Dzip1 and Fam92 form a ciliary transition zone complex with cell type specific roles in Drosophila
- Institut NeuroMyoGène, CNRS UMR 5310, INSERM U1217, Université Claude Bernard Lyon 1, France
- Institute of Human Genetics, Universitätsklinikum Düsseldorf, Heinrich-Heine-Universität Düsseldorf, Germany
- IRCM, INSERM, Université de Montpellier, ICM, France
Figures
Figure 1 with 2 supplements
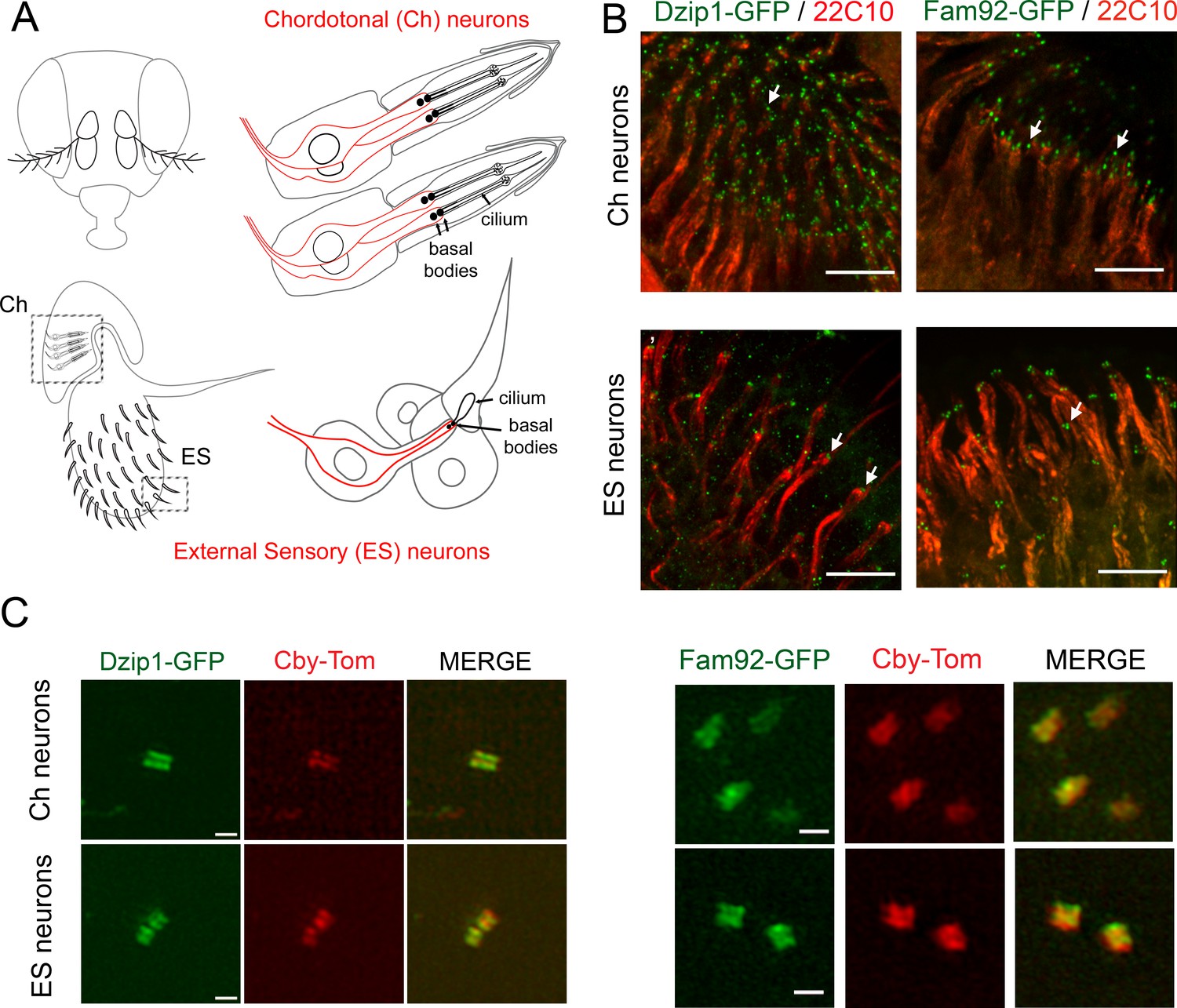
Drosophila Dzip1 and Fam92 are expressed in ciliated sensory neurons and locate at the ciliary transition zone.
(A) Scheme of the two types of sensory organs (or scolopidia) present in the antennae of the Drosophila head. Chordotonal scolopidia are proprioceptors present in the second antennal segment of the Drosophila head and respond to sound vibrations and gravity. Each chordotonal scolopidia is composed of several support cells (gray) and comprise two or three ciliated neurons (red). External sensory organs (or scolopidia) are present under each sensory sensilla of the third antennal segment and react to olfactory and chemical stimuli. In other body parts, ES organs also respond to mechanical stimuli. External sensory scolopidia comprise support cells (gray) and only one ciliated neuron (red). In ciliated sensory neurons, the two centrioles (or proximal and distal basal bodies) stand above each other at the tip of the dendrites. (B) Whole-mount staining of the second and third antennal segment showing neuronal cell bodies and dendrites (22C10), Dzip1-GFP and Fam92-GFP. Dzip1-GFP and Fam92-GFP are present at the tip of the dendrites in each type of neurons. Arrows point both to Dzip1-GFP and Fam92-GFP localization at the tip of the dendrites. (C) 3D-SIM imaging of Ch and ES neurons. Both Drosophila Dzip1 and Fam92 overlap with Cby at the transition zone. Bars = 10 μm for (B); = 0.5 μm for (C).
Figure 1—figure supplement 1
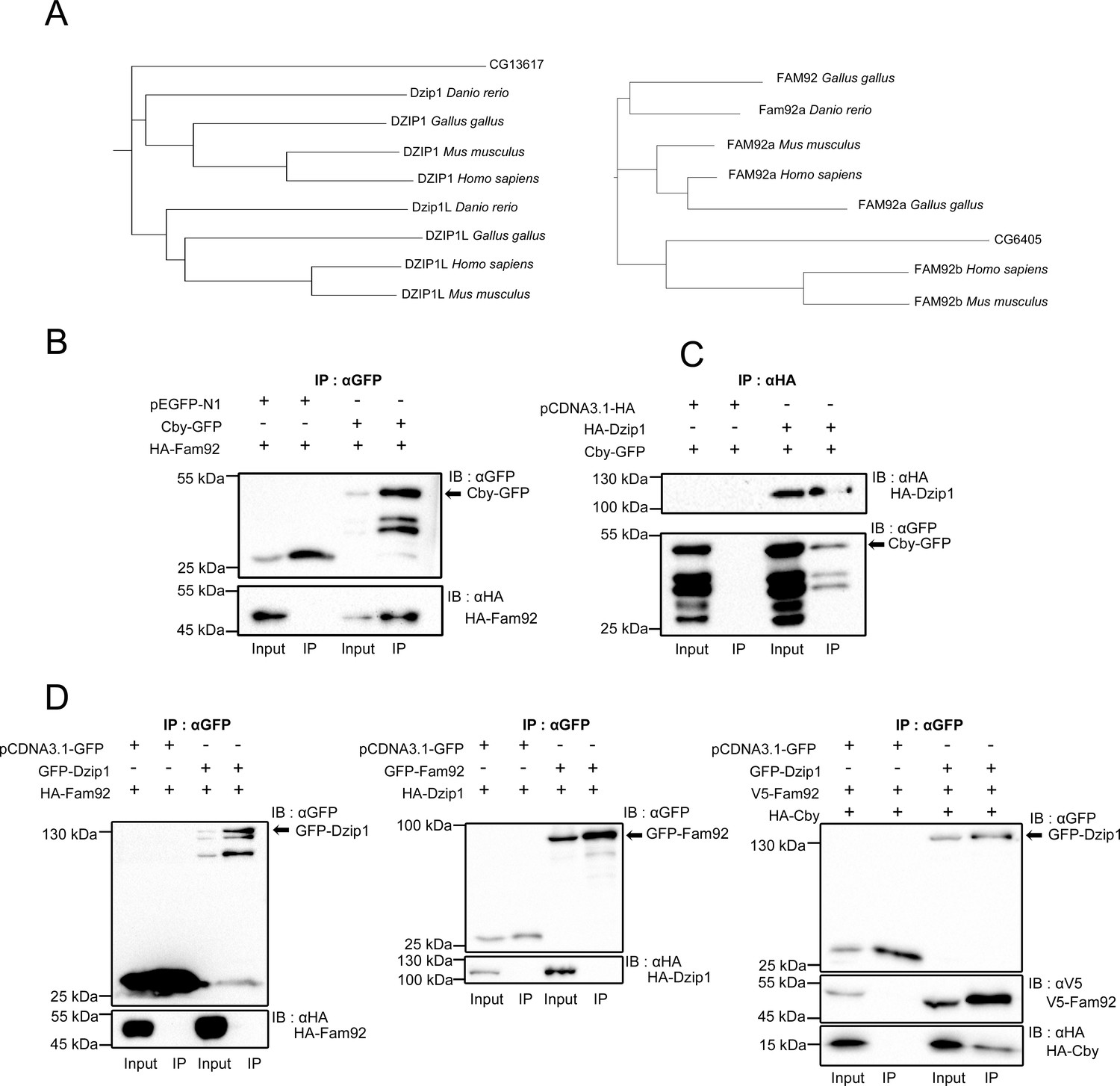
Drosophila Dzip1 and Fam92 interact with Cby.
(A) Phylogenic trees of dzip1 and fam92. dzip1 and fam92 genes are conserved in Drosophila and in protozoa. (B–D) COS-7 or HEK293 cells were transfected with the indicated plasmids and cell lysates were immunoprecipitated with GFP or HA antibodies coupled beads. Both cell lysates (Input) and immunoprecipitates (IP) were detected by western blotting (IB) with GFP, HA or V5 antibodies. Cby-GFP band is indicated by an arrow. (B) Drosophila Fam92 specifically interacts with Drosophila Cby but not with GFP. (C) Drosophila Cby specifically interacts with Drosophila Dzip1 but not with HA epitope. (D) Drosophila Dzip1 specifically interacts with Fam92 only in the presence of Cby. GFP-Dzip1 and GFP-Fam92 are indicated by an arrow.
Figure 1—figure supplement 2
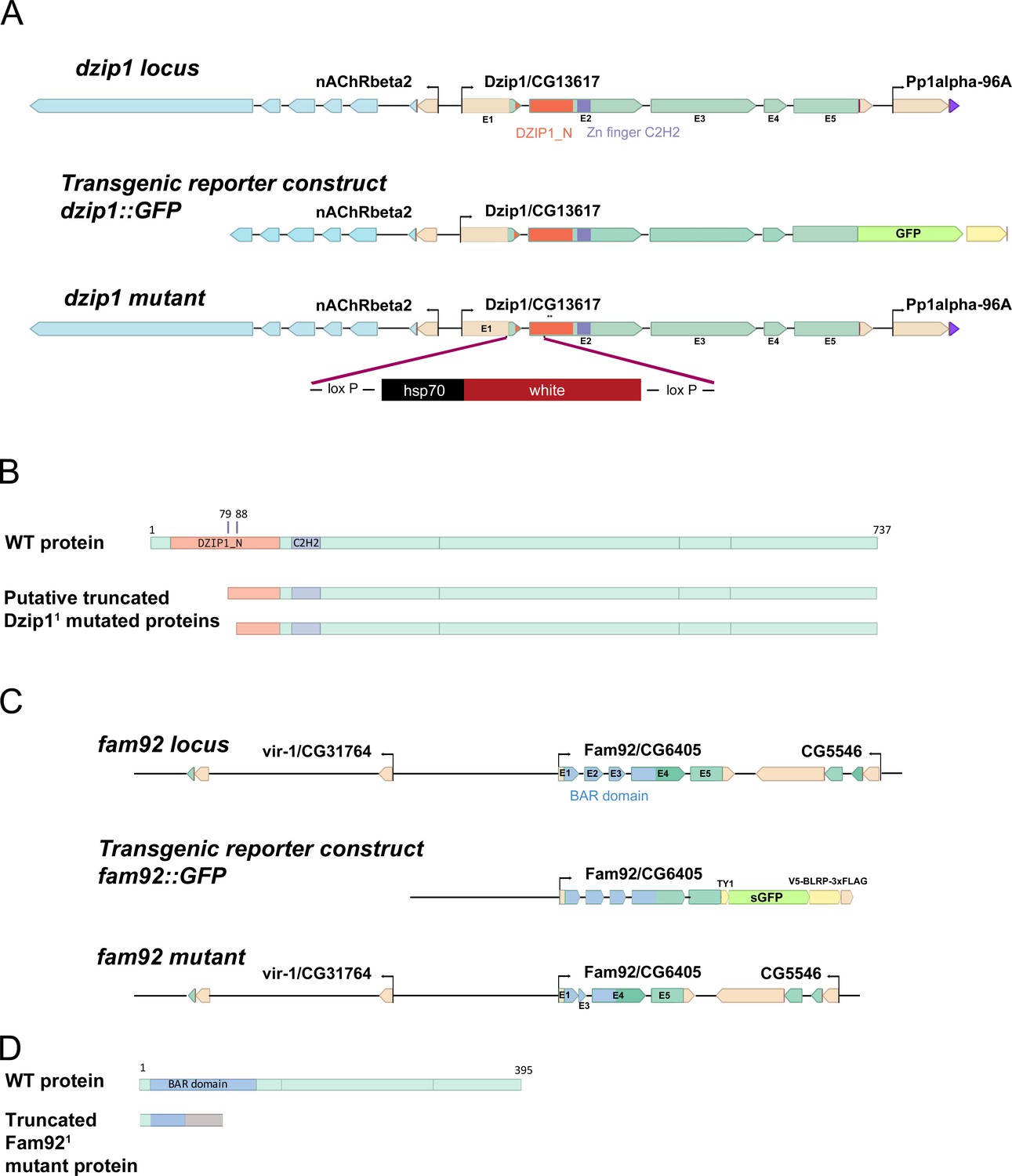
dzip1 and fam92 loci and genetic tools.
(A) Scheme of dzip1 locus, transgenic reporter and dzip11 mutant. dzip1::GFP reporter construct comprises upstream regulatory sequences and the entire coding sequence in frame with GFP. In dzip11 mutant, coding sequences from exon1, including the start codon, to exon two were replaced by the white gene using CRISPR/Cas9-induced homologous recombination. Asterisks indicate the presence of two potential start codons. (B) Scheme of the structure of the WT protein or putative truncated proteins produced by the dzip11 mutant gene. At least 649 aa are retained in the putative Dzip11 mutant protein. The first 79 aa of the WT protein including half of the Dzip1 N-ter conserved domain are absent in the putative Dzip11 mutant protein. (C) Scheme of fam92 locus, transgenic reporter and fam92 mutant. fam92::GFP reporter construct contains a GFP coding sequence in frame with the fam92 coding sequence under the control of its own endogenous promoter. fam921 mutant was obtained by CRISPR/Cas9 cleavage, leading to the complete removal of exon two and part of exon three generating a frameshift in the fam92 coding sequence. (D) Scheme of the structure of the WT protein. The truncated protein produced by the Fam921 mutant allele retains 47 aa from the WT protein and 39 aa translated from the frameshifted open reading frame.
Figure 2

Drosophila Dzip1 and Fam92 are located at the transition zone during spermatogenesis.
(A) Scheme illustrating centriole/basal bodies (BB) behavior during spermatogenesis in male germ cells. In early spermatocytes, two pairs of centrioles (gray) are present in each cell. During spermatocyte maturation, centrioles convert to BB and dock to the plasma membrane while extending a primary like cilium which is entirely decorated with TZ proteins (green and red). In late spermatocytes, the TZ, also described as the primary like cilium, reaches approximatively 500 nm and protrudes at the cell surface. During meiosis, BB with primary like cilia/TZ are internalized, thus retaining the ciliary membrane (yellow) connected to the plasma membrane, hence creating a membrane cap in spermatids. In round spermatid, BB are apposed to the nuclear membrane and the ciliary cap (yellow) connected to the plasma membrane is extended. The ring centriole (green) marks the base of the ciliary cap which is decorated by TZ proteins. At the onset of axoneme elongation, axonemal microtubule nucleation inside the cap extends the ciliary cap/TZ, and as the axoneme grows (black), the ring centriole is pushed away from the basal body. (B–C) Confocal imaging of whole-mount testes showing Dzip1-GFP (B), Fam92-GFP (C), Cby-Tom and Asterless (centrioles). Dzip1 and Fam92 appear together with Cby in early spermatocytes at the tip of centrioles. In elongating spermatids, Drosophila Dzip1 and Fam92 mark the ring centriole (arrows) separating from the BB (asterless, gray). (D) 3D-SIM imaging of male germ cells. Drosophila Dzip1 and Fam92 overlap with Cby in spermatocytes. Plots of the intensity profile of the centrioles along the dotted lines illustrate the overlay between Cby-Tom and Dzip-GFP or Fam92-GFP. Bars = 1 μm.
Figure 3
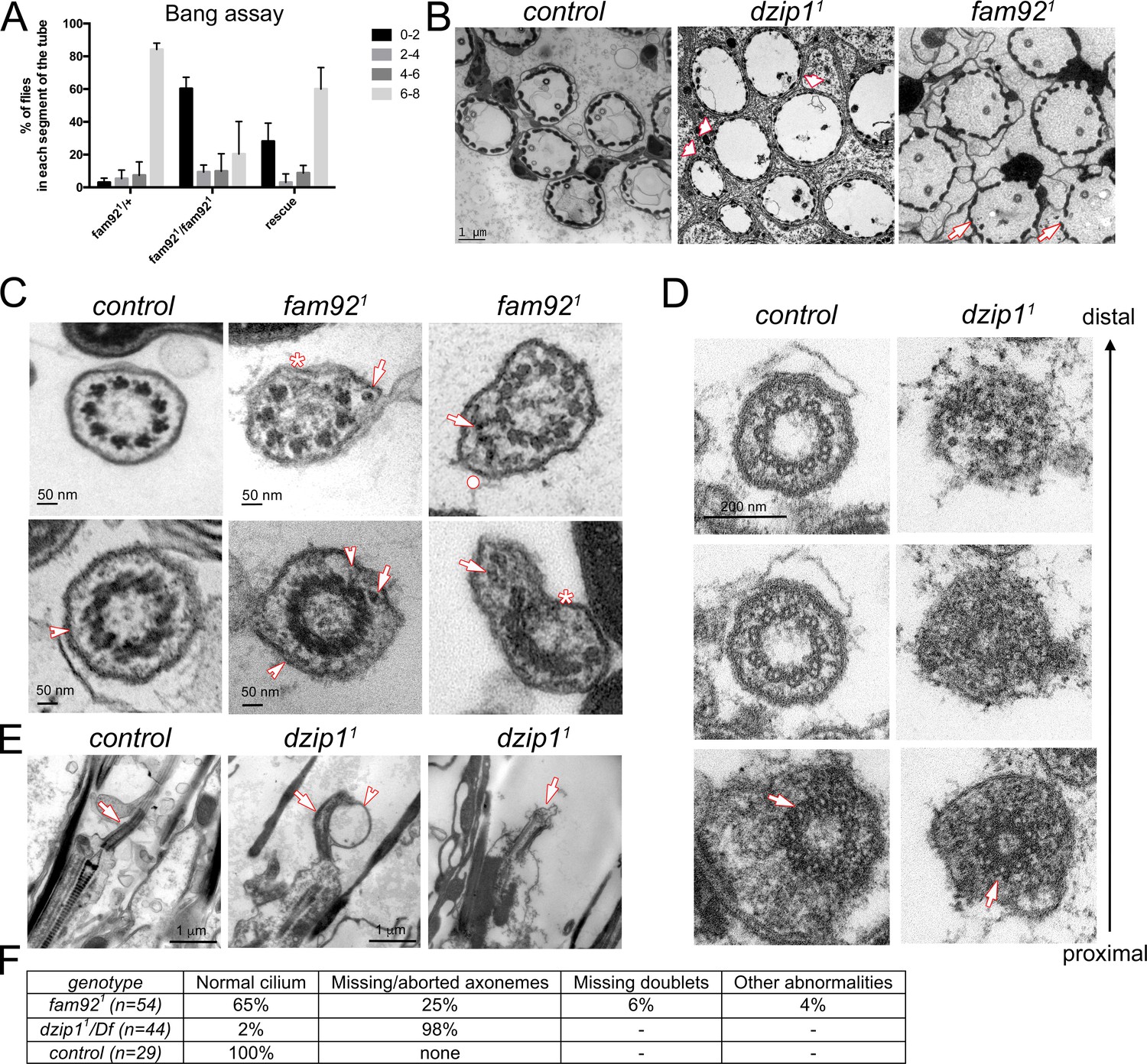
Dzip1 and Fam92 are required for ciliogenesis in Drosophila sensory cilia.
(A) Quantification by bang assay of geotaxis response of fam921 flies compared to control or rescued flies. The percentage of flies that reach a defined level in the tube (0–2, 2–4, 4–6, 6–8) is represented. fam921 flies are unable to climb along the tube compared to control flies. This defect is partially rescued by adding two copies of fam92::GFP transgene (fam921/+ n = 61, fam921/fam921 n = 63, rescue n = 68). (B) EM analysis of cross sections of antennal Chordotonal (Ch) neurons. Whereas two neurons/cilia can be observed in each control scolopidia, cilia are almost completely absent in dzip11 scolopidia (arrowheads). In fam921, reduced number of cilia are observed on several Ch neurons (arrows). (C) Ch neurons cilia ultrastructure of fam921 antennae, showing reduced number of microtubule doublets (asterisks) and/or accumulation of dense material (arrows) underneath the ciliary membrane and deformation of the membrane (dot). Similar defects are also observed on cross sections of the TZ (lower panels). Note that the linkers connecting the axoneme to the membrane are still present (arrowheads). (D) Serial sections of the basal body-transition zone region of Ch neurons in dzip11 compared to control, from the basal body (proximal, lower panels) to the TZ (distal, upper panels). Whereas doublet microtubules are present and symmetrically organized at the basal body level (arrows, lower panels), they fail to elongate along the transition zone which is incompletely assembled. (E) Longitudinal sections of dzip11 showing basal body with aberrant TZ compared to control (arrows). Membrane bulges (arrowhead) along the aberrant dzip11 TZ can also be detected. (F) Quantifications of the cilium defects observed in scolopidia.
Figure 4 with 1 supplement
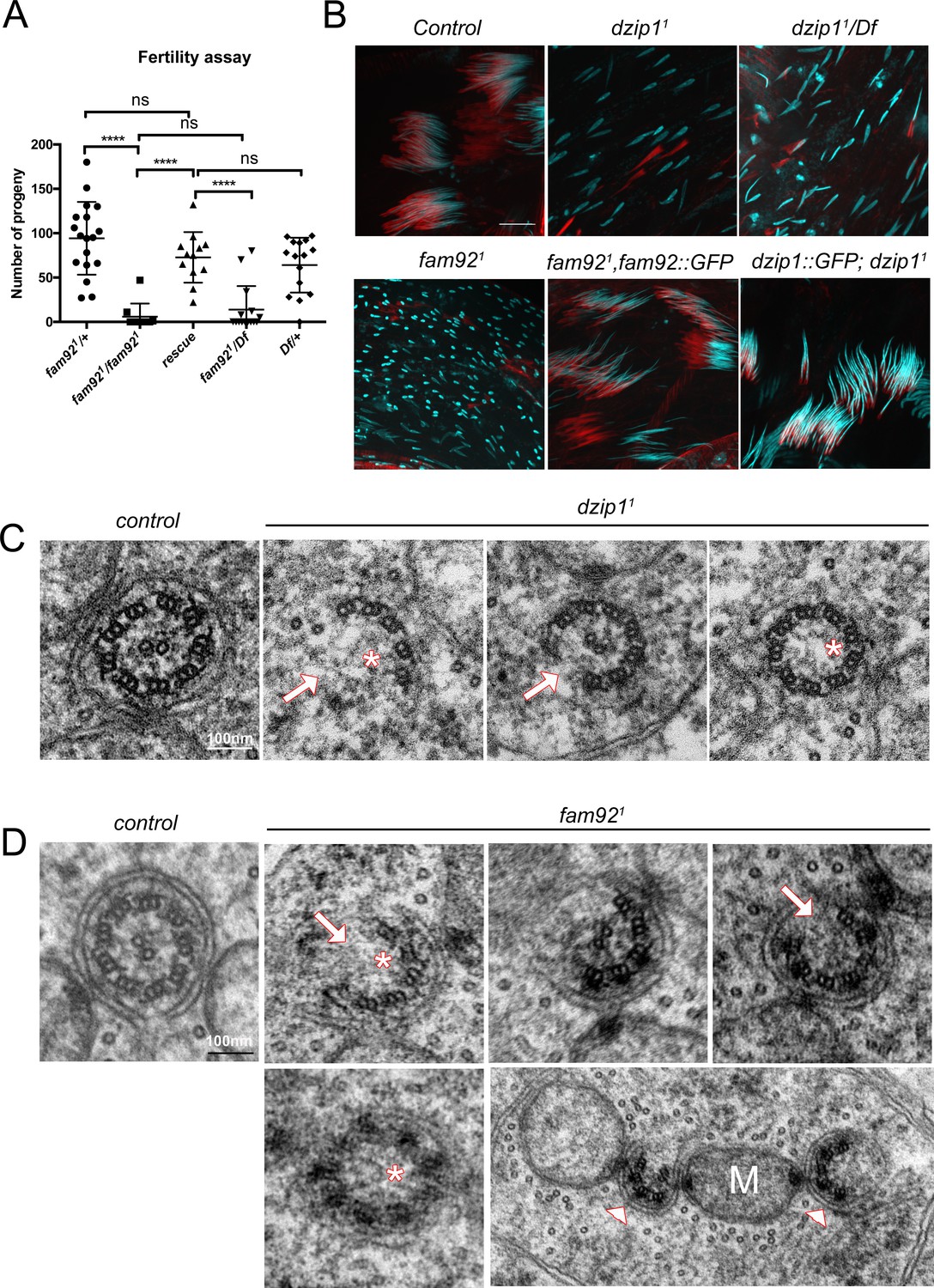
Dzip1 and Fam92 are required for flagella formation.
(A) Fertility assays of fam921 males showing severe reduction of fertility in fam921 or fam921/Df compared to control or rescued flies (fam921/+ n = 18; fam921/fam921 n = 10; rescue n = 15; fam921/Df n = 15; Df/+ n = 16) ****, p<0.0001; ns, p>0,05. (B) Confocal analysis of whole mount testes of control, dzip11, dzip11/Df, fam921 and rescued genotypes stained for nucleus (Hoechst, cyan) and actin cones (phalloidin, red). Nuclei are dispersed in mutant cysts compared to controls, which could reflect axonemal elongation defects. As a consequence, actin cones required for spermatid individualization are mislocalized. Axoneme elongation and spermatid individualization are normal in rescued flies. Bars = 10 μm. (C) EM analysis of cross sections of axoneme of round spermatid. In dzip11, flagella ultrastructure is altered with missing microtubule doublets (arrows) or central pair (asterisks). (D) In fam921 mutant spermatids, most axonemes show altered ultrastructure, with missing microtubule central pairs (asterisks) or doublets (arrows), broken symmetry with each part of the axoneme being relocated along the mitochondria (M, arrowheads).
Figure 4—figure supplement 1
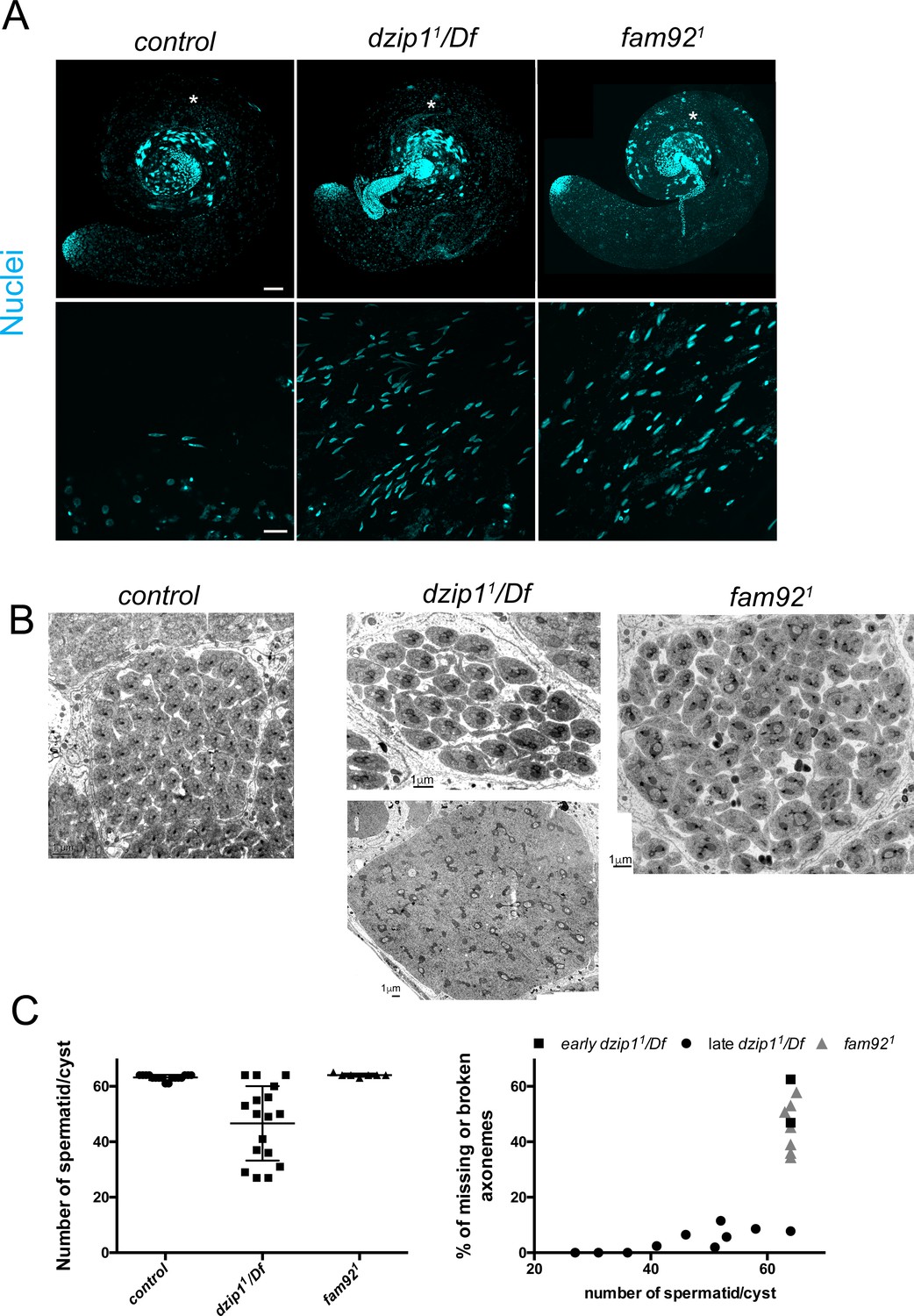
Dzip11 and fam921 testis phenotypes.
(A) Confocal analysis of whole mount testes of control, dzip11/Df, and fam921 showing nuclei dispersion (Hoechst) which is stronger in dzip11/Df than fam921. Magnified view in boxes were taken from the region marked with an asterisk. Bars = 50 μm and 10 μm respectively for upper and lower panel. (B) EM observations of testis cross sections. In control and fam921 cysts, 63 or 64 spermatids are observed in each cysts, whereas a reduced number of spermatid/cysts is observed in older dzip11/Df cysts (upper panel) compared to very early cysts (lower panel). Note that mitochondria are misshaped with apparent cytoplasmic inclusions. (C) Axonemal defects are present in dzip11/Df and fam921 cysts. In dzip11/Df, more defects are observed in young cysts compared to old cysts.
Figure 5 with 1 supplement
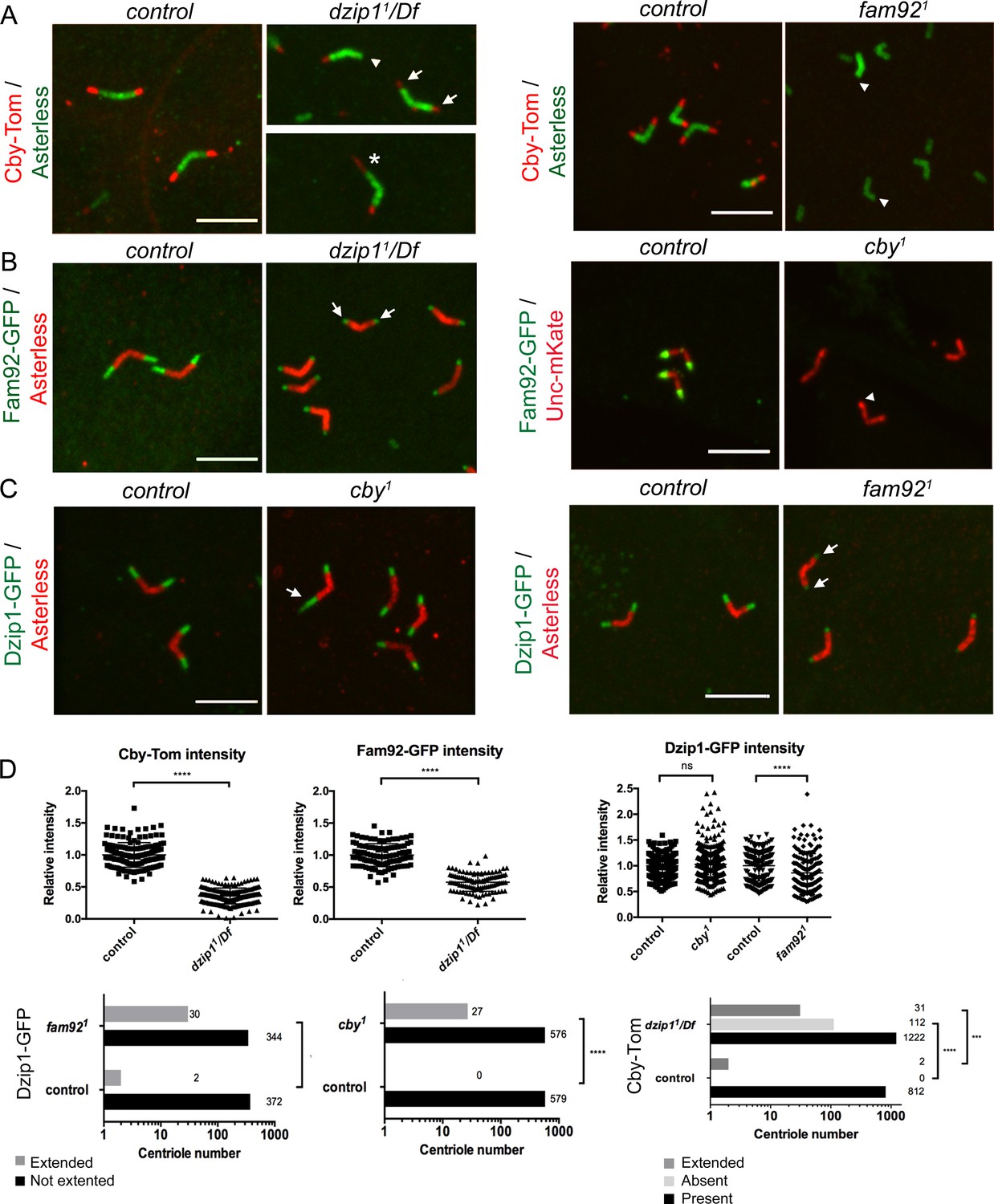
Dzip1, Fam92 and Cby work coordinately at the TZ.
(A) Cby-Tom is present at the tip of the centrioles in control spermatocytes but strongly reduced in dzip11/Df (arrows) and can even be lost at one centriole of the pair (arrowhead), but in a few situations (2.3% see D quantifications below), Cby-Tom domain is extended (asterisk). In fam921 mutant, Cby-Tom is completely lost at the tip of the spermatocyte centrioles (arrowheads). (B) Fam92-GFP is reduced at both centrioles of the pair in dzip11/Df (arrows) compared to control and completely lost at the tip of the centrioles (arrowhead) in cby1 mutants. (C) Dzip1-GFP is strikingly expanded in a few cby1 spermatocyte centrioles (arrow, 4.4% see quantifications in D) and is reduced at the tip of both centrioles in fam921 testes (arrows). Bars = 5 μm. (D) Quantifications of the distribution of Dzip1-GFP, Fam92-GFP and Cby-Tom in the different mutant contexts. ****, p<0.0001; ***, p<0.001; ns, p>0,05.
Figure 5—figure supplement 1
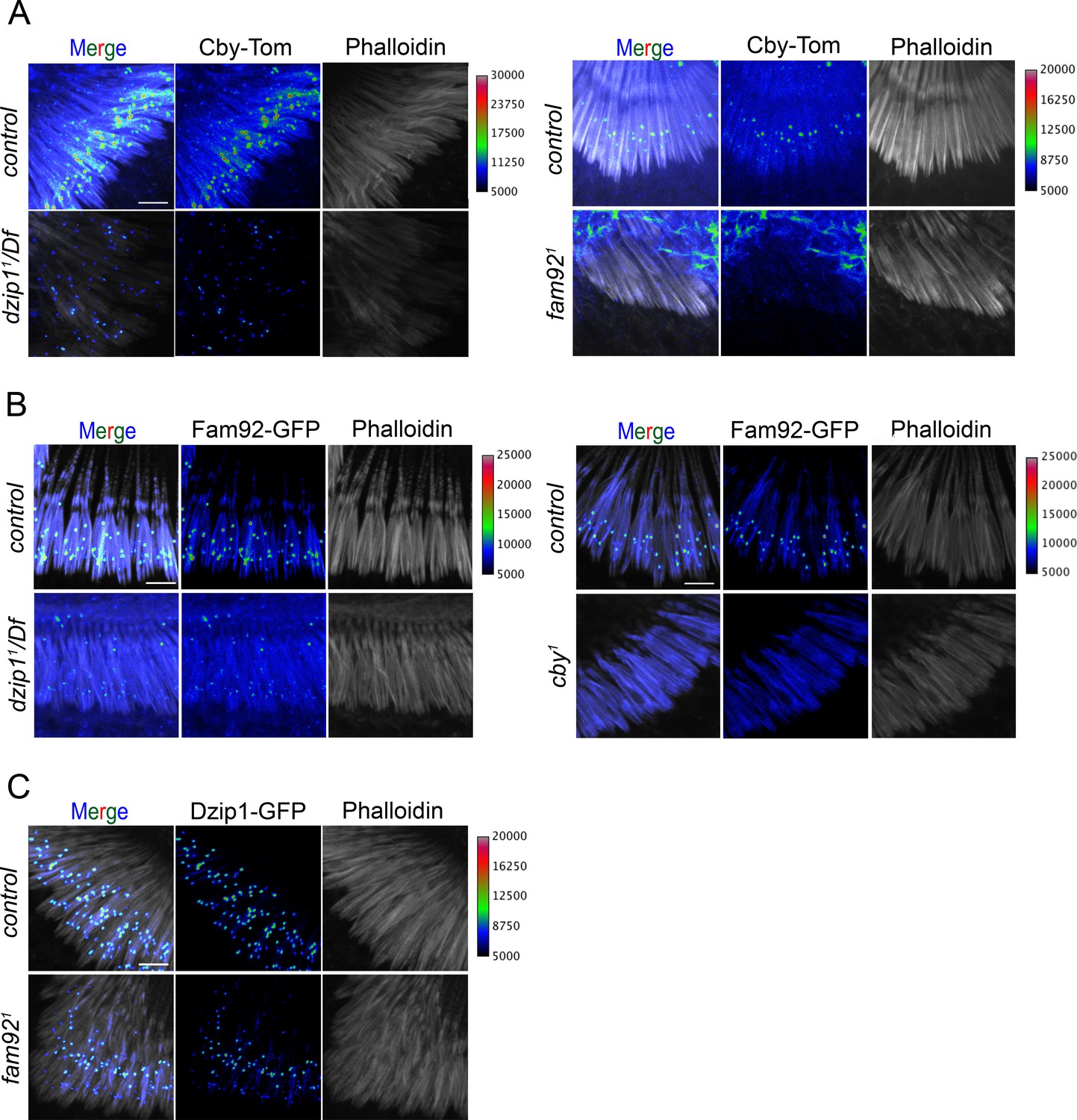
Dzip1, Fam92 and Cby are cooperatively recruited at the TZ in Drosophila sensory cilia.
Confocal analysis of whole mount second antennal segment from dzip11/Df, fam921 or cby1 mutants. Actin staining (phalloidin) reveals scolopale rods that enclose cilia of chordotonal neurons. (A) Whereas Cby-Tom is located at the transition zone at the proximal region of the the scolopidia (phalloidin), it is strongly reduced in dzip11/Df and completely lost in fam921. (B) Fam92-GFP is strongly reduced at the transition zone in dzip11/Df and completely lost in cby1 antennae. (C) Localization of Dzip1-GFP is only slightly affected in fam921. To facilitate visualization of fluorescent signal intensity, GFP or Tomato signals were presented using a pixel intensity heat map with corresponding intensity values presented on the right of each panel. Bars = 5 μm.
Figure 6 with 2 supplements
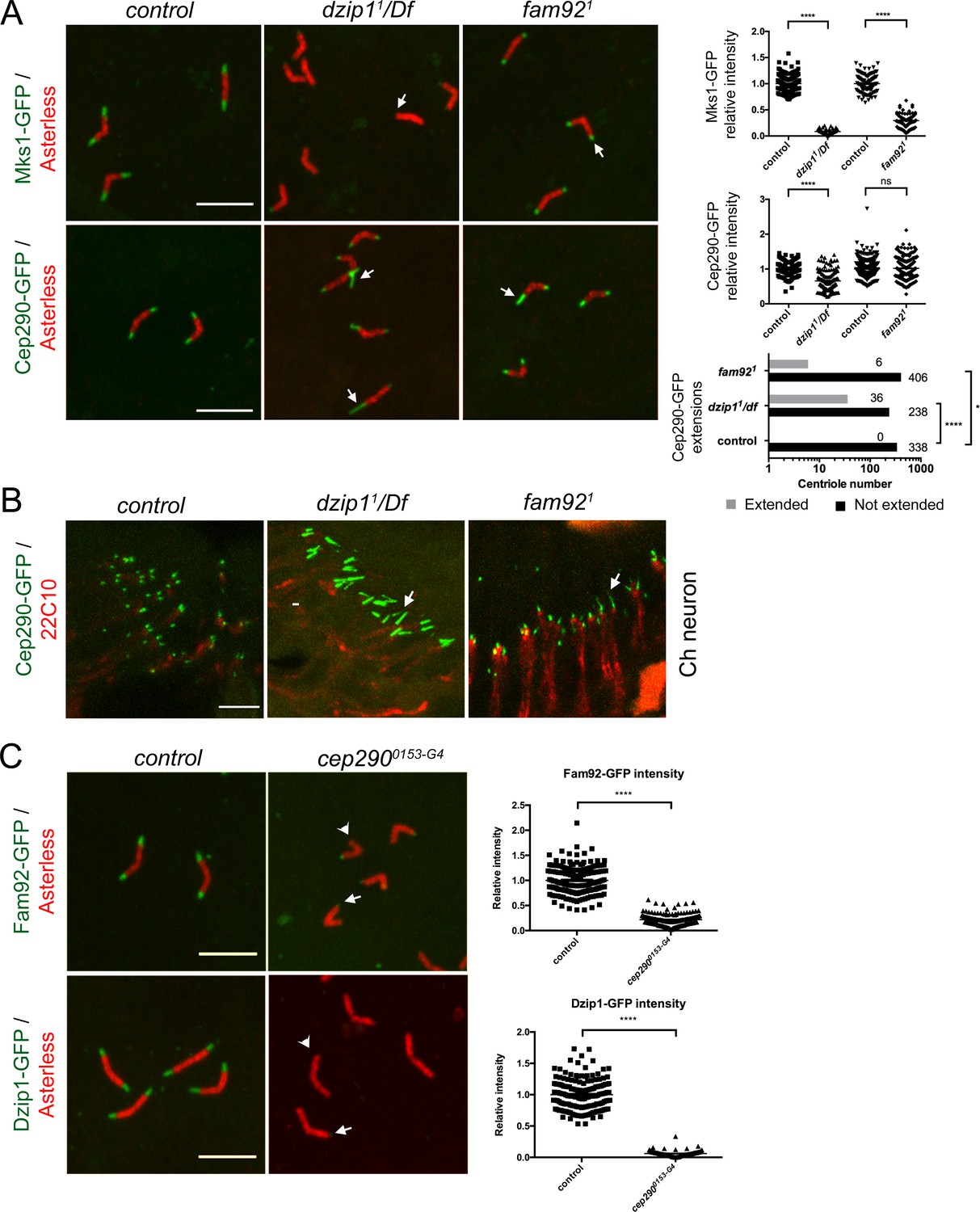
Dzip1 and Fam92 organize the transition zone downstream of Cep290.
(A) In spermatocytes, Mks1-GFP is severely reduced at the ciliary cap or lost in dzip11/Df and fam921 (arrows, quantifications on right graph). Cep290-GFP domain is expanded in dzip11/Df (arrows), but less affected in fam921 (quantifications on right graph). (B) Cep290-GFP is expanded in chordotonal cilia of dzip11/Df or fam921 antennae (arrows). (C) Fam92-GFP and Dzip1-GFP are strongly reduced (arrows) or lost (arrowheads) at the tip of the centrioles in cep2900153-G4 spermatocytes (quantifications are illustrated on the graphs). ****, p<0.0001; *, p<0.01; ns, p>0.05. Bars = 5 μm.
Figure 6—figure supplement 1
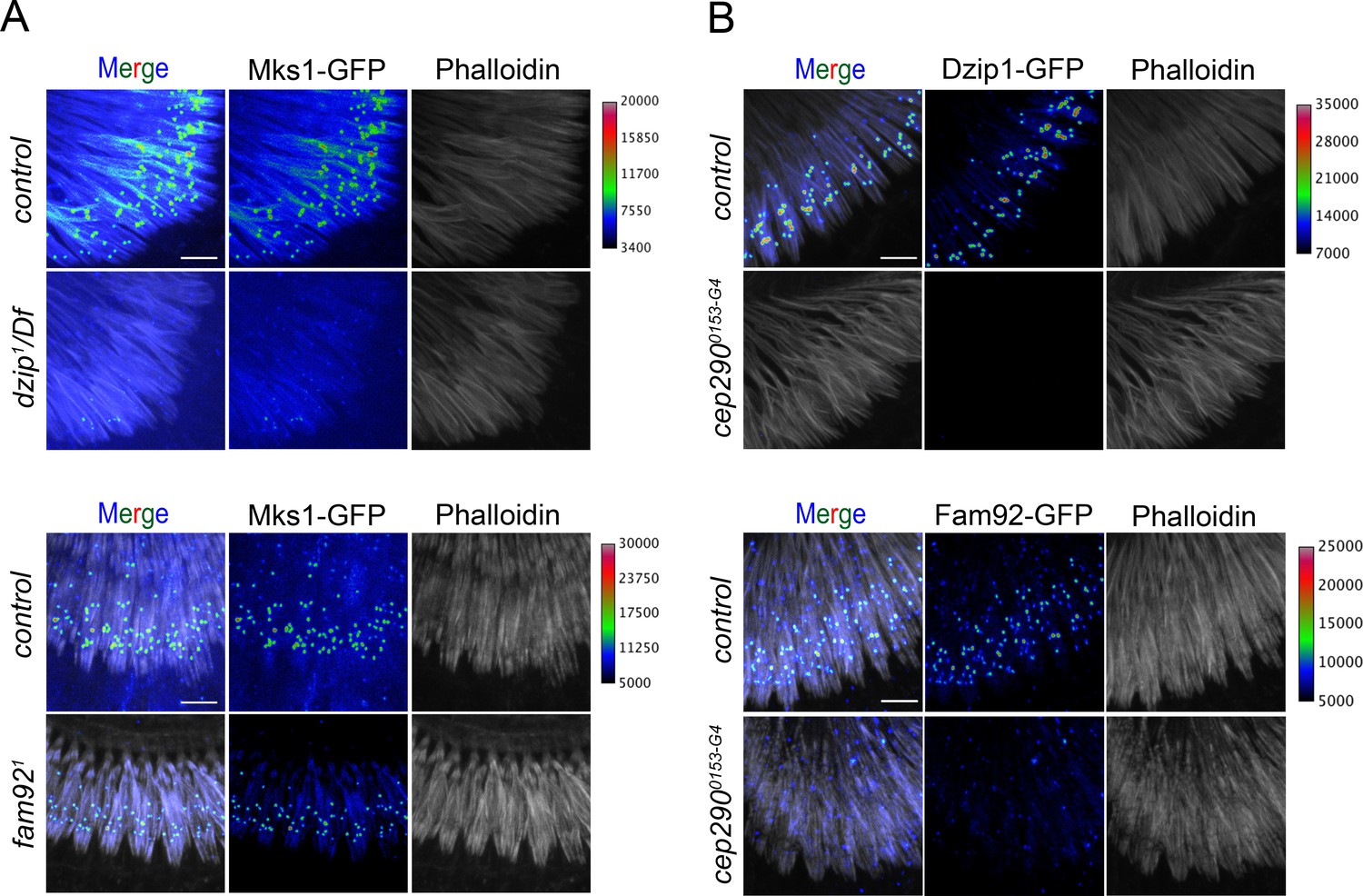
Dzip1 and Fam92 organize the TZ downstream of Cep290 in sensory cilia.
(A) In dzip11/Df, Mks1-GFP is strongly reduced at the transition zone at the base of the scolopidia (phalloidin) and slightly reduced in fam921. (B) Dzip1-GFP and Fam92-GFP are completely lost at the TZ in chordotonal cilia in cep2900153-G4. To facilitate visualization of fluorescent signal intensity, GFP or Tomato signals were presented using a pixel intensity heat map with corresponding intensity values presented on the right of each panel. Bars = 5 μm.
Figure 6—figure supplement 2
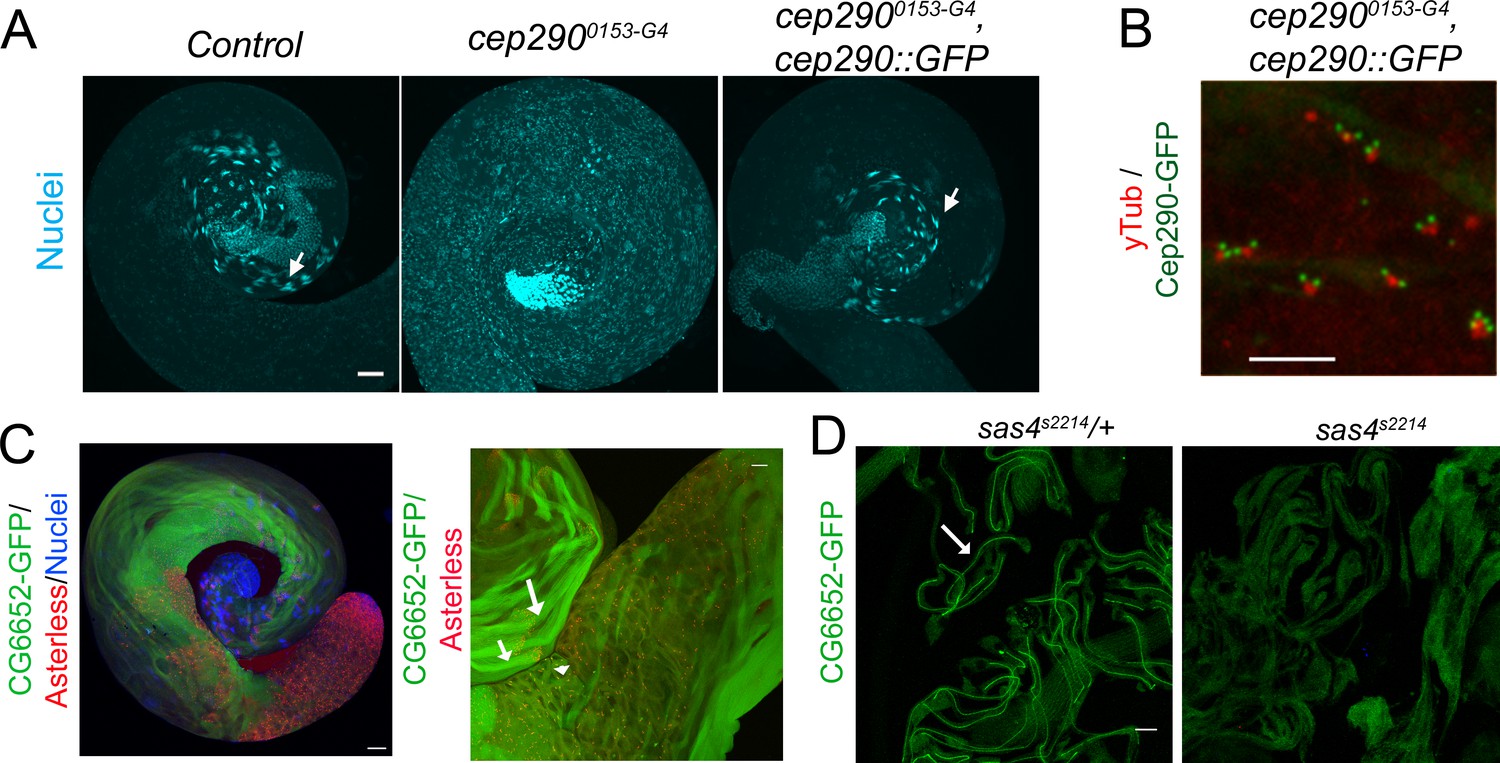
cep2900153-G4 is a strong hypomorphic allele and CG6652 labels the axoneme.
(A) In control spermatocytes, nuclear clusters are clearly observed (arrow). In cep2900153-G4, no nuclear clusters are detected and all nuclei are dispersed along the cyst indicating strong defects in axoneme elongation. This phenotype is reverted in rescued flies (arrow). (B) Cep290-GFP fusion protein localization at the transition zone in cep2900153-G4 at the tip of the centriole (γ-tubulin) in spermatocytes. Bars = 50 μm (A); = 5 μm (B). (C) Whole mount testes expressing CG6652-GFP and labeled with anti-Asterless antibody. At lower magnification left, CG6652-GFP is detectable mostly along spermatid tails. At higher magnification, CG6652-GFP is observed at the tip of centrioles in spermatocytes (arrowhead) and along the flagella in early (short arrow) and late (long arrow) elongated spermatids. Scale bar = 50 μm. (D) In sas4s2214 mutant testes with no centrioles, spermatids elongate but do not exhibit any axonemes (Basto et al., 2006) and CG6652-GFP is not detected compared to the control situation where CG6652-GFP decorates the entire axoneme (arrow). Scale bar = 10 μm.
Figure 7
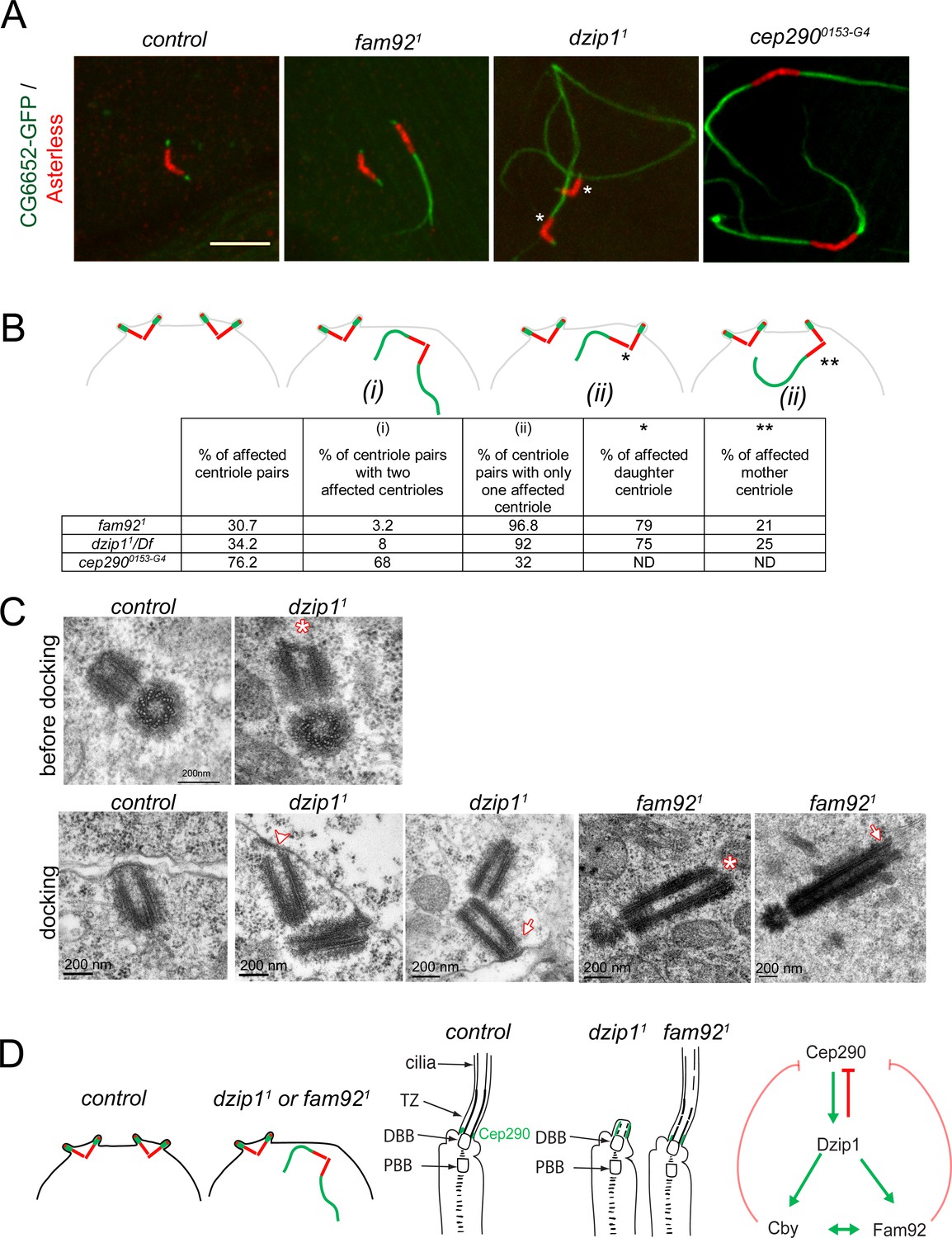
Dzip1 and Fam92 are required for both centriolar docking and proper axonemal formation during Drosophila spermatogenesis.
(A) Confocal imaging of whole-mount testes showing aberrant extensions of axonemes labeled by CG6652-GFP in fam921, dzip11 and cep2900153-G4 spermatocytes. Asterisks point to the centriole in each pair that can be unambiguously assigned as the daughter. Bar = 5 μm (B) Quantifications of the penetrance of CG6652-GFP labeled aberrant axonemes. Schemes representing the different classes that were quantified in the table: (i) two centrioles of the pair are affected; (ii) only one centriole of the pair is affected, either the daughter* or the mother**. (C) EM analysis of cross and longitudinal sections of centrioles in spermatocytes. In dzip11, centriole docking is impaired with partial docking of the centriole (arrowhead) or docking to the plasma membrane of only one centriole of the pair (arrow). Centrioles present an altered/irregular cap, compared with control (asterisk). In fam921, undocked centrioles are also observed in spermatocytes and show irregular distal end (asterisk) or microtubules extending from the tip (arrow). (D) Scheme of the consequences of dzip1 and fam92 loss of function on basal body and cilium assembly in Drosophila ciliated tissues. Whereas basal body anchoring and TZ assembly is affected in male germ cells, only TZ and cilium assembly is affected in sensory cilia. In addition, Cep290-GFP is systematically expanded in dzip1 mutant and occasionally expanded in fam92 mutant chordotonal neurons. The scheme on the right summarizes the functional interactions observed between Dzip1, Cby, Fam92 and Cep290. Red arrows are inhibitory interactions and green arrows define positive interactions.
Videos
Video 1
Real time imaging of dzip11adult flies.
Video 2
Real time imaging of dzip11/Df adult flies.
Tables
Key resources table
| Reagent type (species) or resource | Designation | Source or reference | Identifiers | Additional information |
|---|---|---|---|---|
| Gene (Drosophila melanogaster) | cby | Enjolras et al., 2012 | FLYB: FBgn0067317 | |
| Gene (Drosophila melanogaster) | fam92/CG6405 | This study | FLYB: FBgn0032428 | |
| Gene (Drosophila melanogaster) | dzip1/CG13617 | This study | FLYB: FBgn0039201 | |
| Strain, strain background (Escherichia coli) | DH5alpha | Thermo Fisher Scientific | 18265017 | |
| Genetic reagent (D. melanogaster) | Cep2900153-G4 mutant strain | Bloomington Drosophila Stock Center | BDSC: 62671; FLYB: FBst0062671 RRID:BDSC_62671 | FlyBase symbol: w1118; PBac{IT.GAL4}cep2900153-G4 |
| Genetic reagent (D. melanogaster) | Df(3R)Exel8178 | Bloomington Drosophila Stock Center | BDSC : 7993 FBab0038335 RRID:BDSC_7993 | |
| Genetic reagent (D. melanogaster) | Df(2L)Exel6033 | Bloomington Drosophila Stock Center | BDSC : 7516 FLYB : FBab0037871 RRID:BDSC_7516 | |
| Genetic reagent (D. melanogaster) | sas4s2214 mutant strain | Basto et al., 2006 | BDSC : 12119 FLYB : FBal0196943 RRID:BDSC_12119 | |
| Genetic reagent (D. melanogaster) | cby1 mutant strain | Enjolras et al., 2012 | FLYB : FBal0270281 | |
| Genetic reagent (D. melanogaster) | dzip11 mutant strain | This study | Section Materials and methods “Generation of dzip11 and fam921 alleles” | |
| Genetic reagent (D. melanogaster) | fam921 mutant strain | This study | Section Materials and methods “Generation of dzip11 and fam921 alleles” | |
| Genetic reagent (D. melanogaster) | Unc::mkate | Vieillard et al., 2016 | FLYB : FBal0324713 | |
| Genetic reagent (D. melanogaster) | Cby::Tom | Vieillard et al., 2016 | FLYB : FBal0270280 | |
| Genetic reagent (D. melanogaster) | Cep290::GFP | Basiri et al., 2014 | FLYB : FBal0301636 | |
| Genetic reagent (D. melanogaster) | CG6652::GFP | Vieillard et al., 2016 | FlyB: FBal0324714 | |
| Genetic reagent (D. melanogaster) | mks1::GFP | Vieillard et al., 2016 | FLYB : FBal0324710 | |
| Genetic reagent (D. melanogaster) | fam92::GFP | This study | Section Materials and methods “Plasmids and Drosophila reporter gene constructs” | |
| Genetic reagent (D. melanogaster) | dzip1::GFP | This study | Section Materials and methods “Plasmids and Drosophila reporter gene constructs” | |
| Cell line (Mus musculus) | IMCD3 | ATCC | CRL-2123 | |
| Cell line (Homo-sapiens) | HEK293 | ATCC | CRL-1573 | |
| Cell line (Cercopithecus aethiops) | COS-7 | ATCC | CRL-1651 | |
| Antibody | Mouse monoclonal anti-Futsch/22C10 | DHSB | AB_528403 | IF (1:250) |
| Antibody | Rabbit polyclonal anti-GFP | Abcam | AB6556 | IF(1:1000) WB (1:10000) |
| Antibody | Guinea pig polyclonal anti-Asterless | Klebba et al., 2013 | IF (1:50000) | |
| Antibody | Rat polyclonal anti-Asterless | McLamarrah et al., 2018 | IF (1:50000) | |
| Antibody | Rat monoclonal anti-HA (clone 3F10) | Roche | 11867423001 | WB (1:5000) |
| Antibody | Goat polyclonal anti-rabbit Alexa 488 | Invitrogen | A11008 | IF (1:1000) |
| Antibody | Goat polyclonal anti-rabbit Alexa 647 | Invitrogen | A21244 | IF (1:1000) |
| Antibody | Goat polyclonal anti-guinea pig Alexa 488 | Invitrogen | A11073 | IF (1:1000) |
| Antibody | Goat polyclonal anti-guinea pig Alexa 594 | Invitrogen | A11076 | IF (1:1000) |
| Antibody | Donkey polyclonal anti-guinea pig Alexa 647 | Invitrogen | 706-605-148 | IF (1:1000) |
| Antibody | Goat polyclonal anti-rat Alexa 488 | Invitrogen | A11006 | IF (1:1000) |
| Antibody | Goat polyclonal anti-rat Alexa 555 | Invitrogen | A21434 | IF (1:1000) |
| Antibody | Goat polyclonal anti rat Alexa 647 | Invitrogen | A21247 | IF (1:1000) |
| Antibody | Mouse monoclonal anti-V5 | Invitrogen | R960-25 | WB (1:5000) |
| Antibody | Goat polyclonal anti-rabbit-HRP | Biorad | 170–6515 | WB (1:10000) |
| Antibody | Goat polyclonal anti-mouse-HRP | Biorad | 170–6516 | WB (1:3000) |
| Antibody | Goat polyclonal anti-rat-HRP | Sigma | A5795-1ML | WB (1:20000) |
| Antibody | Mouse monoclonal anti-γ-Tubulin | Sigma | GTU88 | IF (1:500) |
| Chemical compound, drug | phalloidin FluoProbes 547 | Interchim | FP-AZ0330 | IF (1:200) |
| Chemical compound, drug | phalloidin FluoProbes 505 | Interchim | FP-AZ0130 | IF (1:200) |
| Commercial assay or kit | GFP-TRAP | Chromotek | gta-100 | |
| Commercial assay or kit | Mouse monoclonal Anti-HA-agarose antibody | Sigma | A2095 | |
| Commercial assay or kit | S-protein agarose | Merck | 69704 | |
| Commercial assay or kit | Gibson Assembly Master Mix | New England Biolabs, Inc | E5510S | |
| Recombinant DNA reagent | pBFv-U6.2 | Kondo and Ueda, 2013 Nig-Fly | FLYB: FBmc0003127 | |
| Recombinant DNA reagent | pBFv-U6.2B | Kondo and Ueda, 2013 Nig-Fly | FLYB: FBmc0003128 | |
| Recombinant DNA reagent | pG-LAP3 vector | Torres et al., 2009 | Addgene#79704 | |
| Recombinant DNA reagent | pEGFP-N1 | Clontech | Cat #6085–1 |
Additional files
-
Supplementary file 1
Proteins identified by mass spectrometry from two rounds of LAP-Tag-CBY1 purification.
Proteins in red were specifically retrieved in both replicates with LAP-CBY1 but not with LAP-GFP. Proteins in black were specifically retrieved in only one replicate with LAP-CBY1 but not with LAP-GFP. DZIP1L and FAM92A1 are highlighted in yellow
- https://cdn.elifesciences.org/articles/49307/elife-49307-supp1-v1.xlsx
-
Supplementary file 2
Primers used in this study.
- https://cdn.elifesciences.org/articles/49307/elife-49307-supp2-v1.docx
-
Transparent reporting form
- https://cdn.elifesciences.org/articles/49307/elife-49307-transrepform-v1.docx
Download links
A two-part list of links to download the article, or parts of the article, in various formats.
Downloads (link to download the article as PDF)
Open citations (links to open the citations from this article in various online reference manager services)
Cite this article (links to download the citations from this article in formats compatible with various reference manager tools)
Dzip1 and Fam92 form a ciliary transition zone complex with cell type specific roles in Drosophila
eLife 8:e49307.
https://doi.org/10.7554/eLife.49307




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}