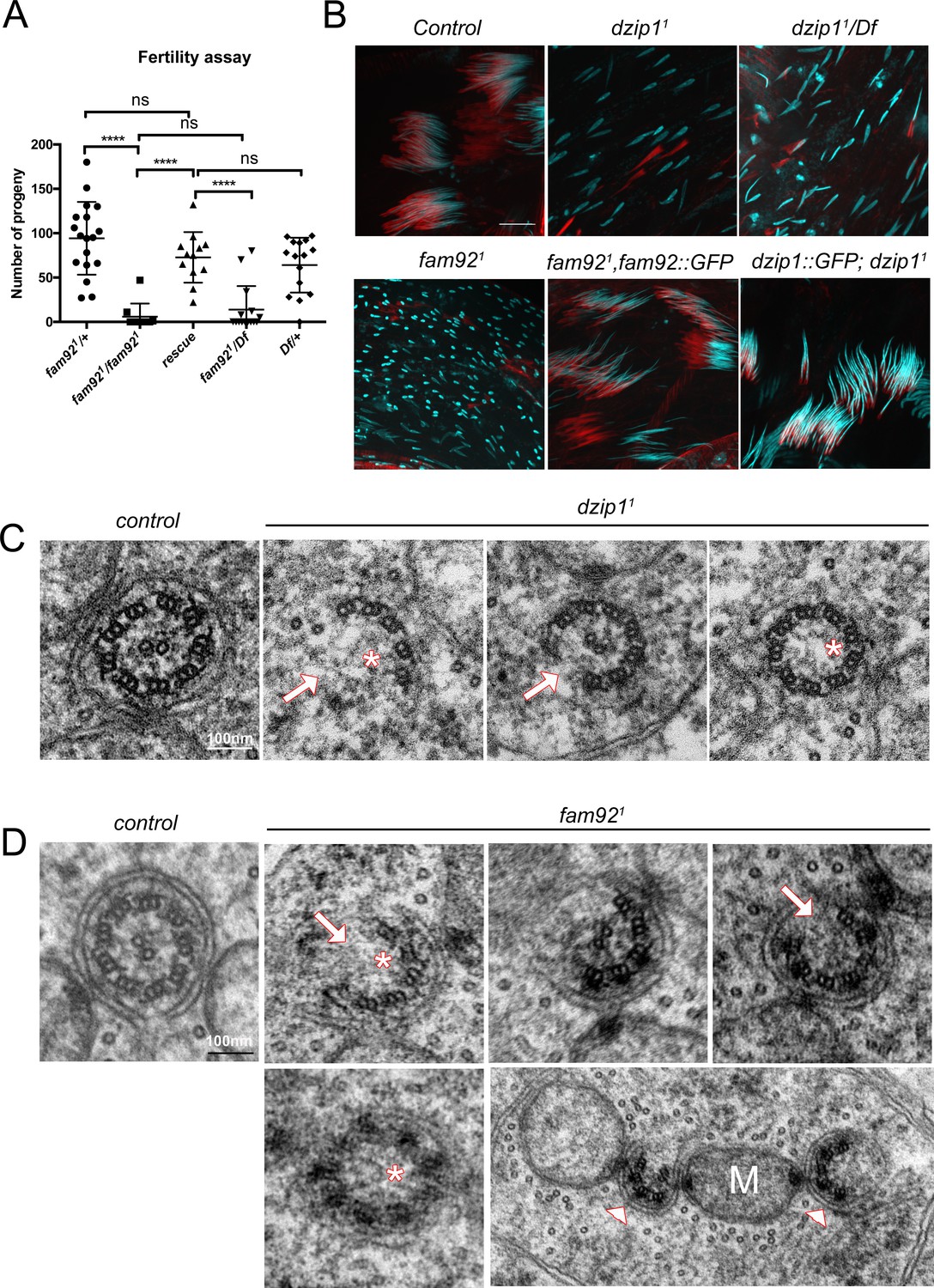
Dzip1 and Fam92 are required for flagella formation.
(A) Fertility assays of fam921 males showing severe reduction of fertility in fam921 or fam921/Df compared to control or rescued flies (fam921/+ n = 18; fam921/fam921 n = 10; rescue n = 15; fam921/Df n = 15; Df/+ n = 16) ****, p<0.0001; ns, p>0,05. (B) Confocal analysis of whole mount testes of control, dzip11, dzip11/Df, fam921 and rescued genotypes stained for nucleus (Hoechst, cyan) and actin cones (phalloidin, red). Nuclei are dispersed in mutant cysts compared to controls, which could reflect axonemal elongation defects. As a consequence, actin cones required for spermatid individualization are mislocalized. Axoneme elongation and spermatid individualization are normal in rescued flies. Bars = 10 μm. (C) EM analysis of cross sections of axoneme of round spermatid. In dzip11, flagella ultrastructure is altered with missing microtubule doublets (arrows) or central pair (asterisks). (D) In fam921 mutant spermatids, most axonemes show altered ultrastructure, with missing microtubule central pairs (asterisks) or doublets (arrows), broken symmetry with each part of the axoneme being relocated along the mitochondria (M, arrowheads).
Taken together, these results demonstrate that Fam92 and Dzip1 are necessary at the ciliary TZ for cilia formation both in sensory neurons and male germ cells.
Dzip1 and Fam92 form a strictly ordered functional module with Cby
To determine the functional hierarchy between Dzip1, Fam92 and Cby, we analyzed their respective localization in dzip11 or fam921 or previously described cby1 mutants (Figure 5 and Figure 5—figure supplement 1).
Figure 5 with 1 supplement see all
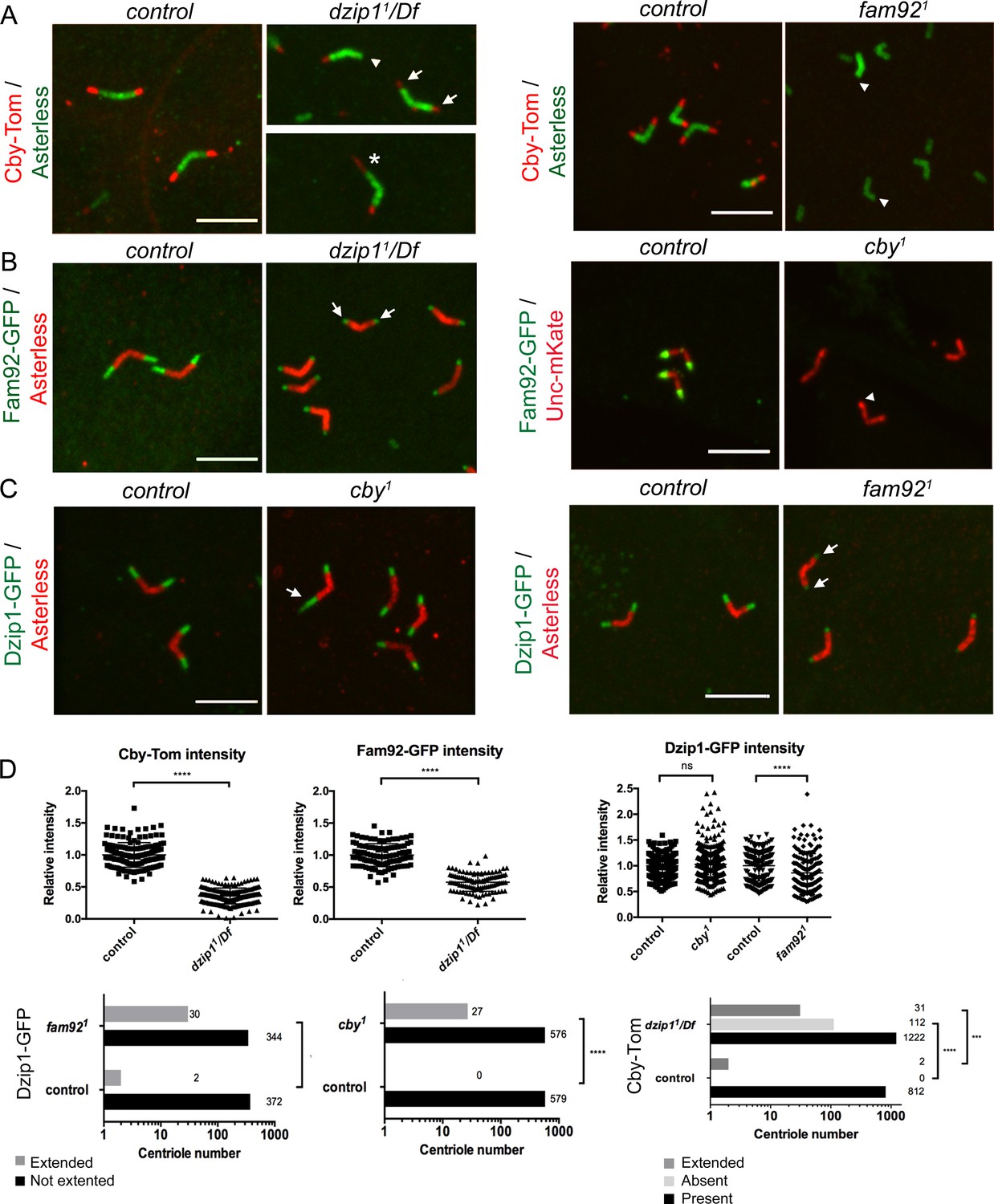
Dzip1, Fam92 and Cby work coordinately at the TZ.
(A) Cby-Tom is present at the tip of the centrioles in control spermatocytes but strongly reduced in dzip11/Df (arrows) and can even be lost at one centriole of the pair (arrowhead), but in a few situations (2.3% see D quantifications below), Cby-Tom domain is extended (asterisk). In fam921 mutant, Cby-Tom is completely lost at the tip of the spermatocyte centrioles (arrowheads). (B) Fam92-GFP is reduced at both centrioles of the pair in dzip11/Df (arrows) compared to control and completely lost at the tip of the centrioles (arrowhead) in cby1 mutants. (C) Dzip1-GFP is strikingly expanded in a few cby1 spermatocyte centrioles (arrow, 4.4% see quantifications in D) and is reduced at the tip of both centrioles in fam921 testes (arrows). Bars = 5 μm. (D) Quantifications of the distribution of Dzip1-GFP, Fam92-GFP and Cby-Tom in the different mutant contexts. ****, p<0.0001; ***, p<0.001; ns, p>0,05.
Cby-Tom is almost completely lost from ciliary TZ in dzip11/Df sensory neurons (Figure 5—figure supplement 1A). In spermatocytes, Cby-Tom signal is strongly reduced at the tip of centrioles (Figure 5A,D) but, in rare occasions, expanded at one centriole of the pair (asterisk, 2%). The amount of Fam92-GFP is severely reduced in dzip11/Df sensory cilia base (Figure 5—figure supplement 1B) and in spermatocytes (Figure 5B,D, arrows). Altogether, these observations indicate that Dzip1 is required at the basal body/TZ to recruit or stabilize both Cby and Fam92.
In fam921 mutant flies, Dzip1-GFP is slightly reduced at the distal tip of basal bodies in sensory cilia (Figure 5—figure supplement 1C). In fam921 spermatocytes, Dzip1-GFP domain is sometimes expanded (8.3% of cases, Figure 5D), but the overall Dzip1-GFP expression is also slightly reduced (Figure 5C, arrows, 5D). Cby-Tom is completely lost in both fam921 ciliated tissues (Figure 5A, arrowheads and Figure 5—figure supplement 1A). Hence, Fam92 is required to recruit Cby, but not Dzip1, at the TZ. As well, in cby1 tissues, Fam92-GFP completely disappears from basal bodies (Figure 5B, arrowhead and Figure 5—figure supplement 1B), indicating that Cby and Fam92 stabilize each other at the basal body. In cby1 mutant spermatocytes, an extended domain of Dzip1-GFP is occasionally observed (Figure 5C, arrows, 4.5%, 5D). However, no overall difference in Dzip1-GFP intensity was observed in cby1 mutant testes (Figure 5C–D) or antennae (not shown) compared to controls.
These observations establish a functional hierarchy for Dzip1, Fam92 and Cby: Dzip1 is required to recruit or stabilize both Fam92 and Cby at centrioles, but does not depend on Fam92 or Cby for its targeting to centrioles, the latter only restricting Dzip1 to the proximal TZ. Conversely, Fam92 and Cby mutually depend on each other to localize at the TZ.
Dzip1 and Fam92 restrict Cep290 to the proximal TZ
To understand how Dzip1 and Fam92 organize the ciliary base, we investigated their functional relationships with other core TZ components. We first investigated the behavior of Mks1, a component of the core conserved MKS complex (Weatherbee et al., 2009). In Drosophila, Mks1 is required to assemble the MKS complex, but removal of MKS components leads to only very mild defects of cilia assembly (Vieillard et al., 2016; Pratt et al., 2016). In dzip11/Df and fam921 mutant flies, Mks1-GFP is lost or reduced at the TZ in both ciliated tissues (Figure 6A, arrows, and Figure 6—figure supplement 1A). Dzip1 and Fam92 are hence important for the recruitment or stabilization of the MKS module at the TZ.
Figure 6 with 2 supplements see all
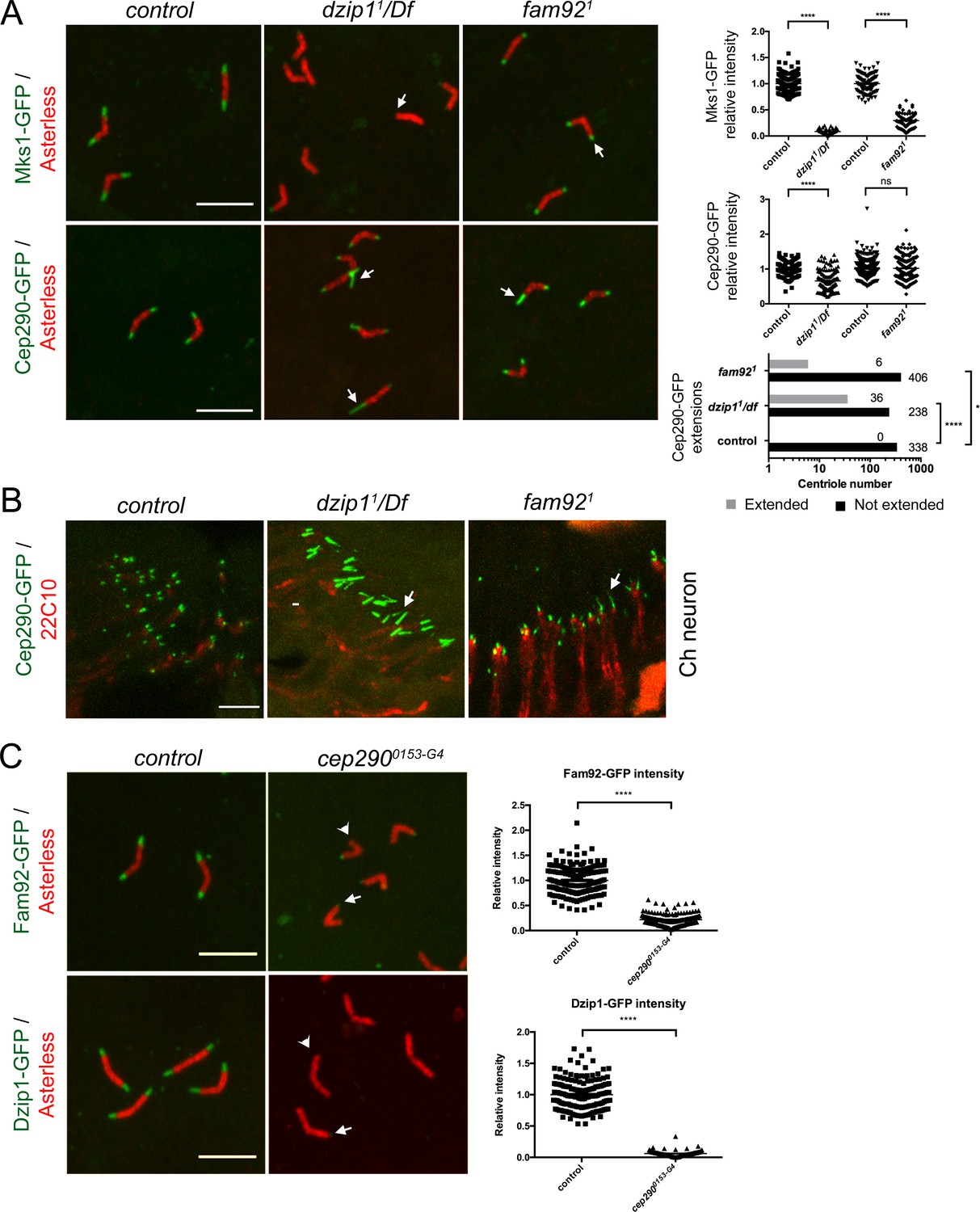
Dzip1 and Fam92 organize the transition zone downstream of Cep290.
(A) In spermatocytes, Mks1-GFP is severely reduced at the ciliary cap or lost in dzip11/Df and fam921 (arrows, quantifications on right graph). Cep290-GFP domain is expanded in dzip11/Df (arrows), but less affected in fam921 (quantifications on right graph). (B) Cep290-GFP is expanded in chordotonal cilia of dzip11/Df or fam921 antennae (arrows). (C) Fam92-GFP and Dzip1-GFP are strongly reduced (arrows) or lost (arrowheads) at the tip of the centrioles in cep2900153-G4 spermatocytes (quantifications are illustrated on the graphs). ****, p<0.0001; *, p<0.01; ns, p>0.05. Bars = 5 μm.
Cep290 is a core conserved TZ component which plays a prominent role in TZ assembly in many organisms, including Drosophila (Craige et al., 2010; Rachel et al., 2015; Li et al., 2016a; Basiri et al., 2014). In flies, Cep290 is located at the base of the TZ in sensory neurons and is a critical component of the migrating ring centriole during spermatogenesis (Basiri et al., 2014). In the absence of Dzip1, besides a small but significant reduction of Cep290-GFP on spermatocyte centrioles, we observed striking Cep290-GFP expanded domains, both in spermatocytes and in Ch neurons (Figure 6A–B, arrows). Around 13% of centrioles in spermatocytes and a majority in antennae showed an expanded Cep290-GFP domain (Figure 6A–B). In fam921 mutants, we also observed a few Cep290-GFP expanded domains in spermatocytes and antennae, but with no significant difference in overall Cep290-GFP intensity (Figure 6A–B, arrows).
To further understand the relationships between Cep290, Dzip1 and Fam92, we took advantage of a strong cep290 hypomorphic mutant, cep2900153-G4. This mutant shows severe uncoordination and completely disorganized spermatid cysts with dispersed nuclei (Figure 6—figure supplement 2A), which is a consequence of the severe axonemal elongation defects observed in this mutant (not shown). The phenotypes were completely rescued by two copies of cep290::GFP (Figure 6—figure supplement 2A–B). We observed that in cep2900153-G4 flies, Dzip1 and Fam92 are strongly reduced or lost at the TZ, both in spermatocytes (Figure 6C) and in sensory neurons (Figure 6—figure supplement 1B).
These observations demonstrate that Cep290 is required during TZ assembly to recruit Dzip1 and Fam92, which in turn restrict Cep290 scaffolding to the proximal part of the TZ.
Dzip1 and Fam92 facilitate basal body docking to the plasma membrane in spermatocytes
Defects in BB anchoring to the plasma membrane in spermatocytes lead to aberrant growth of axonemal microtubules (Vieillard et al., 2016). We used the specific axonemal marker CG6652-GFP that only labels axonemal microtubules in flies (Figure 6—figure supplement 2C–D). With this marker, we observed aberrant growth of axonemal microtubules in spermatocytes, with graded severities increasing from fam921, dzip11 to cep2900153-G4 mutant flies (Figure 7A–B). 30% of centrioles showed CG6652 extensions in fam921 testes, 34% in dzip11/Df and 76% in cep2900153-G4. More strikingly, whereas 68% of the centriole pairs present extensions from both centrioles in cep2900153-G4 spermatocytes, we observed that in fam921 testes, 96% of the centriole pairs present microtubule extensions on only one centriole. This asymmetric penetrance of the phenotype is related to centriole age. Indeed, among 24 centriole pairs for which mother and daughter identities could be unequivocally assigned, 19 daughter centrioles (79%) and only five mother centrioles (21%) were affected. As well, in dzip11/Df testes, among 40 centriole pairs, we found 30 daughter centrioles (75%) and 10 mother centrioles (25%) affected by the absence of Dzip1 (Figure 7A–B).
Figure 7
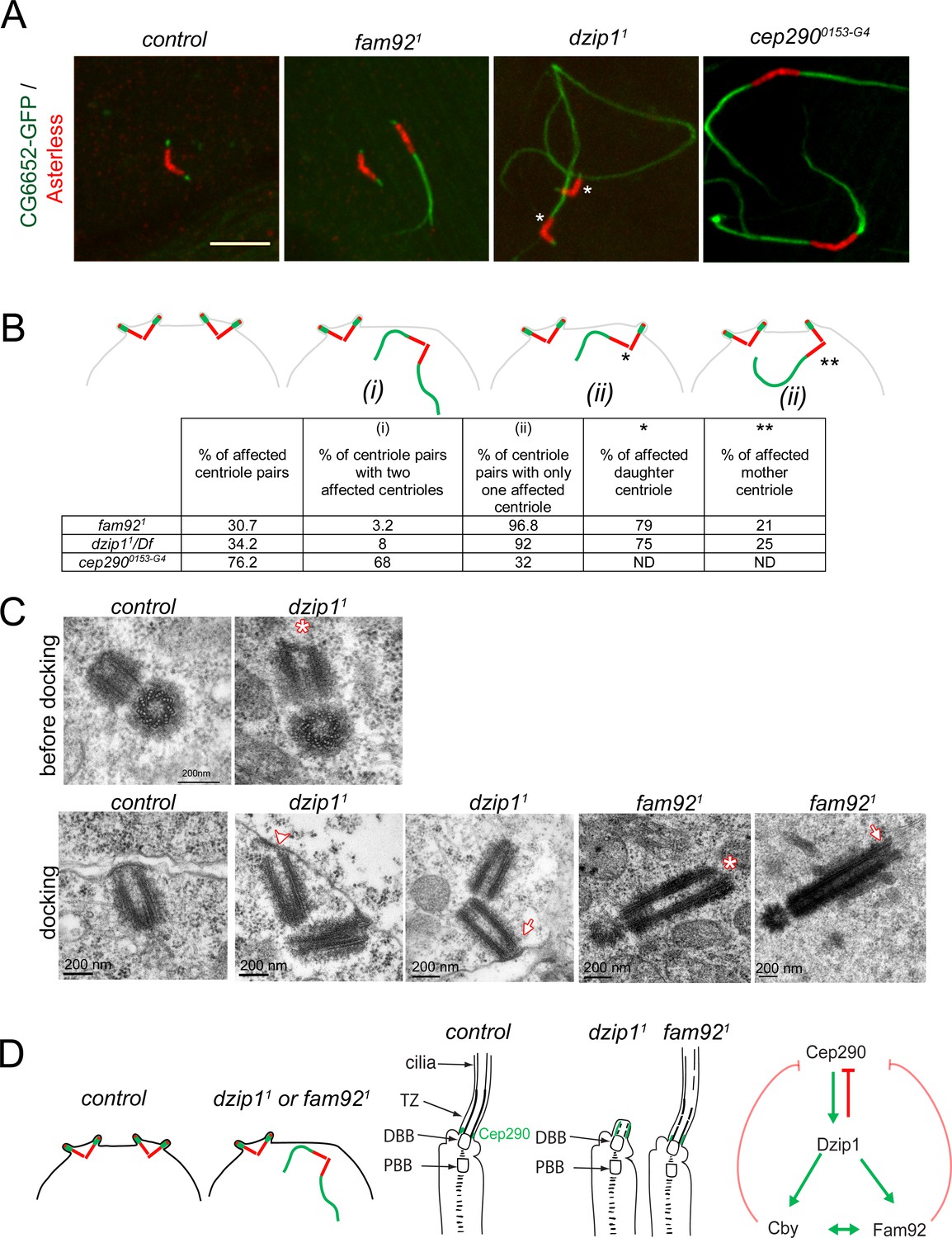
Dzip1 and Fam92 are required for both centriolar docking and proper axonemal formation during Drosophila spermatogenesis.
(A) Confocal imaging of whole-mount testes showing aberrant extensions of axonemes labeled by CG6652-GFP in fam921, dzip11 and cep2900153-G4 spermatocytes. Asterisks point to the centriole in each pair that can be unambiguously assigned as the daughter. Bar = 5 μm (B) Quantifications of the penetrance of CG6652-GFP labeled aberrant axonemes. Schemes representing the different classes that were quantified in the table: (i) two centrioles of the pair are affected; (ii) only one centriole of the pair is affected, either the daughter* or the mother**. (C) EM analysis of cross and longitudinal sections of centrioles in spermatocytes. In dzip11, centriole docking is impaired with partial docking of the centriole (arrowhead) or docking to the plasma membrane of only one centriole of the pair (arrow). Centrioles present an altered/irregular cap, compared with control (asterisk). In fam921, undocked centrioles are also observed in spermatocytes and show irregular distal end (asterisk) or microtubules extending from the tip (arrow). (D) Scheme of the consequences of dzip1 and fam92 loss of function on basal body and cilium assembly in Drosophila ciliated tissues. Whereas basal body anchoring and TZ assembly is affected in male germ cells, only TZ and cilium assembly is affected in sensory cilia. In addition, Cep290-GFP is systematically expanded in dzip1 mutant and occasionally expanded in fam92 mutant chordotonal neurons. The scheme on the right summarizes the functional interactions observed between Dzip1, Cby, Fam92 and Cep290. Red arrows are inhibitory interactions and green arrows define positive interactions.
We analyzed the ultrastructure of the centrioles and primary like cilium in the initial stages of elongation (Figure 7C). We observed irregular distal end (asterisk) of centrioles in dzip11 testes at young spermatocytes stages before docking (upper panel). Whereas centrioles dock in polar spermatocytes before reaching their full size in control testes, undocked centrioles were observed in dzip11. Either both centrioles of the pair were undocked or partially docked (n = 2, lower panel) (Figure 7C, arrowhead) or only one centriole of the pair, the mother, was docked (n = 3) (Figure 7C, arrow). These results show that Dzip1 is required to cap the centriole distal end and foster its anchoring to the plasma membrane. As well, we observed undocked centrioles in fam921 mutant spermatocytes, with irregular distal end (asterisk) or with microtubules extending from the distal end (Figure 7C, arrow), illustrating the role of Fam92 in controlling centriolar distal growth and docking to the plasma membrane.
Altogether, these observations demonstrate that Dzip1 and Fam92 are required for the proper distal elongation of basal bodies and their membrane anchoring in Drosophila spermatocytes. This docking is required to regulate TZ elongation (Figure 7D). In addition, our results show that in male germ cells, although all centrioles have the capacity to generate a ciliary cap, a functional asymmetry of the mother and the daughter centrioles is revealed by their ability to dock to the plasma membrane in absence of key TZ proteins.
Discussion
Our work establishes the functional hierarchy of Dzip1, Fam92 and Cby, which thus define an intrinsic transition zone module for the initiation of ciliogenesis in Drosophila. Dzip1 is required to recruit Fam92 and Cby at the basal body distal end, allowing proper TZ assembly. Dzip1 and Fam92 act downstream of the core TZ protein Cep290 but also regulate its accumulation at the distal basal body. Our work sheds light on tissue specific variations in the initiation of ciliogenesis in the two Drosophila ciliated tissues. In testes, basal body docking strictly depends on the integrity of Dzip1 and Fam92. However, in sensory neurons alterations of the latter complex affect only TZ assembly and cilia formation but not BB docking. Last, our work demonstrates that Dzip1 and Fam92 control the distal elongation of basal bodies and their docking to the plasma membrane, which thus regulate the onset and proper elongation of axonemal microtubules in Drosophila male germ cells.
Conservation of TZ assembly pathways in eukaryotes
In vertebrates, two orthologs have been described for Dzip1 and Fam92 and three for Cby. DZIP1, DZIP1L, FAM92a and b also interact with CBY1 in vertebrates (Wang et al., 2018; Breslow et al., 2017; Ye et al., 2014) indicating a conservation of the interacting capacities of the family members during evolution. The precise hierarchy between members of the complex has not been established in mammals, but depletion of CBY1 prevents the recruitment of FAM92a/b to the centriole (Li et al., 2016b) and DZIP1L was shown to act upstream of CBY1 (Wang et al., 2018; Keller et al., 2009).
In vertebrates, mutations in members of this module lead to ciliogenesis defects of various severities and with different phenotypic outcomes (Wolff et al., 2004; Tay et al., 2010; Glazer et al., 2010; Li et al., 2016b; Breslow et al., 2017) Dzip1 or Dzip1L KO mice die during embryogenesis (Wang et al., 2018; Wang et al., 2013), whereas Fam92a-/- mice show skeletal defects (Schrauwen et al., 2019) and Cby1 KO mice exhibit differentiation defects of motile ciliated epithelia (Burke et al., 2014; Voronina et al., 2009). This phenotypic variability is not expected if all three proteins only act together in a functional module at the TZ, as demonstrated by our work in Drosophila. This suggests that the different mouse paralogs of Cby and Fam92 may have acquired specialized ciliogenic functions in mouse. However, we also observed that dzip11 and fam921 mutant phenotypes show small differences indicating specific functions of each proteins. For instance, in both ciliated tissues, dzip11 hypomorphic mutant phenotype is more severe than fam921, suggesting that Dzip1 has additional functions that are not solely mediated by Fam92.
Despite the conserved role of Dzip1, Cby and Fam92 in TZ and cilia assembly from Drosophila to humans, no homologs could be detected in the genomes of C. elegans. This could be linked to the diversification of cilia function, with both motile and immotile cilia being present from Drosophila to humans, in contrast to C. elegans, where only one subtype of cilia that are immotile is found. Another possible explanation could be that DZIP1/FAM92/CBY are associated with specific signaling or developmental functions in animals that still need to be understood.
Tissue specific functional properties of the TZ proteins in BB docking
This work emphasizes the essential role of Dzip1 and Fam92 in building the ciliary transition zone in the two types of ciliated tissues of Drosophila. Strikingly, it also reveals tissue specific function of these proteins in priming basal body/membrane docking in Drosophila testes. This reveals intrinsic differences in the mechanisms that link basal body to membranes in Drosophila ciliated tissues. In mammals, basal body docking requires transition fibers built from the distal appendages on the mother centriole prior to docking (Wei et al., 2015). In Drosophila, distal appendages have not been observed on centrioles, but structures similar to transition fibers are described at the base of sensory cilia (Ma and Jarman, 2011; Vieillard et al., 2016), whereas only scarce links could be observed in male germ cells (Gottardo et al., 2018; Roque et al., 2018). These differences could explain why destabilization of the TZ leads to basal body anchoring defects in spermatocytes, but not in sensory neurons. This structural characteristic of the spermatocyte TZ is likely to be related to its specific functional properties. Indeed, whereas the TZ is stably built at the ciliary base in sensory neurons, it shows a dynamic behavior during sperm flagella extension, separating from the basal body and migrating along the growing end of the axoneme (Basiri et al., 2014; Avidor-Reiss et al., 2017; Baker et al., 2004; Wei et al., 2008). In addition, in spermatocytes, basal bodies have a dynamic behavior, being first docked at the plasma membrane and next internalized during meiosis (Pasmans and Tates, 1971; Fabian and Brill, 2012). This could induce mechanical constraints on basal bodies that would increase their sensitivity to TZ disruption. In agreement with this hypothesis, when TZ maturation was challenged by modulating membrane phospholipids (Gupta et al., 2018), BB were released from the plasma membrane during meiosis, but their initial docking was not impaired. However, we observed by EM, that BB fail to initially dock in significant occurrences (8 among 13) in dzip11 and fam921 mutant spermatocytes, indicating that Dzip1 and Fam92 are at least involved in the initial steps of BB docking. Previous observations of another strong hypomorphic cep290mecH allele showed docked basal bodies in spermatocytes and spermatids (Basiri et al., 2014). In cep2900153-G4 mutant, we did not quantify the number of docked versus undocked basal bodies in spermatocytes, but we observed up to 76% of aberrant axonemal growth, suggesting that basal body to membrane attachment is compromised in this mutant.
Differences in the organization of the ciliary base associated with variations in the distribution and function of several centriolar and TZ proteins have been documented in Drosophila ciliated cells (Jana et al., 2018). However, none of these identified differences help to explain the behavioral properties of BB docking and TZ dynamics that we have identified in the two ciliated Drosophila tissues. Hence, additional screens for specific factors of basal-body docking or TZ assembly either in sensory neurons or male germ cells need to be designed.
Mother and daughter centriole asymmetry in ciliogenesis
In all our observations, there is a striking phenotypic difference between the mother and daughter centrioles in spermatocytes. In all mutants examined (i.e. dzip1, fam92 and cby) we observed a more penetrant defect on the daughter centriole than the mother. Thus, although the 2 centrioles of each pair are able to form cilia, the daughter centriole appears more sensitive to transition zone perturbations. We do not have molecular explanations for these intrinsic differences of mother and daughter centrioles in spermatocytes. In Drosophila sensory neurons, centrobin plays a critical role in maintaining the daughter centriole fate, precluding its capacity to build a cilium (Gottardo et al., 2015). In sperm cells, centrobin is required for the formation of the C tubule (Reina et al., 2018), which also plays a critical role in TZ assembly (Gottardo et al., 2018). However, in spermatocytes centrobin is equally distributed at the base of both the mother and daughter centrioles and does not show a functional asymmetry. Recently, a transient microtubule based structure that anchors the base of the mother centriole on one of the centriole pair at the onset of meiosis was identified, but its function is unknown (Riparbelli et al., 2018). This structure could stabilize the mother centriole and favor its attachment to the membrane, but the function of this microtubule rootlet in Drosophila spermatocytes needs further investigations. The intrinsic difference of the mother versus daughter centrioles could also be related to the timing of centriole docking during spermatogenesis (Gottardo et al., 2018), where the mother centriole was shown to dock first. This timing difference would be sufficient to better stabilize the TZ of the mother centriole, and hence explain the phenotypic differences observed in our study. There are other situations in the animal kingdom were both mother and daughter centrioles build a cilium. Among the best studied are the bi-flagellated Chlamydomonas and the peculiar case of multiple ciliated epithelia, where numerous de novo centrioles are assembled just at the onset of ciliogenesis. Interestingly, in mammals, CBY1 was shown to play a critical role in multiple ciliated cells to allow proper docking of the multiple basal bodies to the plasma membrane (Burke et al., 2014). It is tempting to speculate that the increased susceptibility of multiple ciliated cells to CBY1 loss, is related to a particular status of these de novo centrioles, as we observed for daughter centrioles in male germ cells. More work will be needed to understand the molecular determinants of the mother versus daughter centriolar functional asymmetry in Drosophila male germ cells.
Specific regulatory role of the ciliary cap in Drosophila male germ cells
We observed that defects of TZ assembly and/or basal body docking lead to aberrant elongation of axonemal microtubules as revealed by the specific Drosophila axonemal marker CG6652-GFP. These abnormal elongation defects appear only in late G2 phase, just at the onset of meiosis, indicating that specific signals, yet to be identified, enable centrioles to start axonemal elongation at the onset of meiosis. The membrane cap restricts this capacity by inhibiting axonemal growth until the round spermatid stage, where a second signal turns on axonemal elongation and TZ migration. Among the candidate proteins recruited by the membrane cap which may coordinate axonemal assembly are microtubule depolymerizing kinesins as previously proposed (Vieillard et al., 2016). This membrane cap could also be involved in stabilizing centriolar capping proteins such as CP110 or CEP97, which in mammals need to be removed from centrioles to allow ciliary assembly (Spektor et al., 2007). However, there are no clear evidence in Drosophila that these proteins need to or are specifically removed before axonemal elongation (Galletta et al., 2016; Franz et al., 2013; Delgehyr et al., 2012). Our observations indicate that regulation of axonemal assembly and cell cycle regulation are tightly linked in these dividing cells, but the effectors of this control still need to be identified.
In conclusion, our work demonstrates the critical role of the conserved Dzip1/Fam92/Cby module downstream of Cep290 in initiating the assembly of the ciliary transition zone in flies. It also reveals key tissue specific differences in basal body docking pathways in Drosophila.
Materials and methods
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene (Drosophila melanogaster) | cby | Enjolras et al., 2012 | FLYB: FBgn0067317 | |
| Gene (Drosophila melanogaster) | fam92/CG6405 | This study | FLYB: FBgn0032428 | |
| Gene (Drosophila melanogaster) | dzip1/CG13617 | This study | FLYB: FBgn0039201 | |
| Strain, strain background (Escherichia coli) | DH5alpha | Thermo Fisher Scientific | 18265017 | |
| Genetic reagent (D. melanogaster) | Cep2900153-G4 mutant strain | Bloomington Drosophila Stock Center | BDSC: 62671; FLYB: FBst0062671 RRID:BDSC_62671 | FlyBase symbol: w1118; PBac{IT.GAL4}cep2900153-G4 |
| Genetic reagent (D. melanogaster) | Df(3R)Exel8178 | Bloomington Drosophila Stock Center | BDSC : 7993 FBab0038335 RRID:BDSC_7993 | |
| Genetic reagent (D. melanogaster) | Df(2L)Exel6033 | Bloomington Drosophila Stock Center | BDSC : 7516 FLYB : FBab0037871 RRID:BDSC_7516 | |
| Genetic reagent (D. melanogaster) | sas4s2214 mutant strain | Basto et al., 2006 | BDSC : 12119 FLYB : FBal0196943 RRID:BDSC_12119 | |
| Genetic reagent (D. melanogaster) | cby1 mutant strain | Enjolras et al., 2012 | FLYB : FBal0270281 | |
| Genetic reagent (D. melanogaster) | dzip11 mutant strain | This study | Section Materials and methods “Generation of dzip11 and fam921 alleles” | |
| Genetic reagent (D. melanogaster) | fam921 mutant strain | This study | Section Materials and methods “Generation of dzip11 and fam921 alleles” | |
| Genetic reagent (D. melanogaster) | Unc::mkate | Vieillard et al., 2016 | FLYB : FBal0324713 | |
| Genetic reagent (D. melanogaster) | Cby::Tom | Vieillard et al., 2016 | FLYB : FBal0270280 | |
| Genetic reagent (D. melanogaster) | Cep290::GFP | Basiri et al., 2014 | FLYB : FBal0301636 | |
| Genetic reagent (D. melanogaster) | CG6652::GFP | Vieillard et al., 2016 | FlyB: FBal0324714 | |
| Genetic reagent (D. melanogaster) | mks1::GFP | Vieillard et al., 2016 | FLYB : FBal0324710 | |
| Genetic reagent (D. melanogaster) | fam92::GFP | This study | Section Materials and methods “Plasmids and Drosophila reporter gene constructs” | |
| Genetic reagent (D. melanogaster) | dzip1::GFP | This study | Section Materials and methods “Plasmids and Drosophila reporter gene constructs” | |
| Cell line (Mus musculus) | IMCD3 | ATCC | CRL-2123 | |
| Cell line (Homo-sapiens) | HEK293 | ATCC | CRL-1573 | |
| Cell line (Cercopithecus aethiops) | COS-7 | ATCC | CRL-1651 | |
| Antibody | Mouse monoclonal anti-Futsch/22C10 | DHSB | AB_528403 | IF (1:250) |
| Antibody | Rabbit polyclonal anti-GFP | Abcam | AB6556 | IF(1:1000) WB (1:10000) |
| Antibody | Guinea pig polyclonal anti-Asterless | Klebba et al., 2013 | IF (1:50000) | |
| Antibody | Rat polyclonal anti-Asterless | McLamarrah et al., 2018 | IF (1:50000) | |
| Antibody | Rat monoclonal anti-HA (clone 3F10) | Roche | 11867423001 | WB (1:5000) |
| Antibody | Goat polyclonal anti-rabbit Alexa 488 | Invitrogen | A11008 | IF (1:1000) |
| Antibody | Goat polyclonal anti-rabbit Alexa 647 | Invitrogen | A21244 | IF (1:1000) |
| Antibody | Goat polyclonal anti-guinea pig Alexa 488 | Invitrogen | A11073 | IF (1:1000) |
| Antibody | Goat polyclonal anti-guinea pig Alexa 594 | Invitrogen | A11076 | IF (1:1000) |
| Antibody | Donkey polyclonal anti-guinea pig Alexa 647 | Invitrogen | 706-605-148 | IF (1:1000) |
| Antibody | Goat polyclonal anti-rat Alexa 488 | Invitrogen | A11006 | IF (1:1000) |
| Antibody | Goat polyclonal anti-rat Alexa 555 | Invitrogen | A21434 | IF (1:1000) |
| Antibody | Goat polyclonal anti rat Alexa 647 | Invitrogen | A21247 | IF (1:1000) |
| Antibody | Mouse monoclonal anti-V5 | Invitrogen | R960-25 | WB (1:5000) |
| Antibody | Goat polyclonal anti-rabbit-HRP | Biorad | 170–6515 | WB (1:10000) |
| Antibody | Goat polyclonal anti-mouse-HRP | Biorad | 170–6516 | WB (1:3000) |
| Antibody | Goat polyclonal anti-rat-HRP | Sigma | A5795-1ML | WB (1:20000) |
| Antibody | Mouse monoclonal anti-γ-Tubulin | Sigma | GTU88 | IF (1:500) |
| Chemical compound, drug | phalloidin FluoProbes 547 | Interchim | FP-AZ0330 | IF (1:200) |
| Chemical compound, drug | phalloidin FluoProbes 505 | Interchim | FP-AZ0130 | IF (1:200) |
| Commercial assay or kit | GFP-TRAP | Chromotek | gta-100 | |
| Commercial assay or kit | Mouse monoclonal Anti-HA-agarose antibody | Sigma | A2095 | |
| Commercial assay or kit | S-protein agarose | Merck | 69704 | |
| Commercial assay or kit | Gibson Assembly Master Mix | New England Biolabs, Inc | E5510S | |
| Recombinant DNA reagent | pBFv-U6.2 | Kondo and Ueda, 2013 Nig-Fly | FLYB: FBmc0003127 | |
| Recombinant DNA reagent | pBFv-U6.2B | Kondo and Ueda, 2013 Nig-Fly | FLYB: FBmc0003128 | |
| Recombinant DNA reagent | pG-LAP3 vector | Torres et al., 2009 | Addgene#79704 | |
| Recombinant DNA reagent | pEGFP-N1 | Clontech | Cat #6085–1 |
All primer sequences are described in Supplementary file 2.
Plasmids and Drosophila reporter gene constructs
Request a detailed protocolCBY1 coding sequence was PCR amplified from mouse ependymal primary cell cDNA with primers F-CBY1 and R-CBY1. PCR product was cloned into pDONR223 (Invitrogen) and then subsequently Gateway cloned into the pG-LAP3 vector (Torres et al., 2009; gift from P. Jackson, Addgene#79704). This vector contains the double EGFP-TEV-S peptide tag in N- terminus allowing a two step affinity purification.
Coding sequences of Drosophila cby, fam92/CG6405 and dzip1/CG13617 were obtained by PCR on cDNA from testis. Cby cDNA was cloned in pEGFP-N1 (Clontech) in frame with GFP (primers F- CbyGFP/R-CbyGFP) and in pCDNA3.1-HA (gift from S. Khochbin, Institut Albert Bonniot, Grenoble ; primers F-HACby/R-HACby). cDNA of fam92/CG6405 and dzip1/CG13617 were cloned in pCDNA3.1-HA with primers F-Fam92HA/R-Fam92HA and F-Dzip1HA/R-Dzip1HA, respectively. pCDNA3.1-GFP, GFP-Fam92 and GFP-Dzip1 were obtained by replacement of the HA tag with EGFP. V5-Fam92 was obtained by replacement of the HA tag with V5 tag.
Drosophila reporter gene construct of dzip1/CG13617 was obtained by cloning 1.6 kb upstream regulatory sequences and the entire coding sequence (primers F-Dzip1GFPrep/R-Dzip1GFPrep) in frame with GFP of pJT61, a pattB plasmid with an extra EGFP-6xMycTag-SV40polyA cassette (Vieillard et al., 2016). The 4 kb fragment of fam92/CG6405 including 1.4 kb of upstream regulatory sequences, the entire coding sequence fused to the multipurpose tag cassette including GFP and the 3'UTR was obtained by PCR with primer F-Fam92GFPrep/R-Fam92GFPrep on fosmid from the FlyFos library (Sarov et al., 2016). The resulting constructs were integrated in the 53B2 VK00018 landing site on the second chromosome by PhiC31 integrase (BestGene). All transgenic lines were obtained from BestGene Inc.
Generation of dzip11 and fam921 alleles
Request a detailed protocolThe dzip11 allele (CG13617) was generated by CRISPR/Cas9 induced homologous directed repair (Gratz et al., 2015). Two gRNA were selected using the http://crispor.tefor.net/crispor.py website: 5’- CCCGTTTCACGGACCATCTG CGG −3’ and 5’-GTTTCCAGCACTGTGCCCAG TGG −3’ (protospacer adjacent motifs are underlined). Oligos were phosphorylated by T4PNK (New England Biolabs, Inc) and annealed. Double-stranded 5’ gRNA and 3’ gRNA were cloned in the BbsI site of pBFv-U6.2 and pBFv-U6.2B vectors, respectively (Kondo and Ueda, 2013). 5’ gRNA was further subcloned in the EcoRI–NotI sites of pBFv-U6.2B to express the two gRNAs from one vector.
The 5’ and 3’ homology arms (1.7 kb and 2.3 kb respectively) were amplified by PCR (primers F-5’armDzip1/R-5’armDzip1 and F-3’armDzip1/R-3’armDzip1) and cloned, respectively, into the pJT38 plasmid (pRK2 plasmid [Huang et al., 2008], with an attB cassette) using Gibson Assembly Master Mix (New England Biolabs, Inc). The two vectors (gRNAs and homology arms) were injected into vasa::Cas9 embryos. Flies were crossed to w; TM2, e Ubx/TM6B, e Hu Tb virgin females and the offspring were screened for red-eyed-flies. Homologous recombination was checked by PCR.
The fam921 allele (CG6405) was generated by CRISPR/Cas9 induced deletion. Two gRNA were selected as before: 5’- CATAAGACCTTGCAGATATC GGG −3’ and 5’- GGCTGTCATAGCGCGGGATA AGG −3’ (protospacer adjacent motifs are underlined). Double-stranded phosphorylated 5’ gRNA and 3’ gRNA were cloned into the pBFv-U6.2B plasmid. The vector was integrated into the 89E11 VK00027 landing site on the third chromosome by PhiC31 integrase (BestGene). Surviving G0 males were individually crossed to y2 v1 virgins. A single male transformant from each cross was mated to y2 cho2 v1; Pr Dr/TM6C, Sb Tb virgins.
Males carrying a U6-double gRNA transgene were crossed to nos-Cas9 females to obtain male founder animals. Each male founder was crossed to three virgin females y2 cho2 v1 ; Sco/CyO. Deletion in the fam92 locus was characterized by PCR with primers F-Fam92KO/R-Fam92KO on genomic DNA from the resultant offspring. Flies showing a 294 bp deletion in the fam92 locus were selected for further studies.
Cell culture and transfection
Request a detailed protocolAll reagents for cell culture were purchased from Thermo Fisher Scientific. IMCD3 cells (murine Inner Medullary Collecting Duct cells, ATCC CRL-2123 a gift from A. Benmerah, Institut Imagine, Paris) were cultured in DMEM/HAM'S F12 medium supplemented with 10% FBS, 1X non-essential amino acids, 100 U/ml penicillin-streptomycin and 100 µg/ml hygromycin. COS-7 cells (ATCC CRL-1651) or HEK293 (ATCC CRL-1573) were maintained in DMEM medium containing 10% FBS, 1X non-essential amino acids and 100 U/ml penicillin-streptomycin. Cells were tested negative for mycoplasma.
Transfections were performed using jetPRIME (Polyplus transfection) according to the manufacturer’s instructions.
Stable cell lines
Request a detailed protocolIMCD3 cell lines expressing LAP-CBY1 or control LAP-GFP were created using the Flp-In system kit (Invitrogen) and established method (Torres et al., 2009).
Tandem affinity purification and mass spectrometry of CBY1-interacting proteins
Request a detailed protocolTwo rounds of purification of CBY1-protein complexes were performed. Cells were seeded in 35 15 cm dishes and grown to confluence before serum starvation for 24 hr to induce primary cilia formation. Cells were washed once with ice-cold PBS1X and purification was performed as described previously (Nachury, 2008). Briefly, cells were collected by scraping and resuspended in 11 ml lysis buffer containing HEPES pH7.5 50 mM, EGTA pH8 1 mM, MgCl2 1 mM, KCl 300 mM, Glycerol 10%, NP-40 0.31% and 1X protease inhibitor cocktail (Roche). CBY1-complexes were purified using GFP-TRAP (Chromotek). S-Tag-CBY1 was cleaved off GFP beads by TEV cleavage and the eluate was further purified on S-protein agarose (Merck) and eluted in 100 μl 2X Laemmli Sample Buffer.
For protein identification, samples were separated by 10% SDS-PAGE. Gels were stained with Coomassie and cut into four slices. Each gel slice was washed, reduced with 10 mM DTT, alkylated with 55 mM iodoacetamide, and subjected to in-gel trypsin digestion overnight (Trypsin Protease, MS Grade Pierce, Thermo Fisher Scientific). The extracted tryptic peptides were cleaned up using OMIX C18 100 µl pipette tips (Agilent), and lyophilized before being reconstituted for the LC-MS/MS analysis.
The peptides were separated using an Eksigent Ultra nano-LC HPLC system coupled with an AB Sciex Triple TOF 5600 mass spectrometer. The LC separations were performed using a Discovery Bio Wide Pore HPLC column (C18, 3 µm, 100 × 5 mm). The mobile phases used were 0.1% formic acid in water (A) and 100% acetonitrile with 0.1% formic acid. The gradient elution steps were performed with a flow rate of 5 µl/min as follows: 0–40% B for 106 min, 40–80% for 5 min, and then 80% B for an additional 5 min. All data were acquired using Analyst software (AB Sciex) in the data dependent mode. Peptide profiling was performed using a mass range of 350–1600 Da, followed by a MS/MS product ion scan from 100 to 1500 Da. For each survey MS1 scan (accumulation time of 250 msec), MS/MS spectra (accumulation time of 75 msec per MS/MS) were obtained for the 30 most abundant precursor ions. The protein identification was performed with the ProteinPilot Software 5.0 (AB Sciex). The MS/MS spectra obtained were searched against the mouse UniProt Proteome database (release 2015_09 with 46470 proteins). The search parameters for tryptic cleavage and accuracy are built-in functions of the software. The other data analysis parameters were as follows: sample type: identification; Cys-alkylation: Iodoacetamide; Digestion: Trypsin; Instrument: TripleTOF 5600; Special factor: gel based ID, and biological modifications; Species: Homo sapiens; Search effort: Thorough ID. Proteins comprising one or more peptides with a high confidence score (95%) and a low false discovery rate (FDR) (estimated local FDR of 1%) were considered positively identified.
Co-IP and western blotting
Request a detailed protocolCo-IP assays were performed using transfected COS-7 cells or HEK293 cells, harvested in ice-cold PBS1X. Cell pellet was resuspended in either lysis buffer (NaCl 150 mM, Tris-HCl pH 7.2 50 mM, NP-40 1%, Desoxycholate Na 1%) or milder lysis buffer (NaCl 150 mM, Tris-HCl pH 7.2 50 mM, NP-40 0.5%, glycerol 10%) with 1X protease inhibitor cocktail (Roche) and incubated for 1 hr at 4°C under agitation. Cell lysates were cleared by centrifugation at 15 000 g for 20 min at 4°C. Supernatants were incubated with either GFP-TRAP (Chromotek) or HA-coupled beads (Sigma) as indicated for 1 or 2 hr at 4°C. The beads were collected and washed five times with 1 ml of ice-cold lysis buffer before SDS-PAGE. Eluates were loaded on 10 or 12% SDS-PAGE. After migration, proteins were transferred onto a P 0.45 µm PVDF or 0.45 µm nitrocellulose Amersham Hybond membranes (GE Healthcare) and immunoblotted with according antibodies. IP were repeated three times with anti-GFP for Cby-GFP/HA-Fam92 and once each (anti-HA or anti-GFP) for Cby-GFP/HA-Dzip1 and IP was performed once with anti-GFP for GFP-Dzip1/V5-Fam92/HA-Cby.
Primary antibodies used for western blotting were the following: rat anti-HA (1:5000; Roche), rabbit anti-GFP (1:10000; Abcam) and mouse anti-V5 (1:5000; Invitrogen). HRP-conjugated secondary antibodies were the following: goat anti-rabbit (1:10000; Biorad), goat anti-mouse (1:3000; Biorad) and goat anti-rat (1:20000; Sigma). Membranes were visualized using ECL prime from GE Healthcare.
Fly stocks and maintenance
Request a detailed protocolThe cby1 mutant and cby::Tomato transgene were previously described (Enjolras et al., 2012). The cep290::GFP transgene was a gift from T. Avidor- Reiss (University of Toledo, USA; Blachon et al., 2009; Basiri et al., 2014). CG6652::GFP, mks1::GFP and unc::mKate transgenes have been described in Vieillard et al. (2016). sas4s2214 mutant flies were kindly provided by R. Basto (Basto et al., 2006). Flies were raised on standard media between 21°C and 25°C. Df(3R)Exel8178 (dzip1 deficiency), Df(2L)Exel6033 (fam92 deficiency) were obtained from the Bloomington Stock Center and uncover the 95F8-−96A6 and 33E4-−33F2 cytological interval respectively. cep2900153-G4 (Bloomington Drosophila Stock Center) harbors a piggy-bac insertion in the beginning of exon 11.
Immunofluorescence and confocal microscopy
Request a detailed protocolTestes from young adult flies or pupae were dissected in PBS1X, fixed 15 min in PBS1X/PFA 4% and either whole mounted or squashed between coverslip and slide and frozen in liquid nitrogen. Coverslip was removed and slides were soaked 2 min in ethanol 100% at −20°C. Testes were treated 15 min in PBS1X/Triton 0.1% (PBT) and blocked 2 hr in PBT/BSA 3%/NGS 5%.
Primary antibodies were incubated in blocking buffer overnight at 4°C. Samples were then washed in PBS1X and incubated 2 hr in secondary antibodies diluted in PBS1X. Slides were washed in PBS1X and rinsed in ultrapure water. Slides were mounted using Vectashield containing Hoescht 1:1000.
Antennae were processed as previously described (Vieillard et al., 2015). Briefly, Drosophila heads from 38 to 45 hr pupae were dissected in PBS1X, fixed 1 hr in PBS1X/PFA 4% and washed in PBS1X. Antennae were permeabilized 1 hr in PBS1X/Triton 0.3%, blocked 1 hr in PBS1X/Triton 0.3%/BSA 3%/NGS 5% and incubated in primary antibodies diluted in blocking solution for 48 hr at 4°C. Samples were washed three times in PBS1X and incubated in secondary antibodies diluted in PBS1X for 48 hr at 4°C. Antennae were washed three times in PBS1X and mounted in Vectashield.
For the whole mount staining of antennae, Drosophila heads from 38 to 45 hr pupae were dissected in PBS1X/0.3% Triton X-100 and fixed in in PBS1X/0.3% Triton X-100/4% PFA for 1 hr at RT. After a few rinses, samples were incubated 1 hr at RT in PBS1X/0.3% Triton X-100/BSA 0.1% and then in phalloidin FluoProbes 505 or 547 (Interchim) diluted 1:200 in PBS1X/0.3% Triton X-100/BSA 0.1% at 4°C for 48 hr. The samples were washed 3 times for 15 min in PBS1X and mounted in Vectashield. All fluorescent observations were performed on at least five different pairs of testes or antennae. Quantifications were performed on at least three independent experiments.
Most slides were imaged using either the IX83 microscope from Olympus equipped with an iXon Ultra 888 EMCDD camera from Andor and the IQ3 software from Andor. The PlanApo N Apochromat 60 × 1.42 NA objective from Olympus was used for all acquisitions. Some slides were imaged using an SP5X confocal laser scanning microscope (Leica Biosystems) equipped with the Application Suite software (Leica Biosystems). An HCX Plan-Apochromat CS 63 × 1.4 NA objective (Leica Biosystems) was used for all acquisitions.
All images were processed with ImageJ. Figures were created with Adobe photoshop. Only contrasts and offset were adjusted.
3D-SIM microscopy
Request a detailed protocolTestes or antennae were squashed on 12 diameter round coverslip with a 44 × 60 mm overlaying coverslip. Immunofluorescence protocols were the same as above using PFA. Images were acquired using the Elyra PS.1 system from Zeiss (Carl Zeiss, AG, Jena) equipped with a PCO edge 5.5 camera and the ZEN 2012 SP2 software (black edition). The objective used for all acquisitions is a Plan-apochromat 63 × 1.4 NA.
Antibodies
The antibodies used were the following: mouse anti-Futsch/22C10 (1:250; DSHB = 22C10), mouse anti-γ-Tubulin (1:500; Sigma), rabbit anti-GFP (1:1000; Abcam), guinea pig and rat anti-Asterless (1:50000; gift from C. Rogers, University of Arizona, Tucson, USA; Klebba et al., 2013; McLamarrah et al., 2018).
The following secondary antibodies were used (all at 1:1000 dilution): goat anti-mouse Alexa 594, goat anti-rabbit Alexa 488 or Alexa 647, goat anti-guinea pig Alexa 488 or Alexa 594, goat anti-rat Alexa 488 or Alexa 555 or Alexa 647 (Invitrogen) and donkey anti-guinea pig Alexa 647 (Jackson Immuno Research).
Quantifications
Request a detailed protocolBang assays were performed as previously described (Enjolras et al., 2012). Approximately 20 staged female flies were placed in graduated tubes (8 cm) and banged on the table at t = 0. The % of flies that reached each quarter of the tube was counted after 30 s. three different batchs were quantified for each genotype.
Quantifications of mother versus daughter centrioles were based on the principle that the daughter centriole, being nucleated from the mother centriole, is orthogonally positioned on the lateral wall of the mother centriole. Quantifications of TZ protein intensities was performed using ImageJ and by measuring the sum of pixel intensity in a defined region encompassing the centrioles. Background intensity was measured by measuring the sum of pixel intensity in an area close to and devoid of centrioles and then subtracted to TZ protein intensities.
Statistics
Results of fluorescence intensity quantifications are represented as scatter plots with the mean and SD on all figures. Statistical significance was determined by two-tailed unpaired parametric Student’s t test (Figure 4A, fertility assay; Figure 5D, Cby-Tom; Figure 6A, Cep290-GFP in fam921) or non-parametric Mann-Withney’s test when variances were not comparable (all other intensity quantifications of Figure 5D and Figure 6A–C). Results of phenotypic proportions are represented as contingency bar graph and statistical significance was determined by two-tailed Chi-square (Figures 5D and 6A). (Prism six software; ns, p>0.05; *, p≤0.05; **, p≤0.01; ***, p≤0.001; ****, p≤0.0001).
Electron microscopy
Request a detailed protocolSamples were processed as previously described (Enjolras et al., 2012; Vieillard et al., 2015). Observations were performed on at least two independent tissue samples.
Data availability
All data generated or analysed during this study are included in the manuscript and supporting files.
References
-
Transition zone migration: a mechanism for cytoplasmic ciliogenesis and postaxonemal centriole elongationCold Spring Harbor Perspectives in Biology 9:a028142.https://doi.org/10.1101/cshperspect.a028142
-
The ciliopathies: an emerging class of human genetic disordersAnnual Review of Genomics and Human Genetics 7:125–148.https://doi.org/10.1146/annurev.genom.7.080505.115610
-
Making sense of cilia in disease: the human ciliopathiesAmerican Journal of Medical Genetics Part C: Seminars in Medical Genetics 151C:281–295.https://doi.org/10.1002/ajmg.c.30231
-
Chibby promotes ciliary vesicle formation and basal body docking during airway cell differentiationThe Journal of Cell Biology 207:123–137.https://doi.org/10.1083/jcb.201406140
-
CEP290 tethers flagellar transition zone microtubules to the membrane and regulates flagellar protein contentThe Journal of Cell Biology 190:927–940.https://doi.org/10.1083/jcb.201006105
-
The ciliary transition zone: from morphology and molecules to medicineTrends in Cell Biology 22:201–210.https://doi.org/10.1016/j.tcb.2012.02.001
-
Drosophila chibby is required for basal body formation and ciliogenesis but not for wg signalingThe Journal of Cell Biology 197:313–325.https://doi.org/10.1083/jcb.201109148
-
Drosophila spermiogenesis: big things come from little packagesSpermatogenesis 2:197–212.https://doi.org/10.4161/spmg.21798
-
CP110 exhibits novel regulatory activities during centriole assembly in DrosophilaThe Journal of Cell Biology 203:785–799.https://doi.org/10.1083/jcb.201305109
-
Asterless is required for centriole length control and sperm developmentThe Journal of Cell Biology 213:435–450.https://doi.org/10.1083/jcb.201501120
-
The zn finger protein Iguana impacts hedgehog signaling by promoting ciliogenesisDevelopmental Biology 337:148–156.https://doi.org/10.1016/j.ydbio.2009.10.025
-
Centrioles in flies: the exception to the rule?Seminars in Cell & Developmental Biology 21:163–173.https://doi.org/10.1016/j.semcdb.2009.07.001
-
The ciliary transition zone: finding the pieces and assembling the gateMolecules and Cells 40:243–253.https://doi.org/10.14348/molcells.2017.0054
-
The cilium-like region of the Drosophila spermatocyte: an emerging flagellum?Journal of Cell Science 126:5441–5452.https://doi.org/10.1242/jcs.136523
-
CRISPR-Cas9 genome editing in DrosophilaCurrent Protocols in Molecular Biology 111:1–20.https://doi.org/10.1002/0471142727.mb3102s111
-
Phosphatidylinositol 4,5-bisphosphate regulates cilium transition zone maturation in Drosophila MelanogasterJournal of Cell Science 131:jcs218297.https://doi.org/10.1242/jcs.218297
-
BAR Domain-Containing FAM92 proteins interact with Chibby1 to facilitate ciliogenesisMolecular and Cellular Biology 36:2668–2680.https://doi.org/10.1128/MCB.00160-16
-
An ordered pattern of Ana2 phosphorylation by Plk4 is required for centriole assemblyThe Journal of Cell Biology 217:1217–1231.https://doi.org/10.1083/jcb.201605106
-
BookCytodifferentiation During Spermatogenesis in Drosophila Melanogaster: An Electron Microscope StudyThe University of Michigan.
-
Centrobin is essential for C-tubule assembly and flagellum development in Drosophila Melanogaster spermatogenesisThe Journal of Cell Biology 217:2365–2372.https://doi.org/10.1083/jcb.201801032
-
Unique properties of Drosophila spermatocyte primary ciliaBiology Open 2:1137–1147.https://doi.org/10.1242/bio.20135355
-
FAM92A underlies nonsyndromic postaxial polydactyly in humans and an abnormal limb and digit skeletal phenotype in miceJournal of Bone and Mineral Research 34:375–386.https://doi.org/10.1002/jbmr.3594
-
Hemingway is required for sperm flagella assembly and ciliary motility in DrosophilaMolecular Biology of the Cell 25:1276–1286.https://doi.org/10.1091/mbc.E13-10-0616
-
Imaging cilia in Drosophila MelanogasterMethods in Cell Biology 127:279–302.https://doi.org/10.1016/bs.mcb.2014.12.009
-
Transition zone assembly and its contribution to axoneme formation in Drosophila male germ cellsThe Journal of Cell Biology 214:875–889.https://doi.org/10.1083/jcb.201603086
-
Inactivation of chibby affects function of motile airway ciliaThe Journal of Cell Biology 185:225–233.https://doi.org/10.1083/jcb.200809144
-
Centrosomal protein DZIP1 regulates hedgehog signaling by promoting cytoplasmic retention of transcription factor GLI3 and affecting ciliogenesisJournal of Biological Chemistry 288:29518–29529.https://doi.org/10.1074/jbc.M113.492066
-
A mouse model for Meckel syndrome reveals Mks1 is required for ciliogenesis and hedgehog signalingHuman Molecular Genetics 18:4565–4575.https://doi.org/10.1093/hmg/ddp422
-
Depletion of plasma membrane PtdIns(4,5)P2 reveals essential roles for phosphoinositides in Flagellar biogenesisJournal of Cell Science 121:1076–1084.https://doi.org/10.1242/jcs.024927
-
The essential roles of transition fibers in the context of ciliaCurrent Opinion in Cell Biology 35:98–105.https://doi.org/10.1016/j.ceb.2015.04.015
Article and author information
Author details
Funding
Agence Nationale de la Recherche (DIVERCIL)
- Jean-André Lapart
- Elisabeth Cortier
- Jean-Luc Duteyrat
- Céline Augière
- Julie Jerber
- Joëlle Thomas
- Bénédicte Durand
Fondation pour la Recherche Médicale (FRM DEQ20131029168)
- Jean-André Lapart
- Elisabeth Cortier
- Jean-Luc Duteyrat
- Céline Augière
- Julie Jerber
- Joëlle Thomas
- Bénédicte Durand
Université Claude Bernard Lyon 1 (PhD Fellowship)
- Céline Augière
The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Acknowledgements
We thank T Avidor-Reiss for the cep290::GFP Drosophila stock and C Rogers for sharing the rat and guinea pig anti-Asterless. Many thanks to E Goillot for helpful discussions and M NGassa for technical assistance. This project was supported by a grant from the Fondation pour la Recherche Médicale, FRM DEQ20131029168 and the Agence Nationale de la Recherche (ANR Divercil). JAL was supported by the FRM DEQ20131029168. CA was supported by a PhD fellowship from the University Claude Bernard Lyon-1. We thank the PLATIM, CiQLE and CTµ platforms of the LyMIC for confocal and EM microscopy. We are grateful to Elodie Chatre for helpful discussions.
Mass spectrometry experiments were carried out using facilities of the Plaform ‘Proteomic Imaging and molecular Interactions’ (PP2I) of Montpellier.
Copyright
© 2019, Lapart et al.
This article is distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use and redistribution provided that the original author and source are credited.
Metrics
-
- 2,020
- views
-
- 247
- downloads
-
- 28
- citations
Views, downloads and citations are aggregated across all versions of this paper published by eLife.
Citations by DOI
-
- 28
- citations for umbrella DOI https://doi.org/10.7554/eLife.49307
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Dzip1 and Fam92 form a ciliary transition zone complex with cell type specific roles in Drosophila
eLife 8:e49307.
https://doi.org/10.7554/eLife.49307




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}